
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change , 24 August 2023
Sec. Pests, Pathogens and Invasions
Volume 6 - 2023 | https://doi.org/10.3389/ffgc.2023.1229456
This article is part of the Research Topic Foliar, Shoot, Stem and Rust Diseases of Trees IUFRO 2022 View all 11 articles
 Tamara Sánchez-Gómez1,2*
Tamara Sánchez-Gómez1,2* Steven J. Harte3
Steven J. Harte3 Paula Zamora1,2,4
Paula Zamora1,2,4 Matéo Bareyre1,5Julio Javier Díez1,2Baudilio Herrero6
Matéo Bareyre1,5Julio Javier Díez1,2Baudilio Herrero6 Jonathan Niño-Sánchez1,2
Jonathan Niño-Sánchez1,2 Jorge Martín-García1,2
Jorge Martín-García1,2Introduction and main objective: Bursaphelenchus xylophilus, commonly known as pine wood nematode (PWN), is considered one of the greatest threats to European and Asian pines. Regarding its management, most efforts have been directed toward control measures for the major vector (Monochamus spp.) and screening for genetic resistance in its hosts. However, an integrated pest management strategy which also implements pinewood nematode control is currently lacking. The aim of this study was to evaluate the nematicidal effect of two Beauveria species, a genus well known for its entomopathogenic activity.
Summary methodology: For this purpose, in vitro antagonism tests of fungi (Beauveria bassiana and B. pseudobassiana) and the mycotoxin beauvericin (C45H57N3O9) on B. xylophilus populations were conducted. Finally, the production of beauvericin in B. bassiana and B. pseudobassiana strains was quantified by high-performance liquid chromatography - mass spectrometry (HPLC-MS).
Results and discussion: Both the B. bassiana and B. pseudobassiana fungal species and the mycotoxin beauvericin showed a clear nematicidal effect on B. xylophilus populations, substantially reducing their survival rate and even attaining 100% mortality in one case. HPLC-MS analysis confirmed and quantified the production of beauvericin by B. bassiana and demonstrated for the first-time beauvericin production in B. pseudobassiana.
Final conclusion: These findings highlight the potential of Beauveria species and the mycotoxin beauvericin to be implemented in an integrated pest management strategy to control both nematode and vector.
Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) poses a serious threat, endangering coniferous forests worldwide. This pathogen, commonly known as pine wood nematode (PWN) and belonging to the group of plant-parasitic nematodes (PPNs), causes decay of afflicted trees, leading to the so-called pine wilt disease (PWD) and eventually, tree mortality. Although PWN is thought to originate in North America, PWD was firstly detected in the early twentieth century in Japan and quickly spread to other Asian countries (Kim et al., 2020). In the late 1990s it was introduced into Europe via Portugal (Mota et al., 1999) where it has subsequently caused significant forest damage. PWD is now found in more than 30% of the Portuguese forest area (De la Fuente et al., 2018) and in several Spanish regions bordering Portugal: Extremadura, Galicia and Castilla y León (Zamora et al., 2015). Although the pathogen currently appears fairly contained, it is listed as a quarantine pest in Europe (List A2) (EPPO, 2016) due to its high pathogenicity and dispersive capacity. Moreover, the impact is predicted to worsen under future climate change scenarios (Hirata et al., 2017; De la Fuente et al., 2018).
The spread of PWN within Europe is linked to insect vectors belonging to genus Monochamus, and more specifically M. galloprovincialis (Coleoptera: Cerambycidae). The pathogen is transmitted via these beetles from dead or dying trees to healthy ones through feeding (Mamiya and Enda, 1972) or oviposition (Edwards and Linit, 1992). Once the nematodes have entered the tree, they lodge in the resiniferous canals and feed on epithelial and parenchymal cells, triggering a host response which leads to disruption of water transport and rapid death by cavitation (Fukuda and Suzuki, 1988; Hara et al., 2006). Thus far, disease control mitigation techniques have been mainly focused on the search for genetic resistance in the host organisms and controlling disease spread by reducing populations of the insect vector. The first reported program for breeding genetic resistance to PWD started in western Japan in 1978, on Japanese black and red pine (Pinus thunbergii and P. densiflora, respectively) and obtained high post-inoculation survival rates in both (Fujimoto and Ohba, 1981; Toda and Kurinobu, 2002). Since then, many resistance programs have been operational, such as on P. massoniana (Liu et al., 2017; Zhu et al., 2021), P. pinaster (Gaspar et al., 2017; Carrasquinho et al., 2018) and P. radiata (Zas et al., 2015; Menéndez-Gutiérrez et al., 2018, 2021) in China, Portugal and Spain, respectively.
In terms of insect vector control, several strategies have been developed to reduce M. galloprovincialis populations such as a trapping method combining pheromone and kairomones (Álvarez-Baz et al., 2016; Galloprotect 2D®), the use of entomopathogenic fungi (Naves et al., 2008; Álvarez-Baz et al., 2015; Petersen-Silva et al., 2015) and the development of an auto-infection device based on the combined use of the attractant and the entomopathogenic fungi (Sacristán-Velasco et al., 2018). To date, only the use of traps with attractants has been implemented more widely, but successful field trials have been performed combining this system with the auto-infection device and powdered formulations of an entomopathogenic fungus (Martín-García, Unpublished data).
Despite progress in other areas, an effective integrated pest management technique for this disease will require nematode control. Progress toward achieving this goal has been made with the effect of fungi and bacteria on different PPNs being studied (Mankau, 1980; Askary, 2015; Abd-Elgawad and Askary, 2018; Migunova and Sasanelli, 2021). For PWN specifically, the most studied antagonistic fungal species is Esteya vermicola, which was first found infecting PWN in Taiwan in 1999 (Liou et al., 1999) and reported as a natural enemy of the nematode. Subsequent laboratory and field tests have corroborated this efficacy as a biological agent against the pathogen (Kubátová et al., 2000; Wang et al., 2008, 2009, 2018; Lin et al., 2013; Pires et al., 2022). Other fungal genera such as Verticillium spp. or Trichoderma spp. have also exhibited nematicidal effect on PWN populations (Maehara and Futai, 2000). More recently, some fungal species within the genera Leptographium and Graphilbum have also been shown to have a nemastatic effect on this pathogen (Vicente et al., 2022). However, little attention has been paid to potential nematicidal effects of the genus Beauveria spp. on PWN, despite significant mortality effect on other nematodes species being thoroughly documented (Youssef et al., 2020; Ye et al., 2021; Karabörklü et al., 2022) and its entomopathogenic effect on M. galloprovincialis is well known (Naves et al., 2008; Álvarez-Baz et al., 2015; Petersen-Silva et al., 2015). Maehara et al. (2007) proved the reduction of PWN transmission from vectors due to Beauveria bassiana was possible; however, this potential control method has not been examined in depth.
Most fungi’s nematicidal properties appear at least in part due to excreting secondary metabolites (mycotoxins) (Anke and Sterner, 1997, 2002; Li et al., 2007; Anke, 2011; Baazeem et al., 2021; Seong et al., 2021). However, some fungi (e.g., E. vermicola) appear to have alternate control mechanisms against the nematode, such as the emission of attractant volatiles similar to the host pine and subsequently the production of specific types of spores (lunate conidia) to trap the attracted nematodes (Kubátová et al., 2000; Lin et al., 2013). It is likely that the antagonistic effect of these fungi on the pathogen is the result of the combination of these two modes of action: nematode-trapping capacity and production of mycotoxins.
The nematicidal mycotoxin mechanism is probably the prime mode of action for the Beauveria genus and the most studied mycotoxin from this genus is beauvericin (BEA). Chemically, BEA is defined as a cyclic hexadepsipeptide consisting of alternating D-α-hydroxy-isovaleryl-(2-hydroxy-3-methylbutanoic acid) and amino acid units (Hamill et al., 1969; Figure 1) and is considered an emerging mycotoxin since it is still neither routinely determined, nor legislatively regulated (Jestoi, 2008; Vaclavikova et al., 2013; EFSA, 2014).
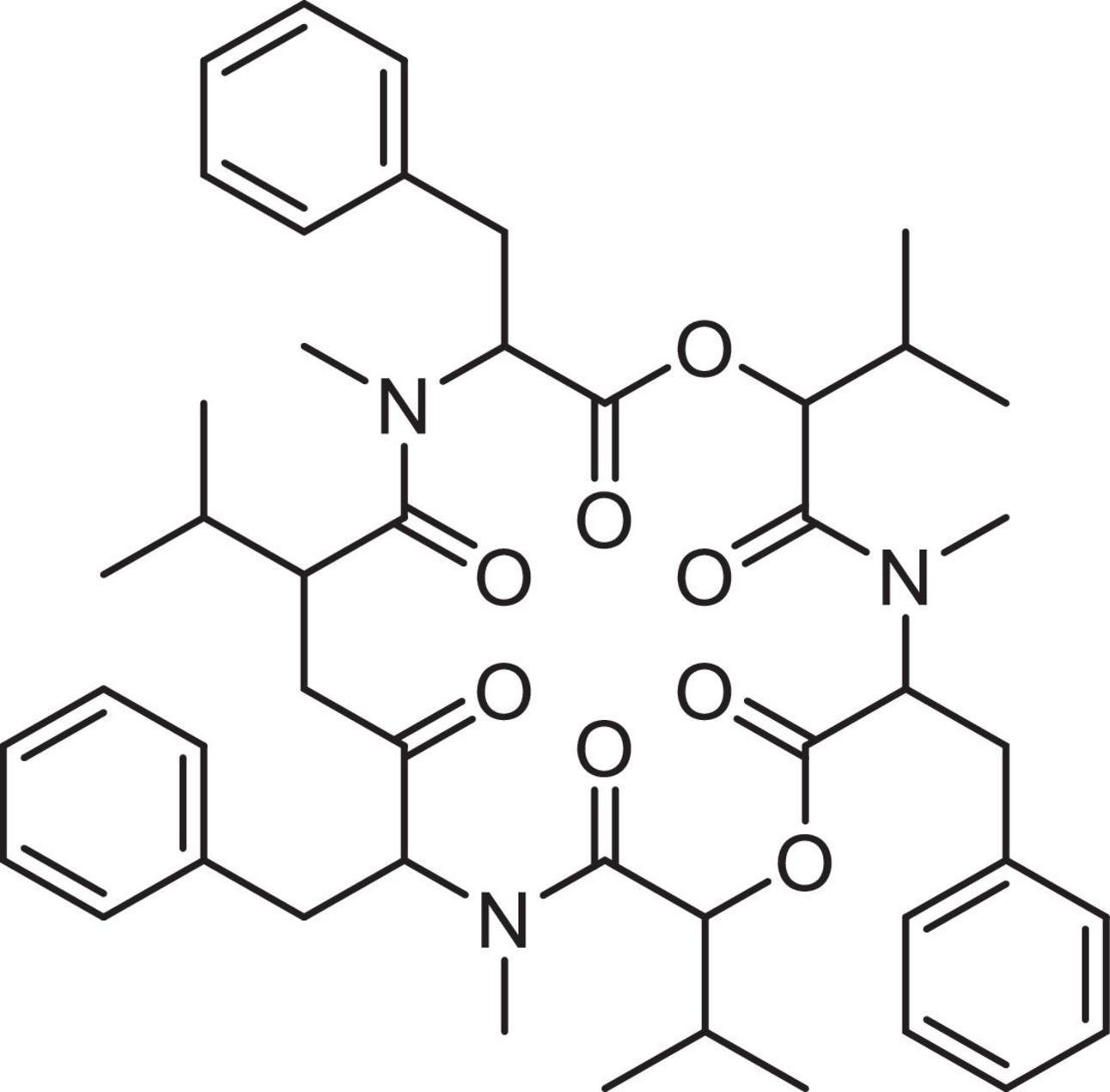
Figure 1. Chemical structure of BEA. Diagram created using the software ChemDraw®.
The compound is not only produced by Beauveria species such as B. bassiana (Hamill et al., 1969), but also by others, for example Fusarium spp (Gupta et al., 1991; Logrieco et al., 2002; Moretti et al., 2002, 2008). In comparison to B. bassiana, B. pseudobassiana has to date few studies identifying and assessing the toxins produced by this species. Berestetskiy et al. (2018) proved that B. pseudobassiana strain BCu22 produces different metabolites depending on the culture medium and demonstrated its insecticidal effect on grain aphid larvae. A subsequent study confirmed the existence of a gene cluster coding for BEA in the strain RGM 2184 of B. pseudobassiana (Altimira et al., 2022). However, none of them were able to corroborate the actual production of BEA by this fungal species. The toxicity of BEA was first tested on the crustacean Artemia salina (Hamill et al., 1969) but soon found to have insecticidal (Grove and Pople, 1980; Gupta et al., 1991; Ganassi et al., 2002; Fornelli et al., 2004; Leland et al., 2005) and nematicidal effects, tested first on Meloidogyne incognita (Mayer, 1995) and later on Caenorhabditis elegans and PWN (Shimada et al., 2010). Additionally, according to the Technical Commission on Contaminants in the Food Chain, acute exposure to BEA does not indicate concern for human health (EFSA, 2014). This fact, together with the nematicidal activity, makes BEA one of the most promising mycotoxins that could be implemented in integrated pest management of PWN.
This evidence suggests that Beauveria species and the mycotoxin BEA could be a potential control option against not only the PWN insect vector (M. galloprovincialis), but also against the nematode itself. The goals of the work presented here were firstly to demonstrate the nematicidal effect of different strains of B. bassiana and B. pseudobassiana on PWN populations, secondly to prove whether BEA has nematicidal effect on PWN and lastly to test whether not only B. bassiana is able to produce BEA, but also potentially B. pseudobassiana.
Two different strains of B. bassiana (EABps 11/01-Mg* and 95b) and one of B. pseudobassiana (MG-BU-17-001) were used for the assays.
*Note: EF1α and ITS classification of several strains of Beauveria species (B. bassiana and B. pseudobassiana) were modified in GenBank a few years ago (see accession numbers AY531938.1 and AY531931). So, the isolate EABps 11/01-Mg was wrongly identified by Álvarez-Baz et al. (2015) as B. pseudobassiana based on the homology of the isolates AY531938.1 and AY531931 (previously identified as B. pseudobassiana and currently B. bassiana). However, we have again analyzed the isolate EABps 11/01-Mg (EF1α and ITS) and checked in GenBank and it should be classified as B. bassiana.
Esteya vermicola CBS 115803 (supplied by the Czech Collection of Microorganisms), which has already demonstrated nematicidal effect on PWN (Wang et al., 2016; Pires et al., 2022) was used as negative-growth control. A non-sporulating Botrytis cinerea isolate, which is used to grow the PWN colonies for fostering its reproduction, was used as positive-growth control. All the isolates were cultivated using potato dextrose agar medium (PDA Scharlau, Spain).
The nematode strain was B. xylophilus CSF-N-1 and was grown using glass vials [28 mm (Ø), 43 mm (h), 25 mL (V)] with medium composed of hulled barley and non-sporulating B. cinerea (Supplementary Figure 1A). The vials were kept at room temperature with the cover ajar, always with oxygen flow inward, and under continuous darkness.
Thirty glass vials with 3 g hulled barley and 3 mL distilled water inside were autoclave sterilized and separated into 6 treatments (B. bassiana 95b, B. bassiana EABps 11/01-Mg, B. pseudobassiana MG-BU-17-001, B. cinerea, E. vermicola CCM 8247 and mock inoculated). A mycelium plug of each fungus was introduced into each vial (except for mock inoculated treatment, in which a plug of PDA was added) and left to grow for 13 days. After this growth period, a 100 μL aliquot with 270 nematodes (B. xylophilus CSF-N-1) was added to each vial and they were kept for a further 13 days at 25°C in continuous darkness.
The extraction of the nematodes from the vials was conducted according to a slightly modified version of the Baermann (1917) funnel technique. A tea filter (M-size, Finum®) was placed with the content of each vial on an autoclaved beaker (100 mL). The content was covered with autoclaved distilled water, so that the nematodes pass through the filter and remain in a clean aqueous suspension. After 24 h, they were washed with distilled water using a nylon sieve of 20 μm mesh size (NY-0073-Labopolis, Spain). The nematodes were retained by the sieve, washed with a washing bottle and finally resuspended into 55 mm (Ø) petri dishes. Three aliquots (100 μL) of each replicate were counted using a counting grid and binocular loupe to determine nematode concentration (Supplementary Figures 1B, C, D).
Considering the nematicidal effect of the metabolite beauvericin naturally extracted from strains of Fusarium bulbicola on PWN is already published (Shimada et al., 2010), we decided to test the effect of BEA commercial (Merck Life Science S.L.U, USA) on populations of B. xylophilus. Solutions of 0.1 mM, 1 mM and 2 mM of BEA with 5% dimethyl sulfoxide (DMSO) were prepared with eight replicates for each concentration by dividing each stock solution into eight 1.5 mL Eppendorf tubes. For the negative controls, another eight replicates were prepared with 5% DMSO only. A 75 μL nematode suspension (400 nm/mL) was added to each tube and live nematodes were then counted. A small hole was made in the top of each tube (with a heated entomological pin) to ensure oxygen flow, and the rack was covered with aluminum foil for continuous dark conditions. The assay was incubated at 25°C for 48 h, each tube was vortex mixed once per day to resuspend the nematodes. After 2 days, the contents of each tube were poured on a grid and the live nematodes counted.
Four replicates of each strain were grown on PDA plates and kept in an incubator at 28°C for 21 days. The PDA plates were subsequently cut into 2 cm cube pieces and all cubes from each PDA plate was placed in individual 150 mL conical flasks. MeOH (100 mL) was then added to each flask and the flasks sonicated for 15 min at 40°C. The organic extract was removed, and the solid media washed with MeOH (50 mL). The organic extracts were combined, filtered, and dried under vacuum. The subsequent oily extract was dissolved in 80% MeOH (7 mL), filtered again, and transferred to a sample vial. The organic solvent was removed under N2 and the aqueous mixture was frozen at −75°C before being dried using a freeze drier. All extract samples were weighed and dissolved in 90% MeOH in a 20 mg/mL concentration.
Samples were analyzed using an Agilent 1260 series HPLC system with attached 1200 series diode array detector and MSD-XT single quad mass spectrometer with attached atmospheric pressure chemical ionisation (APCI) source. The LC-MS used a Phenomenex Kinetex, 2.6 μm, C18, 75 mm × 2.1 mm column with a three solvent system A: H2O, B: acetonitrile and C: 2% formic acid in acetonitrile. The solvent method had a constant flow rate of 0.4 ml/min and started with 90/5/5 initial mix which increases to 45/50/5 over 1 min, then rises again to 5/90/5 over 7 min (8 min total) and held for a further 1 min. The system was allowed re-equilibrate to starting conditions before the next sample was run. BEA was identified in samples using retention time, UV spectra and mass spectrometry. BEA was quantified using UV absorbance (190–210 nm), with a calibration curve obtained using a beauvericin standard purchased from Cayman Chemical Company (concentrations 0.2, 0.1, 0.05 and 0.025 mg/mL).
One-way analysis of variance (ANOVA) and multiple comparison procedures were performed to test the effects of fungi and commercial BEA on nematode populations and to test the concentrations of BEA extracted from each Beauveria strain by HPLC. The ANOVA assumptions (normality and homogeneity of variances) were tested in each analysis by the Shapiro and Bartlett tests. When neither assumption was violated (concentrations of BEA extracted from each Beauveria strain), classical one-way ANOVA and Tukey’s honestly significant difference (HSD) tests was applied. When at least one of these assumptions was violated (effects of fungi and commercial BEA on nematode populations) robust statistical methods were applied (García-Pérez, 2010). In particular, heteroscedastic one-way ANOVAs were performed using the generalized Welch procedure and a 0.1 trimmed mean transformation. These analyses were carried out using the Wilcox’ robust statistics (WRS2) package implemented in the R software environment (R Core Team, 2021).
Growth of B. xylophilus populations were strongly inhibited by the presence of Beauveria species (F = 28.29, p < 0.001). All three Beauveria strains showed a clear nematicidal effect on B. xylophilus populations, with no significant differences between the treatments (Figure 2). The nemastatic or nematicidal effect of B. bassiana has already demonstrated on other nematode species such as M. incognita (Kepenekci et al., 2017; Youssef et al., 2020; Karabörklü et al., 2022) and C. elegans (Ye et al., 2021). Thus, our results are congruent with previous evidence and confirm the potential biocontrol agent role of this fungus on B. xylophilus. To our knowledge, this is the first study that demonstrates the nematicidal activity of B. pseudobassiana. However, considering the phenotypic proximity of both Beauveria species (Rehner et al., 2011; Berestetskiy et al., 2018), this is not an unexpected result.
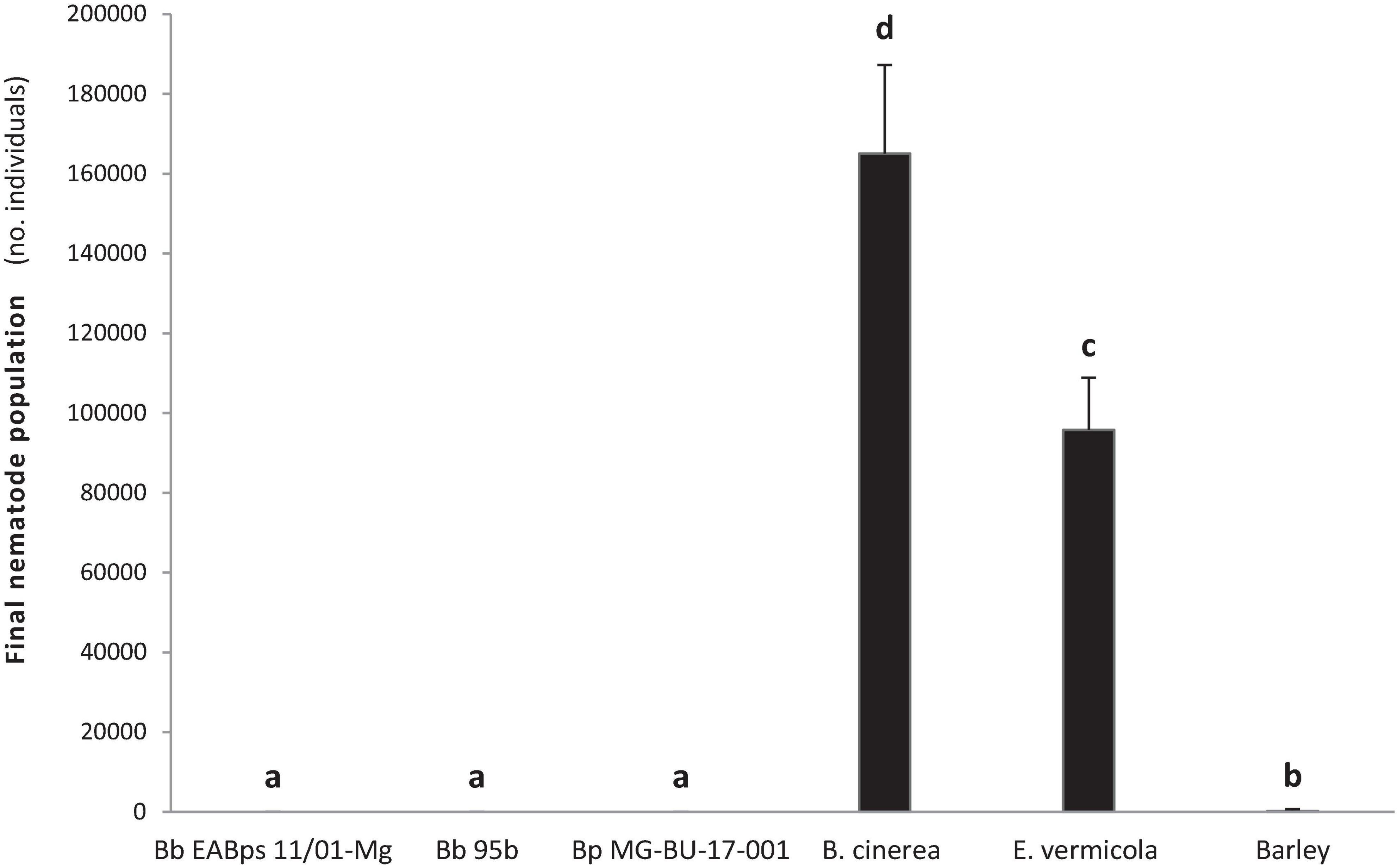
Figure 2. Effect on B. xylophilus populations from B. bassiana and B. pseudobassiana strains after 13 days. Bars with different letters indicate significantly different means (α = 0.05).
Growing methods for B. xylophilus are not standardized, but cultivation with B. cinerea (a non-sporulating isolate) grown on barley is the most widely used (Aikawa and Kikuchi, 2007; Espada et al., 2016; Pimentel et al., 2020) and the one recommended by EPPO (2013). Thus, as predicted, the nematode population in the presence of B. cinerea exponentially increased during the 13-day assay, obtaining a final population of approximately 165,000 individuals (Figure 2), corroborating its role as a positive-growth control. In contrast, the results obtained from the negative-growth control E. vermicola were unexpected. The final population (≈ 96,000 nematodes) from E. vermicola is much larger than obtained in the barley base-growth control (around 2,700 individuals) (Figure 2). The nematophagous effect of this fungus had been previously demonstrated, although this result could be due to the lack of lunate adhesive conidia detected in this study. Previous studies had linked the nematophagous effect to the production of this specific type of conidia (Liou et al., 1999; Kubátová et al., 2000; Wang et al., 2008, 2009, 2018; Lin et al., 2013; Pires et al., 2022).
The results obtained show a clear nematicidal effect of beauvericin on populations of B. xylophilus. While the control treatment had a survival rate (after 48 h) of almost 60%, the BEA treatments survival rates were significantly lower (F = 18.26, p < 0.001). The 0.1 mM BEA treatment attained a survival rate of 35.1%, and 1 mM and 2 mM BEA treatments resulted in 23.5 and 20.8% survival rates, respectively, with no significant differences between them (Figure 3). Compared to control treatment (i.e., deducting the natural mortality), the mortality rates of each BEA treatment were 39.2, 59.2 and 63.9% (0.1, 1, and 2 mM, respectively), demonstrating the nematicidal activity of BEA.
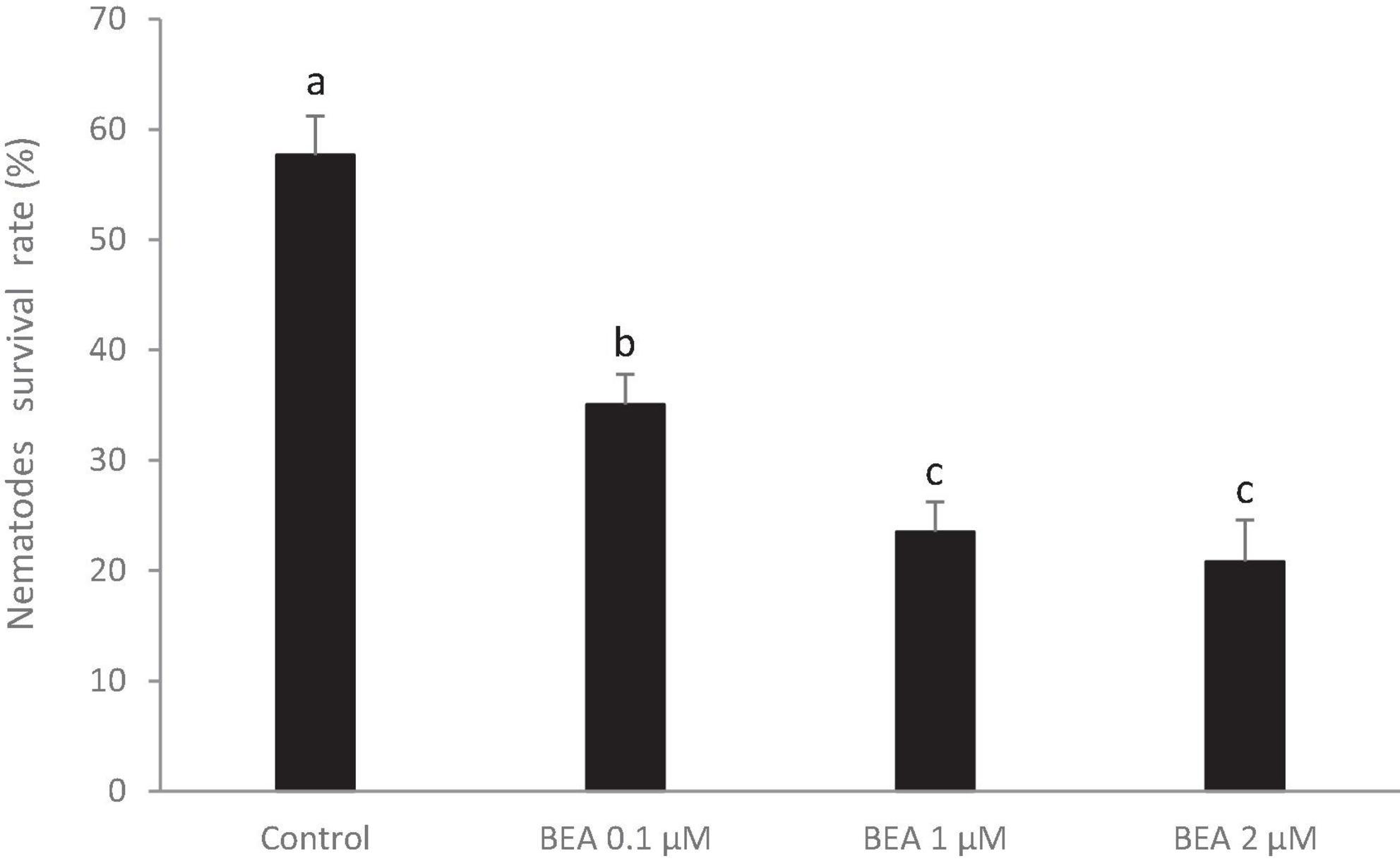
Figure 3. Survival rate of B. xylophilus in control (DMSO 5%) and treatments (BEA + DMSO 5%). Bars with different letters indicate significantly different means (α = 0.05).
The anthelmintic capacity of numerous compounds chemically close to BEA (cyclic-depsipeptides) have been quantified on many occasions (Scherkenbeck et al., 2002; Jeschke et al., 2005; Firakova et al., 2007; Prosperini et al., 2017). More specifically, the disruptive effect of BEA on the model nematode C. elegans (Büchter et al., 2020) and on B. xylophilus (Shimada et al., 2010) has been confirmed. Comparing the results of beauvericin obtained from F. bulbicola (Shimada et al., 2010) with the commercial one used in this study, both exhibited similar linear trends: with higher concentrations of BEA resulting in higher mortality, although this effect seems to stabilize when the concentration approaches 2 mM. In the present study even the lowest BEA treatment (0.1 mM) showed a high nematode mortality (≈ 23% higher than mock treatment), however, Shimada et al. (2010) found only weak nematicidal activity using the same concentration. Of the concentrations of BEA tested, it appears the most suitable one for field application is 1 mM BEA, since it retained a similar nematicidal effect when compared with the 2 mM solution, but with lower product expenditure.
The mechanism of action of BEA on animal cells is still not completely understood, but seems linked to its ionophoric activity, which increases ion permeability in biological membranes and the consequent oxidative stress at molecular level (Mallebrera et al., 2018). In our trials, a brownish “encapsulation” was observed during the final visual assessment of BEA treatments individuals (Figure 4) which could be related to the combination of BEA molecules to the lipids of the nematode epicuticle.
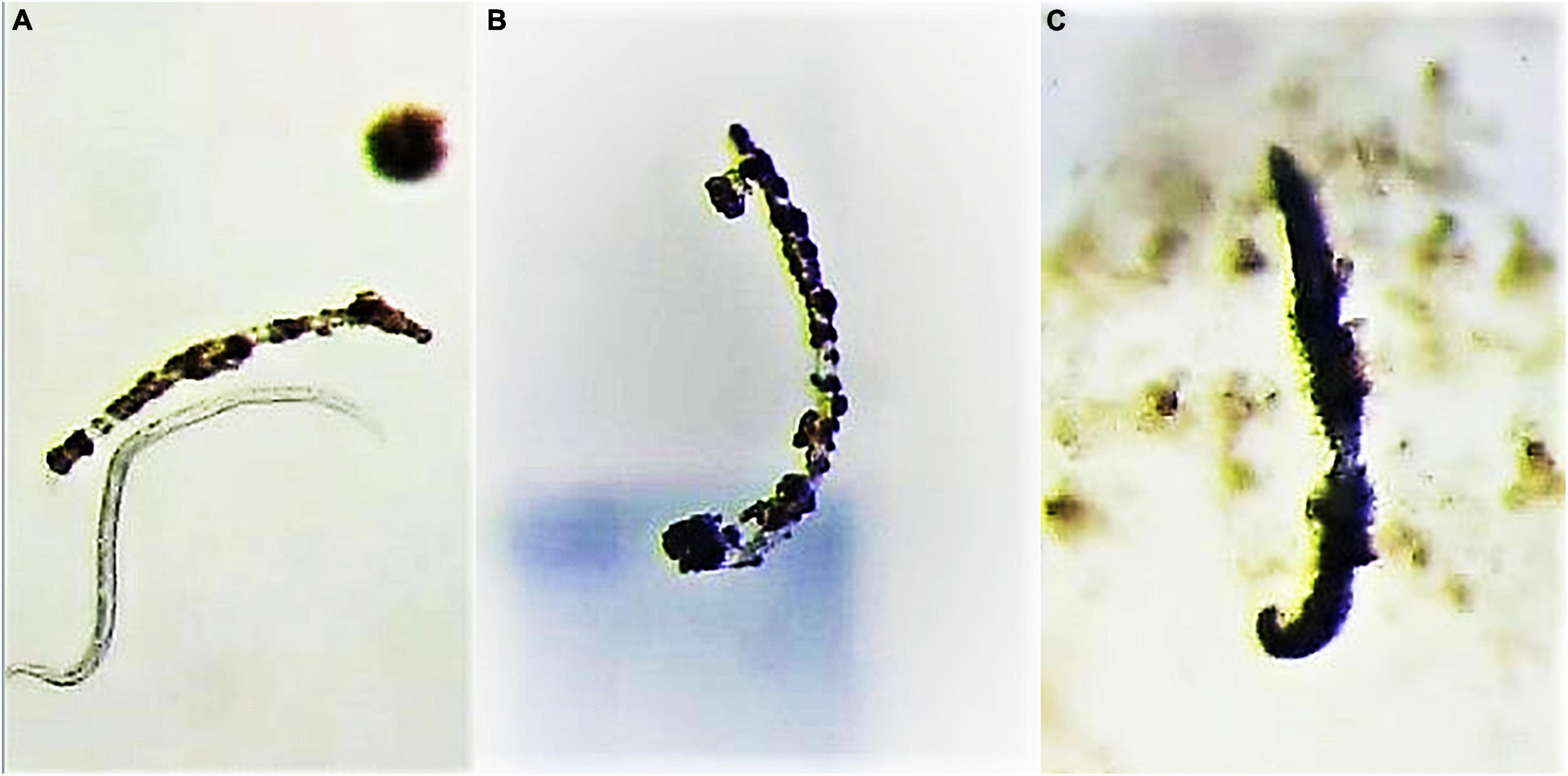
Figure 4. A nematode with a normal appearance together with a “encapsulated” one in 0.1 mM BEA (A) and other “encapsulated” nematodes found in 1 and 2 mM BEA treatments [(B,C), respectively].
Beauvericin (BEA) was positively identified in all three strains using a mixture of UV, MS and HPLC column retention. Shown in Figure 5, is chromatograms of the BEA standard and B. pseudobassiana MG-BU-17-001, in both UV (200 nm) and single ion mode (SIM) MS (m/z = 784.3). BEA was assigned as a single peak [retention time (RT) = 6.85 min (UV) and 6.86 min (MS)] which was found in all experiments. This assignment was further confirmed by comparing the MS spectra (m/z = 100–1,050) of the BEA standard with all three strains showing we believe, for the first time the presence of BEA in B. pseudobassiana (Figure 6).
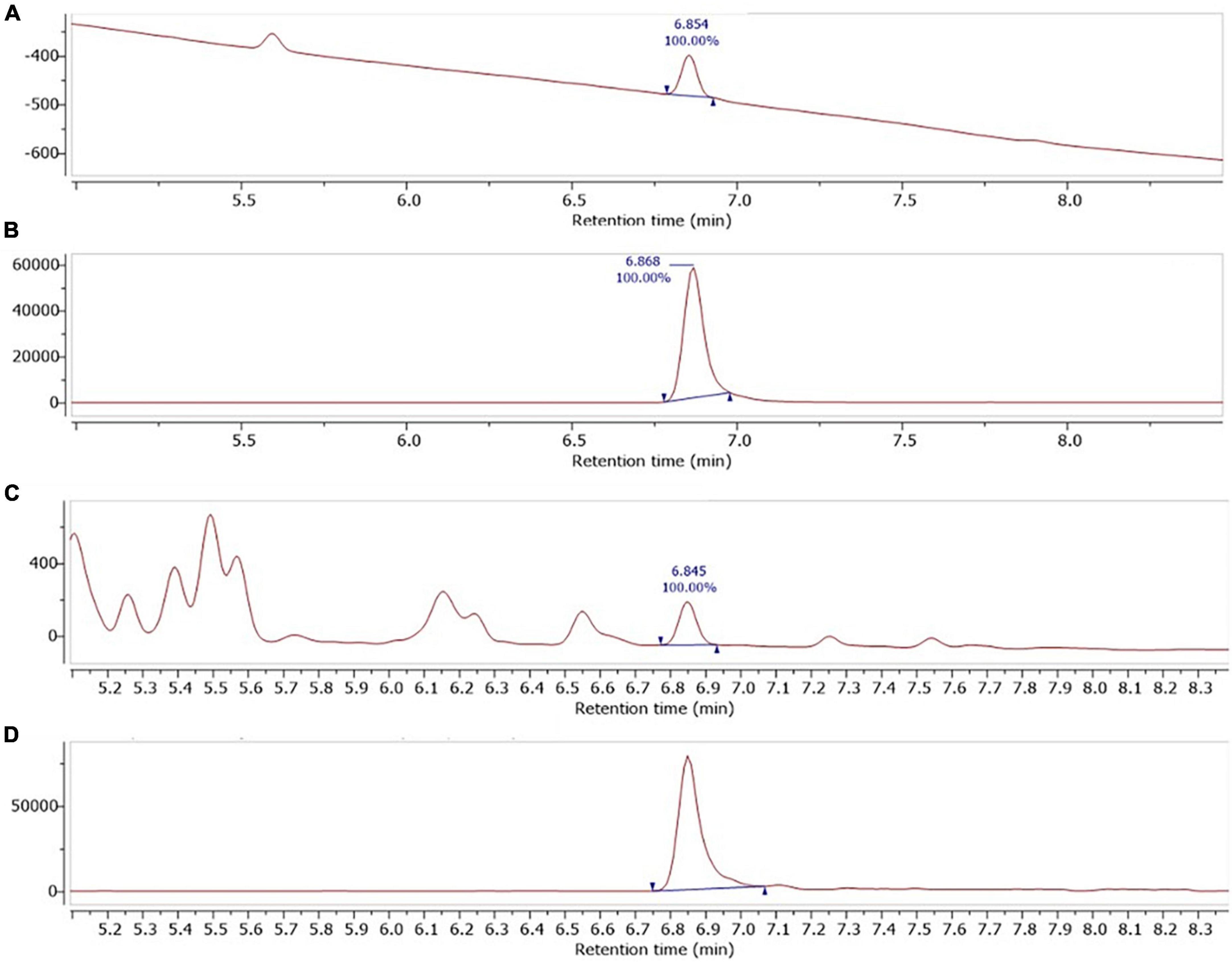
Figure 5. UV diode array detector (DAD) (200 ± 10 nm) and single ion mode (SIM) (784.3) atmospheric pressure chemical ionisation (APCI) chromatograms of, (A,B) BEA standard (10 μg/mL), and (C,D) B. pseudobassiana MG-BU-17-001.

Figure 6. APCI MS spectra for (A) BEA standard (10 μg/mL), and (B) B. pseudobassiana “MG-BU-17-001.”
All three strains tested produced BEA, but significant differences were found between them (F = 10.06, p = 0.02). B. bassiana EABps 11/01-Mg exhibits the highest levels of BEA (151.4 μg/mL), followed by B. bassiana 95b (119.3 μg/mL) and lastly B. pseudobassiana MG-BU-17-001 (56.7 μg/mL) (Figure 7). While previous studies have confirmed the production of BEA by B. bassiana (Hamill et al., 1969; Valencia et al., 2011; Berestetskiy et al., 2018), this assay showed that the B. bassiana strains tested here produced a higher amount of beauvericin than other strains tested by Valencia et al. (2011), which should lead to stronger nematicidal and insecticidal effects. Moreover, to our knowledge, this is the first study that identified and quantified this mycotoxin in extracts of B. pseudobassiana. This finding opens the possibility of using both Beauveria species as potential antagonistic organisms against B. xylophilus. Further studies testing more strains of both species are needed to elucidate whether the differences found in this study are related to intraspecific or interspecific variability. Beyond the field of biopesticides, this outcome has parallel relevance for nutrition and human health, since BEA is classified as an emerging mycotoxin (Jestoi, 2008; Vaclavikova et al., 2013; EFSA, 2014) with high occurrence in food commodities (Al Khoury et al., 2021). It is therefore vital for food monitoring chains to have as much information as possible on all fungal species able to produce BEA.
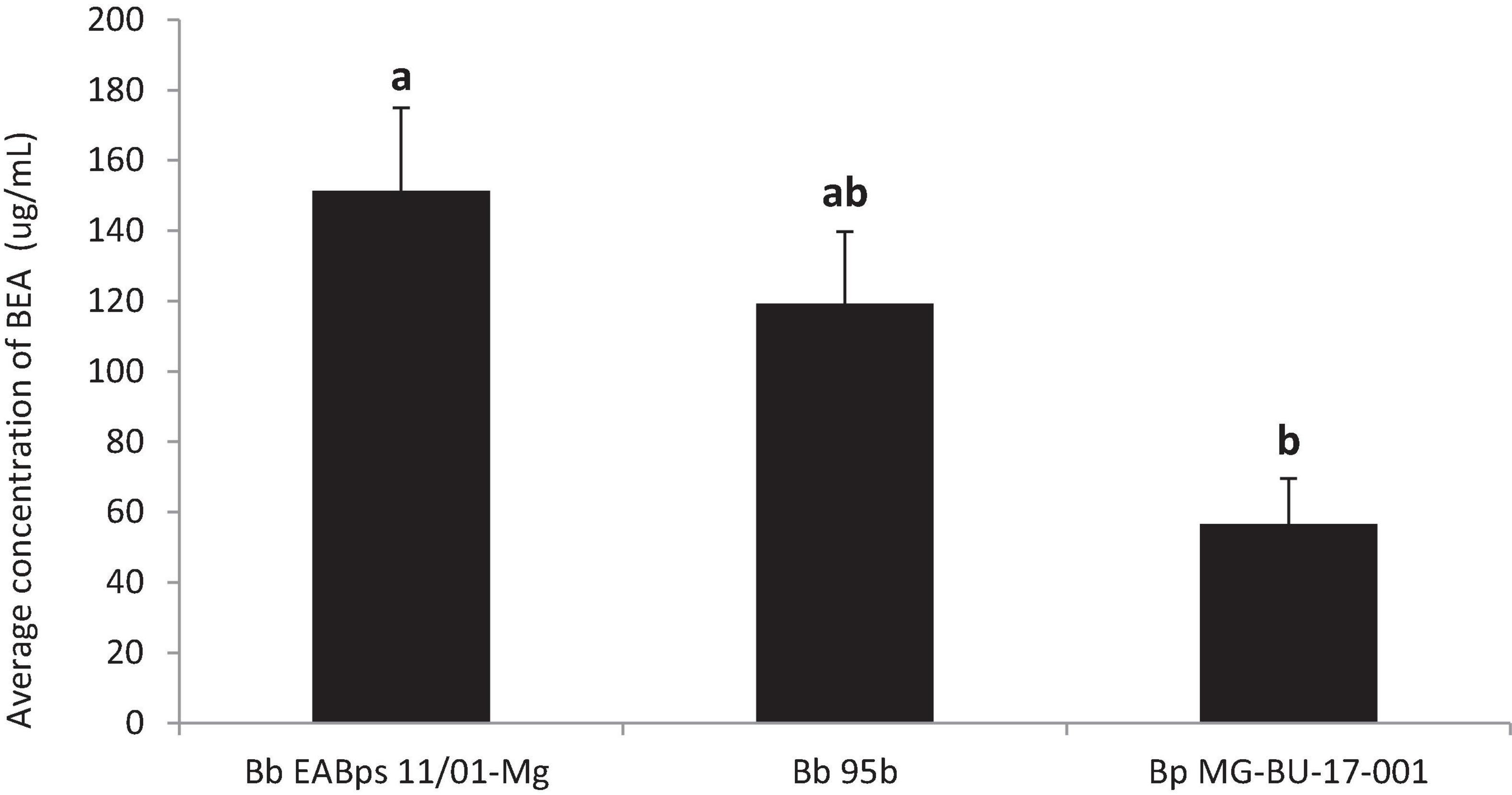
Figure 7. Average concentration of BEA (μg/mL) in extracts obtained from the three strains of Beauveria spp. Bars with different letters indicate significantly different means (α = 0.05).
Taking into account the high risk of spread of B. xylophilus and the serious threat that this species of nematode poses to European pines, the development of an effective tool for controlling the pine wilt disease is essential. Different antagonistic fungi could exert biological control against the nematode or the beetle vector, although ideally a single fungus would be used to control both. This study found that Beauveria species, which had already demonstrated its entomopathogenic activity against M. galloprovincialis (Álvarez-Baz et al., 2015) also exhibited nematicidal activity against B. xylophilus. This finding paves the way for new research into the potential of these fungal strains to be tested in a self-infection device for PWN population control (Sacristán-Velasco et al., 2018) and their subsequent implementation in field conditions.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The manuscript presents research on animals that do not require ethical approval for their study.
JM-G, TS-G, and PZ conceived and designed the experiment. TS-G, PZ, MB, and SH performed the experiments. JM-G analyzed the data. TS-G and JM-G wrote the manuscript. All authors contributed to the article and approved the submitted version.
This publication was supported and financed by the BBVA Foundation (Becas Leonardo a Investigadores y Creadores Culturales 2020, Grant No. ING_0140). The Foundation takes no responsibility for the opinions, statements and contents of this project, which are entirely the responsibility of its authors. JN-S was also supported by the European Union’s Horizon Europe research and innovation programme under the Marie Sklodowska-Curie Actions (MSCA) agreement No. 101068728. This study was also part of the research project “Estrategias para la contención de la enfermedad del marchitamiento del pino, causada por Bursaphelenchus xylophilus: resistencia del hospedante y manejo del insecto vector (NEMOPLAN)” (RTA2017-00012-C02-02) supported by Ministerio de Ciencia, Innovación y Universidades.
We thank to Masaryk University (Brno, Czech Republic) for kindly providing the strain Esteya vermicola CCM 8247. We would also like to thank CIF Lourizán, especially to Dra. Raquel Díaz Vázquez for her collaboration and support in the trials related to the nematode.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor IM is currently organizing a research topic with the author JD.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2023.1229456/full#supplementary-material
Abd-Elgawad, M. M., and Askary, T. H. (2018). Fungal and bacterial nematicides in integrated nematode management strategies. Egypt. J. Biol. Pest Control 28, 1–24. doi: 10.1186/s41938-018-0080-x
Aikawa, T., and Kikuchi, T. (2007). Estimation of virulence of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) based on its reproductive ability. Nematology 9, 371–377. doi: 10.1163/156854107781352007
Al Khoury, C. A., Nemer, N., Tokajian, S., and Nemer, G. (2021). Beauvericin: The journey of a pesticide into a humanized drug. Preprints [Preprint]. doi: 10.20944/preprints202107.0506.v1
Altimira, F., Arias-Aravena, M., Jian, L., Real, N., Correa, P., González, C., et al. (2022). Genomic and experimental analysis of the insecticidal factors secreted by the entomopathogenic fungus Beauveria pseudobassiana RGM 2184. J. Fungi 8:253. doi: 10.3390/jof8030253
Álvarez-Baz, G., Fernández-Bravo, M., Pajares, J., and Quesada-Moraga, E. (2015). Potential of native Beauveria pseudobassiana strain for biological control of pine wood nematode vector Monochamus galloprovincialis. J. Invertebr. Pathol. 132, 48–56.
Álvarez-Baz, G., Gallego, D., Hall, D. R., Jactel, H., and Pajares, J. A. (2016). Combining pheromone and kairomones for effective trapping of the pine sawyer beetle Monochamus galloprovincialis. J. Appl. Entomol. 140, 58–71. doi: 10.1111/jen.12297
Anke, H. (2011). “Insecticidal and nematicidal metabolites from fungi,” in Industrial applications the mycota, Vol 10, ed. M. Hofrichter (Berlin: Springer), 151–163. doi: 10.1007/978-3-642-11458-8_7
Anke, H., and Sterner, O. (1997). Nematicidal metabolites from higher fungi. Curr. Organ. Chem. 1, 361–374.
Anke, H., and Sterner, O. (2002). “Insecticidal and nematicidal metabolites from fungi,” in Industrial applications the mycota, ed. H. D. Osiewacz (Berlin: Springer), 109–127. doi: 10.1007/978-3-662-10378-4_6
Askary, T. H. (2015). “Nematophagous fungi as biocontrol agents of phytonematodes,” in Biocontrol agents of phytonematodes, eds T. Askary and P. Martinelli (Wallingford: CAB International), 81–125. doi: 10.1079/9781780643755.0081
Baazeem, A., Almanea, A., Manikandan, P., Alorabi, M., Vijayaraghavan, P., and Abdel-Hadi, A. (2021). In vitro antibacterial, antifungal, nematicidal and growth promoting activities of Trichoderma hamatum FB10 and its secondary metabolites. J. Fungi 7:331. doi: 10.3390/jof7050331
Baermann, G. (1917). “A simple method for the detection of Ankylostomum (nematode) larvae in soil tests,” in Mededelingen uit het geneeskundig laboratorium te weltevreden, ed. G. Baermann (Batavia: Javasche Boekhandel & Drukkerij), 41–47.
Berestetskiy, A. O., Ivanova, A. N., Petrova, M. O., Prokof’eva, D. S., Stepanycheva, E. A., Uspanov, A. M., et al. (2018). Comparative analysis of the biological activity and chromatographic profiles of the extracts of Beauveria bassiana and B. pseudobassiana cultures grown on different nutrient substrates. Microbiology 87, 200–214. doi: 10.1134/S0026261718020030
Büchter, C., Koch, K., Freyer, M., Baier, S., Saier, C., Honnen, S., et al. (2020). The mycotoxin beauvericin impairs development, fertility and life span in the nematode Caenorhabditis elegans accompanied by increased germ cell apoptosis and lipofuscin accumulation. Toxicol. Lett. 334, 102–109. doi: 10.1016/j.toxlet.2020.09.016
Carrasquinho, I., Lisboa, A., Inácio, M. L., and Gonçalves, E. (2018). Genetic variation in susceptibility to pine wilt disease of maritime pine (Pinus pinaster Aiton) half-sib families. Ann. For. Sci. 75:85. doi: 10.1007/s13595-018-0759-x
De la Fuente, B., Saura, S., and Beck, P. S. (2018). Predicting the spread of an invasive tree pest: The pine wood nematode in Southern Europe. J. Appl. Ecol. 55, 2374–2385. doi: 10.1111/1365-2664.13177
Edwards, O. R., and Linit, M. J. (1992). Transmission of Bursaphelenchus xylophilus through oviposition wounds of Monochamus carolinensis (Coleoptera: Cerambycidae). J. Nematol. 24:133.
EFSA (2014). Scientific opinion on the risks to human and animal health related to the presence of beauvericin and enniatins in food and feed. EFSA J. 12:3802. doi: 10.2903/j.efsa.2014.3802
EPPO (2013). Diagnostics. PM 7/4 (3) Bursaphelenchus xylophilus. EPPO Bull. 43, 105–118. doi: 10.1111/epp.12024
EPPO (2016). EPPO A1 and A2 Lists of pest recommended for regulation as quarantine pests. Paris: European and Mediterranean Plant Protection Organization.
Espada, M., Silva, A. C., Eves van den Akker, S., Cock, P. J., Mota, M., and Jones, J. T. (2016). Identification and characterization of parasitism genes from the pinewood nematode Bursaphelenchus xylophilus reveals a multilayered detoxification strategy. Mol. Plant Pathol. 17, 286–295. doi: 10.1111/mpp.12280
Firakova, S., Proksa, B., and Šturdíková, M. (2007). Biosynthesis and biological activity of enniatins. Die Pharm. Int. J. Pharmaceut. Sci. 62, 563–568. doi: 10.1691/ph.2007.8.7600
Fornelli, F., Minervini, F., and Logrieco, A. (2004). Cytotoxicity of fungal metabolites to lepidopteran (Spodoptera frugiperda) cell line (SF-9). J. Invertebr. Pathol. 85, 74–79. doi: 10.1016/j.jip.2004.01.002
Fujimoto, Y., and Ohba, K. (1981). “The first year results of the breeding of Japanese pines for resistance to the wood nematode,” in Proceeding of the XVII IUFRO World Congress, (Kyoto), 287–291.
Fukuda, K., and Suzuki, K. (1988). Changes of water relation parameters in pine-wood nematode-infested Japanese red pine. J. Jap. For. Soc. 70, 390–394. doi: 10.11519/jjfs1953.70.9_390
Ganassi, S., Moretti, A., Pagliai, A. M. B., Logrieco, A., and Sabatini, M. A. (2002). Effects of beauvericin on Schizaphis graminum (Aphididae). J. Invertebr. Pathol. 80, 90–96. doi: 10.1016/S0022-2011(02)00125-8
García-Pérez, A. (2010). Métodos avanzados de estadística aplicada. métodos robustos y de remuestreo. Madrid: UNED Universidad Nacional a Distancia.
Gaspar, D., Trindade, C., Usié, A., Meireles, B., Barbosa, P., Fortes, A. M., et al. (2017). Expression profiling in Pinus pinaster in response to infection with the pine wood nematode Bursaphelenchus xylophilus. Forests 8:279. doi: 10.3390/f8080279
Grove, J. F., and Pople, M. (1980). The insecticidal activity of beauvericin and the enniatin complex. Mycopathologia 70, 103–105. doi: 10.1007/BF00443075
Gupta, S., Krasnoff, S. B., Underwood, N. L., Renwick, J. A. A., and Roberts, D. W. (1991). Isolation of beauvericin as an insect toxin from Fusarium semitectum and Fusarium moniliforme var. subglutinans. Mycopathologia 115, 185–189. doi: 10.1007/BF00462223
Hamill, R. L., Higgens, C. E., Boaz, H. E., and Gorman, M. (1969). The structure of beauvericin, a new depsipeptide antibiotic toxic to Artemia salina. Tetrahedron Lett. 10, 4255–4258. doi: 10.1016/S0040-4039(01)88668-8
Hara, N., Takeuchi, Y., and Futai, K. (2006). Cytological changes in ray parenchyma cells of seedlings of three pine species infected with the pine wilt disease. Jap. J. Nematol. 36, 23–32. doi: 10.3725/jjn.36.23
Hirata, A., Nakamura, K., Nakao, K., Kominami, Y., Tanaka, N., Ohashi, H., et al. (2017). Potential distribution of pine wilt disease under future climate change scenarios. PLoS One 12:e0182837. doi: 10.1371/journal.pone.0182837
Jeschke, P., Harder, A., Schindler, M., and Etzel, W. (2005). Cyclohexadepsipeptides (CHDPs) with improved anthelmintical efficacy against the gastrointestinal nematode (Haemonchus contortus) in sheep. Parasitol. Res. 97, S17–S21. doi: 10.1007/s00436-005-1440-5
Jestoi, M. (2008). Emerging Fusarium-mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin—A review. Crit. Rev. Food Sci. Nutr. 48, 21–49. doi: 10.1080/10408390601062021
Karabörklü, S., Aydinli, V., and Dura, O. (2022). The potential of Beauveria bassiana and Metarhizium anisopliae in controlling the root-knot nematode Meloidogyne incognita in tomato and cucumber. J. Asia Pac. Entomol. 25:101846. doi: 10.1016/j.aspen.2021.101846
Kepenekci, I., Saglam, H. D., Oksal, E., Yanar, D., and Yanar, Y. (2017). Nematicidal activity of Beauveria bassiana (Bals.-Criv.) Vuill. against root-knot nematodes on tomato grown under natural conditions. Egypt. J. Biol. Pest Control 27:117.
Kim, B. N., Kim, J. H., Ahn, J. Y., Kim, S., Cho, B. K., Kim, Y. H., et al. (2020). A short review of the pinewood nematode, Bursaphelenchus xylophilus. Toxicol. Environ. Health Sci. 12, 297–304. doi: 10.1007/s13530-020-00068-0
Kubátová, A., Novotný, D., Prášil, K., and Mráček, Z. (2000). The nematophagous hyphomycete Esteya vermicola found in the Czech Republic. Czech Mycol. 52, 227–235. doi: 10.33585/cmy.52305
Leland, J. E., McGuire, M. R., Grace, J. A., Jaronski, S. T., Ulloa, M., Park, Y. H., et al. (2005). Strain selection of a fungal entomopathogen, Beauveria bassiana, for control of plant bugs (Lygus spp.) (Heteroptera: Miridae). Biol. Control 35, 104–114. doi: 10.1016/j.biocontrol.2005.06.005
Li, G., Zhang, K., Xu, J., Dong, J., and Liu, Y. (2007). Nematicidal substances from fungi. Recent Patents Biotechnol. 1, 212–233. doi: 10.2174/187220807782330165
Lin, F., Ye, J., Wang, H., Zhang, A., and Zhao, B. (2013). Host deception: Predaceous fungus, Esteya vermicola, entices pine wood nematode by mimicking the scent of pine tree for nutrient. PLoS One 8:e71676. doi: 10.1371/journal.pone.0071676
Liou, J. Y., Shih, J. Y., and Tzean, S. S. (1999). Esteya, a new nematophagous genus from Taiwan, attacking the pinewood nematode (Bursaphelenchus xylophilus). Mycol. Res. 103, 242–248. doi: 10.1017/S0953756298006984
Liu, Q., Wei, Y., Xu, L., Hao, Y., Chen, X., and Zhou, Z. (2017). Transcriptomic profiling reveals differentially expressed genes associated with pine wood nematode resistance in masson pine (Pinus massoniana Lamb.). Sci. Rep. 7:4693. doi: 10.1038/s41598-017-04944-7
Logrieco, A., Rizzo, A., Ferracane, R., and Ritieni, A. (2002). Occurrence of beauvericin and enniatins in wheat affected by Fusarium avenaceum head blight. Appl. Environ. Microbiol. 68, 82–85. doi: 10.1128/AEM.68.1.82-85.2002
Maehara, N., and Futai, K. (2000). Population changes of the pinewood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), on fungi growing in pine-branch segments. Appl. Entomol. Zool. 35, 413–417. doi: 10.1303/aez.2000.413
Maehara, N., He, X., and Shimazu, M. (2007). Maturation feeding and transmission of Bursaphelenchus xylophilus (Nematoda: Parasitaphelenchidae) by Monochamus alternatus (Coleoptera: Cerambycidae) inoculated with Beauveria bassiana (Deuteromycotina: Hyphomycetes). J. Econ. Entomol. 100, 49–53. doi: 10.1093/jee/100.1.49
Mallebrera, B., Prosperini, A., Font, G., and Ruiz, M. J. (2018). In vitro mechanisms of Beauvericin toxicity: A review. Food Chem. Toxicol. 111, 537–545. doi: 10.1016/j.fct.2017.11.019
Mamiya, Y., and Enda, N. (1972). Transmission of Bursaphelenchus lignicolus (Nematoda: Aphelenchoididae) by Monochamus alternatus (Coleoptera: Cerambycidae). Nematologica 18, 159–162. doi: 10.1163/187529272X00395
Mayer, A. (1995). Bekämpfung von pflanzenparasitären Nematoden der Gattung Meloidogyne mit Pilzen und deren Toxinen. Ph.D thesis. Kaiserslautern: University of Kaiserslautern.
Menéndez-Gutiérrez, M., Alonso, M., and Díaz, R. (2021). Assessing genetic variation in resistance to Pinewood Nematode (Bursaphelenchus xylophilus) in Pinus radiata D. Don Half-Sib Families. Forests 12:1474. doi: 10.3390/f12111474
Menéndez-Gutiérrez, M., Alonso, M., Toval, G., and Díaz, R. (2018). Testing of selected Pinus pinaster half-sib families for tolerance to pinewood nematode (Bursaphelenchus xylophilus). Forestry 91, 38–48. doi: 10.1093/forestry/cpx030
Migunova, V. D., and Sasanelli, N. (2021). Bacteria as biocontrol tool against phytoparasitic nematodes. Plants 10:389. doi: 10.3390/plants10020389
Moretti, A., Belisario, A., Tafuri, A., Ritieni, A., Corazza, L., and Logrieco, A. (2002). “Production of beauvericin by different races of Fusarium oxysporum f. sp. melonis, the Fusarium wilt agent of muskmelon,” in Mycotoxins in plant disease, eds A. Logrieco, J. A. Bailey, L. Corazza, and B. M. Cooke (Dordrecht: Springer), 661–666. doi: 10.1007/978-94-010-0001-7_8
Moretti, A., Mulé, G., Ritieni, A., Láday, M., Stubnya, V., Hornok, L., et al. (2008). Cryptic subspecies and beauvericin production by Fusarium subglutinans from Europe. Int. J. Food Microbiol. 127, 312–315. doi: 10.1016/j.ijfoodmicro.2008.08.003
Mota, M., Burgermeister, W., Braasch, H., Sousa, E., Penas, A. C., Metge, K., et al. (1999). First report of Bursaphelenchus xylophilus in portugal and in Europe. Nematology 1, 727–734. doi: 10.1163/156854199508757
Naves, P. M., Sousa, E., and Rodrigues, J. M. (2008). Biology of Monochamus galloprovincialis (Coleoptera, Cerambycidae) in the pine wilt disease affected zone, Southern Portugal. Silva Lusitana 16, 133–148.
Petersen-Silva, R., Inácio, L., Henriques, J., Naves, P., Sousa, E., and Pujade-Villar, J. (2015). Susceptibility of larvae and adults of Monochamus galloprovincialis to entomopathogenic fungi under controlled conditions. Int. J. Pest Manag. 61, 106–112. doi: 10.1080/09670874.2015.1016472
Pimentel, C. S., Firmino, P. N., and Ayres, M. P. (2020). Comparison of methods to obtain and maintain cultures of the pinewood nematode, Bursaphelenchus xylophilus. J. For. Res. 25, 101–107. doi: 10.1080/13416979.2020.1745989
Pires, D., Vicente, C. S., Inácio, M. L., and Mota, M. (2022). The potential of Esteya spp. for the biocontrol of the Pinewood Nematode, Bursaphelenchus xylophilus. Microorganisms 10:168. doi: 10.3390/microorganisms10010168
Prosperini, A., Berrada, H., Ruiz, M. J., Caloni, F., Coccini, T., Spicer, L. J., et al. (2017). A review of the mycotoxin enniatin B. Front. Public Health 5:304. doi: 10.3389/fpubh.2017.00304
R Core Team (2021). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Rehner, S. A., Minnis, A. M., Sung, G. H., Luangsa-ard, J. J., Devotto, L., and Humber, R. A. (2011). Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 103, 1055–1073. doi: 10.3852/10-302
Sacristán-Velasco, A., Bravo, M. D. C. F., Moraga, E. Q., and Alonso, J. A. P. (2018). Control biológico del vector del nematodo de la madera del pino Monochamus galloprovincialis Olivier mediante autoinfección con el hongo entomopatógeno Beauveria pseudobassiana SA Rehner & Humber. Cuadernos Soc. Española Ciencias For. 44, 147–168.
Scherkenbeck, J., Jeschke, P., and Harder, A. (2002). PF1022A and related cyclodepsipeptides-a novel class of anthelmintics. Curr. Top. Med. Chem. 2, 759–777. doi: 10.2174/1568026023393624
Seong, J., Shin, J., Kim, K., and Cho, B. K. (2021). Microbial production of nematicidal agents for controlling plant-parasitic nematodes. Process Biochem. 108, 69–79. doi: 10.1016/j.procbio.2021.06.006
Shimada, A., Fujioka, S., Koshino, H., and Kimura, Y. (2010). Nematicidal activity of beauvericin produced by the fungus Fusarium bulbicola. Zeitschrift Für Naturforschung C 65, 207–210. doi: 10.1515/znc-2010-3-407
Toda, T., and Kurinobu, S. (2002). Realized genetic gains observed in progeny tolerance of selected red pine (Pinus densiflora) and black pine (P. thunbergii) to pine wilt disease. Silvae Genet. 51, 42–44.
Vaclavikova, M., Malachova, A., Veprikova, Z., Dzuman, Z., Zachariasova, M., and Hajslova, J. (2013). ‘Emerging’mycotoxins in cereals processing chains: Changes of enniatins during beer and bread making. Food Chem. 136, 750–757. doi: 10.1016/j.foodchem.2012.08.031
Valencia, J. W. A., Gaitán Bustamante, A. L., Jiménez, A. V., and Grossi-de-Sá, M. F. (2011). Cytotoxic activity of fungal metabolites from the pathogenic fungus Beauveria bassiana: An intraspecific evaluation of Beauvericin production. Curr. Microbiol. 63, 306–312. doi: 10.1007/s00284-011-9977-2
Vicente, C. S., Soares, M., Faria, J., Espada, M., Mota, M., Nóbrega, F., et al. (2022). Fungal communities of the pine wilt disease complex: Studying the interaction of ophiostomatales with Bursaphelenchus xylophilus. Front. Plant Sci. 13:908308. doi: 10.3389/fpls.2022.908308
Wang, C. Y., Fang, Z. M., Sun, B. S., Gu, L. J., Zhang, K. Q., and Sung, C. K. (2008). High infectivity of an endoparasitic fungus strain, Esteya vermicola, against nematodes. J. Microbiol. 46:380. doi: 10.1007/s12275-007-0122-7
Wang, C. Y., Fang, Z. M., Wang, Z., Gu, L. J., Sun, B. S., Zhang, D. L., et al. (2009). High infection activities of two Esteya vermicola isolates against pinewood nematode. Afr. J. Microbiol. Res. 3, 581–584.
Wang, C. Y., Yin, C., Fang, Z. M., Wang, Z., Wang, Y. B., Xue, J. J., et al. (2018). Using the nematophagous fungus Esteya vermicola to control the disastrous pine wilt disease. Biocontrol. Sci. Technol. 28, 268–277. doi: 10.1080/09583157.2018.1441369
Wang, H., Chu, H., Xie, Q., Dou, Q., Feng, H., Yang, C., et al. (2016). Variation in sporulation of four Esteya vermicola isolates and their infectivity against pinewood nematode. Sci. Silvae Sin. 52, 139–146. doi: 10.11707/j.1001-7488.20160917
Ye, S., Shang, L., Xie, X., Cao, Y., and Chen, C. (2021). Optimization of in vitro culture conditions for production of Cordyceps bassiana spores (Ascomycetes) and the effect of spore extracts on the health of Caenorhabditis elegans. Int. J. Med. Mushrooms 23, 44–55. doi: 10.1615/IntJMedMushrooms.2021038683
Youssef, M., El-Nagdi, W., and Lotfy, D. E. (2020). Evaluation of the fungal activity of Beauveria bassiana, Metarhizium anisopliae and Paecilomyces lilacinus as biocontrol agents against root-knot nematode, Meloidogyne incognita on cowpea. Bull. Natl. Res. Centre 44, 1–11. doi: 10.1186/s42269-020-00367-z
Zamora, P., Rodríguez, V., Renedo, F., Sanz, A. V., Domínguez, J. C., Pérez-Escolar, G., et al. (2015). First report of Bursaphelenchus xylophilus causing pine wilt disease on Pinus radiata in Spain. Plant Dis. 99:1449. doi: 10.1094/PDIS-03-15-0252-PDN
Zas, R., Moreira, X., Ramos, M., Lima, R. M. R., Da Silva, M. N., Solla, A., et al. (2015). Intraspecific variation of anatomical and chemical defensive traits in Maritime pine (Pinus pinaster) as factors in susceptibility to the pinewood nematode (Bursaphelenchus xylophilus). Trees 29, 663–673. doi: 10.1007/s00468-014-1143-6
Keywords: pine wilt disease, integrated management, biological control, nematophagous fungi, fungal toxins
Citation: Sánchez-Gómez T, Harte SJ, Zamora P, Bareyre M, Díez JJ, Herrero B, Niño-Sánchez J and Martín-García J (2023) Nematicidal effect of Beauveria species and the mycotoxin beauvericin against pinewood nematode Bursaphelenchus xylophilus. Front. For. Glob. Change 6:1229456. doi: 10.3389/ffgc.2023.1229456
Received: 26 May 2023; Accepted: 11 August 2023;
Published: 24 August 2023.
Edited by:
Isabel Alvarez Munck, Forest Service (USDA), United StatesReviewed by:
Claudia S. L. Vicente, University of Évora, PortugalCopyright © 2023 Sánchez-Gómez, Harte, Zamora, Bareyre, Díez, Herrero, Niño-Sánchez and Martín-García. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tamara Sánchez-Gómez, dGFtYXJhLnNhbmNoZXpAdXZhLmVz
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.