
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ethol., 01 April 2025
Sec. Adaptation and Evolution
Volume 4 - 2025 | https://doi.org/10.3389/fetho.2025.1540458
This article is part of the Research TopicFarm Animal EthologyView all articles
 Thiago Bernardino1,2*
Thiago Bernardino1,2* Leandro Sabei2
Leandro Sabei2 Marisol Parada Sarmiento2,3
Marisol Parada Sarmiento2,3 Denis Sato2
Denis Sato2 Sharacely de Souza Farias2
Sharacely de Souza Farias2 Tauana Maria Carlos Guimarães de Paula2
Tauana Maria Carlos Guimarães de Paula2 Gabriela Rezende Marques4
Gabriela Rezende Marques4 Adroaldo José Zanella2
Adroaldo José Zanella2Boars housed in commercial conditions are often housed in pens or in individual crates, and are subjected to low-stimulus environments, which may affect the expression of normal species-specific behaviors and compromise its health and welfare. Breeding boars’ behaviors has been poorly reported in the scientific literature, as well as their physiology and its circadian rhythm of cortisol. Moreover, an environmental enrichment for breeding boars, as far as we know, has not been studied or reported in literature, lacking its impact on how it can improve their welfare. Thus, we aimed to evaluate the behavior and salivary cortisol physiology of breeding boars housed in three different conditions: crates, pens and enriched pens. For the enriched pens the boars were brushed, they were showering and they received hay, twice daily. All animals were video recorded, and we collected saliva samples in the morning and afternoon to measure the circadian rhythm of cortisol. The boars housed in crates showed more (5.23 seconds) abnormal behaviors (head weaving, biting bars, licking floor, sham-chewing, drinker manipulation, and licking empty trough) than boars housed in pens (2.47 seconds) and in enriched pens (2.93 seconds). In addition, the boars housed in the enriched pens did have significant differences in salivary cortisol through the day (morning and afternoon assessment), different from boars from other treatments. We demonstrated that breeding boars that received environmental enrichment showed indicators of better welfare (behavior and salivary cortisol ratio), suggesting that this strategy should be considered as a protocol to improve their welfare.
Boars housed in commercial conditions are often housed in pens or in individual crates (Knox, 2016), and are subjected to low-stimulus environments, which may affect the expression of normal species-specific behaviors and compromise its health (Teles et al., 2017). The behavior of boars has not been reported in literature as detailed as the behavior of sows, piglets, or finishers. It is known that the temperature, photoperiod, and nutrition can affect the reproductive performance of boars; however, there is limited information about semen collection frequency and housing condition in previous studies (Flowers, 2015). Generally speaking, heat stress for boars can compromise semen features which may affect fertility (Einarsson et al., 2008), even when exposed for a shor period of 72 hours (see review from Flowers, 2022). Moreover, heat stress affect testicular function, which may disrupt the blood-testes barrier integrity, changing the production of important proteins, as well as increasing DNA damage (Flowers, 2022). Semen from boars housed in a facility with a controlled environment temperature showed higher motility and more sperms cells with normal acrosomes, when contrasted with boars without proper temperature control environment (Corcuera et al., 2002). In addition, in this same research, the authors provided straw bedding, which was beneficial in increasing the same sperm parameters during winter season. To our knowledge, there is limited information in scientific literature about the influence of housing conditions in the welfare and physiology of boars, while this subject has been extensively reported in pregnant sows (Schwarz et al., 2021).
Crates are a well-reported source of chronic stress for adult sows, affecting cortisol concentrations (Hemsworth, 2018), which has been used for monitoris HPA axis function in pigs in the past decades (see review by Wolf et al., 2020). Moreover, this type of housing is commonly used to house breeding boars (Knox, 2016). Crates can compromise the ability of the animals to seek social contact, to explore, and to show natural behavior such as rooting, separating clean and dirty areas, exploratory behaviors, among others (Broom, 1986). Moreover, crates with a concrete floor provide an inadequate environment, since boars can only interact with food, metal bars, and with their own feces (Petak et al., 2010). In semi-natural conditions, boars can spend 27% of their daytime grazing (Stolba and Wood-Gush, 1989), indicating a relevant behavior for adult males. A valuable alternative that should be provided in this scenario is environmental enrichment. Environmental enrichment is one used of the tools to improve the welfare of pigs. In sows, the use of environmental enrichment has been studied for its impact to their own physiology and welfare, as well as for its offspring (Tatemoto et al., 2019). Tactile stimulation are powerful mediators of positive states in several species, and it has been reported enjoyable for calves (Horvath et al., 2020), it can reduce stress or frustration for animals housed in poor environments (DeVries et al., 2007), and can be used as environmental enrichment (Wilson et al., 2002). However, the use of environmental enrichment for boars, as far as we know, has been poorly investigated. Teles et al. (2017) reported that crated boars with a coffee husk bed changed their resting behavior and showed higher salivary cortisol concentration than boars kept without coffee husk bedding material. Likewise, using the same bedding material for boars housed in pens showed a reduction in subjective motility (Teles et al., 2016). There are different types of environmental enrichment, which can affect differently the animals, activating their sense of sight, sense of taste, sense of smell, sense of hearing, and others (Kittawornra and Zimmerman, 2010). There are reports showing that pig has shown preferences for toys that can be chewed, such as ropes, chains, chopped straws, which stimulates their exploratory behavior (Tavares et al., 2023). However, the scientific community has not dedicated attention to this issue, especially for breeding boars (Teles et al., 2017). Thus, we decided to use a combination of different environmental enrichment for boars; tactile and sensorial stimulus, and social since they could interact with the boars housed in pens beside.
Another possibility to improve the social environment of breeding boars is keeping the animals in groups. Kunavongkrit et al. (2005) reported that it is difficult to house boars in groups, unless electronic feeding is provided. Another issue is regarding sodomy and aggressive behaviors, which could compromise performance and welfare of the animals (Kunavongkrit et al., 2005). A study carried out by Hemsworth and Beilharz (1979) showed that the social restriction during rearing environment is important for the boar sexual behavior before mating. The authors found that boar reared individually showed less courting behavior and achieved fewer copulations. In addition, a previous study evaluated group housed boars, with 4-5 animals per pen (Cordoba-Dominguez et al., 1991). The boars were reared together from an early age, and the authors reported few undesirable behaviors within this management scenario. Previous research showed that group housing for boars is a good tool to improve social interactions and welfare (Cordoba-Dominguez et al., 1991). It has been hypothesized that more opportunities for social behavior, such as snout-snout contact with another boar, would be a good alternative to improve welfare (Corcuera et al., 2002), avoiding solid walls. The European legislation (Council of the European Union, Directive 2008/120/EC, December, 2009) stands that breeding boars must have the opportunity to hear, smell and see other pigs, which does not allow boars housed in solid walls pens.
One of the consequences for pigs that experience individual or inappropriate housing is the possible activation of different stress mechanisms. Salivary cortisol sampling can provide key information about the hypothalamic-pituitary-adrenal (HPA) axis activity in pigs as it represents the free, unbound fraction, hence biologically active, glucocorticoid, if samples are taken in the morning and afternoon (Zanella et al., 1998). Moreover, salivary cortisol is a biomarker that offer real time information without the drawbacks of serum cortisol, since the sampling may be a several stressor, which could confound the HPA axis assessment (Giergiel et al., 2021). High cortisol levels may compromise reproductive efficiency and fertility in gilts (Tsuma et al., 1996). However, to the best of our knowledge, little information is available about salivary cortisol levels in boars.
The aim of the present study was to evaluate the impact of three different housing conditions for boars on their behavior and salivary cortisol concentration.
This study was approved by the Committee on Ethics in Animal Use (CEUA) of the School of Veterinary Medicine and Animal Science (FMVZ), University of São Paulo (USP) under protocol number 3612010616.
In the beginning of the study, we used 30 boars, 10 for each treatment. However, for healthy reasons, 3 boars were excluded from this study. Thus, 27 boars (F1 large white x landrace) 10 months old, were fed twice daily, 7am and 1pm, with 2.8kg of concentrate per day, and had ad libitum access to a nipple drinker. The diet consisted in corn, soybean meal and mineral premix (3300 kcal per kg, calculated value). Prior to treatment allocation, all animals were housed in individual pens, regardless of the treatment, measuring 3.85 meters x 1.2 meters. Afterwards, animals were allocated in one of the assigned three housing conditions, distributed in the treatments based their seminal quality, measured using Computer Assisted Sperm Analysis (CASA) and sperm morphology. Boars with good, medium, and low semen quality were evenly allocated to groups, which were subsequently randomly assigned to the treatments.
The three housing conditions consisted in a) boars kept in the same pens (n = 9), b) identical pens with an enrichment protocol (n = 9), and c) gestation crates (n = 9). In the enriched pens, environmental enrichment was provided twice daily, one hour after feeding (8am and 2pm). The enrichment included brushing the animals for two minutes using a broom (45.7 cm in length, with 10 cm hard nylon bristles, showering the animals with room temperature water for 30 seconds (rubber hose, and supplying 1,000 grams of Tifton 85 hay per day as rooting material. Personnel familiar to the animals, who had no negative interactions with them, conducted all enrichment activities.
All additional information regarding treatments, management routine, semen sampling, treatment allocation, and testis physiology can be found in Bernardino et al. (2022).
Boar behavior was recorded with cameras (Intelbras VHD 1220 B – G4), in full high-definition quality, and stored in a hard drive disk until further analyses. In order to cover all barn, we used 2 cameras per barn, in opposite walls, totaling 8 cameras. We analyzed six minutes each hour, from 7am until 5pm. The behavior was analyzed by four different trained observers, at week 0 (prior to the treatment allocation), week 4 (30 days after treatment assignment), and week 8 (60 days after treatment assignment). The behavioral observation was performed by a combination of scan sampling, followed by a focal animal and continuous recording (Martin and Bateson, 2007), with each animal observed for 2 uninterrupted minutes. Each boar was observed three times in each hour of the day, totaling 54 minutes per day of observation. Six trained observers had access to the videos and, in a randomized way, gathered data from the studied animals. All collection periods consisted in two consecutive days to avoid interference of possible events in the behavioral data (e.g. on days D-1, D0, D29, D30, D59, and D60).
An ethogram was developed, based on the ethograms validated by Zonderland et al. (2004), Bernardino et al. (2021), and Tatemoto et al. (2019). Table 1 has the behavioral variables used in the behavioral observation protocol. We obtained the variable abnormal behavior by the sum of the following behaviors: head weaving, biting bars, licking floor, sham-chewing, drinker manipulation, and licking empty trough.
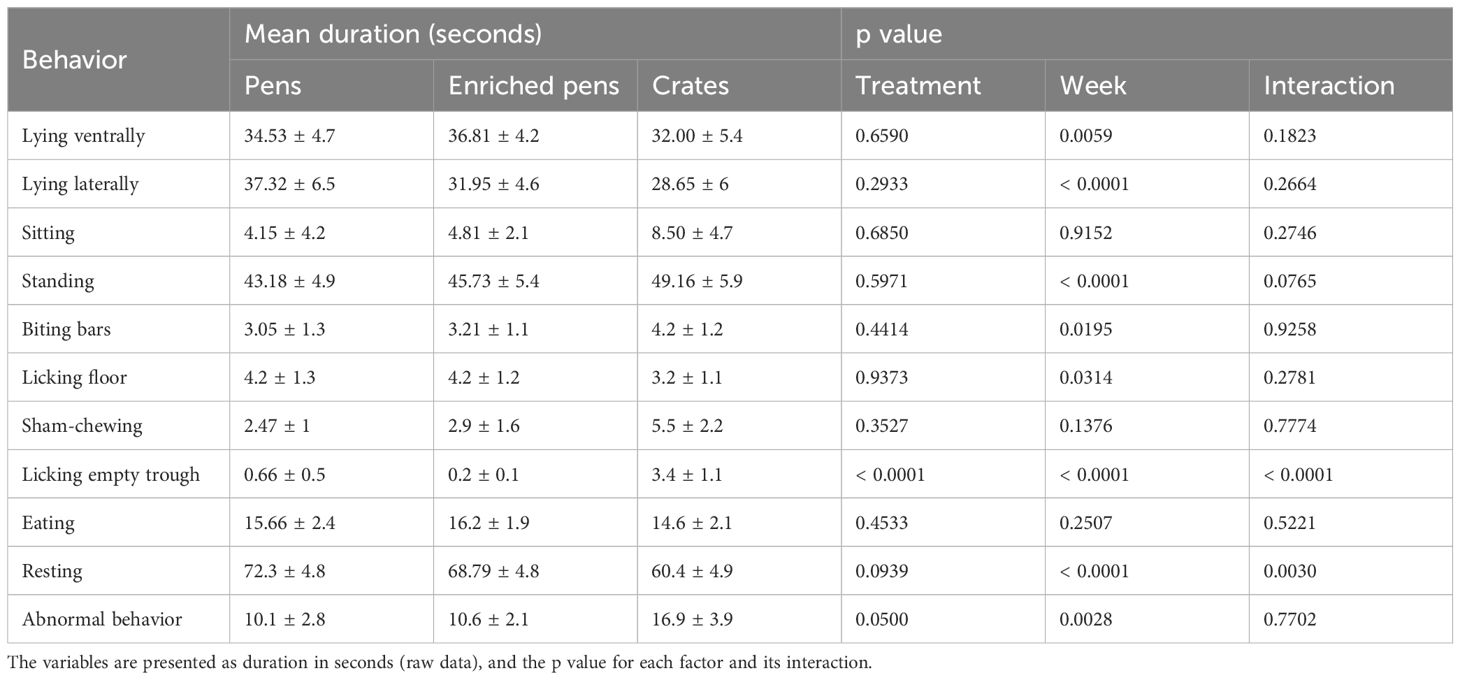
Table 1. Mean duration, standard error mean, and probability of the analyzed behaviors of boars housed in three different conditions.
We followed the methodology of Siegford et al. (2008) and Tatemoto et al. (2019), which consisted in presenting hydrophilic cotton, two roller-shaped units (Cremer®, number 1, non-sterile, starch and chloride free), tied to a dental floss with long tips (Hillo, 500m, no flavor) and offered to the animals. The boar chewed the cotton until it was saturated with saliva. The first sample was discarded; we repeated the protocol to collect only recently produced saliva. After the second sample was collected, it was placed in a 15 ml conic tube (Kasvi®), identified with the animal’s number and time of collection. Subsequently, the tube was packed in an ice cube box until the end of the collection, and then frozen at -20°C until processing. The thawing was performed in a container with ice. After the complete thawing of the sample, it was centrifuged for two minutes at 1000 x g (Celm Combate), and then the supernatant was aliquoted into 1.5 ml micro tubes and again frozen at -20°C until analysis. This process assists in the removal of mucins and other components that may interfere with the analysis protocol. We used a cortisol enzyme immunoassay, without extraction, to measure the cortisol concentration (Palme and Möstl, 1997). All samples were analyzed in duplicates. The sensitivity of the EIA was 0.2 pg/well. To the best of our knowledge, this is the first manuscript to quantify salivary cortisol in breeding boars. Thus, our references were mainly from sows.
Saliva was collected before feeding time (6:30-7:00am) and in the afternoon (6:00-6:30pm) during the following periods of behavioral observations. Samples from two consecutive days were pooled for AM and PM collection times. In addition, we calculated the AM/PM ratio in salivary cortisol samples. To determine the AM/PM ratio, we used the salivary cortisol concentration from the morning and divided it for the cortisol concentration found in the afternoon. This methodology was used in humans for distinct approaches (Han et al., 2019).
All the data were analyzed with the package Statistical Analysis System 9.4 (SAS Inst., Inc., Cary, NC). Initially, the data were checked for the presence of discrepant information (outliers) and we verified the residual normality, through the Shapiro-Wilk test. After those approaches, the data were analyzed by ANOVA with PROC GLIMMIX of SAS using a treatment as principal effect over the time and a block (seminal quality) as a random effect. In addition, the command REPEATED was added to the model, to analyze the time effect. Thus, we had the effect of treatment, the effect of time, and the interaction between these two factors.
For all analyses, among the 15 different covariance structures were tested and the one that best fits the statistical model was chosen, based on the lower value of the Akaike Index Correction Criterion (AICC) (Wang and Goonewardene, 2004). When necessary, for a post-hoc test, the Tukey’s test was used.
We used this approach for the behavior variables and AM/PM ratio. For the variable salivary cortisol concentration, we added the time of collection in the model (morning and afternoon). Thus, for this variable, the boar was considered as plot, the week as sub-plot, and the time of collection was classified as a sub-sub plot.
The means and the standard error mean (SEM) were obtained from raw data. For all statistical analyses performed, a 5% level of significance was adopted.
We found several differences among treatments on behavior, which are summarized in Table 1. Most differences found were caused by time, however, there were several interactions between treatment and time.
For the behavior Licking Empty Trough, we found an interaction between treatment and time (p value = < 0.0001). We found that the boars housed in crates showed this behavior for longer periods at week 8, after treatment allocation. In addition, we did not identify differences between the treatment pens and enriched pens (Figure 1).
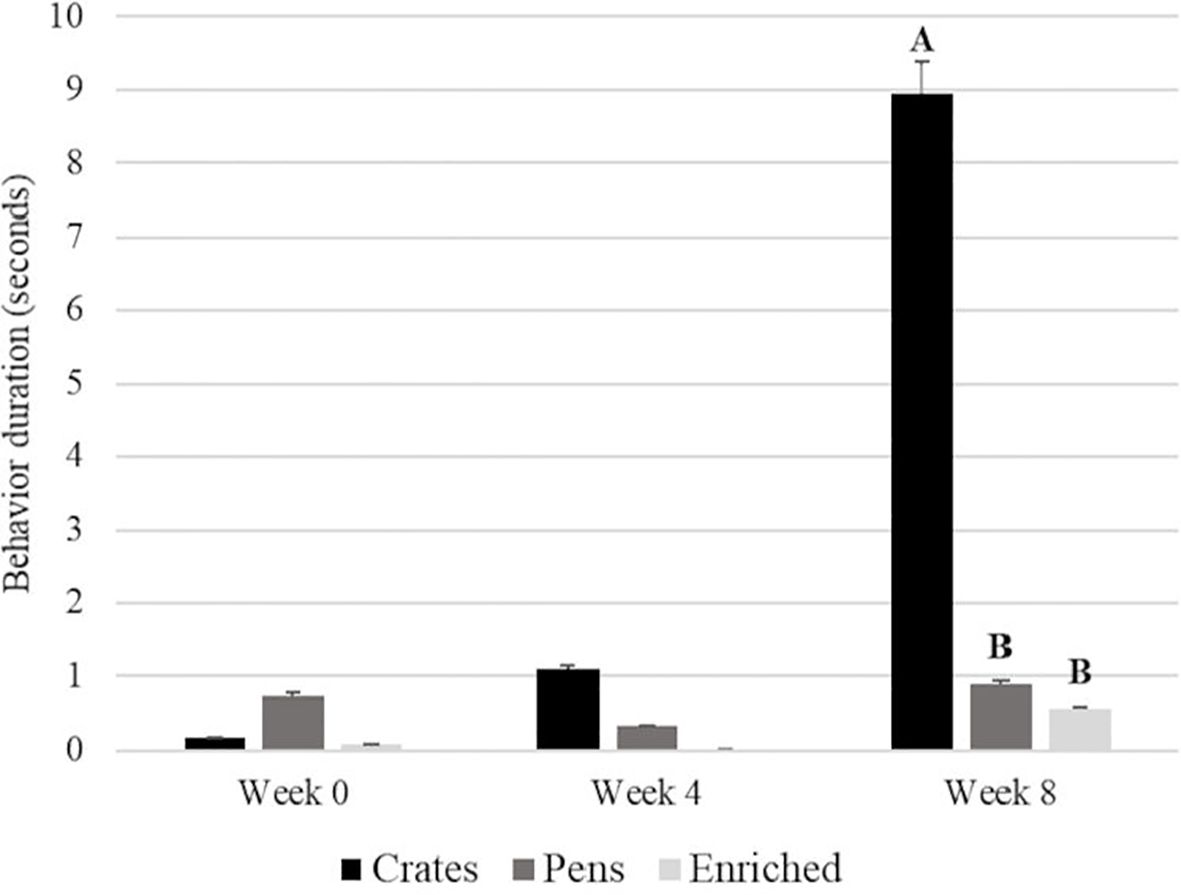
Figure 1. The treatment and time (week) interaction for licking empty trough behavior for boars housed in three different conditions. Different letters mean difference among treatments (p <0.05), and no letters mean no difference.
We found a treatment and time effect for abnormal behavior, but no interaction. The boars housed in crates showed longer abnormal behavior when compared with boars housed in pens and enriched pens (Figure 2).
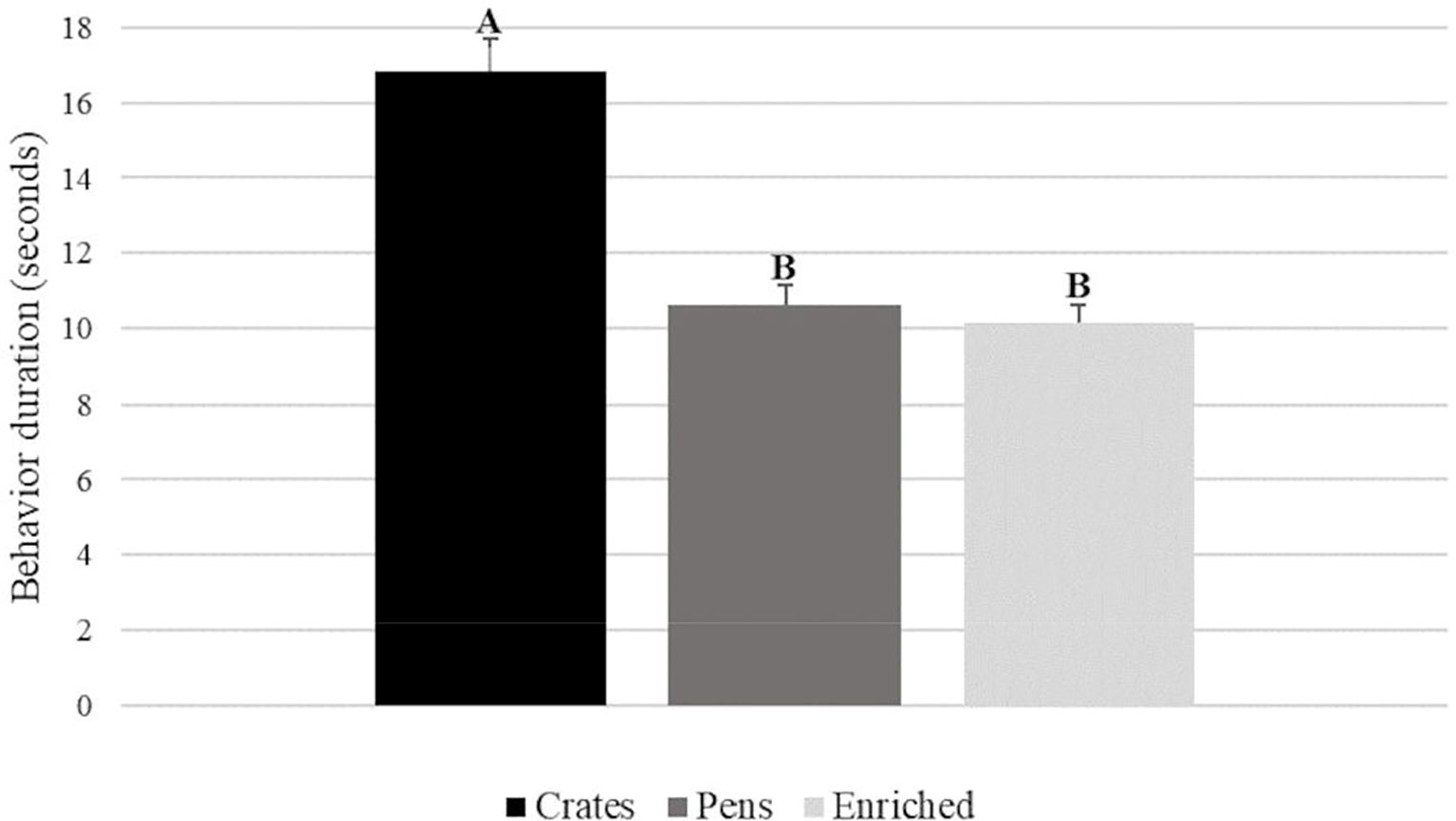
Figure 2. Abnormal behavior duration of boars housed in three different conditions, during 8 weeks interval. Equal letters mean no difference among treatments.
The principal effects of our model in salivary cortisol concentration are presented in Table 2. We identified a week, time, and interaction of treatment with week and time effects. The cortisol concentration data is summarized in Figure 3.
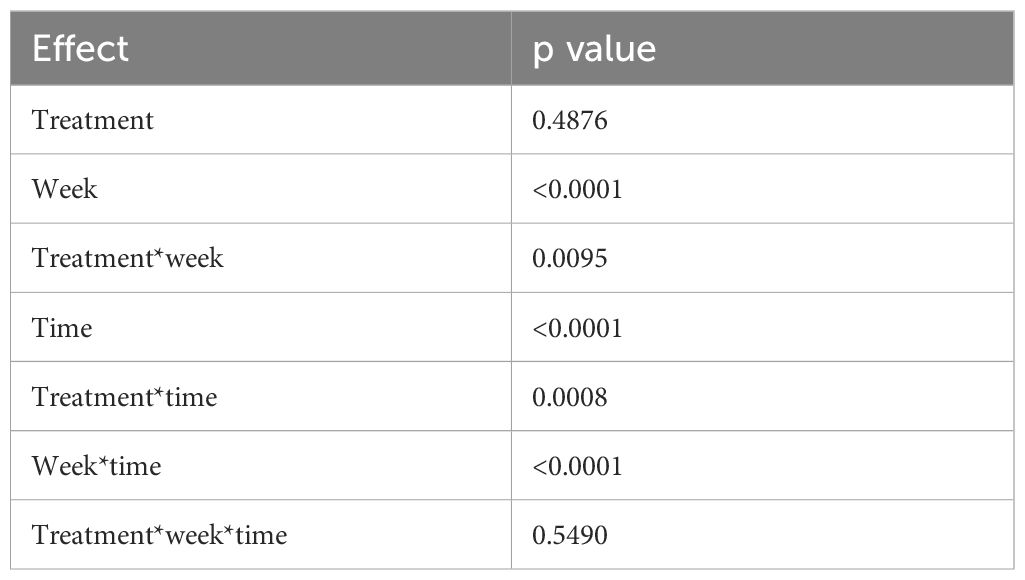
Table 2. p value of principal effects of the treatment, week, time, and the interaction among factors.
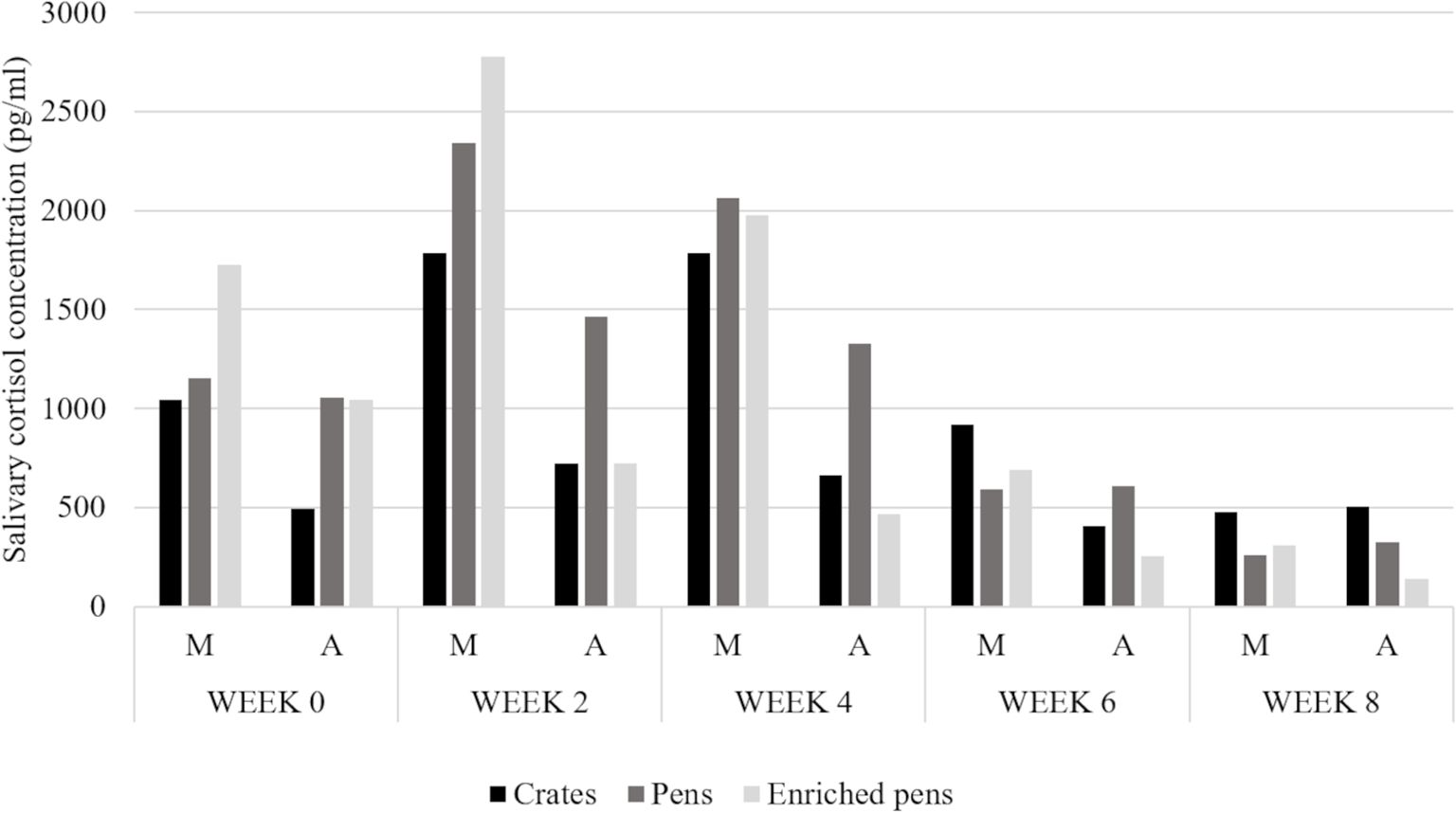
Figure 3. Salivary cortisol concentration of boars kept in three different housing conditions, over a period of 8 weeks, morning (M), and afternoon (A) sampling.
After the Tukey-Kramer test, for the interaction between treatment and week, we found that the salivary cortisol concentration did not differ among treatments at week 8, irrespective of the time of collection (morning or afternoon). For the interaction between week and time, we found that only at week 8, there was no difference among time. In other words, the results from morning and afternoon were not different. For all other weeks, the data from morning samples were different from afternoon, as expected. As we did not find a treatment or a treatment, week, and time interaction, these results will not be detailed discussed.
According to our methodology, the AM/PM ratio showed intriguing results (Figure 4). The AM/PM salivary cortisol concentration was not different in the sampling before treatment allocation (p = 0.3835) and in the week 6 (p = 0.1252). Boars housed in enriched pens showed a higher salivary cortisol concentration ratio in different moments, week 2 (p = 0.05), week 4 (p = 0.0486) and week 8 (p = 0.0322).
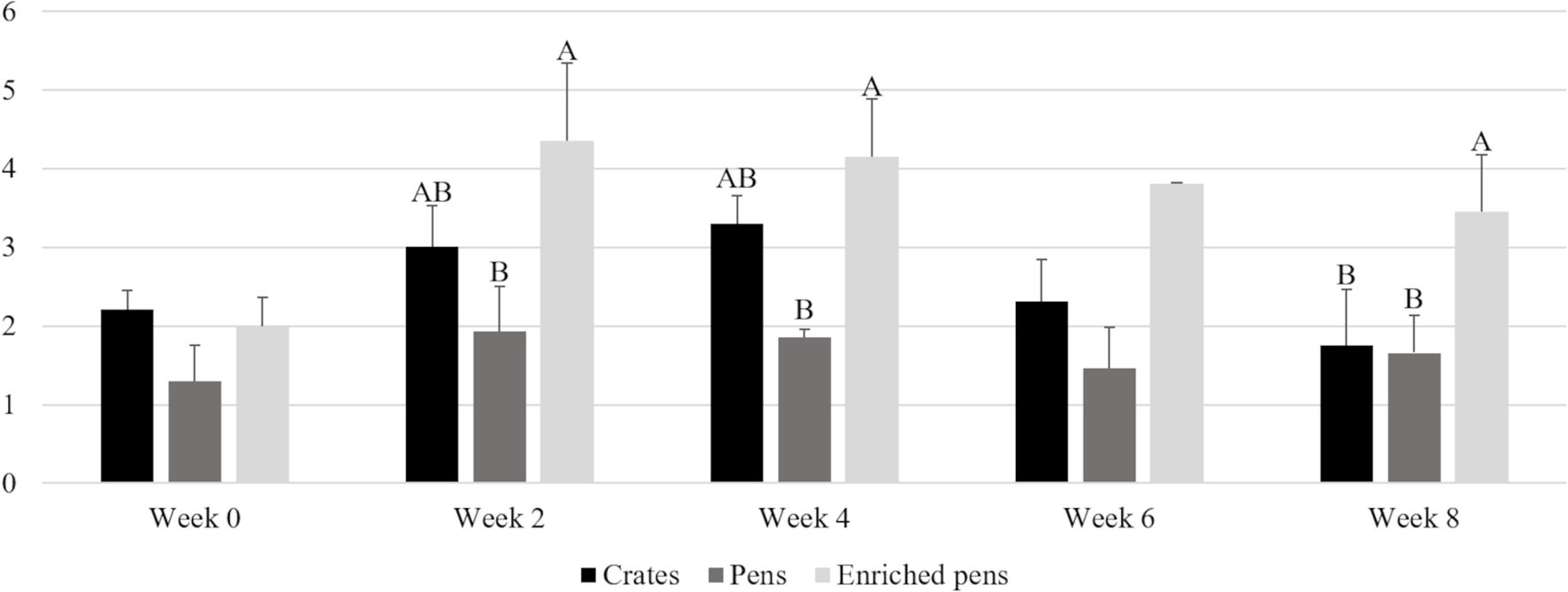
Figure 4. AM/PM ratio of salivary cortisol concentration in boars kept in three different housing conditions. Different letters mean p < 0.05.
We observed that boars housed in crates licked the empty trough longer at 8 weeks after treatment allocation (see Figure 1), when compared to other treatments. Moreover, when we created the combined variable abnormal behavior and contrasted it, we identified that during the completely experimental period, boars housed in crates performed more abnormal behavior than penned and enriched penned boars. The performance of abnormal behaviors is reported as a consequence of the inability to carry natural behaviors, associated with frustration, feed restriction, among others (D’Eath et al., 2009; MaChado et al., 2017; Mason, 1991; Zhang et al., 2017). Moreover, behavior is considered a welfare indicator (Mason, 1991; Zhang et al., 2017) which could reflect the psychological (Tatemoto et al., 2019) and afective states (Mellor et al., 2020). Mellor et al. (2020) mentioned that some unpleased experiences may not be attenuated by changes in behavior and physiology, and can greatly compromise the welfare state. Therefore, we can consider that the boars housed in crates manifested a behavioral pattern, which indicated compromised welfare. This was observed during the entire studied period, as shown in Figure 2. Oral behaviors are very important for pigs thus this result is concerning regarding housing conditions for boars. The most farms in the US and in Brazil, major pork producers, uses crate to house boars, which compromises their welfare, their physiology and their testis health (Bernardino et al., 2022). Thus, we recommend avoiding housing boars in crates, since it compromises boars’ welfare and health.
A healthy cortisol circadian pattern for pigs, as diurnal animals, consist in higher concentration in the morning, decreasing during the course of the day (Koopmans et al., 2005; Ruis et al., 1997). We observed this pattern for the studied period, except for the week 8. We believe that maybe, at that time for crated boars, the HPA axis was near reaching the end of resistance or the beginning of exhaustion phase, according to the general adaption syndrome (Selye, 1950). Few manuscripts explore for how long adult boars can reach the exhaustion phase of their HPA axis. Since we did not measured salivary cortisol concentration to evaluate this syndrome, this is just speculation regarding the possible mechanisms for our results. Crated and penned boars did not show great differences between morning and afternoon salivary cortisol concentration. Only enriched penned boars showed this expected pattern for cortisol, higher levels in morning comparing with afternoon. In addition, the last week of the experiment showed lower environmental temperature, which also influenced the superficial scrotum mean temperature (Bernardino et al., 2022). All animals from all treatments showed higher salivary cortisol concentrations in the morning compared with the afternoon sampling.
We studied breeding males, which is a topic with few reports regarding cortisol patterns for this category. It was already demonstrated that males showed a different hair cortisol concentration than females (Bergamin et al., 2019). The AM/PM ratio is a reflection of a proper function of the HPA axis, associated with health outcomes (Adam and Kumari, 2009; Stone et al., 2001; Šupe-Domić et al., 2016). In the AM/PM ratio, as observed in Figure 4, boars from enriched pens showed a higher AM/PM ratio in the week 2, 4, and 8. These results indicate that these animals had a well-defined circadian rhythm, since they showed higher levels of cortisol concentration in morning sampling and lower concentration in the afternoon. One of the possible explanations for this finding is because every day in the morning and afternoon, we provided the enrichment protocol for treated boars. Brushing, short baths, and hay could play an important role in the organization of the circadian rhythm, added to higher levels of a positive human-animal interaction during the enrichment protocol. Also, the differences observed in the behavior could be a consequence of this organized circadian rhythm, since the cortisol rhythm is correlated with others hormones rhythms and behaviors, such as insulin and glucose, which can influence exploratory behavior for foraging (Han et al., 2019; Koopmans et al., 2005; Ruis et al., 1997; Stone et al., 2001).
Improving housing conditions for breeding boars, has the potential to enhance their welfare and physiology, potentially affecting the subsequent generations. Previous studies within our research group showed compelling evidence indicating that the housing conditions for boars can modulate survival, aggressive behavior and nociception responsiveness in their offspring (Sabei et al., 2023a), as well as their emotional states (Sabei et al., 2023b). Therefore, we need to ensure that these boars are housed in a positive environment, with positive consequences to their welfare, physiology and for their offspring.
Research involving boars in commercial settings is highly constrained due to their intense and specific management routine. Consequently, this study was conducted under controlled research conditions, which introduces certain limitations. A key limitation is the age of the boars, as older animals may respond differently to the treatments. We hypothesized that, in a long-term context, older boars might exhibit significantly different patterns in terms of abnormal oral behaviors, inactivity, and salivary cortisol concentrations. Nonetheless, even a relatively short housing period of 8 weeks in contrasting environments can markedly influence their behavior, circadian cortisol rhythm, reproductive physiology (Bernardino et al., 2022), and the characteristics of their offspring (Sabei et al., 2023a).
When combining, the behavioral and salivary cortisol concentration results showed how the treatments influenced these parameters. Boars housed in crates showed more abnormal behaviors than penned or enriched penned boars. Boars housed in the enriched pens did have significant differences in salivary cortisol through the day (morning and afternoon assessment). Environmental enrichment with brushing, short bath, and hay can affect the behavioral and the cortisol circadian rhythm in breeding boars, and should be considered as a protocol to improve their welfare.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
This study was approved by the Committee on Ethics in Animal Use (CEUA) of the School of Veterinary Medicine and Animal Science (FMVZ), University of São Paulo (USP) under protocol number 3612010616. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent wasobtained from the owners for the participation of their animals in this study.
TB: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. LS: Investigation, Writing – original draft, Writing – review & editing. MPS: Investigation, Writing – original draft, Writing – review & editing. DS: Writing – original draft, Writing – review & editing, Investigation. SSF: Writing – original draft, Writing – review & editing, Investigation. TMCGP: Writing – original draft, Writing – review & editing, Investigation. GRM: Writing – original draft, Writing – review & editing, Investigation. AJZ: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. We are thankful to São Paulo Research Foundation (FAPESP) to the grant (Process number 2017/05604-2) to TB., and for the regular project grant (Process number 2018/01082-4). LS scholarship was given by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brazil (CAPES) (CAPES / PROEX Funding Code 88887.626403/2021-00). MPS scholarship was given in the framework of the Project “Demetra” (Dipartimenti di Eccellenza 2018e2022, CUP_C46C18000530001), funded by the Italian Ministry for Education, University and Research. AJZ received a grant number 16457/2021-3; Chamada CNPq Nº4/2021 - Bolsa de Produtividade em Pesquisa - PQ.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adam E. K., Kumari M. (2009). Assessing salivary cortisol in large-scale, epidemiological research. Psychoneuroendocrinology 34, 1423–1436. doi: 10.1016/j.psyneuen.2009.06.011
Bergamin C., Comin A., Corazzin M., Faustini M., Peric T., Scollo A., et al. (2019). Cortisol, DHEA, and sexual steroid concentrations in fattening pigs’ hair. Animals 9, 345. doi: 10.3390/ani9060345
Bernardino T., Carvalho C. P. T., Batissaco L., Celeghini E. C. C., Zanella A. J. (2022). Poor welfare compromises testicle physiology in breeding boars. PloS One 17, e0268944. doi: 10.1371/journal.pone.0268944
Bernardino T., Tatemoto P., de Moraes J. E., Morrone B., Zanella A. J. (2021). High fiber diet reduces stereotypic behavior of gilts but does not affect offspring performance. Appl. Anim. Behav. Sci. 243, 105433. doi: 10.1016/j.applanim.2021.105433
Broom D. M. (1986). Indicators of poor welfare. Br. Vet. J. 142, 524–526. doi: 10.1016/0007-1935(86)90109-0
Corcuera B. D., Hernández-Gil R., De Alba Romero C., Martín Rillo S. (2002). Relationship of environment temperature and boar facilities with seminal quality. Livest. Prod. Sci. 74, 55–62. doi: 10.1016/S0301-6226(01)00286-X
Cordoba-Dominguez J., Dunne J. H., Cliff A. H., Macpherson O., Menaya C. M., Vidal J. M. A., et al. (1991). “Evaluation of the behaviour of mature boars housed in groups of four,” in Proceedings of the British Society of Animal Production, 65–65. doi: 10.1017/s0308229600020158
Council of the European Union (2009). Official Journal of the Council of the European Union. Off. J. Eur. Union. 2, 9.
D’Eath R. B., Tolkamp B. J., Kyriazakis I., Lawrence A. B. (2009). ‘Freedom from hunger’ and preventing obesity: the animal welfare implications of reducing food quantity or quality. Anim. Behav. 77, 275–288. doi: 10.1016/j.anbehav.2008.10.028
DeVries T. J., Vankova M., Veira D. M., von Keyserlingk M. A. G. (2007). Short communication: usage of mechanical brushes by lactating dairy cows. J. Dairy Sci. 90, 2241–2245. doi: 10.3168/jds.2006-648
Einarsson S., Brandt Y., Lundeheim N., Madej A. (2008). Stress and its influence on reproduction in pigs: a review. Acta Vet. Scand. 50, 48. doi: 10.1186/1751-0147-50-48
Flowers W. L. (2015). Factors affecting the efficient production of boar sperm. Reprod. Domest. Anim. 50, 25–30. doi: 10.1111/rda.12529
Flowers W. L. (2022). Factors affecting the production of quality ejaculates from boars. Ani. Repr. Scie. 246, 106840. doi: 10.1016/j.anireprosci.2021.106840
Giergiel M., Olejnik M., Jablonski A., Posyniak A. (2021). The markers of stress in swine oral fluid. J. Vet. Res. 65, 487–495. doi: 10.2478/jvetres-2021-0065
Han Y., Han L., Dong M. M., Sun Q. C., Zhang Z. F., Ding K., et al. (2019). Preoperative salivary cortisol AM/PM ratio predicts early postoperative cognitive dysfunction after noncardiac surgery in elderly patients. Anesth. Analg. 128, 349–357. doi: 10.1213/ANE.0000000000003740
Hemsworth P. H. (2018). Key determinants of pig welfare: Implications of animal management and housing design on livestock welfare. Anim. Prod. Sci. 58, 1375–1386. doi: 10.1071/AN17897
Hemsworth P. H., Beilharz R. G. (1979). The influence of restricted physical contact with pigs during rearing on the sexual behaviour of the male domestic pig. Anim. Prod. 29, 311–314. doi: 10.1017/S0003356100023564
Horvath K. C., Toaff-Rosenstein R. L., Tucker C. B., Miller-Cushon E. K. (2020). Measuring behavior patterns and evaluating time-sampling methodology to characterize brush use in weaned beef cattle. J. Dairy Sci. 103, 8360–8368. doi: 10.3168/jds.2020-18419
Kittawornra A., Zimmerman J. J. (2010). Toward a better undertanding of pig behavior and pig welfare. Ani. Health Reser. Rev. 12, 25–32. doi: 10.1017/s1466252310000174
Knox R. V. (2016). Artificial insemination in pigs today. Theriogenology 85, 83–93. doi: 10.1016/j.theriogenology.2015.07.009
Koopmans S. J., van der Meulen J., Dekker R., Corbijn H., Mroz Z. (2005). Diurnal rhythms in plasma cortisol, insulin, glucose, lactate and urea in pigs fed identical meals at 12-hourly intervals. Physiol. Behav. 84, 497–503. doi: 10.1016/j.physbeh.2005.01.017
Kunavongkrit A., Suriyasomboon A., Lundeheim N., Heard T. W., Einarsson S. (2005). Management and sperm production of boars under differing environmental conditions. Theriogenology 63, 657–667. doi: 10.1016/j.theriogenology.2004.09.039
Machado S. P., Caldara F. R., Foppa L., De Moura R., Gonçalves L. M. P., Garcia R. G., et al. (2017). Behavior of pigs reared in enriched environment: Alternatives to extend pigs attention. PloS One 12. doi: 10.1371/journal.pone.0168427
Martin P., Bateson P. (2007). Measuring Behaviour: an introductory guide, 3rd ed (Cambridge: Cambridge University Press).
Mason G. J. (1991). Stereotypies and suffering. Behav. Processes 25, 103–115. doi: 10.1016/0376-6357(91)90013-P
Mellor D. J., Beausoleil N. J., Littlewood K. E., McLean A. N., McGreevy P. D., Jones B., et al. (2020). The 2020 Five Domains Model: including human-animal interactions in assessments of animal welfare. Animals 10, 1870. doi: 10.3390/ani10101870
Palme R., Möstl E. (1997). Measurement of cortisol metabolites in faeces of sheep as a parameter of cortosol concentration in blood. Z Saugetierkd – Int. J. Mamm. Biol. 62, 192–197.
Petak I., Mrljak V., Tadić Z., Krsnik B. (2010). Preliminary study of breeding boars’ welfare. Vet. Arh. 80, 235–246.
Ruis M. A. W., Te Brake J. H. A., Engel B., Ekkel E. D., Buist W. G., Blokhuis H. J., et al. (1997). The circadian rhythm of salivary cortisol in growing pigs: Effects of age, gender, and stress. Physiol. Behav. 62, 623–630. doi: 10.1016/S0031-9384(97)00177-7
Sabei L., Bernardino T., Parada Sarmiento M., Barbosa B. S., Farias S., de S., et al. (2023a). Life experiences of boars can shape the survival, aggression, and nociception responses of their offspring. Front. Anim. Sci. 4. doi: 10.3389/fanim.2023.1142628
Sabei L., Parada Sarmiento M., Bernardino T., Çakmakçı C., Farias S., de S., et al. (2023b). Inheriting the sins of their fathers: boar life experiences can shape the emotional responses of their offspring. Front. Anim. Sci. 4. doi: 10.3389/fanim.2023.1208768
Schwarz T., Małopolska M., Nowicki J., Tuz R., Lazic S., Kopyra M., et al. (2021). Effects of individual versus group housing system during the weaning-to-estrus interval on reproductive performance of sows. Animal 15. doi: 10.1016/j.animal.2020.100122
Selye H. (1950). Stress and the general adaptation syndrome. Br. Med. J. 1950, 11 1383–1392. doi: 10.1136/bmj.1.4667.1383
Siegford J. M., Rucker G., Zanella A. J. (2008). Effects of pre-weaning exposure to a maze on stress responses in pigs at weaning and on subsequent performance in spatial and fear-related tests. Appl. Anim. Behav. Sci. 110, 189–202. doi: 10.1016/j.applanim.2007.03.022
Stolba A., Wood-Gush D. G. M. (1989). The behaviour of pigs in a semi-natural environment. Anim. Prod. 48, 419–425. doi: 10.1017/S0003356100040411
Stone A. A., Schwartz J. E., Smyth J., Kirschbaum C., Cohen S., Hellhammer D., et al. (2001). Individual differences in the diurnal cycle of salivary free cortisol: A replication of flattened cycles for some individuals. Psychoneuroendocrinology 26, 295–306. doi: 10.1016/S0306-4530(00)00057-3
Šupe-Domić D., Milas G., Drmić Hofman I., Rumora L., Martinović Klarić I. (2016). Daily salivary cortisol profile: Insights from the Croatian Late Adolescence Stress Study (CLASS). Biochem. Med. 26, 408–420. doi: 10.11613/BM.2016.043
Tatemoto P., Bernardino T., Alves L., Cristina de Oliveira Souza A., Palme R., José Zanella A. (2019). Environmental enrichment for pregnant sows modulates HPA-axis and behavior in the offspring. Appl. Anim. Behav. Sci. 220. doi: 10.1016/j.applanim.2019.104854
Tavares M. C. M. S., Arno A., Silveira R. M. F., Lara I. A. R., Salvador M. L., Freitas S. M., et al. (2023). Evaluation of environmental enrichment on thermophysical responses, carcass, traits, and meat quality of finishing pigs. Trop. Anim. Health Prod. 55, 235. doi: 10.1007/s11250-023-03634-z
Teles M. C., Pereira B. A., Pessoa Rocha L. G., Resende C. O., Rodrigues V. V., Pereira L. J., et al. (2016). Semen quality and reproductive performance of boars kept in pens containing conventional coffee husk as a floor covering. Rev. Bras. Zootec. 45, 365–371. doi: 10.1590/S1806-92902016000700002
Teles M. C., Pereira B. A., Rabelo S. S., Pontelo T. P., Chaves B. R., Ferreira R. A., et al. (2017). Effects of coffee husk as floor covering on the behavior of boars. Rev. Bras. Zootec. 46, 883–889. doi: 10.1590/S1806-92902017001200002
Tsuma V. T., Einarsson S., Madej A., Kindahl H., Lundeheim N., Rojkittikhun T. (1996). Endocrine changes during group housing of primiparous sows in early pregnancy. Acta Vet. Scand. 37, 481–490. doi: 10.1186/BF03548088
Wang Z., Goonewardene L. A. (2004). The use of mixed models in the analysis of animal experiments with repeated measures data. Can. J. Anim. Sci. 84, 1–11. doi: 10.4141/A03-123
Wilson S. C., Mitlöhner F. M., Morrow-Tesch J., Dailey J. W., McGlone J. J. (2002). An assessment of several potential enrichment devices for feedlot cattle. Appl. Anim. Behav. Sci. 76, 259–265. doi: 10.1016/S0168-1591(02)00019-9
Wolf T. E., Mangwiro N., Fasina F. O., Ganswindt A. (2020). Non-invasive monitoring of adrenocortical function in female domestic pigs using saliva and faeces as sample matrices. PloS One 15, e0234971. doi: 10.1371/journal.pone.0234971
Zanella A., Brunner P., Unshelm J., Mendl M., Broom D. (1998). The relationship between housing and social rank on cortisol, β-endorphin and dynorphin (1–13) secretion in sows. Appl. Anim. Behav. Sci. 59, 1–10. doi: 10.1016/S0168-1591(98)00115-4
Zhang M. Y., Li X., Zhang X. H., Liu H. G., Li J. H., Bao J. (2017). Effects of confinement duration and parity on stereotypic behavioral and physiological responses of pregnant sows. Physiol. Behav. 179, 369–376. doi: 10.1016/j.physbeh.2017.07.015
Keywords: breeding males, circadian rhythm, cortisol, environmental enrichment, pigs, salivary cortisol, welfare
Citation: Bernardino T, Sabei L, Parada Sarmiento M, Sato D, Farias SdS, Paula TMCGd, Marques GR and Zanella AJ (2025) Behavior and physiology of boars housed in three different environments. Front. Ethol. 4:1540458. doi: 10.3389/fetho.2025.1540458
Received: 05 December 2024; Accepted: 06 March 2025;
Published: 01 April 2025.
Reviewed by:
Dorota Witkowska, University of Warmia and Mazury in Olsztyn, PolandCopyright © 2025 Bernardino, Sabei, Parada Sarmiento, Sato, Farias, Paula, Marques and Zanella. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thiago Bernardino, dGJlcm5hcmRpbm8uYXdAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.