
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Sci., 24 June 2024
Sec. Drylands
Volume 12 - 2024 | https://doi.org/10.3389/fenvs.2024.1398726
 Feiyan Zhao1
Feiyan Zhao1 Zhongju Meng1*Xiaomeng Ren2
Zhongju Meng1*Xiaomeng Ren2 Xiaohong Dang1,3Shiling Shi1Peng Li1Hailong Ding1Jingjie Guo1
Xiaohong Dang1,3Shiling Shi1Peng Li1Hailong Ding1Jingjie Guo1Inner Mongolia’s grassland is one of the most complete natural grasslands for ecological diversity and biodiversity conservation in the world. However, its degradation has seriously affected the plateau’s sustainable development and grassland restoration and reconstruction. To reveal the response of soil-plant synergistic relationship to different degrees of degradation, we selected three degrees of degradation (total coverage of vegetation:20%–30%, LD), moderate degradation (total coverage of vegetation:10%–20%, MD) and severe degradation (total coverage of vegetation:<10%, SD) on typical grasslands in Inner Mongolia (3 transects per degraded plot, three replicates per transect, a total of 27 quadrats). Correspondence between vegetation and soil under different degrees of degradation was obtained by correlation analysis. Soil physical properties influence degradation degree much more than soil depth. Among them, the most obvious total nitrogen (TN) decreased by 251.63% and 125.81% under moderate degradation (MD) and severe degradation (SD) respectively, compared with light degradation (LD). When the degree of degradation increased, the number of species reduced, and when compared to light degradation, moderate degradation and severe degradation decreased by 52.38% and 66.67%, respectively. The declining trend in the number of species was also indicated by Simpson, Margalef, and Shannon-Winener in addition to the Pielou index. The synergy between plant diversity index and soil organic matter (SOM), available potassium (AK) and bacteria was relatively strong.
Grassland is a valuable ecosystem and natural resource in China. It has multiple ecological functions such as water conservation, windbreak and sand fixation, soil and water conservation, soil improvement and agricultural products (Gieselman et al., 2013); the grassland area is more than 1/5 of the global land area. According to Du and Cong (2024), nearly 90% of natural grasslands in China have been degraded to varying degrees since the turn of the 21st century, resulting in serious damage to ecosystem structure and function. Located in the northern Frontier of the motherland, the Inner Mongolia grassland area is an important green ecological security barrier. In addition to being a part of the central grasslands of Eurasia, Inner Mongolia grassland is one of the world’s most complete natural grasslands when it comes to ecological diversity and biodiversity. However, due to increasingly serious climate change and human disturbance, coupled with the predatory demand for grassland resources in the process of urbanization, the species tend to be simplified and the structure and function gradually decline (Jiang et al., 2023).
Grassland degradation is caused by multiple factors (Liu et al., 2018). Overgrazing and climate change are believed to be the primary causes of grassland degradation. By analyzing remote sensing data, it was found that the contribution of overgrazing to alpine grassland degradation on the Qinghai-Tibet Plateau was 19.90%, which was much lower than the contribution of climate change to grassland degradation of 56.74% (Xu et al., 2017). In contrast, climate change affects grasslands (Li et al., 2019). In a sense, the diversity of a plant community is the direct embodiment of community structure and function (Wang et al., 2001). Clarifying community diversity can not only determine the species composition, structure, and function within the region; at the same time, it can explore the interaction between community and environment, which is critically important with regard to the conservation and utilisation of botanical resources, community succession, and ecological restoration (Dong et al., 2020). At the moment, climate change is greatly affecting the assortment of plants, and the decrease in plant diversity will surely have an impact on the stability of the ecosystem (Song et al., 2022). The ecological transition zone consists of a transition area of two vegetation types. A gradual change is occurring in the environmental factors and plant communities. The structure, function, and dynamics of communities are quite complex and sensitive to human disturbances as well as climate change (Chen and Chen, 1998). The interaction between vegetation and soil is one of the most important topics in ecological research (Xie et al., 2011). Because these factors affect plant species’ survival and settlement, recent studies have demonstrated that topography and soil habitat conditions have played a significant role in determining the structure and composition of local plant communities at the local scale (Huggett, 1995). Throughout space and time, soil characteristics vary greatly and are highly related to the distribution of plants (Zhang et al., 2019a). Land degradation is a combination of landscape fragmentation, surface sloughing, decreased productivity, and increased noxious weeds (Li et al., 2013). The majority of research indicates that as degradation increases, vegetation cover, biomass, and diversity indicators generally show a declining trend (Chen et al., 2015; She et al., 2021; Li et al., 2020). Further, it has been established that as degradation increases, biomass and diversity indices signal a rise followed by a fall, with a peak at moderate degradation. During the degradation process, soil physicochemical characteristics deteriorate, and these characteristics are typically manifested as a decline in soil fertility, an increase in soil capacity, and a decrease in soil water content and porosity. Additionally, the carbon cycle of the ecosystems that contribute to the composition of the community is likely to be affected, and sandy landforms may also develop (Zhou et al., 2012). Plant diversity increases along riverbanks and in uplands in China’s arid desert region as water levels rise. This is mostly due to changes in soil water salinity, pH, and nutrients (Zhang et al., 2016; Hu et al., 2021). Found that the pattern of regional variability in vegetation features dependent on density was strikingly similar to the pattern of spatial variability in plant-effective water in the upper 4 m of soil (Ferreira et al., 2007).
This semi-arid typical grassland represents the most representative semi-arid grassland in China with a total area of 11.3265 million hm2 (Liu, 2013). This is a defense line for the prevention and control of desertification land continuing to move north. This study aims to examine the differences in soil chemical and physical attributes, soil microbial population, community of plants composition, and their connections in various degraded grasslands found in both normal grassland areas and sandy transition zones. The specific objective is to analyse the response relationship between degraded grasslands and soil properties in this particular area. As a consequence of the interplay between soil, plants, and microorganisms, an analysis is conducted of the internal mechanisms contributing to grassland degradation. This approach provides a theoretical basis for the rehabilitation and reconstruction of deteriorated grasslands.
This experiment was conducted in the semi-arid desert steppe of Xilinhot, a city in Inner Mongolia, which is located in the northeastern part of China. The coordinates for this steppe are 43°22′-43°29′N and 114°45′-115°04′E. It has a moderate semiarid climate with an average annual temperature of 0.6°C and 260 mm of rainfall. It was found that a dominant species of chestnut soil is prevalent in the study area. The most common types of natural plants are as follows: Leymus chinensis, Artemisia frigida (Ferreira et al., 2007) (Tables 1, 2; Figure 1).
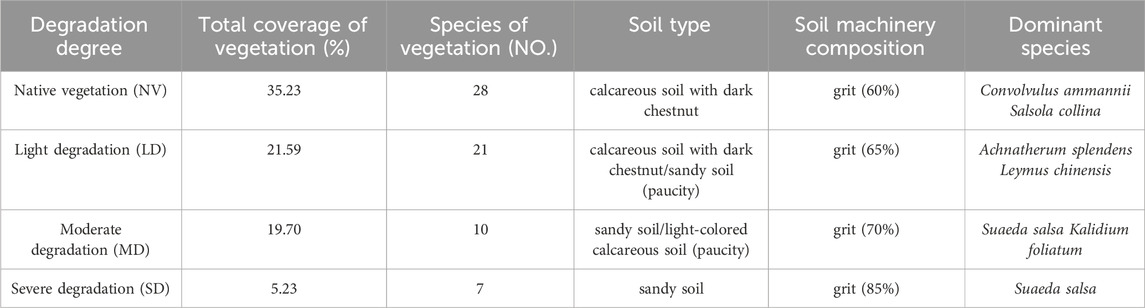
Table 1. Vegetation characteristics of different degraded plots in the thematic areas.
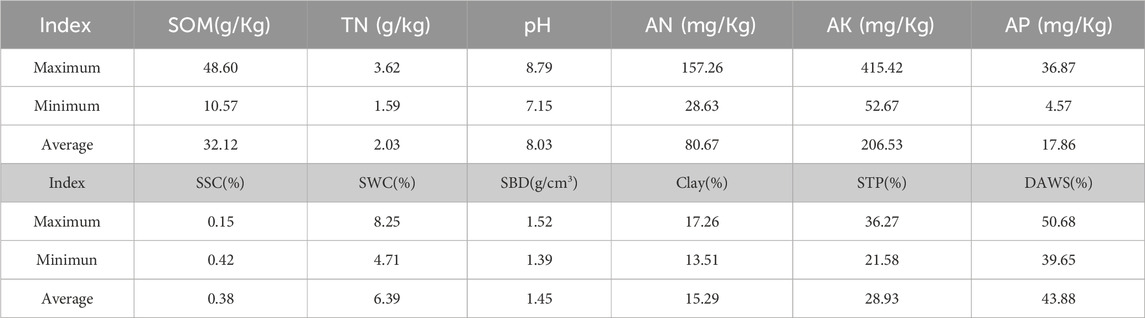
Table 2. Soil characteristics of different degraded plots in the thematic areas.
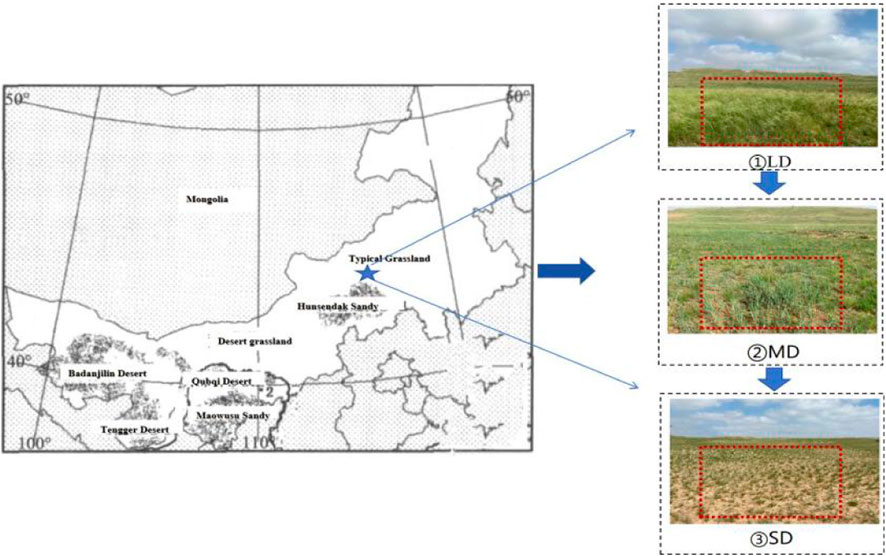
Figure 1. Overview map of the study area. Note: The left side of the picture comes from the literature (Li, 2019), and the right side is the real picture.
Grassland degradation was the focus of research beginning in the year 2023. Three grassland degradation gradients (three sample strips in each degraded sample plot) were included with three replications in a randomised block design to create 27 paddocks (5 × 5 m). For this investigation, three treatments were used: light degradation (LD, total vegetation coverage: 20%–30%), moderate degradation (MD, total vegetation coverage: 10%–20%), and severe degradation (SD, total vegetation coverage:<10%). There is no interference in the experimental plots. Since 2018, the experimental plots have implemented an enclosure policy, and no grazing measures have been taken. There is no significant difference in the topography and slope of the three plots, of which LD:200 × 100 m, MD:100 m × 100 m, SD:100 × 100 m. The linear distance between LD and MD is 20 km, and the linear distance between MD and SD is 50 km (Tables 1, 2).
The sampling was finished by the end of August 2023. Three 1 × 1 m quadrats were used to survey the plant communities within each plot in order to calculate the aboveground biomass and abundance of each species. Each quadrat was examined to identify the plant species present, and the number of bunches (for bunchgrasses) or stems (for rhizomatous grasses) of each species was recorded. All living plants were cut down to ground level and thereafter subjected to a 48-h period of drying in an oven at a temperature of 65°C This process was carried out to ensure a uniform weight measurement.
Late in the month of August, soil samples were collected from the upper 40 cm of the soil profile using an auger with a diameter of 5 cm. There were no precipitation events in the week before and after the experimental sampling. The soil samples were divided into two layers, 0–20 cm and 20–40 cm. This occurred after the plant samples had been collected. This occurred after the plant samples had been collected. In the introduction of the treatments, three soil cores were taken from each plot along a diagonal line. The cores were then merged to form a single sample. Nine soil samples were collected, of which three treatments were tested and three repeats were performed. The soil samples were evenly distributed, one part of which was put into plastic bags for the determination of soil physical and chemical properties, and the other part was stored at low temperature for the determination of microbial quantity. For the purpose of physicochemical examination, half of the samples were dried in the air, while the other half were screened through a 2 mm screen to eliminate any roots that were present. This was done before the analysis of the total nitrogen (TN) concentration in the soil. Following that, the samples were digested using the Kjeldahl method. An analyzer called the TOC-5000A, which was created by Shimadzu Corporation in Kyoto, Japan, was utilized in order to ascertain the concentration of soil organic matter (SOM). A procedure known as water extraction potential was utilized in order to determine the pH of the substrate. Oven drying was the process that was utilized in order to ascertain the soil water content (SWC). A technique known as the wet sieving method (DAWS) was utilized in order to separate the soil water-stable aggregates. For the purpose of determining the total salt content of the soil, the residue method (STP) was utilized. Clay was used in the process of conducting particle size analysis. Electrical conductivity was used as the basis for the calculation of the salt content of the soil. The ring knife gravimetric method was utilized in order to ascertain the soil bulk density (SBD). The diffusion method was utilized in order to ascertain the alkaline nitrogen content of the soil. The antimony colorimetric method (AP) was utilized in order to ascertain the effective phosphorus and molybdenum levels observed in the soil. The NH4Ac leaching-flame photometric method (AK) was utilized in order to quantify the amount of potassium that has a rapid onset of action that was present in the soil. It was determined that nutrient agar (NA), potato dextrose agar (PDA), and corn steep agar (CSA) were the most suitable mediums for the cultivation of actinomycetes, fungi, and bacteria, respectively. The plate colony counting method was utilized in order to determine the amount of actinomyces, bacteria, and fungus (Bao, 2000). Combining a number of soil samples with 10 mL of water resulted in the preparation of a suspension that had a dry weight that was equivalent to 1 g. In order to dilute the soil suspension, Gao’s No. One medium for bacteria, potato-sucrose medium for fungi, and media for actinomycetes were utilized (Lin, 2010). The dilutions that were utilized were 10–2, 10–3, and 10–4 levels.
In this study, Microsoft Excel 2019 was used to preliminarily sort out the data obtained from field investigation and indoor measurement (The data are accurate to 0.01). SPSS was used to test the homogeneity of Levene variance (p > 0.05). Duncan test was used for homogeneity of variance, and Tamhane’s T2 test (IBM SPSS Statistics 23) was used for heterogeneity of variance. One-way analysis of variance and least significant difference method were used for statistics and analysis, and α = 0.05 was used as the significant level of difference. The soil properties in different soil layers under three degradation degrees were counted and compared. Combined with the significance of relevant ecological indicators, Pearson was used to analyze the linear relationship between the three experimental variables, and Originlab 2019 was used for visualization (Table 3).
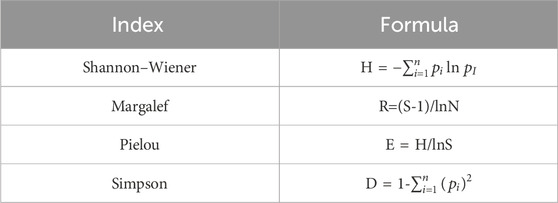
Table 3. The calculation formula of α-diversity index.
Deterioration has a substantial impact on the physical characteristics of the soil; however, their interaction has no effect on the amount of clay in the soil. As compared to the LD, all deterioration levels adversely affected the soil’s physical properties. In contrast to MD and LD, SD had significantly higher soil physical characteristics, with the exception of SWC. Using a more precise analysis, the STP and DAWS values are much greater in SD than in LD at the 0–20 cm depth of the soil layer. In the 20–40 cm layer, the eigenvalue performance pattern is consistent with previous results (Figure 2).

Figure 2. Chemical properties of soils in layers with different levels of degradation. Note: Different letters indicate significant differences (p < 0.05) among different treatments.
It was demonstrated that the chemical characteristics of the soil were greatly influenced by the depth of the soil (p < 0.05). There is a significant correlation between soil depth and soil organic matter, as well as pH, TN, AN, and AP (p < 0.05). In the 0–20 cm and 20–40 cm soil levels, there was no significant difference in soil pH between MD and SD (p > 0.05). However, the soil organic matter content of LD and MD was significantly higher than that of SD in each soil layer. In addition, there was no statistically significant difference in soil pH. Based on the thickness of the soil layer extending from 0–20 cm, LD had a higher total nitrogen concentration than both MD and SD, namely, 251.63% and 125.81%. Further, SD had a greater AN than MD and SD in the soil layer with a height between 0 and 20 cm, respectively. The MD, SD, and LD proved recognizable from one another in any of the soil layers evaluated. In the soil layer between 0 and 20 cm, MD and SD had a lower AP concentration when compared to LD by 235.52% and 271.03%, respectively (Figure 3).
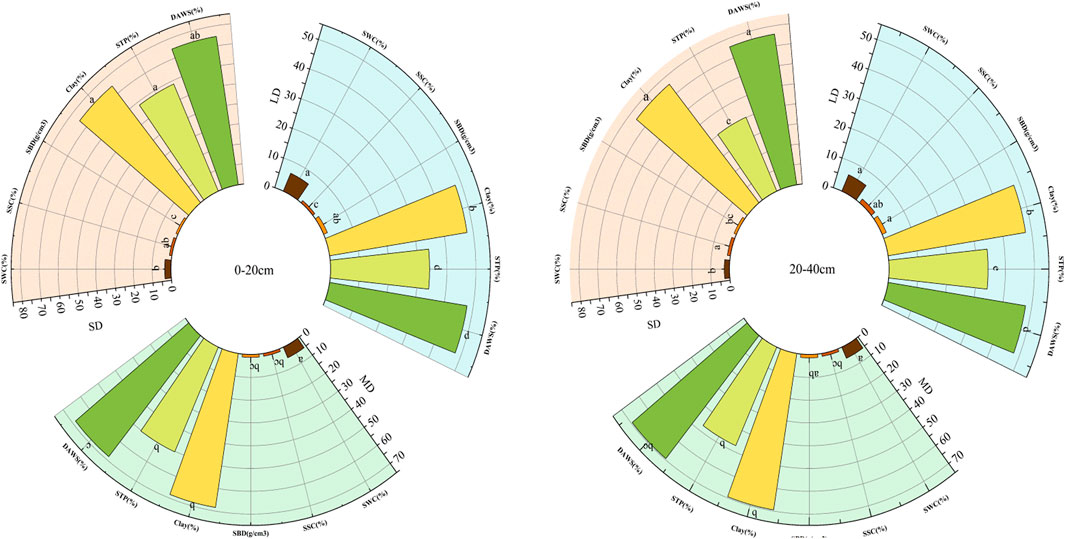
Figure 3. Physical properties of soils in layer swith different levels of degradation. Different letters indicate significant differences (p < 0.05) among different treatments.
The soil bacteria, fungi, and actinomycetes were greatly affected by the depth of the soil and its correlation with the level of degradation (p < 0.05). With the exception of the fungus, the soil microbial populations exhibited a declining trend as soil depth increased (Figure 4). There were notable differences in the amount of soil bacterial biomass between the treatments in the 20–40 cm soil layer. The order of biomass was as follows: MD (24.57 × 105 cfu/g) > LD (11.72 × 105 cfu/g) > SD (8.10 × 105 cfu/g). Conversely, the populations of soil fungi exhibited irregular trends. The greatest numbers of soil fungi are found in the 20–40 cm soil layer. At every stage of degradation, there were major variations in soil actinomycetes between treatments in the 0–20 cm and 20–40 cm soil layers (p < 0.05). The biomass of actinomycetes was shown to be higher in the MD > LC > SD order, with a difference of 150.40%–210.81% compared to the SD (p < 0.05).
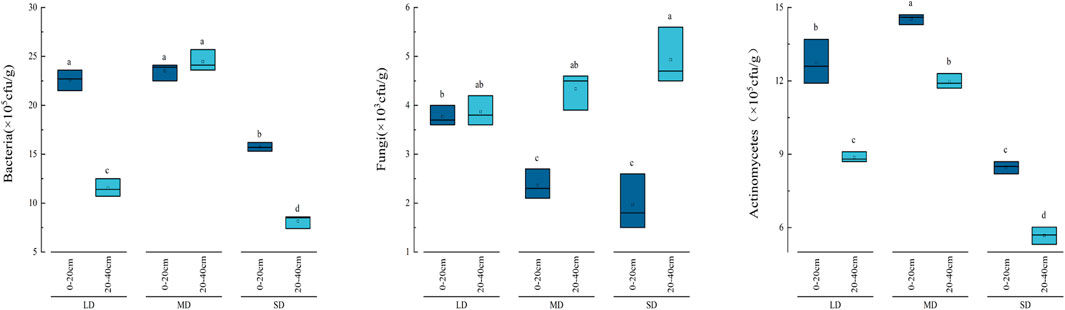
Figure 4. The number of soil microorganisms in different soil layers with different degrees of degradation.
`Statistically, this area is home to nine families, 24 genera, and 28 species of plant groups, all of which are angiosperms; among them, there are 17 species of dicotyledonous plants in seven families and 14 genera, accounting for 60.71% of the total angiosperms. Monocotyledonous plants have two families, 10 genera, and 11 species, accounting for 39.29%, and dicotyledonous plants have obvious advantages. As a natural evolutionary unit in plant taxonomy, family is an important way to reveal the characteristics of plant diversity in this area (Liao et al., 1994). Therefore, the number of species contained in the typical community plants in the semi-arid Aeolian sandy grassland area was counted. The results showed that there was a small family (6–10 species), which was Poaceae (8 genera and eight species), accounting for 11.11% and 28.57% of the total families and species in the area, respectively. There are four families and 16 species of oligotypic families (two to five species), accounting for 44.44% and 57.15% of the total families and species in this area, respectively. There were four families of single species (including one species), accounting for 44.44% and 14.29% of the total families and species in this area. Genus is an important research object reflecting the phylogenetic process and diversity characteristics of plants (Cox and Moore, 2007). The number of species in 24 genera of seed plants in typical communities in sandy grassland was counted. The results showed that there were mainly single species (including one species), a few species (two to five species), and two types. There were three genera and seven species of oligotypic genera (two to five species), accounting for 12.50% and 25.00% of the total number of genera, respectively. There are 22 genera of single species, such as Agropyron, Cleistogenes squarrosa, Plantago, etc., accounting for 87.50% and 75.00% of the total genera and total species, respectively. They account for the highest proportion in all types of genera, and occupy an absolute advantage in the plant composition of the area, constituting the main body of the plant genera in the area (Table 4).
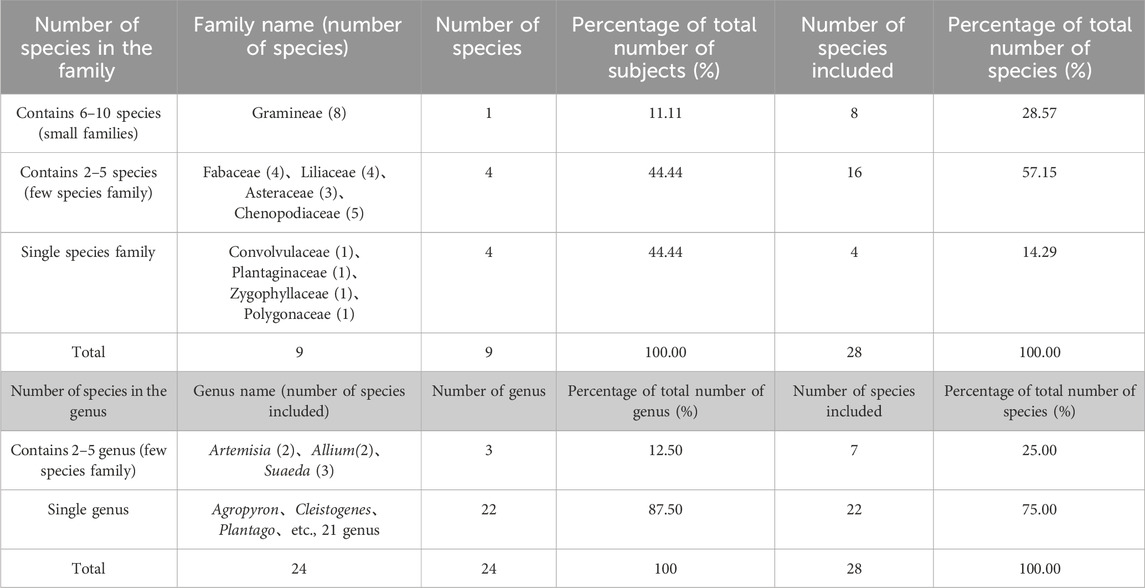
Table 4. Diversity in the composition of plant families and genera.
Table 5 illustrates how the number of species within the plant community differed throughout treatments. For this investigation, a total of 28 species were gathered. Out of all of them, LD has the most plant species (21). Each of the three treatments had a different dominant species: in LD, Achnatherum splendens, Leymus chinensis, and Allium tenuissimum were the dominant species with dominance percentages of 16.72%, 14.93%, and 13.73%; in MD, Suaeda glauca, Kalidium foliated, and Tragus racemosus were the dominant species with dominance percentages of 34.78%, 26.09%, and 8.70%, respectively. The Simpson index did not reveal any significant differences between LD and SD (Figure 5).
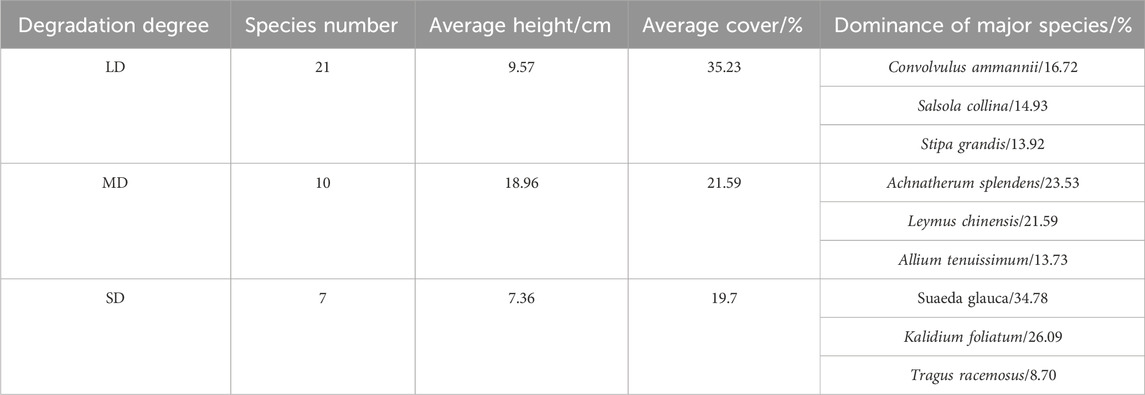
Table 5. Species composition of typical degraded grasslands.
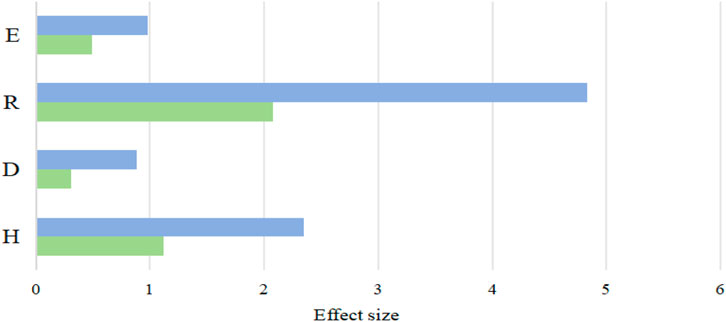
Figure 5. Species diversity. Note: blue represents the maxi value, green represents the min value. E: Pielou, R: Margalef, H: Shannon-Wiener, D: Shannon-Wiener.
Overall, the present research found that soil environmental parameters had a non-significant negative connection with the Pielou index. Nevertheless, there was a positive association (p < 0.05) between the Shannon-Weiner index and pH, SOM, TN, AK, AN, AP, SWC, SSC, SBD, Clay, DAWS, fungi, and actinomycetes. Additionally, there were notable negative connections (p < 0.05) between the index and STP and microorganisms. Lastly, pH, SOM, TN, AK, AN and AP were positively correlated with the Margalef index (p < 0.05).
The results of this investigation revealed the following: the Pielou index exhibited a non-significant negative correlation with soil environmental factors in their entirety; the Shannon-Weiner index demonstrated a statistically significant positive correlation with pH, SOM, TN, AK, AN, AP, SWC, SSC, SBD, Clay, and DAWS, fungi and actinomycetes; and STP and bacteria exhibited significant negative correlations (p < 0.05). The Margalef index determined to be significantly positively correlated with pH, SOM, TN, AK, AN, AP, SAW, STP, and bacteria (p < 0.05) (Figure 6).
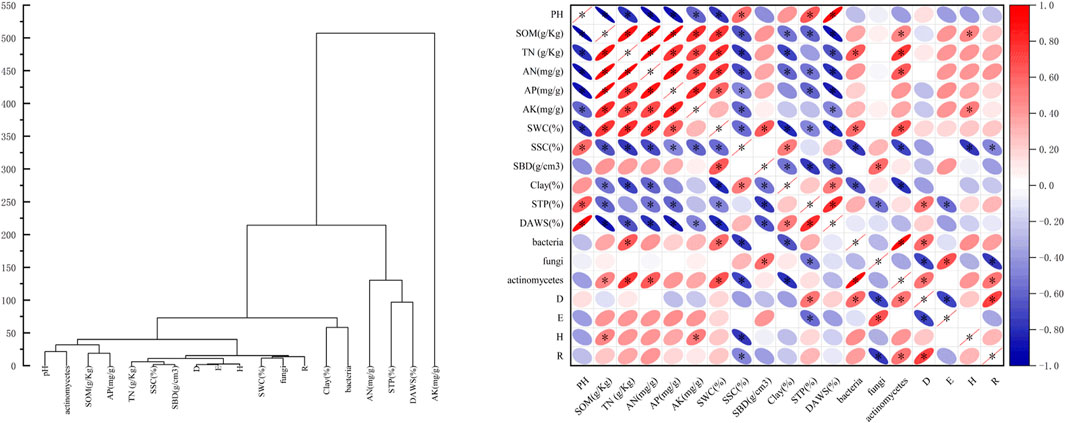
Figure 6. Correlation analysis between soil factors.
Plant species richness decreased with increasing degree of grassland degradation, which is consistent with the results of previous studies on grassland degradation and plant community characterization (Cui et al., 2022), indicating the vulnerability and sensitivity of plant community response to degraded grassland. It may be that most of the grass and sedge species could not survive due to the tendency of the degradation from light to heavy degradation and the occurrence of sloughing or even crusting on the ground surface, which transformed the adapted species into salinity and drought tolerant ones. Uniformity is considered an effective indicator of the degree of resource availability, with lower uniformity indicating underutilization of resources and higher indicating that resources are fully and uniformly utilized; it increased with increasing degradation in this study, but the change was less pronounced. This may be due to the shrinkage of functional spatial extent with increasing degradation degree, thus increasing the degree of evenness of species in the limited functional space (Sheng, 2015). Vegetation community height and cover are the core indicators for assessing productivity, and in this study, we found that the cover of the vegetation community became lower and lower with the increase of degradation, but the average height showed a tendency of increasing and then decreasing; the results of the cover were consistent with most of the results, and the differences in height may be due to the differences in plant traits caused by the differences in families and genera in the process of vegetation community succession (Zhou et al., 2021). An essential metric for assessing the stability of communities or ecosystems is the community species diversity index, which shows the structural level of species composition in the community (Zhang et al., 2019b; Ji et al., 2021). In areas that consist of wind-blown sand, changes in the status of dominant species and typical grasslands have an important effect on the species diversity and community composition of grassland ecosystems. Consistent with the results of previous studies, there is a significant decrease in plant height and cover with increasing degradation. The findings of this investigation are, for the most part, in agreement with the findings of the study that is now being conducted. The peak value of species diversity during the stage of heavy degradation may be attributed to the alpine meadow ecosystem’s initial stability during that phase. However, as the degree of degradation increases, grasses and other high-quality forage grasses become scarcer, and the invasion of Asteraceae and poisonous weed species increases, the community’s structure and diversity change. Furthermore, the preference for greater diversity in the community contributes to this phenomenon. Seed dispersal, it is easy to form the number of severely degraded grassland species is significantly more than the number of moderately degraded grassland species (Wang et al., 2015). It has also been demonstrated that as the degree of degradation increases, Margalef and Shannon-Winener identified a pattern of successive increases and decreases, and determined that the region with the greatest value is the one that is moderately deteriorated (Hao et al., 2020). Additionally, the species richness surpasses that of the area with moderate degradation.
According to the study’s findings, which were consistent with those of earlier investigations, the soil in the study area was neutral to slightly alkaline (between 6.3 and 9.7) (Joyce, 2014). This far exceeds the optimum pH for forage growth, resulting in low nutrient effectiveness and unfavorable growth. Meanwhile, microbial communities will be pressured by environmental stress when pH is neutral so soil microbiology such as nutrient cycling will be affected. pH shows an increasing trend with the degree of degradation, which may be due to the destruction of soil organic acids as a result of strong UV exposure in low-covered areas (Kolos and Banaszuk, 2013). Elevated soil salinity levels will result in reduced nutrient material resource utilization by plants, thereby exacerbating the vulnerability of already endangered species and ultimately contributing to a shift in species diversity (Wei et al., 2022) (Degradation of alpine wetlands and its effects on soil parameters and plant community characteristics). Consistent with previous research, the degree of degradation leads to a reduction in the compost content of the soil (Rebelo et al., 2018). This may be due to the fact that plant diversity is limited in heavily degraded areas and above-ground biomass is reduced, which in turn provides less substrate for nutrient cycling; in addition, the vegetation cover is low in heavily degraded areas, which makes it easy to erode the nutrients in the surface layer. Consistent with the findings of numerous studies, the association between soil organic matter and N content and vegetation modification and species diversity is positive (Hu et al., 2016; Jimenz-ballesta et al., 2018; Zhao et al., 2020; Yuan and Jiang, 2021). As the degree of degradation increases, there is also a tendency to earn the pH value to rise. It may be linked to the enhanced evaporation of water in the rising region and a decline in vegetation cover, which accelerates the speed of evaporation. This phenomenon could potentially be attributed to the elevated altitude and reduced vegetation cover, both of which contribute to heightened evaporation rates. Moreover, the evaporation process facilitates the accumulation of salts along the capillary water, which subsequently elevate the pH value of the soil (Li et al., 2018; Sun et al., 2020, Peng et al., 2020; She et al., 2021; Shao et al., 2022).
Soil bacteria are the dominant group of soil microorganisms and play an important role in nutrient recycling (li et al., 2017). In addition to soil fungi, the number of soil microorganisms decreased with the increase of soil depth, which may be due to the fact that the soil was not completely destroyed. The soil surface layer is the area with the largest distribution of plant roots. A large number of vegetation litters gather in the surface layer and decompose to produce soil organic matter. Soil organic matter is the material source of microbial decomposition and absorption. Coupled with relatively suitable water, heat and ventilation conditions, it is conducive to the growth and high-speed reproduction of microorganisms and promotes the dynamic changes of material and energy in grassland ecosystems (Wu et al., 2013). At the same time, it is opposite to the competition law of fungal community formation, but it is easy to achieve system balance. Through correlation analysis, it was found that there was a significant correlation between plant community characteristics and soil physical and chemical properties (p < 0.0.5). Among them, soil bulk density was negatively correlated with vegetation diversity, indicating that the increase of soil bulk density limited the transport of nutrients and water in soil, directly affected the absorption of nutrients by plants, and was not conducive to the growth and development of plants (Zou et al., 2023). Soil total nitrogen, soil total phosphorus and soil water content directly determine the height of vegetation and the level of biodiversity (Genga et al., 2022).
The physical and chemical characteristics of the soil are significantly altered in typical degraded grasslands. In damaged grasslands, soil microbial communities were also changed, with bacteria and actinomycetes predominating. The typical steppe’s soil properties declined as soil depth increased during the same degradation stage, in addition to pH and soil microbial abundance. When the degree of deterioration increased, the same soil depth saw a decline in the characteristics of the soil, particularly in the areas of 0–20 cm where TN, AN, SWC, STP, and DAWC decreased dramatically. Therefore, it is critical to fortify surface soil protection in order to preserve the soil properties of typical grassland areas. In this area, there are 28 species of plants that are divided into nine families and 24 genera. Apart from Pielou’s index, the diminishing variation pattern with increasing degradation was also observed in Simpson, Margalef, Shannon-Winener, and Winener’s index. There was a positive correlation (p < 0.05) between the plant community diversity and SOM, AK, and bacteria, and a negative correlation (p < 0.05) with SSC and fungi. Consequently, altering the aforementioned four soil qualities can improve the diversity of community structure during the restoration and reconstruction of the typical steppe.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
FZ: Conceptualization, Investigation, Software, Writing–original draft. ZM: Writing–original draft, Resources. XR: Resources, Writing–original draft. XD: Methodology, Writing–original draft. SS: Software, Writing–original draft. PL: Formal Analysis, Writing–original draft. HD: Investigation, Writing–original draft. JG: Resources, Writing–original draft.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was supported by the National Natural Science Foundation of China [42067015]; Desert Ecosystem Conservation and Restoration Innovation Team [BR22-13-03].
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Bao, S. D. (2000). Soil and agricultural chemistry analysis. Beijing, China: China Agriculture Press.
Chen, L. L., Shi, J. J., Wang, Y. L., Ma, Y. S., Dong, Q. M., and Hou, X. K. (2015). Study on different degraded degrees grassland community structure characteristics of the alpine area. Acta Agrestia Sin. 24, 210. doi:10.11733/jissn.1007-0435.2016.01.029
Chen, X. D., and Chen, Z. X. (1998). The Determination of ecotone and thecharacteristics of biome on ordos plateau. Acta Phytoecol. Sin. 22, 312.
Cox, C. B., and Moore, P. D. (2007). Biogeography. Zhao tieqiao trans. Beijing: Higher Education Press.
Cui, H. W., Wang, C., Wang, X. T., Liu, Z., Liu, K., Chen, S., et al. (2022). The loss of above-and belowground biodiversity in degraded grasslands drives the decline of ecosystem multifunctionality. Appl. Soil Ecol. 172, 104370. doi:10.1016/j.apsoil.2021.104370
Dong, X., Xin, Z. M., Duan, R. B., Huang, Y. R., Jian, K. M., and Ma, H. F. (2020). Species diversity of typical shrubs and niches of dominant shrub species in the Ulanbuh Desert. Arid Zone Res. 37, 1009. doi:10.13866/j.azr.2020.04.22
Du, Z. Y., and Cong, N. (2024). Responses of vegetation and soil characterisitics to degraed grassland under different degrees on the Qinghai-Tibet Plateau. Acta Ecol. Sin. 44, 2504–2024. doi:10.20103/j.stxb.202303310628
Ferreira, J. N., Bustamante, M., Garcia-Montiel, D. C., Caylor, K. K., and Davidson, E. A. (2007). Spatial variation in vegetation structure coupled to plant available water determined by two-dimensional soil resistivity profiling in a Brazilian savanna. Oecologia 153, 417–430. doi:10.1007/s00442-007-0747-6
Genga, Y. P., Zhou, S., Yang, K., Tang, M. K., Zeren, Y. Z., Yang, T. Y., et al. (2022). Community characteristics and soil physical and chemical properties under differentdegraded grassland in Zoige,Sichuan. J. Southwest Minzu Univ. Sci. Ed. 48, 369. doi:10.11920/xnmdzk.2022.04.003
Gieselman, T. M., Hodges, K. E., and Vellend, M. (2013). Human-induced edges alter grassland community composition. Biol. Conserv. 158, 384–392. doi:10.1016/j.biocon.2012.08.019
Hao, A. H., Xue, X., Peng, F., You, Q. G., Liao, J., Duan, H. C., et al. (2020). Different vegetation and soil degradation characteristics of a typical grassland in the Qinghai-Tibetan Plateau. Acta Ecol. Sin. 40, 964. doi:10.5846/stxb201809162019
Hu, D., Lu, G. H., Wang, H. F., Yang, Q., and Cai, Y. (2021). Response of desert plant diversity and stability to soil factors based on water gradient. ACTA Ecol. SIN. 41, 6738. doi:10.5846/stxb20200617158
Hu, Z. M., Li, S. G., Guo, Q., Niu, S. L., He, N. P., Li, L. H., et al. (2016). A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China. Glob. Change Biol. 22, 1385–1393. doi:10.1111/gcb.13133
Ji, B., He, J. L., Wang, Z. J., and Jiang, Q. (2021). Characteristics and composition of vegetation carbon storage in natural grassland in Ningxia,China. Chin. J. Appl. Ecol. 32, 1259–1268. doi:10.13287/j.1001-9332.202104.005
Jiang, Y. D., LiuZheng, X. Y., Guo, W. X., and Zhao, T. Q. (2023). Degradation status of grassland resources and exploration of ecological restoration in Inner Mongolia. Grassl. Prataculture 35 (57).
Jimenz-ballesta, R., Garcia-Navarro, F. J., Martin-consuegra, S. B., delos Reyes, C. P., Ortiz-villajos, J. A. A., and Miguel, M. F. S. (2018). The impact of the storage of nutrients and other trace elements on the degradation of a wetland. Int. J. Environmrntal Rasearch 12 (87), 87–100. doi:10.1007/s41742-018-0072-4
Joyce, C. B. (2014). Ecological consequences and restoration potential of abandoned wet grasslands. Ecol. Eng. 66 (91), 91–102. doi:10.1016/j.ecoleng.2013.05.008
Kolos, A., and Banaszuk, P. (2013). Mowing as a tool for wet meadows restoration: effect of long-term management on species richness and composition of sedge-dominated wetland. Ecol. Eng. 55 (23), 23–28. doi:10.1016/j.ecoleng.2013.02.008
Li, J. H., Yang, G. J., and Wang, S. P. (2020). Vegetation and soil characteristics of degraded alpine meadows on the Qinghai-Tibet Plateau,China: a review. Chin. J. Appl. Ecol. 31, 2109–2118. doi:10.13287/j.1001-9332.202006.002
Li, J. M. (2019). The study of impact of earth surface on sand and dust storm in midwestern inner monglia. Nanjing unverisity Inf. Science&Technology.
Li, L., Zhang, F. P., Feng, Q., Wang, H. W., Wei, Y. F., Li, X. J., et al. (2019). Responses of grassland to climate change and human activities in the area around Qinghai Lake. Chin. J. Ecol. 38, 1157. doi:10.13292/j.1000-4890.201904.032
Li, R., Liu, M. X., Zhang, C., Zhao, R. D., and Shao, P. (2017). Distribution characteristics of soil microbial communities along different slope direction of Gannan sub-alpine meadows. Ecol. Environ. Sci. 26, 1884. doi:10.16258/j.cnki.1674-5906.2017.11.009
Li, W., Wang, J. L., Zhang, X. J., Shi, S. L., and Gao, W. X. (2018). Effect of degradation and rebuilding of artificial grasslands on soil respiration and carbon and nitrogen pools on an alpine meadow of the Qinghai-Tibetan Plateau. Ecol. Eng. 111, 134–142. doi:10.1016/j.ecoleng.2017.10.013
Li, X. L., Gao, J., Brierley, G., Qiao, Y. M., Zhang, J., and Yang, Y. W. (2013). Rangeland degradation on the Qinghai-Tibet Plateau: implications for rehabilitation. Land Degrad. 24 (72), 72–80. doi:10.1002/ldr.1108
Liao, W. B., Zhang, H. D., and Zhong, M. J. (1994). Floristic geographical analysis of the subtropical flora of Guangdong Province. Ecol. Sci. 1 (47).
Lin, X. G. (2010). Principleand methods of soil microbiology research. Beijing, China: Higher Education Press.
Liu, A. R., Yang, T., Xu, W., Shangguan, Z. J., Wang, J. Z., Liu, H. Y., et al. (2018). Status, issues and prospects of belowground biodiversity on the Tibetan alpine grassland. Biodivers. Sci. 26, 972–987. doi:10.17520/biods.2018119
Liu, J. (2013). Study on impact of mowing and grazing on vegetation and soilof typical steppe in Xilingol, Inner Mongolia. Chin. Acad. ofAgricultural.
Peng, F., Xue, X., Li, C., Lai, C., Sun, J., Tsubo, M., et al. (2020). Plant community of alpine steppe shows stronger association with soil properties than alpine meadow alongside degradation. Sci. Total Environ. 733, 139048. doi:10.1016/j.scitotenv.2020.139048
Rebelo, A. J., Emsens, W. J., Meire, P., and Esler, K. J. (2018). The impact of anthropogenically induced degradation on the vegetation and biochemistry of South African palmiet wetlands. Wetl. Ecol. Manag. 26, 1157–1171. doi:10.1007/s11273-018-9638-3
Shao, J. X., Liu, Y. H., Ma, F., and Wei, W. D. (2022). Meta-analysis of physical and chemical properties of shallow soils in degraded alpine grasslands. Acta Agrestia Sin. 30 (1). doi:10.11733/j.issn.1007-0435.2022.06.007
She, Y. D., Yang, X. Y., Ma, L., Zhang, Z. H., Wang, D. J., Huang, X. T., et al. (2021). Study on the characteristics and interrelationship of plant community and soil in degraded alpine meadow. Acta Agrestia Sin. 29 (62). doi:10.11733/j.issn.1007-0435.2021.Z1.008
Sheng, L. (2015). Plant commtity functional diversity along degradation gradients in an alpine meadow. yunnnan.
Song, Z. B., Xin, Z. M., and Zhu, Y. J. Characteristics of shrub communities in desert steppe ecotone of Inner Mongolia, China. Journal of Desert Research, 42,104, 2022. doi:10.7522/j.issn.1000-694X.2021.00128
Sun, Z., Wang, Y. B., Liu, G. H., et al. (2020). Heterogeneity analysis of soil particle size distribution in the process of degradation of alpine meadow in the permafrost regions based on multifractal theory. J. Glaciol. Geocryol. 37, 980. doi:10.7522/j.issn.1000-0240.2015.0109
Wang, D. P., Ji, S. Y., and Chen, F. X. (2001). A review on the species diversity of plant community. Chin. J. Ecol. 55. doi:10.13292/j.1000-4890.2001.0079
Wang, X. X., Dong, S. K., Sherman, R., Liu, Q. R., Liu, S. L., Liu, Y. Y., et al. (2015). A comparison of biodiversity–ecosystem function relationships in alpine grasslands across a degradation gradient on the Qinghai–Tibetan Plateau. Rangel. J. 37 (45), 45. doi:10.1071/rj14081
Wei, J. J., Qin, R. M., Zhang, Z. H., Li, Y. L., Deng, Y. F., Ma, L., et al. (2022). Characteriscand interrelationship of plant community and soil properties in different degraded alpine grassland. Acta Ecol. Sin. 30, 3035. doi:10.11733/j.issn.1007-0435.2022.11.020
Wu, X., Li, H. X., Fu, B. J., Jin, T. T., and Liu, G. H. (2013). Study on soil characteristics of alpine grassland in different degradation levels in headwater regions of three rivers in China. Chin. J. Grasssl. 1673.
Xie, Y. Z., Xu, D. M., Shi, X. F., Qi, T. Y., Liu, L. D., and Wang, D. P. (2011). Spatial pattern of soil moisture and vegetation attributes along the criticalarea of desertification in Southern MuUs Sandy Land. ACTA Ecol. SIN. 31, 2697.
Xu, H. J., Wang, X. P., and Zhang, X. X. (2017). Impacts of climate change and human activities on the aboveground production in alpine grasslands: a case study of the source region of the Yellow River, China. Arabian J. Geosciences 10 (1), 17. doi:10.1007/s12517-016-2801-3
Yuan, Z. Q., and Jiang, X. J. (2021). Vegetation and soil covariation, not grazing exclusion, control soil organic carbon and nitrogen in density fractions of alpine meadows in a Tibetan permafrost region. Catena 196, 104832. doi:10.1016/j.catena.2020.104832
Zhang, D. J., Qi, Q., Tong, S. Z., Wang, X. H., An, Y., Zhang, M. Y., et al. (2019b). Soil degradation effects on plant diversity and nutrient in tussock meadow wetlands. J. soil Sci. Plant Nutr. 19, 535–544. doi:10.1007/s42729-019-00052-9
Zhang, Q., Shao, M., Jia, X., and Wei, X. (2019a). Changes in soil physical andchemical properties after short drought stress in semi-humid forests. Geoderma 338, 170–177. doi:10.1016/j.geoderma.2018.11.051
Zhang, X. N., Yang, X. D., and Lü, G. H. (2016). Diversity patterns and response mechanisms of desert plants to the soil environment along soil water and salinity gradients. Acta Ecol. Sin. 36, 3206.
Zhao, C. Z., Zhang, H., Song, C. P., Zhu, J. K., and Shabala, S. (2020). Mechanisms of plant responses and adaptation to soil salinity. Innov. (Camb). 1 (69), 100017. doi:10.1016/j.xinn.2020.100017
Zhou, H. K., Zhao, X. Q., Wen, J., Chen, Z., Yao, B. Q., Yang, Y. W., et al. (2012). The characteristics of soil and vegetation of degenerated alpine steppe in the Yellow River Source Region. Acta Pratac 21 (1).
Zhou, T. C., Hou, G., Sun, J., Zong, N., and Shi, P. (2021). Degradation shifts plant communities from S- to R-strategy in an alpine meadow, Tibetan Plateau. Tibetan Plateau Sci. Total Environ. 800, 149572. doi:10.1016/j.scitotenv.2021.149572
Keywords: typical grasslands, degradation, soil physicochemical properties, plant community diversity, ecological restoration
Citation: Zhao F, Meng Z, Ren X, Dang X, Shi S, Li P, Ding H and Guo J (2024) Changes in plant-soil synergistic patterns along grassland degradation gradients in northern China. Front. Environ. Sci. 12:1398726. doi: 10.3389/fenvs.2024.1398726
Received: 12 March 2024; Accepted: 13 May 2024;
Published: 24 June 2024.
Edited by:
Heping Liu, Washington State University, United StatesReviewed by:
Shaokun Wang, Northwest Institute of Eco-Environment and Resources (CAS), ChinaCopyright © 2024 Zhao, Meng, Ren, Dang, Shi, Li, Ding and Guo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhongju Meng, bWVuZ3pob25nanVAMTI2LmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.