
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change , 01 July 2022
Sec. Forest Ecophysiology
Volume 5 - 2022 | https://doi.org/10.3389/ffgc.2022.914199
 Arivoara Rabarijaona1
Arivoara Rabarijaona1 Stéphane Ponton1*
Stéphane Ponton1* Didier Bert2Alexis Ducousso2Béatrice Richard1
Didier Bert2Alexis Ducousso2Béatrice Richard1 Joseph Levillain1
Joseph Levillain1 Oliver Brendel1
Oliver Brendel1Context: As a widespread species, sessile oak (Quercus petraea) populations occupy a wide range of ecological conditions, with large gradients of soil water availability. Drought acclimation involves a plastic increase in water-use efficiency (WUE), a trait that is easily measured using the carbon isotope composition (δ13C). However, the question remains whether WUE is an adaptive trait that impacts the fitness of trees in natural environments.
Objectives and Methods: To investigate whether WUE was a drought-adaptive trait, we studied a sample of 600 trees originating from 16 provenances, grown for 21 years in a common garden. Intrinsic WUE (WUEi), estimated from tree ring δ13C, was compared among and within populations for three climatically contrasted years. The adaptive character of WUEi was evaluated by relating population mean WUEi, as well as its plasticity to drought, to the pedoclimatic conditions of their provenance sites. The contribution of WUEi to tree and population fitness was finally assessed from the relationship between WUEi and tree radial growth (GI).
Results: Significant differences in WUEi were found among populations but a much larger variability was observed within than among populations. The population WUEi of the juvenile oak trees growing in the relatively mesic conditions of the common garden showed no relationship with a modeled water deficit index for the provenance sites. However, a higher population WUEi plasticity to severe drought was related to a higher proportion of silt and carbon and a lower proportion of sand in the soil of the provenance sites. In response to severe drought, populations with a higher increase in WUEi showed a lower decrease in GI. Populations with lower GI reduction were from sites with higher vapor pressure deficit in May–July (VPD). For the wet year only, populations with a higher WUEi also had a higher GI.
Conclusion: The correlations observed at the common garden site between (i) population means of WUEi plasticity to drought and soil texture of the provenance sites, and (ii) GI plasticity to drought and VPD, suggested a local adaptation of sessile oak.
Drought-induced diebacks of forest trees are commonly observed and affect nearly every forest biome in all parts of the world (Hartmann et al., 2018b). Extreme drought events are expected to be more frequent and intensive in the forthcoming decades due to climate change (Spinoni et al., 2018), and those observed between 2015 and 2018 in Europe have no equivalent in the past two millennia (Büntgen et al., 2021). In Europe, the loss of forest area due to drought-induced mortality has been estimated at 500,000 ha between 1987 and 2016 (Senf et al., 2020). Many forest tree species are affected, especially when they grow under limiting environmental conditions (Allen et al., 2010; Urli et al., 2015). Although considered as rather drought-resistant (Friedrichs et al., 2009; Härdtle et al., 2013; Kunz et al., 2018; Perkins et al., 2018), sessile oak [Quercus petraea (Mattuschka) Lieblein, 1784] already experienced recent diebacks in the most continental part of its distribution area (Petritan et al., 2021) and is expected to suffer future declines on sites with warmer and drier conditions (Bert et al., 2020).
Over the past millennia, migration allowed white oak species, as many others, to respond to climate changes caused by the glacial-interglacial cycles (Kremer et al., 2002; Petit et al., 2002; Cheddadi et al., 2005), with a mean migration rate estimated at 1.5 km per year for the last glacial migration (Brewer et al., 2005). However, climate change models predict an unprecedentedly rapid temperature change over the next 100 years, which would require a much faster migration pace. Assisted migration (Aitken and Bemmels, 2016; Sáenz-Romero et al., 2016) could accelerate range shifts through human intervention; however, this requires detailed information about genetic adaptive variation within the species. As a widespread species, sessile oak populations occupy a wide range of ecological conditions with different local selection pressures favoring local adaptation and resulting in genetic differentiation, as observed by Kremer and Petit (1993). Such genetic differentiation can include genetic diversity for phenotypic plasticity, which contributes to responses to short-term changes in environmental conditions. Higher phenotypic plasticity is expected in tree populations from sites subjected to more variable conditions over time, and especially to larger amplitudes of pedoclimatic conditions (Aitken et al., 2008).
Species with a large diversity in functional traits show a high capacity to adapt and/or acclimate to increasing aridity (Anderegg et al., 2021). The adaptive character of traits is commonly established by linking the among-population differences in traits observed in a common garden with the difference in climate or geographical characteristics of each population (Wang et al., 2010; Aitken and Bemmels, 2016; Sáenz-Romero et al., 2019; Bert et al., 2021). For sessile oak, significant among-population differentiation was observed for radial growth (Bert et al., 2020), height and survival (Grotehusmann and Schönfelder, 2011; Sáenz-Romero et al., 2017; Mátyás, 2021), and leaf functional traits (Vitasse et al., 2009a; Bruschi, 2010; Bresson et al., 2011; Torres-Ruiz et al., 2019). Leaf flushing and senescence dates were earlier for sessile oak populations originating from warmer sites (Vitasse et al., 2009b). Bruschi (2010) showed that the temperature and precipitation of the provenance sites contributed to the among-population variation in sessile oak height, root–shoot ratio, leaf size, and thickness. However, other studies reported no significant differentiation among sessile oak populations, established along an aridity gradient, for leaf unfolding and senescence (Vitasse et al., 2010), leaf area, carbon and nitrogen isotope discrimination, leaf C and N content, stomatal density, xylem pressure inducing 50% loss of hydraulic conductivity due to embolism, vein density and wood density (Torres-Ruiz et al., 2019). These discrepancies illustrate the complex relationship between traits observed in a common garden and provenance site conditions, partly due to the much lower diversity observed among than within sessile oak populations (Bresson et al., 2011), which has also been observed for many other tree species (Hamrick et al., 1992; Ramirez-Valiente et al., 2010; Aranda et al., 2017).
Water-use efficiency (WUE) is a functional trait expressing at the tree level the trade-off between biomass production and water loss by transpiration. Because WUE reflects the link between tree carbon assimilation and water loss, studying among-population variation in WUE allows characterizing tree’s strategies to face drought events (Brendel and Epron, 2022). The corresponding trait at the leaf level is called intrinsic water-use efficiency (WUEi), i.e., the ratio between net CO2 assimilation (A) and stomatal conductance for water vapor (gsw). WUEi is linked to the carbon isotope composition (δ13C, ‰) of the photosynthesis products (Farquhar and Richards, 1984). WUEi and δ13C are positively related, i.e., when WUEi increases, δ13C values become less negative. Although post-photosynthetic fractionation processes have been identified between leaf carbon assimilates and wood (Gessler et al., 2014), tree-ring δ13C is considered as a valid estimator of foliar WUEi integrated over the growing season and at the tree level (Ponton et al., 2002; Roussel et al., 2009). To our knowledge, the only study assessing variation among populations of sessile oak did not detect any significant differences in δ13C measured in leaves sampled in mid-June (Torres-Ruiz et al., 2019), whereas significant differences were observed for many other forest tree species (Zhang and Marshall, 1995; Ferrio et al., 2003; Aranda et al., 2010). Several studies on broadleaved and conifer species observed a negative correlation between populations WUEi (directly calculated from gas exchange measurements or estimated with δ13C, or both) and precipitation at their provenance sites (Li et al., 2000; Cregg and Zhang, 2001; Bekessy et al., 2002; Zhang et al., 2005; Duan et al., 2009), suggesting higher WUEi for populations from drier sites. Conversely, a lower WUEi for populations from the drier sites was observed in conifer species (Aitken et al., 1995; Guehl et al., 1996; Nguyen-Queyrens et al., 1998; Fan et al., 2008) but also in Castanea sativa (Lauteri et al., 2004; Eriksson et al., 2005) and Quercus ilex (Ferrio et al., 2003). These discrepancies among observations might suggest that there is no simple relationship between adaptation to dry environments and WUE. However, as suggested by Brendel and Epron (2022), the relationship between population mean WUE and pedoclimatic characteristics of provenance sites could be an indicator of different drought adaptation strategies. Further, most relationships between population mean WUE and site conditions are based on meteorological data only, whereas the soil water availability depends strongly on the soil characteristics, which should be the main selection force for adaptation to soil drought.
Plant fitness is associated with several traits linked to reproduction and survival, including leaf phenology and vertical and radial growth (Gárate-Escamilla et al., 2019). Therefore, tree fitness could be linked to WUEi by associating it with radial growth. The direction of this relationship allows some inferences about its underlying cause. A variation in photosynthetic capacity among populations would affect both their WUEi and productivity in the same direction, i.e., an increase in photosynthetic capacity could increase WUEi, resulting in less negative values of δ13C, and could also result in an increase in radial growth, at least under optimal conditions of water availability, light intensity, CO2 concentration, and temperature. Thus, a variation in photosynthetic capacity would result in a positive correlation between WUEi and growth. In contrast, a variation in stomatal conductance would result in an inverse relationship between WUEi and radial growth: a lower stomatal conductance increases WUEi (and results in more negative values of δ13C) but would either decrease growth (because of stomatal limitations of A) or result in no relationship. Summarizing studies about intra-specific association between WUEi and tree productivity, Fardusi et al. (2016) showed that, for shrubs and conifers, WUEi is highly and positively correlated to total biomass production (tested only on seedlings) and tree height but only slightly correlated to tree diameter. However, no clear relationships were observed for broadleaved tree species.
Using δ13C measured in tree rings as a proxy of WUEi, this study aimed at testing whether WUEi or its plasticity (based on WUEi from wet and dry years) could be considered an adaptive trait for environments with prolonged summer droughts. For this purpose, young sessile oak trees originating from 16 populations over Europe and planted in a common garden (Sillegny, France) were examined. In addition, in order to improve the environmental characterization of the original environments, each population was visited to analyze the physico-chemical characteristics of the soil and model the water balance of each population.
• (1) We aimed to characterize the among-population diversity of WUEi for sessile oak and to link it with the pedoclimatic conditions of the provenance sites. We hypothesized, that, in the common garden of Sillegny, where the pedoclimatic conditions are in average mesic, populations from the driest sites would display the lowest WUEi, as observed for Q. ilex (Ferrio et al., 2003).
• (2) We also sought to assess the plasticity of WUEi to different soil water conditions (selected years) and to identify any plasticity difference among populations. We hypothesized that populations from dry sites, where annual environmental conditions are more variable, would exhibit larger plasticity in WUEi than populations from wetter sites, as observed for Q. suber (Matías et al., 2019).
• (3) And finally, we wanted to assess the relationship between WUEi and growth as an estimator of fitness. We hypothesized that trees and populations with higher WUEi showed higher radial growth only when the soil water availability was unlimited.
Open-pollinated seeds were collected in 1989 from one population in each of 64 populations across the distribution area of sessile oak [Quercus petraea (Mattuschka) Lieblein, 1784] (Sáenz-Romero et al., 2017; Torres-Ruiz et al., 2019; Bert et al., 2020). After sowing and raising in a nursery, 3-year-old seedlings were planted in March 1993 in a common garden in Sillegny, France (lat. 48° 59’ 24” N, long. 6° 7’ 56” E, alt. 201 m). A complete randomized block design was used to account for within-site variation, and incomplete micro-blocks were nested in the blocks. Each micro-block consisted of eight plots of 24 oaks from the same population planted 1.75 m × 3 m apart (initial stand density: 1905 trees.ha–1). Survival, height, and diameter at breast height were measured four times between 1993 and 2015. In 2015, the first stand thinning was realized during which trees from 16 among the 64 populations were sampled. The populations were selected according to two criteria: (i) maximizing the variability of pedoclimatic conditions among population sites (Figure 1 and Supplementary Table 1), and (ii) maximizing the variability of growth performances (height and diameter) among transplanted populations at Sillegny. In addition, because genetic variability has been completely reshaped by the last postglacial recolonization, in particular by local selection pressures (Kremer, 2000), the choice of the selected populations was restricted to those belonging to a unique lineage originating from an Iberian refugium (Petit et al., 2002).
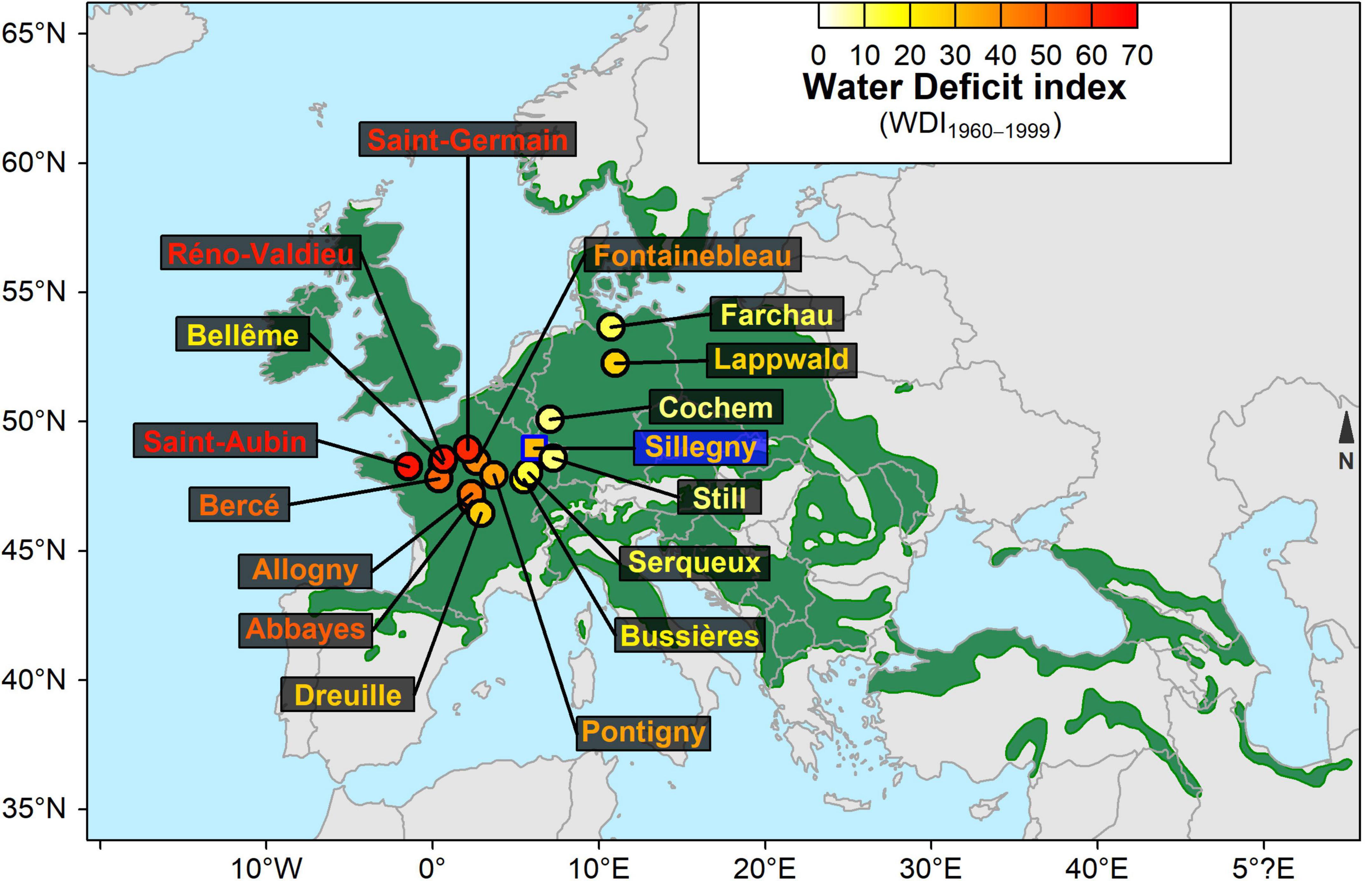
Figure 1. Distribution map of the 16 provenance sites of sessile oak (Quercus petraea). The green area represents the current distribution area of sessile oak in Europe (Caudullo et al., 2017). A square indicates the common garden location (Sillegny). The color associated with each site refers to the mean value of water deficit index calculated for the period 1960 – 1999 (WDI1960–1999). The Abbayes population is also called Soudrain in some databases.
Annual soil moisture in the 16 provenance sites and in the common garden was quantified by Water Deficit Index (WDI, dimensionless) estimated by modeling daily water balance with BILJOU© (Granier et al., 19991). WDI integrates the precocity, the duration, and the intensity of the annual soil water deficit, which is considered to occur when the daily soil relative extractable water (REW) drops below a threshold of 40% (Granier et al., 1999). Daily meteorological data, soil properties, and forest stand characteristics (i.e., evaporative surface) are needed to run BILJOU©.
Each provenance site was visited in 2016–2017 in order to collect soil for a detailed analysis. For both common garden and provenance sites, the soil profile and the rooting depth were described. At least three soil sampling points were taken to verify soil plot homogeneity. Soil samples from each sampling point were then grouped by soil horizon for analysis at the LAS laboratory (INRAE). Particle size distribution analysis (NF X31-107) and organic carbon (ISO 14235) were measured to calculate the plant-available water capacity (PAWC) of the soil using the pedotransfer function of Al Majou et al. (2007). Bulk density was also calculated from organic carbon measurements using the pedotransfer function of Manrique and Jones (1991). In addition, pH water and pH KCl (ISO 10390), Cation Exchange Capacity (CEC, ISO 23470), and nitrogen (ISO 13878) were measured to characterize differences in soil fertility among sites.
The different horizon depths and thicknesses were harmonized by applying a spline approach (Bishop et al., 1999) to obtain a topsoil (0–30 cm) and a subsoil (>30 cm-rooting depth) layers. Splines were fitted to the data using the mpspline function in the GSIF package in R (Hengl et al., 2017). As previously described in Jonard et al. (2017), stocks of organic carbon and nitrogen of topsoil and subsoil were calculated by taking into account dry bulk density, concentrations of organic carbon and nitrogen, the thickness of this layer, and the volume of coarse element (>2 mm).
The daily meteorological data (wind speed at 2 m, precipitation, average temperature, relative humidity, and sunshine duration) were obtained by requesting the SAFRAN meteorological analysis system for French sites (Quintana-Seguí et al., 2008) and Deutscher Wetterdienst (DWD2) for German sites, from the 1st January 1960 to the 31th December 1999. A maximum LAI (Leaf Index Area) of 6 m2 of foliage per m2 of soil was set for each stand based on visual evaluation in the common garden site and estimation in the provenance sites. For each site, the average days of budburst and leaf fall were estimated with the CASTANEA model (Dufrêne et al., 2005) and were inputted into BILJOU© to restrict the calculation of WDI to the leafy period. For each year from 1960, an annual WDI value was modeled, and the average over the 1960–1999 period (WDI1960–1999) was used to characterize the long-term mean pedoclimate of each site. Saint-Aubin (WDI1960–1999 = 65.75) was the driest site whereas Cochem (WDI1960–1999 = 8.90) was the wettest one (Supplementary Table 1). The drier sites (i.e., high WDI1960–1999 values) appeared to be also those with the largest range of interannual variations in WDI (Supplementary Figure 1). In order to compare with other studies, two other drought indices were also used to characterize the sites (Supplementary Table 1): aridity index (AI) and summer heat:moisture index (SHM). Both require only meteorological data. AI was calculated as the ratio between mean annual precipitation and mean annual potential evapotranspiration (Torres-Ruiz et al., 2019), and SHM as the ratio between the mean temperature of the warmest month and the mean summer precipitation (May to September) divided by 1,000 (George et al., 2020). In our study, WDI1960–1999 appeared only slightly correlated with AI and SHM (R2 = 0.373, p = 0.012 and R2 = 0.378, p = 0.112, respectively; n = 16) while AI and SHM were highly correlated (R2 = 0.804, p < 0.0001, n = 16).
In order to evaluate tree plasticity of WUEi in response to water availability, three tree rings corresponding to years with contrasted WDI were analyzed. In the common garden of Sillegny, the canopy started to close after 2007, resulting in a declining trend in radial growth for all trees. As competition for light would strongly affect δ13C (Stokes et al., 2010), the selection of tree rings was restricted to years before 2007. Based on WDI in Sillegny, the years 2000, 2003, and 2005 were selected (Figure 2). In contrast with the wet 2000 year (WDI = 2.7), 2003 was a severe dry year, with a long and strong drought period (130 days) resulting in an annual WDI of 68.7, while 2005 appeared moderately dry with a drought period of similar length (135 days) but less intense (WDI = 49.4).
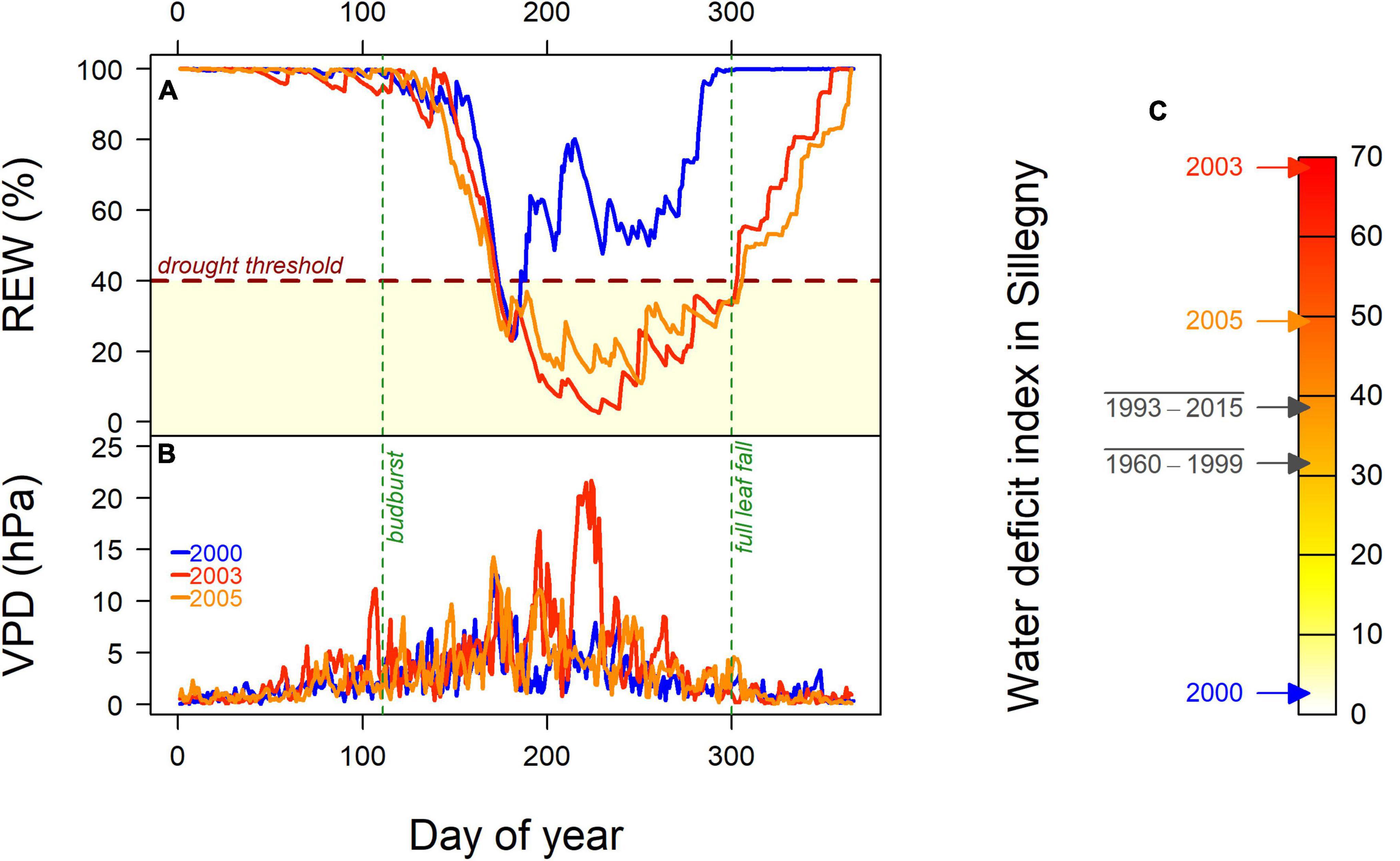
Figure 2. (A) Course of the daily soil Relative Extractable Water (REW, %) calculated by BILJOU© and (B) daily Vapor Pressure Deficit (VPD, hPa) in the common garden of Sillegny in 2000, 2003, and 2005. The red horizontal dashed line represents the drought threshold at 40% of REW. The date of budburst (DoY 111) and full leaf fall (DoY 300) are indicated with dotted vertical green. The annual Water Deficit Index (WDI), which cumulated daily values from budburst to leaf fall, is shown (C) for the three selected years and for the periods 1993–2015 (i.e., the period of tree growth in the common garden) and 1960–1999 (to be compared to the values calculated for the provenance sites; see Supplementary Table 1).
The sampling was done during the winter of 2014–2015 when the first thinning operation was planned to remove one in two trees within the common garden plots. Stem disks were collected from a selection of these felled trees. For each population, 30 trees were selected among the different plots, except for four populations (Pontigny, Allogny, Bussières, and Serqueux) where 60 trees were selected to better characterize the diversity within populations (Supplementary Table 2). The selection focused primarily on the tree social status, visually assessed before sampling, in 2014 (thus after canopy closure), favoring when possible dominant trees to avoid additional microclimate variability which could affect δ13C (McDowell et al., 2011) and growth (Trouvé et al., 2015; Alfaro-Sánchez et al., 2020). However, the constraints of systematic thinning forced the selection of some codominant (37%) and suppressed trees (14%) in order to reach the required sample size for each population (Supplementary Table 2). Finally, 595 trees belonging to 16 populations were sampled from 148 single populations plots dispersed over 66 micro-blocks, nested within four large blocks (Supplementary Figure 2).
From each stem disk, two opposing laths were cut and shaped with a double-bladed saw. Each lath was used for wood micro-density, radial growth, and carbon isotope composition (δ13C) measurements. Wood micro-density profile was first obtained by exposing the 2 mm-thick laths to X-rays. Ring widths (RW, mm) were measured manually from the obtained wood micro-density profile. The RW series were cross-dated with specific pointer years (i.e., narrow rings in 1995, 1998, and 2003 and wide rings in 2004 and 2007). For a complete description, see Bert et al. (2020). In order to remove any growth trends (mainly due to ontogenic and stand characteristics) and only keep the inter-annual variations, each individual rings series was detrended with a cubic smoothing spline (with a wavelength cutoff of 0.5 and a rigidity of 20 years). The remaining first-order auto-correlation was removed with dplR package (Bunn, 2008) to obtain a detrended, dimensionless radial growth index (GI, dimensionless), zero-centered, and normally distributed. In short, RW was the preferred variable for comparison among populations and social status, and GI was retained when comparing years.
Tree rings corresponding to the years 2000, 2003, and 2005 were cut with a scalpel under a binocular and were milled 1.5 min at 30 Hz in a ball mill (MM200, Retsch GmbH, Haan, Germany). Subsamples of 1 ± 0.2 mg were measured with a continuous-flow elemental analyzer (vario ISOTOPE cube, Elementar, Hanau, Germany) coupled with an isotope ratio mass spectrometer (IsoPrime 100, Isoprime Ltd, Cheadle, United Kingdom). Carbon isotope composition is expressed in delta notation (δ13C, ‰) relative to the Pee Dee Belemnite standard. The precision of the δ13C measurements, calculated as the standard deviation of 234 repetitions of a laboratory-working standard, was 0.11‰.
In tree-ring isotopes studies, analysis on cellulose was sometimes preferred as compared to whole wood, whose varying composition of cellulose and lignin could affect the δ13C signal (Weigt et al., 2015). In order to evaluate this potential bias, a preliminary test was run on a subsample of 10 individuals selected for a large range of wood micro-density (as a surrogate for lignin content), including three measurements per tree (i.e., rings 2000, 2003, and 2005). Alpha-cellulose was extracted from whole wood using a variant of the Jayme/Wise methods, as described in Richard et al. (2014), which uses successive steps: solvents and boiled water to remove extractives (extractives-free wood), then acidified sodium chlorite to digest the lignin, and finally alkaline solution to remove the hemicelluloses and produce the “analytical α-cellulose.” The lignin content was carried out on dry extractives-free wood sample and determined by a gravimetric analysis of 72% sulfuric acid-insoluble lignin (Klason lignin), according to the ASTM D1106-96(2007), Standard Test Method for Acid-Insoluble Lignin in Wood (ASTM International, West Conshohocken, PA, 2007, United States3). δ13C was measured on whole wood, α-cellulose and lignin. Compared to α-cellulose, an average depletion in 13C of 0.83‰ and 2.77‰ were observed in whole wood and Klason lignin, respectively. However, differences between (i) wood and α-cellulose and (ii) lignin and α-cellulose were constant (i.e., the slope of their relationship was not significantly different from 1: p = 0.730 and p = 0.975, respectively) and not significantly related to the lignin content of the samples (Supplementary Figure 3). Accordingly, we decided to perform the δ13C analyses on whole wood.
For both δ13C and GI, a plasticity index was calculated to quantify the change in δ13C and GI between the wet year and the moderate or severe dry years. The difference of δ13C (GI) values between 2005 and 2000, hereafter named δ13C2005–2000 (GI2005–2000), characterized the plasticity of δ13C (GI) to moderate drought, whereas the plasticity of δ13C (GI) to severe drought was calculated as the difference between δ13C (GI) values in 2003 and 2000, hereafter named δ13C2003–2000 (GI2003–2000). The term “drought intensity,” used in this paper, refers to these two responses (to moderate and severe drought compared to the control year). The larger the plasticity index of δ13C or of GI, the more δ13C increased or GI decreased in response to the drought compared to the wet year. It is then expected that the plasticity index of δ13C in response to drought will be positive, whereas the plasticity index of GI will be negative.
All statistical analyses were performed with R-environment (R Core Team, 20224). Variations in trait expressions and plasticity were tested with a linear mixed model using the nlme package (Pinheiro et al., 2020). The interactions between factors were first included in the model, which was later simplified by minimizing AIC (Akaike Information Criteria) to select the most parsimonious model:
With:
• Yijklm = observed trait or trait plasticity index of the tree m from the population i, block k, and micro-block l for the year j or in response to the drought intensity j compared to the control year (2000);
• μ = overall mean;
• Pi = population effect of the ith population;
• Tj = effect of the jth year (2000, 2003, 2005) or of the jth pair (2000–2003, 2000–2005);
• (Pi*Tj) = effect of the interaction between population and year or between population and drought intensity;
• Sm = social status in 2014 effect of the mth tree;
• Bk = block random effect of the kth block;
• Mkl = micro-block random effect at the lth micro-block nested within the kth block;
• Iklm = tree random effect of the mth tree nested in the lth micro-block nested within the kth block;
• εijklm = error term.
The contribution of fixed and random factors in the model was, respectively, evaluated by marginal and conditional r-squared (Nakagawa et al., 2017), computed with the MuMIn package (Barton, 2020). Because the design of this study is unbalanced (i.e., different proportions of dominant, co-dominant, and suppressed trees among populations), the Tukey post-hoc test was done on the estimated marginal means (Searle et al., 1980) obtained by using the emmeans package. All mean values of traits and plasticity indices hereafter are therefore estimated marginal means (emmeans). These marginal mean values of traits and plasticity indices per population were correlated with the average WDI1960–1999 and pedoclimatic parameters of provenance sites over the period 1960–1999 (summarized in Supplementary Table 1). The correlations between δ13C and growth index and between their plasticity indices were tested at both population and individual levels (for each population separately).
Overall, the social status had a significant effect on both δ13C and RW (Table 1), with no significant interaction with the year (not shown). The dominant trees displayed a significantly 0.21‰ less negative δ13C than the codominant trees that were not significantly different from suppressed trees (Table 2). The dominant trees also had a higher RW than the codominant trees, which in turn had significantly higher values than the suppressed trees. In contrast, δ13C and GI plasticity indices (i.e., the variation of trait expression between dry and wet years) were not significantly affected by social status (Table 1).
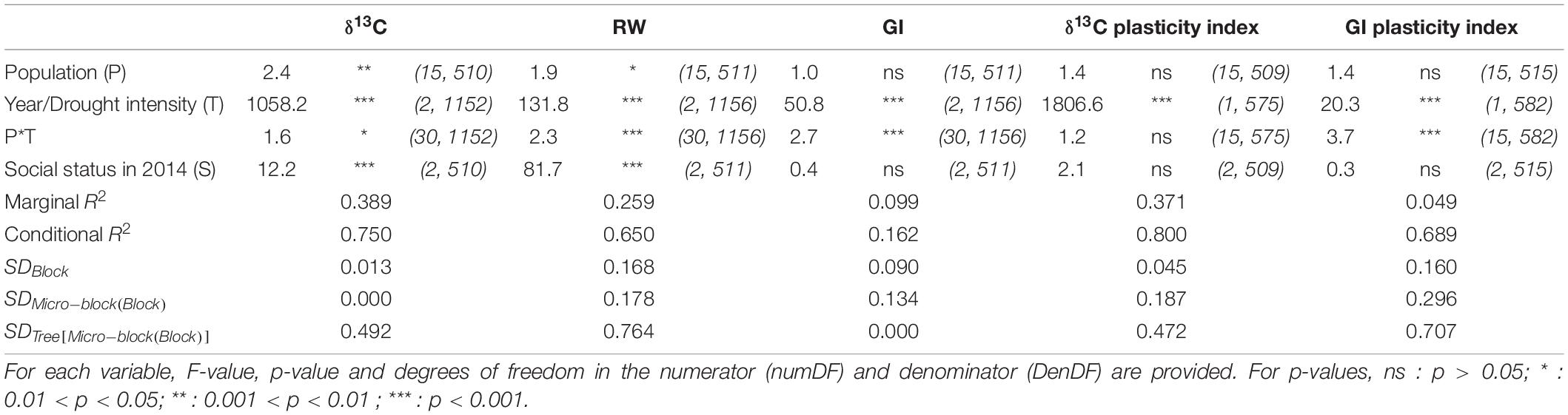
Table 1. Variable significance and model performance for δ13C, ring width (RW), growth index (GI), δ13C plasticity index, and GI plasticity index.
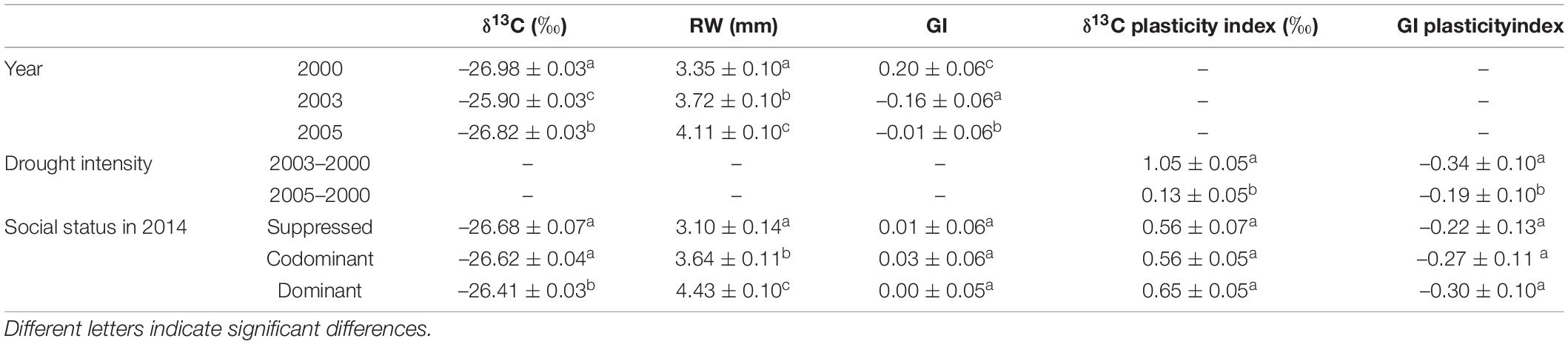
Table 2. Estimated marginal means (emmeans ± SE) and Tukey test group by year and by social status in 2014 for δ13C, ring width (RW), and growth index (GI); by drought intensity and by social status in 2014 for the plasticity indices of δ13C and GI.
The year effect was significant for δ13C (Table 1). The least negative δ13C values were observed in 2003, corresponding to the driest year (WDI = 68.7; Figure 2), and the most negative values in 2000, when WDI was the lowest, i.e., 2.7 (with an average difference of 1.08‰; Table 2). The moderately dry year 2005 (WDI = 49.4) led to δ13C values closer to those of 2000 (+0.16‰), although significantly different (Table 2). The plasticity index of δ13C was significantly higher when the drought intensity was severe than moderate (Table 2).
The year effect also influenced RW (Table 1), whose values showed an overall increase until 2007 (Table 2 and Supplementary Figure 4). GI, which by construction showed interannual variations from which longer-term trends were removed, showed the lowest mean value in 2003 and the highest in 2000 (with a difference of 0.36; Table 2). In 2005, GI mean was just slightly negative (–0.01; Table 2) and significantly different from 2003 and 2000. The plasticity index of GI was significantly higher when the drought intensity was severe than moderate (Table 2).
The population effect on δ13C was significant, although moderate, and there was only a weak evidence for an effect of the interaction with the year (p = 0.024; Table 1). In 2000, when the water deficit was almost non-existent, the trees from Fontainebleau displayed significantly less negative δ13C than those from Dreuille and Farchau (with a difference of 0.61–0.63‰; Supplementary Table 3). In 2003, during the driest studied year, only the trees from Farchau had significantly more negative δ13C than those from Pontigny and Bercé (a difference of 0.58–0.62‰; Supplementary Table 3 and Figure 3). In the moderately dry year 2005, the trees from Saint-Germain and Farchau displayed significantly more negative δ13C values than those from Still (a difference of 0.61–0.64‰; Supplementary Table 3). Overall, Farchau showed consistently the most negative δ13C. The δ13C values observed in 2003 were strongly and positively correlated with those observed in 2005 (R = 0.84; Figure 4) whereas correlations between 2000 values and 2003 and 2005 appeared lower, although significant (R = 0.68 and R = 0.55, respectively).
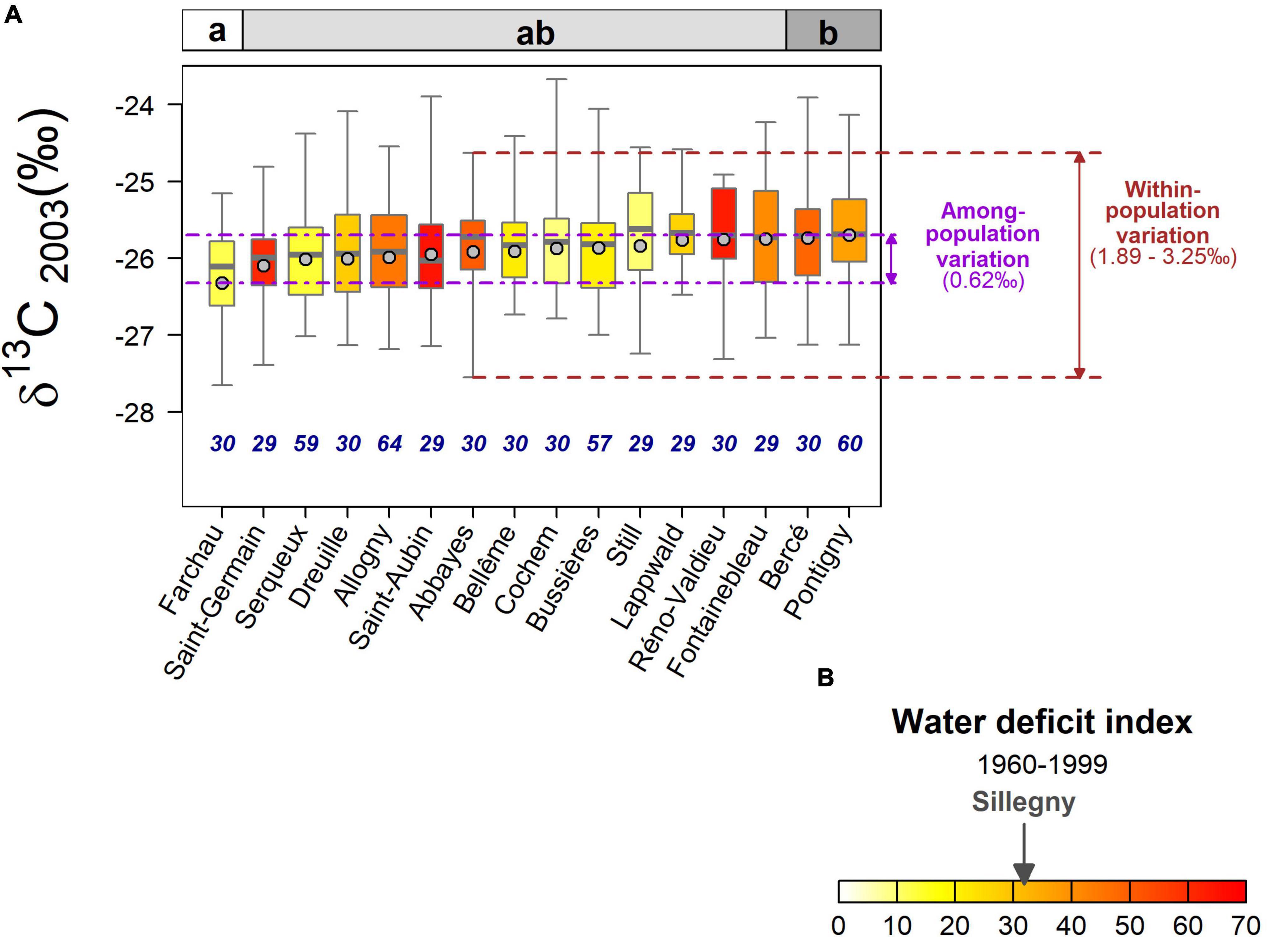
Figure 3. (A) Within- and among-population variations of the 2003 tree ring δ13C (‰). Populations are ordered from the left to the right by increasing least-square mean values of δ13C (gray circles). The bottom and the top of the box are the first and the third quartile, respectively, and the bold line represents the median. The whiskers are extending from minimum to maximum values for each population. The box colors indicate the average water deficit index (dimensionless) calculated over the 1960–1999 period (B) (see Figure 1 and Supplementary Table 1). Different letters in the upper panel indicate significant differences among populations. For each population, the box width is proportional to the sample size (n), which is also displayed below each corresponding box.
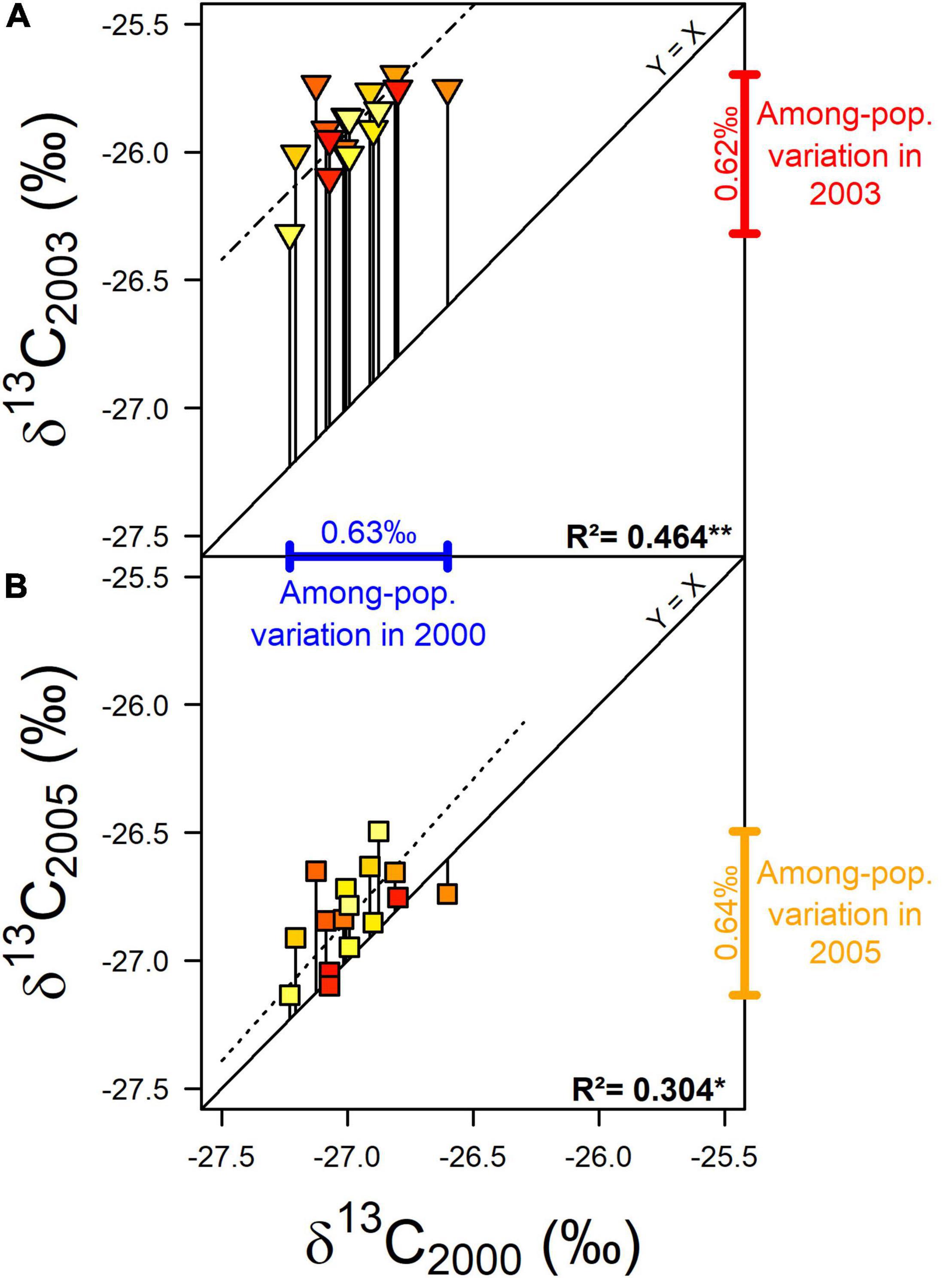
Figure 4. Population δ13C in 2003 (A; severely dry year, triangles) and in 2005 (B; moderately dry year, squares) compared to δ13C in 2000 (wet year). The vertical lines between the Y = X line and each point represent the change of δ13C between dry and wet years (δ13C plasticity index) for each population. Dotted lines are the type II regression lines between δ13C in 2003 and 2000 (A) and δ13C in 2005 and 2000 (B). The corresponding coefficient of determination (R2; n = 16) and significance level (*: 0.01 < p < 0.05; **: 0.001 < p < 0.01) are also shown. Point colors indicate the average water deficit index (dimensionless) calculated over the 1960–1999 period in each provenance site (see Figure 1 and Supplementary Table 1).
The population effect was low, yet significant, on RW but not on GI (Table 1). A significant interaction with the year effect was observed for both RW and GI (Table 1). Particularly, if no significant difference in GI was detected among populations in 2000, trees from Fontainebleau had significantly lower GI in 2003 compared to ten other populations and higher GI in 2005 than those from Dreuille (Supplementary Table 3). For all traits, the within-population variations were always higher than the among-population variations (Supplementary Table 4).
No statistically significant differences of the δ13C plasticity indices were observed among populations (Table 1 and Supplementary Table 3), and the drought intensity × population interaction was also not significant (Table 1). Regarding GI plasticity indices, there was globally no significant effect of the population but its interaction with drought intensity effect appeared significant (Table 1). Indeed, significant population differences in GI plasticity were detected in response to the severe drought while the response to the moderate drought proved to be uniform among populations. In response to the severe drought, trees from Fontainebleau displayed a larger increase in GI compared to those from Abbayes, Bercé, and Lappwald (Supplementary Table 3).
Based on WDI1960–1999, the common garden of Sillegny appeared in a median position with eight drier and eight wetter provenance sites (Figure 1). More negative δ13C values were observed on populations with both lower and higher WDI1960–1999 values than Sillegny (Figure 3). Regardless of the year or drought intensity considered, no significant relationship was observed between δ13C – or δ13C plasticity – and the WDI1960–1999 of the provenance sites (p > 0.3). No other pedoclimatic parameters described in Supplementary Table 1, including AI and SHM, were significantly related to δ13C. The plasticity in δ13C between the wet and the moderately dry year (δ13C2005–2000) was also not significantly correlated to any pedoclimatic parameters. However, the increase in population mean δ13C between the wet and the severely dry year (δ13C2003–2000) was positively correlated to the stock of organic carbon (Corg) in the topsoil (the first 30 cm of depth, see Supplementary Table 5) at their provenance sites (respectively, R2 = 0.464, p = 0.0037). Whether in topsoil or subsoil (Supplementary Table 5), population δ13C2003–2000 was also positively correlated to proportion of silt (Figure 5; topsoil: R2 = 0.438, p = 0.0052; subsoil: R2 = 0.492, p = 0.0025) and negatively correlated to the proportion of sand in the soil at their provenance sites (topsoil: R2 = 0.335, p = 0.0189; subsoil: R2 = 0.300, p = 0.0279). No significant relationship was found between the pedoclimate of the provenance sites and the radial growth (i.e., RW and GI) of the populations in the common garden. However, positive correlations were found between the mean population plasticity indices of GI to drought (GI2003–2000 and GI2005–2000) and the average air vapor pressure deficit from May to July for the period 1960–1999 (D0507) of the provenance sites (respectively, R2 = 0.289, p = 0.0318 and R2 = 0.338, p = 0.0182). No significant relationship was found between the population means of the measured traits (i.e., δ13C, GI, and their plasticity indices) and the remaining soil characteristic parameters described in Supplementary Table 5 (Root, C/N, pHH2O, pHKCl, CEC, N stock, and PAWC).
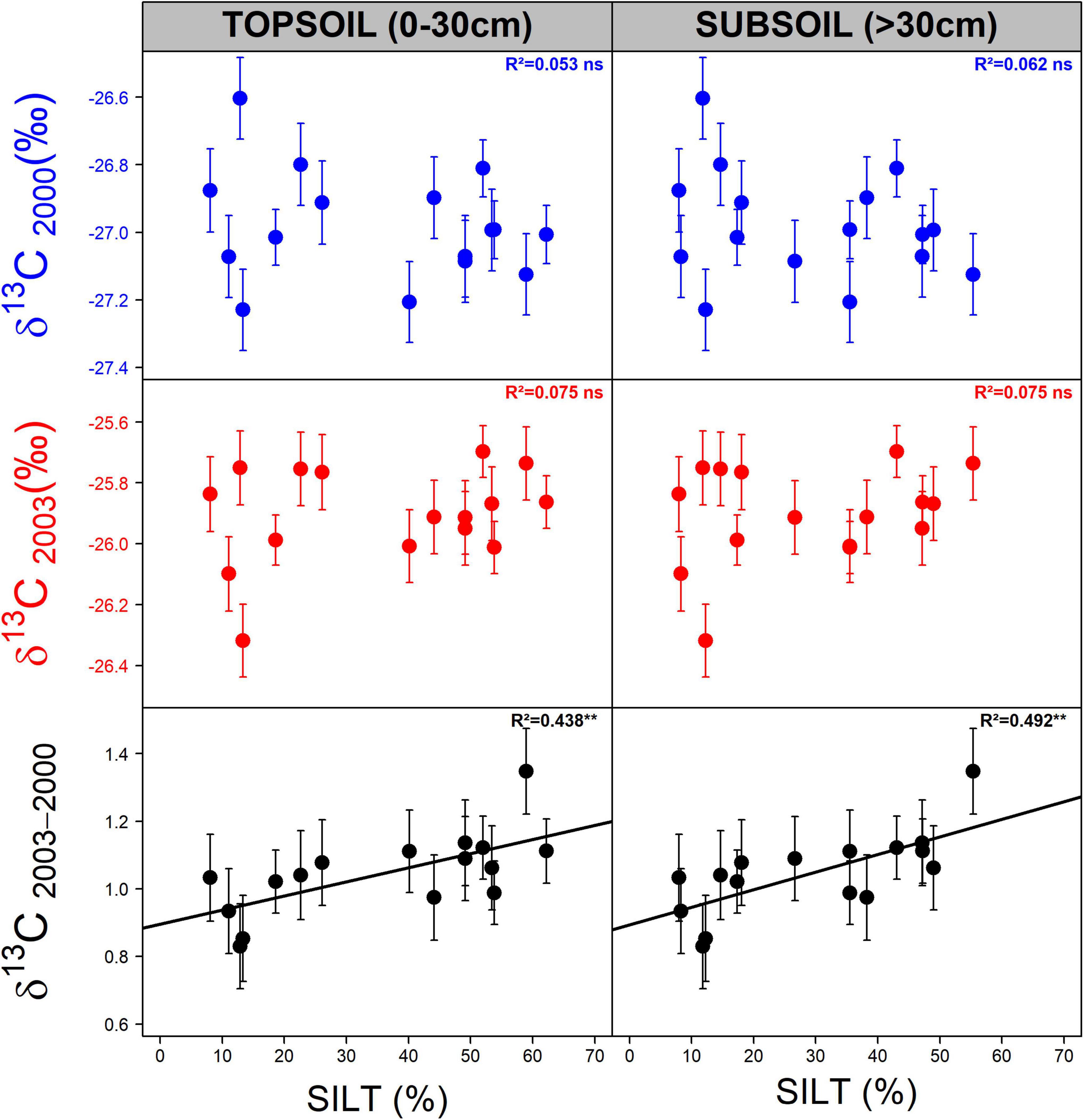
Figure 5. Relationship between population δ13C in 2003 (red), δ13C in 2000 (blue) and δ13C2003–2000 (plasticity of δ13C to severe drought) and the percentage of silt in the soil of their provenance sites. Topsoil represents the first 30 cm of the soil (in depth), and subsoil represents the profile below 30 cm down to the rooting depth. The error bars represent the standard error of the mean. For each correlation, the coefficient of determination (R2; n = 16) and the significance level (ns: p > 0.05; **: 0.001 < p < 0.01) are shown.
When analyzing variations in traits and plasticity indices with the mixed model, the conditional R2 was always approximately twice the marginal R2 (Table 1), which suggests that random effects explained as much variability as fixed effects. Among random effects, the tree effect (in the micro-block nested in the block) had the largest standard deviation for δ13C, RW, δ13C plasticity index, and GI plasticity index (Table 1). Because the detrending process included the centering of the data for each tree independently, the standard deviation due to tree effect was, as expected, zero for GI (Table 1).
During the wet 2000 year, the populations with the highest growth index also displayed the less negative δ13C (R2 = 0.343; p = 0.0171; Figure 6). In 2000, even if no significant difference was detected among populations for GI, the trees from Dreuille, which showed the most negative δ13C mean value, showed a slightly negative GI, and the trees from Fontainebleau, which presented the least negative δ13C mean value, showed the highest GI mean value (Supplementary Table 2). Especially, trees from Fontainebleau, which showed the less negative δ13C mean value in 2000, showed the lowest GI in 2003. During dry years, in 2003 and 2005, the among-population relationships between δ13C and GI were not significant (p > 0.65). However, a positive significant correlation was found among populations between the plasticity indices of δ13C2003–2000 and GI2003–2000 (R2 = 0.429; p = 0.0059). It is also noticeable that populations with high δ13C in the wet 2000 year tended to have low GI in the dry 2003 years (R2 = 0.293; p = 0.0303).
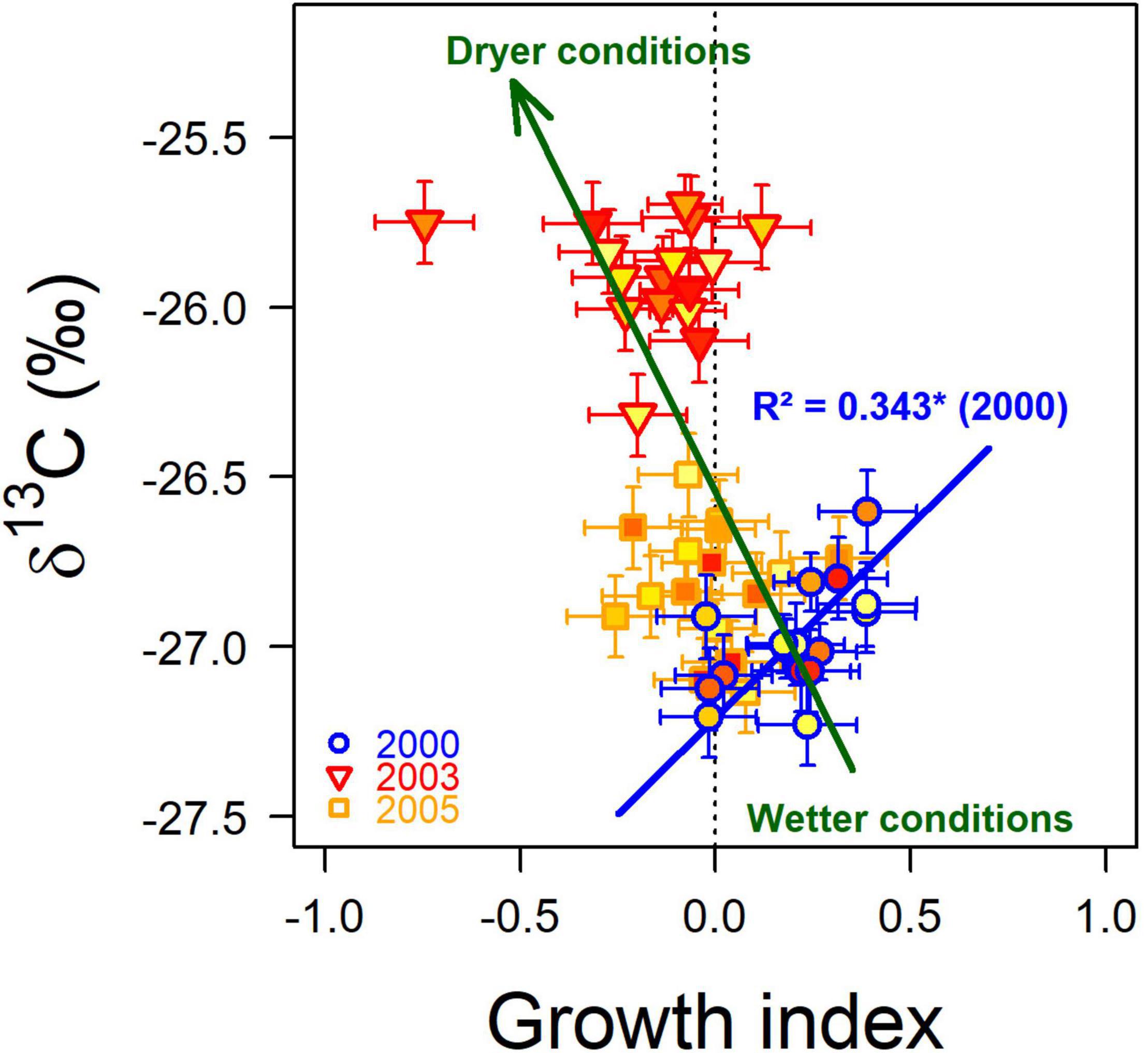
Figure 6. Correlation between the least-square means of δ13C and growth index (GI) in 2000 (blue circles), 2003 (red triangles), and 2005 (orange squares). The error bars represent the standard error of the mean. The blue line indicates the significant among-population relationship between the two traits in 2000 (n = 16) with its coefficient of determination (R2) and significance level (*: 0.01 < p < 0.05). The green arrow represents the direction of δ13C and growth index variations in response to drought. The type II regression between the two traits in 2003 and 2005 is not shown because not significant. Symbol fill colors indicate the average water deficit index (dimensionless) calculated over the 1960 – 1999 period (Figures 1, 3).
At the within-population level, correlations between δ13C and GI in 2000 were significant for the four populations where 60 trees were sampled (instead of 30 for the other populations) as well as the Lappwald population (0.103 < R2< 0.295, 0.0004 < p < 0.0166). A positive correlation between δ13C2003–2000 and GI2003–2000 was observed within 10 out 16 populations (including all populations with n ≈ 60, 0.118 < R2< 0.379, 0.000001 < p < 0.0219).
Trees, as long-lived organisms, need to be able to respond to their environment by adjusting their physiology and their development through plasticity (Scheiner, 1993). By measuring functional traits on tree rings, one is able to observe retrospectively the plastic responses of trees over the years and relate them to environmental variables. A significant relationship between wood carbon isotope composition (δ13C) and WUEi was demonstrated for white oak species (Ponton et al., 2002; Roussel et al., 2009).
In our study, the moist year 2000 was considered as a control compared to the moderately dry year 2005 and the severely dry year 2003. δ13C2003–2000 and δ13C2005–2000, used as plasticity indices of δ13C in response to severe and moderate drought, respectively, were as expected positive (Table 2). Indeed, for most plants and under most circumstances, a decrease in soil water availability is immediately followed by stomatal closure, resulting in an increase in WUEi and δ13C (Farquhar and Sharkey, 1982; Farquhar et al., 1989; Brendel and Epron, 2022), due to the non-linear relationship between A and gsw (Supplementary Figure 5). However, the δ13C values observed in 2005 were much closer (although significantly higher) to those of the wet year 2000 than those of the dry year 2003. It has been demonstrated, especially for sessile oak, that the stomatal closure occurs gradually below a drought threshold of 40% REW (Bréda et al., 1993, 1995; Triboulot et al., 1996). In 2005, the drought lasted longer than in 2003 but the daily REW never dropped below 10% contrary to 2003 with 33 consecutive days below 10% (Figure 2). In addition, air vapor pressure deficit (VPD) reached very high values in August 2003 (>15 hPa) whereas much lower ones were observed at the same period in both 2005 and 2000 (Figure 2). A less negative δ13C in 2003 was therefore expected but the relatively low value observed in 2005, although clearly marked by a water deficit, remained surprising. It could suggest that for these young oak trees, REW needs to decrease below 10% for a strong stomatal reaction.
Compared to 2000, the growth index in 2003 was reduced. The concomitant increase in δ13C in 2003 could indicate either a stomatal closure or an increase in A, the latter being not likely to happen under the estimated soil water deficit. Thus, although a slight decrease in A (hidden by a much larger decrease in gsw) cannot be ruled out, the main reason for the decrease in radial growth might be growth inhibition at low water potentials (Fatichi et al., 2014).
Using a common garden, with the assumption that all trees grow in a homogeneous environment, allows separating the effects of genetic diversity from those of the environment among the studied populations (Wang et al., 2010). Although significant differentiations in δ13C were detected among populations for the three studied years (Table 1), only a few populations differed significantly from the others, with only one (i.e., Farchau) being remarkable for three years with the lowest WUEi (Supplementary Table 3). The δ13C range of 0.6‰ observed among the studied populations was remarkably constant over the three years, despite their different drought intensities. This variation of 0.6‰ among δ13C populations found in sessile oak seems to be low compared to the 2‰ observed in Castanea sativa (Lauteri et al., 2004). However, such small but significant differences among populations have also been observed in other oak species (0.2‰ for Quercus suber, Matías et al., 2019; 0.6‰ for Quercus variabilis, Sun et al., 2016; 0.8‰ for Quercus douglasii, Rice et al., 1993) and for Fagus sylvatica (0.2–1‰, Rose et al., 2009; Robson et al., 2012; Knutzen et al., 2015). In contrast to these studies where δ13C was measured in leaves, our study is, to our knowledge, the only one to use wood δ13C to assess WUEi diversity among populations in the Fagaceae family. While measurements in leaves introduce variability due to the position of the harvested leaves (Le Roux et al., 2001), measuring δ13C in wood has the advantage of integrating the functioning of the entire canopy (Ponton et al., 2002; Roussel et al., 2009).
Because the selection was restricted to populations from the Iberian post-glaciation recolonization lineage (lineage B, Petit et al., 2002), our population selection did not cover the entire distribution area of sessile oak and excluded the more drought-exposed southern populations. However, Torres-Ruiz et al. (2019) who compared a different selection of populations from the same common garden of Sillegny, including different lineages and populations from Ireland and southern France, did not detect any significant differences in δ13C among populations. This supports Kremer’s (2016) statement that there is no difference in local adaptation between white oak lineages from different glacial refugia.
Detecting a relationship between variations in functional traits among populations and variations in the average environmental conditions at their site of origin would support the hypothesis that variations in these traits have evolved under different local selection pressures. Of all the studies detecting significant population differences for δ13C or A/gsw on trees, negative (Li et al., 2000; Cregg and Zhang, 2001; Bekessy et al., 2002; Zhang et al., 2005; Duan et al., 2009) as well as positive (Aitken et al., 1995; Guehl et al., 1996; Nguyen-Queyrens et al., 1998; Ferrio et al., 2003; Lauteri et al., 2004; Eriksson et al., 2005; Fan et al., 2008) correlations have been detected between drought indices and population δ13C. This might suggest that there is not a simple relationship between the mean WUEi of a population and its adaptation to a seasonal dry environment (Brendel and Epron, 2022). In the current study, the pedoclimate of each provenance site was characterized in detail by using the BILJOU© water balance model (Granier et al., 1999), which includes soil features in addition to climatic parameters. The lack of significant correlation between the δ13C of the 16 populations and the pedoclimatic indices calculated by the model, as well as the raw atmospheric variables and soil characteristics, leads to the rejection of our first hypothesis, stipulating that populations from the driest origins would display the lowest WUEi. Similar observations were made for Quercus ilex (Gimeno et al., 2009) and Quercus robur (George et al., 2020) and for conifer species in the Pinaceae and Cupressaceae families (Zhang and Cregg, 1996).
The estimation of the environmental constraints in the populations based on meteorological data from a relatively recent period (1960–1999) might be temporally disconnected from the actual genetic differentiation, which would have been ongoing over several generations, thus probably several centuries earlier (Saleh et al., 2021). Nevertheless, significant relationships were established for sessile oak between soil water availability at provenance sites and other traits such as height (Bruschi, 2010; Torres-Ruiz et al., 2019; but also see Arend et al., 2011), root–shoot ratio, leaf size, and thickness (Bruschi, 2010) or leaf unfolding and specific leaf area (Torres-Ruiz et al., 2019), demonstrating that sessile oak populations show differentiation in functional traits related to drought adaptation. Bert et al. (2020), in a study partly based on the same experimental design as our study, found that the correlation between radial growth of sessile oak populations and water balance was lower at Sillegny than at the other two common gardens studied, exposed to drier conditions. Therefore, the lack of relationship between populations WUEi and pedoclimatic conditions at the provenance sites might be related to the relatively low-stress environment of the Sillegny common garden.
The phenotypic plasticity of a trait to an environmental change can also be subject to selection and considered as an adaptive trait (Via et al., 1995), resulting in population differences in plasticity, related to environmental differences among provenance sites. In our study, no significant population effect was detected for the plasticity indices of δ13C to drought (Table 1 and Supplementary Table 3), suggesting that the within-population variance was larger than the among-population differences. However, a significant interaction between drought and population effects was observed for δ13C. Moreover, the population means of δ13C2003–2000 plasticity were correlated to soil texture. No such correlation was detected for the modeled population WDI1960–1999, which is based on relatively recent meteorological data. Soil texture is constant over a much longer time period, and might better reflect past selection pressures, which might have resulted in population differences. Populations with a lower δ13C2003–2000 plasticity were originating from sites with a lower proportion of silt (Figure 6) and organic carbon (in the topsoil) and a higher proportion of sand. These results are consistent with observations made by Rellstab et al. (2016) who showed a significant association between genetic variation within and across three oak species (including sessile oak) and soil characteristics related to soil aeration and clay content. To our knowledge, there are only few publications that have investigated among-population differences in plasticity of WUEi to drought. A significant among-population difference in WUEi plasticity to drought was found for Quercus suber (Matías et al., 2019), where only the southern population had shown a significant δ13C increase to drought, whereas the northern population did not. However, Gimeno et al. (2009) did not detect a significant population x drought interaction on direct estimates of WUEi on Quercus ilex leaves, suggesting no among-population difference in WUEi plasticity, whereas significant population differences were shown for absolute values of WUEi.
Our results suggest that populations from provenance sites on soils with more draining soil texture (i.e., more sand/less silt and soil carbon) showed a less plastic response of WUEi to the severe 2003 drought at Sillegny, presumably by maintaining their stomata more open. Maintaining stomata open during drought could indicate either higher tolerance for low water potentials, or a larger root system with a lower leaf surface/root surface ratio maintaining the soil water supply. The populations from the provenance sites with more sandy soils might well have evolved toward larger root systems [as for example shown by Matías et al. (2019) for more Southern populations of Q. suber] and thereby acquired a different response strategy to soil water deficit compared to populations from provenance sites with a high plant-available water capacity of the soil.
Further, populations originating from provenance sites with the driest atmosphere, based on May-July mean values, decreased less their growth (GI) in the dry years 2003 and 2005 at Sillegny compared to the wet year (2000). This, again, suggests that populations originating from drier sites showed less response to drought in the mesic conditions of Sillegny. Such an among-population variation in the plasticity to VDP provides an interesting outlook, considering the predicted increase in VPD with the ongoing climate change and the concurrently increasing risk of mortality (Grossiord et al., 2020).
Both results, the correlation of mean population plasticity of δ13C with soil texture and of mean population plasticity GI with atmospheric humidity, suggest that populations from drier environments have evolved different drought response strategies than populations from wetter environments. These differences resulted in a similar variation in WUEi and growth plasticity for the severe drought year, as GI2003–2000 and δ13C2003–2000 were significantly correlated. Whereas during the moderate drought year 2005, a more pronounced reduction in GI was observed compared to the smaller increase in δ13C (Table 2 and Figure 6). It is likely that under this moderate drought level, leaf gas exchanges were maintained whereas stem growth was decreased (Bogeat-Triboulot et al., 2007).
For woody species, within-population variation in traits is generally larger than among-population variation (Hamrick et al., 1992; Bresson et al., 2011), as also found in this study for δ13C as an estimator of WUEi. The experimental design of the common garden in blocks and micro-blocks was made to minimize as much as possible the variation in environmental conditions among trees to improve the detection of genetic variations. However, in addition to the influence of genetic factors on WUEi, that of variations in the microenvironment, including those generated by differences in social status, cannot be excluded (McDowell et al., 2003; Niccoli et al., 2020).
In our study, dominant trees expectedly expressed slightly less negative δ13C (higher WUEi) values than co-dominant and suppressed trees, probably due to higher overall assimilation rates (Forrester, 2019; Vitali et al., 2021). The attribution of the social status was done in 2014, when the canopy of the plantation was closed, whereas the δ13C measurements were done on annual rings before canopy closure. However, height measurements taken in 2002 showed that the trees identified as dominant in 2014 were already taller in 2002 (data not shown). Overall, this could suggest that the higher WUEi of dominant trees is not only due to the direct physiological impacts of competition for light, but might also indicate that the stronger growth of dominant trees might be related to higher photosynthetic capacity. However, no significant social status effect (Table 1) and no interaction with drought intensity were found for the plasticity index of δ13C to drought, supporting the idea that trees with different social status reacted similarly to the studied drought intensities.
In our study, more phenotypic variability in sessile oak δ13C (thus WUEi) was found among individuals within the same population than among populations. The among-sibling variation in δ13C in one full-sib family of Quercus robur was more than 3‰ (similar to the range observed within populations in our study, Supplementary Table 3), and was strongly genetically controlled (Brendel et al., 2008). Large within-population variability in WUEi was also reported for Fagus sylvatica (Aranda et al., 2017) and Quercus suber (Ramirez-Valiente et al., 2010). One reason for this large within-population variability in WUEi may be the low heritability of WUEi, demonstrated by Alexandre et al. (2020a) for sessile oak. Alexandre et al. (2020b) found that δ13C, unlike tree growth and some leaf morphological traits, was unresponsive to contemporary selection in sessile oak and attributed the considerable variations among individuals to adaptive divergence to drought driven by diversifying selection. This diversifying selection of WUEi for sessile oak may explain why no linear relationship was found between populations WUEi and the pedoclimatic conditions at their provenance sites.
Based on the hypothesis that during the wet year (i.e., in 2000), the stomata were overall more open than during the dry years and did therefore restrict A less, and if light resources were not limiting, the observed among-population variation in WUEi would have more likely resulted from a variation in photosynthetic capacity, rather than from variations in stomatal conductance (Supplementary Figure 5) as suggested by Aranda et al. (2017) for Fagus sylvatica. There was no significant effect of population on GI, however, the observed significant population/year interaction would suggest a diversity of growth plasticity for drought among populations. A similar result was observed in a much larger study, in which Sillegny was one of the common gardens studied (Bert et al., 2020). In 2000, populations with high δ13C also displayed a high growth index (Figure 6). A similar relationship was also observed at the within-population level and was statistically significant for populations with 60 individuals sampled, suggesting diversity among but also within populations for photosynthetic capacity. Establishing a relation between WUEi and radial growth appears complex because carbon from leaf photosynthetic assimilation (A) is redistributed into different compartments of the tree and only a portion is used to build tree rings (Hartmann et al., 2018a). In addition, ring width and δ13C are the final results of different dynamic processes, including post-photosynthetic fractionations between leaf assimilates and wood (Gessler et al., 2014), and the use of assimilates from previous years to build earlywood in oak species (Vincent-Barbaroux et al., 2019).
The relationship between δ13C and growth index disappeared during dry years (Figure 6), likely because carbon assimilation is restricted by stomatal closure, and therefore masking any population differences in photosynthetic capacity. It has also been shown that during dry conditions, wood growth can be disconnected from carbon assimilation due to carbon storage processes, allocation, and remobilization (Pflug et al., 2015). Our third hypothesis, stipulating that trees and populations with a higher WUEi only showed a higher radial growth when the soil water availability was unlimited, is confirmed.
In this study, the within-species variability in sessile oak WUEi was evaluated by comparing the δ13C values of 16 populations growing in the common garden of Sillegny. The among-population variation in δ13C (0.6‰) was low compared to the inter-annual variation in δ13C (1‰), and especially compared to the within-population variation in δ13C (2–4‰). The δ13C measurements over three contrasted years with different soil drought levels showed similar plasticity of WUEi to drought among populations. Nevertheless, it was found that the among-population variation in WUEi plasticity correlated with soil characteristics at the provenance sites. In addition, the plasticity in the growth index correlated with summer atmospheric humidity. Both results could suggest a local adaptation of sessile oak in terms of drought response strategies. Moreover, we observed a large variability in WUEi within populations. This large variability, observed in a common environment, could suggest a large underlying genetic diversity, which could facilitate the adaptation of sessile oak to future climate changes. However, to substantiate this conclusion, it will be necessary for future research programs to partition observed variation into genetic and environmental causes.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
AR: data curation, formal analysis, investigation, methodology, and writing—original draft. SP: conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, project administration, supervision, and writing—original draft. DB: investigation, validation, and writing—original draft. AD: resources. BR: investigation, methodology, resources, validation, and writing—original draft. JL: investigation, methodology, resources, validation, and writing—original draft. OB: conceptualization, formal analysis, funding acquisition, investigation, methodology, project administration, supervision, and writing—original draft. All authors contributed to the article and approved the submitted version.
This study was part of the interdisciplinary «H2Oak» research project (ANR-14-CE02-0013 project) aiming to investigate the diversity for adaptative traits related to water use in two European white oaks (Quercus robur and Quercus petraea). This work was supported by a grant overseen by the French National Research Agency (ANR) as part of the “Investissements d’Avenir” program (ANR-11-LABX-0002-01, Lab of Excellence ARBRE).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Our gratitude turns to Patrick Behr, Fabrice Bonne, Bruno Garnier, Thierry Paul, Vincent Rousselet, and Erwin Thirion (SILVA), Frédéric Lagane, Patrick Reynet, and Raphaël Ségura (BIOGECO) for their help in the tree sampling. We are grateful to Cyrielle Visine and Nina Potts (SILVA) for the sample preparation and to Christian Hossann (SILVA) for the isotopic analysis. We thank Vincent Badeau (SILVA) for his help in the field and, together with Nathalie Bréda (SILVA), for their advice regarding the water balance modeling. We would like to thank SILVATECH (Silvatech, INRAE, 2018. Structural and functional analysis of tree and wood Facility, doi: 10.15454/1.5572400113627854E12) from UMR 1434 SILVA, 1136 IAM, 1138 BEF and 4370 EA LERMAB from the research center INRAE Grand-Est Nancy for the carbon stable isotope analysis. SILVATECH facility is supported by the French National Research Agency through the Laboratory of Excellence ARBRE (ANR-11-LABX-0002-01). We also thank the USC INRAE Lermab for the facility in the laboratory to extract the lignin.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2022.914199/full#supplementary-material
Aitken, S. N., and Bemmels, J. B. (2016). Time to get moving: assisted gene flow of forest trees. Evol. Appl. 9, 271–290. doi: 10.1111/eva.12293
Aitken, S. N., Kavanagh, K. L., and Yoder, B. J. (1995). Genetic variation in seedling water-use efficiency as estimated by carbon isotope ratios and its relationship to sapling growth in douglas-fir. For. Genet. 2, 199–206.
Aitken, S. N., Yeaman, S., Holliday, J. A., Wang, T., and Curtis-McLane, S. (2008). Adaptation, migration or extirpation: climate change outcomes for tree populations. Evol. Appl. 1, 95–111. doi: 10.1111/j.1752-4571.2007.00013.x
Al Majou, H., Bruand, A., Duval, O., and Cousin, I. (2007). Variation of the water-retention properties of soils: validity of class-pedotransfer functions. Comptes Rendus Geosci. 339, 632–639. doi: 10.1016/j.crte.2007.07.005
Alexandre, H., Truffaut, L., Ducousso, A., Louvet, J.-M., Nepveu, G., Torres-Ruiz, J. M., et al. (2020a). In situ estimation of genetic variation of functional and ecological traits in Quercus petraea and Q. robur. Tree Genet. Genomes 16:32. doi: 10.1007/s11295-019-1407-9
Alexandre, H., Truffaut, L., Klein, E., Ducousso, A., Chancerel, E., Lesur, I., et al. (2020b). How does contemporary selection shape oak phenotypes? Evol. Appl. 13, 2772–2790. doi: 10.1111/eva.13082
Alfaro-Sánchez, R., Valdés-Correcher, E., Espelta, J. M., Hampe, A., and Bert, D. (2020). How do social status and tree architecture influence radial growth, wood density and drought response in spontaneously established oak forests? Ann. For. Sci. 77:49. doi: 10.1007/s13595-020-00949-x
Allen, C. D., Macalady, A. K., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 259, 660–684. doi: 10.1016/j.foreco.2009.09.001
Anderegg, L. D. L., Loy, X., Markham, I. P., Elmer, C. M., Hovenden, M. J., HilleRisLambers, J., et al. (2021). Aridity drives coordinated trait shifts but not decreased trait variance across the geographic range of eight Australian trees. New Phytol. 229, 1375–1387. doi: 10.1111/nph.16795
Aranda, I., Alía, R., Ortega, U., Dantas, ÂK., and Majada, J. (2010). Intra-specific variability in biomass partitioning and carbon isotopic discrimination under moderate drought stress in seedlings from four Pinus pinaster populations. Tree Genet. Genomes 6, 169–178. doi: 10.1007/s11295-009-0238-5
Aranda, I., Bahamonde, H. A., and Sánchez-Gómez, D. (2017). Intra-population variability in the drought response of a beech (Fagus sylvatica L.) population in the southwest of Europe. Tree Physiol. 37, 938–949. doi: 10.1093/treephys/tpx058
Arend, M., Kuster, T., Gunthardt-Goerg, M. S., and Dobbertin, M. (2011). Provenance-specific growth responses to drought and air warming in three European oak species (Quercus robur, Q. petraea and Q. pubescens). Tree Physiol. 31, 287–297. doi: 10.1093/treephys/tpr004
Barton, K. (2020). MuMIn: Multi-Model Inference. R package version 1.43.17. Available online at: https://cran.r-project.org/package=MuMIn.
Bekessy, S. A., Sleep, D., Stott, A., Menuccini, M., Thomas, P., Ennos, R. A., et al. (2002). Adaptation of monkey puzzle to arid environments reflected by regional differences in stable carbon isotope ratio and allocation to root biomass. For. Genet. 9, 63–70.
Bert, D., Le Provost, G., Delzon, S., Plomion, C., and Gion, J.-M. (2021). Higher needle anatomic plasticity is related to better water-use efficiency and higher resistance to embolism in fast-growing Pinus pinaster families under water scarcity. Trees 35, 287–306. doi: 10.1007/s00468-020-02034-2
Bert, D., Lebourgeois, F., Ponton, S., Musch, B., and Ducousso, A. (2020). Which oak provenances for the 22nd century in Western Europe? dendroclimatology in common gardens. PLoS One 15:e0234583. doi: 10.1371/journal.pone.0234583
Bishop, T. F. A., McBratney, A. B., and Laslett, G. M. (1999). Modelling soil attribute depth functions with equal-area quadratic smoothing splines. Geoderma 91, 27–45. doi: 10.1016/S0016-7061(99)00003-8
Bogeat-Triboulot, M.-B., Brosché, M., Renaut, J., Jouve, L., Le Thiec, D., Fayyaz, P., et al. (2007). Gradual soil water depletion results in reversible changes of gene expression, protein Pro?les, ecophysiology, and growth performance in populus euphratica, a poplar growing in arid regions. Plant Physiol. 143, 876–892. doi: 10.1104/pp.106.088708
Bréda, N., Cochard, H., Dreyer, E., and Granier, A. (1993). Water transfer in a mature oak stand (Quercus petraea): seasonal evolution and effects of a severe drought. Can. J. For. Res. 23, 1136–1143. doi: 10.1139/x93-144
Bréda, N., Granier, A., Barataud, F., and Moyne, C. (1995). Soil water dynamics in an oak stand. Plant Soil 172, 17–27.
Brendel, O., and Epron, D. (2022). Are differences among forest tree populations in carbon isotope composition an indication of adaptation to drought? Tree Physiol. 42, 26–31. doi: 10.1093/treephys/tpab143
Brendel, O., Le Thiec, D., Scotti-Saintagne, C., Bodénès, C., Kremer, A., and Guehl, J.-M. (2008). Quantitative trait loci controlling water use efficiency and related traits in Quercus robur L. Tree Genet. Genomes 4, 263–278. doi: 10.1007/s11295-007-0107-z
Bresson, C. C., Vitasse, Y., Kremer, A., and Delzon, S. (2011). To what extent is altitudinal variation of functional traits driven by genetic adaptation in European oak and beech? Tree Physiol. 31, 1164–1174. doi: 10.1093/treephys/tpr084
Brewer, S., Hélyalleaume, C., Cheddadi, R., de Beaulieu, J.-L., Laurent, J.-M., and Le Cuziat, J. (2005). Postglacial history of Atlantic oakwoods: context, dynamics and controlling factors. Bot. J. Scotl. 57, 41–57. doi: 10.1080/03746600508685084
Bruschi, P. (2010). Geographical variation in morphology of Quercus petraea (Matt.) Liebl. as related to drought stress. Plant Biosyst. – Int. J. Deal. Asp. Plant Biol. 144, 298–307. doi: 10.1080/11263501003672462
Büntgen, U., Urban, O., Krusic, P. J., Rybníèek, M., Koláø, T., Kyncl, T., et al. (2021). Recent European drought extremes beyond Common Era background variability. Nat. Geosci. 14, 190–196. doi: 10.1038/s41561-021-00698-0
Caudullo, G., Welk, E., and San-Miguel-Ayanz, J. (2017). Chorological maps for the main European woody species. Data Brief 12, 662–666. doi: 10.1016/j.dib.2017.05.007
Cheddadi, R., de Beaulieu, J.-L., Jouzel, J., Andrieu-Ponel, V., Laurent, J.-M., Reille, M., et al. (2005). Similarity of vegetation dynamics during interglacial periods. Proc. Natl. Acad. Sci. U S A. 102, 13939–13943. doi: 10.1073/pnas.0501752102
Cregg, B. M., and Zhang, J. W. (2001). Physiology and morphology of Pinus sylvestris seedlings from diverse sources under cyclic drought stress. For. Ecol. Manag. 154, 131–139. doi: 10.1016/S0378-1127(00)00626-5
Duan, B., Li, Y., Zhang, X., Korpelainen, H., and Li, C. (2009). Water deficit affects mesophyll limitation of leaves more strongly in sun than in shade in two contrasting Picea asperata populations. Tree Physiol. 29, 1551–1561. doi: 10.1093/treephys/tpp085
Dufrêne, E., Davi, H., François, C., Le Maire, G., Le Dantec, V., and Granier, A. (2005). Modelling carbon and water cycles in a beech forest. Part I: model description and uncertainty analysis on modelled NEE. Ecol. Model. 185, 407–436. doi: 10.1016/j.ecolmodel.2005.01.004
Eriksson, G., Jonsson, A., Lauteri, M., and Pliura, A. (2005). Genetic variation in drought response of Castanea sativa Mill. seedlings. Acta Hortic. 693, 247–254. doi: 10.17660/ActaHortic.2005.693.30
Fan, S., Grossnickle, S. C., and Russell, J. H. (2008). Morphological and physiological variation in western redcedar (Thuja plicata) populations under contrasting soil water conditions. Trees 22, 671–683. doi: 10.1007/s00468-008-0225-8
Fardusi, M. J., Ferrio, J. P., Comas, C., Voltas, J., Resco de Dios, V., et al. (2016). Intra-specific association between carbon isotope composition and productivity in woody plants: a meta-analysis. Plant Sci. 251, 110–118. doi: 10.1016/j.plantsci.2016.04.005
Farquhar, G. D., and Sharkey, T. D. (1982). Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 33, 317–345. doi: 10.1146/annurev.pp.33.060182.001533
Farquhar, G. D., Ehleringer, J. R., and Hubick, K. T. (1989). Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40, 503–537.
Farquhar, G., and Richards, R. (1984). Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Funct. Plant Biol. 11:539. doi: 10.1071/PP9840539
Fatichi, S., Leuzinger, S., and Körner, C. (2014). Moving beyond photosynthesis: from carbon source to sink-driven vegetation modeling. New Phytol. 201, 1086–1095. doi: 10.1111/nph.12614
Ferrio, J. P., Florit, A., Vega, A., Serrano, L., and Voltas, J. (2003). Δ13C and tree-ring width reflect different drought responses in Quercus ilex and Pinus halepensis. Oecologia 137, 512–518. doi: 10.1007/s00442-003-1372-7
Forrester, D. I. (2019). Linking forest growth with stand structure: tree size inequality, tree growth or resource partitioning and the asymmetry of competition. For. Ecol. Manag. 447, 139–157. doi: 10.1016/j.foreco.2019.05.053
Friedrichs, D. A., Trouet, V., Büntgen, U., Frank, D. C., Esper, J., Neuwirth, B., et al. (2009). Species-specific climate sensitivity of tree growth in Central-West Germany. Trees 23, 729–739. doi: 10.1007/s00468-009-0315-2
Gárate-Escamilla, H., Hampe, A., Vizcaíno-Palomar, N., Robson, T. M., and Benito Garzón, M. (2019). Range-wide variation in local adaptation and phenotypic plasticity of fitness-related traits in Fagus sylvatica and their implications under climate change. Glob. Ecol. Biogeogr. 28, 1336–1350. doi: 10.1111/geb.12936
George, J., Theroux-Rancourt, G., Rungwattana, K., Scheffknecht, S., Momirovic, N., Neuhauser, L., et al. (2020). Assessing adaptive and plastic responses in growth and functional traits in a 10-year-old common garden experiment with pedunculate oak (Quercus robur L.) suggests that directional selection can drive climatic adaptation. Evol. Appl. 13, 2422–2438. doi: 10.1111/eva.13034
Gessler, A., Ferrio, J. P., Hommel, R., Treydte, K., Werner, R. A., and Monson, R. K. (2014). Stable isotopes in tree rings: towards a mechanistic understanding of isotope fractionation and mixing processes from the leaves to the wood. Tree Physiol. 34, 796–818. doi: 10.1093/treephys/tpu040
Gimeno, T. E., Pias, B., Lemos-Filho, J. P., and Valladares, F. (2009). Plasticity and stress tolerance override local adaptation in the responses of Mediterranean holm oak seedlings to drought and cold. Tree Physiol. 29, 87–98. doi: 10.1093/treephys/tpn007
Granier, A., Bréda, N., Biron, P., and Villette, S. (1999). A lumped water balance model to evaluate duration and intensity of drought constraints in forest stands. Ecol. Model. 116, 269–283. doi: 10.1016/S0304-3800(98)00205-1
Grossiord, C., Buckley, T. N., Cernusak, L. A., Novick, K. A., Poulter, B., Siegwolf, R. T. W., et al. (2020). Plant responses to rising vapor pressure deficit. New Phytol. 226, 1550–1566. doi: 10.1111/nph.16485
Grotehusmann, H., and Schönfelder, E. (2011). Comparison of French and German sessile oak (Quercus petraea (Matt.) Liebl.) provenances. Silvae Genet. 60, 186–196. doi: 10.1515/sg-2011-0025
Guehl, J. M., Nguyen-Queyrens, A., Loustau, D., and Ferhi, A. (1996). “Genetic and environmental determinants of water-use efficiency and carbon isotope discrimination in forest trees,” in Paper Presented at the Eurosilva Contribution to Forest tree Physiology, Editions Colloques de l’INRA, eds M. Bonnet-Masimbert and H. Sandermanns (Paris).
Hamrick, J. L., Godt, J. W., and Sherman-Broyles, S. L. (1992). Factors influencing levels of genetic diversity in woody plant species. New For. 6, 95–124.
Härdtle, W., Niemeyer, T., Assmann, T., Aulinger, A., Fichtner, A., Lang, A., et al. (2013). Climatic responses of tree-ring width and δ13C signatures of sessile oak (Quercus petraea Liebl.) on soils with contrasting water supply. Plant Ecol. 214, 1147–1156. doi: 10.1007/s11258-013-0239-1
Hartmann, H., Adams, H. D., Hammond, W. M., Hoch, G., Landhäusser, S. M., Wiley, E., et al. (2018a). Identifying differences in carbohydrate dynamics of seedlings and mature trees to improve carbon allocation in models for trees and forests. Environ. Exp. Bot. 152, 7–18. doi: 10.1016/j.envexpbot.2018.03.011
Hartmann, H., Moura, C. F., Anderegg, W. R. L., Ruehr, N. K., Salmon, Y., Allen, C. D., et al. (2018b). Research frontiers for improving our understanding of drought-induced tree and forest mortality. New Phytol. 218, 15–28. doi: 10.1111/nph.15048
Hengl, T., Kempen, B., Heuvelink, G., Malone, B., and Hannes, R. (2017). GSIF: Global Soil Information Facilities. R Package Version 0.5-4 [software]. Available online at: http://gsif.r-forge.r-project.org/
Jonard, M., Nicolas, M., Coomes, D. A., Caignet, I., Saenger, A., and Ponette, Q. (2017). Forest soils in France are sequestering substantial amounts of carbon. Sci. Tot. Env. 574, 616–628. doi: 10.1016/j.scitotenv.2016.09.028
Knutzen, F., Meier, I. C., and Leuschner, C. (2015). Does reduced precipitation trigger physiological and morphological drought adaptations in European beech (Fagus sylvatica L.)? comparing provenances across a precipitation gradient. Tree Physiol. 35, 949–963. doi: 10.1093/treephys/tpv057
Kremer, A. (2000). Changements climatiques et diversité génétique. Rev. For. Francaise 52, 91–98. doi: 10.4267/2042/5408
Kremer, A. (2016). Microevolution of European temperate oaks in response to environmental changes. C. R. Biol. 339, 263–267. doi: 10.1016/j.crvi.2016.04.014
Kremer, A., and Petit, R. (1993). Gene diversity in natural populations of oak species. Ann. Sci. For. 50, 186–202. doi: 10.1051/forest:19930717
Kremer, A., Kleinschmit, J., Cottrell, J., Cundall, E. P., Deans, J. D., Ducousso, A., et al. (2002). Is there a correlation between chloroplastic and nuclear divergence, or what are the roles of history and selection on genetic diversity in European oaks? For. Ecol. Manag. 156, 75–87. doi: 10.1016/S0378-1127(01)00635-1
Kunz, J., Löffler, G., and Bauhus, J. (2018). Minor European broadleaved tree species are more drought-tolerant than Fagus sylvatica but not more tolerant than Quercus petraea. For. Ecol. Manag. 414, 15–27. doi: 10.1016/j.foreco.2018.02.016
Lauteri, M., Pliura, A., Monteverdi, M. C., Brugnoli, E., Villani, F., and Eriksson, G. (2004). Genetic variation in carbon isotope discrimination in six European populations of Castanea sativa mill. originating from contrasting localities. J. Evol. Biol. 17, 1286–1296. doi: 10.1111/j.1420-9101.2004.00765.x
Le Roux, X., Bariac, T., Sinoquet, H., Genty, B., Piel, C., Mariotti, A., et al. (2001). Spatial distribution of leaf water-use efficiency and carbon isotope discrimination within an isolated tree crown. Plant Cell Environ. 24, 1021–1032. doi: 10.1046/j.0016-8025.2001.00756.x
Li, C., Berninger, F., Koskela, J., and Sonninen, E. (2000). Drought responses of Eucalyptus microtheca provenances depend on seasonality of rainfall in their place of origin. Funct. Plant Biol. 27, 231–238. doi: 10.1071/PP99056
Manrique, L. A., and Jones, C. A. (1991). Bulk density of soils in relation to soil physical and chemical properties. Soil Sci. Soc. Am. J. 55:476.
Matías, L., Pérez-Ramos, I. M., and Gómez-Aparicio, L. (2019). Are northern-edge populations of cork oak more sensitive to drought than those of the southern edge? Environ. Exp. Bot. 163, 78–85. doi: 10.1016/j.envexpbot.2019.04.011
Mátyás, C. (2021). Adaptive pattern of phenotypic plasticity and inherent growth reveal the potential for assisted transfer in sessile oak (Quercus petraea L.). For. Ecol. Manag. 482:118832. doi: 10.1016/j.foreco.2020.118832
McDowell, N. G., Bond, B. J., Dickman, L. T., Ryan, M. G., Whitehead, D., Meinzer, F. C., et al. (2011). “Relationships between tree height and carbon isotope discrimination,” in Size- and Age-Related Changes in Tree Structure and Function. Tree Physiology, eds B. Lachenbruch and T. Dawson (Dordrecht: Springer), doi: 10.1007/978-94-007-1242-3_10
McDowell, N., Brooks, J. R., Fitzgerald, S. A., and Bond, B. J. (2003). Carbon isotope discrimination and growth response of old Pinus ponderosa trees to stand density reductions. Plant Cell Environ. 26, 631–644. doi: 10.1046/j.1365-3040.2003.00999.x
Nakagawa, S., Johnson, P. C. D., and Schielzeth, H. (2017). The coefficient of determination R2 and intra-class correlation coefficient from generalized linear mixed-effects models revisited and expanded. J. R. Soc. Interface 14:20170213. doi: 10.1098/rsif.2017.0213
Nguyen-Queyrens, A., Ferhi, A., Loustau, D., and Guehl, J.-M. (1998). Within-ring δ13C spatial variability and interannual variations in wood cellulose of two contrasting provenances of Pinus pinaster. Can. J. For. Res. 28, 766–773.
Niccoli, F., Pelleri, F., Manetti, M. C., Sansone, D., and Battipaglia, G. (2020). Effects of thinning intensity on productivity and water use efficiency of Quercus robur L. For. Ecol. Manag. 473:118282. doi: 10.1016/j.foreco.2020.118282
Perkins, D., Uhl, E., Biber, P., du Toit, B., Carraro, V., Rötzer, T., et al. (2018). Impact of Climate Trends and Drought Events on the Growth of Oaks (Quercus robur L. and Quercus petraea (Matt.) Liebl.) within and beyond their natural range. Forests 9:108. doi: 10.3390/f9030108
Petit, R. J., Brewer, S., Bordács, S., Burg, K., Cheddadi, R., Coart, E., et al. (2002). Identification of refugia and post-glacial colonisation routes of European white oaks based on chloroplast DNA and fossil pollen evidence. For. Ecol. Manag. 156, 49–74. doi: 10.1016/S0378-1127(01)00634-X
Petritan, A. M., Petritan, I. C., Hevia, A., Walentowski, H., Bouriaud, O., and Sánchez-Salguero, R. (2021). Climate warming predispose sessile oak forests to drought-induced tree mortality regardless of management legacies. For. Ecol. Manag. 491:119097. doi: 10.1016/j.foreco.2021.119097
Pflug, E. E., Siegwolf, R., Buchmann, N., Dobbertin, M., Kuster, T. M., Günthardt-Goerg, M. S., et al. (2015). Growth cessation uncouples isotopic signals in leaves and tree rings of drought-exposed oak trees. Tree Physiol. 35, 1095–1105. doi: 10.1093/treephys/tpv079
Pinheiro, J., Bates, D., DebRoy, S., and Sarkar, D. R Core Team. (2020). nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-149 [software]. Available online at: https://CRAN.R-project.org/package=nlme.
Ponton, S., Dupouey, J.-L., and Bréda, N. (2002). Comparison of water-use efficiency of seedlings from two sympatric oak species: genotype x environment interactions. Tree Physiol. 22, 413–422. doi: 10.1093/treephys/22.6.413
Quintana-Seguí, P., Le Moigne, P., Durand, Y., Martin, E., Habets, F., Baillon, M., et al. (2008). Analysis of near-surface atmospheric variables: validation of the SAFRAN analysis over France. J. Appl. Meteorol. Climatol. 47, 92–107. doi: 10.1175/2007JAMC1636.1
R Core Team. (2022). R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing.
Ramirez-Valiente, J. A., Sanchez-Gomez, D., Aranda, I., and Valladares, F. (2010). Phenotypic plasticity and local adaptation in leaf ecophysiological traits of 13 contrasting cork oak populations under different water availabilities. Tree Physiol. 30, 618–627. doi: 10.1093/treephys/tpq013
Rellstab, C., Zoller, S., Walthert, L., Lesur, I., Pluess, A. R., Graf, R., et al. (2016). Signatures of local adaptation in candidate genes of oaks (Quercus spp.) with respect to present and future climatic conditions. Mol. Ecol. 25, 5907–5924. doi: 10.1111/mec.13889
Rice, K. J., Gordon, D. R., Hardison, J. L., and Welker, J. M. (1993). Phenotypic variation in seedlings of a “keystone” tree species (Quercus douglasii) : the interactive effects of acorn source and competitive environment. Oecologia 96, 537–547. doi: 10.1007/BF00320511
Richard, B., Quilès, F., Carteret, C., and Brendel, O. (2014). Infrared spectroscopy and multivariate analysis to appraise α-cellulose extracted from wood for stable carbon isotope measurements. Chem. Geol. 381, 168–179. doi: 10.1016/j.chemgeo.2014.05.010
Robson, T. M., Sánchez-Gómez, D., Cano, F. J., and Aranda, I. (2012). Variation in functional leaf traits among beech provenances during a Spanish summer reflects the differences in their origin. Tree Genet. Genomes 8, 1111–1121. doi: 10.1007/s11295-012-0496-5
Rose, L., Leuschner, C., Köckemann, B., and Buschmann, H. (2009). Are marginal beech (Fagus sylvatica L.) provenances a source for drought tolerant ecotypes? Eur. J. For. Res. 128, 335–343. doi: 10.1007/s10342-009-0268-4
Roussel, M., Le Thiec, D., Montpied, P., Ningre, N., Guehl, J.-M., and Brendel, O. (2009). Diversity of water use efficiency among Quercus robur genotypes: contribution of related leaf traits. Ann. For. Sci. 66, 408–408. doi: 10.1051/forest/2009010
Sáenz-Romero, C., Kremer, A., Nagy, L., Újvári-Jármay, É, Ducousso, A., Kóczán-Horváth, A., et al. (2019). Common garden comparisons confirm inherited differences in sensitivity to climate change between forest tree species. PeerJ 7:e6213. doi: 10.7717/peerj.6213
Sáenz-Romero, C., Lamy, J.-B., Ducousso, A., Musch, B., Ehrenmann, F., Delzon, S., et al. (2017). Adaptive and plastic responses of Quercus petraea populations to climate across Europe. Glob. Change Biol. 23, 2831–2847. doi: 10.1111/gcb.13576
Sáenz-Romero, C., Lindig-Cisneros, R. A., Joyce, D. G., Beaulieu, J., St. Clair, J. B., and Jaquish, B. C. (2016). Assisted migration of forest populations for adapting trees to climate change. Rev. Chapingo Ser. Cienc. For. Ambiente 22, 303–323. doi: 10.5154/r.rchscfa.2014.10.052
Saleh, D., Chen, J., Leplé, J., Leroy, T., Truffaut, L., Dencausse, B., et al. (2021). Genome-wide evolutionary response of European oaks during the Anthropocene. Evol. Lett. 6, 4–20. doi: 10.1002/evl3.269
Scheiner, S. M. (1993). Genetics and evolution of phenotypic plasticity. Annu. Rev. Ecol. Syst. 24, 35–68. doi: 10.1146/annurev.es.24.110193.000343
Searle, S. R., Speed, F. M., and Milliken, G. A. (1980). Population marginal means in the linear model: an alternative to least squares means. Am. Stat. 34, 216–221. doi: 10.2307/2684063
Senf, C., Buras, A., Zang, C. S., Rammig, A., and Seidl, R. (2020). Excess forest mortality is consistently linked to drought across Europe. Nat. Commun. 11:6200. doi: 10.1038/s41467-020-19924-1
Spinoni, J., Vogt, J. V., Naumann, G., Barbosa, P., and Dosio, A. (2018). Will drought events become more frequent and severe in Europe? Int. J. Climatol. 38, 1718–1736. doi: 10.1002/joc.5291
Stokes, V. J., Morecroft, M. D., and Morison, J. I. L. (2010). Comparison of leaf water use efficiency of oak and sycamore in the canopy over two growing seasons. Trees – Struct. Funct. 24, 297–306. doi: 10.1007/s00468-009-0399-8
Sun, X., Kang, H., Chen, H. Y. H., Du, B., Yin, S., Zhou, X., et al. (2016). Phenotypic plasticity controls regional-scale variation in Quercus variabilis leaf δ13C. Trees 30, 1445–1453. doi: 10.1007/s00468-016-1380-y
Torres-Ruiz, J. M., Kremer, A., Carins Murphy, M. R., Brodribb, T., Lamarque, L. J., Truffaut, L., et al. (2019). Genetic differentiation in functional traits among European sessile oak populations. Tree Physiol. 39, 1736–1749. doi: 10.1093/treephys/tpz090
Triboulot, M., Fauveau, M., Bréda, N., Label, P., and Dreyer, E. (1996). Stomatal conductance and xylem-sap abscisic acid (ABA) in adult oak trees during a gradually imposed drought. Ann. Sci. For. 53, 207–220. doi: 10.1051/forest:19960204
Trouvé, R., Bontemps, J. D., Seynave, I., Collet, C., and Lebourgeois, F. (2015). Stand density, tree social status and water stress influence allocation in height and diameter growth of Quercus petraea (Liebl.). Tree Physiol. 35, 1035–1046. doi: 10.1093/treephys/tpv067
Urli, M., Lamy, J.-B., Sin, F., Burlett, R., Delzon, S., and Porté, A. J. (2015). The high vulnerability of Quercus robur to drought at its southern margin paves the way for Quercus ilex. Plant Ecol. 216, 177–187. doi: 10.1007/s11258-014-0426-8
Via, S., Gomulkiewicz, R., De Jong, G., Scheiner, S. M., Schlichting, C. D., and Van Tienderen, P. H. (1995). Adaptive phenotypic plasticity: consensus and controversy. Trends Ecol. Evol. 10, 212–217. doi: 10.1016/S0169-5347(00)89061-8
Vincent-Barbaroux, C., Berveiller, D., Lelarge-Trouverie, C., Maia, R., Máguas, C., Pereira, J., et al. (2019). Carbon-use strategies in stem radial growth of two oak species, one Temperate deciduous and one Mediterranean evergreen: what can be inferred from seasonal variations in the δ13C of the current year ring? Tree Physiol. 39, 1329–1341. doi: 10.1093/treephys/tpz043
Vitali, V., Klesse, S., Weigt, R., Treydte, K., Frank, D., Saurer, M., et al. (2021). High-frequency stable isotope signals in uneven-aged forests as proxy for physiological responses to climate in Central Europe. Tree Physiol. 41, 2046–2062. doi: 10.1093/treephys/tpab062
Vitasse, Y., Bresson, C. C., Kremer, A., Michalet, R., and Delzon, S. (2010). Quantifying phenological plasticity to temperature in two temperate tree species. Funct. Ecol. 24, 1211–1218. doi: 10.1111/j.1365-2435.2010.01748.x
Vitasse, Y., Delzon, S., Bresson, C. C., Michalet, R., and Kremer, A. (2009a). Altitudinal differentiation in growth and phenology among populations of temperate-zone tree species growing in a common garden. Can. J. For. Res. 39, 1259–1269. doi: 10.1139/X09-054
Vitasse, Y., Delzon, S., Dufrêne, E., Pontailler, J.-Y., Louvet, J.-M., Kremer, A., et al. (2009b). Leaf phenology sensitivity to temperature in European trees: do within-species populations exhibit similar responses? Agric. For. Meteorol. 149, 735–744. doi: 10.1016/j.agrformet.2008.10.019
Wang, T., O’Neill, G. A., and Aitken, S. N. (2010). Integrating environmental and genetic effects to predict responses of tree populations to climate. Ecol. Appl. 20, 153–163. doi: 10.1890/08-2257.1
Weigt, R. B., Bräunlich, S., Zimmermann, L., Saurer, M., Grams, T. E. E., Dietrich, H. P., et al. (2015). Comparison of δ18O and δ13C values between tree-ring whole wood and cellulose in five species growing under two different site conditions. Rapid Commun. Mass Spectrom. 29, 2233–2244. doi: 10.1002/rcm.7388
Zhang, J. W., and Cregg, B. M. (1996). Variation in stable carbon isotope discrimination among and within exotic conifer species grown in eastern Nebraska. USA. For. Ecol. Manag. 83, 181–187. doi: 10.1016/0378-1127(96)03723-1
Zhang, J. W., and Marshall, J. D. (1995). Variation in carbon isotope discrimination and photosynthetic gas exchange among populations of pseudotsuga menziesii and pinus ponderosa in different environments. Funct. Ecol. 9:402. doi: 10.2307/2390003
Keywords: local adaptation, soil texture, vapor pressure deficit, phenotypic plasticity, wood carbon isotope composition, social status, relative extractable water, drought response strategy
Citation: Rabarijaona A, Ponton S, Bert D, Ducousso A, Richard B, Levillain J and Brendel O (2022) Provenance Differences in Water-Use Efficiency Among Sessile Oak Populations Grown in a Mesic Common Garden. Front. For. Glob. Change 5:914199. doi: 10.3389/ffgc.2022.914199
Received: 06 April 2022; Accepted: 02 June 2022;
Published: 01 July 2022.
Edited by:
Henry D. Adams, Washington State University, United StatesReviewed by:
Drew Peltier, Northern Arizona University, United StatesCopyright © 2022 Rabarijaona, Ponton, Bert, Ducousso, Richard, Levillain and Brendel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stéphane Ponton, c3RlcGhhbmUucG9udG9uQGlucmFlLmZy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.