 Wenjun Liang
Wenjun Liang Yonghan Feng
Yonghan Feng Xi Wei
Xi Wei- College of Forestry, Shanxi Agricultural University, Taigu, China
Promoting natural regeneration in artificial forest ecosystems is crucial for sustainable management. Understanding the fundamental mechanisms and drivers of tree regeneration is the prerequisite for promoting it effectively. This study worked with Larix principis-rupprechtii, a species considered difficult to regenerate. Twenty-four sample plots measuring 30 m × 30 m were established, with eight plots at each of the lower, middle, and upper slope positions, respectively. Field investigation and multivariate analysis were performed to uncover the regeneration traits in the plantations with abundant seedlings on the continuous slope. The results revealed that ground diameter and height of the regeneration (RGD and RH) were larger at the lower slope, with significant positive correlations to available nitrogen (contribution rate, CR: 0.858) and slope (CR: 0.652). In contrast, regeneration density (RD), representing the quantity of regeneration, was greater at the middle slope. Its significant impact factors were slope position (CR: −0.648) and herb diversity, represented by Pielou index (CR: 0.961). Stand density had a significant negative effect on regeneration, particularly at the upper slope, with CRs of −0.842 and −0.764 to RGD/RH and RD, respectively. Common contribution was found among the factors, with the largest contribution groups being the topographical and soil factors (CR: 0.358). These findings provide valuable insights into the single species regeneration progress on northern mountainous slopes and offer essential information for developing facilitation methods for the natural regeneration in artificial forests.
1. Introduction
Artificial forests are responsible for more than 50% of the industrial wood supply worldwide (He et al., 2012) and are of great value to nature conservation (El-Keblawy and Ksiksi, 2005). However, the natural regeneration of these forests is often challenging, leading to the formation of overmature forests, declining productivity, significant economic and ecological losses. Conversely, natural regeneration is a cost-effective forest cultivation approach that has a significant research value for maintaining the structure, function, and diversity of artificial forest ecosystems (Sharma et al., 2019; Li et al., 2020). Therefore, the issue of single-species regeneration has garnered considerable attention (Zhao et al., 2010; Liu et al., 2015; Abbas et al., 2021). One of the most economically important temperate coniferous tree species in the high mountains of northern China and similar climatic regions is the Dahurian larch [Larix principis-rupprechtii (LP)]. Its habitat spans elevations from 1,500 to 3,600 m above sea level. The tree species stands out due to its advantageous traits, including a short production cycle, strong cold resistance, poor soil adaptability, and high-quality timber (Ye and Lei, 2019). However, the survival rate of naturally regenerated LP is significantly lower compared to that achieved through artificial regeneration (Liang and Wei, 2020). Implementing thinning practices from early to mature stages in LP forest becomes essential to create a more favorable growth environment for the remaining trees (Ma et al., 2022). It can deeply change both internal factors including seed quality, genetic diversity and structure, adaptability to the environment, as well as external environmental impact factors, such as stand density, root system, herb diversity, soil, and microclimate (Olesinski et al., 2012; Ma et al., 2014). These factors determine its natural regeneration dynamics together.
Natural regeneration of plantations is influenced by a range of biotic and abiotic factors, and their interactive characteristics affect the availability of resources and regeneration conditions (Shen and Nelson, 2018; Liang and Wei, 2021). In regions with water scarcity, identifying success factors for high-quality forest regeneration is crucial. They can aid in vegetation restoration, soil and water conservation, and the rational utilization of natural resources (Kremer and Bauhus, 2020). The overstory and understory composition, structure, and site conditions are considered as significant vegetation factors (Hart and Chen, 2006; Walters et al., 2016; Ali et al., 2019). For instance, regeneration density (RD) is related to stand age (Pham et al., 2022), stand density (Dobrowolska, 2008; Badalamenti et al., 2018), canopy density (Kara and Topaçoğlu, 2018), and distance from adult trees (Moreno-Fernandez et al., 2020). Simultaneously, the herbaceous layer has the highest biodiversity (Gilliam, 2007) and exerts divergent effects on seedling regeneration in different tree types (Collins and Good, 1987; Jensen et al., 2012; Gavinet et al., 2016; Soto et al., 2019). Herbs exhibit positive effects, such as protection from herbivory browsing and serving as a catalyst for regeneration (Randriambanona et al., 2019). On the other hand, they may hinder seed germination or intensify interspecific competition, thus impeding seedling growth (Grubb, 1977; Forbes et al., 2016; Thrippleton et al., 2016). Ground layer disturbance has a considerable impact on gap partitioning of understory vegetation, resulting in structural heterogeneity and biodiversity within forests. These changes, in turn, influence regeneration processes (Fahey and Puettmann, 2007). In light of these, reducing stand density, adjusting forest gaps, and regulating herb diversity have been proposed as effective methods to facilitate seedlings in previous studies (Mayer et al., 2017; Teketay et al., 2018; Hammond and Pokorný, 2020).
Tree regeneration is influenced by soil moisture and properties, including infiltration capacity and nutrient availability. Soil water deficit can hinder root growth, reducing the potential for seedling establishment and growth (Zadworny et al., 2021). The infiltration capacity of the soil determines the distribution of water resources, which can influence the growth and survival of seedlings (Fernández et al., 2019). Soil nutrients, such as organic matter, nitrogen, and phosphorus, can also affect the performance of trees and seedlings, either complementing or limiting their growth on different scales (Harpole et al., 2011; Venanzi et al., 2019; Liu et al., 2020a; Sklodowski, 2020). Nutrients often interact with other factors, such as light (Finzi and Canham, 2000; Valladares et al., 2000). Specifically, increasing nitrogen has been linked to enhancing sapling growth and productivity (Bigelow and Canham, 2007). Moreover, total nitrogen impacts on species differences and plant community development (Soto et al., 2017). Additionally, the nitrogen availability is a critical determinant that influences the spatial and temporal patterns of regeneration (Catovsky and Bazzaz, 2002). The germination, viability, and growth of seedlings are evaluated through growth-chamber, greenhouse, and field experiments (Chang et al., 1991; Davis et al., 2018; de Medeiros-Sarmento et al., 2021). Furthermore, the regeneration success of different tree species varies with different soil types, which exhibit diverse internal properties and spatial distribution of nutrients (Mayer et al., 2017). Soil characteristics also interact with other site factors, such as climate, topography, and vegetation, to shape the composition and structure of plant communities (Lin et al., 2017; Abbas et al., 2021).
Changing climatic factors are key abiotic drivers of species distribution and regeneration dynamics (Lin et al., 2017; Tiscar et al., 2017; Sagra et al., 2018). Nonetheless, the role of non-climatic constraints, such as topography, on tree regeneration is increasingly recognized. Topographic factors, including altitude, aspect, slope, and slope position, can directly or indirectly affect soil and microclimate conditions and influence seedling establishment and growth (Tsujino and Yumoto, 2008; Tingstad et al., 2015; Ameztegui et al., 2016). Particularly, on a uniform slope surface with consistent light, temperature, precipitation, and evaporation in mountainous regions, regeneration success primarily varies with micro-topography (Yu et al., 2013; Bergin and Kimberley, 2014; Thrippleton et al., 2016). Among them, slope position has gained considerable attention. Xu et al. (2020) found that slope position had a more significant effect on planted tree species than thinning intensity. Liu et al. (2020b) demonstrated that soil properties varied along the slope position gradient, which can promote vegetation restoration and ecological sustainability. Silva et al. (2018) reported that slope position influenced soil texture, but not its chemical properties. Conversely, Lin et al. (2017) found no significant changes in soil chemical properties related to slope position. These divergent results suggest that the influences of slope position on vegetation, soil, and regeneration outcomes are not conclusive and may vary with specific contexts and species.
The overarching objective of this study was to evaluate how the factors of vegetation and soil at varying slope positions affected the natural regeneration of a single species. By comparing the factors, we identified several drivers that influenced the growth of regenerated seedlings. We examined the progress of multi-factorial interactions at the slope scale, and our findings had implications for the natural regeneration of artificial forests and ecological environment restoration. An investigation on 24 sampling plots within an LP plantation situated across a continuous slope was conducted in a mountainous area. It aimed to assess the influences of various factors, including trees, herbs, water, soil, and other site conditions, on both the quantity and quality of the LP regeneration. The hypotheses are as follows: (1) high overstory stand density and understory herb diversity may independently or jointly play significant roles in tree regenerating abundance and growth; (2) increases of soil water availability and higher nitrogen content significantly influence the seedlings growth; and (3) slope position serves as a determinant of regeneration, with higher-quality regeneration typically observed at lower slopes due to generally higher soil moisture and fertility.
2. Materials and methods
2.1. Study site
The study is carried out within the forest zone of Guandi Mountain, situated in the Pangquangou Nature Reserve with the area of 10,443.5 ha in western Shanxi Province, China (37°45′–37°55′N, 111°22′–111°33′E; elevation 1,900–2,200 m). The average annual temperature and rainfall are 4.3°C and 822 mm, respectively, as reported by Liu et al. (2007). The dominant soil type is identified as Haplic Luvisol, according to the FAO classification, with an average thickness of about 60–70 cm (Yang et al., 2017). The study area, the size of which is 5.78 ha, has a temperate continental climate, with most precipitation occurring in the summer and early autumn. The area is characterized by long winters with low temperatures and short summers with more rain. The dominant tree species comprise Dahurian larch (LP), Chinese pine (Pinus tabulaeformis), east-Liaoning oak (Quercus wutaishanica), white birch (Betula platyphylla), and dragon spruce (Picea asperata). The main shrub communities include meadow sweets (Spiraea fritschiana), mayberry (Rubus corchorifolius), seabuckthorn (Hippophae rhamnoides), and rose (Rosa bella). The herbaceous layer is mainly composed of maizuru grass (Maianthemum bifolium), suzuran (Convallaria majalis), and carex (Carex spp.).
2.2. Area of study and sample design
A total of 24 sampling plots, each measuring 30 m × 30 m, were established on the northwest slope of Chailu ditch, located in Guandi Mountain. The ditch was located at the central part of the Pangquangou Nature Reserve. It had the average climate, topography and environment of the area and distinct elevation characteristics. The area was characterized by a well-established single-species plantation of LP with tree ages ranging from 42 to 70 years old, which was the natural regeneration gathering place of the main tree species. Therefore, Chailu ditch was selected as the study area because it basically could represent the ecological environment characteristics of the whole reserve, even other similar regions. The sampling plots were situated on the entire continuous slope surface, comprising randomly eight at the lower slope, eight at the middle slope, and eight at the upper slope, respectively (Figure 1). The artificial forest on the slope exhibited clear regeneration characteristics. Furthermore, the micro-topography of the plots was highly heterogeneous, with elevations ranging from 1,970 to 2,080 m above sea level and slope gradients varying from 3 to 32° (refer to Table 1 for detailed information). Inside all the 24 plots, the indicators of vegetation, and soil were measured to test three hypotheses (Table 2).
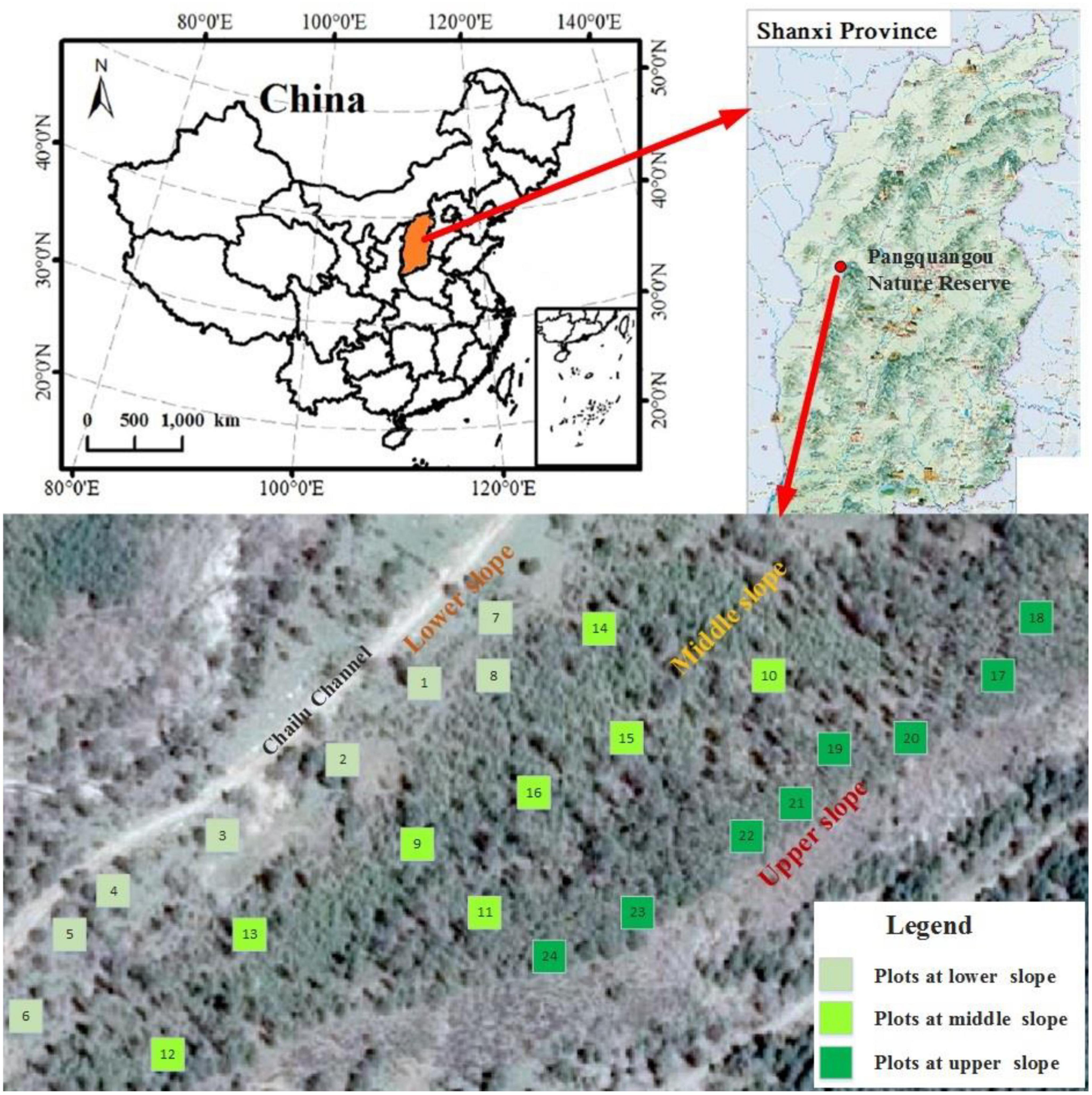
Figure 1. Schematic diagram of sample plots’ distribution.
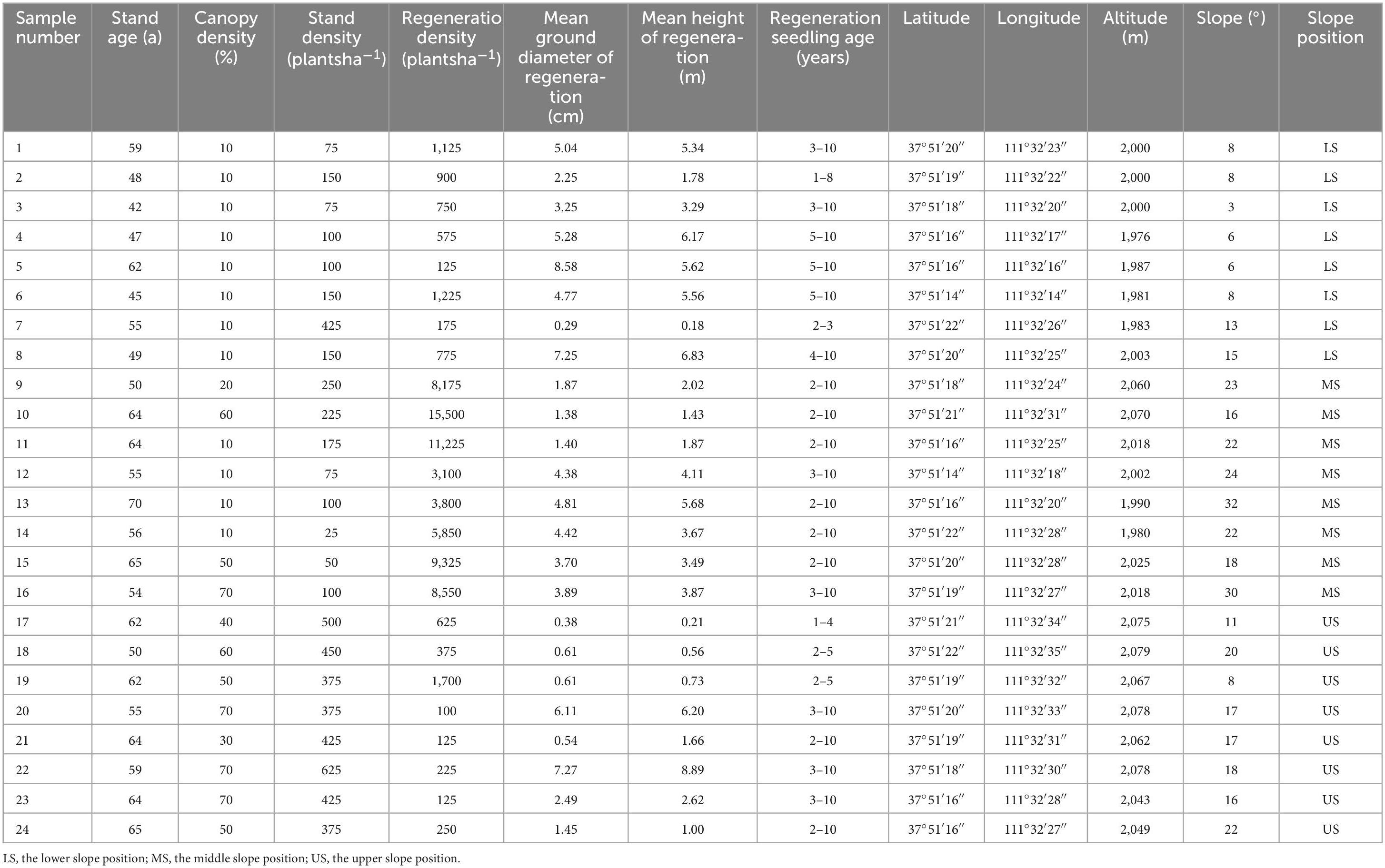
Table 1. Basic conditions of sample plots.
Table 2. Sampling design to test the hypotheses.
2.3. Vegetation investigation
Vegetation inventories were conducted on all 24 sampling plots during the close to natural regeneration of LP plantations from July to August 2020, coinciding with the vigorous summer growing season. The ages of the trees (unit: years) were determined using increment borers and were found to be larger than 40 years. Trees were regarded as seed bearers, whose diameters at breast height of 1.30 m (DBHs, unit: cm) were larger and tree heights (THs, unit: m) were higher than the regeneration seedlings. Canopy density (CD, unit: %) was the ratio of the total area projected by the tree crown to the area of stand which was calculated using drawing crown projection method. Stand density (SD, unit: plantsha–1) were calculated from the forest inventory data. Regeneration seedling was defined as an individual with an age of less than 10 years and a regeneration ground diameter (RGD, unit: cm) of less than 20 cm in all 30 m × 30 m plots. During the investigation, RGD was measured using vernier calipers and regeneration height (RH, unit: m) was measured using rulers. Regeneration density (RD, unit: plantsha–1), meaning the ratios of seedling number to each plot area, was calculated (refer to Table 1 for details).
The understory plant communities were characterized by a paucity of shrubs. Therefore, in each plot, three subplots of 1 m × 1 m were surveyed only for herbaceous vegetation within the forest stands. The species of herbs included maizuru grass (M. bifolium), suzuran (C. majalis), carex (Carex spp.), oriental strawberry (Fragaria orientalis Losina-Losinsk), small red chrysanthemum (Chrysanthemum chanetii H. Lév.), black bupleurum (Bupleurum smithii H. Wolff), common plantain (Plantago asiatica L.), burdock [Trigonotis peduncularis (Trevis.) Benth. ex Baker & S. Moore], white mugwort (Artemisia sacrorum Ledeb.), herba leonuri (Geranium wilfordii Maxim.), poison hemlock (Pedicularis reaupinanta L.), Chinese rhubarb (Euphorbia pekinensis Rupr.), purple gromwell (Viola philippica), Chinese ginseng (Polygonatum sibiricum Delar. ex Redoute), buttercup (Ranunculus japonicus Thunb.), Japanese millet (Poa annua L.), fiddlehead fern (Potentilla chinensis Ser.), chenopodium (Veratrum nigrum L.), etc. The data collected was then used to evaluate the herbaceous diversity and evenness using classic indices, namely the Shannon–Wiener index(herb-H′ = ), Simpson index (herb-D = ), and Pielou index (herb-J = H′/ ln S), following the established formulas (Shannon and Wiener, 1949; Simpson, 1949; Pielou, 1966; Daniel et al., 2019).
2.4. Soil sampling
To analyze the soil characteristics, three random replicated soil profiles were established per plot, with each profile encompassing three different soil depths (0–20, 20–40, and 40–60 cm). The cutting ring method was employed to measure soil moisture content. To mitigate the influence of rainfall on soil moisture, soil sampling was carried out during mostly clear weather conditions or delayed by at least 3 days following any precipitation event. The soil moisture content was determined by subjecting the collected soil samples to oven-drying at 105°C for 24 h. The mean value of the three layers was calculated to represent the soil profile content, and the values of the three replicated samples were averaged to determine the soil moisture content per plot. Furthermore, the soil infiltration rate was measured using the double loop infiltration approach.
Additionally, one soil sample from each layer was placed in a valve bag and air-dried for subsequent analysis of soil nutrient content. The samples from the three soil layers of the same soil profiles were mixed uniformly to assess the soil nutrition content. Soil organic matter was determined by means of the K2Cr2O7 volumetric method after boiling the elimination. Total nitrogen was measured using the DigiPREP TKN system (JK9890, China) following elimination by boiling. Total phosphorus and available nitrogen were quantified through extractions with HClO4-H2SO4 and KCl, respectively. Furthermore, available phosphorus was determined according to Olsen’s method, utilizing a NaHCO3 solution.
2.5. Statistical analyses
The datasets obtained from the plots situated at different slope positions, including the above mentioned trees, regeneration seedling, herbs, soil, and topographical factors, were subjected to the distribution and probability density analysis of the lower, middle, and upper slope position groups, showing with violin plots. Among them, regeneration seedling indicators included RD, height, and ground diameter. Topographical factors contained slope position, elevation, and slope. Soil nutrient availability was assessed through measurements of soil organic matter, total nitrogen, total phosphorus, available nitrogen, and available phosphorus. To gauge the direction and strength of linear correlations between the tree and regeneration seedling indicators, soil, and topographical factors, Pearson’s correlation test was employed. Statistical analyses were conducted using SPSS 21.0 software (IBM Corporation, Armonk, NY, USA). Statistical significance was considered at P ≤ 0.05.
Redundancy analysis (RDA) was used to assess the relationship between soil moisture and nutrients at different slope positions and regeneration indicators (i.e., RGD, RH, and RD). The method can analyze the specific correlations between these variables and identify the factors responsible for the variation in the target variables. RDA was executed using SPSS 21.0 and CANOCO 5.0 software packages (Microcomputer Power, Ithaca, NY, USA), which allowed for the identification of significant variables and their respective contribution rates (CRs).
To further analyze the independent and common contributions of the various environmental factors, we classified them into four groups: herb diversity (including the three herb diversity indexes), stand structure (including stand age, stand density, and canopy density), topographical factors (including slope position, slope, and altitude) and soil factors (including all the above soil indicators). Variance partitioning analysis (VPA) was used to study the independent and common contributions of each group of these environmental factors through the Venn diagram, in order to make up the deficiency of RDA analysis. Figures were generated using Origin 9.0 software (Origin Lab, Northampton, MA, USA) and R-Statistical software (R Core Team, 2008).
3. Results
3.1. Effects of overstory and understory upon natural regeneration
During the growing season of 2020, the number of regenerated seedlings per hectare varied significantly, ranging from 125 to 15,500 across all plots. The changes in RD, height (RH), and ground diameter (RGD) on different slope positions were illustrated in Table 1. The lower, middle, and upper slope positions demonstrated very different RDs with mean values of 706, 8,191, and 441 plantsha–1, respectively. The mean values of RGDs and RHs were 4.59 cm versus 4.3 m at the lower slope, 3.23 cm versus 3.3 m at the middle slope, and 2.43 cm versus 2.7 m at the upper slope. These findings indicate that slope position played a critical role in the growth of LP seedlings. Especially, the middle slope had a higher quantity of seedlings compared to the other two slope positions.
Slope position was found to be a topographic factor that indirectly affected tree regeneration through direct factors, including vegetation in the overstory and herbs in the understory. In this study, we focused on the influence of herb diversity on the regeneration of LP plantations, illustrated in Figure 2. The herbaceous diversity was measured by Shannon–Wiener index, Simpson index, and Pielou index, which highlighted diversity, richness and evenness, respectively. The mean values of the three indices showed some similarity, which were largest on the middle slope, but smaller on the lower and upper slope. The distributions of the three herbaceous diversity indexes had variability in values because of the different meanings of the three diversity indexes and the small sample size. Changes in herb diversity at different slope positions were basically consistent with RD and different from RGD and RH. That emphasized herb diversity could influence number and growth of LP regeneration seedlings.
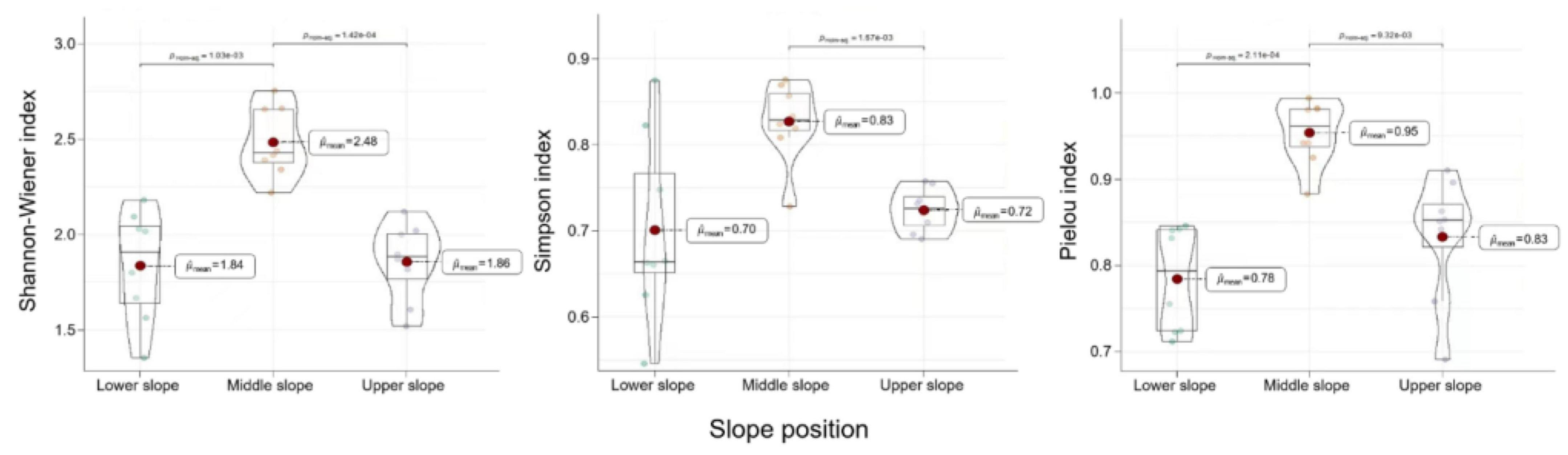
Figure 2. Changes in Shannon–Wiener index, Simpson index, and Pielou index of the herbs in response to slope position variation.
In addition to slope position and herb diversity, stand structure in the overstory was identified as another dominant vegetation factor that directly impacted on regeneration dynamics. The ages of the surveyed forests were between 42 and 70 years old and consisted of near-mature and mature timbers that had undergone selection cutting. To investigate changes in the stand structure and regeneration properties in LP plantations, we measured stand density, canopy density, RD, RGD, and RH, as presented in Figure 3.
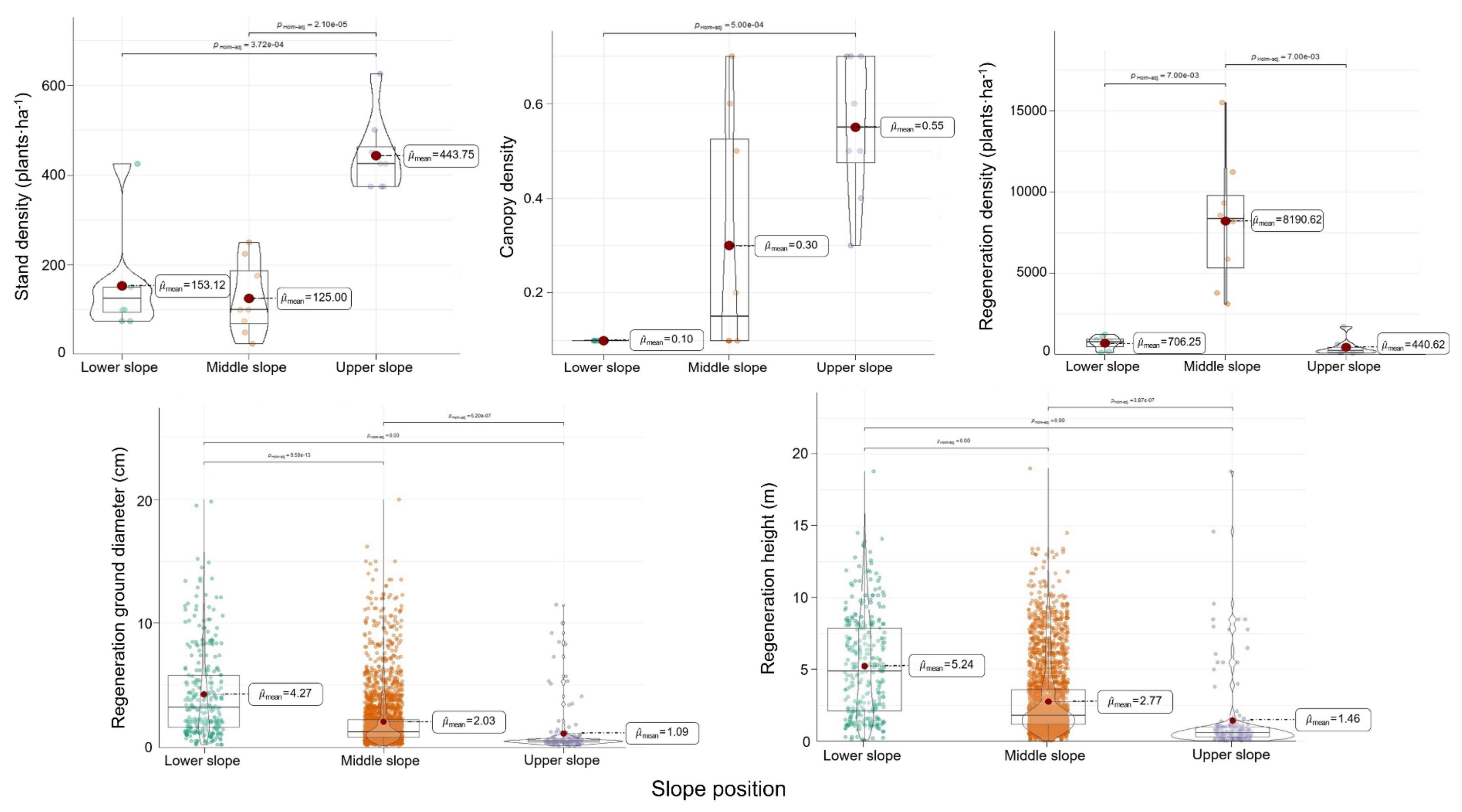
Figure 3. Changes in stand density (plantsha– 1), canopy density, regeneration density (plantsha– 1), regeneration ground diameter (cm), and regeneration height (m) in response to slope position variation.
Variation in structural indicators was evident across different slope positions. The LP at the middle and upper slopes had mean ages of 60 years each (Table 1). Stand age was found to influence the number and growth of LP regeneration seedlings, with no significant effect of slope position. Stand densities and canopy densities of the lower and middle slopes’ plots were lower than the upper slopes, with mean values of 153, 125, and 444 plantsha–1 for stand density, and mean values of 10, 30, and 55% for canopy density, respectively. The distributions of the values in the three slope positions exhibited considerable differences. The middle slope had the largest mean value of RD, where stand densities and canopy densities were moderate. This suggested that lower stand density and canopy density might be beneficial for LP regeneration. The distribution of RGD and RH of the LP regeneration seedlings, as shown in Figure 3 and Appendix Table 1, were higher on the lower and middle slopes compared to the upper slope. It indicated that the biomass of the seedlings, as represented by RGD and RH, grew better under sparse canopies than thick ones, especially on the lower slope.
Successful regeneration of LP plantations was more prominent at the lower and middle slopes, attributed to their higher biomass per plant and larger quantity, respectively. The impact of herb diversity and forest canopy on regeneration seedlings varied across the lower, middle, and upper slope positions. Similar changes in Shannon–Wiener index, Simpson index, and Pielou index of herbs were observed at the lower and middle slopes compared to RD. Interestingly, distinct changes were observed in stand density and canopy density at the lower and middle slopes, along with variations in herb diversity indexes at the upper slope. In addition, lower stand density increased the diversity, suggesting a potential synergy between the two factors for regeneration, referring to Figures 2, 3.
3.2. Differential effects of soil properties on regeneration
Soil factors, comprising soil water and nutrients, were generally recognized to play a critical role in forest regeneration. As depicted in Figure 4, among the surveyed plots, the soil infiltration rates at the lower, middle, and upper slope did not exhibit markedly distinct features, with mean values of 56.78, 66.47, and 72.71 mm/h, respectively. In comparison, the variation of the RDs was more pronounced, with the most abundant regeneration observed at the middle slope. The mean volumetric moisture contents in the 0–60 cm soil depth were 22.11, 22.37, and 25.27% from lower to upper slope, respectively. As a key indicator, the values of soil moisture content at the lower and middle slopes were smaller than that at the upper slope. They were oppositely related to RGD and RH, but not significantly related to RD. Nonetheless, taking into account the regeneration status of the surveyed stands, it appeared that soil moisture did not act as a limiting factor for LP regeneration across the entire slope.
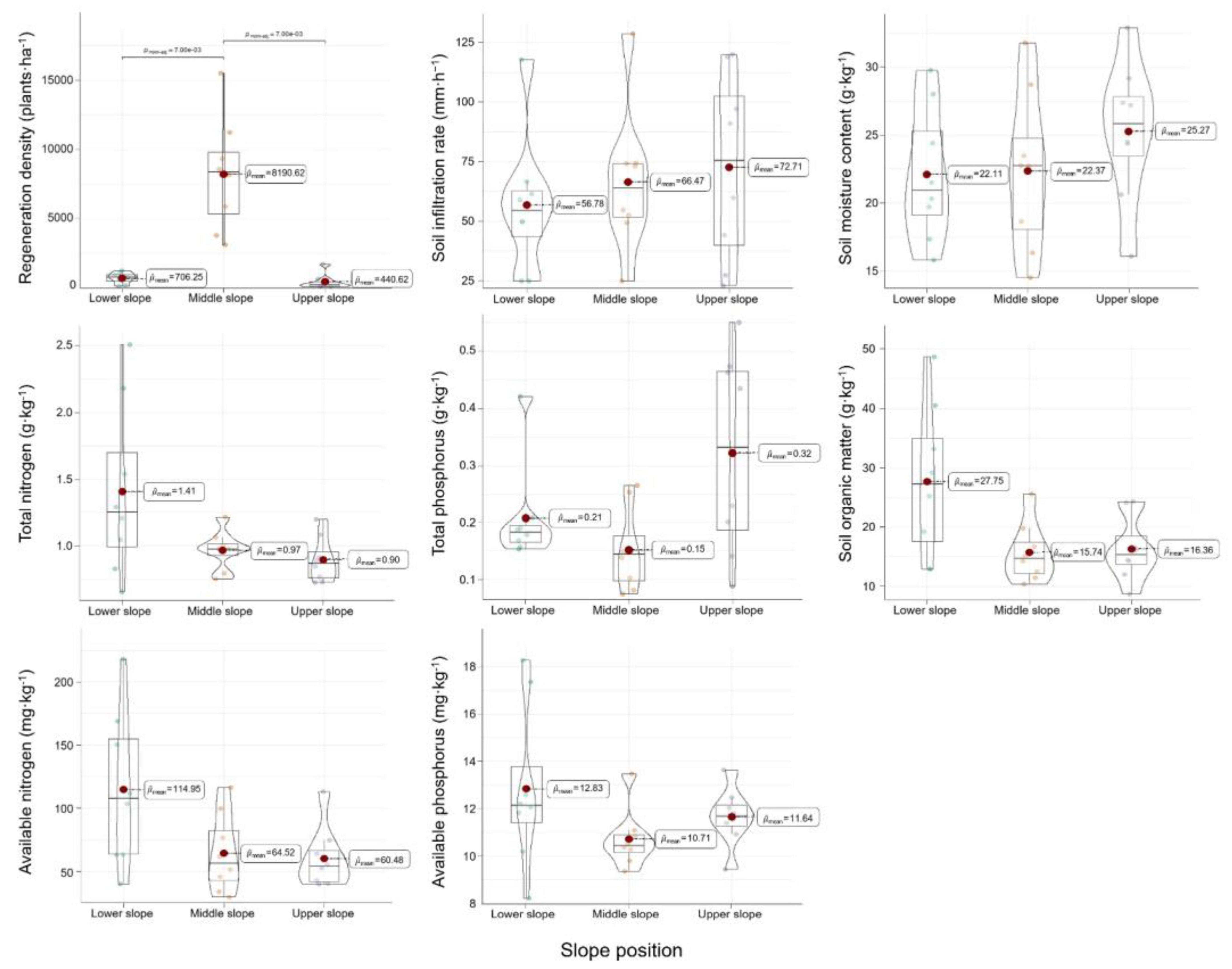
Figure 4. Changes in regeneration diversity in response to the variables of soil water and nutrients at different slope positions.
As predicted, different distributions of the data were observed in most soil chemical properties across the three slope positions by the violin plots (Figure 4 and Appendix Table 1). Soil organic matter, total nitrogen, and available nitrogen exhibited similar distributions, with higher concentrations at the lower slope and lower concentrations at the upper slope. Mean soil organic matter values from lower to upper slope were 27.75, 15.74, and 16.36 g/kg, respectively. Mean total nitrogen values were 1.41, 0.97, and 0.90 g/kg, and mean available nitrogen values were 114.95, 64.52, and 60.48 mg/kg, respectively. Total phosphorus and available phosphorus first decreased, then increased with increasing slope position, contrary to the aforementioned three indicators. Accounting for the regeneration of the local population, soil nutrients at the lower slope, with the exception of total phosphorus, were more favorable for the seedlings growth compared to those at the middle and upper slopes, as indicated by the RGD and RH measurements in Figure 3.
The heights and ground diameters of regenerating seedlings were affected by soil indicators, including organic matter, total nitrogen, and available nitrogen, due to their significant variation across the slope. However, RD was not significantly related to soil properties. These findings suggested that the effects of soil water on regeneration were limited across the entire slope. There were no significant relationships between regeneration and individual nutrients such as soil organic matter and nitrogen. A combination of soil water and nutrients may enhance natural regeneration after seed germination at the lower slope, where soil nutrients were relatively high. Notably, the above data and conclusions (referring to Figures 2–4) had variability and needed further confirmation due to the smaller sample size.
3.3. Dominant factors driving regeneration across the continuous slope
Pearson’s correlation analysis revealed significant associations between regeneration and its impact factors along the continuous slope (Figure 5). Notably, the three regeneration indicators exhibited varying correlations. RGD and RH, representing seedling growth, showed a strong positive correlation (CC: 0.939) and were both negatively correlated with the number of seedlings (RD) (CCs: −0.196 and −0.189, respectively).
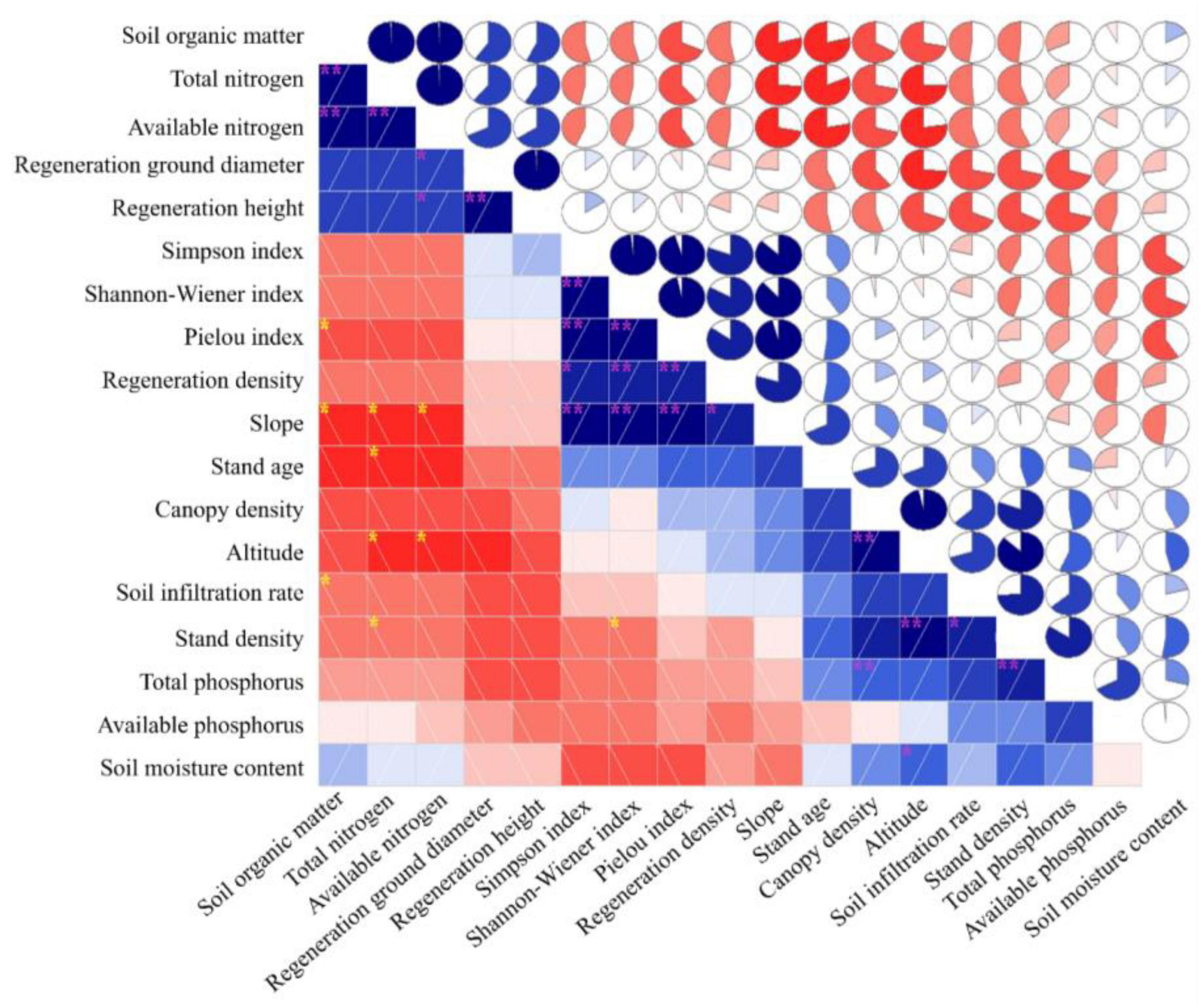
Figure 5. Pearson’s correlation analysis showing the relations between regeneration and the influencing factors including stands, herbs, and soils on the slope surface. The bar and circle symbols use the same color, and the correlation is reflected by the shading and saturation. Bar details: blue with a slash from bottom left to top right denotes two variables that are positively correlated, and red with a slash from top left to bottom right denotes two variables that are negatively correlated. The darker the color is, the greater the correlation between variables. Circle details: blue with the pie chart filled clockwise from 12 o’clock indicates the two variables are positively correlated, and red with a counterclockwise-filled pie chart indicates the two variables are negatively correlated. “**” means the correlation is significant at 0.01 level (P ≤ 0.01); “*” means the correlation is significant at 0.05 level (P ≤ 0.05).
The three herb diversity indexes exhibited significant positive correlations among themselves, which influenced regeneration quantity and quality to varying degrees. Shannon–Wiener index, Simpson index, and Pielou index of herb were not significantly negatively correlated with RGD and RH (CCs ≤ 0.22), but were significantly positively correlated with RD (CCs: 0.613, 0.502, and 0.546, respectively). On the other hand, the correlations between regeneration and stand factors in the overstory were measured. The absolute values of the CCs ranged from 0.095 to 0.372 and were not significant. Stand density had a negative impact on all regeneration variables. Canopy density and stand age had positive correlations with RD and negative correlations with RGD and RH.
Furthermore, several soil chemical properties in the three slope positions exhibited significant correlations with regeneration variables. Specifically, available nitrogen showed a positive correlation with RGD and RH, with coefficients of 0.408 and 0.436, respectively. Soil organic matter and nitrogen affected regeneration similarly, as evidenced by significant relationships between soil organic matter, total nitrogen, and available nitrogen (CCs: 0.929, 0.924, and 0.885, respectively). Additionally, soil moisture content, soil infiltration rate, slope, and altitude were positively correlated with RD, but negatively correlated with RGD and RH.
Based on the significant correlations and differences observed across the lower, middle, and upper slopes, herb diversity and slope had stronger positive interactions with RD, directly influencing regeneration quantity. Conversely, available nitrogen, soil organic matter, and total nitrogen exhibited significant positive correlations with RGD and RH, driving the quality of the regeneration. To identify the most influential factors on regeneration dynamics, the method of RDA was applied (Figure 6).
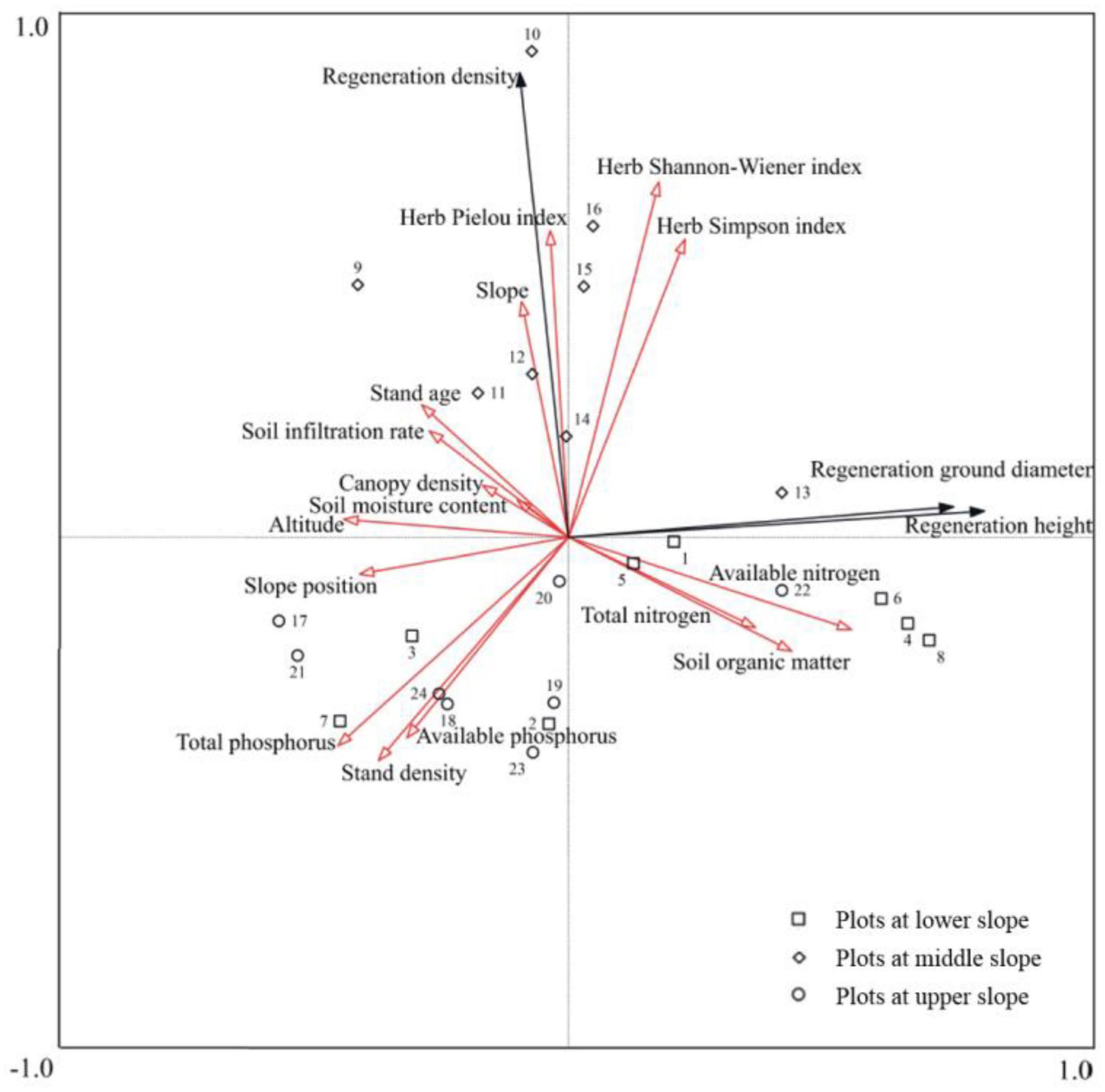
Figure 6. Redundancy analysis of the factors affecting regeneration at the different slope positions.
The results showed that the explanatory variables collectively accounted for 83.1% of the variation, with eigenvalues of 0.412, 0.311, and 0.108, respectively. The individual impact of each factor on LP regeneration was determined by its explanatory value. Notably, available nitrogen, soil moisture content, herb Shannon–Wiener index, and herb Pielou index had the largest explanatory values of 15.9, 14.6, 5.9, and 5.9%, respectively (Table 3). CR, which represented the extent to the factors influenced LP regeneration, was indicated by the length and angle of a red arrow with respect to a black arrow in Figure 6. The dominant factors that positively affected RD were Pielou index, Shannon–Wiener index, and Simpson index, and slope, with CRs of 0.961, 0.943, 0.887, and 0.652, respectively. The key positive contributors to both RGD and RH, indicated by two essential coincident arrows, were available nitrogen (CR: 0.858), soil organic matter (CR: 0.731), and total nitrogen (CR: 0.643). Altitude (CR: −0.775), slope position (CR: −0.648), and soil infiltration rate (CR: −0.601) were the primary negative drivers affecting seedling growth. Stand density, total phosphorus, and available phosphorus made significant negative contributions to regeneration, with CRs of −0.764, −0.712, and −0.537 to RD, and −0.842, −0.729, and −0.633 to RGD and RH, respectively.
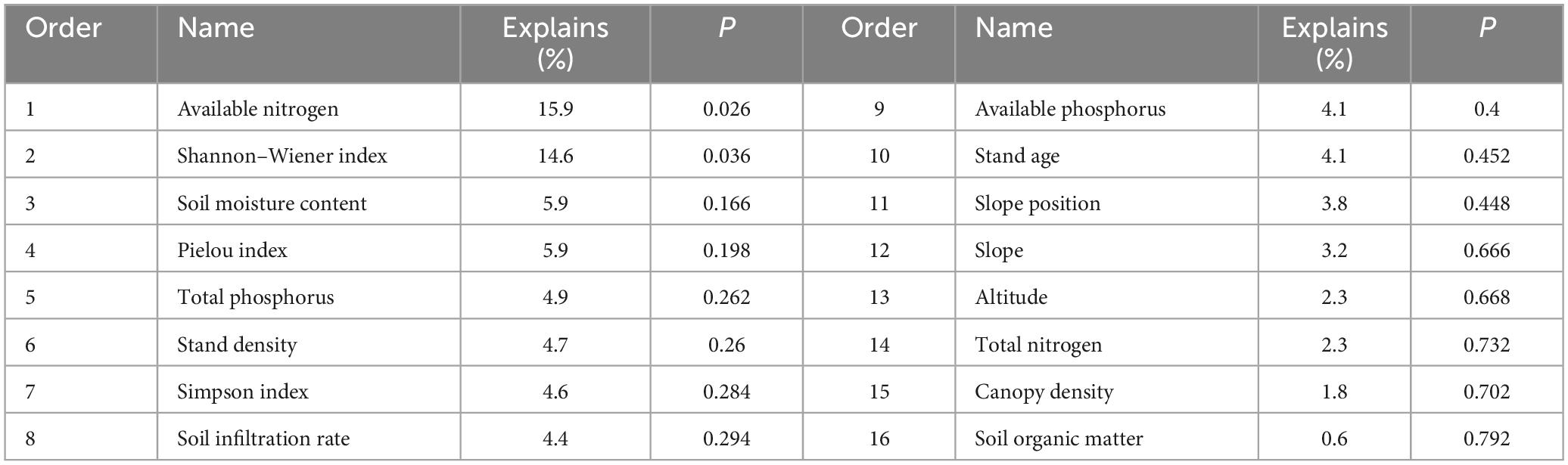
Table 3. Explanatory contribution of the different variables tested for regeneration in the RDA analysis.
The Venn diagram displayed the independent and common contributions of each group of the environmental factors (Figure 7). The combined CR (0.358) of the topographical and soil factors was largest. The factors in the independent group of stand structure had the second larger contribution (0.196). In addition, there were some joint contributions that cannot be ignored among three groups of topographical factors, stand structure, and herb diversity, with the CRs of 0.174, 0.118, 0.114, and 0.053.
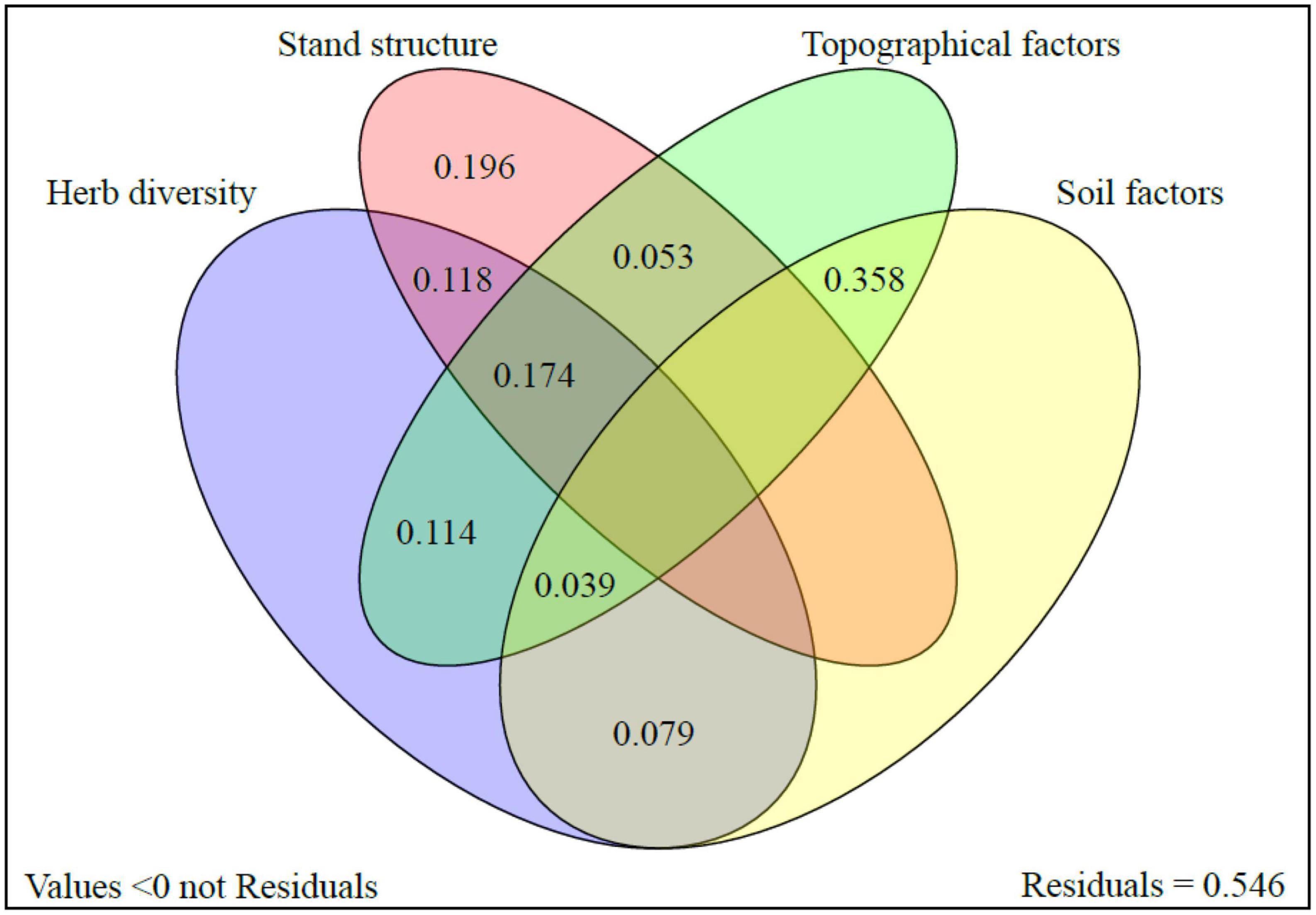
Figure 7. Venn diagram of the factors affecting regeneration.
Based on the extensive feature analysis, Pearson correlation analysis, RDA, and VPA, several dominant factors affecting regeneration were identified. The determinants included soil nitrogen (represented by available nitrogen), herb diversity (represented by Pielou index), soil moisture content, stand density, slope, slope position, and altitude. Notably, their roles in regeneration varied depending on the slope position, as illustrated in the scatter plot distribution in Figure 6 and the Venn diagram in Figure 7. Soil available nitrogen demonstrated a positive impact on RGD and RH, particularly at the lower slope. Conversely, Pielou index of herb and slope positively influenced RD at the middle slope. Interestingly, stand density, slope position, and altitude exhibited negative effects primarily at the upper slope, leading to the reduced LP regeneration in those areas. Moreover, an interesting observation was made that lower stand density contributed to increase the diversity, suggesting that stand structure and herb diversity played a combined role in influencing regeneration, as depicted in Figures 2, 3, 7.
4. Discussion
4.1. Promoting effects of vegetation at different layers on tree regeneration
The ability of single species plantations to sustain themselves through succession has been a topic of recent debate (Ali et al., 2019). The success of seed germination and subsequent regeneration may depend on the tree species, the presence of trees in the overstory and herbs in the understory, with minimal shrub coverage (Sklodowski, 2020). Operations of forest canopy can directly impact on stand density or canopy density, and indirectly alter the internal ecological environment of the stand (Kara and Topaçoğlu, 2018; Picchio et al., 2020), leading to frequent changes in both tree regeneration and understory vegetation (Sharma et al., 2019). Of the regenerated seedlings, those reflected by RD, ground diameter, and height were the most important in terms of the changing progress (Figure 3). The presence of canopy gaps provided a valuable opportunity for seed germination and seedling growth, owing to the low stand densities and canopy densities within the plots, particularly at the lower and middle slopes. Consequently, maintaining a low tree density may facilitate the regeneration of artificial forests, as previously observed (Dobrowolska, 2008; Badalamenti et al., 2018; Moelder et al., 2019), with fewer plants per hectare in many cases (Bütler et al., 2013).
The growth and diversity of herbs in the understory have been identified as important factors affecting the regeneration of seedlings in forest ecosystems. Previous studies reported varying effects of herbs on the regeneration of different tree species (Forbes et al., 2016; Randriambanona et al., 2019). In our study, it was found that all the three herb diversity indices (with strong correlation, Figure 5) showed positive associations with the regeneration of seedlings, due to their similar changes consistent with RD (Figure 2). Pielou index was selected as the representative index of herb diversity. Considering their co-influence, the interaction between the forest canopy, reduced tree density, and increased Pielou index fostered abundant and high quality regeneration. In practical forest management, several effective methods can be employed to promote natural succession of pure plantations. Thinning, selection cutting, and sowing different grass seed mixtures had been demonstrated to be successful strategies in this regard (Mayer et al., 2017; Teketay et al., 2018; Hammond and Pokorný, 2020). These measures facilitated the development of diverse and robust herb communities in the understory, which in turn promoted successful seedling regeneration and the maintenance of forest ecosystem health.
4.2. Soil available nitrogen was a driver to enhance tree regeneration
Soil moisture and fertility have long been considered as critical factors affecting plant growth and development (Yang et al., 2009; Dyderski et al., 2018; Liu et al., 2020b). In semi-arid and arid regions, such as the mountainous area of northern China (Liang and Wei, 2021), soil water availability is particularly emphasized as a potential constraint for plant and seedling survival (Zadworny et al., 2021). Furthermore, soil properties, such as nitrogen availability, have been identified as significant factors limiting regeneration in nutrient-poor environments (Catovsky and Bazzaz, 2002; Soto et al., 2017; Davis et al., 2018; Haruka and Toshiya, 2020). Therefore, we hypothesized that higher soil nitrogen would be closely related to the regeneration of seedlings in the study area.
However, tree regeneration on the typical continuous slope in the mountainous region was not limited by soil water. The reason was likely that the abundance of precipitation during the growing season of 2020, as evidenced by data from https://data.cma.cn/. Soil and topography had a significant common contribution (CR: 0.358 in Figure 7) for regeneration. However, the amounts of available nitrogen and total nitrogen significantly influenced the growth of seedlings, yet their influence on regeneration quantity was not significant surprisingly. The results differed from conventional knowledge (Perry et al., 2008; Haruka and Toshiya, 2020). Among the measured soil factors, available nitrogen emerged as the most significant soil characteristic, showing negative correlations with regeneration quantity and positive correlations with regeneration quality. Notably, available nitrogen, total nitrogen, and organic matter had strong correlations. Therefore, regulating soil available nitrogen can serve as a potential driver to enhance tree regeneration, particularly during wet years.
4.3. Effects of topography on tree regeneration
Slope position, as an indirect factor of vegetation, redistributes water, soil properties, and other resources along the slope position gradients (Wang et al., 2017; Silva et al., 2018; Xu et al., 2020). The present investigation revealed that soil nutrients at the lower slope were substantially greater than those at the middle and upper slopes (Figure 4), consistent with previous findings (Liu et al., 2020a). Nevertheless, the pattern of tree regeneration did not follow that of water and soil, as seedlings were abundant at the middle slope but scarce at the lower and upper slopes. It indicated that the quantity of regeneration was strongly influenced by slope position. The middle slope was found to be more suitable for seed germination owing to the rich herbaceous diversity in the area.
Prior to conducting the slope surface survey, we hypothesized that the lower slope would be the most conducive for regeneration. Our findings surpassed the initial expectations, revealing a clear pattern in the similar climatic conditions. Based on the drivers identified, it was proposed that the middle slope, with its steeper gradient, may possess stronger drainage capabilities when water was plentiful, making it more favorable for seed germination. As such, slope and slope position, key topographic factors, profoundly influenced the regeneration dynamics. Especially, the overall conditions on the middle slope were advantageous for seed germination. It should be noted, however, that in cases of water scarcity, dominant factors may shift and require further investigation.
5. Conclusion
On a continuous slope surface, the single species LP plantation exhibited the highest RD at the middle slope, contrary to our initial hypothesis. On the other hand, RGD and RH were identified as indicators of regeneration quality, indicating successful growth predominantly at the lower slope. A high Pielou index (representing herb diversity) and increasing slope facilitated greater seedling numbers at the middle slope. The higher available nitrogen, identified as the most significant soil property, was positively correlated with seedling growth at the lower slope. Conversely, the high stand density played a negative role in natural regeneration, particularly at the upper slope. In addition, the lower stand density increased the herb diversity. Combined with the largest contribution groups of the topographical and soil factors, the middle slope provided a positive comprehensive condition on the regeneration. These findings offer valuable insights into understanding the progress of single species regeneration for plantation regeneration in northern mountainous slope areas.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
WL: software, writing—original draft, conceptualization, funding acquisition, methodology, writing—review and editing, and supervision. XW: data curation, formal analysis, investigation, writing—original draft, visualization, conceptualization, funding acquisition, methodology, and writing—review and editing. YF, FL, YZ, JZ, JY, and CC: formal analysis, investigation, and visualization. All authors approved the manuscript.
Funding
The study was supported by the National Natural Science Foundation of China (31971644, 31971725, and 31901365), the National Key R&D Program of China (2022YFF1300401), Scientific and Technological Innovation Programs of Higher Education Institutions in Shanxi (2021L105), the Fund for Introduced Talents for Shanxi Agricultural University (2018YJ09), and the Shanxi Provincial Outstanding Doctoral Program for Incentive Funds for Scientific Research Projects (SXYBKY2018032).
Acknowledgments
We would like to thank our colleagues for their comments on this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbas, S., Nichol, J. E., Zhang, J. L., Fischer, G. A., Wong, M. S., and Irteza, S. M. (2021). Spatial and environmental constraints on natural forest regeneration in the degraded landscape of Hong Kong. Sci. Total Environ. 752:141760. doi: 10.1016/j.scitotenv.2020.141760
Ali, A., Dai, D., Akhtar, K., Teng, M., Yan, Z., Urbina-Cardona, N., et al. (2019). Response of understory vegetation, tree regeneration, and soil quality to manipulated stand density in a Pinus massoniana plantation. Glob. Ecol. Conserv. 20:200775. doi: 10.1016/j.gecco.2019.e00775
Ameztegui, A., Coll, L., Brotons, L., and Ninot, J. M. (2016). Land-use legacies rather than climate change are driving the recent upward shift of the mountain tree line in the Pyrenees. Glob. Ecol. Biogeogr. 25, 263–273. doi: 10.1111/geb.12407
Badalamenti, E., Bueno, R. D. S., Campo, O., Gallo, M., Veca, D. S. L. M., Pasta, S., et al. (2018). Pine stand density influences the regeneration of Acacia saligna labill. Hlwendl. And native woody species in a mediterranean coastal pine plantation. Forests 9:359. doi: 10.3390/f9060359
Bergin, D. O., and Kimberley, M. O. (2014). Factors influencing natural regeneration of totara (Podocarpus totara D.Don) on grazed hill country grassland in Northland, New Zealand. N. Zeal. J. For. Sci. 44:13. doi: 10.1186/s40490-014-0013-8
Bigelow, S. W., and Canham, C. D. (2007). Nutrient limitation of juvenile trees in a northern hardwood forest: Calcium and nitrate are preeminent. For. Ecol. Manage. 243, 310–319.
Bütler, R., Lachat, T., Larrieu, L., and Paillet, Y. (2013). “Habitat trees: Key elements for forest biodiversity,” in Integrative approaches as an opportunity for the conservation of forest biodiversity, eds D. Kraus and F. Krumm (Joensuu: European Forest Institute), 84–91.
Catovsky, S., and Bazzaz, F. A. (2002). Nitrogen availability influences regeneration of temperate tree species in the understory seedling bank. Ecol. Applic. 12, 1056–1070.
Chang, P. Y., Lin, M. L., Zhao, S. J., Chang, C., and Guo, L. S. (1991). Promotion of germination of coniferous seeds by snow stratification combined with dehydration-rehydration treatment. N. For. 5, 239–246.
Collins, S. L., and Good, R. E. (1987). The seedling regeneration niche: Habitat structure of tree seedlings in an oak-pine forest. Oikos 48, 89–98.
Daniel, J. A., Ramaraju, K., and Rameshkumar, A. (2019). Comparative studies of mymarid diversity from three different zones of paddy ecosystem in Tamil Nadu, India. ENTOMON 44, 173–182. doi: 10.33307/entomon.v44i3.458
Davis, E. L., Hager, H. A., and Gedalof, Z. (2018). Soil properties as constraints to seedling regeneration beyond alpine treelines in the Canadian Rocky Mountains. Arctic Antarct. Alpine Res. 50:e1415625. doi: 10.1080/15230430.2017.1415625
de Medeiros-Sarmento, P. S., Ferreira, L. V., and Gastauer, M. (2021). Natural regeneration triggers compositional and functional shifts in soil seed banks. Sci. Total Environ. 753:141934. doi: 10.1016/j.scitotenv.2020.141934
Dobrowolska, D. (2008). Effect of stand density on oak regeneration in flood plain forests in Lower Silesia, Poland. Forestry 81, 511–523. doi: 10.1093/forestry/cpn025
Dyderski, M. K., Gazd, A. A., Hachulka, M., Horodecki, P., Kalucka, I. L., Kamczyc, J., et al. (2018). Impacts of soil conditions and light availability on natural regeneration of Norway spruce Picea abies (L.) H. Karst. in low-elevation mountain forests. Ann. For. Sci. 75:91. doi: 10.1007/s13595-018-0775-x
El-Keblawy, A., and Ksiksi, T. (2005). Artificial forests as conservation sites for the native flora of the UAE. For. Ecol. Manage. 213, 288–296. doi: 10.1016/j.foreco.2005.03.058
Fahey, R. T., and Puettmann, K. J. (2007). Ground-layer disturbance and initial conditions influence gap partitioning of understory vegetation. J. Ecol. 95, 1098–1109. doi: 10.1111/j.1365-2745.2007.01283.x
Fernández, C., Ma Fernandez-Alonso, J., and Vega, J. A. (2019). Effects of mastication of burned non-commercial Pinus pinaster Ait. trees on soil compaction and vegetation response. For. Ecol. Manage. 449:117457. doi: 10.1016/j.foreco.2019.117457
Finzi, A. C., and Canham, C. D. (2000). Sapling growth in response to light and nitrogen availability in a southern New England forest. For. Ecol. Manage. 131, 153–165.
Forbes, A. S., Norton, D. A., and Carswell, F. E. (2016). Tree fern competition reduces indigenous forest tree seedling growth within exotic Pinus radiata plantations. For. Ecol. Manage. 359, 1–10. doi: 10.1016/j.foreco.2015.09.036
Gavinet, J., Prevosto, B., and Fernandez, C. (2016). Do shrubs facilitate oak seedling establishment in Mediterranean pine forest understory? For. Ecol. Manage. 381, 289–296. doi: 10.1016/j.foreco.2016.09.045
Gilliam, F. S. (2007). The ecological significance of the herbaceous layer in temperate forest ecosystems. Bioscience 57, 845–858. doi: 10.1641/B571007
Grubb, P. (1977). The maintenance of species-richness in plant communities: The importance of the regeneration niche. Biol. Rev. 52, 107–145.
Hammond, M. E., and Pokorný, R. (2020). Effects of gap size on natural regeneration and micro-environmental soil conditions in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst) dominated mixed forest. Plant Soil Environ. 66, 607–615. doi: 10.17221/397/2020-PSE
Harpole, W. S., Ngai, J. T., Cleland, E. E., Seabloom, E. W., Borer, E. T., Bracken, M. E., et al. (2011). Nutrient co-limitation of primary producer communities. Ecol. Lett. 14, 852–862. doi: 10.1111/j.1461-0248.2011.01651.x
Hart, S. A., and Chen, H. Y. H. (2006). Understory vegetation dynamics of North American boreal forests. Crit. Rev. Plant Sci. 25, 381–397. doi: 10.1080/07352680600819286
Haruka, Y., and Toshiya, Y. (2020). Various scarification treatments produce different regeneration potentials for trees and forbs through changing soil properties. J. For. Res. 25, 41–50. doi: 10.1080/13416979.2019.1706798
He, Y. J., Li, Z. Y., Zhang, P., and Fan, S. H. (2012). Environmental management strategies for global planted forests. World For. Res. 25, 1–7. doi: 10.1007/s11783-011-0280-z
Jensen, A. M., Götmark, F., and Löf, M. (2012). Shrubs protect oak seedlings against ungulate browsing in temperate broadleaved forests of conservation interest: A field experiment. For. Ecol. Manage. 266, 187–193. doi: 10.1016/j.foreco.2011.11.022
Kara, F., and Topaçoğlu, O. (2018). Influence of stand density and canopy structure on the germination and growth of Scots pine (Pinus sylvestris L.) seedlings. Environ. Monit. Assess. 190:749. doi: 10.1007/s10661-018-7129-x
Kremer, K. N., and Bauhus, J. (2020). Drivers of native species regeneration in the process of restoring natural forests from mono-specific, even-aged tree plantations: A quantitative review. Restorat. Ecol. 28, 1074–1086. doi: 10.1111/rec.13247
Li, J., Shi, X. D., Gao, R. M., Li, C., Zhang, Y. Q., and Li, H. Y. (2020). Regeneration and affecting factors of Larix principis-rupprechtii natural secondary forests. J. For. Environ. 40, 588–596. doi: 10.13324/j.cnki.jfcf.2020.06.004
Liang, W. J., and Wei, X. (2020). Factors promoting the natural regeneration of Larix principis-rupprechtii plantation in the Lvliang Mountains of central China. PeerJ 8, e9339. doi: 10.7717/peerj.9339
Liang, W. J., and Wei, X. (2021). Multivariate path analysis of factors influencing Larix principis-rupprechtii plantation regeneration in northern China. Ecol. Indic. 129:107886. doi: 10.1016/j.ecolind.2021.107886
Lin, Y. M., Deng, H. J., Rafay, L., Zhang, G. S., and Fan, C. Z. (2017). Combined effects of climate, restoration measures and slope position in change in soil chemical properties and nutrient loss across lands affected by the Wenchuan Earthquake in China. Sci. Total Environ. 596, 274–283. doi: 10.1016/j.scitotenv.2017.04.034
Liu, J. F., Su, S. J., He, Z. S., Jiang, L., Gu, X. G., Xu, D. W., et al. (2020a). Relationship between Pinus taiwanensis seedling regeneration and the spatial heterogeneity of soil nitrogen in Daiyun Mountain, southeast China. Ecol. Indic. 115:106398. doi: 10.1016/j.ecolind.2020.106398
Liu, R. H., Pan, Y. F., Bao, H., Liang, S. C., Jiang, Y., Tu, H. R., et al. (2020b). Variations in soil physico-chemical properties along slope position gradient in secondary vegetation of the hilly region, Guilin, Southwest China. Sustainability 12:1303. doi: 10.3390/su12041303
Liu, T. Y., Lin, K. C., Vadeboncoeur, M. A., Chen, M. Z., Huang, M. Y., Lin, T. C., et al. (2015). Understorey plant community and light availability in conifer plantations and natural hardwood forests in Taiwan. Appl. Veget. Sci. 18, 591–602. doi: 10.1111/avsc.12178
Liu, Y., Han, S. J., Zhou, Y. M., and Li, X. F. (2007). Soil and root respiration under elevated CO2 concentrations during seedling growth of Pinus sylvestris var, Sylvestriformis. Pedosphere 17, 660–665. doi: 10.1016/S1002-0160(07)60079-4
Ma, Y., Cheng, X. Q., Kang, F. F., and Han, H. R. (2022). Effects of thinning on soil aggregation, organic carbon and labile carbon component distribution in Larix principis-rupprechtii plantations in North China. Ecol. Indic. 139:108873. doi: 10.1016/j.ecolind.2022.108873
Ma, Y. C., Piao, S. L., Sun, Z. Z., Lin, X., Wang, T., Yue, C., et al. (2014). Stand ages regulate the response of soil respiration to temperature in a Larix principis-rupprechtii plantation. Agric For. Meteorol. 184, 179–187. doi: 10.1016/j.agrformet.2013.10.008
Mayer, M., Matthews, B., Rosinger, C., Sanden, H., Godbold, D. L., and Katzensteiner, K. (2017). Tree regeneration retards decomposition in a temperate mountain soil after forest gap disturbance. Soil Biol. Biochem. 115, 490–498. doi: 10.1016/j.soilbio.2017.09.010
Moelder, A., Sennhenn-Reulen, H., Fischer, C., Rumpf, H., Schoenfelder, E., Stockmann, J., et al. (2019). Success factors for high-quality oak forest (Quercus robur, Q. petraea) regeneration. For. Ecosyst. 6:49. doi: 10.1186/s40663-019-0206-y
Moreno-Fernandez, D., Ledo, A., Canellas, I., and Montes, F. (2020). Strategies for modeling regeneration density in relation to distance from adult trees. Forests 11:120. doi: 10.3390/f11010120
Olesinski, J., Lavigne, M. B., and Krasowski, M. J. (2012). Fine-root dynamics change during stand development and in response to thinning in balsam fir (Abies balsamea L. Mill.) forests. For. Ecol. Manage. 286, 48–58. doi: 10.1016/j.foreco.2012.08.047
Perry, D. A., Oren, R., and Hart, S. C. (2008). Soil: The fundamental resource, forest ecosystems. Baltimore: Johns Hopkins University Press.
Pham, V. V., Ammer, C., Annighöfer, P., and Heinrichs, S. (2022). Tree regeneration characteristics in limestone forests of the Cat Ba National Park, Vietnam. BMC Ecol. Evolut. 22:6. doi: 10.1186/s12862-021-01957-9
Picchio, R., Mederski, P. S., and Tavankar, F. (2020). How and how much, do harvesting activities affect forest soil, regeneration and stands? Curr. For. Rep. 6, 115–128. doi: 10.1007/s40725-020-00113-8
Pielou, E. C. (1966). The measurement of diversity in different types of biological collections. J. Theoret. Biol. 13, 131–144.
R Core Team (2008). R: A language and environment for statistical computing, Vol. 1, 2010. Vienna: R Foundation for Statistical Computing. Available online at: https://www.R-project.org/
Randriambanona, H., Randriamalala, J. R., and Carriere, S. M. (2019). Native forest regeneration and vegetation dynamics in non-native Pinus patula tree plantations in Madagascar. For. Ecol. Manage. 446, 20–28.
Sagra, J., Ferrandis, P., Plaza-Alvarez, P. A., Lucas-Borja, M. E., Gonzalez-Romero, J., Alfaro-Sanchez, R., et al. (2018). Regeneration of Pinus pinaster Aiton after prescribed fires: Response to burn timing and biogeographical seed provenance across a climatic gradient. Sci. Total Environ. 637, 1550–1558. doi: 10.1016/j.scitotenv.2018.05.138
Shannon, C., and Wiener, W. (1949). The mathematical theory of communication. Chicago, IL: University of Illinois press.
Sharma, L. N., Shrestha, K. B., and Maren, I. E. (2019). Tree regeneration in gap-understory mosaics in a subtropical Shorea robusta (Sal) forest. J. For. Res. 30, 2061–2068. doi: 10.1007/s11676-018-0747-x
Shen, C. C., and Nelson, A. S. (2018). Natural conifer regeneration patterns in temperate forests across the Inland Northwest, USA. Ann. For. Sci. 75:54. doi: 10.1007/s13595-018-0724-8
Silva, A. D., de Resende, A. S., Chaer, G. M., Goncalves, F. L. A., and Campello, E. F. C. (2018). Hill slope form and position influence in soil characteristics and natural regeneration of abandoned pastures areas. Cienc. Florest. 28, 1239–1252. doi: 10.5902/1980509833366
Sklodowski, J. (2020). Two directions of regeneration of post-windthrow pine stands depend on the composition of the undergrowth and the soil environment. For. Ecol. Manage. 461:117950. doi: 10.1016/j.foreco.2020.117950
Soto, D. P., Jacobs, D. F., Salas, C., Donoso, P. J., Fuentes, C., and Puettmann, K. J. (2017). Light and 657 nitrogen interact to influence regeneration in old-growth Nothofagus-dominated forests in 658 south-central Chile. For. Ecol. Manage. 384, 303–313. doi: 10.1016/j.foreco.2016.11.016
Soto, D. P., Puettmann, K. J., Fuentes, C., and Jacobs, D. F. (2019). Regeneration niches in Nothofagus-dominated old-growth forests after partial disturbance: Insights to overcome arrested succession. For. Ecol. Manage. 445, 26–36. doi: 10.1016/j.foreco.2019.05.004
Teketay, D., Kashe, K., Madome, J., Kabelo, M., Neelo, J., Mmusi, M., et al. (2018). Enhancement of diversity, stand structure and regeneration of woody species through area exclosure: The case of a Mopane woodland in northern Botswana. Ecol. Process. 7:5. doi: 10.1186/s13717-018-0116-x
Thrippleton, T., Bugmann, H., Kramer-Priewasser, K., and Snel, L. R. S. (2016). Herbaceous understorey: An overlooked player in forest landscape dynamics? Ecosystems 19, 1240–1254. doi: 10.1007/s10021-016-9999-5
Tingstad, L., Olsen, S. L., Klanderud, K., Vandvik, V., and Ohlson, M. (2015). Temperature, precipitation and biotic interactions as determinants of tree seedling recruitment across the tree line ecotone. Oecologia 179, 599–608. doi: 10.1007/s00442-015-3360-0
Tiscar, P. A., Candel-Perez, D., Estrany, J., Balandier, P., Gomez, R., and Lucas-Borja, M. E. (2017). Regeneration of three pine species in a Mediterranean forest: A study to test predictions from species distribution models under changing climates. Sci. Total Environ. 584, 78–87. doi: 10.1016/j.scitotenv.2017.01.150
Tsujino, R., and Yumoto, T. (2008). Seedling establishment of five evergreen tree species in relation to topography, sika deer (Cervus nippon yakushimae) and soil surface environments. J. Plant Res. 121, 537–546. doi: 10.1007/s10265-008-0184-6
Valladares, F., Martinez-Ferri, E., Balaguer, L., Perez-Corona, E., and Manrique, E. (2000). Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: A conservative resource-use strategy? N. Phytol. 148, 79–91. doi: 10.1046/j.1469-8137.2000.00737.x
Venanzi, R., Picchio, R., Grigolato, S., and Latterini, F. (2019). Soil and forest regeneration after different extraction methods in coppice forests. For. Ecol. Manage. 454:117666. doi: 10.1016/j.foreco.2019.117666
Walters, M. B., Farinosi, E. J., Willis, J. L., and Gottschalk, K. W. (2016). Managing for diversity: Harvest gap size drives complex light, vegetation, and deer herbivory impacts on tree seedlings. Ecosphere 7:e01397. doi: 10.1002/ecs2.1397
Wang, Y. B., Wang, Y. H., Xiong, W., Yao, Y. Q., Zhang, T., and Li, Z. H. (2017). Variation in the sap flow velocity of Larix principis-rupprechtii and its impact factors in different slope positions in a semi-arid region of Liupan mountains. Sci. Silvae Sinic. 53, 19–20. doi: 10.11707/j.1001-7488.20170602
Xu, X. L., Wang, X. J., Cleary, M., Wang, P., Lu, N. N., Sun, Y. J., et al. (2020). Slope position rather than thinning intensity affects arbuscular mycorrhizal fungi (AMF) community in Chinese fir plantations. Forests 11:273. doi: 10.3390/f11030273
Yang, X. Q., Han, Y. Z., Li, L., Chen, X., and You, J. (2009). The effect of heterogeneous spatial distribution of soil nitrogen on regeneration of Larix principis -rupprechtii seedlings in typical naturally -regenerated montane forests of Northern China. Acta Ecol. Sin. 29, 4656–4664. doi: 10.1007/978-1-4020-9623-5_5
Yang, X. Q., Zhang, P., Wu, Q. T., Yan, H. B., Yu, M. K., Wang, G., et al. (2017). Effects of simulated wind load on leaf photosynthesis and carbohydrate allocation in eight Quercus Species. J. Biobased Mater. Bioener. 11, 652–657. doi: 10.1166/jbmb.2017.1721
Ye, S. L., and Lei, G. Y. (2019). Rules of changes in soil nutrients and enzyme activities of Larix principis-rupprechtii in different forest ages. Asian Agric. Res. 11, 54–58. doi: 10.19601/j.cnki.issn1943-9903.2019.4.011
Yu, F., Wang, D. X., Shi, X. X., Yi, X. F., Huang, Q. P., and Hu, Y. N. (2013). Effects of environmental factors on tree seedling regeneration in a pine-oak mixed forest in the Qinling Mountains, China. J. Mount. Sci. 10, 845–853. doi: 10.1007/s11629-013-2548-1
Zadworny, M., Mucha, J., Jagodzinski, A. M., Koscielniak, P., Lakomy, P., Modrzejewski, M., et al. (2021). Seedling regeneration techniques affect root systems and the response of Quercus robur seedlings to water shortages. For. Ecol. Manage. 479:118552. doi: 10.1016/j.foreco.2020.118552
Zhao, X. L., Li, J. J., and Li, H. J. (2010). Effects of vegetation restoration type on soil carbon, nitrogen, and microbial quantity in Guandi Mountain. Chin. J. Ecol. 29, 2102–2110. doi: 10.13292/j.1000-4890.2010.0341
Appendix

Appendix Table 1. The parameters of the violin diagrams.
Keywords: tree regeneration, tree density, herb diversity, soil property, Larix principis-rupprechtii
Citation: Liang W, Feng Y, Li F, Zhao Y, Zhu J, Yao J, Chang C and Wei X (2023) Drivers of vegetation and soil determine natural regeneration of a single plantation at different slope positions. Front. For. Glob. Change 6:1167284. doi: 10.3389/ffgc.2023.1167284
Received: 16 February 2023; Accepted: 03 August 2023;
Published: 23 August 2023.
Edited by:
Pablo Donoso, Austral University of Chile, ChileReviewed by:
María Florencia Urretavizcaya, National Scientific and Technical Research Council (CONICET), ArgentinaDaniel Soto, Universidad de Aysén, Chile
Copyright © 2023 Liang, Feng, Li, Zhao, Zhu, Yao, Chang and Wei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xi Wei, d2VpeGk4NjA4MjZAMTYzLmNvbQ==