 Shunsuke Tei
Shunsuke Tei Ayumi Kotani
Ayumi Kotani Atsuko Sugimoto3,4,5
Atsuko Sugimoto3,4,5 Nagai Shin
Nagai Shin- 1Department of Forest Management, Forestry and Forest Products Research Institute, Tsukuba, Japan
- 2Graduate School of Bioagricultural Sciences, Nagoya University, Nagoya, Japan
- 3Arctic Research Center, Hokkaido University, Sapporo, Japan
- 4Faculty of Environmental Earth Science, Hokkaido University, Sapporo, Japan
- 5Global Station for Arctic Research, Global Institution for Collaborative Research and Education, Hokkaido University, Sapporo, Japan
- 6Research Institute for Global Change, Japan Agency for Marine-Earth Science and Technology (JAMSTEC), Yokohama, Japan
Terrestrial forest ecosystems are crucial to the global carbon cycle and climate system; however, these ecosystems have experienced significant warming rates in recent decades, whose impact remains uncertain. This study investigated radial tree growth using the tree-ring width index (RWI) for forest ecosystems throughout the Northern Hemisphere to determine tree growth responses to autumn climate change, a season which remains considerably understudied compared to spring and summer, using response function and random forest machine learning methods. Results showed that autumn climate conditions significantly impact the RWI throughout the Northern Hemisphere. Spatial variations in the RWI response were influenced by geography (latitude, longitude, and elevation), climatology, and biology (tree genera); however, geographical and/or climatological characteristics explained more of the response compared to biological characteristics. Higher autumn temperatures tended to negatively impact tree radial growth south of 40° N in regions of western Asia, southern Europe, United State of America and Mexico, which was similar to the summer temperature response found in previous studies, which was attributed to temperature-induced water stress.
Introduction
Forest ecosystems play a vital role in the global carbon cycle (Pan et al., 2011) and climate systems (e.g., Bonan, 2008). However, since the early 1970s, forest ecosystems across the Northern Hemisphere have experienced varying warming rates. In particular, the warming rates across northern high-latitude regions are higher than anywhere else on earth, a phenomenon known as Arctic amplification (e.g., Serreze and Barry, 2011). This amplified warming has a potentially significant impact on forest ecosystems (e.g., Tei et al., 2019a, d; Tei and Sugimoto, 2020).
An important role of forest ecosystems is mitigating the accumulation of atmospheric CO2 (Fan et al., 1998; Bousquet et al., 2000). However, there is significant uncertainty regarding the magnitude and location of the forest ecosystem CO2 sink (Goodale et al., 2002; Ciais et al., 2010). Consequently, understanding spatial variations in the forest ecosystem response to climate change over a vast biome or region with significant gradients in geographic, climatological, and biological characteristics are crucial for accurate projections of the terrestrial carbon cycle and global climate.
A dendroecological analysis helps study the forest ecosystem response to a changing environment (e.g., Walker et al., 2015; Tei et al., 2015, 2019b). Tree-ring width indices (RWI) are useful long-term indicators of historical forest productivity due to the frequently observed close relationship of the RWI with forest-level gross primary production (GPP) and/or net ecosystem exchange (NEE) (e.g., Xu et al., 2017; Tei et al., 2019c). A recent synthetic analysis of RWI data from the International Tree-Ring Data Bank (ITRDB) confirmed the widespread negative sensitivity of radial tree growth to warming in circumboreal forests (Tei et al., 2017a, b), which is particularly prevalent in dry continental regions, such as the interiors of Alaska and Canada, southern Europe, and central-eastern Siberia. These researches have indicated a one-year time lag in the forest ecosystem response to climate change in these regions, which is considered to be an adaptation of the forest ecosystem to a severely water-stressed environment (e.g., Chapin et al., 1990; McDowell et al., 2008; Genet et al., 2010; Wiley and Helliker, 2012).
To date, broad-scale RWI analysis (e.g., Tei et al., 2017a, b) and satellite remote sensing studies (e.g., Wang et al., 2011; Buermann et al., 2014) have mainly focused on tree radial growth/ecosystem responses to climate change during the spring and/or summer. However, recent studies have reported that climate variables in other seasons can significantly affect tree radial growth and ecosystem activity in the northern high-latitude region (Tei and Sugimoto, 2018). While there have been recent improvements in our understanding of the importance of cold season climate drivers (e.g., Wu et al., 2018), there is still a lack of information regarding the spatial distribution of seasonal climate variables that control temporal variations in RWI. Furthermore, there is a lack of understanding with respect to whether the tree radial growth response to climate change varies by tree genera or species because very few RWI studies have evaluated differences in the climate change response and the genus or species scale (Lloyd and Bunn, 2007).
In this study, we investigate the geographical (latitude, longitude, and/or elevation), climatological and biological (genus and/or species) characteristics of the tree radial growth response to climate change during the autumn over a wide region/biome with significant geographical, climatological and biological gradients. We achieve this by applying statistical response function analysis and the random forest machine learning method to RWI data and autumn climate variables, including temperature, precipitation, and cloud cover, over the past 120 years.
Materials and Methods
Tree-Ring Data
We collected raw tree-ring-width chronologies from 2894 ITRDB sites (1 accessed during spring–winter 2020) located in the Northern Hemisphere and with a chronology that started before 1950 and ended after 1990. The standard RWI chronology was generated for each site by using R (R Core Team, 2017) and the “dplR” package. We detrended the growth patterns associated with tree age and natural disturbances in the raw time series, derived from the single dominant tree species for each site, using cubic smoothing splines and, subsequently, averaging the standardized ring widths among the samples. Spline fits with a length of 128 years were applied to raw time series. Generally, standardization methods are varied among chronologies/sites; however, in this study, we applied spline fitting to standardize the chronologies across all the sites, which was done because our approach is conservative, and the method is regarded as relatively site-insensitive (Cook, 1985; Cook and Kairiukstis, 1990).
The quality of RWI was assessed using the expressed population signal (EPS). The EPS is a measure of how well a finite sample of tree-ring data represents an infinite population chronology, and an EPS value of 0.85 is considered a threshold of acceptable statistical quality for dendrochronological analysis (Wigley et al., 1984). The EPS values for 2167 of the 2894 sites were higher than 0.85, indicating a high degree of common variability between individual trees. Therefore, for subsequent analysis, we used the RWI for these 2167 sites.
Climate Data
Gridded monthly near-surface average temperature, cloud cover (the fraction of the sky obscured by clouds), and total precipitation datasets from the Climate Research Unit (CRU TS 4.01; 1901–2016; 0.5° × 0.5°; Harris et al., 2014) were used to derive monthly climate records from grids over the RWI sites.
Response Function Analysis
To evaluate the impact of a target climate variable (among temperature, precipitation, and cloud cover) in autumn independently from other seasons, i.e., winter, spring and summer, and other climate variables, we applied response function (RF) analysis for RWI and seasonally averaged climate variables for the 2167 RWI sites over the vast biome/region of the Northern Hemisphere. In this RF analysis, the climate conditions of the current four seasons (from winter to autumn) and previous summer and autumn were used because existing RWI and satellite image studies have shown that the climate conditions of the previous year substantially affected tree radial growth, especially in arid regions (Kagawa et al., 2003, 2006; Tei et al., 2013a; Tei and Sugimoto, 2018). The period when both RWI and climate data were available (1901–2016) was used for the analysis.
RF represents a result of principal component (PCs) analysis in which climate variables are decorrelated (Fritts, 1976; Cook and Kairiukstis, 1990). In the RF analysis, the multi-collinearity among seasonal climatic parameters (3 climate variables with six seasons: previous summer and autumn, and current winter, spring, summer, and autumn) was reduced by extracting principal components (PCs). The eigenvectors with relatively low values calculated from the seasonal climate variables are excluded. The relationships between RWI and the PCs of the seasonal climate variables were back-transformed to relationships with the seasonal climate variables, i.e., the coefficient of RF analyses. The RF analysis was applied using R and the “treeclim” package.
With reference to previous studies of plant seasons in boreal/arctic ecosystems (e.g., Morozumi et al., 2020; Nagai et al., 2020), we assumed that the definitions of each season varied depending on the latitude of the RWI site. For sites located between 0 and 60° N, spring was considered to be March, April, and May; summer was June, July, and August; autumn was September, October, and November; and winter was December, January, and February. For sites located between 60 and 67° N, spring was considered to be April and May; summer was June, July, and August; autumn was September and October; and winter was November, December, January, February, and March. For sites located north of 67° N, spring was considered to be May and June; summer was July; autumn was August and September; and winter was October, November, December, January, February, March, and April.
Relationship Between RWI Response to Autumn Climate Change and Latitude/Climate Condition
We used coefficients from the RF analysis for each RWI site to investigate how geographical (latitude, longitude, and elevation), climatological (temperature, precipitation, and cloud cover), and biological (tree genus and species) characteristics impacted the RWI response to autumn climate change. We used the climate conditions throughout the year instead of autumn to consider the typical climate condition of each RWI site. In addition, only those RWI sites with significant RF coefficients for autumn climate variables were included in the analysis. To ensure a sufficient number of sites for each tree genus and species, we further limited our analysis to those RWI sites with tree genera observed at 90 or more sites and species observed at 5 or more sites. We then averaged the RWI site latitude, annual averaged temperature and cloud cover, annual cumulative precipitation, and RF coefficients by tree species and plotted the relationships to identify the most meaningful relationships. The tree genera and species included in our analysis are summarized in Table 1.
Table 1. Tree genera (tree species) that were observed at more than 90 (5) tree-ring width index (RWI) sites that had a significant response function coefficient for autumn climate variables (temperature, precipitation, and cloud cover).
Random Forest Algorithm
To quantify the relative importance of geographical, climatological, and biological characteristics on the RWI response to each autumn climate variable, we applied the random forest algorithm to the filtered RWI data (refer to section “Relationship Between RWI Response to Autumn Climate Change and Latitude/Climate Condition”). Then, we computed the mean decrease in the Gini importance for each geographical, climatological and biological characteristic after training. The random forest algorithm was applied using R and the “randomForest” package.
Random forest is a supervised learning algorithm, an ensemble of decision trees (Zhao, 2012) commonly trained using the bootstrap aggregation or “bagging” method. The algorithm builds multiple decision trees and merges them to achieve an overall result, generally providing a more accurate and stable prediction. In recent years, the random forest method has also been applied to carbon cycle research in forest ecosystems (e.g., Cai et al., 2020).
A positive attribute of the random forest algorithm is that it is easy to measure the relative importance of each feature (Gini importance). Because the random forest is an ensemble of individual decision trees, Gini importance can be leveraged to calculate the mean decrease in Gini, which is a measure of variable importance for estimating a target variable. This is an effective measure of the importance of a variable in estimating the value of the target variable across all trees that make up the forest. A higher mean decrease in Gini indicates remarkable variable importance.
Geographical, climatological, and biological characteristics were used as explanatory variables to ensure the reliability of the random forest models. Climatological characteristics included annual averaged temperature and cloud cover and annual cumulative precipitation. A total dataset for each climate variable was randomly divided into two datasets, comprising 80 and 20% of the total number of RWI sites. The former was prepared for random forest model training, while the latter was used for model verification. Finally, we confirmed that the RF coefficient for each climate variable in the autumn season, estimated from each random forest model using geographical, climatological and biological characteristics, showed significant correlation with the coefficients in the verification dataset (temperature: r = 0.77, p < 0.01; precipitation: r = 0.49, p < 0.01; cloud cover: r = 0.73, p < 0.01), demonstrating the reliability of our model. Finally, we built an additional similar random forest model for the widely distributed Pinus genus and confirmed the reliability of each random forest model (temperature: r = 0.72, p < 0.01; precipitation: r = 0.40, p < 0.01; cloud cover: r = 0.63, p < 0.01).
Results
Reginal Characteristics of the RWI Response to Autumn Climate Change
Figures 1A–C shows the significant RF coefficients for each autumn climate variable (temperature, precipitation, and cloud cover). We found that positive (negative) coefficients for autumn temperature were dominant over higher (lower) latitude regions (Figure 1A). The negative RF coefficients were distributed mainly in the United States of America and are mainly derived from the autumn temperature in the previous year (Figures 2A, 3A). Interestingly, almost no significant RF coefficients with autumn temperature were observed at sites north of latitude 50° N for the Siberian and Asian regions (Figure 1A). The opposite spatial pattern of the RF coefficients with temperature was observed for cloud cover (Figure 1C). Positive RF coefficients were distributed mainly in the United States of America (Figure 1C), and were mainly derived from autumn cloud cover in the previous year (Figures 2C, 3C). The RF coefficients for autumn precipitation were positive over a broader geographic region than temperature and cloud cover (Figure 1B), derived mainly from autumn precipitation in the previous year (Figures 2B, 3B).
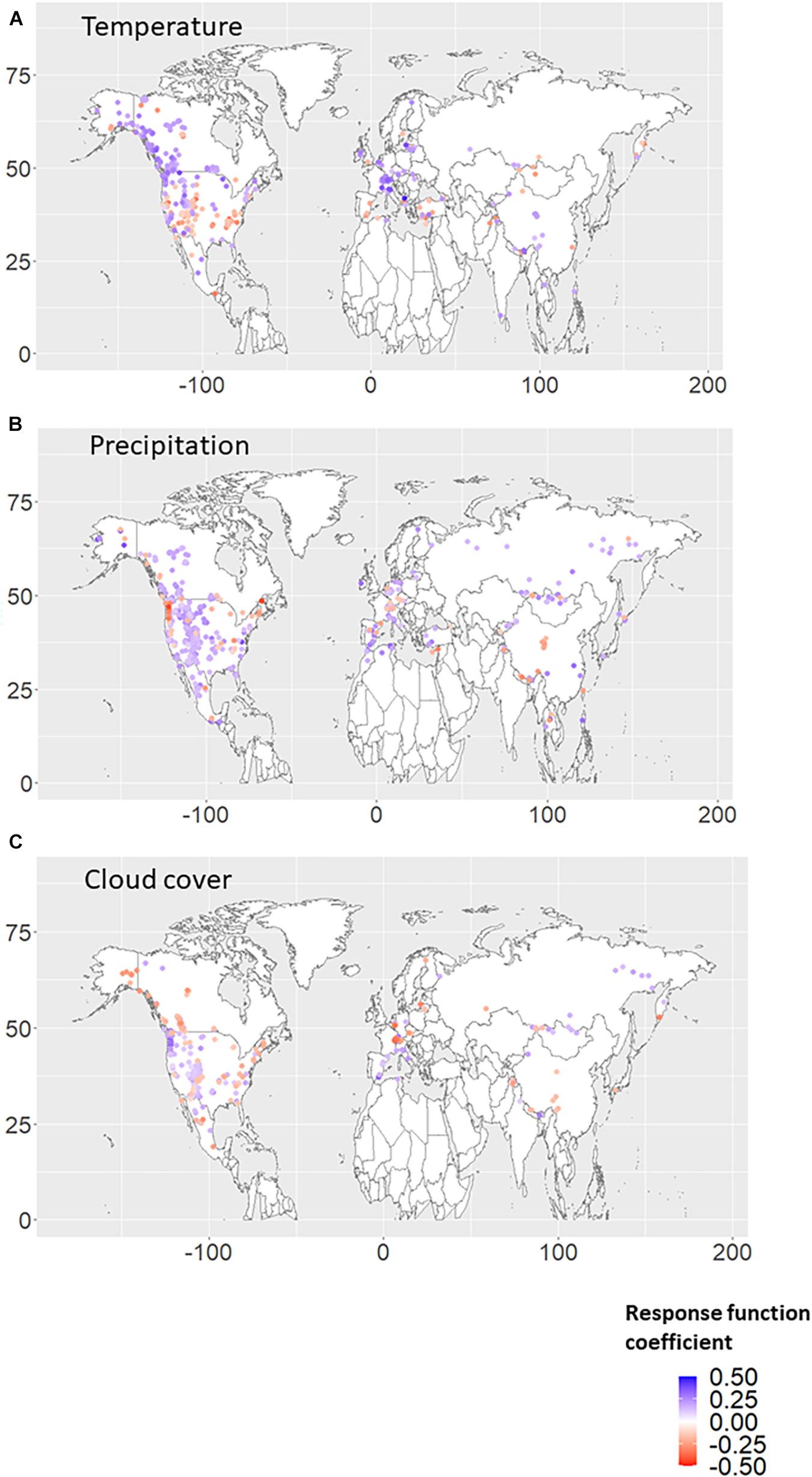
Figure 1. Response function (RF) coefficients between RWI and climate variables, i.e., (A) temperature, (B) precipitation, and (C) cloud cover, in the autumn season. The period when both RWI and climate data existed in duplicate during 1901–2016 was used for the analysis. Only the RWI sites where the RF coefficient was significant (p < 0.10) are plotted. Climate conditions from the current autumn and previous autumn were used for the RF analysis. If RF coefficients were significant in both the previous and current autumn, the larger absolute value is plotted.
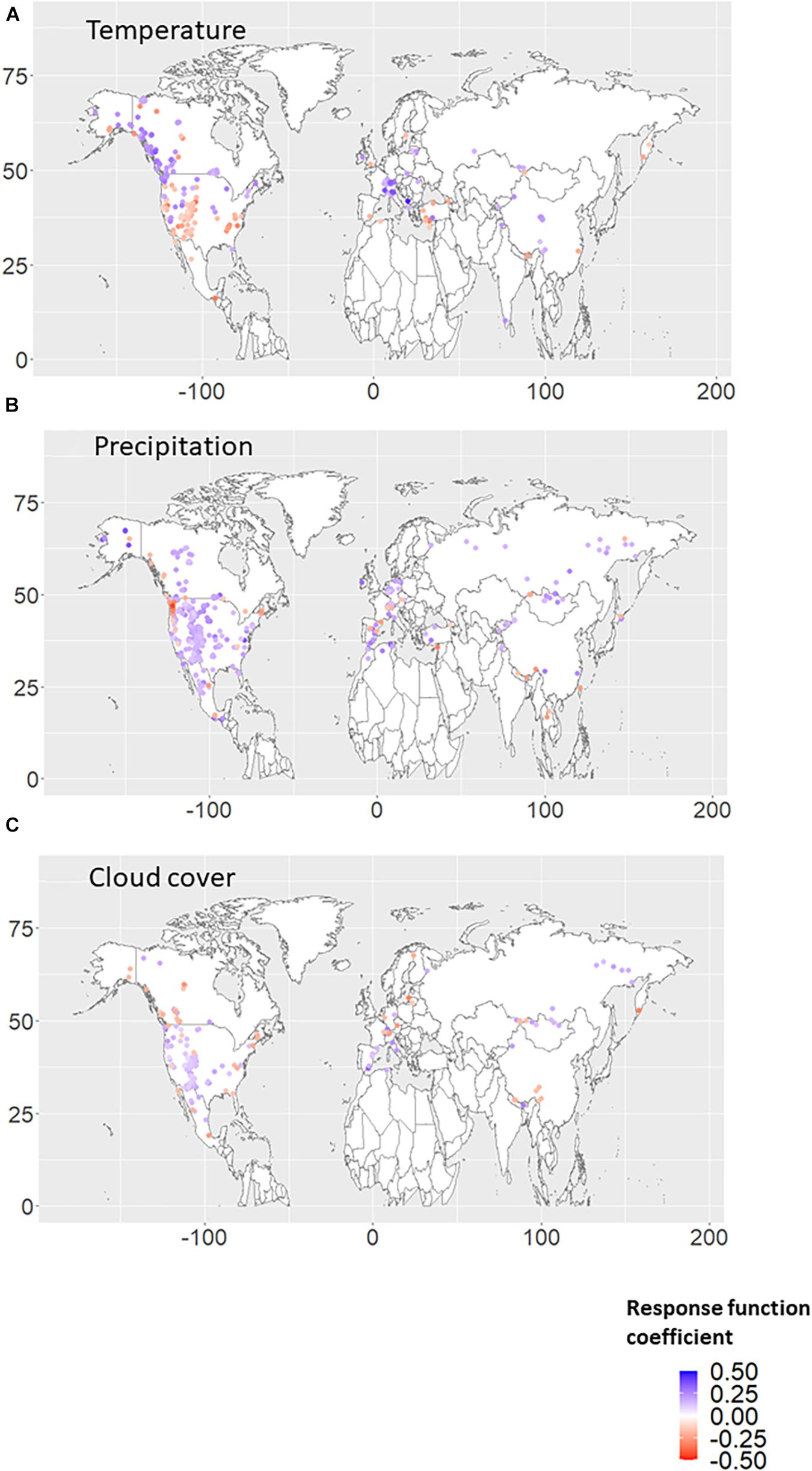
Figure 2. Response function (RF) coefficients between RWI and climate variables, i.e., (A) temperature, (B) precipitation, and (C) cloud cover, in the autumn season of the previous year. The period when both RWI and climate data existed in duplicate during 1901–2016 was used for the analysis. Only the RWI sites where the RF coefficient was significant (p < 0.10) are plotted.
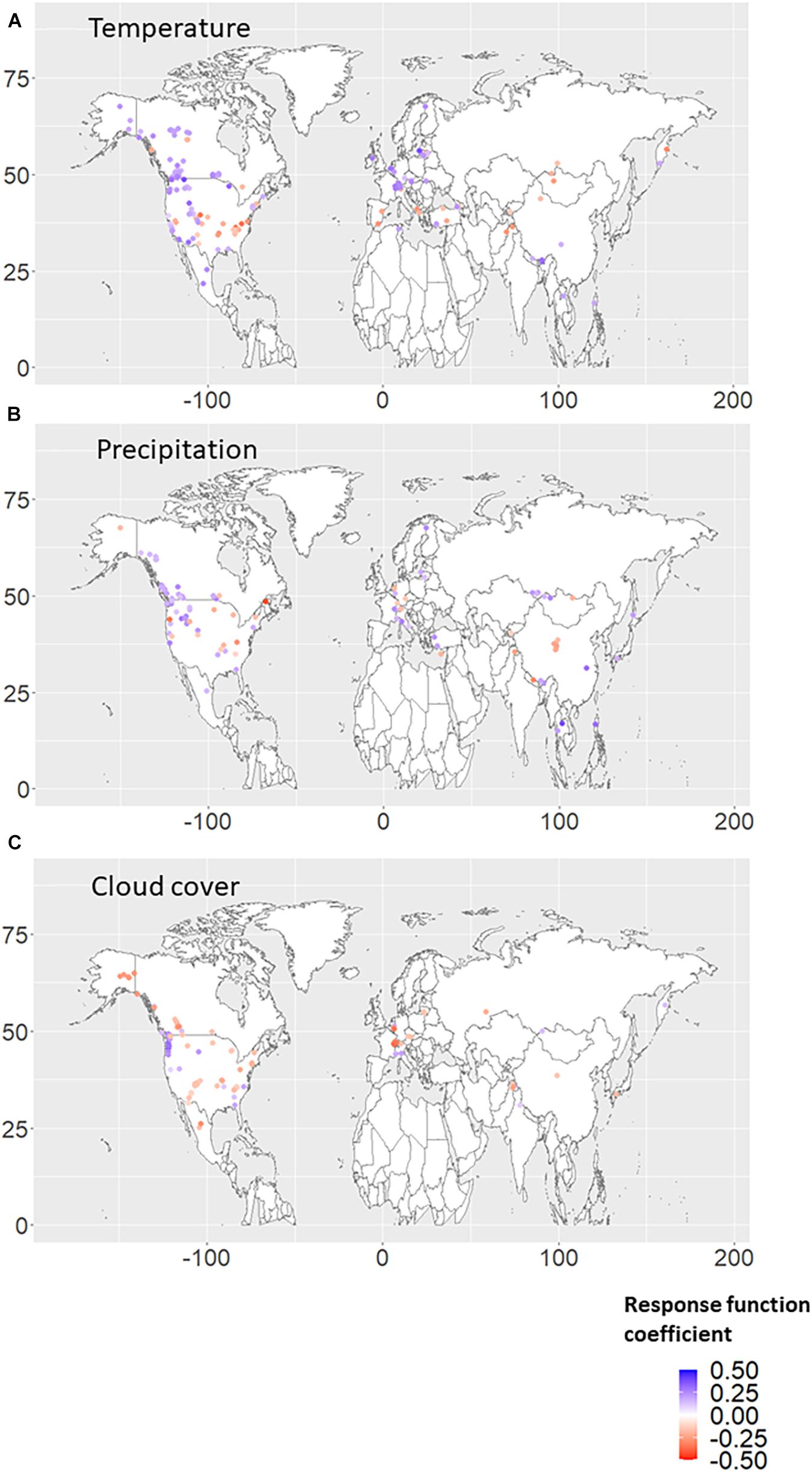
Figure 3. Response function (RF) coefficients between RWI and climate variables, i.e., (A) temperature, (B) precipitation, and (C) cloud cover, in the autumn season of the current year. The period when both RWI and climate data existed in duplicate during 1901–2016 was used for the analysis. Only the RWI sites where the RF coefficient was significant (p < 0.10) are plotted.
Table 2, based on the results from Figure 1, summarizes the percentage of sites showing positive or negative coefficients for each climate variable by region. Positive RWI responses to autumn temperature were more dominant in Europe (except for southern Europe), eastern Asia, and Canada, whereas negative responses were more dominant in western Asia, southern Europe, the United States of America, and Mexico. A positive RWI response to autumn precipitation was predominant in all regions. The RWI responses to cloud coverage were more regional, with RWI sites showing predominantly negative responses in Europe. These negative responses might be related to moist environments from moist westerly air in the European region.
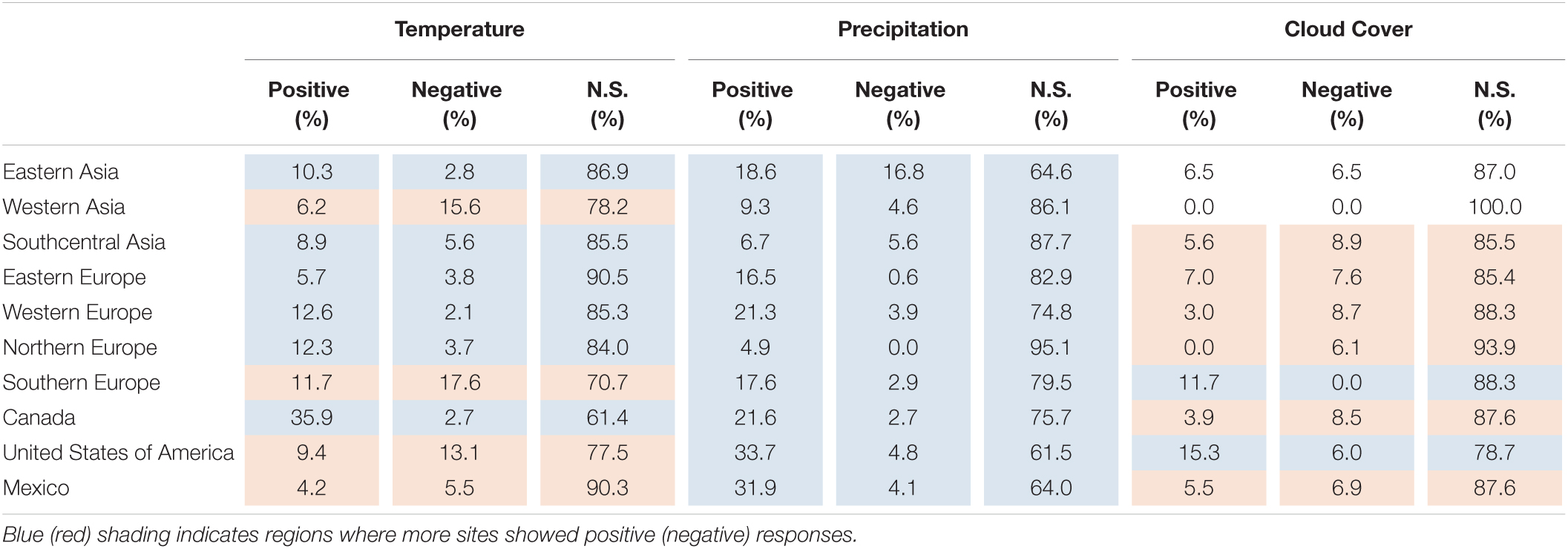
Table 2. Percentage of RWI sites with significant positive or negative response function coefficients for each autumn climate variable (temperature, precipitation, and cloud cover) summarized by region.
Biological Characteristics of the RWI Response to Autumn Climate Change
Table 3, which is also based on the results from Figure 1, summarizes the percentage of RWI sites showing positive or negative regression coefficients by tree genus. Positive RWI responses to autumn temperature and precipitation were more dominant for most tree genera; however, negative responses to autumn temperature and precipitation were more common for Pinus and Pseudotsuga, and Juniperusa and Tsuga, respectively. Conversely, negative RWI responses to the autumn cloud cover were more dominant for most tree genera; however, positive responses were more common for Pinus, Tsuga, and Pseudotsuga.
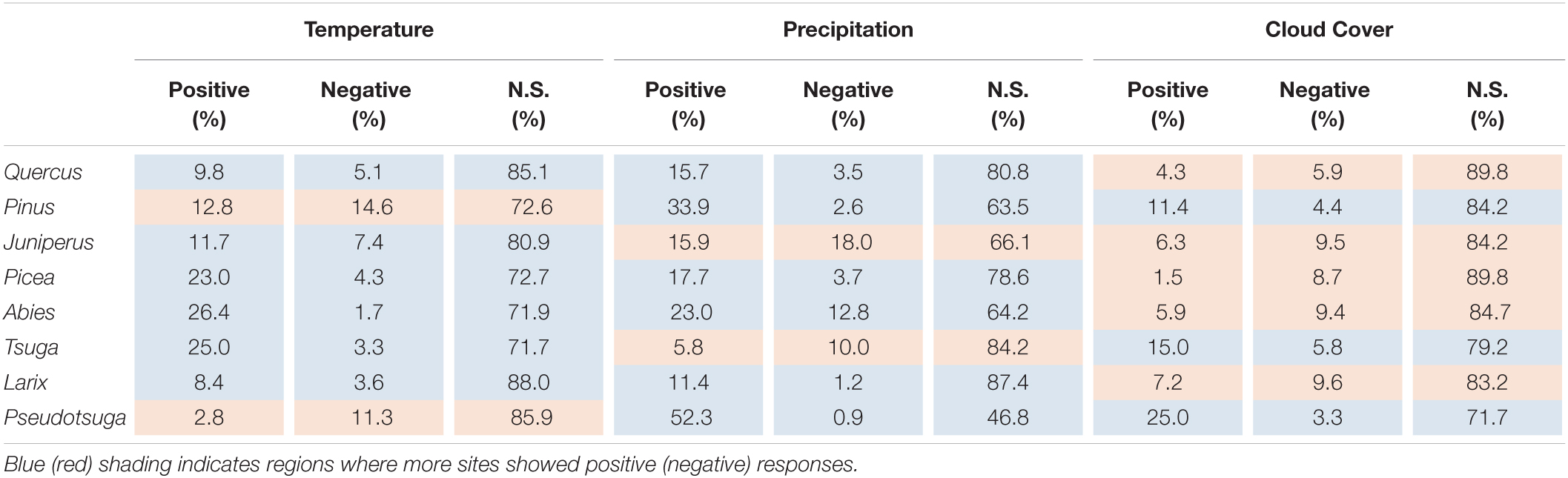
Table 3. Percentage of RWI sites with significant positive or negative response function coefficients for each autumn climate variable (temperature, precipitation, and cloud cover) summarized by tree genera.
Variations in RWI Response to Autumn Climate Change With Latitude
We observed clear relationships between the latitude of the RWI site and the RF coefficients of each climate variable during autumn (Figure 4). Except for the Juniperus genus, negative RF coefficients for autumn temperature were dominant for the sites lower than 40° N, mainly consisting of Pinus trees (Figure 4A). Autumn precipitation showed a positive RF coefficient, but the variability varied widely, especially around 50° N (Figure 4B). In addition, the positive RF coefficients tended to be stable at the south of latitude 40° N. Unlike the response to autumn temperature and precipitation, the RWI response to autumn cloud cover in relation to the RWI site latitude was unclear but tended to show a positive RF coefficient for the sites at the south of 40° N (Figure 4C).
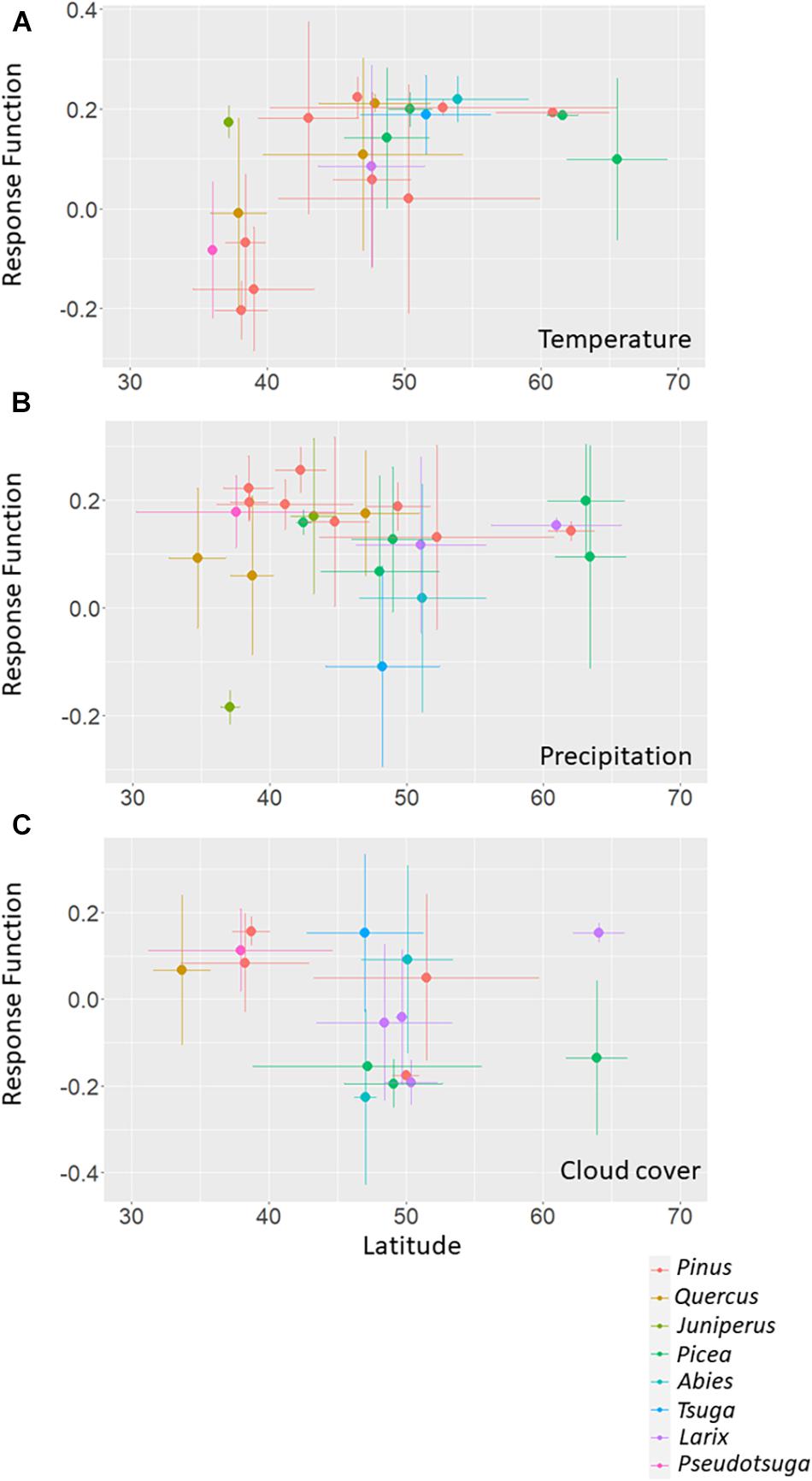
Figure 4. Relationship between the latitude and the response function (RF) coefficients for (A) temperature, (B) precipitation, and (C) cloud cover, in autumn season. Both the site latitude and RF coefficient are averaged by tree species. Colors indicate tree genera. To ensure a sufficient number of sites for each tree genera and species, we limited our analysis to RWI sites with tree genera observed at 90 or more sites and with tree species observed at 5 or more sites from those sites with significant RF coefficients for each autumn climate variable.
Pinus trees covered a wide range of latitudes and showed positive and negative relationships between species-based averaged site latitude and the RF coefficients for autumn temperature, and precipitation and cloud cover, respectively. Nevertheless, this relationship was less clear for other tree genera.
Variations in RWI Response to Autumn Climate Change With the Climatic Condition
We also observed relationships between RF coefficients of RWI with autumn climate variables and the annual average or sum of the climate variables (Figures 5A–C). The negative RF coefficients for autumn temperature were dominant for the sites with annually averaged temperatures exceeding 5 °C, especially for Pinus trees (Figure 5A). Autumn precipitation tends to show a positive RF coefficient with RWI and shows a negative relationship with annual cumulative precipitation (Figure 5B). Unlike the response to autumn temperature and precipitation, the RWI response to autumn cloud cover in relation to the annual averaged cloud cover is unclear but tends to show a positive RF coefficient for the sites with annual averaged cloud cover lower than 55% (Figure 5C).
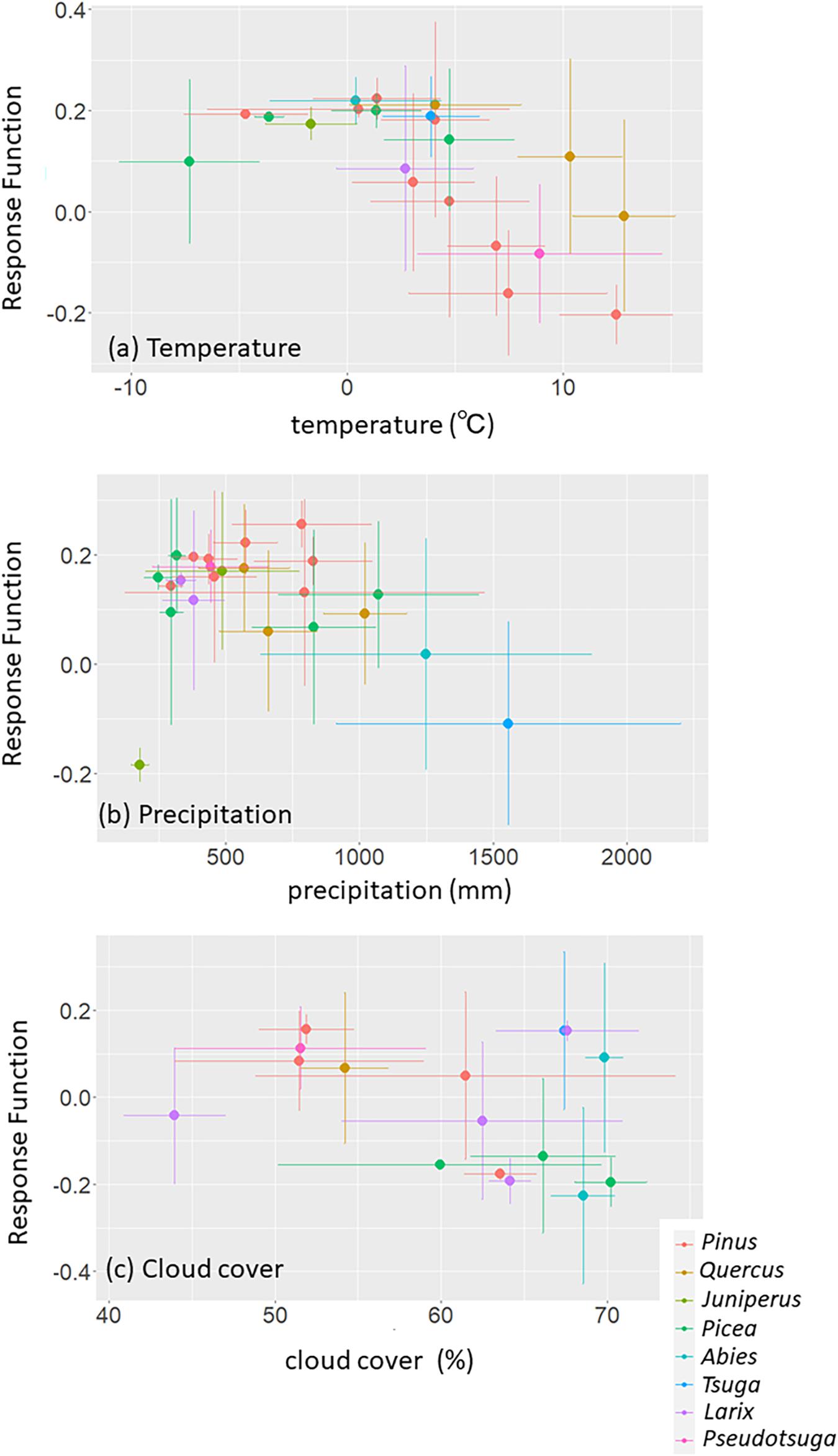
Figure 5. Relationship between the climatic condition and the response function (RF) coefficients for (A) temperature, (B) precipitation, and (C) cloud cover, in autumn season. We used annual averaged temperature/cloud cover and annual cumulative precipitation. Both the site climatic condition and coefficient are averaged or cumulative by tree species. Colors indicate tree genera. To ensure a sufficient number of sites for each tree genera and species, we limited our analysis to RWI sites with tree genera observed at 90 or more sites and with tree species observed at 5 or more sites from those sites with significant RF coefficients for each autumn climate variable.
Importance of Geographical, Climatological, and Biological Characteristics on RWI Response to Autumn Climate Change
The reliability of the random forest model differed for each autumn climate variable, with temperature having the highest reliability (r = 0.77, p < 0.01), followed by cloud cover (r = 0.73, p < 0.01) and precipitation (r = 0.49, p < 0.01). Conversely, the importance (i.e., the mean decrease in the Gini coefficient) of each explanatory variable was relatively similar for all climate variables (Table 4), with geographical (latitude, longitude, and elevation) or climatological (annual averaged temperature/cloud cover, and annual cumulative precipitation) characteristics having the highest importance, and followed by biological characteristics (tree genus). In other words, latitude and annual cumulative precipitation have the highest explanatory power for the RWI response to autumn temperature/cloud cover and precipitation, respectively, whereas the genus has the lowest explanatory power for all climate variables.
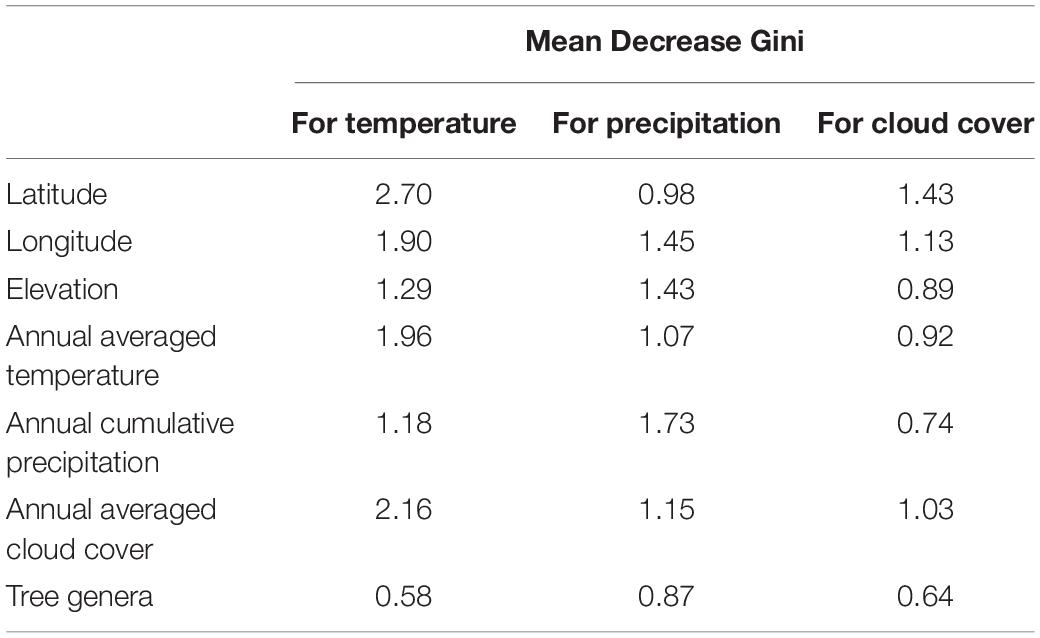
Table 4. Mean Gini coefficient decrease for geographical (latitude, longitude, and elevation), climatological (annual averaged temperature and cloud cover, and annual cumulative precipitation), and biological (tree genus) characteristics in random forest models explaining response function coefficients of the RWI with autumn climate variables.
When the random forest analysis was limited to the Pinus genera with a broad latitudinal distribution, the tree species significantly impacted the model, especially concerning the RWI response to autumn temperature (Table 5). We could not develop such random forest model for other tree genera due to the small number of tree species included in the model.
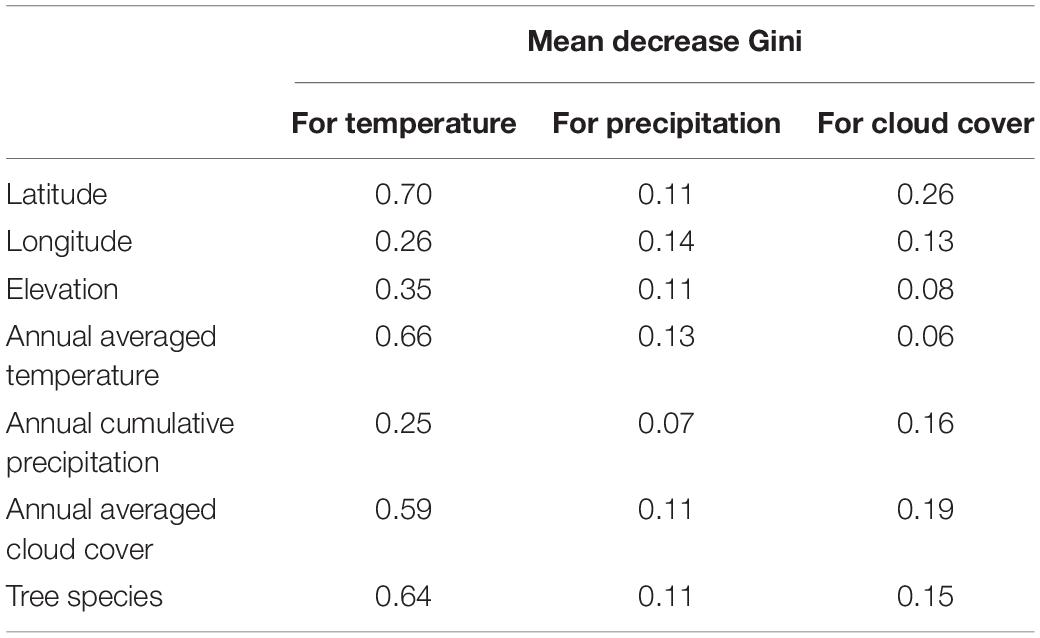
Table 5. Mean Gini coefficient decrease for geographical (latitude, longitude, and elevation), climatological (annual averaged temperature and cloud cover, and annual cumulative precipitation), and biological (tree species) characteristics in random forest models limited to the Pinus tree genus explaining the response function coefficients of the RWI with autumn climate variables.
Discussion
Negative Impacts of Autumn Temperature on RWI in Lower Latitude Regions Across the Northern Hemisphere
The RWI response to autumn climate change showed evident regional characteristics (Figure 1 and Table 2), and the response changed significantly with latitude (Figure 4) and climate condition (Figure 5). Negative responses of RWI to autumn temperature were dominant south of 40° N (western Asia, southern Europe, United State of America and Mexico; Table 2), where Pinus was the dominant genus (Figure 4A). There are several possible attributions for this negative response, including direct temperature stress (D’Arrigo et al., 2004), temperature-induced drought stress (Barber et al., 2000), temperature-induced respiratory carbon losses (Piao et al., 2008; Liu et al., 2018), and changes in the timing of autumn leaf senescence (Jeong et al., 2011). Previous studies also stated that the previous autumn–current spring climate affects tree physiology through changes in the timing of spring-snow melting and/or nitrogen availability in the following summer (e.g., Kirdyanov et al., 2003; Sidorova et al., 2009).
However, considering the dominant positive response of the RWI to autumn precipitation across lower latitude sites (Figures 1B, 4B), temperature-induced drought stress seems to be the most reasonable cause of the negative RWI response to autumn temperature. The observed inverse pattern of RF coefficient for RWI with autumn temperature and cloud coverage (Figures 1A,C) may also support this hypothesis. If there are many cloudy days, evapotranspiration of water from the soil surface might be suppressed. However, it should be noted that this interpretation is for sites with relatively warm environments (Figure 5A) mainly distributed south of 40° N latitude, not the entire Northern Hemisphere high latitudes.
Similar patterns of the RWI response to autumn temperature (Figures 1–5A) have been reported with respect to vegetation activity responses to summer climate change across southern boreal forests and continental dry climate regions (e.g., Nikolaev et al., 2009; Silva et al., 2009; Dulamsuren et al., 2010; Andreu-Hayles et al., 2011; Tei et al., 2014, 2017b). These response patterns were also attributed to temperature-induced drought stress caused by a reduction in the stomatal conductance of plants under limited soil moisture availability (Barber et al., 2000). Compared to summer, moisture stress could be more severe in the late summer and autumn, especially in dry regions. More water is usually available for vegetation in spring due to the infiltration of snowmelt water, which then gradually decreases from spring to summer and autumn (Sugimoto et al., 2003). Consequently, the impact of soil moisture on stomatal conductance is low during the spring and then increases in late summer and autumn (Tei et al., 2013b). The authors reported that the signal of the severe water stress in the late growing season had been recorded in the latewood ring δ13C chronologies of larch trees in eastern Siberia. In addition, latewood ring δ18O for pine trees in southwestern China reportedly correlates with autumn precipitation (Fu et al., 2017).
It is also should be mentioned that the significant negative (positive) RF coefficients of the RWI with autumn temperature (precipitation and cloud cover) tend to be calculated from the climate variables of the previous year (Figure 2). Previous dendroecological studies (e.g., Tei et al., 2017b; Tei and Sugimoto, 2018) has indicated such a one-year time lag in the response of the forest ecosystem to climate change in continental climate regions over circumboreal forests, which is regarded as the adaptation of the forest ecosystem to the severely water-stressed environment (e.g., Chapin et al., 1990; McDowell et al., 2008; Genet et al., 2010; Wiley and Helliker, 2012).
Juniperus trees were common south of 40° N; however, they showed an opposite response pattern to Pinus; that is, positive and negative responses to autumn temperature and precipitation, respectively (Figures 4A,B), likely because Juniperus tend to grow at higher elevations (2000–4500 m a.s.l.) compared to other tree genera. Due to the lower temperatures at higher altitudes (Figure 5A), tree growth is considered to be limited by temperature. Positive vegetation response to warming in cool environments has been frequently reported in RWI (Tei et al., 2017a, b) and normalized difference vegetation index studies (e.g., Buermann et al., 2014; Wu et al., 2015). Furthermore, northward and upward shifts of tree lines under warming conditions in forest ecosystems have also been reported (Shiyatov et al., 2005, Devi et al., 2008).
There is a growing body of evidence to suggest that the response of tree radial growth to summer temperature tends to be positive in higher latitude regions such as the Arctic, whereas it tends to be negative in lower latitude regions such as southern boreal forests and continental dry climate regions (Tei et al., 2017b; Tei and Sugimoto, 2020). In the current study, we investigated the spatial distribution of tree radial growth responses to autumn climate change and obtained results corresponding with those of the summer.
Less Substantial Impact of Tree Genera on RWI Response to Autumn Climate Change
It is unclear whether the RWI response to climate change varies by tree genus or species. Lloyd and Bunn (2007) reported on different tree radial growth responses to climate change (considering all seasons, but not revealing the impact of each season) by tree species over a circum-boreal forest (>55° N). They found that Picea (Picea abies, Picea glauca, Picea mariana, and Picea obovata) and Pinus banksiana tended to show inverse responses to temperature, whereas Larix (Larix gmelinii and Larix siberica), Pinus (Pinus sylvestris and Pinus sitchensis), and Tsuga mertensiana tended to show a positive response. However, our study focused on the autumn climate and a target region covering the entire Northern Hemisphere and did not show the same tree species characteristics. Instead, we showed predominantly negative responses to autumn temperature from several species of Pinus trees below 40° N (Figure 4A).
These results indicate that tree radial growth responses to climate change may differ with the season, and the impact of the genus is less critical compared to other geographic and climatological factors, although we did show that some characteristics of the RWI response to autumn climate change were due to genera differences (Table 3). This interpretation is strongly supported by the low importance of genera in the random forest models (Table 4).
We observed a more sensitive response to autumn temperature and precipitation with latitude and climate conditions for Pinus than other genera (Figures 4A,B, 5A,B). Furthermore, when the random forest model was applied to Pinus only, the importance of tree species was improved, especially for temperature (Table 5). We could not develop such an individual random forest model for other tree genera due to the small number of tree species included in the model. In our study, Pinus covered a wider latitude range compared to other tree genera (Figure 4); therefore, more observation points for other tree genera are needed to determine whether the higher importance of tree species is a natural characteristic of the Pinus genus, or it could be applied to the other genera.
The distribution of RWI sites is biased toward Europe and North America. Therefore, it is likely that our findings will have some regional bias. Further efforts to fill the gaps in RWI data, such as in central and western Canada and Siberia, are required. Notwithstanding, this study contributes to our understanding of the RWI-based tree radial growth response to autumn climate change, which has received less research interest compared to the spring and summer seasons.
Conclusion
We investigated the RWI for forest ecosystems in a vast biome/region of the Northern Hemisphere and observed spatial variations in the tree radial growth response to autumn climate change with respect to geographical, climatological, and biological gradients. Our results showed that warmer autumn temperatures tended to negatively affect tree radial growth south of 40° N in regions such as western Asia, southern Europe, United State of America and Mexico, which was attributed to temperature-induced water stress in line with the findings of previous studies that focused on the summer season. Furthermore, it was clear that geographical and climatological characteristics had a more significant impact on the RWI response compared to biological characteristics. Further efforts to fill spatial gaps in the RWI data, for example, in central and western Canada and Siberia, are required to better understand the tree radial growth response to climate change, resulting in more accurate future projections of forest dynamics and global climate change.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author Contributions
ST, AK, and NS designed the study. ST and AS collected and analyzed the data. ST, AK, AS, and NS wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was supported partly by the Grant for Joint Research Program of the Japan Arctic Research Network Center and KAKENHI (Grant Nos. 20K19950).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
References
Andreu-Hayles, L., Planells, O., Gutierrez, E., Muntan, E., Helle, G., Anchukaitis, K. J., et al. (2011). Long tree-ring chronologies reveal 20th century increases in water-use efficiency but no enhancement of tree growth at five Iberian pine forests. Glob. Change Biol. 17, 2095–2112. doi: 10.1111/j.1365-2486.2010.02373.x
Barber, V. A., Juday, G. P., and Finney, B. P. (2000). Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 405, 668–673. doi: 10.1038/35015049
Bonan, G. B. (2008). Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science 320, 1444–1449. doi: 10.1126/science.1155121
Bousquet, P., Peylin, P., Ciais, P., Le Quéré, C., Friedlingstein, P., and Tans, P. P. (2000). Regional changes in carbon dioxide fluxes of land and oceans since 1980. Science 290, 1342–1346. doi: 10.1126/science.290.5495.1342
Buermann, W., Parida, B., Jung, M., MacDonald, G. M., Tucker, C. J., and Reichstein, M. (2014). Recent shift in Eurasian boreal forest greening response may be associated with warmer and drier summers. Geophys. Res. Lett. 41, 1995–2002. doi: 10.1002/2014GL059450
Cai, J., Xu, K., Zhu, Y., Hu, F., and Li, L. (2020). Prediction and analysis of net ecosystem carbon exchange based on gradient boosting regression and random forest. Appl. Energy 262:114566. doi: 10.1016/j.apenergy.2020.114566
Chapin, F., Schultz, E., and Mooney, H. (1990). The ecology and economics of storage in plants. Ann. Rev. Ecol. Syst. 21, 423–447. doi: 10.1146/annurev.es.21.110190.002231
Ciais, P., Canadell, J. G., Luyssaert, S., Chevallier, F., Shvidenko, A., Poussi, Z., et al. (2010). Can we reconcile atmospheric estimates of the Northern terrestrial carbon sink with land-based accounting? Curr. Opin. Env. Sust. 2, 225–230. doi: 10.1016/j.cosust.2010.06.008
Cook, E. R. (1985). A Time Series Analysis Approach To Tree-Ring Standardization. Ph. D. Dissertation. Tucson: The University of Arizona Press.
Cook, E. R., and Kairiukstis, A. (1990). Methods of Dendrochronology: Applications in the Environmental Sciences. Dordrecht: Kluwer Academic Press.
D’Arrigo, R. D., Kaufmann, R. K., Davi, N., Jacoby, G. C., Laskowski, C., Myneni, R. B., et al. (2004). Thresholds for warming-induced growth decline at elevational tree line in the Yukon Territory. Canada. Global Biogeochemical Cycles 18, 1–7.
Devi, N., Hagedorn, F., Moiseev, P., Bugmann, H., Shiyatov, S., Mazepa, V., et al. (2008). Expanding forests and changing growth forms of Siberian larch at the Polar Urals tree line during the 20th century. Glob. Change Biol. 14, 1581–1591. doi: 10.1111/j.1365-2486.2008.01583.x
Dulamsuren, C., Hauck, M., Khishigjargal, M., Leuschner, H. H., and Leuschner, C. (2010). Diverging climate trends in Mongolian taiga forests influence growth and regeneration of Larix siberica. Oecologia 163, 1091–1102. doi: 10.1007/s00442-010-1689-y
Fan, S., Gloor, M., Mahlman, J., Pacala, S., Sarmiento, J., Takahashi, T., et al. (1998). A large terrestrial carbon sink in north America implied by atmospheric and oceanic carbon dioxide data and models. Science 282, 442–446. doi: 10.1126/science.282.5388.442
Fu, P. L., Grießinger, J., Gebrekirstos, A., Fan, Z. X., and Bräuning, A. (2017). Earlywood and latewood stable carbon and oxygen isotope variations in two pine species in southwestern china during the recent decades. Front. Plant Sci. 7:2050. doi: 10.3389/fpls.2016.02050
Genet, H., Bre´da, N., and Dufre^ne, E. (2010). Age-related variation in carbon allocation at tree and stand scales in beech (Fagus sylvatica L.) and sessile oak (Quercus petraea (Matt.) Liebl.) using a chronosequence approach. Tree Physiol. 30, 177–192. doi: 10.1093/treephys/tpp105
Goodale, C. L., Apps, M. J., Birdsey, R. A., Field, C. B., Heath, L. S., Houghton, R. A., et al. (2002). Forest carbon sinks in the Northern Hemisphere. Ecol. Appl. 12, 891–899. doi: 10.1890/1051-0761(2002)012[0891:fcsitn]2.0.co;2
Harris, I., Jones, P. D., Osborn, T. J., and Lister, D. H. (2014). Updated high-resolution grids ofmonthly climatic observations – the CRU TS3.10 dataset. Int. J. Climatol. 34, 623–642. doi: 10.1002/joc.3711
Jeong, S. J., Ho, C. H., Gim, H. J., and Brown, M. (2011). Phenology shifts at start vs. end of growing season in temperate vegetation over Northern Hemisphere for the period 1982-2008. Glob. Change Biol. 17, 2385–2399. doi: 10.1111/j.1365-2486.2011.02397.x
Kagawa, A., Naito, D., Sugimoto, A., and Maximov, T. C. (2003). Effects of spatial and temporal variability in soil moisture on widths and δ13C values of eastern Siberiantree rings. J. Geophys. Res. 108:4500. doi: 10.1029/2002JD003019
Kagawa, A., Sugimoto, A., and Maximov, T. C. (2006). Seasonal course of translocation, storage and remobilization of C-13 pulse-labeled photoassimilate in naturally growing Larix gmelinii saplings. New Phytol. 171, 793–804. doi: 10.1111/j.1469-8137.2006.01780.x
Kirdyanov, A., Hughes, M. K., Vaganov, E., Schweingruber, F., and Silkin, P. (2003). The importance of early summer temperatures and date of snow melt for tree growth in Siberian subarctic. Trees 17, 61–69. doi: 10.1007/s00468-002-0209-z
Liu, D., Piao, S., Wang, T., Wang, X., Wang, X., Ding, J., et al. (2018). Decelerating autumn CO2 release with warming induced by attenuated temperature dependence of respiration in northern ecosystems. Geophys. Res. Lett. 45, 5562–5571. doi: 10.1029/2018gl077447
Lloyd, A. H., and Bunn, A. G. (2007). Responses of the circumpolar boreal forest to 20th century climate variability. Environ. Res. Lett. 2, 45013–45013.
McDowell, N. G., Pockman, W. T., Allen, C. D., Breshears, D. D., Cobb, N., Kolb, T., et al. (2008). Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol. 178, 719–739. doi: 10.1111/j.1469-8137.2008.02436.x
Morozumi, T., Sugimoto, A., Suzuki, R., Nagai, S., Kobayashi, H., Tei, S., et al. (2020). Photographic records of plant phenology and spring river flush timing in a river lowland ecosystem at the taiga-tundra boundary, north-eastern Siberia. Ecol. Res. 35, 717–723, doi: 10.1111/1440-1703.12107
Nagai, S., Kotani, A., Morozumi, T., Kononov, A. V., Petrov, R. E., Shakhmatov, R., et al. (2020). Detection of long-term year-to-year spring and autumn bio-meteorological variations in Siberian ecosystems. Polar Sci. 25:100534. doi: 10.1016/j.polar.2020.100534
Nikolaev, A. N., Fedorov, P. P., and Desyatkin, A. R. (2009). Influence of climate and soilhydrothermal regime on radial growth of Larix cajanderi and Pinus sylvestris in Central Yakutia. Russia Scand. J. For. Res. 24, 217–226. doi: 10.1080/02827580902971181
Pan, Y., Birdsey, R. A., Fang, J., Houghton, R., Kauppi, P., Kurz, W. A., et al. (2011). A large and persistent carbon sink in the world’s forests. Science 333, 988–993. doi: 10.1126/science.1201609
Piao, S., Ciais, P., Friedlingstein, P., Peylin, P., Reichstein, M., Luyssaert, S., et al. (2008). Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 451, 49–52. doi: 10.1038/nature06444
R Core Team (2017). R: A Language And Environment For Statistical Computing. Vienna: R Foundation for Statistical Computing.
Serreze, M. C., and Barry, R. G. (2011). Processes and impacts of Arctic amplification: a research synthesis. Glob. Planet. Change 77, 85–96. doi: 10.1016/j.gloplacha.2011.03.004
Shiyatov, S. G., Terent’ev, M. M., and Fomin, V. V. (2005). Spatiotemporal dynamics of forest-tundra communities in the Polar Urals. Russ. J. Ecol. 36, 69–75. doi: 10.1007/s11184-005-0051-9
Sidorova, O. V., Siegwolf, R. T. W., Saurer, M., Shashkin, A. V., Knorre, A. A., Prokushkin, A. S., et al. (2009). Do centennial tree-ring and stable isotope trends of Larix gmelinii (Rupr.) Rupr. Indicate increasing water shortage in the Siberian north? Oecologia 161, 825–835. doi: 10.1007/s00442-009-1411-0
Silva, L. C. R., Anand, M., Oliveira, J. M., and Pillar, V. D. (2009). Past century changes in Araucaria angustifolia (Bertol.) Kuntze water use efficiency and growth in forest and grassland ecosystems of southern Brazil: implications for forest expansion. Glob. Change Biol. 15, 2387–2396. doi: 10.1111/j.1365-2486.2009.01859.x
Sugimoto, A., Naito, D., Yanagisawa, N., Ichiyanagi, K., Kurita, N., Kubota, J., et al. (2003). Characteristics of soil moisture in permafrost observed in East Siberian taiga with stable isotopes of water. Hydrol. Processes 17, 1073–1092. doi: 10.1002/hyp.1180
Tei, S. and Sugimoto, A. (2018). Time lag and negative responses of forest greenness and tree growth to warming over circumboreal forests. Glob. Chang. Biol. 24, 4225–4237. doi: 10.1111/gcb.14135
Tei, S., and Sugimoto, A. (2020). Excessive positive response of model-simulated land net primary production to climate changes over circumboreal forests. Plant Environ. Interact. 1, 102–121. doi: 10.1002/pei3.10025
Tei, S., Morozumi, T., Nagai, S., Takano, S., Sugimoto, A., Shingubara, R., et al. (2019a). An extreme flood caused by a heavy snowfall over the Indigirka River basin in Northeastern Siberia. Hydrol. Process 34, 522–537. doi: 10.1002/hyp.13601
Tei, S., Nagai, S., and Sugimoto, A. (2019b). Effects of climate dataset type on tree-ring analysis: a case study for Siberian forests. Polar Sci. 21, 136–145. doi: 10.1016/j.polar.2018.10.008
Tei, S., Sugimoto, A., Kotani, A., Ohta, T., Morozumi, T., Saito, S., et al. (2019c). Storing and stable relationships between tree-ring parameters and forest -level carbon fluxes in a Siberian larch forest. Polar Sci. 21, 146–157. doi: 10.1016/j.polar.2019.02.001
Tei, S., Sugimoto, A., Liang, M., Yonenobu, H., Matsuura, Y., Oosawa, A., et al. (2017a). Radial growth and physiological response of coniferous trees to Arctic amplification. J. Geophys. Res. 122, 2786–2803. doi: 10.1002/2016JG003745
Tei, S., Sugimoto, A., Yonenobu, H., Hoshino, Y., and Maximov, T. C. (2013a). Reconstruction of summer palmer drought severity index from δ13C of larch tree rings in East Siberia. Quat. Int. 290-291, 275–281. doi: 10.1016/j.quaint.2012.06.040
Tei, S., Sugimoto, A., Yonenobu, H., Kotani, A., and Maximov, T. C. (2019d). Effects of extreme drought and wet events for tree mortality: Insights from tree-ring width and carbon isotope ratio in a Siberian larch forest. Ecohydrology 12:e2143.
Tei, S., Sugimoto, A., Yonenobu, H., Matsuura, Y., Osawa, A., Sato, H., et al. (2017b). Tree-ring analysis and modeling approaches yield contrary response of circumboreal forest productivity to climate change. Glob. Change Biol. 23, 5179–5188. doi: 10.1111/gcb.13780
Tei, S., Sugimoto, A., Yonenobu, H., Ohta, T., and Maximov, T. C. (2014). Growth and physiological responses of larch trees to climate changes deduced from tree ring widths and d13C at two forest sites in eastern Siberia. Polar Sci. 8, 183–195. doi: 10.1016/j.polar.2013.12.002
Tei, S., Sugimoto, A., Yonenobu, H., Yamazaki, T., and Maximov, T. C. (2013b). Reconstruction of soil moisture for the past 100 years in eastern Siberia by using δ13C of larch tree rings. J. Geophys. Res. 118, 1256–1265. doi: 10.1002/jgrg.20110
Tei, S., Yonenobu, H., Sugimoto, A., Ohta, T., and Maximov, T. C. (2015). Reconstructed summer palmer drought severity index since 1850 AD based on δ13C of larch tree rings in eastern Siberia. J. Hydrol. 529, 442–448. doi: 10.1016/j.jhydrol.2015.01.085
Walker, X., Michelle, C. M., and Johnstone, J. F. (2015). Stable carbon isotope analysis reveals widespread drought stress in boreal black spruce forests. Glob. Change Biol. 21, 3102–3113. doi: 10.1111/gcb.12893
Wang, X., Piao, S., Ciais, P., Li, J., Friedlingstein, P., Koven, C., et al. (2011). Spring temperature change and its implication in the change of vegetation growth in North America from 1982 to 2006. Proc. Natl. Acad. Sci. U.S.A. 108, 1240–1245. doi: 10.1073/pnas.1014425108
Wigley, T. M. L., Briffa, K. R., and Jones, P. D. (1984). On the average value of correlated time-series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 23, 201–213. doi: 10.1175/1520-0450(1984)023<0201:otavoc>2.0.co;2
Wiley, E., and Helliker, B. (2012). A re-evaluation of carbon storage in trees lends greater support for carbon limitation to growth. New Phytol. 195, 285–289.
Wu, D. H., Zhao, X., and Liang, S. L. (2015). Time-lag effects of global vegetation responses to climate change. Glob. Change Biol. 21, 3520–3531.
Wu, X., Li, X., Liu, H., Ciais, P., Li, Y., Xu, C., et al. (2018). Uneven winter snow influence on tree growth across temperate China. Glob. Chang Biol. 25, 144–154. doi: 10.1111/gcb.14464
Xu, K., Wang, X., Liang, P., An, H., Sun, H., Han, W., et al. (2017). Tree-ring widths are good proxies of annual variation in forest productivity in temperature forests. Sci. Rep. 7:1945. doi: 10.1038/s41598-017-02022-6
Keywords: tree-ring width index, forest ecosystem, Northern Hemisphere, random forest algorithm, climate change, temperature-induced water stress, machine learning, response function
Citation: Tei S, Kotani A, Sugimoto A and Shin N (2021) Geographical, Climatological, and Biological Characteristics of Tree Radial Growth Response to Autumn Climate Change. Front. For. Glob. Change 4:687749. doi: 10.3389/ffgc.2021.687749
Received: 30 March 2021; Accepted: 03 August 2021;
Published: 26 August 2021.
Edited by:
Sigrid Netherer, University of Natural Resources and Life Sciences Vienna, AustriaCopyright © 2021 Tei, Kotani, Sugimoto and Shin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shunsuke Tei, ZWluNDU3OTA3MDlAYWZmcmMuZ28uanA=