
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change , 15 January 2020
Sec. Forest Disturbance
Volume 2 - 2019 | https://doi.org/10.3389/ffgc.2019.00091
This article is part of the Research Topic Invasions and Impacts of Exotic Species in Forests View all 6 articles
 Alain Roques1,2*
Alain Roques1,2* Juan Shi2,3
Juan Shi2,3 Marie-Anne Auger-Rozenberg1,2Lili Ren2,3
Marie-Anne Auger-Rozenberg1,2Lili Ren2,3 Sylvie Augustin1,2You-qing Luo2,3
Sylvie Augustin1,2You-qing Luo2,3Forests are increasingly threatened worldwide by the establishment of non-native species arriving from other continents with globalization of trade and international travels. We compiled comprehensive lists of non-native insects associated to woody plants in Europe and China to compare their invasive patterns between these two regions. By 2019, a total of 449 species have colonized forest, ornamental and fruits trees and shrubs in Europe whereas only 77 species were reported in China, a likely underestimated number because studies concentrated on economically-important species. Approximately 6.1 new species have been detected per year since 2000 in Europe vs. only 1.9 in China. Europe and China also exhibited distinct patterns in the invasion sources. Most non-native species recorded in Europe originated from Asia (37.2%), and more especially from China (22.9%) whereas Europe was only a minor source for the insect invasions in China (9.1%), which predominantly originated from other parts of Asia (40.3%). These different patterns likely reflect the dissimilar trends of the trade in ornamental plants. Taxonomic and guild composition of non-native insects in both regions confirmed that ornamental trade of plants for planting including fruit trees played a larger role than timber and wood trade in shaping the non-native entomofaunas associated to woody plants. Hemiptera, and subsequently sap suckers, largely dominated the non-native entomofauna in both regions. However, there were striking differences in the range of hosts these species colonized. Although fruit trees, and especially Citrus, are the most important hosts in both regions, tropical trees including palms were also highly colonized in China whereas deciduous trees, shrubs and conifers were proportionally more affected in Europe. Quite a half of the non-native insect species did not switch from their original, non-native host to natives ones in Europe whereas only a minority of these insect invaders still sticked to their original, non-native host in China. It may be due to the more important presence in China of native woody plants taxonomically close to the original host.
Along with globalization of economies, expanding world-wide trade, and climate change, the rate at which humans translocate species beyond their native ranges has substantially increased during the last centuries with no sign of saturation (Seebens et al., 2017), the more as new source species pools become accessible due to continuous changes in trade routes and imported goods (Seebens et al., 2018). This process leads to permanent additions to local floras and faunas, which are susceptible to affect ecosystem functioning and economies (Vilà et al., 2010; Simberloff et al., 2013). Since insects represent a large part of these non-native species translocated from one continent to another (DAISIE, 2009), forests like other terrestrial ecosystems are increasingly threatened by the establishment and spread of non-native insects worldwide (Brockerhoff and Liebhold, 2017). Regional assemblages of such non-native insect species have recently been characterized for 20 world regions, including North America, Japan and Europe (Liebhold et al., 2016), showing that plant diversity drives global patterns of insect invasions (Liebhold et al., 2018).
In Europe, the unprecedented building-up of the pan-European database of non-native species from the DAISIE (Delivering Alien Species Inventories in Europe; www.europe-aliens.org) project launched in 2008 allowed to show that the rate of establishment of non-native insect species has nearly doubled over the last few decades (DAISIE, 2009). Indeed, it increased from an average of 10.9 species per year for the period 1950–1974 to an estimated 19.6 species per year for 2000–2008 (Roques et al., 2010). Moreover, this recent influx of non-native insects appears to involve predominately accidentally-introduced herbivores and, among them, ca. 75% associated to woody plants (Roques, 2010). This is probably related to recent changes in trade practices, especially the increased magnitude in the movement of live plants globally (Liebhold et al., 2012; Eschen et al., 2014; Essl et al., 2015). Thus, Roques (2015) estimated the number of species associated with woody plant newly arriving per year in Europe to have increased from 1.8 during the first half of the 20th century to 6.7 during 2000–2014, to finally result in the presence of 385 non-native species on the continent in 2014 (Eschen et al., 2015). At country level, Matošević and Pajač Živković (2013) noticed a similar acceleration in Croatia with 6.4 new species establishing per year on woody plants during the period 2007–2012 vs. only 1.8 during 2002–2007.
Extensive surveys of the non-native entomofauna associated with woody plants carried out in North America and New Zealand revealed similar trends in invasion patterns (Brockerhoff and Liebhold, 2017). In the USA, Aukema et al. (2010) estimated the detection of new species at about 2.5 per year for a total of 455 established non-native species, involving 64 families and eight orders. In an earlier study, Langor et al. (2009) found 419 non-native species on woody plants in Canada but noticed that their arrival rate may have decreased following the Quarantine legislation enacted in 1976 in this country. In both North America and Europe, hemipteran species largely dominate the non-native entomofauna associated to woody plants, representing 41–53% of the total species (Langor et al., 2009; Aukema et al., 2010; Roques, 2015).
However, such surveys do not appear to be as extensive in other countries where studies tended to concentrate on economically- important species, and the resulting numbers of established non-native species may not reflect reality. Thus, Schühli et al. (2016) only listed 22 non-native insect species associated with woody plants, mostly eucalypts, in Brazil. In China, Ji et al. (2011) reported 34 non-native forest pests established by 2008. They however noticed that among the 207 species found on Eucalypts planted in China, none originating from Australia were present until the record in 2007 of the blue gum chalcid, Leptocybe invasa. Then, in a tentative inventory of invasive species in China, Xu et al. (2012a) provided a list of 92 non-native insects, of which 40 species associated to woody plants. Among 125 non-native insect species related to agriculture and forestry, Wan and Yang (2016) recorded 19 species associated with forests in 2014. The authors also showed that the cumulative number of non-native insect species tended to grow exponentially, with 70% of the species recorded after 1950. In 2017, Lu and Sun (2017) gave a list of 27 non-native forest pests but their work essentially described invasive patterns of economically- important case studies; i.e., fall webworm (Hyphantria cunea), Sirex woodwasp (Sirex noctilio), Blue gum chalcid (Leptocybe invasa), and Loblolly pine mealybug (Oracella acuta). More recently, Xian et al. (2018) listed 52 new invasive insect species having affected agricultural and forestry ecosystems of China during the last 20 years whereas Qi and Lu (2018) recorded 23 non-native species of which 17 associated with woody plants in the tropical provinces of China. These successive lists of non-native insects established on woody plants in China thus provided variable data on both the species considered and the dates and places of first record in the country. Additional species were recently recorded, e.g., the platypodid Euplatypus parallelus restricted to Hainan Island (Li et al., 2018), but others appeared missing in these lists although may-be introduced long ago, e.g., the eucalyptus tortricid Strepsicrates semicanella (Nair, 2001; Deng et al., 2011) or the eucalypt psyllid, Blastopsylla occidentalis (Yen et al., 2013).
As Wan and Jiang (2017) noticed in their book “Biological invasions and its management in China,” the numbers of non-native species in China was thus probably underestimated due to the overlooking of species with low density and rarely collected at early stages of invasions, or with no substantial consequences on human health and/or on economy. For example, only four non-native ant species were reported by Wan and Yang (2016) whereas Guénard and Dunn (2012) identified 15 ant species as clearly non-native to China, 32 others being listed as either potentially non-native or may-be native to some regions of China. These problems potentially create some bias in using lists of non-native species. This is not proper to China. In 1994, Mattson et al. (1994) listed 368 non-native phytophagous species on woody plants in USA, but 16 years later Aukema et al. (2010) revised this total to 455by. The gap was even more important between the records from Mattson et al. (2007), who only listed 109 non-native insects on woody plants in Europe, and these from Roques (2015) who reported 7 years later the establishment of 3 times more species in the same continent. The differences partly resulted from an increasing number of arrivals during the meantime. However, comparing Mattson's list with the DAISIE data also revealed that this list has underestimated the numbers of established non-native species in minor groups such as seed chalcids, scales and midges. Therefore, the hypothesis of Mattson et al. (2007) about the legacy of the European Pleistocene/Holocene crucible leading to fewer immigrant phytophagous insects on woody plants in Europe than in North America seems highly disputable at present.
Since Asia, and more especially China, has become the major supplier of non-native insects to Europe during the last decades (Roques, 2010, 2015), probably in relation with the recent strong increase in the trade of plants for planting from China to Europe (a 6-fold increase between 2000 and 2010; Eschen et al., 2014), it appeared interesting to check whether the invasive patterns differ between these two parts of Eurasia. Therefore, the objectives of the present paper are (i) to provide comprehensive checklists of the non-native insect species associated to woody plants in Europe and China, and (ii) to compare between the two regions the invasive patterns of such non-native species in origin, taxa and feeding guilds, and their variations over time.
We compiled comprehensive lists of non-native insect species associated with woody plants in Europe and China. Only the species developing on trees, shrubs, and vines, for at least part of their life cycle, were considered. We excluded ants as well as species damaging bananas (Musa spp. and others), bamboos and Agavaceae, but we considered those damaging monocots showing a kind of woody trunk such as palms (Arecaceae) and Dracenacae. The insect species only observed in greenhouses but not in the wild were also excluded.
For Europe, we considered all species established in each of the 41 countries of geographic Europe, including major islands or archipelagoes (Balearics, Corsica, Canaries, Crete, Faeroe, Ionian islands, Madeira, North Aegean islands, Sardinia, Sicily, South Aegean islands, Svalbard) but not the Azores that were considered too much distant. We first built a list of non-native insects by extracting the species associated to woody plants mentioned in the tables supplied in the different chapters of the two volumes of the book “Alien terrestrial arthropods of Europe” (Roques et al., 2010), which was an output of the DAISIE project. For each insect order, the corresponding table included data on the plant hosts, insect native range, date and country of first record in Europe, colonized habitat, and invaded countries. We thus screened Sauvard et al. (2010) for Curculionidae including bark beetles, Cocquempot and Lindelöw (2010) for Cerambycidae, Beenen and Roques (2010) for Chrysomelidae, Denux and Zagatti (2010) for other coleopteran families, Rabitsch (2010) for Heteroptera true bugs, Mifsud et al. (2010) for Aleyrodidae, Phylloxeroidea, and Psylloidea and Hemiptera Auchenorrhyncha, Pellizzari and Germain (2010) for Coccoidea scales, Coeur d'acier et al. (2010) for Aphididae, Skuhravá et al. (2010) for Diptera, Lopez-Vaamonde et al. (2010) for Lepidoptera, Rasplus et al. (2010) for Hymenoptera, Reynaud (2010) for Thysanoptera, and Rasplus and Roques (2010) for Isoptera and Phasmatodea. Then, we updated this list up to July 2019 by retrieving information about recent arrivals of non-native insect species from the EASIN catalog (European Alien Species Information Network), (Katsanevakis et al., 2015; http://easin.jrc.ec.europa.eu) and EPPO Informations Services (https://gd.eppo.int/). Finally, using the browsers Google Scholar and Science Direct we completed the list through a literature search for papers published between 2008 and 2019 matching a combination of the following keywords: Insect AND (Forest OR Tree OR Shrub OR Ornamental) AND (Non-native OR Invasive OR Alien OR Exotic) AND (Europe OR Albania OR Austria OR Belgium OR Bosnia OR Britain OR Belarus OR Bulgaria OR Croatia OR Cyprus OR Czech Republic OR Denmark OR Estonia OR European Russia OR Finland OR France OR FYRM OR Germany OR Greece OR Hungary OR Ireland OR Italy OR Latvia OR Lithuania OR Luxembourg OR Malta OR Montenegro OK Netherlands OR Poland OR Portugal OR Romania OR Serbia OR Slovakia OR Slovenia OR Spain OR Sweden OR Ukraine OR United Kingdom). The last query was applied 15 July 2019.
The database we finally constructed (Supplementary Material S1) thus included the following data: 1- Species taxonomy (Order, family); 2- Status (a- non-native; b- cryptogenic); 2- Date of first record in Europe; 3- Country of first record in Europe; 4- Origin of the species (a- Africa; b- China; c- Asia other than China; d- Australasia; e- Northern America; f- Central and Southern America; g- Tropical; h- Unknown); 5- Host categorization (a- attacks only non-native plants; b- has switched on native plants); 6- Host group (a- conifers; b- deciduous; c- shrubs; d- fruit trees including Citrus; e- eucalypts; f- exotic legume trees; g- palms, dracenacae and cycads; h- other tropical woody plants; i- drywood); 7- feeding guild (a- external defoliator; b- bud/shoot feeder; c- leaf miner; d- sap sucker; e- gall maker; f- xylophagous insect; g- root feeder; h- flower feeder; i- fruit insect; j- seed insect; k- unknown); 8- references.
For China, we considered all species established in mainland China, including Hainan and Hong-Kong but excluded those only established in Taiwan. We first visited the China invasive species database (http://www.chinaias.cn/wjPart/index.aspx), then we used the lists from Ji et al. (2011), Xu et al. (2012a), Wan and Yang (2016), Lu and Sun (2017), Qi and Lu (2018), and Xian et al. (2018) but similarly as for Europe we additionally retrieved information about recent arrivals of non-native species from bibliographic databases research using the Chinese research browser CNKI (www.cnki.net) and Science Direct. We used the same keyword combination as before but replacing Europe and its constitutive countries by the keyword “China.” From the former lists, we excluded the species which appeared to be agricultural pests but not related to woody plants, namely Liryomiza sativae, Trialeurodes vaporarium and Opogona sacchari, as well ants (Solenopsis spp.) and termites except the drywood pest Incisitermes minor, but we kept the fruit flies (Bactrocera spp.) because they are damaging tree fruits. We also deleted species that appeared to be only present in Taiwan (Aegus philippinensis mentionned in Wan and Yang, 2016), species only present under glasshouses but not in the wild (Phenacoccus philippinensis mentionned in Xian et al., 2018 but only observed under glasshouses in Beijing City; Trencheva et al., 2010; Wang and Wu, 2010), and species first identified as non-natives but later shown to correspond to a native species (pear aphid, Aphanostigma piry, mentioned in Xu et al., 2012b, but corresponding to A. iakusuiense; Tao, 1999). Whereas, it was rather easy to consider arrivals in Europe of non-native species originating from other continents, it was difficult to categorize a number of southeastern Asian species as non-native in China because their native range may include part of Southern China. We finally decided to exclude from the list of non-native species two teak defoliators, the teak skeletoniser, Eutectona machaeralis (Pyralidae) (Nair, 2001) and Hyblaea puera (Hyblaeidae) (Chen and Wu, 1984), because these species are largely distributed throughout south Asia and because teak was introduced at least 100 years ago in China (Nair, 2001). The database of the non-native species established in China on woody plants (Supplementary Material S2) was finally built in a similar way as the European one, with the same categories except for 3- Date of first record in China, 4- Province of first record in China, and 5- Origin where Europe replaced China.
The distribution pattern of the non-native insect species within regions of origin, taxonomic orders, host groups and feeding guilds was compared between Europe and China. The similarity of the distribution between the two regions was tested using chi-square tests of homogeneity with simulated Monte Carlo p-values (Hope, 1968) due to low sample size. The distribution was considered to differ when P < 0.05. Then, Bayesian Multinomial-Dirichlet models were fitted to compare more specifically differences between each group in the two regions. The temporal trends in new records of non-native insect species by 10-year periods were compared between Europe and China by fitting log-linear models of the form log(1 + N) = a t + b, with t the date of record and N the number of newly-recorded species during the preceding 10 years. The parameter a is the exponential rate of increase, each 10-year period, N + 1 being increased by a factor e10 a. Similarly as in Seebens et al. (2017), the time interval 2010–2019 was excluded from the calculations because these recent data are likely to be influenced by a reduced sampling intensity due to delays in detection and reporting of new non-native species. All analyses were performed using R© version 3.6.1 (R Core Team, 2019).
A total of 449 non-native species (of which 27 cryptogenic ones) associated with woody plants were recorded in Europe by July 2019 (Figure 1A; Supplementary Material S1) whilst in China this total only amounted 77 species of which two cryptogenic ones (Figure 1A; Supplementary Material S2). An exponential acceleration in the establishment records was observed during the last 50 years in Europe, with a mean of ca. 6.1 new records per year during the period 2000–2019 to be compared to 2.4 records per year during 1950–1970 (Figure 1B). This acceleration was also observed in China, but at a slower rate (1.9 new records per year during 2000–2019 vs. 0.2 during 1950–1970) (Figure 1B). Over the whole period of observation, the exponential rates of increase were comparable between Europe (a = 0.014; 95% CI: 0.010, 0.018; R2 = 0.81; Figure 1C) and China (a = 0.018; 95% CI: 0.003, 0.03; R2 = 0.43; Figure 1C), corresponding approximately to an average 15–20% increase (e10 a ≈ 1.15 − 1.20) in the number of new records every 10 years.
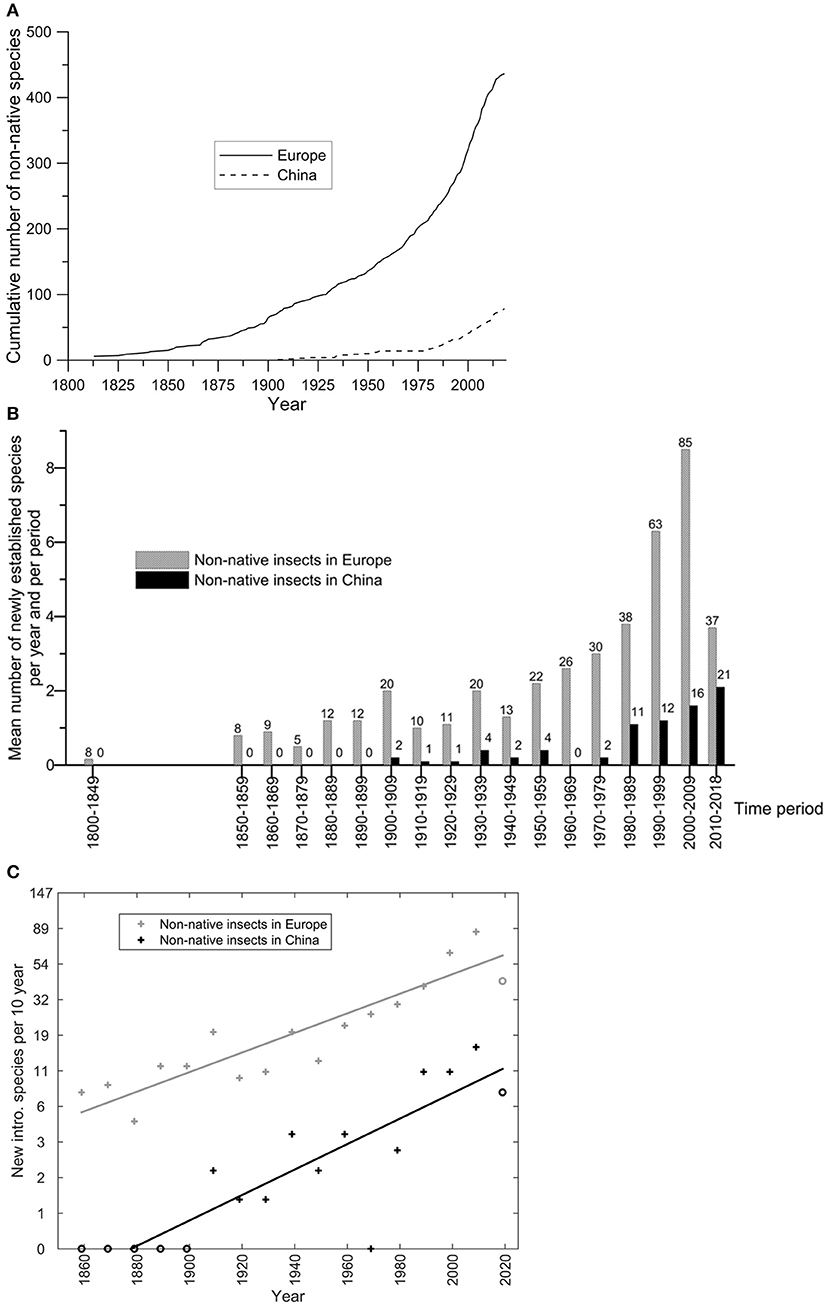
Figure 1. (A) Cumulative number of detections (i.e., new establishments) of non-native insect species associated to woody plants over time in Europe and China; (B) Temporal changes in the mean number of new records per year of non-native species associated to woody plants having established in Europe and China up to 2019. The number above each bar indicates the total number of non-native species observed per 10-year period; (C) Fits of log-linear models log (1 + N) = a t + b on the data shown in (B). The circles correspond to the data that have not been taken into account during the fitting procedure. The y-axis is represented in log-scale, the linear trend thus indicating an exponential increase.
The origin of the non-native species significantly differed between Europe and China (Figure 2; chi-square test of homogeneity, p < 0.000; Supplementary Material S3). Most of the non-native species observed in Europe on woody plants originated from Asia (37.2% of the total), largely outcompeting North America (25.8%) whereas the other continents gave minor inputs (Figure 2). Species originating from China dominated the non-native species arriving from Asia (103 species vs. 64 for the other Asian countries). The predominance of Asian species, and especially the Chinese ones, over the species coming from North America was observed since the 1980s (Figure 3). The non-native species recorded in China predominantly came from other Asian regions (i.e., southeast Asia and Japan; 40.3%), followed by Central and South America (20.8%) whereas North America and Europe contributed much less (14.3 and 9.1% of the total, respectively) (Figure 2). The predominance in China of non-native species originating from other Asian regions over the North American ones was also observed since the 1970s (Figure 3).
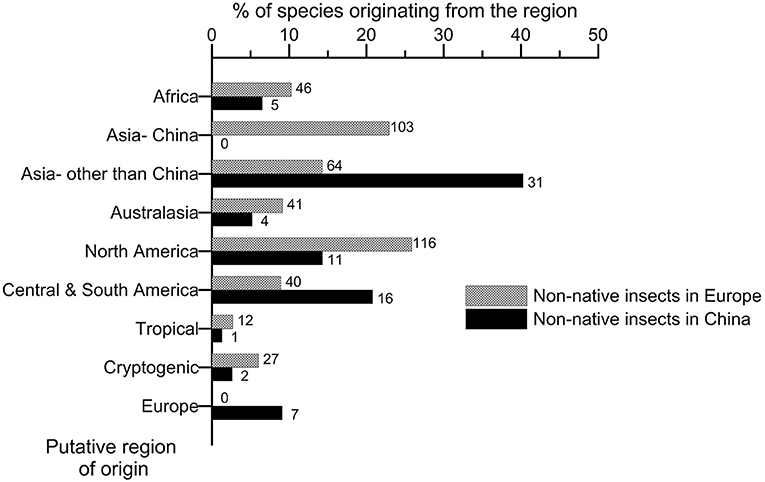
Figure 2. Comparison of the regions of origin of the non-native species associated to woody plants having established in Europe and China up to 2019. The number on the right of each bar indicates the total number of non-native species observed per region of origin.
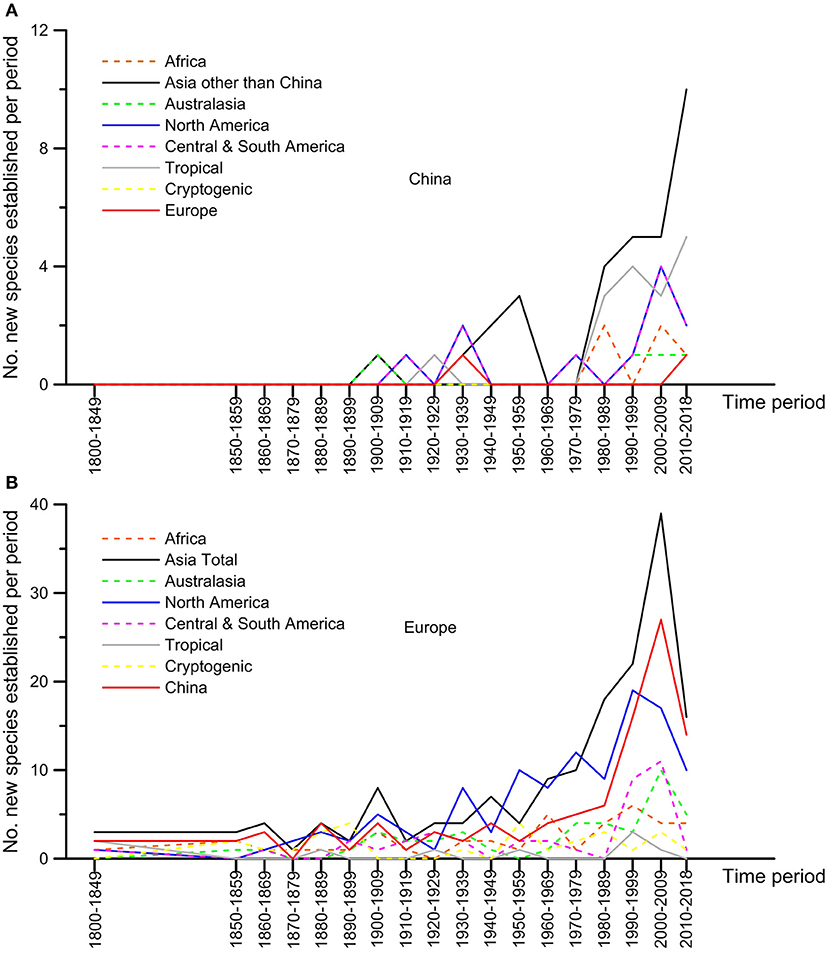
Figure 3. Temporal changes in the regions of origin of the non-native species associated to woody plants having established in Europe and China up to 2019.
The taxonomic composition of the non-native entomofaunas associated to woody plants did not differ significantly between Europe and China when insect orders were considered (Figure 3; chi-square test of homogeneity, p = 0.230, Supplementary Material S4). In both cases, Hemiptera dominated, accounting for 53.2% of the total in Europe and 42.9% in China, followed by Coleoptera (21.4 and 24.7%, respectively). In contrast, large differences existed at family level in Hemiptera (Figure 4). The dominant family in Europe, Aphididae (61 species accounting for 13.6% of the total species), had a limited contribution in the Chinese records (four species accounting for 5.2% of the total). Non-native Diaspididae scales were also quite absent in China (2 vs. 50 species in Europe; 2.6 vs. 11.1 of the total species), scales being mostly represented by Pseudococcidae (13.0% of the total vs. 4.7% in Europe). However, the contribution of another major family, Curculionidae, including weevils and bark and ambrosia beetles was quite similar in both Europe (9.8%) and China (7.8%). Tephritidae fruit flies also appeared an important family in China (7.8% of the total) whereas it was very minor in Europe (1.3%). In both regions, no significant change in the taxonomic composition of the non-native entomofauna was observed during the last century, with a constant dominance of Hemipteran species (Supplementary Material S5).
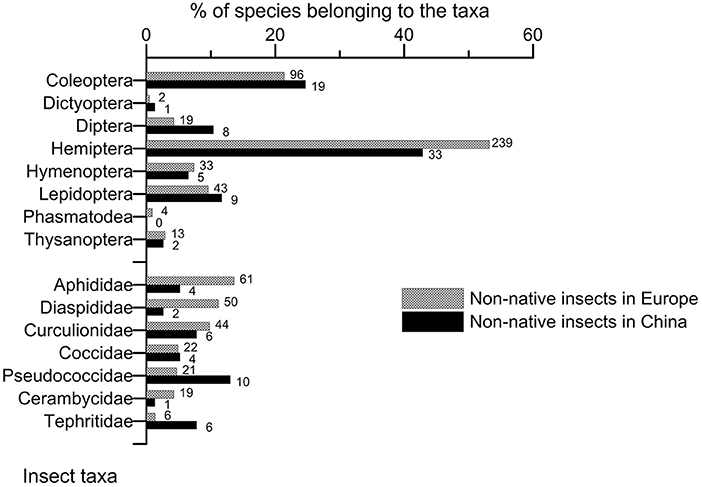
Figure 4. Comparison of the relative importance of the different taxonomic orders and main families in the non-native entomofauna associated to woody plants having established in Europe and China up to 2019. The number on the right of each bar indicates the total number of non-native species observed per taxa.
The feeding guilds exhibited by the non-native entomofaunas did not differ significantly between Europe and China (Figure 5; chi-square test of homogeneity, p = 0.262, Supplementary Material S6), essentially because sap suckers largely dominated in both regions (55.5 and 45.5% of the total species in Europe and China, respectively). However, this guild was proportionally more important in Europe than in China, especially since the 1960s (Supplementary Material S7). Two feeding guilds appeared proportionally more represented in China, external defoliators (14.3. vs. 7.6% in Europe and China, respectively) and species attacking tree fruits (7.8 vs. 2.7% in Europe) whilst the proportion of xylophagous species was nearly equal (19.5 vs. 16.3% in Europe and China, respectively) (Figure 5).
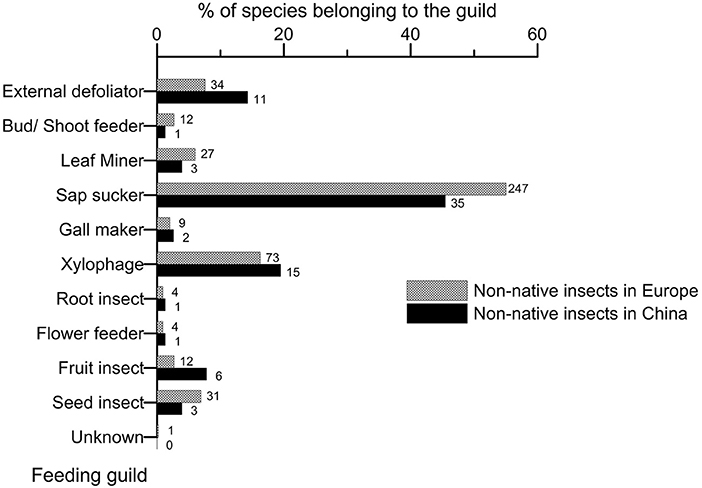
Figure 5. Comparison of the feeding guild shown by the non-native species associated to woody plants having established in Europe and China up to 2019. The number on the right of each bar indicates the total number of non-native species observed per guild. Note that a species may belong to several guilds depending on its development stages.
In Europe, a large part of the non-native species yet sticked to their non-native host (45.2% of the total species; Figure 6) whereas most succeeded in switching to native hosts in China (81.8%). The colonized host groups significantly differed between Europe and China (Figure 7; chi-square test of homogeneity, p = 0.031), essentially because of the relative colonization of fruit trees (Supplementary Material S8). Although fruit trees were more colonized than other woody plants in both regions, the related insect species accounted for 40.6% of the total species in China but only for 29.2% in Europe. Ornamental shrubs and deciduous trees were quite equally colonized by ca. 23% each of the non-native species in Europe whereas palms and ornamental shrubs represented each 17–19% of the hosts in China. Palms appeared proportionally more colonized in China since the 1990s (Supplementary Material S9). In both regions, non-native species were less recorded on conifers (16.3% in Europe and 10.9% in China) than on the previous groups of woody plants.
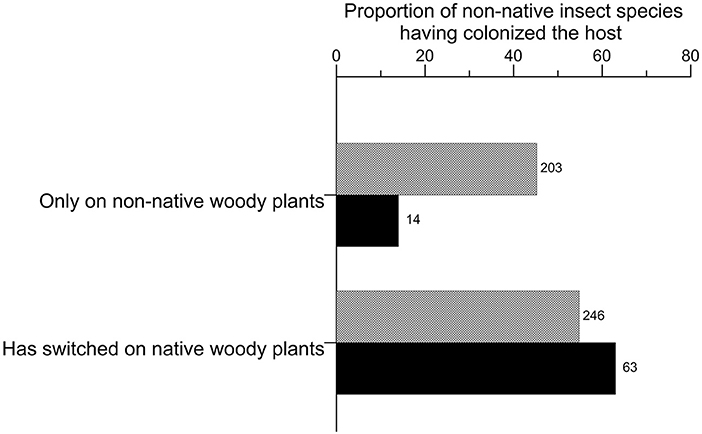
Figure 6. Switching processes of non-native species associated to woody plants having established in Europe and China up to 2019. The number on the right of each bar indicates the total number of non-native species observed per guild.
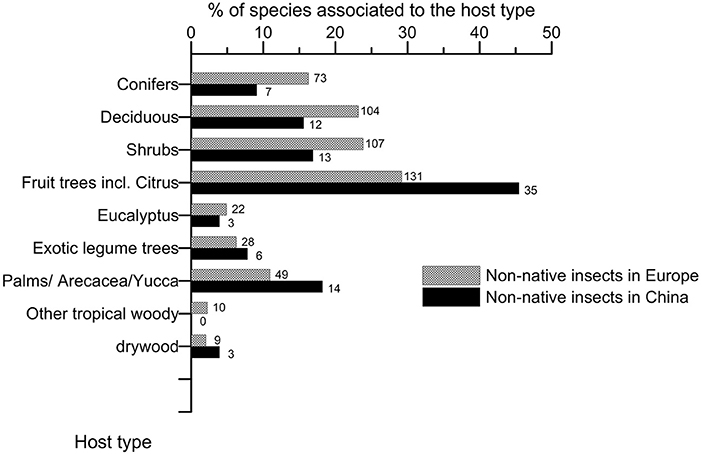
Figure 7. Comparison of the host types colonized by the non-native species associated to woody plants having established in Europe and China up to 2019. The number on the right of each bar indicates the total number of non-native species observed per host type. Note that a species may colonize several types of hosts.
Two-third (66.5%) of the first records of the non-native species in Europe were located in the southern countries (297 records, and essentially mainland Italy- 107 records and mainland France- 80 records), followed by western Europe (87 records, essentially Great Britain- 47 records), Central Europe (20 records), Eastern Europe (25 records), and Northern Europe (7 records). In China, when the location was known more than 50% of the first records also concerned the southern part of the country; i.e., the island of Hainan (13 records) and the provinces of Guangdong (12 records plus 3 records in Hong Kong), Guangxi (5 records) and Yunnan (5 records). However, the north-western province of Xinjiang was also the first place where several non-native species related to fruit trees were first observed (5 records).
Our study confirmed the exponential increase in the establishment of non-native insect species related to woody plants in Eurasia during the last decades. This has already been noticed by several authors in both Europe and China (Roques, 2010, 2015; Eschen et al., 2015; Wan and Yang, 2016) but the numbers of species indicated by these authors appeared largely underestimated, and thus probably dampered the observation of the actual increase. Actually, we recorded 449 non-native species related to woody plants in Europe instead of the 385 ones noticed by Eschen et al. (2015). For China, the number of 77 non-native species associated to woody plants we listed is much larger than previously indicated by Ji et al. (2011), Xu et al. (2012a), Wan and Yang (2016), Lu and Sun (2017), and Xian et al. (2018). Actually, there are probably much more non-native species already present in China, but yet undetected such as the north American seed bug, Leptoglossus occidentalis, which has invaded Korea and has been intercepted by the quarantine services in Tianjin, north-eastern China (Zhu, 2010). The discrepancy between the 471 non-native insect species recorded in all ecosystems of the nearby mainland Japan (Yamanaka et al., 2015) and the total of 92 and 125 non-native insect species reported in China by Xu et al. (2012a) and Wan and Yang (2016), respectively, is also indicative of a probable under-estimation.
Unlike the clear exponential increase shown by the non-native species in Europe during the last 50 years, the invasion process seemed more fluctuating in China. According to Wan and Yang (2016), the timing of the introduction of invasive insect pests of agricultural lands and forests of China is closely related to several milestones in the national policy and economic development of the country. These authors noticed that about 1/3 of the species have invaded the country before the founding of the People's Republic of China in 1949. Then, they observed a collapse in new records during the next 30 years (1950–1979), followed by large increases during the periods after the implementation of the landmark Reform and Opening in 1978 and China's admission to the World Trade Organization in 2001. Our data showed exactly the same process when non-native insects linked to woody plants were considered separately from agricultural pests.
Europe and China exhibited distinct patterns in the sources of the invasions by non-native insects associated to woody plants. Most non-native species recorded in Europe originated from Asia, and more especially from China. In contrast, Europe was only a minor source for the insect invasions in China. These two different patterns likely reflects the dissimilar trends of the trade in ornamental plants. The 6-fold increase between 2000 and 2010 in the trade in live plants from China to Europe could be interpreted as a proxy for propagule pressure (Eschen et al., 2014), the more as the climatic similarity of the Chinese regions of plant production for the European market means that the likelihood of any arriving pests establishing in Europe is high. Most of the non-native species recorded on woody plants in China also originated mainly from Asia but came predominantly from southeastern Asia. In contrast, Xian et al. (2018) reported a more important contribution of North America (27.8%) than Asia (24.7%) when species affecting agriculture are merged with those affecting forestry. The increase in importing woody plants, especially subtropical/tropical plants, from countries of southeast Asia such as Malaysia, Thailand, and Indonesia (Hu, 2017) probably relied the invasion process for species affecting woody plants, the more as that a high proportion of species were first detected in coastal southern provinces and on Hainan island where climate conditions are quite similar to these of other countries of southeast Asia. This is coherent with the data from Qi and Lu (2018), who already listed 17 non-native species associated to woody tropical plants in the tropical provinces of China. Lu et al. (2018) also proposed that Taiwan and Hong-Kong could considered as stepping-stones for invasion to mainland China, citing the example ofa drywood termite (Incisitermes minor) brought to Hong-Kong from Japan along with timber and which then spread to Zhejiang. More globally, Wan and Yang (2016) noted that the spatial distribution of invasive insect pests in China showed a general pattern of gradually declining concentration from southeast to northwest, mainly because of increased opportunities for the introduction of invasive species by sea due to rapid development of coastal economies (Eastern China's international trade accounted for 86.5% of the national total in 2013). The presence of large sea ports as well as favorable climate conditions may also explain why a large majority of the non-native species arrived through the southern areas in Europe, too.
Although the contribution of North America was fewer in both cases, this continent supplied several highly invasive insect pests in China such as the fall webworm (Hyphantria cunea), the red turpentine beetle (Dendroctonus valens) or the Loblolly pine mealybug (Oracella acuta) (Lu and Sun, 2017). However, it appears at present that the analysis of the origin of the invasions must be more cautious. Genetic analyses of the invasion routes for some species in Europe revealed increasingly complex patterns, involving multiple introductions (e.g., the box tree moth, Cydalima perspectalis; Bras et al., 2019) and/or bridgehead processes (e.g., the Asian long-horned beetle, Anoplophora glabripennis; Javal et al., 2019; the north American seed bug, Leptoglossus occidentalis, Lesieur et al., 2019). Chu et al. (2019) also compiled the genetic analyses of invasion routes carried out for a number of non-native species related to woody plants in China; e.g., Cydia pomonella (Men et al., 2013), Frankliniella occidentalis (Duan et al., 2013), Viteus vinifoliae (Sun et al., 2009), Dendroctonus valens (Chen et al., 2013).
The predominance of Hemiptera in the non-native entomofauna of both regions is striking but coherent with the observations done in the USA and Canada where Hemiptera accounted for 41.5 and 53%, respectively, of the non-species on woody plants (Langor et al., 2009; Aukema et al., 2010). This apparently worldwide success of Hemiptera may be related to their small size, especially of their eggs, which may allow them to more easily escape detection than larger, more conspicuous insects but also to the parthenogenetic habits of high percent of them, especially aphids and scales (Langor et al., 2009). However, the limited numbers of aphids listed in the Chinese records strongly suggests that this kind of insects has not yet really been targeted for detection as non-natives in the country.
Due to the relative importance of hemipterans in both non-native entomofaunas, the sap sucker feeding guild was largely dominant. However, foliage feeders only represented a minor feeding guild in contrast to the USA where defoliators accounted for 34% of the non-native species on woody plants (Aukema et al., 2010). Xylophagous insects also presented a rather minor contribution, between 16 and 20% of the total species, although some species are important forest pests in Europe (Asian long-horned beetles, Anoplophora spp.; Javal et al., 2019) and China (Dendroctonus valens and Sirex noctilio; Lu and Sun, 2017). An important difference between China and Europe consisted in the relative importance of the fruit feeders, a much larger proportion of non-native species being observed in China. It likely relied on the presence in southern China of fruits of tropical woody plants, which are not occurring in Europe.
Guild composition in both regions confirmed that ornamental trade of plants for planting including fruit trees played a larger role than timber and wood trade in shaping the non-native entomofaunas associated to woody plants (Matošević and Pajač Živković, 2013; Eschen et al., 2015). Actually, insects arriving via plant trade can relatively easily establish because they arrive on their host, which is often planted outside (Aukema et al., 2010). There were striking differences in the range of hosts colonized by the non-native species in China and Europe. Quite a half of the non-native insect species did not switch from their original, non-native host to natives ones in Europe whereas only a minority of these species stayed on the original, non-native host in China. It may be due to the more important presence in China of native woody plants taxonomically close to the original host, especially in the southern areas for the non-native insects arriving from tropical southeastern Asia. Although fruit trees, and especially Citrus, are the most colonized hosts in both regions, tropical trees including palms were also highly colonized in China. This is consistent with the high number of records in southern provinces with subtropical and tropical climate conditions. The rather large colonization of shrubs, more important in Europe, reflects the importance of the ornamental trade. Deciduous trees were proportionally more colonized in Europe than in China, the European situation appearing similar to the one observed in Canada (Langor et al., 2009).
Essl et al. (2011) have shown that the full effects of current socioeconomic patterns on the numbers of alien species can be delayed by several decades, resulting in what has been called an “invasion debt.” Considering the fast economic growth of China, and the rapidly increasing levels of trade, tourism and transport, it is very likely that the country will face huge larger problems from invasive species in the future (Xu et al., 2012a), and has already accumulated an “invasion debt.” For instance, the near absence in Eucalyptus plantations of non-native insects from Australia, except Leptocybe invasa and Blastopsylla occidentalis, appears rather amazing considering the actual spread of eucalypt gall-makers and foliage feeders worldwide (Hurley et al., 2016). Similarly, Roques et al. (2003) listed 39 insect species, mostly seed chalcids in the genus Megastigmus (Hymenoptera), but also midges (Diptera), as potential seed-borne invaders on Chinese conifers. Their invasion success was suspected to depend on the presence of native Chinese conifers that are congeneric with the original host, or on the presence of the original host as an exotic planted in China.
The present data we compiled for Europe showed that the legacy of the European crucible hypothesized by Mattson et al. (2007) is no more reflecting the reality, when a total of 449 non-native species related to woody plants is compared to the 455 species recorded in the USA by Aukema et al. (2010) and the 419 ones recorded in Canada by Langor et al. (2009). Trends in historical accumulation of such non-native species in Europe and North America also appeared similar (Brockerhoff and Liebhold, 2017). For the future, the present increase in trade volume from China to Europe appears to be a good indicator for the most likely source of (new) non-native insects related to woody plants in the years to come (Eschen et al., 2015). Moreover, there are many new plant species and varieties in China that could be marketed in Europe (Zhao and Zhang, 2003), these new commercial markets constituting an additional invasion risk.
All datasets generated for this study are included in the article/Supplementary Material.
AR and JS prepared the database. AR did the calculations. AR, JS, M-AA-R, LR, SA, and YL contributed to the writing of the manuscript.
This work was supported by the Special Fund for Forest Scientific Research in the Public Welfare of China (Grant No. 201504304). We also acknowledge funding by INRA of the International Associated Laboratory IFOPE. The study was also part of the HOMED project, which received funding from the European Union's Horizon 2020 research and innovation programme under grant agreement no. 771271.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We greatly acknowledge Julien Papaïx and Lionel Roques (INRA Avignon) for their invaluable help in the statistical analyses.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2019.00091/full#supplementary-material
Supplementary Material S1. Database of the non-native insect species associated to woody plants established in Europe up to July 2019.
Supplementary Material S2. Database of the non-native insect species associated to woody plants established in China up to July 2019.
Supplementary Material S3. Comparison of the distribution of the regions of origin of the non-native species between Europe (black) and China (gray). Bayesian estimations of probability vectors of the multinomial distribution through a Multinomial-Dirichlet model. For the analysis, a category Europe-China has been considered to test for the symmetry of the invasion process. The posterior medians (closed circles) along with the 95% credibility intervals (vertical lines) are displayed.
Supplementary Material S4. Comparison of the relative importance of the different taxonomic orders in the non-native entomofauna between Europe (black) and China (gray)- Bayesian estimations of probability vectors of the multinomial distribution through a Multinomial-Dirichlet model. The posterior medians (closed circles) along with the 95% credibility intervals (vertical lines) are displayed.
Supplementary Material S5. Temporal changes in the taxonomic composition of the non-native entomofaunas associated to woody plants having established in Europe and China up to 2019.
Supplementary Material S6. Comparison of the feeding guilds shown by the non-native species associated to woody plants having established in Europe (black) and China (gray) -Bayesian estimations of probability vectors of the multinomial distribution through a Multinomial-Dirichlet model. The posterior medians (closed circles) along with the 95% credibility intervals (vertical lines) are displayed.
Supplementary Material S7. Temporal changes in the feeding guilds of the non-native species associated to woody plants having established in Europe and China up to 2019.
Supplementary Material S8. Comparison of the host types colonized by the non-native species associated to woody plants having established in Europe (black) and China (gray) up to 2019-Bayesian estimations of probability vectors of the multinomial distribution through a Multinomial-Dirichlet model. The posterior medians (closed circles) along with the 95% credibility intervals (vertical lines) are displayed.
Supplementary Material S9. Temporal changes in the host types colonized by the non-native species associated to woody plants having established in Europe and China up to 2019.
Aukema, J. E., McCullough, D. G., Von Holle, B., Liebhold, A. M., Britton, K., and Frankel, S. J. (2010). Historical accumulation of nonindigenous forest pests in the continental US. BioScience 60, 886–897. doi: 10.1525/bio.2010.60.11.5
Beenen, R., and Roques, A. (2010). Leaf and Seed Beetles (Coleoptera, Chrysomelidae). Chapter 8.3. In: Roques, A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 267–292. doi: 10.3897/biorisk.4.52
Bras, A., Avtzis, D. N., Kenis, M., Li, H. M., Vétek, G., Bernard, A., et al. (2019). A complex invasion story underlies the fast spread of the invasive box tree moth (Cydalima perspectalis) across Europe. J. Pest Sci. 92, 1187–1202. doi: 10.1007/s10340-019-01111-x
Brockerhoff, E. G., and Liebhold, A. M. (2017). Ecology of forest insect invasions. Biol. Inv. 19, 3141–3159. doi: 10.1007/s10530-017-1514-1
Chen, F., Luo, Y. Q., Li, J. G., Zhao, H. Q., Zong, S. X., and Shi, J. (2013). Rapid detection of red turpentine beetle (Dendroctonus valens Leconte) using nested PCR. Entomol. Am. 119, 7–13. doi: 10.1664/11-RA-010R.1
Chen, Z. Q., and Wu, S. X. (1984). Preliminary observations on Hyblaea puera Cramer. Insect Knowl. (Kunchong Zhishi). 21, 161–163.
Chu, D., Qu, W. M, and Guo, L. (2019). Invasion genetics of alien insect pests in China, Research progress and future prospects. J. Integr. Agric. 18, 748–757. doi: 10.1016/S2095-3119(17)61858-6
Cocquempot, C., and Lindelöw, Å. (2010). Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1. In: Roques, A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 193–218. doi: 10.3897/biorisk.4.56
Coeur d'acier, A., Pérez Hidalgo, N., and Petrović-Obradović, O. (2010). Aphids (Hemiptera, Aphididae). Chapter 9.2. In: Roques, A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 435–474. doi: 10.3897/biorisk.4.57
Deng, X. H., Chen, L. S., Ruan, Z. Y., Wang, M., and Fan, X. L. (2011). First record of the pest Strepsicrates semicanella (Lepidoptera, Tortricidae) of Wax-Apple from China. J. S. China Agric.Univ. 2, 55–56. doi: 10.7671/j.issn.1001-411X.2011.02.013
Denux, O., and Zagatti, P. (2010). Coleoptera families other than Cerambycidae, Curculionidae sensu lato, Chrysomelidae sensu lato and Coccinelidae. Chapter 8.5. In: Roques, A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 315–406. doi: 10.3897/biorisk.4.61
Duan, H. S., Yu, Y., Zhang, A. S., Guo, D., Tao, Y. L., and Chu, D. (2013). Genetic diversity and inferences on potential source areas for the introduced western flower thrips, Frankliniella occidentalis in Shandong, China based on mitochondrial and microsatellite markers. Flor. Entomol. 96, 963–972. doi: 10.1653/024.096.0334
Eschen, R., Holmes, T., Smith, D., Roques, A., Santini, S., and Kenis, M. (2014). Likelihood of establishment of tree pests and diseases based on their worldwide occurrence as determined by hierarchical cluster analysis. For. Ecol. Manag. 315, 103–111. doi: 10.1016/j.foreco.2013.12.021
Eschen, R., Roques, A., and Santini, A. (2015). Taxonomic dissimilarity in patterns of interception and establishment of alien arthropods, nematodes and pathogens affecting woody plants in Europe. Div. Distrib. 21, 36–45. doi: 10.1111/ddi.12267
Essl, F., Bacher, S., Blackburn, T. M., Booy, O., Brundu, G., et al. (2015). Crossing frontiers in tackling pathways of biological invasions. Bioscience 65, 769–782. doi: 10.1093/biosci/biv082
Essl, F., Dullinger, S., Rabitsch, W., Hulme, P. E., Hülber, K., Jarošík, V., et al. (2011). Socioeconomic legacy yields an invasion debt. Proc. Natl. Acad. Sci. U.S.A. 108, 203–207. doi: 10.1073/pnas.1011728108
Guénard, B., and Dunn, R. R. (2012). A checklist of the ants of China. Zootaxa 3558, 1–77. doi: 10.11646/zootaxa.3558.1.1
Hope, A. C. A. (1968). A simplified Monte Carlo significance test procedure. J. R. Statist. Soc. B 30, 582–598. doi: 10.1111/j.2517-6161.1968.tb00759.x
Hu, K. E. (2017). Study on quarantine supervision of wooden packaging of imported goods after the implementation of ISPM N°15 (Master thesis). Beijing: Beijing Forestry University.
Hurley, B., Garnas, J., Wingfield, M., Branco, M., Richardson, D., and Slippers, B. (2016). Increasing numbers and intercontinental spread of invasive insects on eucalypts. Biol. Inv.18, 921–933. doi: 10.1007/s10530-016-1081-x
Javal, M., Lombaert, E., Tsykun, T., Courtin, C., Kerdelhué, C., Prospero, S., et al. (2019). Deciphering the worldwide invasion of the Asian long-horned beetle: a recurrent invasion process from the native area together with a bridgehead effect. Mol. Ecol. 28, 951–967. doi: 10.1111/mec.15030
Ji, L. Z., Wang, Z., Wang, X. W., and An, L. L. (2011). Forest insect pest management and forest management in China, an overview. For. Env. Manag. 48, 1107–1121. doi: 10.1007/s00267-011-9697-1
Katsanevakis, S., Deriu, I., D'Amico, F., Nunes, A. N., Pelaez Sanchez, S., Crocetta, F., et al. (2015). European Alien Species Information Network (EASIN), supporting European policies and scientific research. Manag. Biol. Inv. 6, 147–157. doi: 10.3391/mbi.2015.6.2.05
Langor, D., DeHaas, L., and Foottit, R. (2009). Diversity of nonnative terrestrial arthropods on woody plants in Canada. Biol. Inv. 11, 5–19. doi: 10.1007/s10530-008-9327-x
Lesieur, V., Lombaert, E., Guillemaud, T., Courtial, B., Strong, W., Roques, A., et al. (2019). The rapid spread of Leptoglossus occidentalis in Europe: a bridgehead Invasion. J. Pest Sci. 92, 189–200. doi: 10.1007/s10340-018-0993-x
Li, Y., Zhou, X., Lai, S. C., Yin, T., Ji, Y. C., Wang, S. P., et al. (2018). First Record of Euplatypus parallelus (Coleoptera, Curculionidae) in China. Fla. Entomol. 101, 141–143. doi: 10.1653/024.101.0127
Liebhold, A. M., Brockerhoff, E. G., Garrett, L., Parke, J., and Britton, K. (2012). Live plant imports, the major pathway for the forest insect and pathogen invasions of the US. Front. Ecol. Environ. 10, 135–143. doi: 10.1890/110198
Liebhold, A. M., Yamanaka, T., Roques, A., Augustin, S., Chown, S. L., Brockerhoff, E. G., et al. (2016). Global compositional variation among native and non-native regional insect assemblages emphasizes the importance of pathways. Biol. Inv. 18, 893–905. doi: 10.1007/s10530-016-1079-4
Liebhold, A. M., Yamanaka, T., Roques, A., Augustin, S., Chown, S. L., Brockerhoff, E. G., et al. (2018). Plant diversity drives global patterns of insect invasions. Sci. Rep. 8:12095. doi: 10.1038/s41598-018-30605-4
Lopez-Vaamonde, C., Agassiz, D., Augustin, S., De Prins, J., De Prins, W., Gomboc, S., et al. (2010). Lepidoptera- Chapter 11. In Roques A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 603–668. doi: 10.3897/biorisk.4.50
Lu, J. B., Li, S. P., Wu, Y. J., and Jiang, L. (2018). Are Hong Kong and Taiwan stepping-stones for invasive species for the mainland of China? Ecol. Evol. 8, 1966–1973. doi: 10.1002/ece3.3818
Lu, M., and Sun, J. H. (2017). “Biological Invasions in forest ecosystems in China”, in Biological Invasions and its Management in China, eds. F. H. Wan and M. X. Jiang (Dordrecht: Springer Business Media B.V.), 53–66.
Matošević, D., and Pajač Živković, I. (2013). Alien phytophagous insect and mite species on woody plants in Croatia. Šumarski List 3–4, 191–205. doi: 10.31298/sl
Mattson, W. J., Niemelä, P., Millers, I., and Inguanzo, Y. (1994). Immigrant Phytophagous Insects on Woody Plants in the United States and Canada: An Annotated List. Report No. NC-169, US Department of Agriculture.
Mattson, W. J., Vanhanen, H., Veteli, T., Sivonen, S., and Niemel,ä, P. (2007). Few immigrant phytophagous insects on woody plants in Europe: Legacy of the European crucible? Biol. Inv. 9, 957–974. doi: 10.1007/s10530-007-9096-y
Men, Q. L., Chen, M. H., Zhang, Y. L., and Feng, J. N. (2013). Genetic structure and diversity of a newly invasive species, the codling moth, Cydia pomonella (L.) (Lepidoptera, Tortricidae) in China. Biol. Inv. 2, 447–458. doi: 10.1007/s10530-012-0299-5
Mifsud, D., Cocquempot, C., Mühlethaler, R., Wilson, M., and Streito, J. C. (2010). Other Hemiptera Sternorrhyncha (Aleyrodidae, Phylloxeroidea, and Psylloidea) and Hemiptera Auchenorrhyncha. Chapter 9.4. In: Roques, A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 511–552. doi: 10.3897/biorisk.4.63
Nair, K. S. S. (2001). Pest Outbreaks in Tropical Forest Plantations, is There a Greater Risk for Exotic Tree Species? Bogor Barat: Centre for International Forestry Research (CIFOR).
Pellizzari, G., and Germain, J.-F. (2010). Scales (Hemiptera, Superfamily Coccoidea). Chapter 9.3. In: Roques, A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 475–510; doi: 10.3897/biorisk.4.45
Qi, G. J., and Lu, L. H. (2018). Species characteristics and invasion status of major harmful alien insects in the tropic area of China since 2000. J. Env. Entomol. 40, 749–757.
R Core Team (2019). R Version 3.6.1 “Action of the Toes”. Vienna: The R Foundation for Statistical Computing.
Rabitsch, W. (2010). True Bugs (Hemiptera, Heteroptera). Chapter 9.1. In: Roques A et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 407–403. doi: 10.3897/biorisk.4.44
Rasplus, J.-Y., and Roques, A. (2010). Dictyoptera (Blattodea, Isoptera), Orthoptera, Phasmatodea and Dermaptera. Chapter 13.3. In: Roques, A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 807–831. doi: 10.3897/biorisk.4.68
Rasplus, J.-Y., Villemant, C., Paiva, M. R., Delvare, G., and Roques, A. (2010). Hymenoptera. Chapter 12. In: Roques, A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 669–776. doi: 10.3897/biorisk.4.55
Reynaud, P. (2010). Thrips (Thysanoptera). Chapter 13.1. In: Roques, A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 767–791. doi: 10.3897/biorisk.4.59
Roques, A. (2010). Alien forest insects in a warmer world and a globalized economy, impacts of changes in trade, tourism and climate on forest biosecurity. N. Z. J. For. Sci. 40, 77–94.
Roques, A. (2015). Drivers and pathways of forest insect invasions in Europe, can we predict the next arrivals? Atti Accad. Naz. Ital. Entomol. 53, 145–150.
Roques, A., Rasplus, J. Y., Rabistch, W., Lopez-Vaamonde, C., Kenis, M., Nentwig, W., et al. (2010). Alien terrestrial arthropods of Europe. BioRisk 4, 1–1024.
Roques, A., Sun, J. H., Auger- Rozenberg, M. A., and Hua, O. (2003). Potential invasion of China by exotic insect pests associated with tree seeds. Biodiv. Conserv.12, 2195–2210. doi: 10.1023/A:1024592716172
Sauvard, D., Branco, M., Lakatos, F., Faccoli, M., and Kirkendall, L. R. (2010). Weevils and Bark Beetles (Coleoptera, Curculionoidea). Chapter 8.2. In: Roques, A. et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 219–266. doi: 10.3897/biorisk.4.64
Schühli, G. S. E., Penteado, S. C., Barbosa, L. R., Reis Filho, W., and Iede, E. T. (2016). A review of the introduced forest pests in Brazil. Pesqui. Agropec. Bras. 51, 397–406. doi: 10.1590/S0100-204X2016000500001
Seebens, A., Blackburn, T. M., Dyer, E. E., Genovesi, P., Hulme, P. E., Jeschke, J. M., et al. (2018). The global rise in emerging alien species results from increased accessibility of new source poolS. Proc. Natl. Acad. Sci. U.S.A. 115, E2264–E2273. doi: 10.1073/pnas.1719429115
Seebens, H., Blackburn, T. M., Dyer, E. E., Genovesi, P., Hulme, P. E., Jeschke, J. M., et al. (2017). No saturation in the accumulation of alien species worldwide. Nat. Commun. 8:71314435. doi: 10.1038/ncomms14435
Simberloff, D., Martin, J. L., Genovesi, P., Maris, V., Wardle, D. A., Aronson, J., et al. (2013). Impacts of biological invasions, what's what and the way forward. Trends Ecol. Evol. 28, 58–66. doi: 10.1016/j.tree.2012.07.013
Skuhravá, M., Martinez, M., and Roques, A. (2010). Diptera. Chapter 10. RoquesA et al. (Eds) Alien terrestrial arthropods of Europe. BioRisk 4, 553–602 doi: 10.3897/biorisk.4.53
Sun, Q. H., Chen, Y. C., Wang, H. B., Downie, D. A., and Zhai, H. (2009). Origin and genetic diversity of grape phylloxera in China. Acta Entomol. Sin. 8, 885–894. Available online at: http://www.insect.org.cn/CN/Y2009/V52/I8/885
Tao, C. C. C. (1999). List of Aphidoidea (Homoptera) of China. Taichung city: Taiwan Agricultural Research Institute.
Trencheva, K., Trenchev, G., Tomov, R., and Wu, S. A. (2010). Non-indigenous scale insects on ornamental plants in Bulgaria and China: a survey. Entomol. Hell. 19, 114–123. doi: 10.12681/eh.11578
Vilà, M., Basnou, C., Pyšek, P., Josefsson, M., Genovesi, P., Gollasch, S., et al. (2010). How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 8, 135–144. doi: 10.1890/080083
Wan, F. H., and Jiang, M. X. (2017). Biological Invasions and Its Management in China, Vol. 1 (Dordrecht: Springer Science + Business Media B.V.).
Wan, F. H., and Yang, N. W. (2016). Invasion and management of agricultural alien insects in China. Annu. Rev. Entomol. 61, 77–98 doi: 10.1146/annurev-ento-010715-023916
Wang, S., and Wu, S. A. (2010). A new record species of the Genus Pseudococcus Westwood (Hemiptera: Pseudococcidae) from China. Acta Zootax. Sinica 35, 240–246.
Xian, X. Q., Wang, R., Guo, J. Y., Liu, W. X., Zhang, G. F., et al. (2018). Analysis of new invasive alien species in China's agricultural and forestry ecosystems in recent 20 years. Plant Prot. 44, 168–175. doi: 10.16688/j.zwbh.2018332
Xu, H., Chen, K., Ouyang, Z. Y., Pan, X. B., and Zhu, S. F. (2012a). Threats of invasive species for China caused by expanding international trade. Environ. Sci. Tech. 46, 7063–7064. doi: 10.1021/es301996x
Xu, H., Qiang, S., Genovesi, P., Ding, H., Wu, J., Meng, L., et al. (2012b). An inventory of invasive alien species in China. NeoBiota 15, 1–26. doi: 10.3897/neobiota.15.3575
Yamanaka, T., Morimoto, N., Nishida, G. M., Kiritani, K., Moriya, S., and Liebhold, A. M. (2015). Comparison of insect invasions in North America, Japan and their islands. Biol. Inv. 17, 3049–3061. doi: 10.1007/s10530-015-0935-y
Yen, A. L., Burckhardt, D., and Cen, Y.-J. (2013). The occurrence of the Australian Blastopsylla occidentalis Taylor (Hemiptera, Psylloidea) in China. Dongwu Fenlei Xuebao 38, 436–439.
Zhao, L., and Zhang, D. (2003). Ornamental plant resources from China. Acta Horticult. 620, 365–375. doi: 10.17660/ActaHortic.2003.620.46
Keywords: non-native, insect, woody plants, China, Europe
Citation: Roques A, Shi J, Auger-Rozenberg M-A, Ren L, Augustin S and Luo Y-q (2020) Are Invasive Patterns of Non-native Insects Related to Woody Plants Differing Between Europe and China? Front. For. Glob. Change 2:91. doi: 10.3389/ffgc.2019.00091
Received: 15 August 2019; Accepted: 19 December 2019;
Published: 15 January 2020.
Edited by:
Alberto Santini, Italian National Research Council (IPSP-CNR), ItalyReviewed by:
René Eschen, Cabi, SwitzerlandCopyright © 2020 Roques, Shi, Auger-Rozenberg, Ren, Augustin and Luo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alain Roques, YWxhaW4ucm9xdWVzQGlucmEuZnI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.