 Hongwei Zhang1,2
Hongwei Zhang1,2 Lihui Tian
Lihui Tian Eerdun Hasi
Eerdun Hasi
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Sci. , 16 February 2023
Sec. Drylands
Volume 11 - 2023 | https://doi.org/10.3389/fenvs.2023.1119605
This article is part of the Research Topic Hydrological Connectivity and Sustainable Watershed Management in a Changing Environment View all 13 articles
Different types of dunes cause habitat differences, which can affect vegetation growth. In turn, water utilized by vegetation leads to the differential distribution of soil moisture in different dunes. However, vegetation–soil dynamics in the alpine desert of the Qinghai–Tibet Plateau remain poorly understood. We chose the largest section of desert along the eastern shore of Qinghai Lake as a study area to test the hypotheses that plant community characteristics and soil physicochemical properties show differences in different sand-binding communities, in addition to soil moisture content, based on methods of field investigations and in situ observations. The main results were as follows:1) Plant community species diversity and herb coverage increased with the stabilization status of dunes; meanwhile, coverage increased with the age of the added artificial semi-fixed dunes. 2) Surface deposits were mainly composed of medium sand, leading to low organic matter and nutrient content in the study area; this being different from other deserts in northern China. 3) Soil moisture’s seasonal variation was consistent with rainfall seasonality and lacked significant interannual differences, while its vertical distribution was influenced by precipitation infiltration. 4) Soil crust forms beneath specific species, such as leguminous plants in the study area, which should be effective at preventing wind erosion. Our findings will facilitate a mechanistic understanding of plant–soil–water relationships in alpine deserts and provide timely information for screening introduced species for enhancing sand-fixation effectiveness.
Desertification has attracted much international attention as one of the most serious land degradation problems, and establishing suitable land rehabilitation projects in sensitive areas would be the best strategy for slowing desertification worldwide (D’Odorico et al., 2013; Huang et al., 2020). Vegetation restoration and reconstruction have become the most important and effective measures and methods for mitigating desertification in drylands (Li et al., 2014; Qi et al., 2018). Shrubs are often used as priority individuals for planting in desertification restoration projects, as they can generally maintain a high coverage rate and strong protective performance under the most severe wind erosion conditions in spring and winter. Once pioneer plant species have been established on mobile dunes, aeolian sediments tend to be fine-grained and biological crusts begin to appear under the canopy (Zuo et al., 2008; Li et al., 2013). Yet, vegetation coverage undergoes a self-thinning process that declines during the succession of vegetation from artificial ecosystems to natural ecosystems (Wang et al., 2005). A previous study showed that vegetation coverage is close to being maximally coordinated with local soil moisture carrying capacity, rendering soil moisture a key limiting factor leading to shrubs’ mortality and leaving herbs to gradually become the dominant species in a semi-arid desert in China (Li et al., 2014). Nevertheless, that would be a long-term succession process, one that is still poorly understood in alpine deserts. There is no doubt that vegetation–soil dynamics are the principal focus of study for understanding the stability of a desert ecosystem, where characteristics of vegetation and soil properties strongly interact with each other (Maxwell et al., 2018). On one hand, artificial measures can weaken aeolian activities and promote organic matter’s increase on the surface due to the interception of dust; on the other hand, soil structure can influence the rainfall infiltration rate and pattern of plants’ water and nutrient absorption by roots (Noy-Meir, 1979; Silva and Lambers, 2018). Therefore, understanding the dynamics of both the plant community and soil at the initial restoration stage is useful to gain insight into the evolution of alpine desert ecosystems.
Qinghai Lake is the largest inland saltwater lake in China. It is not only the convergence zone of China’s three major climate types but also a buffer zone between the Qaidam Basin and Huangshui Valley in the Qinghai–Tibet Plateau (Li et al., 2018). Providing ecological security for the northeastern Qinghai–Tibet Plateau, and a natural barrier blocking off desertification that is spreading eastward, Qinghai Lake plays a paramount ecological role in the stable development of the Qinghai–Tibet Plateau. Currently distributed around Qinghai Lake is about 134 295 ha of desert, which has expanded by 2.2% annually since the 1950s (Wu et al., 2019a). Desertification of the Qinghai Lake watershed provides a hotspot for research investigating the influence of climate change on the Tibetan Plateau (Li et al., 2018). The local Qinghai Province government has encouraged afforestation and conducted several sand-fixation projects to impede the spread of mobile dunes along the nearby Qinghai–Tibet Railway and Eastern Surrounding Road (Tian et al., 2019). Desertification has been effectively curbed by a relatively stable pattern of “fixed lowlands,” “semi-fixed dunes,” and “mobile dunes” from the lakeshore to the foothills of Riyue Mountain, the eastern boundary of Qinghai Lake.
Recent studies in this area have mainly focused on desert formation, dynamic changes of desertification, desertification control measures, soil particle composition, soil moisture dynamics, effective control and evaluation of straw checkerboards, and individuals’ effects on reducing wind-driven sand erosion (Tian et al., 2015; Tian et al., 2019; Tian et al., 2020), in addition to water use sources of afforestation species (Wu et al., 2016; Wang et al., 2022). Work by Wu H et al. (2019) has analyzed species characteristics (richness and traits) and other afforestation community features over several years. Earlier, Lu et al. (2013) focused on the effect of rainfall on replenishing soil moisture in newly afforested areas. Yet, little attention has been paid to the interaction between community features and soil properties in the alpine desert ecosystem of the Qinghai–Tibet Plateau, especially in the Qinghai Lake watershed.
To fill these knowledge gaps, here we illustrate community characteristics and soil physicochemical properties during growing seasons in an alpine semi-arid desert in the northeastern region of the Tibetan Plateau. The tested hypotheses were that 1) plant community diversity and coverage and soil physicochemical properties showed differences in different sand-binding communities; 2) soil moisture content was higher in natural dunes than in afforested dunes. This study’s findings will facilitate a mechanistic understanding of plant–soil–water relationships in alpine desert ecosystems, while also providing information for screening introduced species for sand-fixation effectiveness according to local soil conditions and soil water content. This could have far-reaching significance for the ecological engineering of desertification control in the Qinghai–Tibet Plateau.
The study was conducted at the eastern shore of Qinghai Lake, which belongs to the Ketu Wind Prevention and Sandfixation Experimental Range (hereafter, WPSER; 36°46.9′ N, 100°46.8′ E; 3,224 m; for more details about WPSER, refer to Tian et al. (2019) and Tian et al. (2020)). Mega-dunes and continuous mobile dunes are distributed across the WPSER. After 5 years of treatment, the amount of vegetation covering the mobile dunes, semi-fixed dunes, and fixed dunes has increased by 30.4%, 20.2%, and 11.6%, respectively (Tian et al., 2015).
The WPSER is situated within an alpine semi-arid climatic zone, having a relatively wet summer and cold winter. Based on data from the Haiyan County National Meteorological Station, located ca. 20 km away from the WPSER, the annual mean temperature is 0.93°C, with the lowest mean monthly temperature occurring in January (–13.1°C) and the highest occurring in July (12.4°C). The annual average precipitation is 438.6 mm, approximately 80% of which falls between July and September (Wang et al., 2022). Precipitation and temperature have both increased since 2004, especially from 2010 onward (Cui and Li, 2015; Wu H et al., 2019). The WPSER is a low-wind energy environment whose aeolian activity is strongest from late autumn to early spring, being controlled by the north-west wind direction. The threshold wind velocity for sand movement on a mobile dune is 6.5 m s−1, but it is greater than 8 m s−1 on afforestation dunes (Tian et al., 2020). The natural shrub species distributed in natural fixed dunes are mainly Artemisia ordosica and Caragana jubata, while natural herbs are Carex qinghaiensis and Achnatherum splendens. The soil in the study area consists of sand without a typical soil development sequence. The growing season of vegetation begins in May and ends in September or early October.
In spring 2011, we selected five dunes in the WPSER: a natural fixed dune (SD), a natural semi-fixed dune (SSD), a mobile dune (MD), and two dunes with artificial measures (SJD and WLD). Their corresponding plots are summarized in Table 1. We investigated vegetation features and soil properties on the top of each dune from 2011 through 2013.
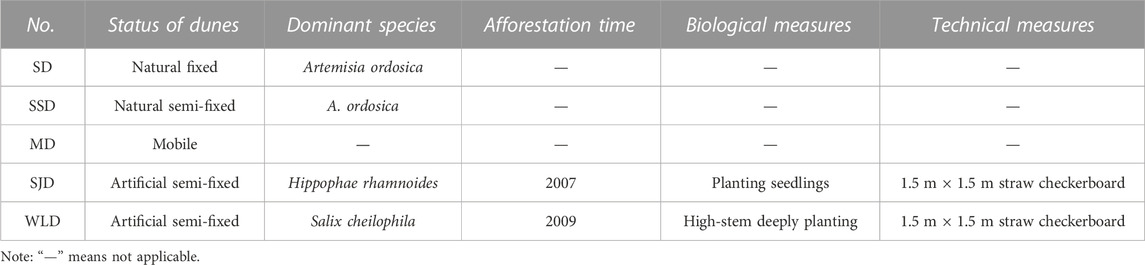
TABLE 1. Study sites in the WPSER.
Three randomized 5 m × 5 m plots were established at the top of each dune type. Then, traditional measurements in ecology were used to survey the shrub species and their canopy and height in each plot. According to these data, we calculated shrub coverage and defined the dominant species in each plot community. Within fixed shrub plots, we set up five quadrats (each 1 m × 1 m: one in the center, four at the corners) and then measured the height and number of herbaceous species to calculate herb coverage. In this study, the Simpson index D, Shannon–Wiener index H, and Pielou evenness index E were used to measure the species diversity of communities, as follows:
where pi is the relative importance value of species i (relative height plus relative coverage) and S is the total number of species i in the plot or quadrat, that is, an abundance index.
A total of 75 soil cores were collected from all 15 plots to a depth of 80 cm, by digging sections from each plot in triplicate. The depth profile was divided into five layers from the surface to 80 cm depth: 0–10 cm, 10–20 cm, 20–40 cm, 40–60 cm, and 60–80 cm. Soil samples were analyzed in the Key Laboratory of Environmental Change and Natural Disaster [ECND] at Beijing Normal University for particle size, organic matter content (OM), and available nitrogen (N), phosphorus (P), and potassium (K) content. The fractions of sand, silt, and clay were obtained following the standard defined by Blott and Pye (2001): sand (0.063 < d < 2.00 mm), silt (0.002 < d < 0.063 mm), and clay (d < 0.002 mm). Organic matter content was determined by the method of Walkley and Black (1934).
Soil moisture data have been collected in the WPSER using an automatic sensor network. This includes the aforementioned five dune (plot) sites, where it has been operating since August 2009. Capacitance-type soil moisture probes (ECH2O-5, Decagon Devices Inc., USA) were variously installed in five soil horizons according to five layers of soil samples (mentioned previously). All the cables were carefully buried below the depth of the installed probes and then laid horizontally away from each probe location. We obtained continuous 10-min time-series data from August 2009 through December 2013. In this study, we only used soil moisture data for the entire year, from 2011 to 2013, for consistency with the data obtained for the investigated vegetation and soil properties.
The variation in community diversity or plant coverage among the five dune types was distinguished by one-way analysis of variance (ANOVA) combined with a least significant difference (LSD). ANOVAs were also used to detect differences in soil particle size, organic content, and bulk density for each community. The alpha = 0.05 significance level was set for the statistical analyses, all of which were performed in SPSS 26.0 software (SPSS Inc., Chicago, IL, USA). All figures were drawn in the Origin 2018 software (OriginLab, Northampton, MA, USA).
As seen in Table 2, there were nine families and 13 species at the study sites after afforestation. These consisted of five species of Leguminosae, two species of Poaceae, and other families that were composed of only one species. Except for Carex tristachya, Artemisia ordosica, Astragalus adsurgens, and Astragalus polycladus, all other species had been artificially planted. Since the 1980s, Hippophae rhamnoides has been most commonly used in afforestation projects in the WPSER. Starting in 2007, Salix cheilophila, Pinus sylvestris, Potentilla fruticosa, and Sabina vulgaris were planted to implement China’s 11th Five-Year Plan in the study area. The greater the dune fixation, the higher its community richness and number of perennial herb species, as shown in Table 3. Salix stems were deep-planted in 2009 on WLD and then replanted with H. rhamnoides, P. fruticosa, and P. sylvestris; breeding of Caragana korshinskii, A. ordosica, and Hedysarum scoparium was undertaken in 2010. Therefore, the increased richness was human-induced.
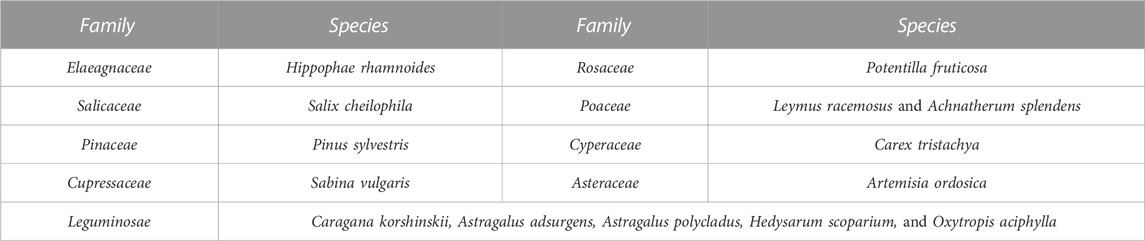
TABLE 2. Species composition of plant communities on the sites (in accordance with family classification).
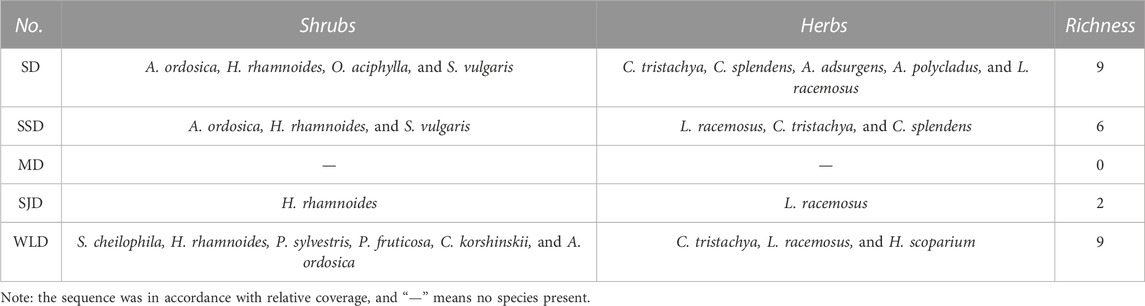
TABLE 3. Species and richness of plant communities on the study sites (2013).
Table 4 shows that the greater a dune’s fixation, the higher the species diversity in its community. However, there were no significant differences among dune types in terms of H nor for evenness (E) and richness (D) (p > 0.05). With each subsequent fixation year, the species diversity increased. Because artificial species were abundant in the WLD, its α-diversity index exceeded that of other dune types, but S. cheilophila remained the dominant species.
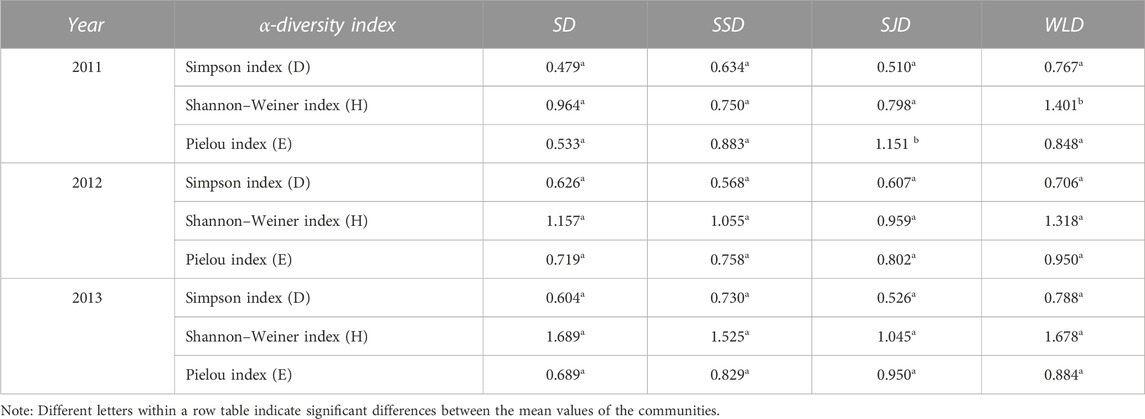
TABLE 4. α-diversity of communities on study sites (2011–2013).
Afforestation stabilized mobile dune surfaces, which provided a favorable condition for the recovery of natural species. As Figure 1 shows, the total vegetation cover on artificial semi-fixed dunes (WLD and SJD sites) reached more than 40% in 2013. The vegetation cover exhibited an increasing trend during the study period. Meanwhile, the vegetation cover on artificial mobile dunes in the initial restoration years was similar to that on natural semi-fixed dunes. In addition, the vegetation cover was low due to lower precipitation in 2012, except for natural fixed dunes.
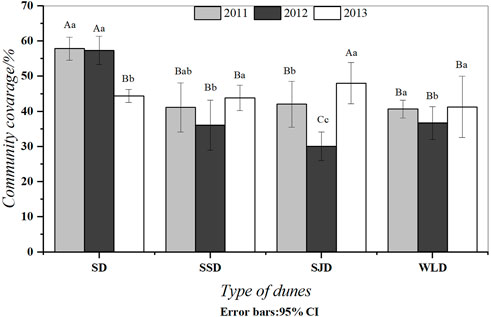
FIGURE 1. Coverage dynamics of different dunes. Note: Differing capital letters indicate that coverage in different communities was significantly different (p < 0.05), while differing lower-case letters indicate that the coverage of the same communities in different years was significantly different (p < 0.05). The same letter indicates no significant difference (p > 0.05).
A. ordosica was not only the dominant species in SD and SSD but also the most abundant naturally established species in the study area. As seen in Table 5, according to an overall survey of its vegetation recovery level, A. ordosica increased in cover with elapsed years of recovery growth, though its density varied negligibly while its plant height increased. The A. ordosica growing on SJD came from seedling afforestation in 2009, whereas the A. ordosica on WLD had high coverage and arose from both artificial seedlings and natural recovery.
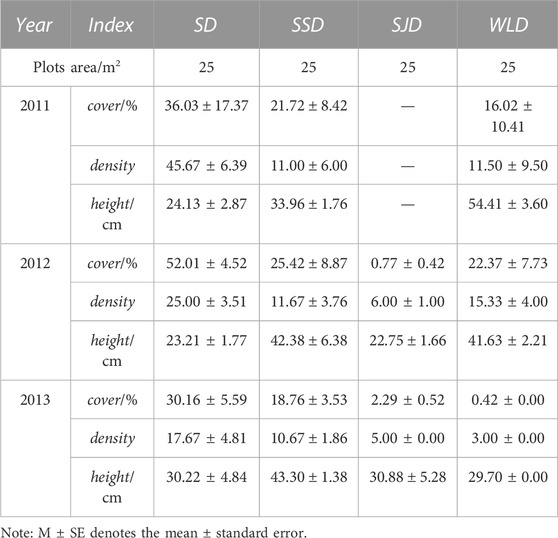
TABLE 5. Features of Artemisia ordosica in different dunes (M±SE).
Table 6 shows that on SD, there was minimal clay, whose concentrations were <1%. The silt was the most evident conversion feature that emerged during the evolution from sand to the soil, being closely related to sediment nutrients. Meanwhile, it was also the main substance carried by wind in the sand–wind flow. The silt was more prominent in SD than in the other dune types (p < 0.05). Fine and medium sand were the main particles in the sediments and nearly equal in the content on SD, whereas medium sand predominated in mobile dunes more than fixed dunes. However, coarse sand was not regular between dunes.
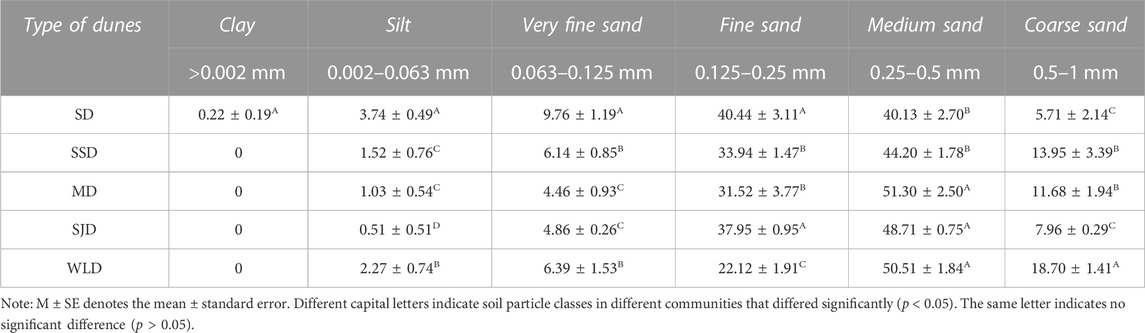
TABLE 6. Particle size composition (M±SE, %) in the surface 10 cm of deposits in different dunes.
As Figure 2 shows, SD had the highest organic matter content at 4.56 g kg−1. Organic matter content was closely related to clay content. There were significant variations in organic matter content among SSD, SJD, and WLD (p < 0.05) while it was different between SD and MD (p > 0.05).
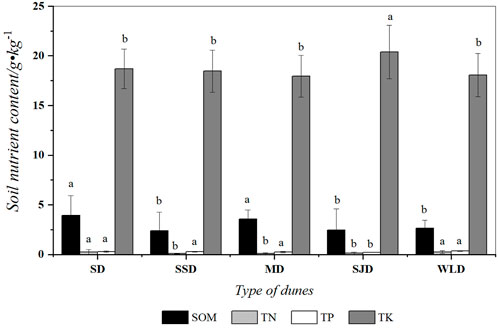
FIGURE 2. Organic matter and nutrient content 80 cm below the surface of deposits in dunes examined. Note: different letters indicate organic matter and nutrient content differed significantly between communities (p < 0.05); the same letter indicates no significant difference (p > 0.05).
Soil nutrient content was relatively low in the WPSER. The average content of available nitrogen, phosphorus, and potassium was 0.07 g kg−1, 0.002 g kg−1, and 0.17 g kg−1, respectively. The content of available nitrogen in SD exceeded that in other dune types (p < 0.05). In this region, phosphorus would be negligible, being found only on SD in a very small amount, i.e., 0.004 g kg−1. There was no significant difference (p > 0.05) in potassium content among the dune types, except for SJD.
Analyzing the bulk density of the soil in the dune types (Figure 3) revealed little difference between them (p > 0.05), with an average content of 1.54–1.67 g cm−3. This was attributable to higher vegetation amelioration on dunes, which may also be associated with the functional types of species; e.g., soil amelioration that was significantly superior in H. rhamnoides with rhizobia than S. cheilophila.
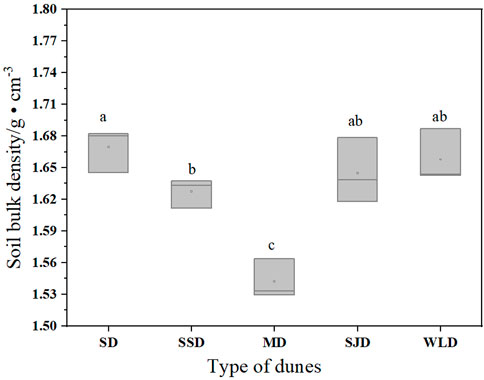
FIGURE 3. Soil bulk density 80 cm below the surface of deposits in dunes examined in the WPSER. Note: different letters indicate soil bulk density differed significantly between communities (p < 0.05); the same letter indicates no significant difference (p > 0.05).
Soil moisture content (SMC) was the highest in SD and SSD in the study area, and its value was more than or similar to 9% from 2011 to 2013 (Figure 4). Meanwhile, the volumetric moisture content in the other three dunes was below 9%. There were no significant interannual differences for the same dune type (p > 0.05). Nevertheless, within the same year, SMC was different among various dunes; in general, that of SJD was the lowest.
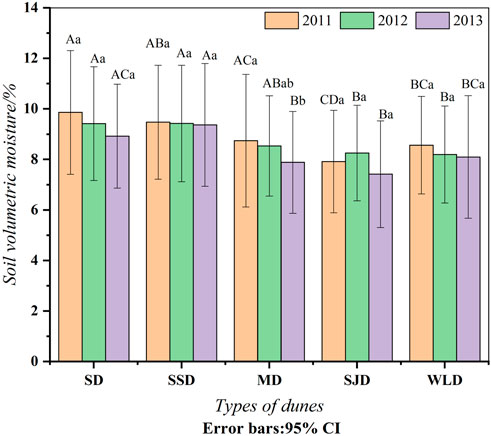
FIGURE 4. Soil moisture dynamics from 2011 to 2013 in dunes examined. Note: different capital letters indicate the SWC differed significantly between communities (p < 0.05), while different small letters mean the SWC of the same communities varied significantly from year to year (p < 0.05). The same letter indicates no significant difference (p > 0.05).
As seen in Figure 5, from December to the following February, a freezing period occurred with low soil temperatures and the lowest soil moisture. However, soil moisture remained relatively stable during the winter. In March, soil thawing and snow melting led to a rapid replenishment of soil moisture. Plants began germinating in early April when soil water consumption was still low via transpiration. Thus, soil moisture increased at the onset of vegetation growth. Whereas H. rhamnoides consumed a large amount of water during its early growth stage, soil moisture at the SJD site was lower in April. In May, rainfall increased, followed by a gradual increase in vegetation transpiration, when soil moisture increases were unlikely to occur. From July to August, plants were in a vigorous phase of the growing season, distinguished by a stronger evapotranspiration rate and peaks in yearly rainfall. Rainfallaugmented soil moisture and soil moisture loss via evaporation were almost equal. Therefore, soil moisture remained stable at this stage. In September, plants underwent a decline in their growth. Soil moisture was at its yearly peak because of the large amount of soil water stored and a small quantity of rainfall supplement. Then, soil moisture exhibited a sharp decline due to the little precipitation in October. At this point, plants have completed a year of growth and begin to go dormant, and the soil begins to freeze.
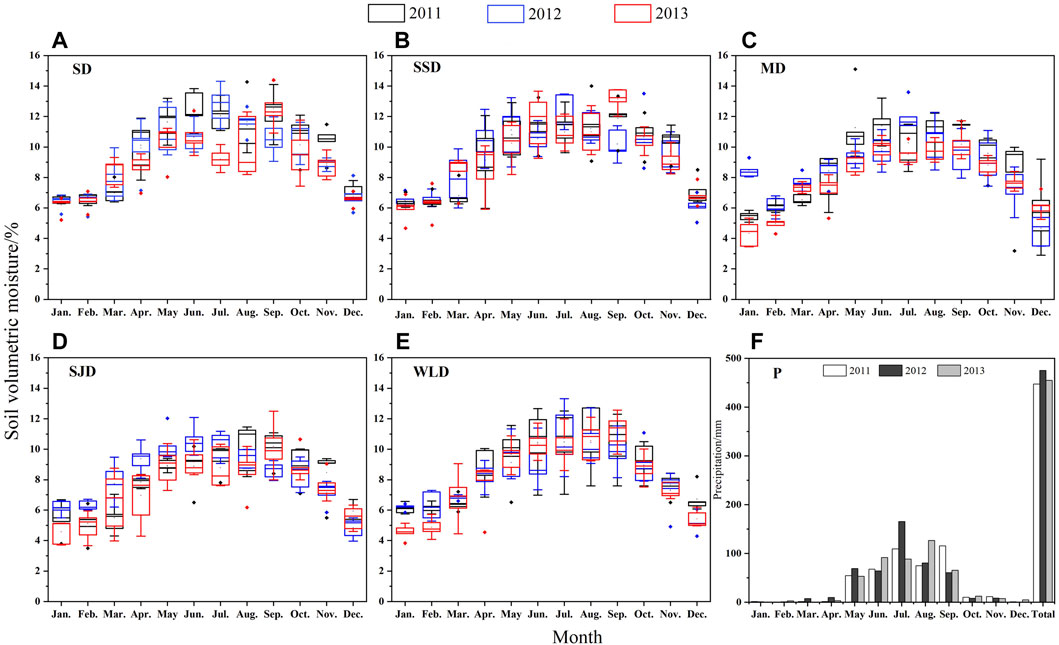
FIGURE 5. Seasonal variation of soil moisture dynamics 80 cm below the surface from 2011 to 2013 in different dunes (A–E) and precipitation (F) in the study area.
According to Figure 6, the vertical depth of soil moisture in the different dune types varied predictably. Differences in annual precipitation led to significant interannual variability in SWC at 20 cm depth. Contrasting the vertical differentiation of soil moisture from 2011 to 2013, the soil moisture content was highest at the 10–20 cm depth in dunes except for MD, which attained its greatest value at the 20–40 cm depth in both 2011 and 2012 but at the 60–80 cm depth in 2013. Overall, the point of the highest soil moisture content was deeper in 2013 than in the first two years, except for SJD and SSD, whose interannual variation was not pronounced.
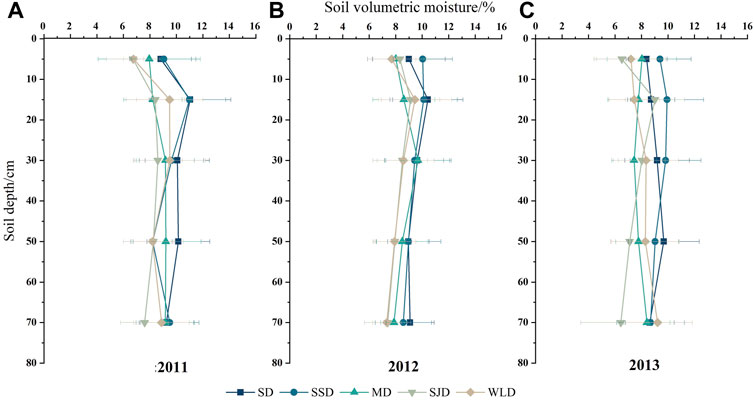
FIGURE 6. Vertical soil moisture variation 80 cm below the surface between 2011 and 2013 in different dunes.
Table 7 describes the crusts present on the soil surface at SD, SJD, and WLD. Soil crusts developed under O. aciphylla, A. ordosica, and H. rhamnoides canopies, covering up to 70% of the surface with a 0.5–1.0 cm thickness at SD. Biological crusts were covered by the two former plant species, whereas a physical crust formed under the third species. There were no crusts under any herbs. Atop SSD, A. ordosica nebkhas formed without any soil crusts. No crusts were yet found on MD.
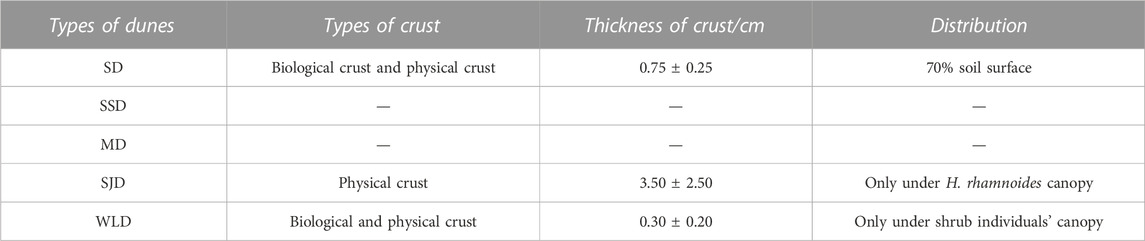
TABLE 7. Features of crusts in different types of dunes in the WPSER.
Straw checkerboard barriers placed atop SJD had an average height of 2–3 cm. H. rhamnoides were planted in the northwest checkerboard in 2007, where they grew well with few dry stems. A hard physical crust has formed under the canopy of that species since 2011. Crust thickness—including the thickness of the adhesive sand—was 0.5–6 cm.
In 2009, we deeply planted S. cheilophila with tall stems on WLD and then replanted with H. rhamnoides and P. fruticosa in 2010. The biological crust thickness was 0.1–0.2 cm in the A. ordosica nebkhas. Underneath S. cheilophila plants, we found biological crusts with a thickness of 0.2–0.5 cm. The physical crust was 1.5 cm thick under H. rhamnoides.
In drylands, abiotic processes such as wind- and water-driven erosion significantly influence surface processes (Turnbull et al., 2012). Before artificial measures were taken, sand movements were extremely strong on mobile dunes because of forceful winds in winter and spring. Once plantations were established on dunes, wind-related hazards decreased considerably (Tian et al., 2020).
Soil texture is a key factor in the coupled relationships between climate, soil, and vegetation, which affect soil erosion, soil accumulation, and soil physicochemical processes (Fernandez-Illescas et al., 2001; Renne et al., 2019). In other words, soil particle composition is one of the most important soil physical properties because it directly affects soil moisture transport and vegetation distribution patterns (Pan and Wang, 2009; Bayat et al., 2015). Medium and fine sands were the dominant particles in the study area (Table 6), whose content was more than 72.36% at some dune sites. Also, sand has a stronger infiltration capacity for precipitation than either silt or clay, thus creating an “inverse texture effect” in arid and semi-arid ecosystems (Noy-Meir, 1973; Porporato et al., 2002). Overall, the coarser the sand, the greater the water infiltration capacity that is exploitable by plant individuals in a water-limited environment (Pan et al., 2015). In our study area, the particles of natural fixed dunes were significantly finer than those of mobile or semi-fixed dunes, both artificial and natural, which ultimately resulted in less infiltration into deeper soils, allowing direct uptake by vegetation for survival (Wang et al., 2022).
Soil is the most basic and important part of an ecosystem, and fertile soil is an important guarantee for the healthy and sustainable development of plant communities (Noy-Meir, 1973). Our analysis of the physical and chemical characteristics of soil in the alpine desert uncovered a low content of organic matter and nutrients, particularly phosphorus, and the deposits mainly consisted of fine sand or medium sand with little clay present (Table 6; Figure 2). However, in this study, plantations on the artificial semi-fixed dunes were in an early stage of succession, and their deposits were still coarse in soil texture. With more years of vegetation restoration, the physical and chemical properties of soil become more robust indicators of soil development (Yu and Jia, 2014); good examples of this include soil texture refinement, soil organic matter, and nutrient content increases, soil bulk density decreases, and biological crust development.
The groundwater level at a dune’s topmost position is above 10 m in depth in our study area (Wang et al., 2022). Furthermore, there were few rainfall events from January to April, so precipitation had little effect on soil water content. However, the amount of soil water replenishment via soil melting increases significantly in May. Moreover, Qinghai Lake is located in the distribution area of seasonally frozen soil. The high-water zone formed by gaseous water condensation in this seasonally frozen soil, namely, a cold-generated wet sand layer, is beneficial for the growth of desert plants (Wan et al., 2005). However, in this area, the depth of the groundwater in the sand dunes is too significant for gaseous water transport to be utilized by artificial sand fixation vegetation.
Desertification control measures consist mainly of biological measures, or these are combined with mechanical measures, and soil moisture is one of the main limiting factors for vegetation restoration in the study area (Lu et al., 2013). Precipitation quantity and seasonality interact with soils to determine the spatiotemporal patterning of plant-available water in the soil, thereby influencing the differential success of plant growth forms in water-limited systems (Renne et al., 2019). Soil moisture seasonal dynamics in our study area could be divided into three periods. The first is a stage where stagnant water is frozen from December to the following February when the soil moisture content is low and relatively stable. The second stage lasts from March to June when losses in soil moisture are compensated by increases in soil moisture. The third and final stage occurs during a period of dehydration from July to November when high soil moisture levels gradually decrease. Overall, the highest soil moisture occurs in summer and autumn, when plants need water the most to maintain their growth, as proven by Rodriguez-Iturbe et al. (1999).
Community features and soil properties are mutually reinforcing (Maxwell et al., 2018). On one side, vegetation cover reduces radiation and wind speed at the soil surface, thus reducing evaporation (Porporato et al., 2002). On the other side, once the vegetation has established on a mobile dune, leafy plants begin to control, to some extent, the loss of their own resources and create a favorable microenvironment for other plants and animals (Noy-Meir, 1973). Furthermore, vegetation also affects soil particle size composition, mainly in two ways. First, soil layers under the influence of different vegetation will have different root systems and their associated rhizosphere secretions, which in turn change the physicochemical properties of soil and its particle composition (Zhu et al., 2014). Second, the capacity of vegetation to intercept rainfall and lessen surface soil erosion depends on the canopy density and coverage of plants, leading to different retention capacities of surface soil particles (Gu et al., 2018). By comparing the particle size characteristics of the five dune types (Table 6), we conclude that artificial measures can significantly impact the soil surface by promoting the deposition of fine particles and by effectively reducing surface wind erosion.
In terms of the vegetation’s vertical structure, its above-ground components, such as plant type, canopy structure, surface cover, and litter, can reduce water erosion by intercepting rainfall and attenuating the energy of raindrops. Furthermore, root systems can slow surface runoff and weaken soil erodibility by improving soil structure and its biophysical properties (Xu et al., 2006). A recent study shows that precipitation in the studied region can strongly influence the moisture content at the surface (0–20 cm), while deeper soil moisture is less replenished (Wang et al., 2022), as was the case in our study.
In addition, vegetation restoration affects soil properties and hydrological processes in drylands by increasing soil organic matter content (Wu et al., 2023). In arid and semi-arid areas, individual plants’ canopies and collective vegetation cover are the main factors affecting soil water content, which is negatively correlated with soil water content in the community (Pan and Wang, 2009). The greater the width of a plant’s canopy, the more water it requires and the greater its water consumption (Wang et al., 2011). Yet, the soil water content of natural fixed dunes in the study area exceeded that of mobile dunes (Figure 4), due to the fast precipitation infiltration rate of mobile dunes and the lack of effective interception of precipitation by ground vegetation and shielding of surface evaporation by the canopy. Seasonal variation in soil moisture was similar among the dune types, but differences in water consumption among individual plants can also lead to changes in soil water content (Tian et al., 2021).
The shade and higher surface soil moisture under a shrub canopy enhance biocrust formation, which can significantly bolster the resistance of the soil surface to wind erosion by increasing the wind friction velocity threshold of the soil (Li et al., 2021). Once the straw checkerboard barrier is established, the prerequisite microenvironment for physical crust formation has been created. When H. rhamnoides is planted in the checkerboard’s corners, rainfall is intercepted by straw, and its mulched leaves form hard black physical crusts. We find that crust formation is strongly related to the kind of vegetation growing locally (Table 7). Furthermore, the length of formed crusts on dunes was generally longer in the north–south direction than the east–west direction, likely due to the prevailing westerly and northwesterly winds that disrupt the presence of crusts (Jiang et al., 2018). Previous research has shown that the amount of biocrust cover has a negative effect on rainfall infiltration, leading to the death of mature shrubs in Chinese temperate deserts (Li et al., 2013). The hydrological roles of biocrusts in the alpine desert warrant further investigation.
Given the short recovery times of three-five years for SJD and WLD, fewer herbs will have settled on these dunes in the WPSER. In winter and spring, the contribution of herbaceous cover to the plant community is almost negligible, unlike that from shrubs and subshrubs, chiefly due to strong winds and low temperatures that hasten graminoid mortality. For example, the above-ground segments of L. racemosus and C. tristachya dried out in October and were later blown off in winter; hence, both species had to produce new branches from their roots in May of the following year. As a result, the Poaceae family has a very low density and contributes little to the composition of the desert community.
With the successive establishment of plantations, higher surface cover from plants leads to excessive moisture and intense local competition among individuals, which eventually leads to poor shrub growth (Yu et al., 2016; Yu et al., 2020). Based on a sound understanding of the spatiotemporal distribution of soil moisture, it is necessary to select appropriate plant species and allocate their densities rationally to achieve robust long-term effects on blocking alpine desert winds and reducing sand erosion (Yu et al., 2017; Li and Sawada, 2022). The differences in plant morphology and community characteristics of dune plantations reflect the ecological adaptations of sand-fixing species in alpine deserts (Wu et al., 2019b). Under similar climatic and soil conditions, species diversity in communities is determined by physiological and innate ecological adaptation mechanisms. Nonetheless, afforestation practices and structural patterns deserve more attention in future desert research and management.
An ecosystem is a collection of organic and inorganic components that are open to being influenced by human disturbances (Hill, 1975). In this very complex dune ecosystem, there is a self-regulating, buffering, and repair mechanism operated by the external environment (Noy-Meir, 1979–80). If the pressure from the external environment reaches a tolerable threshold, the vegetation can restore itself to some stable state after normal environmental conditions recover. Once the threshold is exceeded, however, both soil properties and vegetation will likely undergo a series of changes (Zhou et al., 2020). During this process, the alterations to the physical and chemical properties of the soil are not as pronounced as those of the vegetation, but when they do occur, it is difficult to restore them to their original state (Dormaar and Willms, 1990). Soil properties and vegetation are closely related; for example, high levels of ecosystem biodiversity with a complex vegetation composition are resilient, that is, able to withstand external disturbances. By contrast, plantations that are monoculture systems are less resilient and thus easier to change because their resistance to disturbance is minimal. In the study area, the reduced vegetation cover in SD was mainly caused by human damage to the biological crusts and the expansion of bare areas detected by yearly field surveys in spring during the study period (Figure 1). Finally, strong wind erosion occurred in spring and winter, ultimately leading to reductions in vegetation cover. However, soil moisture infiltration may increase, and vegetation could likely recover in the coming years. Thus, plant community management should be carefully carried out in the future.
Regardless of the dominant woody plant species, planting increases soil water content. Previous studies have shown that plants have prominent regulating and storage functions and that the water regulation exercised by plants becomes more pronounced as restoration is prolonged (Li et al., 2014). Afforestation increases vegetation cover and suppresses surface evaporation, thereby augmenting soil water content, which ultimately improves sand-driven wind speed and effectively reduces the impact of wind erosion in alpine deserts. Hence, the reasonable planting of artificial vegetation plays a pivotal role in improving the ecological environment in terms of its soil water content balance during alpine desert restoration.
Overall, vegetation growth and soil features of the alpine desert along the eastern shore of Qinghai Lake have gradually improved after desertification control measures were implemented. The species diversity of the community has increased with the extension of the fixed growth time of the dunes, while the coverage of herbaceous plants has also increased. Soil surface deposits were mainly concentrated in medium sands, whose organic matter and nutrient content are both low. Influenced by precipitation, soil moisture reaches its highest state on natural dunes rather than mobile dunes but lacks interannual variation for all dune types. However, the vertical distribution of soil moisture is seasonal, showing variation from precipitation and exhibiting interannual variability. The dynamic patterns of vegetation and soil properties are similar to those of other deserts in northern China, despite the severe impact of climatic conditions on plant survival in the alpine desert, which makes it more difficult to control the expansion of desertification.
The raw data supporting the conclusions of this article will be made available by the authors without undue reservation.
HZ started the formal analysis and wrote the original draft. LT played a guiding role in conceptualizing the field experiment and editing the manuscript. EH helped with editing and reviewing the manuscript. DZ and WW assisted in the data curation and editing of the manuscript.
This research was funded by the Chinese Western Youngest Scientist Program supported by CAS (2021_1_1), the Leading Talent Program supported by Qinghai Province (2022), the Qinghai Three Rivers Ecological Protection and Construction Phase II (2018-S-1), and the Open Project of the State Key Laboratory of Plateau Ecology and Agriculture, Qinghai University (2022-KF-07).
We thank Dengfeng Zhang for his kind assistance with this study’s field experiments.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations or those of the publisher, editors, or reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Bayat, H., Rastgo, M., Mansouri Zadeh, M., and Vereecken, H. (2015). Particle size distribution models, their characteristics and fitting capability. J. Hydrology 529, 872–889. doi:10.1016/j.jhydrol.2015.08.067
Blott, S. J., and Pye, K. (2001). Gradistat: A grain size distribution and statistics package for the analysis of unconsolidated sediments. Earth Surf. Process. Landforms 26, 1237–1248. doi:10.1002/esp.261
Cui, B. L., and Li, X. Y. (2015). Stable isotopes reveal sources of precipitation in the Qinghai Lake Basin of the northeastern Tibetan Plateau. Sci. Total Environ. 527-528, 26–37. doi:10.1016/j.scitotenv.2015.04.105
D’odorico, P., Bhattachan, A., Davis, K. F., Ravi, S., and Runyan, C. W. (2013). Global desertification: Drivers and feedbacks. Adv. Water Resour. 51, 326–344. doi:10.1016/j.advwatres.2012.01.013
Dormaar, J. F., and Willms, W. D. (1990). Sustainable production from the Rough Fescue Prairie. J. of Soil and Water Conservation 45, 4.
Fernandez-Illescas, C. P., Porporato, A., Laio, F., and Rodriguez-Iturbe, I. (2001). The ecohydrological role of soil texture in a water-limited ecosystem. Water Resour. Res. 37, 2863–2872. doi:10.1029/2000wr000121
Gu, C., Mu, X., Gao, P., Zhao, G., Sun, W., Tatarko, J., et al. (2018). Influence of vegetation restoration on soil physical properties in the Loess Plateau, China. J. Soils Sediments 19, 716–728. doi:10.1007/s11368-018-2083-3
Hill, A. R. (1975). Ecosystem stability in relation to stress caused by human activities. Canadian Geographer 3, 15.
Huang, J., Zhang, G., Zhang, Y., Guan, X., Wei, Y., and Guo, R. (2020). Global desertification vulnerability to climate change and human activities. Land Degrad. Dev. 31, 1380–1391. doi:10.1002/ldr.3556
Jiang, Z.-Y., Li, X.-Y., Wei, J.-Q., Chen, H.-Y., Li, Z.-C., Liu, L., et al. (2018). Contrasting surface soil hydrology regulated by biological and physical soil crusts for patchy grass in the high-altitude alpine steppe ecosystem. Geoderma 326, 201–209. doi:10.1016/j.geoderma.2018.04.009
Li, S., and Sawada, Y. (2022). Soil moisture-vegetation interaction from near-global in-situ soil moisture measurements. Environ. Res. Lett. 17, 114028. doi:10.1088/1748-9326/ac9c1f
Li, X., Hui, R., Tan, H., Zhao, Y., Liu, R., and Song, N. (2021). Biocrust research in China: Recent progress and application in land degradation control. Front. Plant Sci. 12, 751521. doi:10.3389/fpls.2021.751521
Li, X., Yang, X., Ma, Y., Hu, G., Hu, X., Wu, X., et al. (2018). Qinghai Lake basin critical zone observatory on the Qinghai-Tibet Plateau. Vadose Zone J. 17, 1–11. doi:10.2136/vzj2018.04.0069
Li, X., Zhang, Z., Huang, L., and Wang, X. (2013). Review of the ecohydrological processes and feedback mechanisms controlling sand-binding vegetation systems in sandy desert regions of China. Chin. Sci. Bull. 58, 1483–1496. doi:10.1007/s11434-012-5662-5
Li, X., Zhang, Z., Tan, H., Gao, Y., Liu, L., and Wang, X. (2014). Ecological restoration and recovery in the wind-blown sand hazard areas of northern China: Relationship between soil water and carrying capacity for vegetation in the tengger desert. Sci. China Life Sci. 57, 539–548. doi:10.1007/s11427-014-4633-2
Lu, R., Tang, Q., and Zhang, D. (2013). Rainwater infiltration at dunes under various rainfall events in sandy land to the East of Qinghai Lake. J. Desert Res. 33, 6.
Maxwell, T. M., Silva, L. C. R., and Horwath, W. R. (2018). Integrating effects of species composition and soil properties to predict shifts in montane forest carbon-water relations. Proc. Natl. Acad. Sci. U. S. A. 115, E4219–E4226. doi:10.1073/pnas.1718864115
Noy-Meir, I. (1973). desert ecosystems: Environment and producers. Annu. Rev. Ecol. Syst. 4, 25–51. doi:10.1146/annurev.es.04.110173.000325
Pan, Y.-X., and Wang, X.-P. (2009). Factors controlling the spatial variability of surface soil moisture within revegetated-stabilized desert ecosystems of the Tengger Desert, Northern China. Hydrol. Process. 23, 1591–1601. doi:10.1002/hyp.7287
Pan, Y.-X., Wang, X.-P., Zhang, Y.-F., and Hu, R. (2015). Spatio-temporal variability of root zone soil moisture in artificially revegetated and natural ecosystems at an arid desert area, NW China. Ecol. Eng. 79, 100–112. doi:10.1016/j.ecoleng.2015.04.019
Porporato, A., Odorico, P. D., Laio, F., Ridolfi, L., and Rodriguez-Iturbe, I. (2002). Ecohydrology of water-controlled ecosystems. Adv. Water Resour. 25, 1335–1348. doi:10.1016/s0309-1708(02)00058-1
Qi, Y., Chen, T., Pu, J., Yang, F., Shukla, M. K., and Chang, Q. (2018). Response of soil physical, chemical and microbial biomass properties to land use changes in fixed desertified land. Catena 160, 339–344. doi:10.1016/j.catena.2017.10.007
Renne, R. R., Bradford, J. B., Burke, I. C., and Lauenroth, W. K. (2019). Soil texture and precipitation seasonality influence plant community structure in North American temperate shrub steppe. Ecology 100, e02824. doi:10.1002/ecy.2824
Rodriguez-Iturbe, I., D'odorico, P., Porporato, A., and Ridolfi, L. (1999). On the spatial and temporal links between vegetation, climate, and soil moisture. Water Resour. Res. 35, 3709–3722. doi:10.1029/1999wr900255
Tian, L., Wang, H., Zhang, D., Wang, Q., and Liu, R. (2021). Water use patterns for a typical afforested shrub among topographic positions in an alpine desert of Qinghai-Tibet Plateau using stable isotopes tracers. Acta Ecol. Sin. 41, 12.
Tian, L., Wu, W., Zhang, D., Lu, R., and Wang, X. (2015). Characteristics of erosion and deposition of straw checkerboard barriers in alpine sandy land. Environ. Earth Sci. 74, 573–584. doi:10.1007/s12665-015-4059-6
Tian, L., Wu, W., Zhang, D., and Yu, Y. (2020). Airflow field around Hippophae rhamnoides in alpine semi-arid desert. Land 9, 140. doi:10.3390/land9050140
Tian, L., Wu, W., Zhou, X., Zhang, D., Yu, Y., Wang, H., et al. (2019). The ecosystem effects of sand-binding shrub Hippophae rhamnoides in alpine semi-arid desert in the northeastern Qinghai–Tibet Plateau. Land 8, 183. doi:10.3390/land8120183
Turnbull, L., Wilcox, B. P., Belnap, J., Ravi, S., D'odorico, P., Childers, D., et al. (2012). Understanding the role of ecohydrological feedbacks in ecosystem state change in drylands. Ecohydrology 5, 174–183. doi:10.1002/eco.265
Walkley, A. J., and Black, I. A. (1934). Estimation of soil organic carbon by the chromic acid titration method. Soil Sci. 37, 10.
Wan, L., Cao, W., Hu, F., Liang, S., and Jin, X. (2005). Eco-hydrology and eco-hydrogeology. Geol. Bull. China 24, 4.
Wang, H., Tian, L., Zhang, H., Yu, Y., and Wu, H. (2022). Water uptake by Artemisia ordosica roots at different topographic positions in an alpine desert dune on the northeastern Qinghai–Tibet Plateau. Front. Earth Sci. 10. doi:10.3389/feart.2022.686441
Wang, X.-P., Li, X.-R., Xiao, H.-L., and Pan, Y.-X. (2005). Evolutionary characteristics of the artificially revegetated shrub ecosystem in the Tengger Desert, northern China. Ecol. Res. 21, 415–424. doi:10.1007/s11284-005-0135-9
Wang, X. P., Wang, Z. N., Berndtsson, R., Zhang, Y. F., and Pan, Y. X. (2011). Desert shrub stemflow and its significance in soil moisture replenishment. Hydrology Earth Syst. Sci. 15, 561–567. doi:10.5194/hess-15-561-2011
Wu, H., Li, X.-Y., Jiang, Z., Chen, H., Zhang, C., and Xiao, X. (2016). Contrasting water use pattern of introduced and native plants in an alpine desert ecosystem, Northeast Qinghai–Tibet Plateau, China. Sci. Total Environ. 542, 182–191. doi:10.1016/j.scitotenv.2015.10.121
Wu, H., Li, X. Y., Zhang, J., Li, J., Liu, J., Tian, L., et al. (2019). Stable isotopes of atmospheric water vapour and precipitation in the northeast Qinghai-Tibetan Plateau. Hydrol. Process. 33, 2997–3009. doi:10.1002/hyp.13541
Wu, W., Chen, G., Meng, T., Li, C., Feng, H., Si, B., et al. (2023). Effect of different vegetation restoration on soil properties in the semi-arid Loess Plateau of China. Catena 220, 106630. doi:10.1016/j.catena.2022.106630
Wu, W., Zhang, D., Tian, L., Gao, S., Zhang, M., and Zhou, X. (2019a). Erosion-deposition characteristics and fencing life of straw checkerboard in alpine sandy lands. Environ. Eng. Sci. 36, 60–70. doi:10.1089/ees.2018.0155
Wu, W., Zhang, D., Tian, L., Zhang, M., and Zhou, X. (2019b). Features of artificial plant communications from the east sand region of the Qinghai Lake over the last 10 years. Acta Ecol. Sin. 39, 13.
Xu, X., Ma, K., Li, C., Fu, B., Liu, X., Huang, Y., et al. (2006). Research review of the relationship between vegetation and soil loss. Acta Ecologica Sinica 26, 7.
Yu, X., Huang, Y., Li, E., Li, X., and Guo, W. (2017). Effects of vegetation types on soil water dynamics during vegetation restoration in the Mu Us Sandy Land, northwestern China. J. Arid Land 9, 188–199. doi:10.1007/s40333-017-0054-y
Yu, Y., and Jia, Z. Q. (2014). Changes in soil organic carbon and nitrogen capacities of Salix cheilophila Schneid along a revegetation chronosequence in semi-arid degraded sandy land of the Gonghe Basin, Tibet Plateau. Solid Earth. 5, 1045–1054. doi:10.5194/se-5-1045-2014
Yu, Y., Wei, W., Chen, L., Feng, T., Daryanto, S., and Wang, L. (2016). Land preparation and vegetation type jointly determine soil conditions after long-term land stabilization measures in a typical hilly catchment, Loess Plateau of China. J. Soils Sediments 17, 144–156. doi:10.1007/s11368-016-1494-2
Yu, Y., Zhao, W., Martinez-Murillo, J. F., and Pereira, P. (2020). Loess Plateau: From degradation to restoration. Sci. Total Environ. 738, 140206. doi:10.1016/j.scitotenv.2020.140206
Zhou, G., Xia, J., Zhou, P., Shi, T., and Li, L. (2020). Not vegetation itself but mis-revegetation reduces water resources. Sci. China Earth Sci. 51, 8.
Zhu, B.-Q., Yu, J.-J., Rioual, P., and Ren, X.-Z. (2014). Particle size variation of aeolian dune deposits in the lower reaches of the Heihe River basin, China. Sediment. Geol. 301, 54–69. doi:10.1016/j.sedgeo.2013.12.006
Keywords: vegetation communities, soil properties, soil moisture, sand dunes, Qinghai Lake
Citation: Zhang H, Tian L, Hasi E, Zhang D and Wu W (2023) Vegetation–soil dynamics in an alpine desert ecosystem of the Qinghai Lake watershed, northeastern Qinghai–Tibet Plateau. Front. Environ. Sci. 11:1119605. doi: 10.3389/fenvs.2023.1119605
Received: 09 December 2022; Accepted: 03 February 2023;
Published: 16 February 2023.
Edited by:
Hu Liu, Northwest Institute of Eco-Environment and Resources (CAS), ChinaReviewed by:
Dejin Wang, Kunming University of Science and Technology, ChinaCopyright © 2023 Zhang, Tian, Hasi, Zhang and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lihui Tian, bGh0aWFuQHFodS5lZHUuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.