 Nana R. Diakité1*
Nana R. Diakité1* Patricia B. Koffi1
Patricia B. Koffi1 Cyrille K. Konan1Fidèle K. Bassa1
Cyrille K. Konan1Fidèle K. Bassa1 Andrew J. Chamberlin2Mamadou Ouattara1
Andrew J. Chamberlin2Mamadou Ouattara1 Giulio A. De Leo2
Giulio A. De Leo2 Eliezer K. N’Goran1
Eliezer K. N’Goran1- 1Laboratoire de Biologie et Santé, UFR Biosciences, Université Félix Houphouët-Boigny, Abidjan, Côte d’Ivoire
- 2Hopkins Marine Station, Stanford University, Pacific Grove, CA, United States
Background: Schistosomiasis is endemic throughout all regions of Côte d’Ivoire, however, species of the intermediate snail host vary across bioclimatic zones. Hence, a deeper knowledge of the influence of climatic on the life history traits of the intermediate snail host is crucial to understand the environmental determinants of schistosomiasis in a rapidly changing climate. The aim of this study was to run a common garden experiment to assess differences in survival, somatic growth and fecundity of both Bulinus truncatus and Biomphalaria pfeifferi snails collected in three different bioclimatic areas.
Methods: A cross-sectional malacological survey was conducted in February 2021 in the south, center and north of Côte d’Ivoire. We sampled two populations of B. truncatus, the intermediate host snail of Schistosoma haematobium, from northern and central Côte d’Ivoire, and two populations of Bi. pfeifferi, the intermediate host snail for Schistosoma mansoni, from the southern and central regions. Snails collected at the human-water contact sites were brought in the laboratory where they reproduced. The first generation snails (G1) for each population were reared under the same laboratory conditions, i.e., at 24°C–26°C, during 63 days (9 weeks), to estimate survival, growth, and fecundity.
Results: We found that G1 Bulinus snails from the north population showed higher survival and growth rates during our study and higher number of eggs at first reproduction, compared to the ones from the central region. For Bi. pfeifferi, no significant difference in survival rate was observed between G1 snails from the southern and central populations, whereas those from the south exhibited higher growth rates and higher number of eggs per individual at first reproduction than G1 snails from the central population.
Conclusion: Our study provides evidence for heterogeneity in snails’ life-history traits in response to temperature among the populations from the three climatic regions. Further experiments from multiple populations are needed to confirm that snails express traits under optimal conditions, can lead to expansion of their geographical range and hence an increase in the risk of schistosomiasis transmission. Transplantation experiments will be required to assess implications of the changing climate on snails persistence, distribution and abundance.
Introduction
Schistosomiasis is a Neglected Tropical Disease (NTD) that remains a significant global public health problem. Human schistosomiasis causes the second-highest burden of parasitic disease in the world, after malaria (Global Health & Division of Parasitic Diseases and Malaria, 2018). More than 90% of cases occur in Africa and at least 236.6 million people required preventive treatment in 2019 (WHO, 2022). Human infection is caused by six species of trematodes, but the predominant species in Africa are S. haematobium (eggs excreted in the urine) and S. mansoni (eggs excreted in the feces). Schistosome parasites require two obligate hosts to complete their life cycle: specific freshwater snail species of the genus Biomphalaria for S. mansoni and Bulinus for S. haematobium, where parasites reproduce asexually, and a human host (both parasites) or a small mammalian (S. mansoni) where parasites reproduce sexually. Adult parasites lodge in the blood vessels of the definitive mammalian host where they produce eggs that are passed through the urine (for S. haematobium) or the stool (for S. mansoni) of infected people (Nelwan, 2019). Once the eggs are exposed to water, miracidia, one of the two free-living, swimming stages of the parasite, hatch from the eggs and then search an intermediate host snail to penetrate. 3 to 7 weeks later, depending upon water temperature and snail species, infected snails start shedding cercariae, the other free-living stage of the parasite. Cercariae try to penetrate the skin of a mammalian host stepping into the water (Colley et al., 2014).
The obligate snail host thus play a crucial role in the transmission dynamics of schistosomiasis (Allan et al., 2020). Heterogeneity in snail life history and demographic traits may affect parasite transmission in multiple ways. For instance, snail susceptibility to infection decreases with age (Anderson et al., 2021) and, while cercarial shedding rate increases with snail body size, the lifetime production of cercariae increases with snail’s survival rate (Niemann and Lewis, 1990). Therefore, variations in snail life history traits may have implications in the transmission of both human and livestock Schistosoma parasite (Tchuenté and N’Goran, 2009; Webster et al., 2007; Moné et al., 2012; Boon et al., 2018). To better understand how ecological determinants affect schistosomiasis prevalence in humans, it is important to investigate variation in snail life history traits that can influence schistosomiasis transmission across different geographical range.
In Côte d’Ivoire, both intestinal (S. mansoni) and urogenital (S. haematobium) schistosomiasis are endemic. They are transmitted by Biomphalaria pfeifferi and by Bulinus globosus and B. truncatus, respectively (N’Goran, 1987; Tchuenté and N’Goran, 2009). A study carried out in the north and central parts of Côte d’Ivoire has shown that the genus Bulinus transmits not only S. haematobium, but also S. bovis and the hybrid between S. haematobium and S. bovis (Tian Bi et al., 2019; Campbell et al., 2020). Disease and snails are present in Côte d’Ivoire throughout a wide geographical region, however, species and population traits vary across bioclimatic zones (Assaré et al., 2015). The present study hypothesized that important life history traits of the intermediate host snails vary geographically according to climatic zones in Côte d’Ivoire (Altizer et al., 2013; Kalinda et al., 2018). Here we present the results of a common garden experiment to variation in survival, fecundity, and growth rates in response to temperature of first-generation snails from two populations of B. truncatus sampled in north and center, and two populations of Bi. pfeifferi from the south and center of Côte d’Ivoire.
Materials and methods
Study area
Snail sampling was carried out at three sites, each corresponding to a different bioclimatic zone in Côte d’Ivoire. In the northern department of Ouangolodougou samples were collected in Torla (9°35′00′N latitude, 5°11′00′ W longitude), in the central department of Bouaké in Raffierkro (07°34′53′N latitude, 05°01′26′W longitude) and in the southern department of Adzopé in Diasson (6°06′25′N latitude, 3°51′36′W longitude) (Figure 1). Previous work (Diakité et al., 2018; Tian Bi et al., 2019) showed that S. haematobium is endemic in the south-eastern, northern and central sites, whereas S. mansoni is endemic in the central and southern sites (Ouattara et al., 2021). Accordingly, B. truncatus and B. forskalii were mainly observed in the north and the center and B. globosus and Bi. pfeifferi were found in the central and the southern sites.
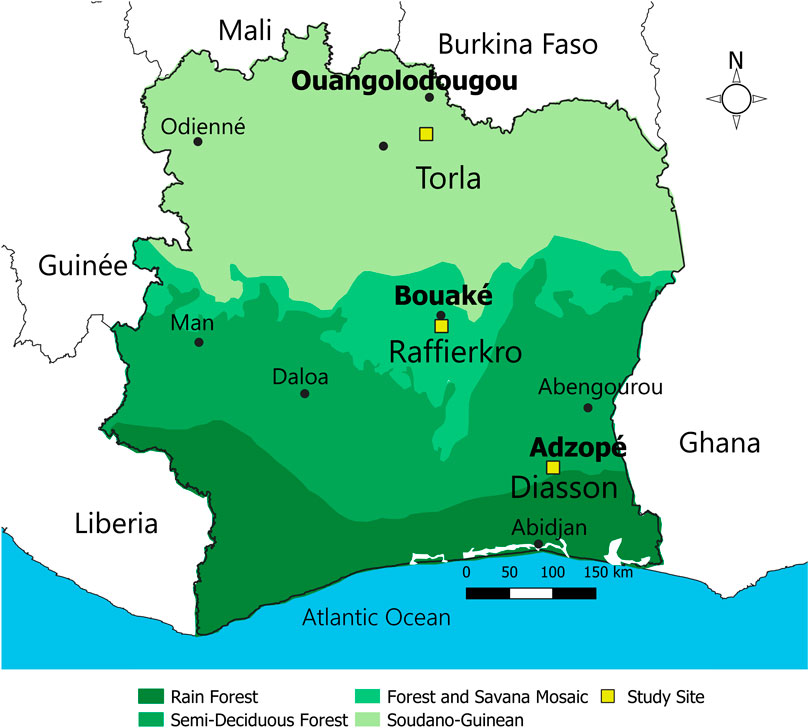
FIGURE 1. Map of Cote d’ Ivoire showing the bioclimatic zones and study sites (Source: https://faolex.fao.org/docs/pdf/Ivc186335.pdf 23-04-24.
The northern zone is characterized by Soudanean climate with two well-defined seasons: the dry season, which lasts from November to March, and the rainy season from April to October. The mean air temperature ranges from 24.2°C to 30.8°C (mean: 27.6°C) and the mean annual precipitation is about 1,000 mm (https://fr.climate-data.org/afrique/cote-d-ivoire). Several small multipurpose dams, constructed to enhance agricultural production and livestock rearing characterize the northern and central parts (Tian bi et al., 2018; Diakité et al., 2021).
The central pre-forest zone is an intermediate climatic zone between the north and the south and is characterized by two seasons: a rainy season from March to October and a dry season the remaining months of the year. The annual average temperature ranges between 24.2°C and 28°C (mean/26.2°C) and the mean annual precipitation is 1,373 mm. The vegetation consists of savannah and forest and there are streams, large and small dams, and semi-permanent rivers.
The southern zone is characterized by a hot and humid climate, with temperature ranging from 24.4°C to 27.2°C (mean: 25.7°C). The average annual precipitation is 1,529 mm and there are four seasons: a short rainy season from September to November, a long dry season from December to March, a long rainy season from April to mid-July and a short dry season from mid-July to August. The vegetation is dense tropical rain forest and streams and many permanent rivers drain this area.
Snail sampling and identification
A cross-sectional survey was conducted to collect snails during the dry season in February 2021, in waterbodies known to harbor these snail species of the three localities: small dams in Torla (north) and Raffierkro (center) and a river in Diasson (south). The sites sampled were permanent water bodies, which have been previously investigated several times throughout the seasons (Tian Bi et al., 2019). Two field collectors sampled snails using scoops and forceps. The snails were identified based on morphological characteristics and a key for snail identification (Danish Bilharziasis Laboratory, 1980; Brown, 1994). Bulinus truncatus population of the north was previously identified by using molecular barcoding (Tian Bi et al., 2019). The specimens collected were transferred to water-soaked cotton in perforated plastic boxes, put in a cooler at about 20°C and transferred to a laboratory at the university Félix Houphouët-Boigny.
Snail rearing
In the laboratory, all intermediate snail host of schistosomiasis underwent the cercarial shedding test (Frandsen and Christensen, 1984). Each snail was put in a glass tube containing 5 mL of mineral water and was exposed to artificial light for 2–4 h. Then, each tube was checked under a stereomicroscope for the presence of cercariae. After several tests over a month, uninfected snails were selected for breeding (Gérard and Théron, 1997). The breeding room temperature was set at 24°C–26°C and the average humidity at 78.59%. We selected specimens of Bi. pfeifferi from the south and center sites and specimens of B. truncatus from the center and north sites. The four snail populations were reared following a common garden approach. Snails sampled from each region, (here referred to as generation G0) were placed together in a 1.5 L transparent plastic tank for acclimatization. Twelve days after, 25 G0 snails randomly chosen from the population were measured (apex to aperture or diameter) using an electronic caliper. To avoid a mixing juvenile (immature) and adult (mature) snails, we selected snails with size equal or larger than 6 mm. Each G0 snail was put in a transparent plastic box containers with 30 mL of mineral water. The snails were fed aquarium fish pellets (one granule per adult snail) and dried or boiled lettuce for juveniles. The water was refreshed twice a week. One week after each individual G0 snail laid eggs, egg capsules were scraped off the box using a plastic spatula and counted under a binocular stereomicroscope. The eggs were then placed in petri dishes containing 20 mL of mineral water, where the eggs hatched and formed the first generation (noted G1). After 14 days (2 weeks), two or four G1 juveniles randomly selected from each of the parental G0 petri dishes were measured using a graph paper (width: 5 cm and length: 5 cm). In total, 50 G1 snails per population were used for further laboratory experiments. G1 juveniles were reared in individual 5 mL glass containers with mineral water which was refreshed weekly and fed boiled lettuce.
Life cycle parameters
Life-history traits (growth, survival, and fecundity) of G1 snails were monitored over 63 days (9 weeks). Each snail was carefully moved using a plastic spatula to an absorbent paper to remove excess water. The size of the shell when the snail laid its first clutch of eggs (size at first reproduction) was measured using a digital caliper. The distance between the apex and the aperture of B. truncatus and the diameter of the shell of Bi. pfeifferi were measured.
The mortality rate of each species was evaluated weekly. The death of an individual was confirmed by observation of one of the following criteria: flesh shrinking in its shell or completely out of the shell, inability to climb the wall of the container, or remaining inside the shell for several days.
The fecundity was assessed weekly by counting the number of capsules per individual, the number of eggs per capsule, the number of eggs per individual and the number of juveniles after hatching. At the end of each week, the egg capsules were removed from the glass; the eggs were counted and put in a new petri dish containing mineral water. Egg capsules were checked twice a day, and the number of hatched individuals (G2 generation) was recorded in order to evaluate the hatching rate per week for each population.
Statistical analysis
Data were entered in an Excel 2016 software spreadsheet (Microsoft Corporation; Redmond, Washington, United States). All analyses were conducted using R Statistical Software (ver. 3.6.3 R core Team, 2020) and R Studio (Rstudio Team, 2020). Region effects on survival of populations were tested using β the Cox regression coefficient. Exp (β) represents the hazard ratio (HR) or risk of death. HR = 0: No effect, HR < 1: Reduction in the hazard, HR > 1: Increase in the hazard regression. The Generalized Linear Model (GLM) tests was used to compare life cycle parameters between populations. Survival rates between B. truncatus and Bi. pfeifferi populations were compared using Cox regression with R package. Age, height, and width at first reproduction as well as growth parameters during the 7 weeks of follow-up were analyzed assuming a Gaussian distribution of error terms. Fecundity at first reproduction and mean fecundity (mean number of eggs over the study time) were analyzed following a Poisson distribution. The hatching rate of individuals was analyzed according to a binomial distribution. The significance threshold (α) for the statistical tests was set at 0.05.
Results
A total of 434 freshwater snails were collected from the study sites. Identification based on shell morphology revealed seven genera. The two species known to be intermediate hosts of schistosomiasis were B. truncatus and Bi. pfeifferi. Out of 212 B. truncatus snails collected, 118 (55.6%) were sampled at the center site and 94 (44.4%) at the northern site. A total of 171 Bi. pfeifferi were sampled, 119 (69.6%) were collected in central Côte d’Ivoire and 52 (30.4%) in the south. The cercariae emergence test revealed a very low infection rate, 1.31% for schistosomes and 1.04% for the others trematodes (xiphidiocercariea, and strigeid cercariae).
Growth of snail’s populations
There was no significant difference in shell height size between G1 bullinid snails from the north and central sites at the first week (center population mean size 2.10 ± 0.18 mm, north population mean size 2.11 ± 0.25 mm, p = 0.805). However, shell width was greater in the central population than in the northern (p < 0.001). At the seventh week of follow-up, shell height in the northern populations was significantly greater than that of the central (5.51 ± 0.34 mm versus 5.31 ± 0.33 mm, p = 0.008) (Table 1). For Bi. pfeifferi, there was a significant size difference (diameter) between G1 snails from the central and southern populations at the first and seventh weeks. The mean size was 1.76 ± 0.21 mm in the south versus 1.5 ± 0.20 mm in the central population at week 1, and 5.12 ± 0.51 mm versus 4.29 ± 0.57 mm, respectively, at week 7 (GLM, p < 0.001) (Table 2).
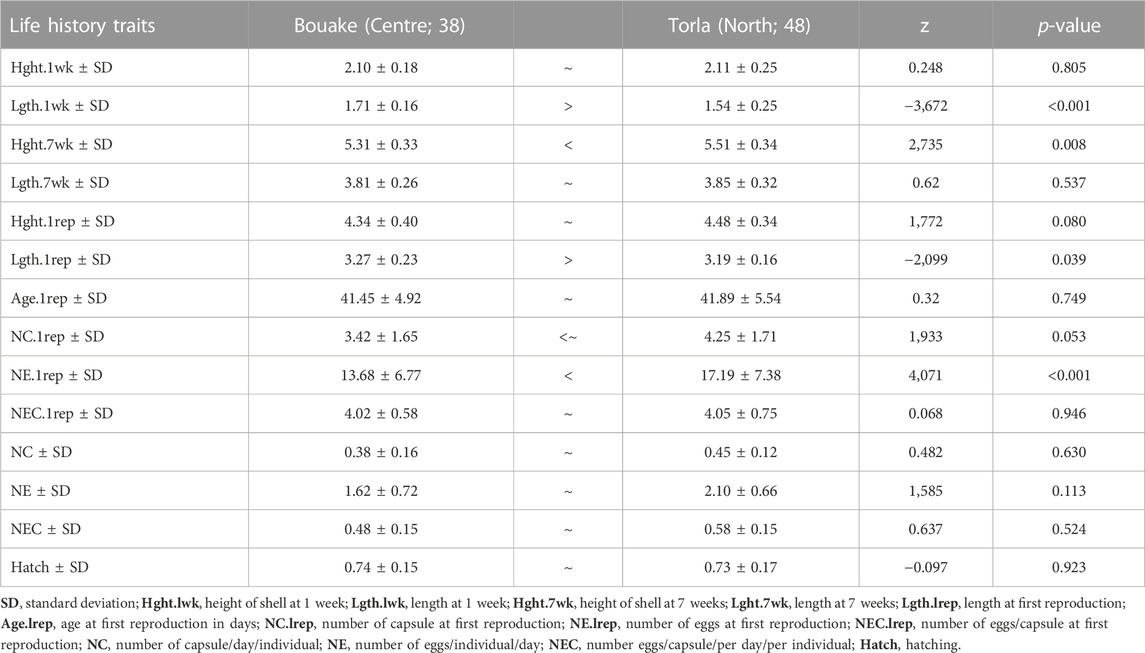
TABLE 1. Growth rate and the reproductive output of two populations of Bulinus truncatus.
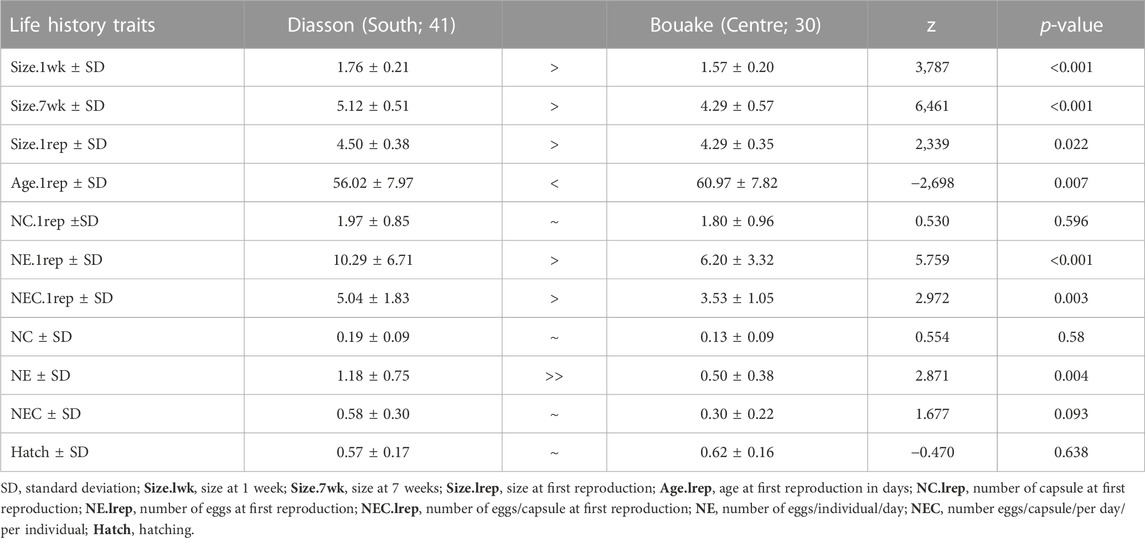
TABLE 2. Growth rate and reproductive output of two Biomphalaria pfeifferi snail populations.
Fecundity and egg hatching
For B. truncatus, the mean number of eggs at first reproduction was the only parameter significantly different between the two G1 populations (Table 1), with northern snails shedding more eggs than central snails (17.19 ± 7.38 versus 13.68 ± 6.77, respectively, p < 0.001). For Bi. pfeifferi, reproduction parameters, namely, age, number of capsules and eggs at the first reproduction and the number of eggs per individual, were different between snails from the central and southern regions (Table 2). Age at the first reproduction and average size for Bi. pfeifferi snails from the south was 56 ± 7.97 days and 4.5 ± 0.38 mm versus 61 ± 7.82 days and 4.29 ± 0.35 mm for snails from the central population, p < 0.001.
Survival of populations
The survival rate during the 7 weeks of the experiment varied by population, climatic zone and species. Survival was 100% in the populations of B. truncatus in the first 38 days, but started decreasing on day 39, and fell to 98% and 76% in the northern and central populations, respectively, on day 60, at the end of the experiment (Figure 2A). Similarly, for the populations of Bi. pfeifferi, the survival rate was 100% during the first 33 days and started decreasing on day 34, ultimately falling to 92% and 86% in the central and southern populations, respectively (Figure 2B). The analyses showed that the effect of the climatic zone on mortality was significantly higher in the central versus the northern populations of B. truncatus (Hazard Ratio HR = 13.12, p = 0.013) presenting a risk of mortality 0.08 times lower in the north than in the center. However, for Bi. pfeifferi no difference in survival was observed.
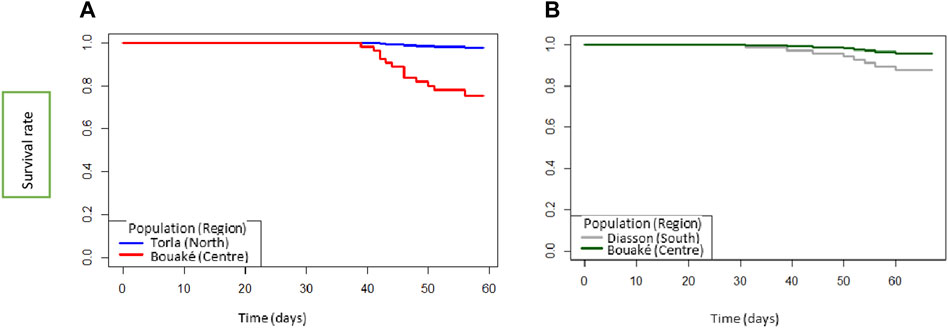
FIGURE 2. Surival curves of (A) Bulinus truncatus populations collected in the north and center and (B) Biomphalaria pfeifferi from the south and the center of Côte d’ Ivoire.
Discussion
In this study we sampled populations of intermediate host snails of Schistosoma from three different climatic regions in Côte d’Ivoire to determine their response to temperature. Specifically, we analyzed variations in survival, growth rates and reproduction parameters of the first generation (G1) snails from populations sampled over a north-to-south geographical gradient, corresponding to a shift from more arid to more humid climate. Specifically, we sampled two populations of B. truncatus snails, one from the north, characterized by an arid climate, and the other from the center, as well as two populations of Bi. pfeifferi, one from center and the other from south, characterized by a more humid climate.
Survival and growth rates
The results of our common garden experiments showed a general decrease in G1 bullinid snails’ survival on a north-to-south gradient but no difference in the survival rate of Bi. pfeifferi from the south and the center. Bulinus snails from the northern region were larger and grew faster than those sampled in central Côte d’Ivoire. Similarly, Biomphalaria snails sampled in the southern region were larger in size and grew faster than Biomphalaria snails sampled in the central region. If this observation is confirmed by further studies using several populations of the same climatic area, that will support the hypothesis of bullinid snails adaptation to local climatic conditions (Viard et al., 1996). Specifically, when exposed to the same climatic conditions of the common garden experiment, bullinid snail populations in the northern region, experiencing harsher climate historically (i.e., more arid climate with extreme temperatures) exhibited increased survival compared to snails from the central populations.
Reproduction parameters of the first generation of snails
Biomphalaria pfeifferi populations of the south were significantly larger and more fertile than the corresponding population from the center. Previous laboratory experiments (Harrison and Shiff, 1966; Kalinda et al., 2017b) have shown that the optimum temperature for snail growth occurs at around 25°C–26°C. Age at first reproduction for both Bi. pfeifferi populations was similar to that reported by El-Hasan (1974) for Bi. Alexandrina in a laboratory experiments at 25°C. However, it is lower than previously reported for Bi. pfeifferi snails from Man, Western Côte d’Ivoire (80–103 days), which were reared in laboratory conditions at 22°C–24°C (Tian Bi et al., 2013). Similarly, B. truncatus from the north and the center were younger at first reproduction (56 and 61 days, respectively), which is lower than that reported by Konan et al. (2022) for snails from the same region reared in a laboratory at 22°C–24°C (69 and 87 days). It is also lower than the age at first reproduction reported for the same species reared at 25°C by El-Hassan (1974). Other studies have shown a marked variation among freshwater snail populations in regards to age, size and fecundity at first reproduction (Evanno et al., 2006; Chapuis et al., 2007; Tian Bi et al., 2013).
In general, we found that G1 Bi. pfeifferi snails from the south grew larger and faster and had greater per capita daily fecundity than snails from the central Côte d’Ivoire population when both populations were reared in laboratory conditions at 24–26°C. We also found that, under the same condition, G1 B. truncatus snails from the northern Côte d’Ivoire grew larger and faster and suffered lower mortality, than G1 snails from the central Côte d’Ivoire. It is generally known that populations living in different climates and environments may evolve different life-history strategies. Theoretical models have predicted, for example, that organisms living in fluctuating environments should reproduce earlier, at a smaller size and produce larger clutch sizes compared to those living in stable habitats (Reznick et al., 2002). Contrary to the above-mentioned prediction, B. truncatus snails from the north, where climatic conditions are harsher, matured at a larger size/age and laid more eggs at first reproduction compared to the central population. One explanation is that size selects for resistance to the stressors (Facon et al., 2004; Chapuis et al., 2012) and a larger size is known to reduce water loss by conferring a better surface-volume ratio (Facon et al., 2004). Selection might therefore, delay the onset of reproduction to a less fragile stage. Other selective factors are parasites or predators and availability of nutritive resources. For example, fish breeding and fish predation may accelerate reproduction onset. Also, habitat openness (open versus closed) and season (end of dry vs. rainy season) has been shown to affect snail population size, migration and mode of colonization (Charbonnel et al., 2002a).
The results of our study highlight that temperature may potentially influence key life history traits, which in turn may affect the distribution, abundance and persistence of freshwater snails, including the obligate intermediate hosts for the parasite causing human schistosomiasis. Our common garden experiment provides supportive evidence that the observed heterogeneities in life history traits might have a genetic basis. Further studies are needed to assess whether the observed heterogeneity in the life history traits are an adaptation to different climatic zones, or simply the result of genetic drift from seasonal population fluctuations, which lead to population bottlenecks. While we believe that genetic drift is unlikely, it cannot be ruled out without the proper experiments. Accordingly, future experiments are needed to cast a light on this crucial issue and should include multiple independent populations sampled in each climatic zones. Whatever the cause is, our common garden experiment provides supporting evidence of variability in hereditable life history traits in populations from different climatic zones, which may have important consequences for schistosomiasis transmission in regards to change climate. Future studies using thermal-sensitive, process-based, mechanistic models of snail-parasite dynamics can clarify how this variability in snail persistence, abundance, and distribution might ultimately influence schistosomiasis transmission risk to the human population.
Conclusion
This study highlights the variability according to the bioclimatic areas of survival, somatic growth and fecundity of intermediate host snail of S. haematobium and S. mansoni, collected in three bioclimatic areas of Côte d’Ivoire. Firstly, the B. truncatus populations from the northern, more arid climatic zone exhibited higher survival than those of central Côte d’Ivoire. Secondly, Biomphalaria pfeifferi from the South were larger, matured faster and had higher fecundity than the population from central Côte d’Ivoire. Understanding the variations of basic life history traits of medically important snails across different ecozones can provide valuable information to improve our understanding of snail population dynamics associated with specific climatic regimes, and, ultimately, to improve efforts to control and, possibly, eliminate schistosomiasis.
Data availability statement
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Author contributions
ND, PK, MO, and EN’G conceived and validated the methodology of the study. CK and PK collected the data. CK performed statistical analysis. The first draft was writing by ND. The manuscript was revised by FB, AC, GD, and EN’G. All authors contributed to the article and approved the submitted version.
Funding
This study was funded by the Belmont Collaborative Forum on Climate, Environment and Health (Award no. 19/23593). ND, PK, MO, EN’G, and CK were funded by the “Programme d’Appui Stratégique à la Recherche Scientifique en Côte d’Ivoire (PASRES).” De Leo and Chamberlin where partially supported by the Belmont Collaborative Forum for Climate, Environment and Health and the US National Science Foundation (ICER-2024383 and DEB-2011179).
Acknowledgments
We gratefully acknowledge Jennifer Giovanoli Evack for having read the manuscript. We also thank all the village leaders for their collaboration in sampling snail populations.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2023.1193239/full#supplementary-material
References
Allan, F., Ame, S. M., Tian-Bi, Y. T., Hofkin, B. V., Webster, B. L., Diakité, N. R., et al. (2020). Snail-related contributions from the schistosomiasis consortium for operational research and evaluation program including xenomonitoring, focal mollusciciding, biological control, and modeling. Am. J. Trop. Med. Hyg. 103 (1), 66–79. doi:10.4269/ajtmh.19-0831
Altizer, S., Ostfeld, R. S., Johnson, P. T., Kutz, S., and Harvell, C. D. (2013). Climate change and infectious diseases: from evidence to a predictive framework. Science 341 (6145), 514–519. doi:10.1126/science.1239401
Anderson, L. C., Loker, E. S., and Wearing, H. J. (2021). Modeling schistosomiasis transmission: the importance of snail population structure. Parasites Vectors 14, 94. doi:10.1186/s13071-021-04587-8
Assaré, R. K., Lai, Y. S., Yapi, A., Tian-Bi, Y. N., Ouattara, M., Yao, P. K., et al. (2015). The spatial distribution of Schistosoma mansoni infection in four regions of western Côte d'Ivoire. Geospat Health 10 (1), 345. doi:10.4081/gh.2015.345
Boon, N. A. M., Van Den Broeck, F., Faye, D., Volckaert, F. A. M., Mboup, S., Polman, K., et al. (2018). Barcoding hybrids: heterogeneous distribution of Schistosoma haematobium × Schistosoma bovis hybrids across the Senegal River Basin. Parasitology 145 (5), 634–645. doi:10.1017/S0031182018000525
Brown, D. S. (1994). Freshwater snails of Africa and their medical importance. London: Taylor & Franch.
Campbell, C. H., Binder, S., King, C. H., Knopp, S., Rollinson, D., Person, B., et al. (2020). SCORE operational research on moving toward interruption of schistosomiasis transmission. Am. J. Trop. Med. Hyg. 103 (1_Suppl. l), 58–65. doi:10.4269/ajtmh.19-0825
Chapuis, E., and Ferdy, J. (2012). Life history traits variation in heterogeneous environment: the case of a freshwater snail resistance to pond drying. Ecol. Evol. 2 (1), 218–226. doi:10.1002/ece3.68
Chapuis, E., Trouve, S., Facon, B., Degen, L., and Goudet, J. (2007). High quantitative and no molecular differentiation of a freshwater snail (Galba truncatula) between temporary and permanent water habitats. Mol. Ecol. 16 (16), 3484–3496. doi:10.1111/j.1365-294X.2007.03386.x
Charbonnel, N., Quesnoit, M., Razatavonjikay, R., Brémond, P., and Jarne, P. (2002a). Evolutionnary aspects of the metapopulation dynamics of Biomphalaria pfeifferi, the host of. Schistosoma Mansoni. J. Evol. Biol. 15, 248–261. doi:10.1046/j.1420-9101.2002.00381.x
Charbonnel, N., Quesnoit, M., Razatavonjizay, R., Brémond, P., and Jarne, P. (2002b). A spatial and temporal approach to microevolutionary forces affecting population biology in the freshwater snail Biomphalaria pfeifferi. Am. Nat. 160 (6), 741–755. doi:10.1086/343875
Colley, D. G., Bustinduy, A. L., Secor, W. E., and King, C. H. (2014). Human schistosomiasis. Lancet (London, Engl. 383 (9936), 2253–2264. doi:10.1016/S0140-6736(13)61949-2
Danish Bilharziasis Laboratory (1980). A practical guide to the identification of Africa Freshwater snails. Malacol. Rev. 13, 95–119.
Diakité, N. R., N'Zi, K. G., Ouattara, M., Coulibaly, J. T., Saric, J., Yao, P. K., et al. (2018). Association of riverine prawns and intermediate host snails and correlation with human schistosomiasis in two river systems in south-eastern Côte d'Ivoire. Parasitology 145 (13), 1792–1800. doi:10.1017/S003118201800135X
Diakité, N. R., Ouattara, M., Bassa, F. K., Coulibaly, J. T., Tian-Bi, Y. T., Meïté, A., et al. (2021). Baseline and impact of first-year intervention on Schistosoma haematobium infection in seasonal transmission foci in the northern and central parts of Côte d'Ivoire. Trop. Med. Infect. Dis. 6 (1), 7. doi:10.3390/tropicalmed6010007
El-Hassan, A. A. (1974). Laboratory studies on the direct effect of temperature on Bulinus truncatus and Biomphalaria alexandrina, the snail intermediate hosts of schistosomes in Egypt. Folia Parasitol. 21 (2), 181–187.
Evanno, G., Castella, E., and Goudet, J. (2006). Evolutionary aspects of population structure for molecular and quantitative traits in the freshwater snail Radix balthica. J. Evol. Biol. 19 (4), 1071–1082. doi:10.1111/j.1420-9101.2006.01098.x
Facon, B., Machline, E., Pointier, J. P., and David, P. (2004). Variation in desiccation tolerance in freshwater snails and its consequences for invasion ability. Biol. Invasions 6, 283–293. doi:10.1023/b:binv.0000034588.63264.4e
Frandsen, F., and Christensen, N. O. (1984). An introductory guide to the identification of cercariae from African freshwater snails with special reference to cercariae of trematode species of medical and veterinary importance. Acta Trop. 41 (2), 181–202.
Gérard, C., and Théron, A. (1997). Age/size- and time-specific effects of Schistosoma mansoni on energy allocation patterns of its snail host Biomphalaria glabrata. Oecologia 112 (4), 447–452. doi:10.1007/s004420050331
Global Health & Division of Parasitic Diseases and Malaria, (2018). Parasites—schistosomiasis. Centers for disease control and prevention. https://www.cdc.gov/parasites/schistosomiasis/index.html.
Harrison, A., and Shiff, C. (1966). Factors influencing the distribution of some species of aquatic snails. S. Afr. J. Sci. 62, 3–258.
Kalinda, C., Chimbari, M., and Mukaratirwa, S. (2017a). Implications of changing temperatures on the growth, fecundity and survival of intermediate host snails of schistosomiasis: a systematic review. Int. J. Environ. Res. Public Health 14 (1), 80. doi:10.3390/ijerph14010080
Kalinda, C., Chimbari, M. J., Grant, W. E., Wang, H.-H., Odhiambo, J. N., and Mukaratirwa, S. (2018). Simulation of population dynamics of Bulinus globosus: effects of environmental temperature on production of Schistosoma haematobium cercariae. PLoS Negl. Trop. Dis. 12 (8), e0006651. doi:10.1371/journal.pntd.0006651
Kalinda, C., Chimbari, M. J., and Mukaratirwa, S. (2017b). Effect of temperature on the Bulinus globosus - Schistosoma haematobium system. Infect. Dis. Poverty 6 (1), 57. doi:10.1186/s40249-017-0260-z
Konan, C. K., Tian-Bi, Y. N. T., Diakité, N. R., Ouattara, M., Coulibaly, J. T., Diabaté, S., et al. (2022). Spatial variation of life-history traits in Bulinus truncatus, the intermediate host of schistosomes, in the context of field application of niclosamide in Côte d’Ivoire. BMC Zool. 7, 7. doi:10.1186/s40850-021-00104-7
Moné, H., Minguez, S., Ibikounlé, M., Allienne, J. F., Massougbodji, A., and Mouahid, G. (2012). Natural interactions between S. haematobium and S. Guineensis in the republic of Benin. Sci. World J. 2012, 1–8. doi:10.1100/2012/793420
Najarian, H. H. (1961). Biological studies on the snail, Bulinus truncatus, in Central Iraq. Bull. World Health Organ 25 (4-5), 435–446.
Nelwan, M. L. (2019). Schistosomiasis: life cycle, diagnosis, and control. Curr. Ther. Res. Clin. Exp. 91, 5–9. doi:10.1016/j.curtheres.2019.06.001
N’Goran, K. E. (1987). Situation épidémiologique des Schistosomoses en zone rurale du centre de la Côte d’Ivoire: influence d’un barrage à vocation agropastorale. Thèse 3ème cycle, Univ. Abidjan-Cocody , N° 109, 108.
Niemann, G. M., and Lewis, F. A. (1990). Schistosoma mansoni: influence of Biomphalaria glabrata size on susceptibility to infection and resultant cercarial production. Exp. Parasitol. 70 (3), 286–292. doi:10.1016/0014-4894(90)90110-x
Ouattara, M., Diakité, N. R., Yao, P. K., Saric, J., Coulibaly, J. T., Assaré, R. K., et al. (2021). Effectiveness of school-based preventive chemotherapy strategies for sustaining the control of schistosomiasis in Coˆte d’Ivoire: results of a 5- year cluster randomized trial. PLoS Negl. Trop. Dis. 15 (1), e0008845. doi:10.1371/journal.pntd.0008845
R Core Team (2020). “R: a language and environment for statistical computing,” in R version 3.6.3 (Vienna, Austria: R Foundation for Statistical Computing).
Reznick, D., Bryant, M. J., and Bashey, F. (2002). r- and K-selection revisited: The role of population regulation in life-history evolution. Ecol. 83, 1509–1520.
Satayathum, S. A., Muchiri, E. M., Ouma, J. H., Whalen, C. C., and King, C. H. (2006). Factors affecting infection or reinfection with Schistosoma haematobium in coastal Kenya: survival analysis during a nine-year, school-based treatment program. Am. J. Trop. Med. Hyg. 75 (1), 83–92.
Stearns, S. C. (1989). Trade-offs in life-history evolution. Funct. Ecol. 3 (3), 259–268. doi:10.2307/2389364
Stensgaard, A.S., Utzinger, J., Vounatsou, P., Hürlimann, E., Schur, N., Saarnak, C., et al. (2013). Large-scale determinants of intestinal schistosomiasis and intermediate host snail distribution across Africa: Does Climate Matter? ActaTropica 128 (2), 378–390. doi:10.1016/j.actatropica.2011.11.010
Tabo, Z., Neubauer, T. A., Tumwebaze, I., Stelbrink, B., Breuer, L., Hammoud, C., et al. (2022). Factors controlling the distribution of intermediate host snails of Schistosoma in Crater Lakesin Uganda: a machine learning approach. Front. Environ. Sci. 10, 871735. doi:10.3389/fenvs.2022.871735
Tchuenté, L. A., and N'Goran, E. K. (2009). Schistosomiasis and soil-transmitted helminthiasis control in Cameroon and Côte d'Ivoire: implementing control on a limited budget. Parasitology 136 (13), 1739–1745. doi:10.1017/S0031182009005988
Tchuenté, L. A., Southgate, V. R., Vercruysse, J., Kaukas, A., Kane, R., Mulumba, M. P., et al. (1997). Epidemiological and genetic observations on human schistosomiasis in Kinshasa, Zaire. Trans. R. Soc. Trop. Med. Hyg. 91 (3), 263–269. doi:10.1016/S0035-9203(97)90068-7
Tian-Bi, Y. N., Jarne, P., Konan, J. N., Utzinger, J., and N'Goran, E. K. (2013). Contrasting the distribution of phenotypic and molecular variation in the freshwater snail Biomphalaria pfeifferi, the intermediate host of Schistosoma mansoni. Heredity 110 (5), 466–474. doi:10.1038/hdy.2012.115
Tian-Bi, Y. T., Ouattara, M., Knopp, S., Coulibaly, J. T., Hürlimann, E., Webster, B., et al. (2018). Interrupting seasonal transmission of Schistosoma haematobium and control of soil-transmitted helminthiasis in northern and central Côte d'Ivoire: a SCORE study protocol. BMC public health 18 (1), 186. doi:10.1186/s12889-018-5044-2
Tian-Bi, Y. T., Webster, B., Konan, C. K., Allan, F., Diakité, N. R., Ouattara, M., et al. (2019). Molecular characterization and distribution of Schistosoma cercariae collected from naturally infected bulinid snails in northern and central Côte d'Ivoire. Parasit. Vectors 12 (1), 117. doi:10.1186/s13071-019-3381-3
Viard, F., Bremond, P., Labbo, R., Justy, F., Delay, B., and Jarne, P. (1996). Microsatellites and the genetics of highly selfing populations in the freshwater snail Bulinus truncatus. Genetics 142 (4), 1237–1247. doi:10.1093/genetics/142.4.1237
Webster, B. L., Tchuem Tchuenté, L. A., and Southgate, V. R. (2007). A single-strand conformation polymorphism (SSCP) approach for investigating genetic interactions of Schistosoma haematobium and Schistosoma guineensis in Loum, Cameroon. Parasitol. Res. 100 (4), 739–745. doi:10.1007/s00436-006-0310-0
Who, (2022). Guideline on control and elimination of human schistosomiasis. Available online: https://www.who.int/publications/i/item/9789240041608.
Keywords: Bulinus truncatus, Biomphalaria pfeifferi, climatic factors, survival, growth, fecundity, Côte d’Ivoire
Citation: Diakité NR, Koffi PB, Konan CK, Bassa FK, Chamberlin AJ, Ouattara M, De Leo GA and N’Goran EK (2023) Variability of biological traits of Bulinus truncatus and Biomphalaria pfeifferi, the intermediate host snails of schistosomiasis, from three climatic zones of Côte d’Ivoire. Front. Environ. Sci. 11:1193239. doi: 10.3389/fenvs.2023.1193239
Received: 27 March 2023; Accepted: 24 November 2023;
Published: 07 December 2023.
Edited by:
Tatenda Dalu, University of Mpumalanga, South AfricaReviewed by:
Gabriel Mouahid, UMR5244 Interactions Hôtes Pathogènes Environnements (IHPE), FranceChristian Albrecht, University, Germany
Thendo Mutshekwa, Rhodes University, South Africa
Copyright © 2023 Diakité, Koffi, Konan, Bassa, Chamberlin, Ouattara, De Leo and N’Goran. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nana R. Diakité, ZGlha25hcm9zZUB5YWhvby5mcg==