 Jason P. Field
Jason P. Field David D. Breshears
David D. Breshears John B. Bradford
John B. Bradford Darin J. Law
Darin J. Law Xiao Feng
Xiao Feng Craig D. Allen
Craig D. Allen- 1School of Natural Resources and the Environment, University of Arizona, Tucson, AZ, United States
- 2Department of Ecology and Evolutionary Biology, University of Arizona, Tucson, AZ, United States
- 3U.S. Geological Survey, Southwest Biological Science Center, Flagstaff, AZ, United States
- 4Department of Geography, Florida State University, Tallahassee, FL, United States
- 5U.S. Geological Survey, Fort Collins Science Center, New Mexico Landscapes Field Station, Los Alamos, NM, United States
Drought and warming increasingly are causing widespread tree die-offs and extreme wildfires. Forest managers are struggling to improve anticipatory forest management practices given more frequent, extensive, and severe wildfire and tree die-off events triggered by “hotter drought”—drought under warmer than historical conditions. Of even greater concern is the increasing probability of multi-year droughts, or “megadroughts”—persistent droughts that span years to decades, and that under a still-warming climate, will also be hotter than historical norms. Megadroughts under warmer temperatures are disconcerting because of their potential to trigger more severe forest die-off, fire cycles, pathogens, and insect outbreaks. In this Perspective, we identify potential anticipatory and/or concurrent options for non-timber forest management actions under megadrought, which by necessity are focused more at finer spatial scales such as the stand level using higher-intensity management. These management actions build on silvicultural practices focused on growth and yield (but not harvest). Current management options that can be focused at finer scales include key silvicultural practices: selective thinning; use of carefully selected forward-thinking seed mixes; site contouring; vegetation and pest management; soil erosion control; and fire management. For the extreme challenges posed by megadroughts, management will necessarily focus even more on finer-scale, higher-intensity actions for priority locations such as fostering stand refugia; assisted stand recovery via soil amendments; enhanced root development; deep soil water retention; and shallow water impoundments. Drought-induced forest die-off from megadrought likely will lead to fundamental changes in the structure, function, and composition of forest stands and the ecosystem services they provide.
Climate change and forests of the future: Managing in the face of uncertainty
“Adaptive strategies include resistance options (forestall impacts and protect highly valued resources), resilience options (improve the capacity of ecosystems to return to desired conditions after disturbance), and response options (facilitate transition of ecosystems from current to new conditions). … Priority-setting approaches (e.g., triage), appropriate for rapidly changing conditions and for situations where needs are greater than the available capacity to respond, will become increasingly important in the future” (Millar et al., 2007).
Introduction
Widespread tree die-offs and extreme wildfires are increasingly being triggered by drought and warming across the globe (IPCCa, 2014; Hartmann et al., 2018; Brando et al., 2019a). Drought-induced and heat-related tree mortality—including “forest die-off” where a large proportion of trees die—has been progressively documented on all forested continents (Allen et al., 2010, 2015; IPCCa, 2014; Hartmann et al., 2018). These changes are related to “hotter droughts”—droughts that are warmer than prior historical droughts due to increased greenhouse gas emissions (Breshears et al., 2005; Allen et al., 2015). A pronounced hastening in tree mortality has experimentally been documented in response to extreme hotter drought (Adams et al., 2009, 2017; Will et al., 2013; Duan et al., 2014, 2015), indicating that tree mortality will be an increasing challenge as warming continues. Furthermore, tree mortality may be additionally exacerbated if drought increases in frequency, duration, and severity as warming continues as projected (Bradford and Bell, 2017; Schwalm et al., 2017; McDowell et al., 2018). Wildfire activity also is being exacerbated by warming and drought, especially in the Western U.S. (Westerling et al., 2006; Williams et al., 2013)—increasing in frequency, severity, and extent in many areas. Indeed, recent increases in extreme wildfire and widespread tree die-off track warming trends and are projected to increase dramatically with projected increases in aridity and temperature (Williams et al., 2013; Brando et al., 2019b). Managing forests that are experiencing extensive tree mortality from hotter drought is challenging, as these systems can undergo potentially irreversible state changes, including ecological cascades and even ecosystem collapse (Allen et al., 2007; Cobb et al., 2017; Ruthrof et al., 2018). For example, ecological cascades triggered by extreme hotter drought and multi-year to multi-decadal megadrought in the Western U.S. could progress through alternate states from climate-disturbed forest, to woodland, shrubland, invasive grassland, and even extremely degraded ecosystems (Allen et al., 2007; Romme et al., 2009; Cobb et al., 2017). The potential for forest stands to shift to alternative states depends heavily on recovery trajectories, which can be altered and improved through including the implementation of non-traditional, finer-scale, more intensive stand-level management practices (Bradford and Bell, 2017; Cobb et al., 2017).
A key traditional tool for trying to address the challenge of hotter drought is selective thinning to maintain forests at lower density to minimize competition and enhance resistance and resilience to short-term drought (D'Amato et al., 2013; Bottero et al., 2017; Gleason et al., 2017). Additional non-traditional forest management practices are being modified to address hotter drought and include intensive, novel management practices to minimize or manage tree die-offs and wildfire events (Millar et al., 2007; Allen et al., 2015; Millar and Stephenson, 2015; Cobb et al., 2017; Stephens et al., 2018).
Increasingly being considered is the option to let forests progress through change and try to somewhat modify that change rather than prevent it (Millar and Stephenson, 2015). This might be particularly relevant for regional-scale megadroughts—multi-year to multi-decadal droughts with levels of aridity that are as dry as the extreme droughts of the twentieth century—which have been documented in long-term tree-ring data and precipitation records throughout many regions worldwide (Ault et al., 2016; Ault and St. George, 2018). Paleoclimate records suggest that megadroughts can lead to rapid, regional-scale ecosystem degradation and collapse (e.g., Godfree et al., 2019). Additionally, megadroughts will likely promote more frequent and extreme wildfires, which can accelerate the transformational processes to non-forest systems (Stevens-Rumann et al., 2018). However, the ecological consequences, including taxonomic breadth, trophic depth, and geographic pattern of these impacts remain largely unknown (Godfree et al., 2019). Recent studies suggest that state-of-the-art climate model simulations tend to underestimate the natural occurrence rate of extreme events, especially since these models cannot represent decadal-to-centennial climate variations comparable in magnitude to paleoclimate reconstructions (Ault et al., 2016; Ault and St. George, 2018). There is growing evidence to suggest that an increase in global temperature of 1.5–3°C above pre-industrial levels will likely increase the magnitude and frequency of extreme drought, as well as the occurrence of megadrought across most global land areas, including more mesic systems (Prudhomme et al., 2014; Cook et al., 2015, 2016, 2019; Naumann et al., 2018).
The occurrence of megadrought at broad sub-continental to continental scales can have devastating socioeconomic and environmental impacts, as has been documented to occur worldwide within the past century (Stahle et al., 2007; Evans et al., 2018; Naumann et al., 2018; Godfree et al., 2019). The ecological consequences of hotter megadroughts may be particularly detrimental and largely irreversible on forested ecosystems—which, in some cases may be more vulnerable to drought than other systems such as grasslands and deserts due to physiological differences among dominant plant functional types and lifeforms (e.g., grasses or annual herbs vs. trees; angiosperms vs. gymnosperms; Slatyer, 1967; Tyree and Zimmermann, 2002; McDowell et al., 2008; Breshears et al., 2016; Eller et al., 2016; O'Sullivan et al., 2017; Choat et al., 2018).
Because megadroughts are spatially extensive, temporally persistent and climatically extreme, management strategies for preserving forests at landscape scales likely will be impractical and infeasible, necessitating management actions that might nudge forest transition to an alternate state of one type that is preferred over another, and/or manage the system post-megadrought (Millar and Stephenson, 2015). However, preservation of existing forest might be feasible at finer scales of stands using higher intensity and potentially novel management.
In this Perspective, we focus on forest management options under megadrought, which are largely constrained to finer-scale, higher-intensity practices. We (1) introduce a scenario framework to discuss potential ecological trajectories and management options; (2) briefly recap traditional forest management options and emerging concepts for dealing with hotter drought events; (3) highlight finer-scale, higher-intensity stand management options that might be used in an attempt to buffer forests from megadrought (extending initial general concepts noted by Millar and Stephenson, 2015); and (4) propose that novel challenges associated with mitigating megadrought impacts need to be considered, evaluated, and researched more extensively to enhance our forest treatment toolbox for more effective forest management under emerging global change stresses.
A Scenario Framework: Contrasting Management for Hotter Drought and Megadrought From Base Conditions
We propose a scenario framework (Figure 1)—based on a case study using semiarid western U.S. forests that are changing rapidly and for which we have a rich knowledge base (e.g., Williams et al., 2013; Allen et al., 2015; Cobb et al., 2017; Breshears et al., 2018)—that considers forest trajectories under warming climate (solid lines) or with megadrought (dashed lines). The trajectories shown in Figure 1 are theoretical examples highlighting possible outcomes based on expert opinion and existing data compiled from available studies and field observations in the semiarid western U.S. The impacts of warming climate alone in these ecosystems are expected to be so great that our strategy for forest management is more focused on first maintaining small refugia and then managing the products of succession in highly disturbed landscapes to the best of our abilities. That is, we expect the challenges ahead, at least in western U.S. forests and probably others as well, are sufficiently great that effective landscape-scale management to maintain current forest cover will become increasingly challenging and that a shift to prioritizing stand-scale high priority locations over landscape-scale preservation will become critical. Ecosystem conditions can cascade through levels of stress and change (Figure 1 left column), from drought stress and hotter drought through loss of biodiversity, soil, and ecosystem services. At the scale of stand refugia, under a warming climate alone, we expect major impacts to forests including ecological cascades, especially under no management (red solid line). When compounded by megadrought, under no management, we expect these cascades to be more rapid and remain in the most degraded state (red dashed line). Using broader-scale, less intensive management tools such as those currently applied, these cascades may be able to be partially arrested in absence of megadrought (black solid lines), but less so with the occurrence of megadrought (black dashed lines). Finally, shifting focus to finer-scale, more intense approaches may result in extensive landscape degradation but will enable stand refugia to persist under warming climate (blue solid line) and perhaps even under megadrought (blue dashed line). Next we carry this framework forward to discuss existing broad-scale options to manage forests under a warming climate with hotter drought and their potential efficacy under megadrought, and then address finer-scale more intensive options that may help protect or create refugia under megadrought.
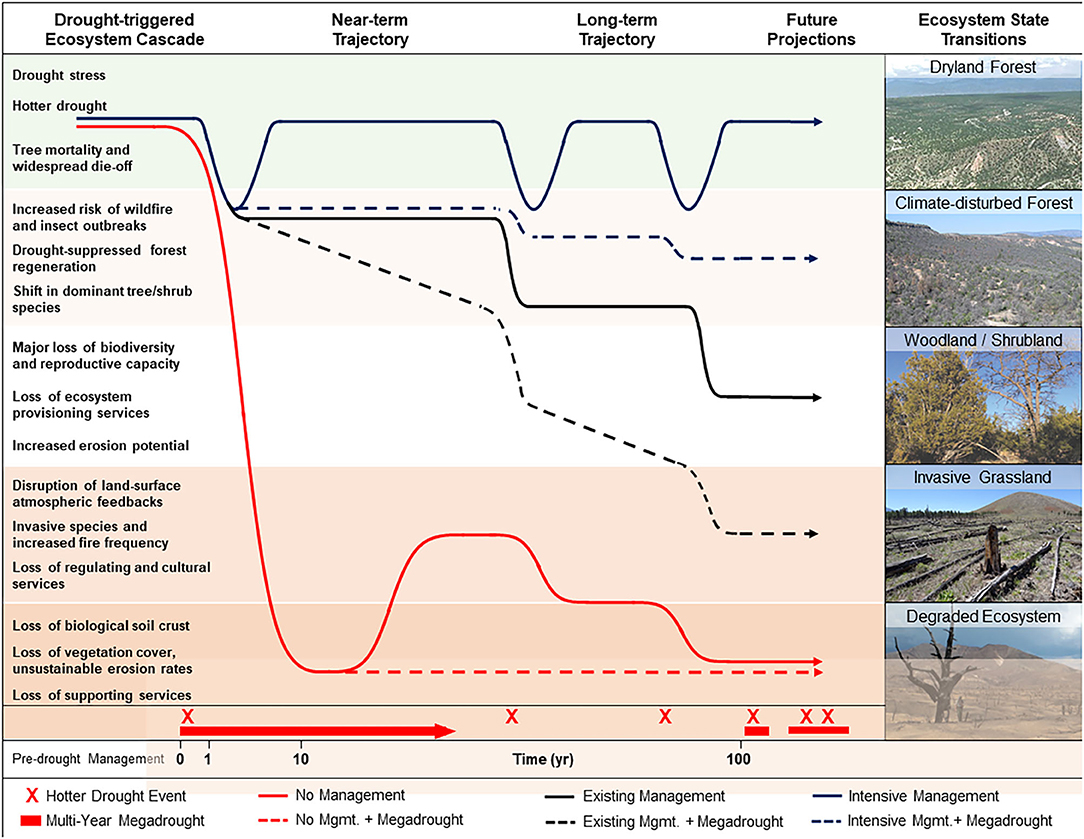
Figure 1. Drought-triggered ecological cascade and management strategies at finer spatial scales to reduce the risk of forest die-off and ecosystem degradation, trajectories are theoretical examples highlighting possible outcomes for dry forests in the Western U.S. based on example references including but not limited to Allen et al. (2010, 2015), Breshears et al. (2011, 2018), Grant et al. (2013), Field et al. (2015), Bradford and Bell (2017), Cobb et al. (2017), and Petrie et al. (2017)—showing trajectories and future projections for unmanaged systems (red), systems managed with existing forest practices (black), intensively managed systems (blue)—for both recurring extreme drought (solid lines) and multidecadal megadrought (dashed lines); more mesic forests would experience different trajectories, particularly including species turnover toward more drought tolerant tree species while still remaining forest (e.g., Brando et al., 2019b). Photo credits: C. D. Allen, J. B. Bradford.
Hotter Droughts: Existing Forest Management Options
Forest managers have been considering how to deal with the effects of hotter drought for over a decade (Millar et al., 2007). A first step in anticipatory management for hotter drought is to accurately identify in advance the forested ecosystems most at risk to drought (Millar and Stephenson, 2015). Identification of vulnerable forest stands can be aided by Earth System Models (Swann et al., 2018); forest health monitoring networks (Hartmann et al., 2018); near-term ecological forecasting of extreme events (Dietze et al., 2018; Redmond et al., 2019); remote sensing (Mu et al., 2013), traits related to drought-induced mortality (Anderegg et al., 2016; Sperry et al., 2016; O'Brien et al., 2017); and ground surveys (Breshears et al., 2005; Redmond et al., 2019).
A second step in managing for hotter drought is to draw on, and modify as feasible, traditional anticipatory forest management practices (Bradford et al., 2018) that would be most effective prior to the onset of a hotter drought event (Table 1). These options are not limited to, but include traditional stand-level practices such as: (1) dispersal of carefully selected seed mixes to reduce erosion and risk of unwanted species like cheatgrass (Bromus tectorum) and buffelgrass (Pennisetum ciliare) and novel techniques such as seed pillows or seed pellets to improve recruitment (Gornish et al., 2015; Davies, 2018; Madsen et al., 2018); (2) selective thinning of stands to reduce competition for water during drought events and minimize drought-drive growth declines that lead to mortality (Bréda et al., 1995; McDowell et al., 2006; Millar and Stephenson, 2015; Andrews et al., 2020); (3) contouring to slow overland flow, increase infiltration, and enhance soil water availability (Panagos et al., 2015); (4) vegetation and pest management—to control pests using pesticides or pheromones and to manipulate vegetation characteristics and demography, including understory vegetation, such as size, age, distribution, and noxious, non-native vs. native species (Millar and Stephenson, 2015); (5) mulching residual thinning debris to enhance soil water storage and reduce erosion (Grant et al., 2013; Xia et al., 2019); and (6) fire management for reducing fuel loads and risk of crown fire (Moreira et al., 2011).

Table 1. Possible management options and objectives for increasing forest resilience to hotter drought and multidecadal megadrought.
Some of the above proposed management practices could also have positive feedbacks on other climate-mediated responses such as making trees less susceptible to insects, pathogens, and wildfire (Allen et al., 2015; Cobb et al., 2017). For example, selective thinning to reduce water stress within a forest stand can increase the amount of soil water available per tree for remaining trees (Zou et al., 2008; Andrews et al., 2020). Although these existing forest management strategies enhance forest resistance and resilience to drought (Table 1), these practices have limits, especially as temperatures rise, and are unlikely to buffer forests against the impacts of multi-year to multi-decadal hotter drought conditions (Figure 1). In particular, maintaining dry forests at low densities may help reduce drought-induced mortality through the middle of the twenty-first century, but this strategy likely will not be sufficient to avoid dramatically higher tree mortality rates in later decades (Bradford and Bell, 2017).
Megadroughts: Focused Actions to Protect Stand Refugia
Although a combination of anticipatory traditional forest practices, in conjunction with emerging management practices for hotter drought, can prove substantial buffers against a hotter drought event, they are unlikely to be sufficient to prevent extreme wildfire and forest die-off at broad scales during megadrought. Traditional practices focused on forest regeneration are less likely to be effective during a megadrought. Forest regeneration is episodic, especially in semiarid forests such as those of the western U.S, and is driven by relatively rare combinations of viable seed, soil moisture, and temperature conditions (Kolb and Robberecht, 1996; Brown and Wu, 2005; Coop and Givnish, 2008; Feddema et al., 2013; Savage et al., 2013; Petrie et al., 2017). Management practices for ensuring forest stand recovery from megadrought could include rapid response to establish trees before noxious unwanted species take hold, targeting seeding or planting during periods of favorable moisture conditions to maximize success (Bradford et al., 2018), and utilizing seed pellet technology (Gornish et al., 2015), tree species, and/or genotypes that will be suitable for future conditions.
Megadrought, then, by necessity may require prioritizing selected stands for finer-scale, high-intensity management (Figure 1), similar in scale to restoration islands (Hulvey et al., 2017). Prioritization of such stands for high-intensity management will likely need to be made on a case-by-case basis, but criteria that could aid in such prioritization include: (i) protecting existing refugia (McDowell et al., 2019); (ii) protecting large old trees due to their disproportionate influence on ecosystems and ecosystem services (Boylan, 2010; Úradníček et al., 2017; Enquist et al., 2020), (iii) protecting culturally important trees and groves (Daniel et al., 2016), including iconic trees and groves in protected areas such as national parks (Grant et al., 2013); (iv) protecting groves that provide key ecosystem services, especially for those practicing subsistence living (Sutherland et al., 2016); (v) protecting genetically distinct and perhaps more climatically resistant populations (Polle et al., 2019), or endangered evolutionarily important species (e.g., Wollemi pine [Wollemia nobilis] in Australia; Woodford, 2012); (vi) protecting groves that form a key part of corridors for fauna and flora (Rosot et al., 2018); and (vii) protecting isolated patches that might be important in reducing seed dispersal distances for revegetation post-disturbance (Shive et al., 2018).
Intensive management options that could be deployed in areas of limited spatial extent for protecting stand refugia during multi-year hotter drought could include options such as soil amendments and agriculture-based technologies (Table 1). Soil amendments, particularly recent advances in nanotechnology-based amendments such as nanochitosan, may provide an additional set of useful tools for forest managers to utilize to help reduce the risk of tree mortality and forest die-back during extreme drought. These practices could also be useful during non-drought periods as well. A growing number of studies, primarily in the agricultural literature, have demonstrated the effectiveness of nanochitosan soil applications for significantly improving the physiological properties of plants including: drought tolerance, increased growth and yield, leaf area, resin production, antifungal and antiviral activity, nutrient use efficiency, seedling establishment, resistance to diseases and pathogens, and production of secondary metabolites to help overcome stressors such as windshear, temperature extremes, osmotic pressure, and attacks by phytophagous (e.g., Sharp, 2013; Katiyar et al., 2015; Behboudi et al., 2018; Kumaraswamy et al., 2018; Li et al., 2019; Table 1). Given the challenges expected with hotter drought and megadrought, a soil amendment such as mulch, fertilizer, organic matter, and possibly even nanochitosan, all have potential as a treatment that might enable longer persistence of trees during hotter drought conditions. Nanochitosan is commercially available and can be applied directly to the plant foliage or to the soil, either at the base of individual trees or as broadcast application for forest stands, with an approximate application cost per hectare comparable to the cost of standard fertilizer applications (Li et al., 2019). Other agriculture-based technologies could provide a set of useful tools for reforestation and afforestation practices and for managing forest stands, especially when drought occurs during recruitment and planting phases. In addition to planting phases, new agriculture-based technologies could significantly improve drought tolerance, as well as many other physiological characteristics, in tree seedlings and saplings grown for nursery stock. For example, the use of a newly developed plant growing structure for use inside nursery containers and for use when planting bare root trees in the ground can significantly enhance (i) root architecture and biomass; (ii) plant growth and yield; (iii) soil aeration, infiltration, and drainage; (iv) plant uptake of nutrients and water; (v) beneficial soil bacterial and fungal populations; and (vi) plant resistance to pathogens, disease, and environmental stressors (Table 1). Incorporating the use of agriculture-based technologies at fine spatial scales to protect stand refugia could lead to novel management practices for enhancing forest stand resistance and resilience during and against the time of multi-year megadroughts.
Additional options for fine-scale, more intensive management practices for protecting stand refugia in strategically-chosen locations during multi-year megadroughts could include (Table 1): (i) using deep soil water retention to maximize refugia, by actively managing surface characteristics to promote the concentration and infiltration of overland flow—which has been shown to be effective in drought and warming experiments where transplanted trees (Law et al., 2019) and established trees (McDowell et al., 2019) have apparently survived for more than a decade under hotter drought conditions due to root access to deep soil water (transplanted trees) and bedrock groundwater (established trees); (ii) shallow-water impoundments, which have been created extensively at landscape scales and are dominated by millions of water bodies smaller than 1 km2–especially low-tech farm impoundments, representing up to 6% of total agricultural land (Downing et al., 2006)—but has also been deployed successfully in forest stands to increase soil moisture and tree growth by up to 50% (e.g., Broadfoot, 1967), although these impoundments would need to be maintained frequently in highly erodible areas; and (iii) supplemental irrigation from other more distant sources, which could include utilizing subsurface drip irrigation and other agriculture-based irrigation practices (Grant et al., 2013). These non-traditional intensive management options are only feasible at fine scales and should be targeted where areas of natural stand refugia already exist. For example, while irrigating large tracts of forest is impractical, using site-specific strategically chosen stands of limited spatial extent where resources are readily accessible could be a feasible management option in extreme situations where the trees at risk are of highest importance. It may be that several specifically chosen stands could undergo site-specific irrigation that would knit together a network of forest preservation. Perhaps one of the most plausible scenarios with respect to supplemental irrigation is digging a few wells to access water deeper than tree roots can extend in a limited area of natural refugia to supply water to trees of high-value for ecosystem services such as aesthetic, cultural or wildlife habitat. Supplemental irrigation to protect stand refugia at fine scales will have site-specific limitations and will likely have important tradeoffs with other economic and ecological considerations such as demand for water during drought (e.g. municipal and agricultural use) and flora and fauna dependence on streamflow and depth to groundwater. Perhaps the greatest value of watering a forest may be to sustain forest stands themselves and all the ecosystem services, and habitats they provide (Grant et al., 2013).
Paths Forward
In conclusion, given the likelihood of extreme, hotter drought events occurring more often and over larger areas, in conjunction with increased likelihood of recurring megadroughts, we recommend that forest managers and policymakers take anticipatory actions to explicitly consider the growing forest vulnerabilities to these emerging climate-induced disturbances, as well as the associated needs for innovative and potentially intensive forest stand management to increase forest resistance and resilience to drought in high-priority stands (Figure 1). To minimize the forest loss and/or degradation that will be promoted by these challenges, forest managers may need to take preemptive action before the onset of a megadrought, building from existing management practices and incorporate new technology to develop the most economical and feasible long-term solutions for mitigating the risks of megadrought and drought-induced forest die-off. We note traditional forest management practices can be helpful to increase forest resistance and resilience and should not be discarded but used in conjunction with more intensive forest management options at finer spatial scales. Given the potential likelihood of more frequent, widespread forest die-off events, the most effective management strategies likely will be focused on guiding recovery and facilitating stand transitions following major die-off events (Millar et al., 2007; Cobb et al., 2017; Petrie et al., 2017; Bradford et al., 2018). Although drivers of forest recovery from drought and disturbances have received less attention than mortality or growth, these recovery and transition trajectories are likely to determine if forest stands impacted by megadroughts experience an important state change. Without stand-scale management intervention under emerging and anticipated hotter drought events and megadroughts, many forested ecosystems worldwide will be increasingly vulnerable to substantial, rapid ecological transformations—accompanied by loss of ecosystem services and biodiversity (e.g., Hessburg et al., 2019). The management strategies outlined in this Perspective—enhancing resilience at fine-scales to protect stand refugia, employing existing forest stand management practices as well as more intensive agriculture-based practices such as irrigation and soil amendments, and using facilitated recovery and transitions for prioritized stands—may help forest managers better deal with now-foreseeable challenges associated with increasingly extreme hotter drought events and megadroughts.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Author Contributions
JF provided an initial rough draft of the manuscript. All authors contributed to identifying management strategies for reducing forest die-off during hotter drought events and multi-year megadroughts, as well as substantial revision and editing of the text, table, and figure.
Funding
This work was supported primarily by NSF EASM-1243125. Additional support was provided by USGS SW Climate Adaptation Science Center Award G20AC00257, NSF EF-1340624, EF-1550756, EAR-1659546, DEB-1824796, USDA McIntire-Stennis ARZT-1390130-M12-222 and the University of Arizona Bridging Biodiversity and Conservation Science program supported by the Arizona Institutes for Resilience.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank James Walworth for discussion on agriculture-based technologies, Nora Caceres for discussion on nanotechnology applications, and Diana Liverman, Julia Cole, and Jonathan Overpeck for coordinating work on megadrought impacts that inspired this work. We thank Jayne Belnap, Julio Betancourt, Don Falk and Jeff Whicker for input on an earlier related version of Figure 1. Any use of trade, firm, or product name is for descriptive purposes only and does not imply endorsement by the U.S. Government.
References
Adams, H. D., Guardiola-Claramonte, M., Barron-Gafford, G. A., Villegas, J. C., Breshears, D. D., Zou, C. B., et al. (2009). Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proc. Natl. Acad. Sci. U.S.A. 106, 7063–7066. doi: 10.1073/pnas.0901438106
Adams, H. D., Zeppel, M. J., Anderegg, W. R., Hartmann, H., Landhäusser, S. M., Tissue, D. T., et al. (2017). A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 1:1285. doi: 10.1038/s41559-017-0248-x
Allen, C. D., Anderson, R. S., Jass, R. B., Toney, J. L., and Baisan, C. H. (2007). Paired charcoal and tree-ring records of high-frequency Holocene fire from two New Mexico bog sites. Int. J. Wildland Fire 17, 115–130. doi: 10.1071/WF07165
Allen, C. D., Breshears, D. D., and McDowell, N. G. (2015). On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 6, 1–55. doi: 10.1890/ES15-00203.1
Allen, C. D., Macalady, A. K., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecol. Manag. 259, 660–684. doi: 10.1016/j.foreco.2009.09.001
Anderegg, W. R., Klein, T., Bartlett, M., Sack, L., Pellegrini, A. F., Choat, B., et al. (2016). Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought-induced tree mortality across the globe. Proc. Natl. Acad. Sci. U.S.A. 113, 5024–5029. doi: 10.1073/pnas.1525678113
Andrews, C. A., D'Amato, A. W., Palik, B., Fraver, J. S., Battaglia, M., and Bradford, J. B. (2020). Stand density moderates growth declines during hot-droughts in arid forests. J. Appl. Ecol. 57, 1089–1102. doi: 10.1111/1365-2664.13615
Ault, T. R., Mankin, J. S., Cook, B. I., and Smerdon, J. E. (2016). Relative impacts of mitigation, temperature, and precipitation on 21st-century megadrought risk in the American Southwest. Sci. Adv. 2:e1600873. doi: 10.1126/sciadv.1600873
Ault, T. R., and St. George, S. (2018). Unraveling the mysteries of the megadrought. Phys. Today 71:44. doi: 10.1063/PT.3.3997
Behboudi, F., Tahmasebi Sarvestani, Z., Kassaee, M. Z., Sanavi, M., Mohamad, S. A., Sorooshzadeh, A., et al. (2018). Evaluation of chitosan nanoparticles effects on yield and yield components of barley (Hordeum vulgare L.) under late season drought stress. J. Water Environ. Nanotechnol. 3, 22–39. doi: 10.22090/JWENT.2018.01.003
Bottero, A., D'Amato, A. W., Palik, B. J., Bradford, J. B., Fraver, S., Battaglia, M. A., et al. (2017). Density-dependent vulnerability of forest ecosystems to drought. J. Appl. Ecol. 54, 1605–1614. doi: 10.1111/1365-2664.12847
Boylan, C. (2010). “Champion and heritage trees of Ireland,” in 22nd IFPRA World Congress (Hong Kong) 15–18.
Bradford, J. B., and Bell, D. M. (2017). A window of opportunity for climate-change adaptation: easing tree mortality by reducing forest basal area. Front. Ecol. Environ. 15, 11–17. doi: 10.1002/fee.1445
Bradford, J. B., Betancourt, J. L., Butterfield, B. J., Munson, S. M., and Wood, T. E. (2018). Anticipatory natural resource science and management for a changing future. Front. Ecol. Environ. 16, 295–303. doi: 10.1002/fee.1806
Brando, P. M., Paolucci, L., Ummenhofer, C. C., Ordway, E. M., Hartmann, H., Cattau, M. E., et al. (2019a). Droughts, wildfires, and forest carbon cycling: a pantropical synthesis. Annu. Rev. Earth Planet. Sci. 47, 555–581. doi: 10.1146/annurev-earth-082517-010235
Brando, P. M., Silvério, D., Maracahipes-Santos, L., Oliveira-Santos, C., Levick, S. R., Coe, M. T., et al. (2019b). Prolonged tropical forest degradation due to compounding disturbances: Implications for CO2 and H2O fluxes. Glob. Chang. Biol. 25, 2855–2868. doi: 10.1111/gcb.14659
Bréda, N., Granier, A., and Aussenac, G. (1995). Effects of thinning on soil and tree water relations, transpiration and growth in an oak forest (Quercus petraea (Matt.) Liebl.). Tree Physiol. 15, 295–306. doi: 10.1093/treephys/15.5.295
Breshears, D. D., Carroll, C. J., Redmond, M. D., Wion, A. P., Allen, C. D., Cobb, N. S., et al. (2018). A dirty dozen ways to die: metrics and modifiers of mortality driven by drought and warming for a tree species. Front. For. Global Change 1:4. doi: 10.3389/ffgc.2018.00004
Breshears, D. D., Cobb, N. S., Rich, P. M., Price, K. P., Allen, C. D., Balice, R. G., et al. (2005). Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. U.S.A. 102, 15144–15148. doi: 10.1073/pnas.0505734102
Breshears, D. D., Knapp, A. K., Law, D. J., Smith, M. D., Tidwell, D., and Wonka, C. L. (2016). Rangeland responses to predicted increases in drought extremity. Rangelands 38, 191–196. doi: 10.1016/j.rala.2016.06.009
Breshears, D. D., López-Hoffman, L., and Graumlich, L. J. (2011). When ecosystem services crash: preparing for big, fast, patchy climate change. Ambio 40, 256–263. doi: 10.1007/s13280-010-0106-4
Broadfoot, W. M. (1967). Shallow-water impoundment increases soil moisture and growth of hardwoods. Soil Sci. Soc. Am. J. 31, 562–564. doi: 10.2136/sssaj1967.03615995003100040036x
Brown, P. M., and Wu, R. (2005). Climate and disturbance forcing of episodic tree recruitment in a southwestern ponderosa pine landscape. Ecology 86, 3030–3038. doi: 10.1890/05-0034
Choat, B., Brodribb, T. J., Brodersen, C. R., Duursma, R. A., López, R., and Medlyn, B. E. (2018). Triggers of tree mortality under drought. Nature 558, 531–539. doi: 10.1038/s41586-018-0240-x
Cobb, R. C., Ruthrof, K. X., Breshears, D. D., Lloret, F., Aakala, T., Adams, H. D., et al. (2017). Ecosystem dynamics and management after forest die-off: a global synthesis with conceptual state-and-transition models. Ecosphere 8:e02034. doi: 10.1002/ecs2.2034
Cook, B. I., Cook, E. R., Smerdon, J. E., Seager, R., Williams, A. P., Coats, S., et al. (2016). North American megadroughts in the common era: reconstructions and simulations. Wiley Interdiscip. Rev. Clim. Change 7, 411–432. doi: 10.1002/wcc.394
Cook, B. I., Seager, R., Williams, A. P., Puma, M. J., McDermid, S., Kelley, M., et al. (2019). Climate change amplification of natural drought variability: the historic mid-twentieth-century north American drought in a warmer world. J. Clim. 32, 5417–5436. doi: 10.1175/JCLI-D-18-0832.1
Cook, E. R., Seager, R., Kushnir, Y., Briffa, K. R., Büntgen, U., Frank, D., et al. (2015). Old world megadroughts and pluvials during the common era. Sci. Adv. 1:e150056. doi: 10.1126/sciadv.1500561
Coop, J. D., and Givnish, T. J. (2008). Constraints on tree seedling establishment in montane grasslands of the Valles Caldera, New Mexico. Ecology 89, 1101–1111. doi: 10.1890/06-1333.1
D'Amato, A. W., Bradford, J. B., Fraver, S., and Palik, B. J. (2013). Effects of thinning on drought vulnerability and climate response in north temperate forest ecosystems. Ecol. Appl. 23, 1735–1742. doi: 10.1890/13-0677.1
Daniel, K. S., Udeagha, A. U., and Jacob, D. E. (2016). Socio-cultural importance of sacred forests conservation in south southern Nigeria. Afr. J. Sustain. Dev. 6, 251–268.
Davies, K. W. (2018). Incorporating seeds in activated carbon pellets limits herbicide effects to seeded bunchgrasses when controlling exotic annuals. Rangeland Ecol. Manag. 71, 323–326. doi: 10.1016/j.rama.2017.12.010
Dietze, M. C., Fox, A., Beck-Johnson, L. M., Betancourt, J. L., Hooten, M. B., Jarnevich, C. S., et al. (2018). Iterative near-term ecological forecasting: needs, opportunities, and challenges. Proc. Natl. Acad. Sci. U.S.A. 115, 1424–1432. doi: 10.1073/pnas.1710231115
Downing, J. A., Prairie, Y. T., Cole, J. J., Duarte, C. M., Tranvik, L. J., Striegl, R. G., et al. (2006). The global abundance and size distribution of lakes, ponds, and impoundments. Limnol. Oceanogr. 51, 2388–2397. doi: 10.4319/lo.2006.51.5.2388
Duan, H. L., Duursma, R. A., Huang, G. M., Smith, R. A., Choat, B., O'Grady, A. P., et al. (2014). Elevated CO2 does not ameliorate the negative effects of elevated temperature on drought-induced mortality in Eucalyptus radiata seedlings. Plant Cell Environ. 37, 1598–1613. doi: 10.1111/pce.12260
Duan, H. L., O'Grady, A. P., Duursma, R. A., Choat, B., Huang, G. M., Smith, R. A., et al. (2015). Drought responses of two gymnosperm species with contrasting stomatal regulation strategies under elevated CO2 and temperature. Tree Physiol. 35, 756–770. doi: 10.1093/treephys/tpv047
Eller, C. B., Lima, A. L., and Oliveira, R. S. (2016). Cloud forest trees with higher foliar water uptake capacity and anisohydric behavior are more vulnerable to drought and climate change. New Phytol. 211, 489–501. doi: 10.1111/nph.13952
Enquist, B. J., Abraham, A. J., Harfoot, M. B., Malhi, Y., and Doughty, C. E. (2020). The megabiota are disproportionately important for biosphere functioning. Nat. Commun. 11:699. doi: 10.1038/s41467-020-14369-y
Evans, N. P., Bauska, T. K., Gázquez-Sánchez, F., Brenner, M., Curtis, J. H., and Hodell, D. A. (2018). Quantification of drought during the collapse of the classic maya civilization. Science 361, 498–501. doi: 10.1126/science.aas9871
Feddema, J. J., Mast, J. N., and Savage, M. (2013). Modeling high-severity fire, drought and climate change impacts on ponderosa pine regeneration. Ecol. Model. 253, 56–69. doi: 10.1016/j.ecolmodel.2012.12.029
Field, J. P., Breshears, D. D., Law, D. J., Villegas, J. C., López-Hoffman, L., Brooks, P. D., et al. (2015). Critical zone services: expanding context, constraints, and currency beyond ecosystem services. Vadose Zone J. 14:vzj2014.10.0142. doi: 10.2136/vzj2014.10.0142
Gleason, K. E., Bradford, J. B., Bottero, A., D'Amato, A. W., Fraver, S., Palik, B. J., et al. (2017). Competition amplifies drought stress in forests across broad climatic and compositional gradients. Ecosphere 8:e01849. doi: 10.1002/ecs2.1849
Godfree, R. C., Knerr, N., Godfree, D., Busby, J., Robertson, B., and Encinas-Viso, F. (2019). Historical reconstruction unveils the risk of mass mortality and ecosystem collapse during pancontinental megadrought. Proc. Natl. Acad. Sci. U.S.A. 116, 15580–15589. doi: 10.1073/pnas.1902046116
Gornish, E. S., Aanderud, Z. T., Sheley, R. L., Rinella, M. J., Svejcar, T., Englund, S. D., et al. (2015). Altered snowfall and soil disturbance influence the early life stage transitions and recruitment of a native and invasive grass in a cold desert. Oecologia 177, 595–606. doi: 10.1007/s00442-014-3180-7
Grant, G. E., Tague, C. L., and Allen, C. D. (2013). Watering the forest for the trees: an emerging priority for managing water in forest landscapes. Front. Ecol. Environ. 11, 314–321. doi: 10.1890/120209
Hartmann, H., Moura, C. F., Anderegg, W. R. L., Ruehr, N. K., Salmon, Y., Allen, C. D., et al. (2018). Research frontiers for improving our understanding of drought-induced tree and forest mortality. New Phytol. 218, 15–28. doi: 10.1111/nph.15048
Hessburg, P. F., Miller, C. L., Parks, S. A., Povak, N. A., Taylor, A. H., Higuera, P. E., et al. (2019). Climate, environment, and disturbance history govern resilience of Western North American forests. Front. Ecol. Evol. 7:239. doi: 10.3389/fevo.2019.00239
Hulvey, K. B., Leger, E. A., Porensky, L. M., Roche, L. M., Veblen, K. E., Fund, A., et al. (2017). Restoration islands: a tool for efficiently restoring dryland ecosystems? Restor. Ecol. 25, S124–S134. doi: 10.1111/rec.12614
IPCCa (2014). “Climate change 2014: impacts, adaptation, and vulnerability. part a: global and sectoral aspects,” in Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds. C. B. Field, V. R. Barros, D. J. Dokken, K. J. Mach, M. D. Mastrandrea, T. E. Bilir, M. Chatterjee, K. L. Ebi, Y. O. Estrada, R. C. Genova, B. Girma, E. S. Kissel, A. N. Levy, S. MacCracken, P. R. Mastrandrea, and L. L. White (Cambridge; New York, NY: Cambridge University Press), p. 1132.
Katiyar, D., Hemantaranjan, A., and Singh, B. (2015). Chitosan as a promising natural compound to enhance potential physiological responses in plant: a review. Indian J. Plant Physiol. 20, 1–9. doi: 10.1007/s40502-015-0139-6
Keeley, J. E., Allen, C. D., Betancourt, J., Chong, G. W., Fotheringham, C. J., and Safford, H. D. (2006). A 21st century perspective on postfire seeding. J. For. 104, 103–104. doi: 10.1093/jof/104.2.103
Kolb, P. F., and Robberecht, R. (1996). High temperature and drought stress effects on survival of Pinus ponderosa seedlings. Tree Physiol. 16, 665–672. doi: 10.1093/treephys/16.8.665
Kumaraswamy, R. V., Kumari, S., Choudhary, R. C., Pal, A., Raliya, R., Biswas, P., et al. (2018). Engineered chitosan based nanomaterials: bioactivities, mechanisms and perspectives in plant protection and growth. Int. J. Biol. Macromol. 113, 494–506. doi: 10.1016/j.ijbiomac.2018.02.130
Law, D. J., Adams, H. D., Breshears, D. D., Cobb, N. S., Bradford, J. B., Zou, C. B., et al. (2019). Bioclimatic envelopes for individual demographic events driven by extremes: plant mortality from drought and warming. Int. J. Plant Sci. 180, 53–62. doi: 10.1086/700702
Li, R., He, J., Xie, H., Wang, W., Bose, S. K., Sun, Y., et al. (2019). Effects of chitosan nanoparticles on seed germination and seedling growth of wheat (Triticum aestivum L.). Int. J. Biol. Macromol. 126, 91–100. doi: 10.1016/j.ijbiomac.2018.12.118
Madsen, M. D., Svejcar, L., Radke, J., and Hulet, A. (2018). Inducing rapid seed germination of native cool season grasses with solid matrix priming and seed extrusion technology. PLoS ONE 13:e0204380. doi: 10.1371/journal.pone.0204380
McDowell, N., Allen, C. D., Anderson-Teixeira, K., Brando, P., Brienen, R., Chambers, J., et al. (2018). Drivers and mechanisms of tree mortality in moist tropical forests. New Phytol. 219, 851–869. doi: 10.1111/nph.15027
McDowell, N., Pockman, W. T., Allen, C. D., Breshears, D. D., Cobb, N., Kolb, T., et al. (2008). Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol. 178, 719–739. doi: 10.1111/j.1469-8137.2008.02436.x
McDowell, N. G., Adams, H. D., Bailey, J. D., Hess, M., and Kolb, T. E. (2006). Homeostatic maintenance of ponderosa pine gas exchange in response to stand density changes. Ecol. Appl. 16, 1164–1182. doi: 10.1890/1051-0761(2006)016[1164:HMOPPG]2.0.CO;2
McDowell, N. G., Grossiord, C., Adams, H. D., Pinzón-Navarro, S., Mackay, D. S., Breshears, D. D., et al. (2019). Mechanisms of a coniferous woodland persistence under drought and heat. Environ. Res. Lett. 14:045014. doi: 10.1088/1748-9326/ab0921
Millar, C. I., and Stephenson, N. L. (2015). Temperate forest health in an era of emerging megadisturbance. Science 349, 823–826. doi: 10.1126/science.aaa9933
Millar, C. I., Stephenson, N. L., and Stephens, S. L. (2007). Climate change and forests of the future: managing in the face of uncertainty. Ecol. Appl. 17, 2145–2151. doi: 10.1890/06-1715.1
Moreira, F., Viedma, O., Arianoutsou, M., Thomas, C., Koutsias, N., Rigolot, E., et al. (2011). Landscape – wildfire interactions in southern Europe: implications for landscape management. J. Environ. Manag. 92, 2389–2402. doi: 10.1016/j.jenvman.2011.06.028
Mu, Q., Zhao, M., Kimball, J. S., McDowell, N. G., and Running, S. W. (2013). A remotely sensed global terrestrial drought severity index. Bull. Am. Meteorol. Soc. 94, 83–98. doi: 10.1175/BAMS-D-11-00213.1
Naumann, G., Alfieri, L., Wyser, K., Mentaschi, L., Betts, R. A., Carrao, H., et al. (2018). Global changes in drought conditions under different levels of warming. Geophys. Res. Lett. 45, 3285–3296. doi: 10.1002/2017GL076521
O'Brien, M. J., Engelbrecht, B. M., Joswig, J., Pereyra, G., Schuldt, B., Jansen, S., et al. (2017). A synthesis of tree functional traits related to drought-induced mortality in forests across climatic zones. J. Appl. Ecol. 54, 1669–1686. doi: 10.1111/1365-2664.12874
O'Sullivan, O. S., Heske, M. A., Reich, P. B., Tjoelker, M. G., Weerasinghe, L. K., Penillard, A., et al. (2017). Thermal limits of leaf metabolism across biomes. Glob. Chang. Biol. 23, 209–223. doi: 10.1111/gcb.13477
Panagos, P., Borrelli, P., Poesen, J., Ballabio, C., Lugato, E., and Meusburge, K. (2015). The new assessment of soil loss by water erosion in Europe. Environ. Sci. Policy 54, 438–447. doi: 10.1016/j.envsci.2015.08.012
Petrie, M. D., Bradford, J. B., Hubbard, R. M., Lauenroth, W. K., Andrews, C. M. and Schlaepfer, D. R. (2017). Climate change may restrict dryland forest regeneration in the 21st century. Ecology 98, 1548–1559. doi: 10.1002/ecy.1791
Polle, A., Chen, S. L., Eckert, C., and Harfouche, A. (2019). Engineering drought resistance in forest trees. Front. Plant. Sci. 9:1875. doi: 10.3389/fpls.2018.01875
Prudhomme, C., Giuntoli, I., Robinson, E. L., Clark, D. B., Arnell, N. W., Dankers, R., et al. (2014). Hydrological droughts in the 21st century, hotspots and uncertainties from a global multimodel ensemble experiment. Proc. Natl. Acad. Sci. U.S.A. 111, 3262–3267. doi: 10.1073/pnas.1222473110
Redmond, M. D., Law, D. J., Field, J. P., Meneses, N., Carroll, C. J. W., Wion, A. P., et al. (2019). Perspective. Targeting extreme events: complementing near-term ecological forecasting with rapid experiments and regional surveys. Front. Environ. Sci. 7:183. doi: 10.3389/fenvs.2019.00183
Romme, W. H., Allen, C. D., Bailey, J. D., Baker, W. L., Bestelmeyer, B. T., Brown, P. M., et al. (2009). Historical and modern disturbance regimes, stand structures, and landscape dynamics in piñon–juniper vegetation of the western United States. Rangeland Ecol. Manag. 62, 203–222. doi: 10.2111/08-188R1.1
Rosot, M. A. D., Maran, J. C., da Luz, N. B., Garrastazú, M. C., de Oliveira, Y. M. M., Franciscon, L., et al. (2018). Riparian forest corridors: a prioritization analysis to the landscape sample units of the Brazilian national forest inventory. Ecol. Indic. 93, 501–511. doi: 10.1016/j.ecolind.2018.03.071
Ruthrof, K. X., Breshears, D. D., Fontaine, J. B., Froend, R. H., Matusick, G., Kala, J., et al. (2018). Subcontinental heat wave triggers terrestrial and marine, multi-taxa responses. Sci. Rep. 8:13094. doi: 10.1038/s41598-018-31236-5
Savage, M., Mast, J. N., and Feddema, J. J. (2013). Double whammy: high-severity fire and drought in ponderosa pine forests of the Southwest. Can. J. For. Res. 43, 570–583. doi: 10.1139/cjfr-2012-0404
Schwalm, C. R., Anderegg, W. R., Michalak, A. M., Fisher, J. B., Biondi, F., Koch, G., et al. (2017). Global patterns of drought recovery. Nature 548, 202–205. doi: 10.1038/nature23021
Sharp, R. (2013). A review of the applications of chitin and its derivatives in agriculture to modify plant-microbial interactions and improve crop yields. Agronomy 3, 757–793. doi: 10.3390/agronomy3040757
Shive, K. L., Preisler, H. K., Welch, K. R., Safford, H. D., Butz, R. J., O'Hara, K. L., et al. (2018). From the stand scale to the landscape scale: predicting the spatial patterns of forest regeneration after disturbance. Ecol. Appl. 28, 1626–1639. doi: 10.1002/eap.1756
Sperry, J. S., Wang, Y., Wolfe, B. T., Mackay, D. S., Anderegg, W. R., McDowell, N. G., et al. (2016). Pragmatic hydraulic theory predicts stomatal responses to climatic water deficits. New Phytol. 212, 577–589. doi: 10.1111/nph.14059
Stahle, D. W., Fye, F. K., Cook, E. R., and Griffin, R. D. (2007). Tree-ring reconstructed megadroughts over North America since AD 1300. Clim. Change 83:133. doi: 10.1007/s10584-006-9171-x
Stephens, S. C., Collins, B. M., Fettig, C. J., Finney, M. A., Hoffman, C. M., Knapp, E. E., et al. (2018). Drought, tree mortality, and wildfire in forests adapted to frequent fire. BioScience 68:77–88. doi: 10.1093/biosci/bix146
Stevens-Rumann, C. S., Kemp, K. B., Higuera, P. E., Harvey, B. J., Rother, M. T., Donato, D. C., et al. (2018). Evidence for declining forest resilience to wildfires under climate change. Ecol. Lett. 21, 243–252. doi: 10.1111/ele.12889
Sutherland, I. J., Gergel, S. E., and Bennett, E. M. (2016). Seeing the forest for its multiple ecosystem services: indicators for cultural services in heterogeneous forests. Ecol. Indic. 71, 123–133. doi: 10.1016/j.ecolind.2016.06.037
Swann, A. L. S., Laguë, M. M., Garcia, E. S., Field, J. P., Breshears, D. D., Moore, D. J. P., et al. (2018). Continental-scale consequences of tree die-offs in North America: identifying where forest loss matters most. Environ. Res. Lett. 13:055014. doi: 10.1088/1748-9326/aaba0f
Tyree, M. T., and Zimmermann, M. H. (2002). Xylem Structure and the Ascent of Sap. New York, NY: Springer. doi: 10.1007/978-3-662-04931-0
Úradníček, L., Šrámek, M., and Dreslerová, J. (2017). Checklist of champion trees in the czech republic. J. Landsc. Ecol. 10, 109–120. doi: 10.1515/jlecol-2017-0020
Westerling, A. L., Hidalgo, H. G., Cayan, D. R., and Swetnam, T. W. (2006). Warming and earlier spring increase western US forest wildfire activity. Science 313, 940–943. doi: 10.1126/science.1128834
Will, R. E., Wilson, S. M., Zou, C. B., and Hennessey, T. C. (2013). Increased vapor pressure deficit due to higher temperature leads to greater transpiration and faster mortality during drought for tree seedlings common to the forest-grassland ecotone. New Phytol. 200, 366–374. doi: 10.1111/nph.12321
Williams, A. P., Allen, C. D., Macalady, A. K., Griffin, D., Woodhouse, C. A., Meko, D. M., et al. (2013). Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Change 3, 292–297. doi: 10.1038/nclimate1693
Woodford, J. (2012). The Wollemi Pine: the Incredible Discovery of a Living Fossil from the Age of the Dinosaurs. Melbourne: Text Publishing.
Xia, L., Song, X., Fu, N., Cui, S., Li, L., Li, H., et al. (2019). Effects of forest litter cover on hydrological response of hillslopes in the Loess Plateau of China. Catena 181:104076. doi: 10.1016/j.catena.2019.104076
Keywords: drought, die-off, forest management, nanochitosan, mortality, extreme events, ecological forecasting, wildfire
Citation: Field JP, Breshears DD, Bradford JB, Law DJ, Feng X and Allen CD (2020) Forest Management Under Megadrought: Urgent Needs at Finer Scale and Higher Intensity. Front. For. Glob. Change 3:502669. doi: 10.3389/ffgc.2020.502669
Received: 04 October 2019; Accepted: 20 November 2020;
Published: 23 December 2020.
Edited by:
Thomas Cordonnier, Institut National de Recherche pour l'Agriculture, l'Alimentation et l'Environnement (INRAE), FranceReviewed by:
Peter Brang, Swiss Federal Institute for Forest, Snow and Landscape Research (WSL), SwitzerlandBenjamin Issac Cook, National Aeronautics and Space Administration (NASA), United States
Copyright © 2020 Field, Breshears, Bradford, Law, Feng and Allen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jason P. Field, anBmaWVsZEBlbWFpbC5hcml6b25hLmVkdQ==