
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Sci. , 25 September 2023
Sec. Freshwater Science
Volume 11 - 2023 | https://doi.org/10.3389/fenvs.2023.1204787
 Kleber Renan de Souza Santos1,2*†
Kleber Renan de Souza Santos1,2*† Guilherme Scotta Hentschke3*†
Guilherme Scotta Hentschke3*† Graciela Ferrari4Ana Paula Dini Andreote5
Graciela Ferrari4Ana Paula Dini Andreote5 Marli de Fátima Fiore5
Marli de Fátima Fiore5 Vitor Vasconcelos3,6Célia Leite Sant’Anna7
Vitor Vasconcelos3,6Célia Leite Sant’Anna7We studied nineteen populations of A. platensis (Microcoleaceae, Cyanobacteria), from Pantanal (Brazil) shallow, saline and alkaline lakes. The lakes are connected to the Paraná and Paraguay Rivers Basins, and during wet seasons, the waters flow towards La Plata River estuary. Morphology of natural populations and cultures were analyzed using optical microscope, and 16S rDNA sequences were used for the BI and ML phylogenetic analysis. The morphological analysis shows that our populations fit in the original description of A. platensis, but with additional aerotopes. Also, it is evident that these structures are facultative and the species is planktonic, rather than benthic without aerotopes, as originally described. The phylogenetic analysis shows our strains in the monophyletic Limnospira clade. Considering that, in this paper we transfer the species A. platensis to the genus Limnospira, based on phylogenetic and morphological data. This new taxonomical combination is supported also by ecological data, and indicates that the species is more related and abundant in Pantanal, than in La Plata Basin, region from where it was originally described. According to our results, L. platensis is planktonic or benthic and typical from saline, alkaline and warm waters.
Arthrospira platensis (Nodst.) Gomont is a Cyanobacteria largely used in industrial scale, as a food supplement, under the commercial name “Spirulina”. Although it is one of the most studied Cyanobacteria, due to its economic importance, its taxonomy is still controversial (Nowicka-Krawczyk et al., 2019). In an important taxonomic revision of the genus Arthrospira (Komárek and Lund, 1990), the authors included Spirulina maxima and S. fusiformis in the planktonic forms (with aerotopes) of Arthrospira, as A. maxima Setchell et Gardner and A. fusiformis (Voronichin) Komárek et Lund. Based on an exsiccate of the type material of A. platensis, held at the British Museum of Natural History, the authors considered this species as benthic, without aerotopes. Some authors disagree and argue that aerotopes are lost in dry material (Tomaselli, 1997) and that the Gomont iconotype does not show all the morphological variability of the species. Because of that, A. platensis hypothetically could present aerotopes and be planktonic.
In this paper, we extensively studied nineteen A. platensis populations from saline and alkaline lakes of Pantanal da Nhecolândia, Brazil. The Pantanal Biome is located in the center of South America and is the greatest wetland in the planet (Allem and Valls, 1987). In the sub-region Pantanal da Nhecolândia, the presence of thousands of shallow lakes (maximum 2 m depth), predominantly rounded are remarkable. These lakes are commonly dominated by cyanobacterial blooms of A. platensis and Anabaenopsis elenkinii Miller. The populations of A. platensis in our samples, fit in the original description of the species, but present facultative aerotopes and are phylogenetic related to the genus Limnospira Nowicka-Krawczyk et al. Consequently, based on phylogenetic, morphological and ecological data, in this paper we propose to transfer A. platensis to the genus Limnospira.
Nineteen water samples were collected in dry and wet seasons from 2004 to 2012, by immersing bottles, in “Pantanal da Nhecolândia”, Brazil, lakes. Located between Paraná and Paraguay Rivers, the region is characterized by the presence of thousands of shallow lakes (Figure 1) with alkaline (pH 9–11) and saline waters with high conductivity (>2000 μS cm-1), dominated by the Cyanobacteria A. elenkinii and “A. platensis”. The lakes are part of the Paraguay River Basin, which is composed by the Paraguay-Paraná-Plata Rivers systems. During the wet seasons, the lakes are connected to this system and the waters flow towards La Plata River estuary (Santos and Sant’Anna, 2010).

FIGURE 1. (A) General view of Pantanal da Nhecolândia lakes, (B) Salina do Meio in Nhumirim Farm (Credits: (A). A.Y. Sakamoto 2001, (B) A. Pott, In: Pott and Pott, 2000).
For the qualitative analysis of A. platensis natural populations, parts of the samples were preserved with 4% formalin and later included in the collection of the State Scientific Herbarium “Maria Eneida P. Kauffman Fidalgo” (SP), in the Institute of Botany, São Paulo, Brazil. For the isolation of strains, the samples were kept under refrigeration in a Styrofoam box with ice, and then transported to the laboratory of the Institute of Botany. The isolation was made using microscope and a micropipette to transfer individuals to tubes with BG-11 (Stanier et al., 1971) liquid media, adjusted to pH 9.5 with NaOH (Santos et al., 2011). The three isolate strains were included in the Institute of Botany Culture Collection (CCIBt), São Paulo, Brazil, and kept under controlled conditions: temperature 23°C, irradiance 40–50 μmol photons m -2 s -1, photoperiod 14-10 h light-dark, and transferred to new flasks every 40 days. In the studied lakes, the parameters temperature, salinity, conductivity and pH were measured with a WTW 340i probe.
Morphological analysis was performed based on nature material using optical microscope. The following characters were analyzed: 1) general trichome morphology and measurements; 2) apical cells morphology and measurements; 3) cells morphology and measurements; 4) number of spirals per trichome; 5) spirals height; 6) distance between spirals; 7) presence of aerotopes; 8) presence of mucilaginous envelope using China Ink.
DNA extraction was performed according to (Fiore et al., 2000). The 16S rDNA and the 16S-23S ITS region amplification was performed using the primers 27F1 (5′-AGAGTTTGATCCTGCTCAG-3′) (Neilan et al., 1997), and 23S30R (5′-CTTCGCCTCTGTGTGCCTAGGT-3′) (Lepère et al., 2000) under the following conditions: Heat 94°C/5 min; 10 cycles 94°C/45s, 57°C/45s, 72°C/2min; 25 cycles 94°C/45s, 54°C/45s, 72°C/2min and final extension 72°C/7min, and then sequenced. The inserts were cloned into “pGEM®-T Easy Vector Systems” (Promega, Madison, WI, EUA), pGEM®–T Easy Vector System (Promega, Madison, WI, United States) according to the supplier’s manual, cloned by heat–shock in E. coli DH5α cells and plated for blue–white selection (Sambrook et al., 1989). Three white colonies for each strain were selected and the plasmid extractions were performed by the alkaline lysis method (Birnboim and Doly, 1979). The gene was sequenced using “Big Dye Terminator” version 3.0 (Applied Biosystems) with the plasmid primers T7 and M13 and the internal primers 357F/357R, 704F/704R and 1114F/1114R (Lane, 1991). The sequences were deposited in NCBI under the accession numbers: Arthrospira SM CCIBt3335 (OR142670), Arthrospira IP CCIBt3336 (OR142671) and Arthrospira PSol CCIBt3254 (OR142672).
The 16S rDNA sequences obtained in this study were aligned using ClustalW, in MEGA11: Molecular Evolutionary Genetics Analysis version 11 (Tamura et al., 2021), with Cyanobacteria reference sequences of Microcoleaceae, Oscillatoriaceae and Desertifilaceae, retrieved from GenBank. The final dataset contained 42 sequences with 1,247 informative sites. The phylogenetic trees were built using Maximum Likelihood and Bayesian Inference analysis. GTR + G + I evolutionary model was selected by MEGA 11. The robustness of ML tree was estimated by bootstrap percentages, using 1,000 replications using IQ-Tree online version v1.6.12 (Trifinopoulos et al., 2016). Bayesian trees were constructed in two independent runs, with four chains each, for 5 × 106 generations, burnin fraction set to 0.25, sample frequency 1,000, using MrBayes (Ronquist et al., 2012) in Cipres Gateway (Miller et al., 2010).
The Bayesian Inference Phylogeny (Figure 2) (average standard deviation of split frequencies: 0.0095), shows strong backbone support and shows our strains L. platensis CCIBt3335, CCIBt3336 and CCIBt3254 strongly supported (BI = 1) in the monophyletic Limnospira clade (Nowicka-Krawczyk et al., 2019), close related to the Microcoleaceae genera Neolyngbya, Lyngbya, Capilliphycus and Limnoraphis (BI = 1), and distant from the Arthrospira clade, represented by the reference strains A. jenneri A10 MDA (Nowicka-Krawczyk et al., 2019). The Maximum Likelihood phylogeny (Figure 3), although presents weak backbone support, corroborates with the BI phylogeny, showing Pantanal strains in the Limnospira clade (boostrap 99), again related to that same Microcoleaceae genera. In both BI and ML trees Arthrospira clade is related to Microcoleus, Tychonema and Okeania.
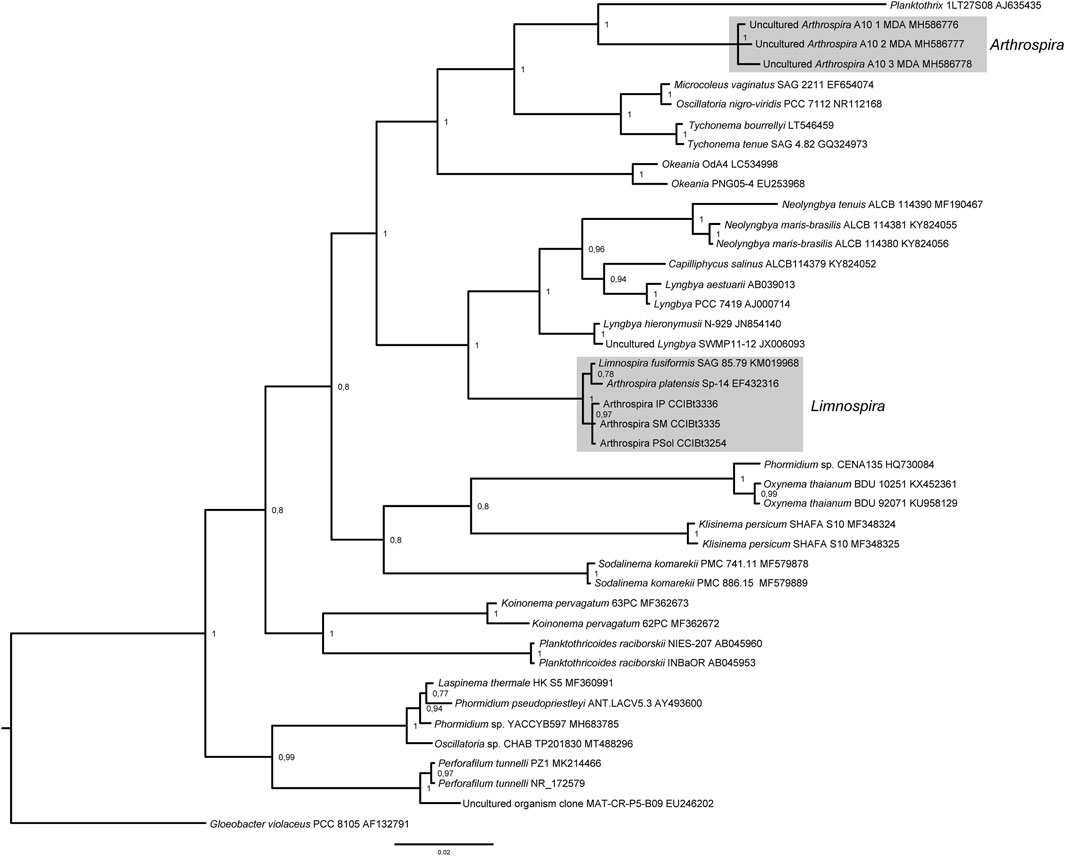
FIGURE 2. 16S rDNA Bayesian phylogenetic analysis. Posterior probabilities are indicated at nodes.
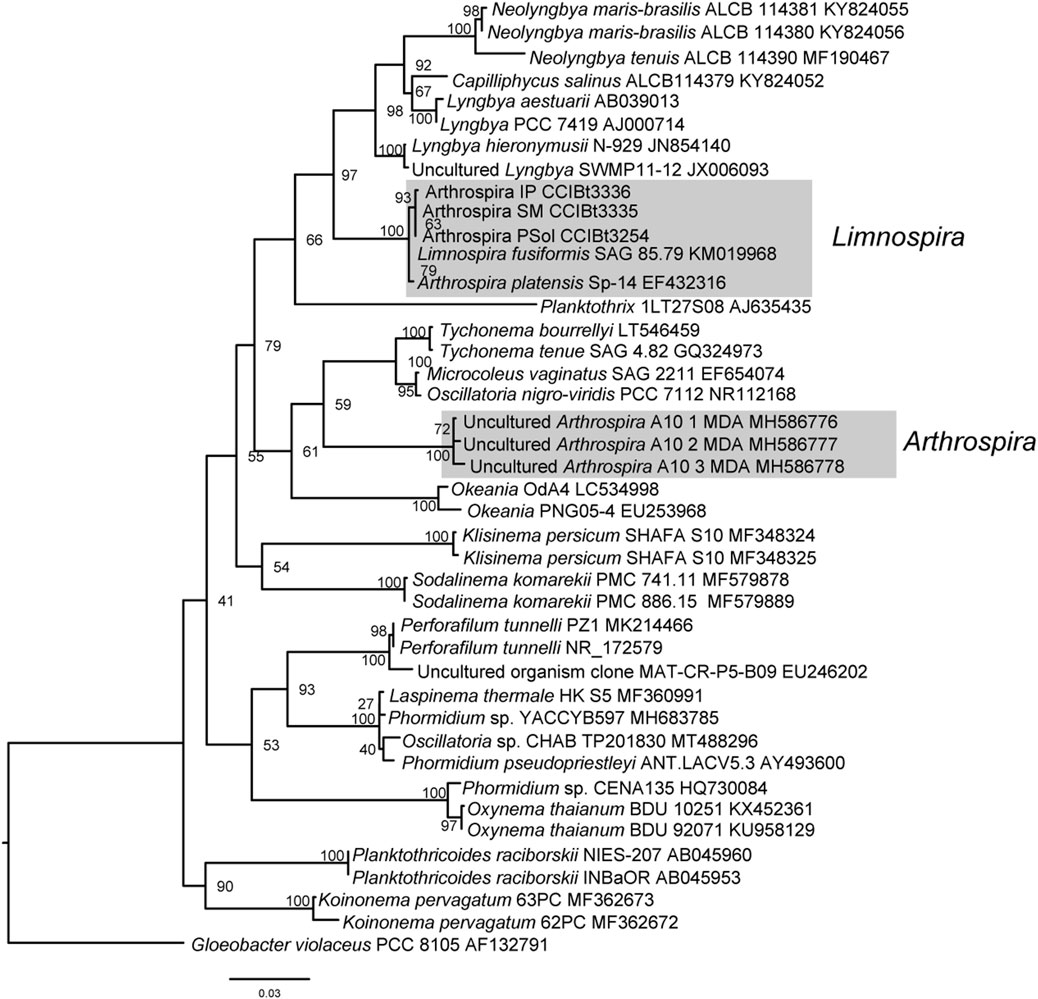
FIGURE 3. 16S rDNA Maximum Likelihood phylogenetic analysis. Bootstrap support is indicated at nodes.
Limnospira platensis (Gomont) K. R. S. Santos and G. S. Hentschke comb. nov. (Figures 4–6).
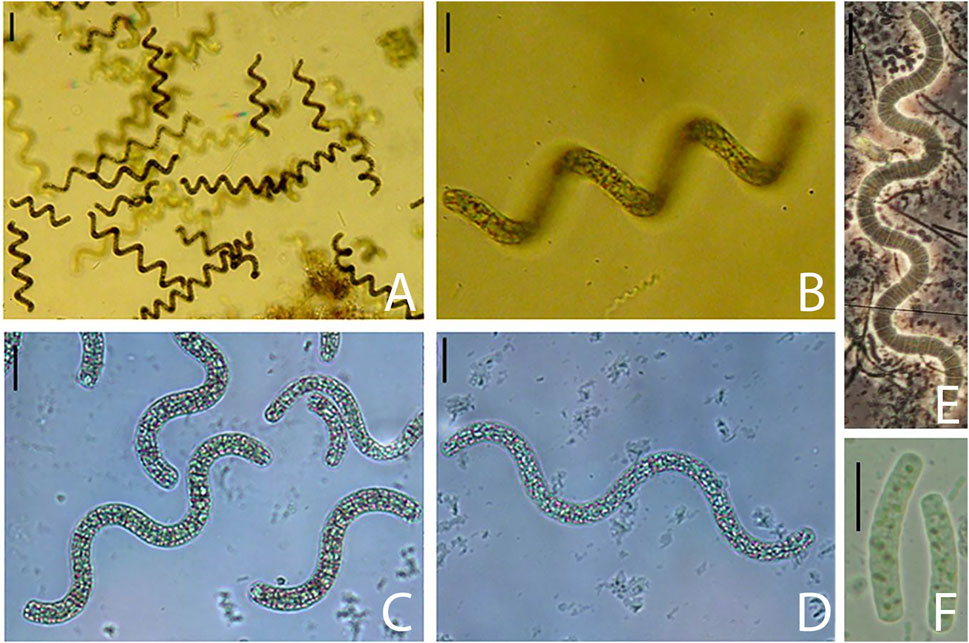
FIGURE 4. (A–D) General aspect of regularly coiled trichomes with aerotopes, (E) Trichome without aerotopes, (F) Hormogonia with granulose protoplasm and without aerotopes. Scales = 10 µm.
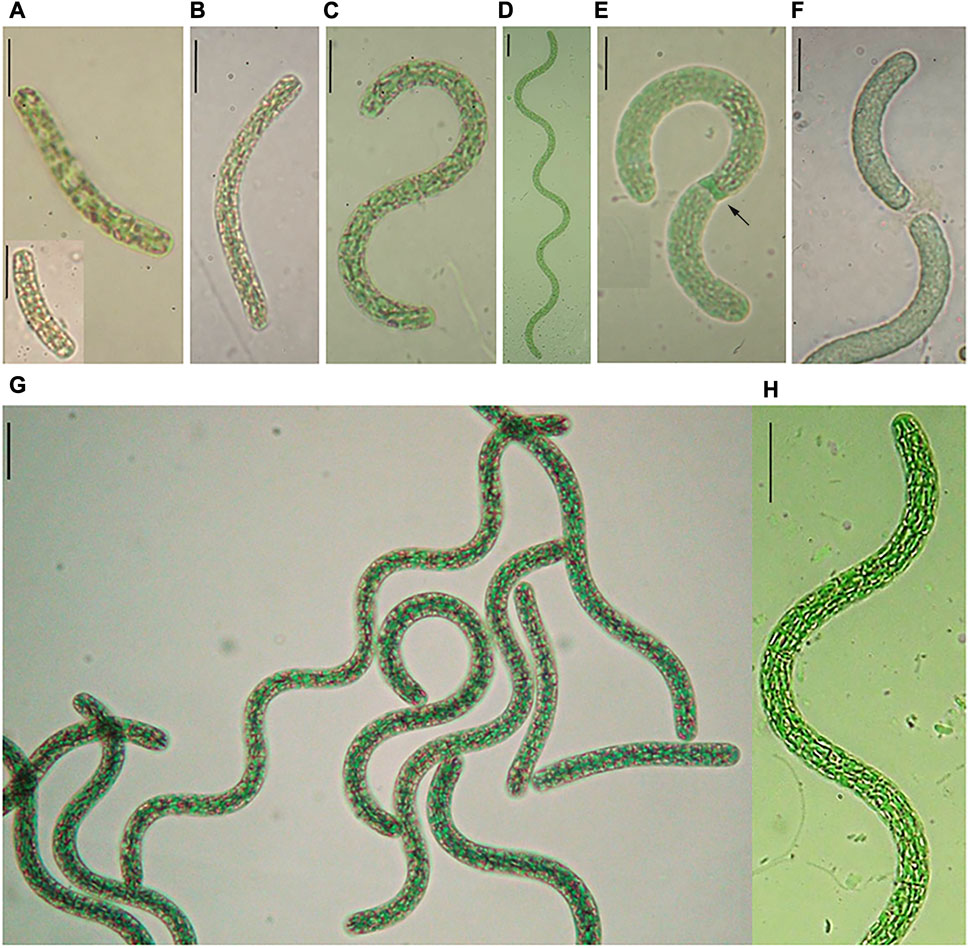
FIGURE 5. (A–F) Life cycle of L. platensis, (A) Hormogonia in nature (Salina do Meio), (B, C) Hormogonia in culture, (D) Adult trichome in culture, (E) Trichome with necridium (arrow), (F) Trichome broken at necridium, (G, H) Trichome in culture.
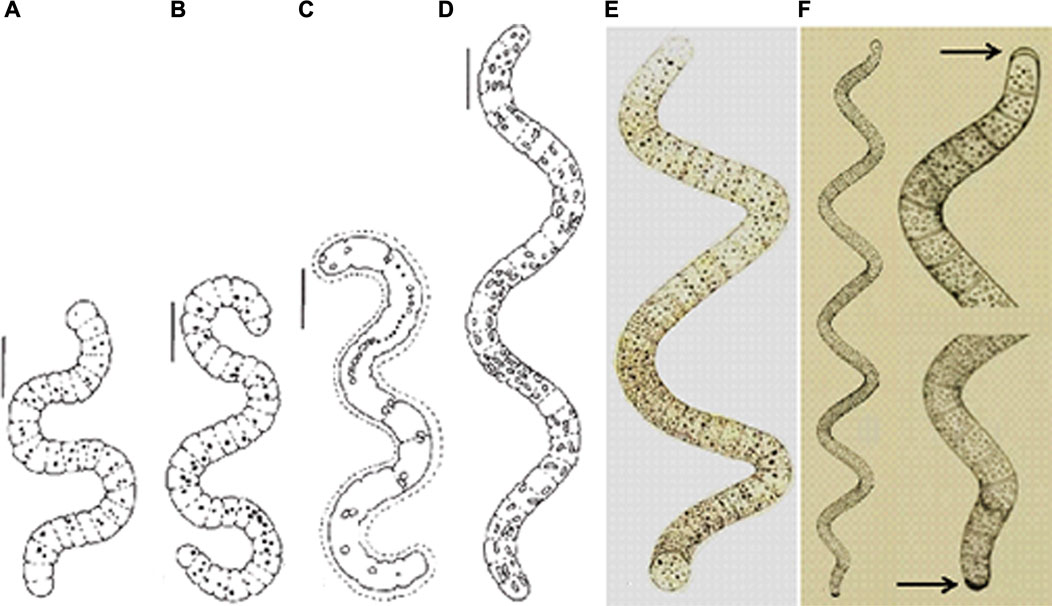
FIGURE 6. (A–D) Morphological variability of L. platensis from Pantanal da Nhecolândia, (A, B) Trichomes with granulose protoplasm, slightly widened and narrowed apexes, (C) Trichome with mucilaginous envelope (rare), (D) Regularly coiled trichome, with aerotopes (Santos and Sant’Anna, 2010), (E) Original illustration of A. platensis (iconotype) showing trichome with rounded terminal cells, without calyptra (Gomont, 1892, pl. 7, fig. 27), (F) Original illustration of A. maxima (iconotype) showing trichome with calyptra (arrows) (Gardner, 1917, pl. 33, Figure 3). Scales = 10 µm.
Basonym: A. platensis Gomont 1892: 247, pl. VII: fig. 27.
Published in: Gomont, M. (1892′1893′). Monographie des Oscillariées (Nostocacées Homocystées). Deuxième partie—Lyngbyées. Annales des Sciences Naturelles, Botanique, Série 7 16: 91-264, pls 1-7.
Trichomes solitary, regularly coiled when adult, constricted or not, 35-151 (−371) μm long, rarely with mucilaginous envelope. Spirals height 16–32.4 μm; distance between spirals 22–40.6 μm. Cells shorter than wide (rarely longer than wide), 1.7–5.3 (−9) μm long and 4–7.8 μm wide. Ratio L/W = 0.5-1.3. Apical cells rounded, slightly narrowed (rarely widened). Cell content blue-green, granulose, with facultative aerotopes.
Habitat: Saline and alkaline lakes, planktonic and more rarely benthic (part of the life cycle).
Reference strain: Arthrospira PSol CCIBt3254 (OR142672)
Studied material: BRAZIL. Mato Grosso do Sul: Corumbá, Pantanal da Nhecolândia, Salina do Meio, 09/25/2005, K.R.S. Santos and C.F.S. Malone (SP390917), 04/22/2006, K.R.S. Santos (SP390919), 08/28/2006, K.R.S. Santos (SP390922), 05/04/2007, K.R.S. Santos (SP390927), 08/19/2009, K.R.S. Santos (SP400654), 10/27/2011, K.R.S. Santos (SP427290), 05/06/2012, C.F.S. Malone and C.L. Sant’Anna (SP427740); Salina Pantanal, 08/16/2009, K.R.S. Santos (SP427747); Salina da Reserva, 08/19/2009, K.R.S. Santos (SP401692), 05/06/2012, C.F.S. Malone and C.L. Sant’Anna (SP427741); Salina da Ponta, 08/19/2009, K.R.S. Santos (SP401691); Salina Pedra do Sol, 08/26/2006, K.R.S. Santos (SP427742), 11/16/2006, K.R.S. Santos (SP427743.
Comments: L. platensis is morphologically different from the other species of the genus, L. fusiformis, L. indica and L. maxima. L. platensis is the only species of the genus without calyptra. Furthermore, L. fusiformis presents irregular spirals, while the spirals of L. platensis are regular. Although the presence of calyptra in L. maxima, this is the most similar species to A. platensis and the differences are discussed below. According to Komárek and Anagnostidis (2005), the relations between L. maxima and L. indica are unclear, and maybe these species are synonyms. When comparing our L. platensis cultures with the nature material, we did not find significative morphological differences between them.
L. platensis was found forming blooms with A. elenkinii Miller in saline and alkaline lakes in “Pantanal da Nhecolândia”. Table 1 shows all the studied samples and the respective physicochemical parameters. The species occurred in Pantanal saline lakes with pH ranging from 9.2 to 10.2, electrical conductivity from 716 to 19020 μS cm-1 and temperature from 22.8 to 33.3°C. In Lagoa Salitrada Campo Dora the species was registered only once, in the dry season (pH 8.4, electrical conductivity 1852 μS cm-1 and temperature 32°C). These data evidence the dependence of the alkaline pH, high electrical conductivity and warm temperatures for the growing of L. platensis. These conditions also indicate that high nutrient levels are fundamental for the species.
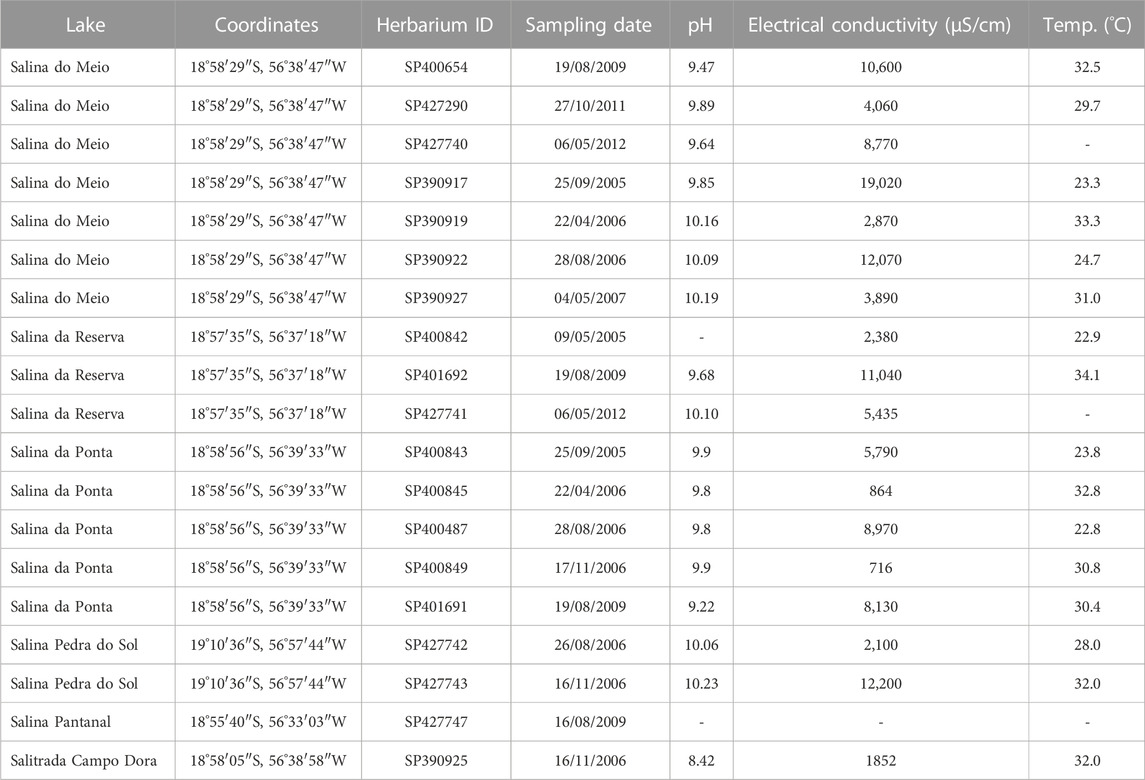
TABLE 1. Sampling location, geographical coordinates, Herbarium identification and physicochemical parameters of Pantanal da Nhecolândia lakes.
Arthrospira and Limnospira are monophyletic genera, morphologically distinguished from each other by the presence of aerotopes in Limnospira (Nowicka-Krawczyk et al., 2019). Aerotopes were commonly observed in our nature samples and isolate strains, and we also observed that this character was facultative in all or our samples and cultures. Apart of the facultative aerotopes, our specimens fit perfectly in the original description of A. platensis (Gomont, 1892). Figure 5 and Table 2 summarize the morphological similarity between our populations and A. platensis, and show the differences between this species and L. maxima, A. platensis presents smaller trichomes and the terminal cells lack calyptra, while L. maxima presents larger trichomes and presents calyptra at the terminal cells.

TABLE 2. Morphological characters of “A. platensis” and its most similar species A. maxima according to different authors.
According to the original description, based on an herbarium exsiccate, A. platensis presents cylindrical trichomes, not narrowed toward the ends or slightly narrowed, 6–8 μm wide; spiral height 26–36 μm, distance between spirals 43–57 μm; cells isodiametric 2–6 μm long with granulose protoplasm. The author does not mention the occurrence of aerotopes, nor the habits benthic or planktonic for this species described from La Plata Basin. A brief indication of the species habit is described as “greenish trichomes, forming a thin layered stratum” (free translation from Latin). These morphological and habit descriptions do not exclude the possibility of the presence of aerotopes and the planktonic habit, but make these characters uncertain, considering that the specimens analyzed were from dried material. The expression “thin layered stratum” does not specify where it was sampled and also, at that time, planktonic material was not analyzed, so the author didn´t know if specimens were also growing in phytoplankton. Consequently, we consider the original description poor and encompassing only a little portion of the morphological and ecological plasticity of the species.
Komárek and Lund (1990), also based on the type material in herbarium exsiccate, considered A. platensis as benthic and without aerotopes. However, to consider the species benthic based on the lack of aerotopes in dry preserved specimens is questionable. Currently it is known that aerotopes collapse in dry material (Tomaselli, 1997), and probably that is the reason why these authors (and Gomont) do not mention the presence of these structures.
Considering that, all of our studied populations from nineteen lakes, fit in the original description of A. platensis. All populations presented facultative aerotopes, and because of that, in this paper, we add this character to the species description. This is in agreement with Jeeji-Bai (1999) genus revision, which also report aerotopes for A. platensis. Moreover, the benthic habit was stated by (Komárek and Lund, 1990) as a consequence of the lack of aerotopes, however, in our observations it is clear that the populations are planktonic.
Our ecological data corroborates with the morphological analysis. Although Pantanal and La Plata Basin are geographically distant, they are connected by the rivers Paraguay and Paraná. Pantanal is a corridor connecting the Amazon and La Plata Basins, and during the rainy seasons, the lakes are connected to these basins. The rivers flow to the South carrying a great diversity of species to La Plata estuary, which presents also high salinity, as observed by us in Pantanal lakes (Alho and Gonçalves, 2005; Junk and Cunha, 2005). Another important fact that can explain the presence of A. platensis in La Plata Basin, is that many birds migrate from this area to Pantanal every year for reproduction, and can carry the microbiota in their feathers. It is impossible to know from which environment the original described population was sampled, but we hypothesize that it could be found in temporary small pools (forming stratum), with higher salinity, pH and/or temperature. Clearly, A. platensis is not typical of La Plata Basin, considering that after the description of this species in 1892, even with many sampling efforts of Argentinian and Uruguayan research groups, no populations of A. platensis were found in this region.
The fact that A. platensis was never reported for La Plata Basin after the original description indicates that this species is more related to the salines of Pantanal, due to its abundance and wide distribution in this Biome. According to our results, L. platensis is typical from alkaline, saline, nutrient rich and warm environments and this can explain why the species is rare in La Plata Basin. The species is reported worldwide, but its actual distribution is unknown, mainly because of misidentification of populations in Europe, America, Africa and Asia (Sili et al., 2012).
According to these statements and our phylogenetic analysis showing our strains in Limnospira clade, we conclude that A. platensis has to be combined to L. platensis. Also, we recommend to use Arthrospira PSol CCIBt3254 (OR142672) as the reference strain for the species.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: GenBank OR142670-OR142672.
KS (Article initiative, generated morphological data, described the species, collaborated in discussion section) GH (Article initiative, wrote the paper) GF (collaborated in discussion section) AA (generated molecular data) MF (generated molecular data) VV (collaborated in discussion section, revised the manuscript) CS’A (Author of the project, revised the manuscript). All authors contributed to the article and approved the submitted version.
GH is funded by Programa de Recuperação e Resiliência Português (PRR), Blue Bioeconomy Innovation Pact, Call 02/C05-I01/2022, Application Number C632741873-00467082. VV thanks to FCT Projects UIDB/04423/2020 and UIDP/04423/2020. KS thanks the financial supports of Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP—Process number 09/51655-1); the Embrapa Pantanal for permission to collect phytoplankton samples in Nhumirim farm, the Dr. Arnaldo Yoso Sakamoto (Universidade Federal de Mato Grosso do Sul—UFMS Campus TrêsLagoas) for the logistical support for the collections expeditions; the Freitag Laboratórios (Timbó/SC, Brazil) for supporting this publication. GF thanks to Laboratorio Tecnologico del Uruguay (LATU) for financial support.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Alho, C. J. R., and Gonçalves, H. C. (2005). Biodiversidade do pantanal: Ecologia and conservação. Campo Grande: UNIDERP.
Allem, A. C., and Valls, J. F. M. (1987). Recursos forrageiros nativos do Pantanal Mato-grossense. Brasília: Empresa Brasileira Agropecuária/Centro Nacional de Recursos Genéticos.
Birnboim, H. C., and Doly, J. (1979). A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 6, 1513–1523. doi:10.1093/nar/7.6.1513
Fiore, M. F., Moon, D. H., Tsai, S. M., Lee, H., and Trevors, J. T. (2000). Miniprep DNA isolation from unicellular and filamentous cyanobacteria. J. Microbiol. Methods 39, 159–169. doi:10.1016/s0167-7012(99)00110-4
Gomont, M. (1892). Monographie des Oscillariées (Nostocacées Homocystées). Deuxième partie. - lyngbyées. Annales des Sciences Naturelles. Botanique 7, 91–264.
Jeeji-Bai, N. (1999). “A taxonomic revision of the genus Arthrospira based on certain new criteria,” in Marine Cyanobacteria. Editors L. Charpy, and A. W. D. Larkum (Monaco: Bulletin of the Institute of Oceanography), 47–52.
Junk, W. J., and Cunha, C. N. (2005). Pantanal: A large South American wetland at a crossroads. Ecol. Eng. 24, 391–401. doi:10.1016/j.ecoleng.2004.11.012
Komárek, J., and Anagnostidis, K. (2005). “Cyanoprokaryota 2. Teil: Oscillatoriales,” in Süsswasserflora von Mitteleuropa 19. Editors B. Büdel, L. Krienitz, G. Gärtner, and M. Schagerl (Elsevier Spektrum Akademischer Verlag), 1–759.
Komárek, J., and Komárková-Legnerová, J. (2002). Contribution to the knowledge of planktic cyanoprokaryotes from central Mexico. Preslia 74, 207–233.
Komárek, J., and Lund, J. W. G. (1990). What is “Spirulina platensis” in fact? Arch. Hydrobiol. Suppl. Algol. Stud. 58, 1–13.
Lane, D. J. (1991). “16S/23S rRNA sequencing,” in Nucleic acid techniques in bacterial systematics. Editors E. Stackebrandt, and M. Goodfellow (Hoboken: Wiley and Sons), 115–175.
Lepère, C., Wilmotte, A., and Meyer, B. (2000). Molecular diversity of Microcystis strains (Cyanophyceae, Chroococcales) based on 16S rDNA sequences. Syst. Geogr. Plants 70, 275–283. doi:10.2307/3668646
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2010). “Creating the CIPRES science gateway for inference of large phylogenetic trees,” in Proceedings of the Gateway computing environments workshop (GCE), 1–8.
Neilan, B. A., Jacobs, D., Dot, T. D., Blackall, L. L., Hawkins, P. R., Cox, P. T., et al. (1997). rRNA sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus microcystis. International union of microbiological societies. Available at: www.microbiologyresearch.org.
Nowicka-Krawczyk, P., Mühlsteinová, R., and Hauer, T. (2019). Detailed characterization of the Arthrospira type species separating commercially grown taxa into the new genus Limnospira (Cyanobacteria). Sci. Rep. 9, 694. doi:10.1038/s41598-018-36831-0
Ronquist, F., Teslenko, M., Van Der Mark, P., Ayres, D. L., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi:10.1093/sysbio/sys029
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular cloning: A laboratory manual. 2nd ed. Cold Harbor: Cold Spring Harbor Laboratory Press.
Santos, K. R. S., Jacinavicius, F. R., and Sant’Anna, C. L. (2011). Effects of the pH on growth and morphology of Anabaenopsis elenkinii Miller (Cyanobacteria) isolated from the alkaline shallow lake of the Brazilian Pantanal. Fottea 11 (1), 119–126. doi:10.5507/fot.2011.012
Santos, K. R. S., and Sant’Anna, C. L. (2010). Cianobactérias de diferentes tipos de lagoas (“salina”, “salitrada” e “baía”) representativas do Pantanal da Nhecolândia, MS, Brasil. Rev. Bras. Botânica 33, 61–83. doi:10.1590/s0100-84042010000100007
Sili, C., Giuseppe, T., and Vonshak, A. (2012). “Arthrospira (Spirulina),” in Ecology of Cyanobacteria II: Their diversity in space and time. Editor B. A. Whitton (Durham: School of Biological and Biomedical Sciences, Durham University), 677–705.
Stanier, R. Y., Kunisawa, R., Mandel, M., and Cohen-Bazire, G. (1971). Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 35, 171–205. doi:10.1128/br.35.2.171-205.1971
Tamura, K., Stecher, G., and Kumar, S. (2021). MEGA11: Molecular evolutionary Genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027. doi:10.1093/molbev/msab120
Tomaselli, L. (1997). “Morphology, ultrastructure and taxonomy of Arthrospira (Spirulina) maxima and Arthrospira (Spirulina) platensis,” in Spirulina platensis (Arthrospira): Physiology, cell-biology and biotechnology. Editor A. Vonshak (London: Taylor and Francis), 79–99.
Keywords: salines, extreme environments, biodiversity, polyphasic approach, new combination
Citation: Santos KRdS, Hentschke GS, Ferrari G, Andreote APD, Fiore MdF, Vasconcelos V and Sant’Anna CL (2023) Molecular, morphological and ecological studies of Limnospira platensis (Cyanobacteria), from saline and alkaline lakes, Pantanal Biome, Brazil. Front. Environ. Sci. 11:1204787. doi: 10.3389/fenvs.2023.1204787
Received: 12 April 2023; Accepted: 07 September 2023;
Published: 25 September 2023.
Edited by:
Allan Douglas Cembella, Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research (AWI), GermanyReviewed by:
Prashant Kumar Singh, Mizoram University, IndiaCopyright © 2023 Santos, Hentschke, Ferrari, Andreote, Fiore, Vasconcelos and Sant’Anna. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guilherme Scotta Hentschke, Z3VpbGhlcm1lLnNjb3R0YUBnbWFpbC5jb20=; Kleber Renan de Souza Santos, c2FudG9za3JzQGdtYWlsLmNvbQ==
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.