 Yan Yang
Yan Yang Kari Klanderud
Kari Klanderud Yinghui Yang3
Yinghui Yang3 Tongzuo Zhang
Tongzuo Zhang Genxu Wang
Genxu Wang- 1Key Laboratory of Mountain Surface Processes and Ecological Regulation, Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu, China
- 2Faculty of Environmental Sciences and Natural Resource Management, Norwegian University of Life Sciences, Ås, Norway
- 3School of Mathematics, Southwest Jiaotong University, Chengdu, China
- 4Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining, China
- 5State Key Laboratory of Hydraulics and Mountain River Engineering, College of Water Resource and Hydropower, Sichuan University, Chengdu, China
The Tibetan Plateau is the most extensive high-elevation grassland on Earth, with the largest expanse of high-elevation permafrost. It is experiencing climate warming that is projected to continue at rates above the global mean, potentially jeopardizing ecosystem functioning. We conducted a broad-scale resampling project in the permafrost region of Tibet to examine if plant production and diversity had changed over time. We recorded vascular plant species occurrences and harvested aboveground biomass at 36 alpine grassland sites in 2008 and 2018. Our results show that aboveground biomass increased for legumes and forbs, but decreased for grasses and sedges, resulting in no overall change in the aboveground biomass during the 10-year period. Our results indicate that functional group abundance may shift from grasses and sedges toward more legumes and forbs, and that species composition is becoming more similar between grassland types, and thus, beta diversity is decreasing in the permafrost region of Tibet.
Introduction
Alpine ecosystems are temperature-limited systems and predicted to be sensitive to climate change (Elmendorf et al., 2015). The projected rate of future warming in the alpine region is also faster than that of the global average (Stocker et al., 2014). Thus, understanding how climate warming affects alpine communities is especially important for our ability to predict impacts of future climate change. An increasing number of studies report shifts in vegetation composition, biomass, and diversity in arctic and alpine tundra (Klein et al., 2004; Walker et al., 2006; Elmendorf et al., 2012a; Elmendorf et al., 2012b; Liu et al., 2018; Yang et al., 2018), and these changes will have cascading effects on the structure and function of alpine ecosystems (Yang et al., 2011; Bjorkman et al., 2018; Niittynen et al., 2020; Wu et al., 2021). For example, experimental warming increases primary productivity and decreases species numbers in the arctic and alpine plant communities (Klein et al., 2004; Elmendorf et al., 2012a; Yang et al., 2015), shifts abundances of different functional groups (Ganjurjav et al., 2016; Liu et al., 2018), and lengthens the growing season leading to advancing leaf and reproduction phenology, and delayed withering (Cleland et al., 2007; Dorji et al., 2013; Meng et al., 2016; Prevéy et al., 2018; Lian et al., 2020). Rapid and accelerating increases in species richness over the past century have been reported in European mountaintops (Steinbauer et al., 2018), and remote sensing shows vegetation change over the same period in alpine ecosystems (Pattison et al., 2015; Zhu et al., 2016; Verbyla and Kurkowski 2019; May et al., 2020), likely as a result of climate warming.
Most resurvey studies have so far been performed in European mountains, and in areas without permafrost, and observations of changes in plant community composition and productivity over time are confined to site-level experiments (Walker et al., 2006; Elmendorf et al., 2012a; Bjorkman et al., 2018; Yang et al., 2018). Interestingly, a synthesis of long-term monitoring studies across multiple alpine and arctic sites found that forb responses to climate warming differed between sites with and without permafrost (Elmendorf et al., 2012b). Thus, there is a need for more knowledge on recent changes in plant community productivity and species richness across larger permafrost regions to understand how changes in such areas may affect carbon cycling and biodiversity under climate change.
Changes in the aboveground net primary productivity (ANPP) is a crucial factor affecting the carbon cycling of terrestrial ecosystems (Oberbauer et al., 2007; Rodriguez et al., 2018), and may influence the carbon feedback between the terrestrial biosphere and the atmosphere (Shen et al., 2015). Over the past decades, many experimental studies have explored how plant community composition and primary productivity respond to climate change (Franklin et al., 2016; Liu et al., 2017; Ma et al., 2017). Regional-scale evidence for changes in plant productivity has mainly come from remote-sensing data (Piao et al., 2006; Pouliot et al., 2009; Gao et al., 2019) and plant community dynamics from plot-based observations with experimental manipulation (Elmendorf et al., 2012b). There are, however, limitations in both remotely sensed data as these do not identify species-level productivity and diversity (Zhang G. et al., 2013), and in experimental manipulations, which do not reflect realistic climate change (Korell et al., 2019). Broad-scale resurvey studies with sufficient replication of sites can detect region-scale changes in plant species richness (Klanderud and Birks 2003; Gottfried et al., 2012; Pauli et al., 2012; Steinbauer et al., 2018), but few works have historic data for biomass (Ding et al., 2017). Observations of changes in plant community composition and productivity over time are confined to site-level experiments (Walker et al., 2006; Elmendorf et al., 2012a; Bjorkman et al., 2018; Yang et al., 2018).
The Tibetan Plateau has the largest expanse of high-elevation permafrost in the world and has unique ecosystems typically characterized by low temperature, low atmospheric pressure, and oxygen concentrations, and are experiencing climate warming that may jeopardize the functioning of the ecosystems (Klein et al., 2014; He et al., 2021). The presence or absence of permafrost determines the structure and nutrient status of the soil and affects plant community structure and production (Yi et al., 2011; Yang et al., 2018). The grasslands in the permafrost region of the Tibetan Plateau support a diverse assemblage of wildlife and pastoralism as a primary livelihood activity (Klein et al., 2014; Rodriguez et al., 2018). A vegetation greening measured by normalized difference vegetation index (NDVI) in the alpine grasslands of the Tibetan Plateau (Xu et al., 2008; Zhong et al., 2010; Peng et al., 2012; Zhang L. et al., 2013; Zhang et al., 2017) suggests increased plant growth over the past decades. Remotely sensed metrics, such as NDVI, is, however, unable to detect finer-scale changes in plant species diversity and biomass of different functional groups, which are important for understanding ecosystem change (Hopping et al., 2018). Thus, to examine recent changes in functional group biomass and species diversity in Tibet, we performed a resampling study of 36 sites surveyed for the first time in 2008. By revisiting the same sites and using methods as similar as possible to the original survey, we repeated the survey in 2018 to examine the following questions: 1) Have plant community diversity and evenness, biomass, cover, and species richness of different functional groups changed during the 10 years? 2) Are there differences in change between grassland types? We tested the hypothesis that climate changes over the past decades increased plant biomass and species richness and diversity in the permafrost region of Tibet, and that graminoids increased more in abundance than forbs, as observed in climate warming experiments in other alpine and arctic regions (Elmendorf et al., 2012a; Yang et al., 2018).
Material and Methods
Study Area
Permafrost on the Tibetan Plateau covers 1.5 million km2, approximately half the area of the plateau. It accounts for 70.5% of the permafrost area in high Asia and 75% of the permafrost in the Northern Hemisphere mountains (Yang et al., 2010). The mean annual air temperature on the plateau ranges from −3.1 to 4.4°C, and the mean annual precipitation ranges from 103 to 694 mm (Ding et al., 2017). During the past three decades, the plateau has experienced an increase in the mean annual air temperature of 0.5°C per decade and soil temperature of 0.6°C per decade at 10 cm depths. Precipitation and soil moisture also increased over the past three decades (Ding et al., 2017). Observational data from monitoring stations on the plateau indicate that the mean annual permafrost temperature at 6.0 m depth increased by 0.43°C from 1996 to 2006 at an average annual rate of 0.04°C (Wu and Zhang 2008), and increased by 0.1°C from 2006 to 2010 at an average annual rate of 0.02°C (Wu et al., 2012). Alpine grasslands, which consist primarily of alpine meadow and alpine steppe vegetation, cover more than two-thirds of the plateau (Xu et al., 2017). In our 36 resurvey sites, the dominant species in the alpine meadow are cold-tolerant sedges, such as Kobresia pygmaea, K. humilis, and K. tibetica, and in the alpine steppe, common species are the grass Stipa purpurea and the sedge Carex moorcroftii. Major soil types are cambisols in the meadow and xerosols in the steppe (Lu et al., 2004).
Vegetation Resampling
To detect changes in plant community biomass and species richness across the Tibetan alpine permafrost regions, we compiled a broad-scale repeated-sampling based on the field campaigns in 2008 and 2018. Both campaigns were conducted during the growing season from 20 July to 20 August. The sampling sites span elevations from 4,300 to 5,130 m, and were set along the Qinghai–Xizang Highway to cover the permafrost area across a 600-km range from the northern boundary of permafrost at Xidatan to the southern boundary at Anduo (Zou et al., 2017). Each of the 36 sites was surveyed in both 2008 and 2018 (Supplementary Figure S1). Plant community survey and biomass sampling was performed using the same sampling scheme in both years. Specifically, the sites were established 1 km away from the highway, and consisted of six 1 × 1 m2quadrats randomly located in a 20 × 20 m2 plot at each site in 2008. In 2018, we established six new 1 × 1 m2 quadrats in the same 20 × 20 m2 plot. We recorded plant species composition and cover in each 1 × 1 m2 quadrat, and harvested live aboveground biomass of each species at peak biomass in both years. We harvested all individuals of each species in each quadrate, and put each species of each quadrate in separate paper bags. All the samples were oven dried at 70°C and weighted on a semi-analytical balance in the lab.
Climate and Grazing Data
The monthly air temperature and precipitation data for each site between 2008 and 2018 were extracted from the nearest grids in the surface gridded product (ground resolution 0.5°) of the China Meteorological Data Service Center of National Meteorological Information Center, China (http://data.cma.cn/data/cdcdetail/dataCode/SURF_CLI_CHN_TEM_MON_GRID_0.5.html;http://data.cma.cn/data/cdcdetail/dataCode/SURF_CLI_CHN_PRE_MON_GRID_0.5.html). Given the potential grazing effects on vegetation during the resurvey period, data on the numbers of grazing livestock were collected by grassland monitoring stations in Qumalai county and Tanggula area for each year from 2008 to 2018, which covered the majority of the resurveyed sites in this study (Supplementary Figure S1).
Statistical Analyses
We calculated the total plant community biomass, number of species, and cover of different functional groups for each plot and year. Diversity, using Shannon’s diversity index (H), was calculated as H = −∑pi (ln [pi]), where pi is the proportion of a particular species cover in plot. Evenness (E) was calculated from diversity (H) and species richness (S) as E = log(H)/ln(S).
We obtained change values of paired samples between 2018 and 2008. We transformed the change values into relative change ratios by dividing the mean values over the study period and then we calculated the estimates and 95% confidence intervals of the change ratios for all the response variables. Thus, significant changes in these variables between 2008 and 2018 are shown if the confidence intervals are not crossing the zero line. Change in aboveground biomass, species richness, plant cover, diversity, and evenness over time was analyzed separately using repeated-measures mixed effects models with the year and the interaction between year and grassland type (alpine meadow and alpine steppe) as fixed effects and plot nested in block as random effects.
To examine if species richness changed more in some plant families than others, we tested difference in species numbers for each family between 2008 and 2018 using a t-test. To determine the effects of year on species composition of all species, we performed permutational multivariate analyses of variance (PERMANOVA) on the species abundance using Bray–Curtis distance matrices. For testing the effect of years between 2008 and 2018, we defined a restricted permutation design with sampling within plots in different years permuted as time series. Furthermore, to visualize differences in community composition between 2008 and 2018, we performed a nonmetric multidimensional scaling analysis using Bray–Curtis distance matrices, three dimensions (k = 3) to meet the stress criterion of <0.2 (Clarke 1993). All analyses were performed in R version 4.0.0 (R Development Core Team, 2019).
Results
There was a gradual decrease in summer precipitation and increase in summer temperature during the study period from 2008 to 2018 across all sites (Figures 1A,B). Although the total aboveground biomass did not change over time (Table 1), some functional groups changed depending on the grassland type (Table 1; Figure 2). Specifically, we found a marginal increase in legume biomass in steppe and forb biomass in both steppe and meadow, and a decreasing tendency in grass and sedge biomass between 2008 and 2018 in both meadow and steppe (Figure 2).
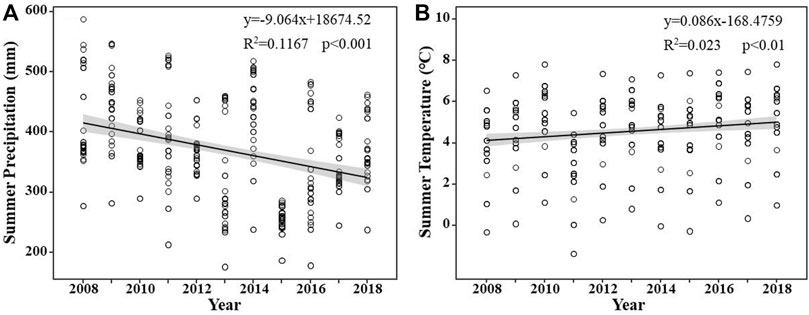
FIGURE 1. Summer precipitation (A) and air temperature (B) between 2008 and 2018 in all the sites we surveyed in the permafrost region of the Tibetan Plateau. Solid circle, alpine meadow; blank circle, alpine steppe.
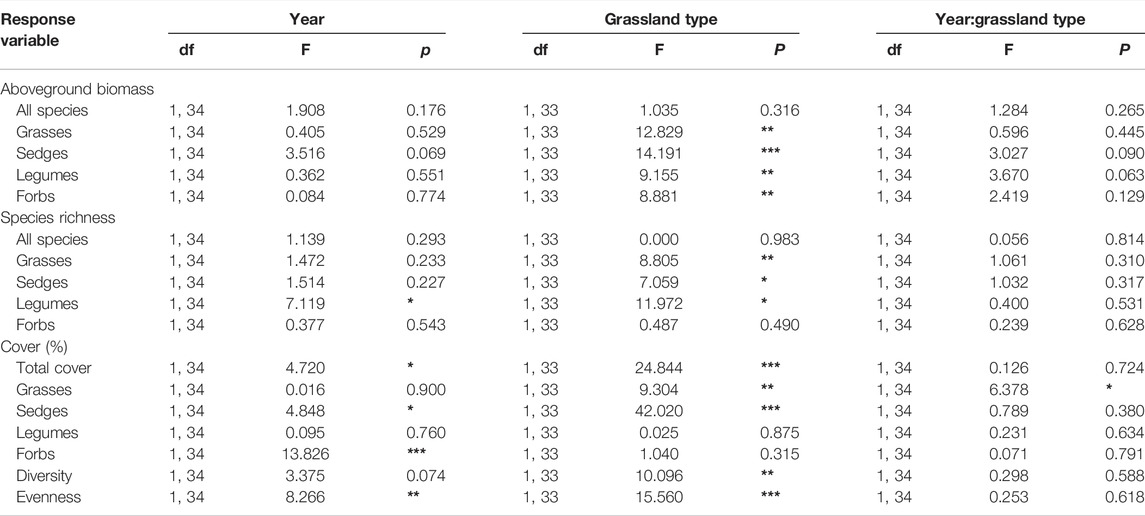
TABLE 1. Results of the mixed effect model testing for the effects of year (2008 vs. 2018), grassland type (meadow vs. steppe), and the interaction between year and grassland type on aboveground biomass, species richness, and plant cover of all species, grasses, sedges, legumes, and forbs, and diversity and evenness. Asterisks denote significant differences at p < 0.05*, p < 0.01**, p < 0.001***.
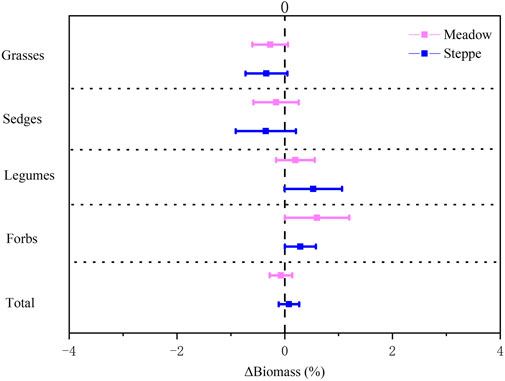
FIGURE 2. Changes in biomass of grasses, sedges, legumes, forbs, and total biomass in alpine meadow (red) and steppe (blue) from 2008 to 2018 in the permafrost region of the Tibetan Plateau. Relative change rate in aboveground biomass (ΔBiomass %) was calculated as the ratio of the absolute change (in units of g m−2) to the mean biomass over the study period. Data are shown as estimate ±95% confidence intervals (CI). Data greater than zero represent a significant increase in the measured variable, and data lower than zero represent a significant decrease between 2008 and 2018.
Total species richness increased at 22 of the 36 sites between 2008 and 2018, with a mean increase of 2.1 species per site, but this was not statistically significant across the grassland types (Table 1; Figure 3). Only the number of legume species decreased significantly over time, particularly in the meadows (Table 1; Figure 3). There was also a tendency toward decreased species richness of grasses and sedges in the meadows, leading to a decrease in total species richness here (Figure 3). In the steppe, on the other hand, forbs and total species richness appeared to increase between 2008 and 2018 (Figure 3). When testing the changes per plant family, there was a significant increase in species richness for Asteraceae, Scrophulariaceae, Gentianaceae, Umbelliferae, Crassulaceae, Papaeraceae, Ranunculaceae, and Liliaceae species between 2008 and 2018 (Supplementary Table S1).
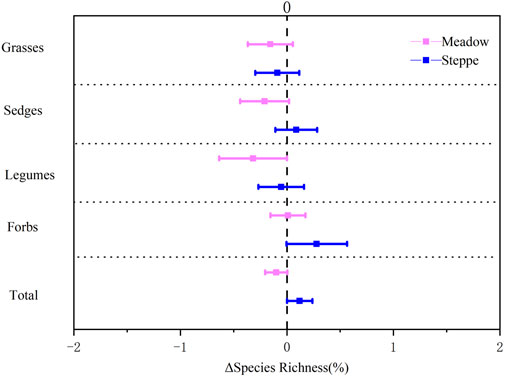
FIGURE 3. Change in species richness of grasses, sedges, legumes, forbs, and all species in alpine meadow (red) and steppe (blue) from 2008 to 2018 in the permafrost region of the Tibetan Plateau. Relative change rate in the number of species (ΔSpecies Richness) was calculated as the ratio of the absolute change (in units of species m−2) to the mean species richness over the study period. Data are shown as estimate ±95% confidence intervals (CI). Data greater than zero represent a significant increase in the measured variable, and data lower than zero represent a significant decrease between 2008 and 2018.
Legume, forb, and total vegetation cover increased over time in both meadow and steppe (Table 1; Figure 4). Diversity appeared to increase in the steppe (Figure 5), whereas evenness increased in both the grassland types (Table 1; Figure 5).
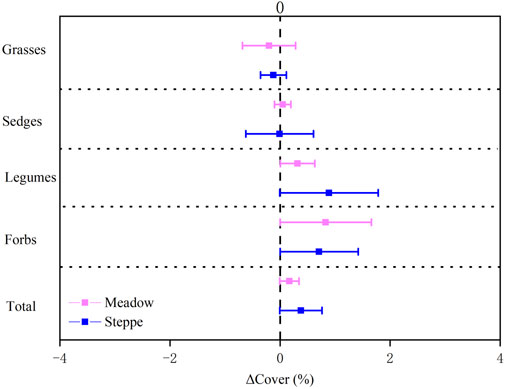
FIGURE 4. Change in grass, sedge, legume, forb, and total plant cover in alpine meadow (red) and steppe (blue) from 2008 to 2018 in the permafrost region of the Tibetan Plateau. The relative change rate in cover (ΔCover, %) was calculated as the ratio of the absolute change (in units of %) to the mean cover value over the study period. Data were shown as estimate ±95% confidence intervals (CI). Data greater than zero represent a significant increase in the measured variable, and data lower than zero represent a significant decrease between 2008 and 2018.
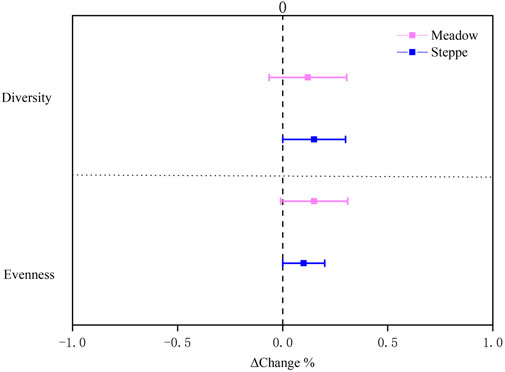
FIGURE 5. Change in diversity and evenness in alpine meadow (red) and steppe (blue) grasslands from 2008 to 2018 in the permafrost region of the Tibetan Plateau. Relative change rate (ΔChange) was calculated as the ratio of the absolute change to the mean value over the study period. Data were shown as estimate ±95% confidence intervals (CI). Data greater than zero represent a significant increase in measured variable, and lower than zero represent a significant decrease between 2008 and 2018.
The NMDS showed that species composition is clearly distinguished between the two grassland types, meadow and steppe, but that species composition is becoming slightly more similar over time (Figure 6; Table 2).
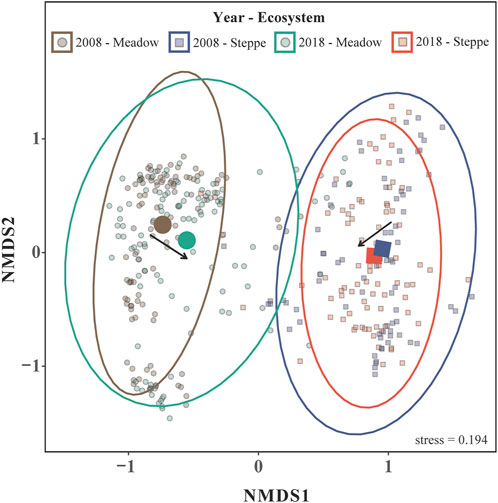
FIGURE 6. Nonmetric dimensional scaling (NMDS) analysis showing differences in plant species composition between 2008 and 2018 for alpine meadow and steppe communities in the permafrost region of the Tibetan Plateau along the first and second NMDS axes.
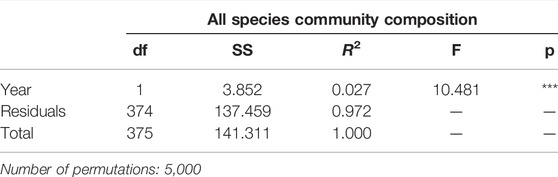
TABLE 2. Results of permutational multivariate analysis of variance (PERMANOVA) using Bray–Curtis distance matrices, testing for the effect of year on community species composition from both 2008 and 2018. Asterisks denote significant differences at p < 0.01 (**), p < 0.001 (***).
Discussion
Our resampling of alpine grasslands in the Tibetan Plateau shows that the total community-level primary production and total species richness did not change over the 10-year study period (Table 1), in contrast to our hypothesis, but vegetation cover and evenness increased. However, the different functional groups showed different tendencies over 10 years, with grasses and sedges tending to decrease, and forbs and legumes increasing in biomass and cover, in contrast to our hypothesis.
The increase of forbs in our study is, interestingly, in line with Elmendorf et al. (2012b), who found that forb abundance increased under climate warming only in sites with permafrost. These results are also in line with the warming experiments in the same mesic meadows, which showed decreased grass and sedge, and increased legume and forb productivity (Yang et al., 2015), likely due to the interspecific competition rather than a direct response to warming. These results may also show compensatory interactions among functional groups (Bai et al., 2004; Connell and Ghedini, 2015; Liu et al., 2017), suggesting that a species increase in abundance is a response to the reduction of another in a fluctuating environment (Naeem and Li 1997). The lack of change in total biomass from 2008 to 2018 in our study is in line with a previous study resampling biomass in the same region (Ding et al., 2017) and in a long-term monitoring experiment in a mesic meadow of northeastern Tibet (Liu et al., 2017; Wang et al., 2020). These results are, however, in contrast to alpine and arctic tundra elsewhere, where increased height of the vegetation canopy in most areas indicate increasing biomass (Elmendorf et al., 2012a; Myers-Smith et al., 2015). One important difference between our study area in Tibet and alpine and arctic tundra elsewhere is the more extensive grazing by livestock in the Tibetan Plateau. This grazing pressure has been relatively stable during the last decades (Supplementary Figure S2), and may have buffered an increase in biomass due to climate change. The basic characteristic of degraded alpine grasslands is decreased numbers of graminoids (Zhou et al., 2005), as we found in our study (Figure 3; Table 1). Many of the forb species that increased in cover (Figure 4; Table 1), such as the Cremanthodium liheare, Saussurea graminea, Leontopodium leontopodioides, Ajania tenuifolia, Pedicularis kansuensis, and Gentiana leucomelaena Maxim., are toxic weeds and not preferred by animals (Zhao 2009; Wang and Kang 2011). Thus, the decrease in grass and sedge biomass observed in our study and others, and the increase in forbs (Figure 2), may lead to a decrease in forage quality in the alpine grasslands of Tibet.
Changes in species richness mainly depended on grassland type in our study (Table 1). Number of grass, sedge, and legume species showed a decreasing tendency, leading to decreased total species richness in the meadow, which is in line with the results of in situ warming experiments reporting decreased species richness in Tibet (Klein et al., 2004; Yang et al., 2018) and in other alpine and arctic regions (Hollister et al., 2005). However, the overall community diversity and evenness increased, and, although not statistically significant, species richness increased in 61% of the sites we resurveyed, most of them in the steppe grasslands. These results are in line with the resurveys of European mountain summits, where the number of species increased during recent decades (Pauli et al., 2012; Steinbauer et al., 2018). The lack of a strong species richness effect may be due to the relatively short period of 10 years in our study. Interestingly, species composition in the two grassland types became more similar during the 10 years (Figure 6; Table 2). This homogenization is in line with a resampling study from alpine Norway, which also showed homogenization in species composition over time (Odland et al., 2010).
The increase in the total plant cover in our study is in line with increased greening measured as NDVI in the alpine grassland of Tibet (Peng et al., 2012; Zhang et al., 2017). The lack of change in the total biomass production, thus, indicates that NDVI cannot simply be translated into biomass.
Summer precipitation decreased and summer temperature increased in the study area during the study period (Figure 1). The observed decreased tendency in grasses and sedges may be due to shallow roots and thus difficulty in acquiring deeper soil water than the more deep-rooted forbs (Klein et al., 2007). Deep soil moisture and temperature have been shown to be the main drivers of legume species richness (Ganjurjav et al., 2016), and the increase in legume biomass in our study over time is in line with site-level experiments, finding that legumes increased in abundance after four years of warming (Peng et al., 2020). Legumes may increase their abundance in sites where soil moisture (Cowles et al., 2016; Ganjurjav et al., 2016) and nitrogen are limited by cryoturbation induced by warming, leading to nitrogen deficiency in the upper soil layer (Chang et al., 2017).
In conclusion, we found that overall biomass and species richness of the alpine grasslands of the permafrost region of Tibet studied here have been relatively stable during the last decade (Table 1). This can be due to the relatively short term of the study, but also due to a potential buffering by the grazing livestock. Homogenization in species composition over time indicated decreased beta diversity, and the observed change tendencies in different functional groups may suggest a decrease in forage quality through a shift from grasses and sedges to less preferred legumes and forbs. Given the important contribution of forage quality to local husbandry, these findings are important when formulating management policies for the permafrost region of Tibet.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author Contributions
YaY and GW collected samples and field data. YaY analyzed data, and YaY, YiY, and HJ drew figures. TZ and YL provided data about grazing and climate. YaY, KK, and GW designed or advised on the experiment and paper writing. YaY and KK led the study and all authors provided edits on the manuscript.
Funding
This study was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (No. XDA20050102 and No. XDA23060601), “Kunlun Talents ⋅ Senior Innovation and Entrepreneurship Talents” project of the People’s Government of Qinghai Province, and the National Natural Science Foundation of China (41571204, 41861134039, 41701081, and 42061013). Our gratitude extends to the administration bureau of three-River-Rource National Park, the Fenghuoshan Field Station of the Northwest Research Institute Co., Ltd. of the China Railway Engineering Corporation and Beiluhe Observation and Research Station on Frozen Soil Engineering and Environment, Northwest Institute of Eco-Environment and Resources, CAS, for their assistance in the field and the Alpine Ecosystem Observation and Experiment Station of Mt. Gongga, CAS, for their assistance with Laboratory work.
Acknowledgments
We thank Prof. Lianwen Zhao, Yao Xiao, and Tao Zhang for helping in early data analysis and field work and Tang Chuanchuan, Yang Kai, and Zeng Fang for helping in the laboratory. We appreciate Prof. Guoying Zhou in the Northwest Institute of Plateau Biology, CAS in the initial field work stage.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2022.932993/full#supplementary-material
References
Bai, Y. F., Han, X. G., Wu, J. G., Chen, Z. Z., and Li, L. H. (2004). Ecosystem Stability and Compensatory Effects in the Inner Mongolia Grassland. Nature 431, 181–184.
Bjorkman, A. D., Myers-Smith, I. H., Elmendorf, S. C., Normand, S., Rüger, N., Beck, P. S. A., et al. (2018). Plant Functional Trait Change across a Warming Tundra Biome. Nature 562 (7725), 57–62. doi:10.1038/s41586-018-0563-7
Chang, R. Y., Wang, G. X., Yang, Y. H., and Chen, X. P. (2017). Experimental Warming Increased Soil Nitrogen Sink in the Tibetan Plateau. J. Geophy. Res. Biogeo. 122.
Clarke, K. R. (1993). Non-parametric Multivariate Analyses of Changes in Community Structure. Austral Ecol. 18, 117–143. doi:10.1111/j.1442-9993.1993.tb00438.x
Cleland, E. E., Chuine, I., Menzel, A., Mooney, H. A., and Schwartz, M. D. (2007). Shifting Plant Phenology in Response to Global Change. Trends Ecol. Evol. 22, 357–365.
Connell, S. D., and Ghedini, G. (2015). Resisting Regime-Shifts: The Stabilising Effect of Compensatory Processes. Trend Ecol. Evol. 30 (9), 513–515.
Cowles, J. M., Wragg, P. D., Wright, A. J., Powers, J. S., and Tilman, D. (2016). Shifting Grassland Plant Community Structure Drives Positive Interactive Effects of Warming and Diversity on Aboveground Net Primary Productivity. Glob. Change Biol. 22, 741–749. doi:10.1111/gcb.13111
Ding, J., Chen, L., Ji, C., Hugelius, G., Li, Y., Liu, L., et al. (2017). Decadal Soil Carbon Accumulation across Tibetan Permafrost Regions. Nat. Geosci. 10, 420–424. doi:10.1038/ngeo2945
Dorji, T., Totland, Ø., Moe, S. R., Hopping, K. A., Pan, J. B., and Klein, J. A. (2013). Plant Functional Traits Mediate Reproductive Phenology and Success in Response to Experimental Warming and Snow Addition in Tibet. Glob. Change Biol. 19, 459–472.
Elmendorf, S. C., Henry, G. H. R., Hollister, R. D., Björk, R. G., Bjorkman, A. D., Callaghan, T. V., et al. (2012a). Global Assessment of Experimental Climate Warming on Tundra Vegetation: Heterogeneity over Space and Time. Ecol. Lett. 15, 164–175. doi:10.1111/j.1461-0248.2011.01716.x
Elmendorf, S. C., Henry, G. H. R., Hollister, R. D., Björk, R. G., Boulanger-Lapointe, N., Cooper, E. J., et al. (2012b). Plot-scale Evidence of Tundra Vegetation Change and Links to Recent Summer Warming. Nat. Clim. Change 2, 453–457. doi:10.1038/nclimate1465
Elmendorf, S. C., Henry, G. H. R., Hollister, R. D., Fosaa, A. M., Gould, W. A., Hermanutz, L., et al. (2015). Experiment, Monitoring, and Gradient Methods Used to Infer Climate Change Effects on Plant Communities Yield Consistent Patterns. Proc. Natl. Acad. Sci. USA. 112, 448–452.
Franklin, J., Serra-Diaz, J. M., Syphard, A. D., and Regan, H. M. (2016). Global Change and Terrestrial Plant Community Dynamics. Proc. Natl. Acad. Sci. U.S.A. 113, 3725–3734. doi:10.1073/pnas.1519911113
Ganjurjav, H., Gao, Q., Gornish, E. S., Schwartz, M. W., Liang, Y., Cao, X., et al. (2016). Differential Response of Alpine Steppe and Alpine Meadow to Climate Warming in the Central Qinghai-Tibetan Plateau. Agric. For. Meteorology 223, 233–240. doi:10.1016/j.agrformet.2016.03.017
Gao, M., Piao, S., Chen, A., Yang, H., Liu, Q., Fu, Y. H., et al. (2019). Divergent Changes in the Elevational Gradient of Vegetation Activities over the Last 30 Years. Nat. Commun. 10, 2970. doi:10.1038/s41467-019-11035-w
Gottfried, M., Pauli, H., Futschik, A., Akhalkatsi, M., Barančok, P., Benito Alonso, J. L., et al. (2012). Continent-wide Response of Mountain Vegetation to Climate Change. Nat. Clim. Change 2 (2), 111–115. doi:10.1038/nclimate1329
He, J.-S., Dong, S., Shang, Z., Sundqvist, M. K., Wu, G., and Yang, Y. (2021). Above-belowground Interactions in Alpine Ecosystems on the Roof of the World. Plant Soil 458, 1–6. doi:10.1007/s11104-020-04761-4
Hollister, R. D., Webber, P. J., and Tweedie, C. E. (2005). The Response of Alaskan Arctic Tundra to Experimental Warming: Differences between Short- and Long-Term Responses. Glob. Change Biol. 11, 525–536. doi:10.1111/j.1365-2486.2005.00926.x
Hopping, K. A., Yeh, E. T., and Harris, R. B.Gaerrang (2018). Linking People, Pixels, and Pastures: a Multi-Method, Interdisciplinary Investigation of How Rangeland Management Affects Vegetation on the Tibetan Plateau. Appl. Geogr. 94, 147–162. doi:10.1016/j.apgeog.2018.03.013
Klanderud, K., and Birks, H. J. B. (2003). Recent Increases in Species Richness and Shifts in Altitudinal Distributions of Norwegian Mountain Plants. Holocene 13 (1), 1–6. doi:10.1191/0959683603hl589ft
Klein, J. A., Harte, J., and Zhao, X.-Q. (2004). Experimental Warming Causes Large and Rapid Species Loss, Dampened by Simulated Grazing, on the Tibetan Plateau. Ecol. Lett. 7, 1170–1179. doi:10.1111/j.1461-0248.2004.00677.x
Klein, J., Harte, J., and Zhao, X. Q. (2007). Experimental Warming, not Grazing, Decreases Rangeland Quality on the Tibetan Plateau. Ecol. Appl. 17 (2), 541–557.
Klein, J. A., Hopping, K. A., Yeh, E. T., Nyima, Y., Boone, R. B., and Galvin, K. A. (2014). Unexpected Climate Impacts on the Tibetan Plateau: Local and Scientific Knowledge in Findings of Delayed Summer. Glob. Environ. Change 28, 141–152. doi:10.1016/j.gloenvcha.2014.03.007
Korell, L., Auge, H., Chase, J. M., Harpole, S., and Knight, T. M. (2019). We Need More Realistic Climate Change Experiments for Understanding Ecosystems of the Future. Glob. Change Biol. 26, 325–327. doi:10.1111/gcb.14797
Lian, X., Piao, S., Li, L. Z. X., Li, Y., Huntingford, C., Ciais, P., et al. (2020). Summer Soil Drying Exacerbated by Earlier Spring Greening of Northern Vegetation. Sci. Adv. 6, eaax0255. doi:10.1126/sciadv.aax0255
Liu, H., Mi, Z., Lin, L., Wang, Y., Zhang, Z., Zhang, F., et al. (2017). Shifting Plant Species Composition in Response to Climate Change Stabilizes Grassland Primary Production. Proc. Natl. Acad. Sci. U. S. A. 115 (16), 4051–4056. doi:10.1073/pnas.1700299114
Liu, H. Y., Mi, Z. R., Lin, L., Wang, Y. H., Zhang, Z. H., Zhang, F. W., et al. (2018). Shifting Plant Species Composition in Response to Climate Change Stabilizes Grassland Primary Production. Proc. Natl. Acad. Sci. USA. 115 (16), 4051–4058.
Lu, H. Y., Wu, N., Gu, Z., Wang, L., Wu, H., Wang, G., et al. (2004). Distribution of Carbon Isotope Composition of Modern Soils on the Qinghai-Tibetan Plateau. Biogeochem 70, 273–297. doi:10.1023/b:biog.0000049343.48087.ac
Ma, Z., Liu, H., Mi, Z., Zhang, Z., Wang, Y., Xu, W., et al. (2017). Climate Warming Reduces the Temporal Stability of Plant Community Biomass Production. Nat. Commun. 8, 15378. doi:10.1038/ncomms15378
May, J. L., Hollister, R. D., Betway, K. R., Harris, J. A., Tweedie, C. E., Welker, J. M., et al. (2020). NDVI Changes Show Warming Increases the Length of the Green Season at Tundra Communities in Northern Alaska: A Fine-Scale Analysis. Front. Plant Sci. 11, 1174. doi:10.3389/fpls.2020.01174
Meng, F., Cui, S., Wang, S., Duan, J., Jiang, L., Zhang, Z., et al. (2016). Changes in Phenological Sequences of Alpine Communities across a Natural Elevation Gradient. Agric. For. Meteorology 224, 11–16. doi:10.1016/j.agrformet.2016.04.013
Myers-Smith, I. H., Elmendorf, S. C., Beck, P. S. A., Wilmking, M., Hallinger, M., Blok, D., et al. (2015). Climate Sensitivity of Shrub Growth across the Tundra Biome. Nat. Clim. Change 5, 887–891. doi:10.1038/nclimate2697
Naeem, S., and Li, S. (1997). Biodiversity Enhances Ecosystem Reliability. Nature 390, 507–509. doi:10.1038/37348
Niittynen, P., Heikkinen, R. K., and Luoto, M. (2020). Decreasing Snow Cover Alters Functional Composition and Diversity of Arctic Tundra. Proc. Natl. Acad. Sci. U. S. A. 117 (35), 21480–21487. doi:10.1073/pnas.2001254117
Oberbauer, S. F., Tweedie, C. E., Welker, J. M., Fahnestock, J. T., Henry, G. H. R., Webber, P. J., et al. (2007). Tundra CO2 Fluxes in Response to Experimental Warming across Latitudinal and Moisture Gradients. Ecol. Monogr. 77, 221–238. doi:10.1890/06-0649
Odland, A., Høitomt, T., and Olsen, S. L. (2010). Increasing Vascular Plant Richness on 13 High Mountain Summits in Southern Norway since the Early 1970s. Arct. Antarct. Alp. Res. 42 (4), 458–470. doi:10.1657/1938-4246-42.4.458
Pattison, R. R., Jorgenson, J. C., Raynolds, M. K., and Welker, J. M. (2015). Trends in NDVI and Tundra Community Composition in the Arctic of NE Alaska between 1984 and 2009. Ecosystems 18, 707–719. doi:10.1007/s10021-015-9858-9
Pauli, H., Gottfried, M., Dullinger, S., Abdaladze, O., Akhalkatsi, M., Alonso, J. L. B., et al. (2012). Recent Plant Diversity Changes on Europe's Mountain Summits. Science 336, 353–355. doi:10.1126/science.1219033
Peng, A., Klanderud, K., Wang, G., Zhang, L., Xiao, Y., and Yang, Y. (2020). Plant Community Responses to Warming Modified by Soil Moisture in the Tibetan Plateau. Arct. Antarct. Alp. Res. 52 (1), 60–69. doi:10.1080/15230430.2020.1712875
Peng, J., Liu, Z., Liu, Y., Wu, J., and Han, Y. (2012). Trend Analysis of Vegetation Dynamics in Qinghai-Tibet Plateau Using Hurst Exponent. Ecol. Indic. 14, 28–39. doi:10.1016/j.ecolind.2011.08.011
Piao, S., Fang, J., Zhou, L., Ciais, P., and Zhu, B. (2006). Variations in Satellite-Derived Phenology in China's Temperate Vegetation. Glob. Change Biol. 12, 672–685. doi:10.1111/j.1365-2486.2006.01123.x
Pouliot, D. A., Rasim, L., and Ian, O. (2009). Trends in Vegetation NDVI From 1km AVHRR Data Over Canada for the Period 1985–2006. Int. J. Remote Sens. 30 (1), 149–168.
Prevéy, J. S., Rixen, C., Rüger, N., Høye, T. T., Bjorkman, A. D., Myers-Smith, I. H., et al. (2019). Warming Shortens Flowering Seasons of Tundra Plant Communities. Nat. Ecol. Evol. 3, 45–52. doi:10.1038/s41559-018-0745-6
Rodriguez, A. M., Jacobo, E. J., and Golluscio, R. A. (2018). Glyphosate Alters Aboveground Net Primary Production, Soil Organic Carbon, and Nutrients in Pampean Grasslands (Argentina). Rangel. Ecol. Manag. 71, 119–125. doi:10.1016/j.rama.2017.07.009
R Development Core Team (2019). R: A Language and Environment for Statistical Computing. Vienna, Austria: R foundation for statistical computing.
Shen, M., Piao, S., Jeong, S.-J., Zhou, L., Zeng, Z., Ciais, P., et al. (2015). Evaporative Cooling over the Tibetan Plateau Induced by Vegetation Growth. Proc. Natl. Acad. Sci. U.S.A. 112 (30), 9299–9304. doi:10.1073/pnas.1504418112
Steinbauer, M. J., Grytnes, J.-A., Jurasinski, G., Kulonen, A., Lenoir, J., Pauli, H., et al. (2018). Accelerated Increase in Plant Species Richness on Mountain Summits Is Linked to Warming. Nature 556, 231–234. doi:10.1038/s41586-018-0005-6
Stocker, T. F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S. K., Boschung, J., et al. (2014). Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge Univ. Press.
Verbyla, D., and Kurkowski, T. A. (2019). NDVI-climate Relationships in High-Latitude Mountains of Alaska and Yukon Territory. Arct. Antarct. Alp. Res. 51 (1), 397–411. doi:10.1080/15230430.2019.1650542
Walker, M. D., Wahren, C. H., Hollister, R. D., Henry, G. H. R., Ahlquist, L. E., Alatalo, J. M., et al. (2006). Plant Community Responses to Experimental Warming across the Tundra Biome. Proc. Natl. Acad. Sci. U.S.A. 103 (5), 1342–1346. doi:10.1073/pnas.0503198103
Wang, L. Y., and Kang, H. J. (2011). Grassland Resources and the Dominated Plants Atlas of Sanjiangyuan Region. Xining, China: Qinghai People’s Publishing House.
Wang, H., Liu, H. Y., Cao, G. M., Ma, Z. Y., Li, Y. K., Zhang, F. W., et al. (2020). Alpine Grassland Plants Grow Earlier and Faster but Biomass Remain Unchanged Over 35 Years of Climate Change. Ecol. Lett. 23, 701–710.
Wu, M. H., Chen, S. Y., Chen, J. W., Xue, K., Chen, S. L., Wang, X. M., et al. (2021). Reduced Microbial Stability in the Active Layer Is Associated with Carbon Loss under Alpine Permafrost Degradation. Proc. Natl. Acad. Sci. U. S. A. 118 (25), e2025321118. doi:10.1073/pnas.2025321118
Wu, Q. B., and Zhang, T. J. (2008). Recent Permafrost Warming on the Qinghai-Tibetan Plateau. J. Geophys. Res. Atmos. 113, D13108. doi:10.1029/2007JD009539
Wu, Q., Zhang, T., and Liu, Y. (2012). Thermal State of the Active Layer and Permafrost along the Qinghai-Xizang (Tibet) Railway from 2006 to 2010. Cryosphere 6, 607–612. doi:10.5194/tc-6-607-2012
Xu, Y., Knudby, A., Ho, H. C., Shen, Y., and Liu, Y. (2017). Warming over the Tibetan Plateau in the Last 55 Years Based on Area-Weighted Average Temperature. Reg. Environ. Change 17, 2339–2347. doi:10.1007/s10113-017-1163-z
Xu, Z. X., Gong, T. L., and Li, J. Y. (2008). Decadal Trend of Climate in the Tibetan Plateau-Regional Temperature and Precipitation. Hydrol. Process. 22 (16), 3056–3065. doi:10.1002/hyp.6892
Yang, M., Nelson, F. E., Shiklomanov, N. I., Guo, D., and Wan, G. (2010). Permafrost Degradation and its Environmental Effects on the Tibetan Plateau: a Review of Recent Research. Earth-Science Rev. 103, 31–44. doi:10.1016/j.earscirev.2010.07.002
Yang, Y., Hopping, K. A., Wang, G., Chen, J., Peng, A., and Klein, J. A. (2018). Permafrost and Drought Regulate Vulnerability of Tibetan Plateau Grasslands to Warming. Ecosphere 9 (5), e02233. doi:10.1002/ecs2.2233
Yang, Y., Luo, Y., and Finzi, A. C. (2011). Carbon and Nitrogen Dynamics during Forest Stand Development: a Global Synthesis. New Phytol. 190, 977–989. doi:10.1111/j.1469-8137.2011.03645.x
Yang, Y., Wang, G., Klanderud, K., Wang, J., and Liu, G. (2015). Plant Community Responses to Five Years of Simulated Climate Warming in an Alpine Fen of the Qinghai-Tibetan Plateau. Plant Ecol. Div. 2 (8), 211–218. doi:10.1080/17550874.2013.871654
Yi, S. H., Zhou, Z. Y., Ren, S. L., Xu, M., Qin, Y., Chen, S. Y., et al. (2011). Effects of Permafrost Degradation on Alpine Grassland in a Semi-Arid Basin on the Qinghai-Tibetan Plateau. Environ. Res. Lett. 6, 045403.
Zhang, G., Zhang, Y., Dong, J., and Xiao, X. (2013). Green-up Dates in the Tibetan Plateau Have Continuously Advanced from 1982 to 2011. Proc. Natl. Acad. Sci. U.S.A. 110 (11), 4309–4314. doi:10.1073/pnas.1210423110
Zhang, L., Guo, H., Ji, L., Lei, L., Wang, C., Yan, D., et al. (2013). Vegetation Greenness Trend (2000 to 2009) and the Climate Controls in the Qinghai-Tibetan Plateau. J. Appl. Remote Sens. 7, 073572. doi:10.1117/1.jrs.7.073572
Zhang, W., Zhou, T., and Zhang, L. (2017). Wetting and Greening Tibetan Plateau in Early Summer in Recent Decades. J. Geophys. Res. Atmos. 122, 5808–5822. doi:10.1002/2017jd026468
Zhong, L., Ma, Y., Salama, M. S., and Su, Z. (2010). Assessment of Vegetation Dynamics and Their Response to Variations in Precipitation and Temperature in the Tibetan Plateau. Clim. Change 103, 519–535. doi:10.1007/s10584-009-9787-8
Zhou, H., Zhao, X., Tang, Y., Gu, S., and Zhou, L. (2005). Alpine Grassland Degradation and its Control in the Source Region of the Yangtze and Yellow Rivers, China. Grassl. Sci. 51, 191–203. doi:10.1111/j.1744-697x.2005.00028.x
Zhu, Z. C., Piao, S., Myneni, R. B., Huang, M., Zeng, Z., Canadell, J. G., et al. (2016). Greening of the Earth and its Drivers. Nat. Clim. Change 6, 791–795. doi:10.1038/nclimate3004
Keywords: productivity, resampling, climate warming, species richness, the Tibetan Plateau
Citation: Yang Y, Klanderud K, Yang Y, Jin H, Lu Y, Zhang T and Wang G (2022) Homogenization in Species Composition and No Change in Aboveground Biomass Across Tibetan Permafrost Regions Over Ten Years. Front. Environ. Sci. 10:932993. doi: 10.3389/fenvs.2022.932993
Received: 30 April 2022; Accepted: 09 June 2022;
Published: 17 August 2022.
Edited by:
Xiaodong Wu, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Dongliang Luo, Northwest Institute of Eco-Environment and Resources (CAS), ChinaZhenfeng Xu, Sichuan Agricultural University, China
Copyright © 2022 Yang, Klanderud, Yang, Jin, Lu, Zhang and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Genxu Wang, d2FuZ2d4QHNjdS5lZHUuY24=