
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Sci. , 25 October 2022
Sec. Freshwater Science
Volume 10 - 2022 | https://doi.org/10.3389/fenvs.2022.1020243
This article is part of the Research Topic Threatened Aquatic Gems: Freshwater Springs and Groundwater-Dependent Ecosystems View all 9 articles
 Adam H. Love1*
Adam H. Love1* Andy Zdon1
Andy Zdon1 Naomi S. Fraga2
Naomi S. Fraga2 Brian Cohen3Maura Palacios Mejia4
Brian Cohen3Maura Palacios Mejia4 Rachel Maxwell1
Rachel Maxwell1 Sophie S. Parker5
Sophie S. Parker5Desert ecosystems are an environment of climatic extremes, to which many forms of life have adapted. These systems are characterized by scarce water and often sparsely vegetated lands where ecological adaptation to arid conditions has its underpinnings in the universal dependency on water to support life. Understanding the ecohydrological similarity of springs in water-limited areas requires integration of multiple lines of evidence from diverse disciplines. A unique dataset of hydrological and ecological characteristics of Mojave and Sonoran Desert spring ecosystems in California has been developed, incorporating a wide range of spring conditions that enables a broad evaluation of similarity in ecohydrological characteristics across springs in the region. The lack of observed correlation between hydrologic and ecological parameters suggests that the springs in the California Desert each represent a somewhat unique ecosystem that has developed in relative isolation from the other springs. These results imply that because of the uniqueness of these desert ecosystems the idea of mitigation compensation or mitigation offset via replacement or substitution can never truly be achieved if/when these spring areas are impacted. Any ecosystem used as a replacement or substitution could not reasonably be expected to be similar in terms of the associated hydrologic and ecologic conditions to the ecosystem lost. While such offsets can represent a coarser view of replacements or substitution, the more specific conditions that support endemic and water-dependent biodiversity cannot be easily replaced or substituted.
Desert ecosystems may appear harsh and unforgiving, because they are characterized by scarce water, but amid these conditions, fragile ecosystems supporting an abundance of biodiversity have the ability to adapt and thrive (Cantonati et al., 2012; Parker, 2021; Fraga 2022). Springs in arid areas often provide the only perennial source of water for aquatic organisms and wide-ranging species (Davis et al., 2017) and are recognized as biodiviersity hotspots (Bogan et al., 2014). These water-dependent conditions and adaptations have been studied across the globe: Europe (Cantonati et al., 2012), Australia (Harris 1992; Kodric-Brown and Brown 1993; Fensham and Fairfax 2003; and Africa (Suhling et al., 2006). Desert springs are known to have high species endemism, rarity, and presence of relict species (Smith, 2012; Fensham et al., 2011; Rossini et al., 2018). As such, desert springs are geographically-small natural features that provide disproportionate ecological importance in comparison with their size (Hunter et al., 2017) based on their role as evolutionary refugia and ecological refuges (Davis et al., 2013; Murphy et al., 2015).
In North America, desert spring are rare (Shepard 1993) and the Mojave and Sonoran Deserts of California (referred to here as the California Desert) are no different than desert springs elsewhere that act as oases for ecosystem survival. While North American desert springs contain the highest rates of local endemism on the continent (Smith, 2012), with 26 taxa known to be endemic to Ash Meadows Wildlife Refuge in Nevada (Sada 1990), published biological data specific to springs in the California portion of this desert is limited. Parker. (2021) demonstrates that springs in the California desert constitute a diverse range of aquatic habitats and groundwater dependent ecosystems, with varied hydrological and ecological conditions. Several rare and endangered plant taxa occur at California desert springs and seeps, including alkali mariposa lily (Calochortus striatus), Amargosa niterwort (Nitrophila mohavensis), and Tecopa bird’s beak (Chloropyron tecopense). Endemic animals such the Amargosa vole (Microtus californicus subsp. scirpensis) and Shoshone pupfish (Cyprinodon nevadensis subsp. shoshone) are restricted to just a few acres of spring-fed habitat within the California desert.
As such, the conservation importance of desert springs in the California Desert and the associated biota is well recognized (Parker, 2021). The process of conservation is aided by understanding if there are key attributes of such ecosystems that are similar across a region that allow for or facilitate their establishment, adaptation and/or survival.
Understanding the ecohydrological similarity of springs in water-limited areas requires integration of multiple lines of evidence from diverse disciplines. Cartwright, 2020 proposed the following spring evaluation model for assessing stable ecohydrological refugia (Figure 1). This model reflects that stable ecohydrological refugia include such general characteristics as 1) highly permeable geologic formations, 2) relatively stable water discharge, 3) stable water quality and long-groundwater residence times, 4) locations far away from or isolated from population centers, 5) high diversity of obligate aquatic, endemic, or relic taxa, and 6) resilience to drought conditions. While the many springs in the California Desert qualitatively reflect these characteristics for stable ecohydrological refugia, a quantitative evaluation of the interrelations of these characteristics has not been performed.
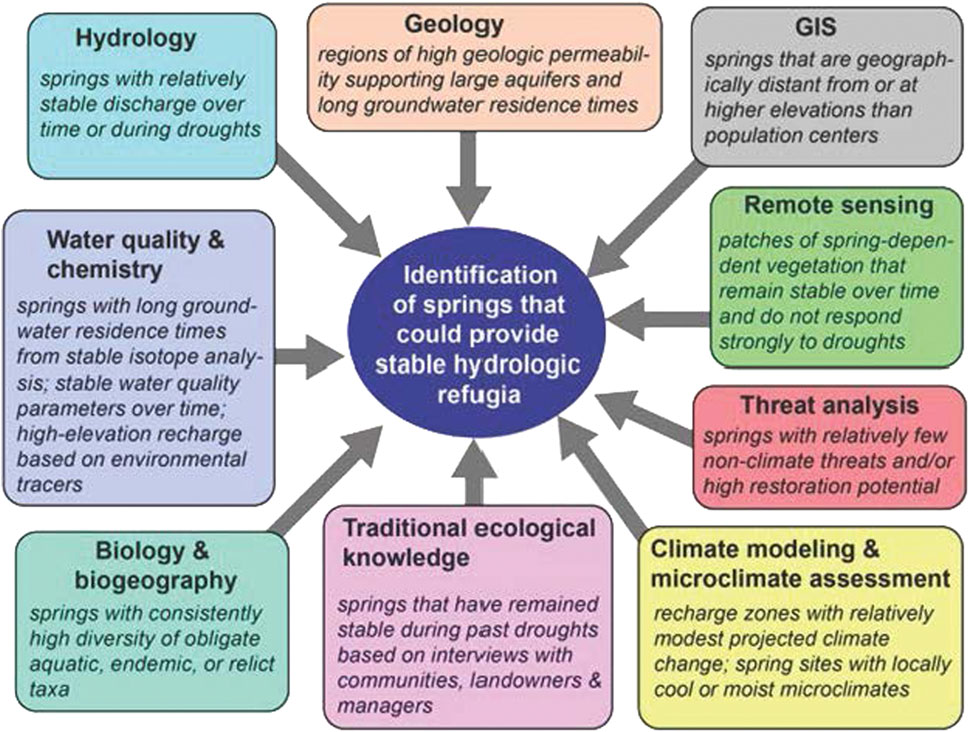
FIGURE 1. Potential lines of evidence for evaluating the ecohydrological similarity of springs in water-limited areas from Cartwright. (2020).
In recent years, a thorough dataset of hydrological and ecological characteristics of California Desert spring ecosystems has been developed that enables the evaluation of similarity in ecohydrological characteristics across springs in the region using criteria consistent with Cartwright, 2020. Information collected reflects 1) water discharge, quality, and residence time, 2) geography, and 3) botanical diversity. This paper reports on the analysis of this dataset, which is the largest dataset known for assessing such characteristics in the California Desert region, and thus the most comprehensive evaluation of the ecohydrological similarity of desert springs in this area.
The springs in the California Desert span a wide variety of source, supply, and quality conditions. Springs range in volume from barely observable to being a major surface water feature, from ambient water to hot springs, from fresh to saline water quality, and natural geologic water seeps to free-flowing water that results from human expansion of a spring resource for water supply purposes. This variety provides a broad spectrum of ecohydrological characteristics that may be used to identify if there are similar traits within and among these spring conditions.
The methodology employed for this evaluation required the collection of ecohydrological data, the use of metadata from botanical field surveys, data dimensional reduction techniques, and statistical evaluation of the ecohydrological data available for comparison. The methodologies utilized for each of these steps is described below.
Four hundred and eighty (480) California Desert springs have been identified and surveyed (Partner Engineering and Science, Inc. 2020). Hydrogeologic field data collected from these springs was based on field data previously collected by Partner Engineering and Science, Inc. (2020), and partially reported in Parker. (2021). Additionally, methodologies for describing spring conditions developed by others (Sada and Pohlman, 2006; Sky Island Alliance, 2012) formed the basis of field descriptions of springs. The hydrogeologic field data collected include spring flow in gallons per minute (gpm), temperature, pH, electrical conductivity, total dissolved solids (TDS), salinity, dissolved oxygen, and isotopic abundances for hydrogen (tritium and deuterium), oxygen, and carbon.
Spring flow was quantified via visual estimation using the “Velocity-area Method” (Gordon, 2004), measuring the time it takes for spring flow to fill a bucket or other container of a known volume, use of a flowmeter, or in the case of Shoshone Spring, measuring outfall characteristics from a pipe of known size (Driscoll 1989). In some cases, such as Borax Spring and Tecopa Hot Spring in the Amargosa Desert region, the spring discharged over a lip or out a pipe which enabled direct measurement of spring flow. At other locations, such as at Crystal Spring, discharge was temporarily captured and channeled into a pipe or a flume to facilitate direct measurement using the bucket filling technique. See Figure 2 for the location of the springs referenced above.
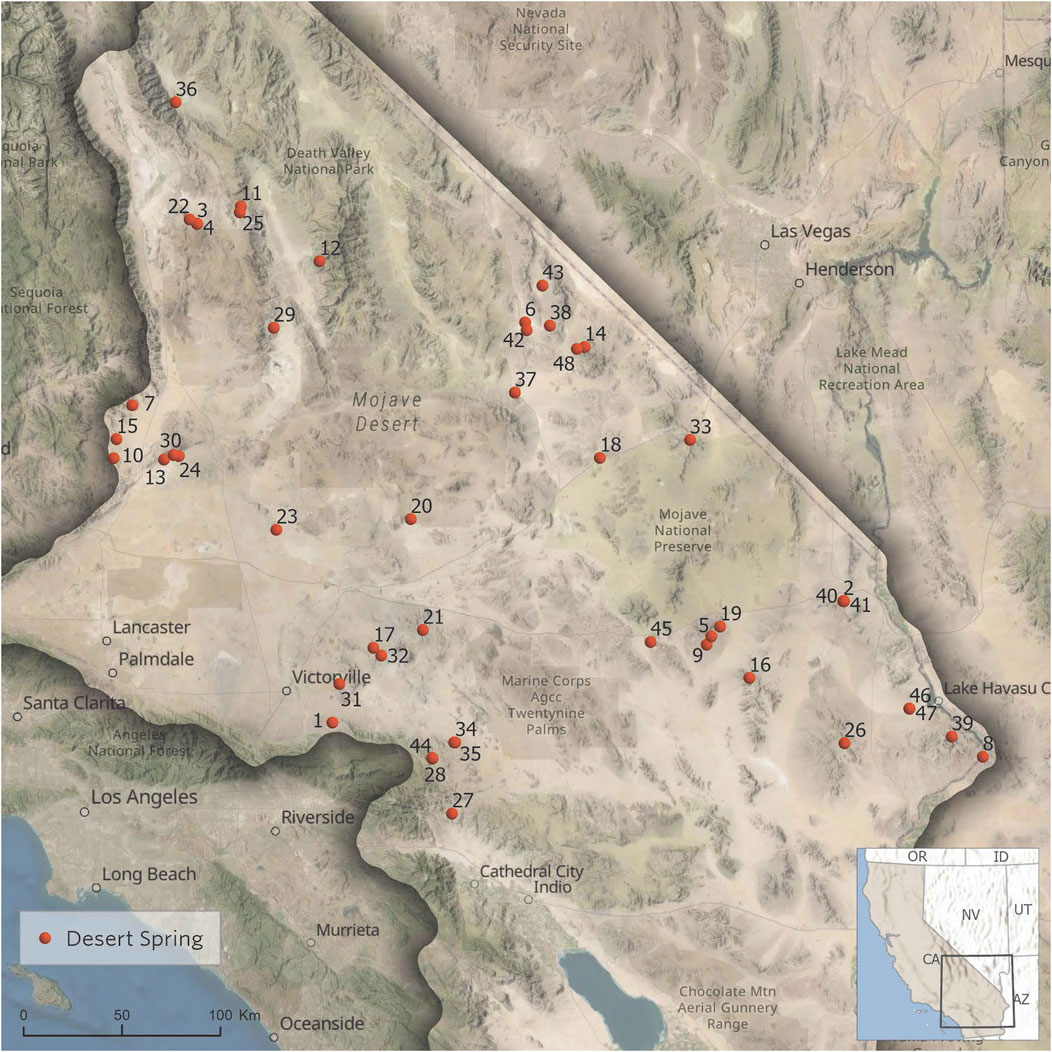
FIGURE 2. Map of California Desert spring locations used in this ecohydrogeological evaluation. Map identification numbers correspond to the spring names in Table 1.
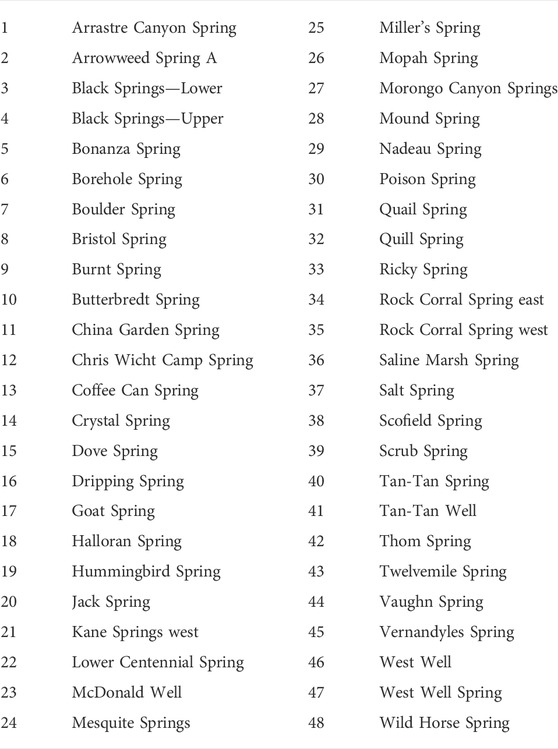
TABLE 1. Names of springs shown in Figure 2.
Note, there are compromises in the implementation of spring flow measurements that can result in under-estimation or over-estimation of free-flowing discharge. Ideally, all the flow from a spring would be fully captured and channeled into a pipe or flume, allowing for much greater accuracy in measurement of flow. Temporarily channeling the spring using a pipe and other non-permanent materials such as mud and rocks can capture most of the flow, but not all, which can lead to inaccuracies in measurement. Measurement of flow using a solid-state flow meter requires estimates of cross-sectional area and the use of one to two flow measurement points as the meter is often large relative to the width of the channel. In addition, flow measurements are only reported for the days that the measurements were actually collected, and potential variability in both seasonal spring flow and longer-term annual trends are reasonably expected. Ultimately, in most cases the spring flow measurements within this paper should be seen as a semi-quantitative estimate for the spring discharge.
Field water quality measurements were made with a Hanna HI991300 Multiparameter Meter (for temperature, pH, conductivity, and total dissolved solids) and a YSI 550A Dissolved Oxygen Meter. Equipment was checked for calibration daily and calibrated (multipoint) as required per equipment guidelines.
Spring water samples were collected from a select group of total springs and wells, for measurement of:
• Stable hydrogen and oxygen isotopes where surface water was present; and,
• Radiocarbon and tritium to evaluate hydrologic residence time (age) of spring waters.
Stable isotope, tritium and radiocarbon analyses were conducted by Isotech Laboratories, Inc. (Isotech) in Champaign, Illinois. Samples for oxygen (δ18O) and deuterium (δD) were collected in 60 ml glass bottles equipped with a conical shaped insert inside the cap that forms an airtight seal when the bottle is closed. Samples were shipped to Isotech where the 18O/16O and D/H ratios were measured as a gas using standardized mass spectrometry methods. Results are reported as a normalization to Standard Mean Ocean Water (SMOW), which is an internationally recognized standard in stable isotope analysis. The normalization converted to standard δ (“del”) notation following the convention:
Where R is the isotope ratio of the sample and Rstd is the ratio of the standard.
For radiocarbon analyses, water samples were collected in 1-L high density polyethylene (HDPE) sample bottles provided by the laboratory. Samples were shipped in a chilled cooler to Isotech where the radiocarbon analyses involved acidification of water to convert dissolved inorganic carbon (DIC) to carbon dioxide (CO2) which was then extracted, purified, and submitted for final analysis by mass spectrometry. Tritium analysis was conducted using the tritium enhanced enrichment (TEE) method to obtain lower reporting limits.
The δ13C results are reported as a normalization to the Vienna Pee Dee Belemnite (VPDB), an internationally recognized standard in δ13C analysis. The 14C content of DIC is reported as a percentage of modern carbon (pmc). 14C decays at a steady rate with a half-life of 5,730 years. Therefore, waters with 50% modern carbon would have an apparent age of 5,730 years, waters with 25% modern carbon would have an apparent age of 11,460 years and so on.
While the limited number of sampled spring locations restricted the age dating of the source water data being used in most of the statistical analyses, the observations of age of the source water characteristics nonetheless provided some additional information regarding similarity of this limited dataset.
Botanical field surveys were conducted at 48 spring locations in the fall of 2018 and summer of 2019, when most wetland plants are in the best condition for plant identification. While 480 desert springs have been surveyed for hydrology in the California Desert, botanical field surveys were limited to 48 based on the availability of funding to support such work. Prior to botanical field surveys, herbarium specimens, literature, and publicly available databases were examined to identify historical records and plant species of conservation concern previously documented across the survey area. The Consortium of California Herbaria (2019) and SEINet (2019) were queried to produce a list of 523 unique historical plant records to establish baseline data of the local vegetation. The springs were selected because they span a broad geography and capture documented hydrological variation. Plant species were identified to the minimum rank possible (species, subspecies, or variety), and observations were excluded if they could only be identified to genus or family. Five hundred and twenty-four (524) herbarium specimens were collected in the field to verify plant identification. The resulting specimens were deposited in the California Botanic Garden herbarium. Plant specimens were not collected if plants did not have flowers or fruits present and could not be confidently identified. Plants were identified using multiple sources including Baldwin. (2012), Jepson Flora project (2022), and Flora of North America Editorial Committee. (1993). Details of field sampling collection, handing, and analysis are described in Fraga (2022).
We located each spring using field-documented coordinates and delineated the areal extent of each spring using “heads-up” digitizing into a polygon feature class within Esri’s ArcGIS Desktop 10.8 using the Albers equal-area conic projection. Digitizing polygons is based on the associated vegetation and footprint visible in NAIP 2018 1-m aerial imagery. The ArcGIS CALCULATE AREA tool enables calculation of the hectares for each polygon, and the ArcGIS GENERATE NEAR TABLE tool enables calculation of the distances between each of the spring polygons, in meters.
We used Spatial Join to associate the centroid of each spring footprint with the Conservation Value from the Mojave Desert Ecoregional Assessment (Randall, 2010), the underlying groundwater basin (Department of Water Resources 2022), and the Desert Renewable Energy Conservation Program designation (Bureau of Land Management 2016).
We calculated a standardized Topographic Position Index (sTPI) on a 10-m elevation dataset for the study area using the methods described by Weiss (2001). This process uses a fine scale approach with an inner radius of 3 m and an outer radius of 12 m. We ran Zonal Statistics As Table on the sTPI for each spring footprint to determine mean sTPI for each spring.
Lastly, we calculated the average summer Normalized Difference Vegetation Index (NDVI) for the years 1985 through 2020 for each spring footprint using the GDE Pulse API (The Nature Conservancy 2021). NDVI is used to quantify vegetation greenness and is useful in understanding vegetation density.
We compiled the hydrologic, ecological, and GIS data in a database (provided in Supplemental Material) that consists of 1100 + entries with 480 unique spring names. Within the database, 48 springs have both hydrologic and botanical data entries. Some springs have hydrologic data from multiple collection dates, and some have data only from a single collection date. For springs with hydrologic data from multiple collection dates, we reduced the data from each spring to a single average hydrologic condition that was the mean of the observations. Botanical observations were simplified into categories of information: mean summer NDVI (Summer NDVIAve), hectares, total number of wetland plant taxa and total number of invasive species.
While ideally ecological descriptions and parameters would be available for the 480 unique springs, this evaluation is limited to the statistical power resulting from the available data from the 48 springs with both hydrologic and botanical data entries. Funding is not currently available for the intensive nature that such ecological field assessment and data collections require to expand the botanical survey database and provide greater statistical power. Therefore, this database reflects the most comprehensive dataset for the California Desert to perform such an evaluation.
We evaluated the quantitative parameters in the ecohydrologic database using the Microsoft Excel XLSTAT statistical package to perform both pairwise correlations and multivariate principal component analyses (PCA). We evaluated direct connections between database elements by performing pairwise linear correlations using Pearson correlation. We then used the PCA results to conduct a data exploration evaluation where we examined the distribution of springs in principal component space for visual similarly. PCA is often used to identify the combination of features for best describing the variance in a data set. For situations where one or a few springs’ characteristics were outliers from the rest of the spring conditions and dominated the factor weighting of the PCA, we eliminated the outlier spring(s) from the dataset and then re-ran and re-evaluated the new subset of springs using PCA.
While other multivariate methods were explored for the identifying similarity in the specific species identified in botanical survey, many of the individual springs have a small number of species that occur in them and those species are not often shared with the other springs. Thus, the ability to use specific species for pattern matching and/or discrimination (e.g. only 6 springs had more than 50 taxa and most had 3–10) across the California Desert or its subregions was limited. The springs surveyed had relatively low alpha diversity at individual sites, but high gamma diversity across all sites and pairwise differences between sites indicated high beta diversity.
The ecoregion of the California Desert spans an area of 103,500 km2 (Omernik 1987). The 48 locations in the database with ecohydrological data occupy an tiny area (approximately 35 km2) of the total landmass of the California Desert. The springs have an average distance of 166 km between springs and only seven pairwise distances between the 48 springs that are below 0.5 km. Thus, the geographically isolated nature of most desert springs in this region is a notable characteristic. Figure 2 shows the locations across the California Desert of each spring used in this ecohydrogeological evaluation.
Pairwise correlations of the geographic, hydrologic and botanical ecohydrologic parameters illustrate expected correlations among geographic/hydrologic variables (longitude and latitude versus δD and δ18O; elevation and temperature) and ecological variables (Summer NVDIAve versus hectares, total wetland plants taxa, and # non-native plants; total wetland plants taxa versus # non-native plants). While correlations between parameters such as spring flow versus vegetation density, wetland size, or plant diversity might have been expected, no meaningful pairwise correlations were observed between hydrogeologic and ecological parameters. Only the modest positive correlation between water conductivity (salinity) versus hectares and the modest negative correlation between elevation versus hectares were different from zero with a significance level alpha = 0.05. Table 2 shows the linear correlation among ecohydrological parameters.
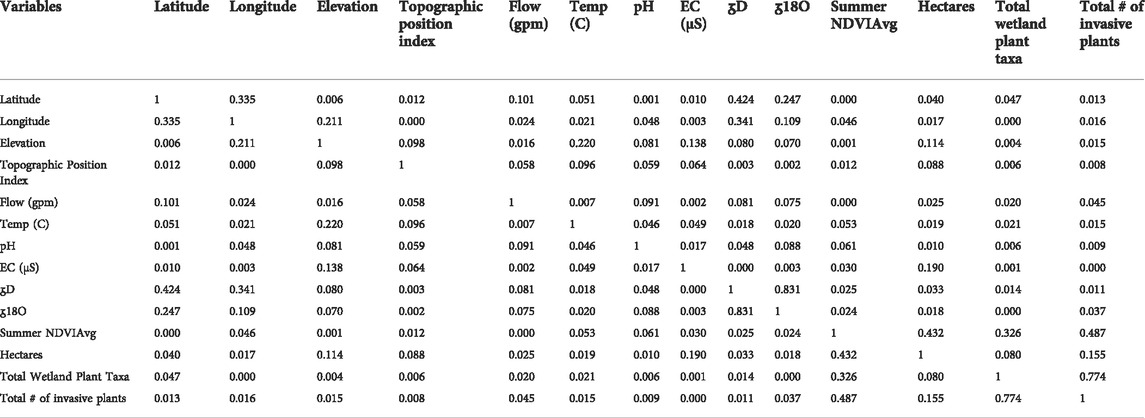
TABLE 2. Linear correlation among ecohydrological parameters for 48 California Desert springs. Values shown are Pearson coefficients of determination (r2).
Multivariate PCA did not reveal any more complex multiparameter correlation among the hydrologic and ecological dataset of these desert springs. As with the linear correlations, correlations of multivariate PCA factors reflected some interdependence each among ecological parameters and hydrologic parameters, but no significant cross dependence relationship was identified. Seventy percent of the cumulative variability among parameters can be accounted for through the first three PCA factors, with ecological parameters having the strongest correlation in the primary set of PCE features (F1) and hydrologic parameters having the strongest correlations in secondary and tertiary set of PCE features (F2 and F3). Table 3 shows the multivariate contribution of the ecohydrological parameters (%) to the associated PCA factors.
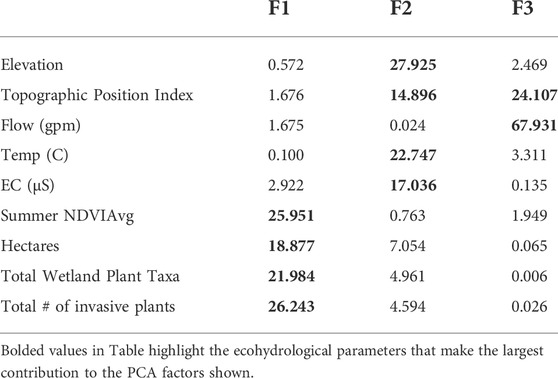
TABLE 3. Multivariate contribution of the ecohydrological parameters (%) for 48 California Desert springs to the associated PCA factors.
Given the vast area of the California Desert, PCA was used to evaluate if the overall set of ecohydrologic parameters reflected any distinctive geographic clustering. Figure 3 shows a PCA biplot of F1 and F2 for ecohydrological dataset of California Desert springs. The distribution of springs within the F1 vs. F2 biplot (accounting for almost 55 percent of the cumulative parameter variability) did not reveal any distinct geographic clustering among the desert springs where a subsection within the California Desert reflected distinct ecohydrological characteristics.
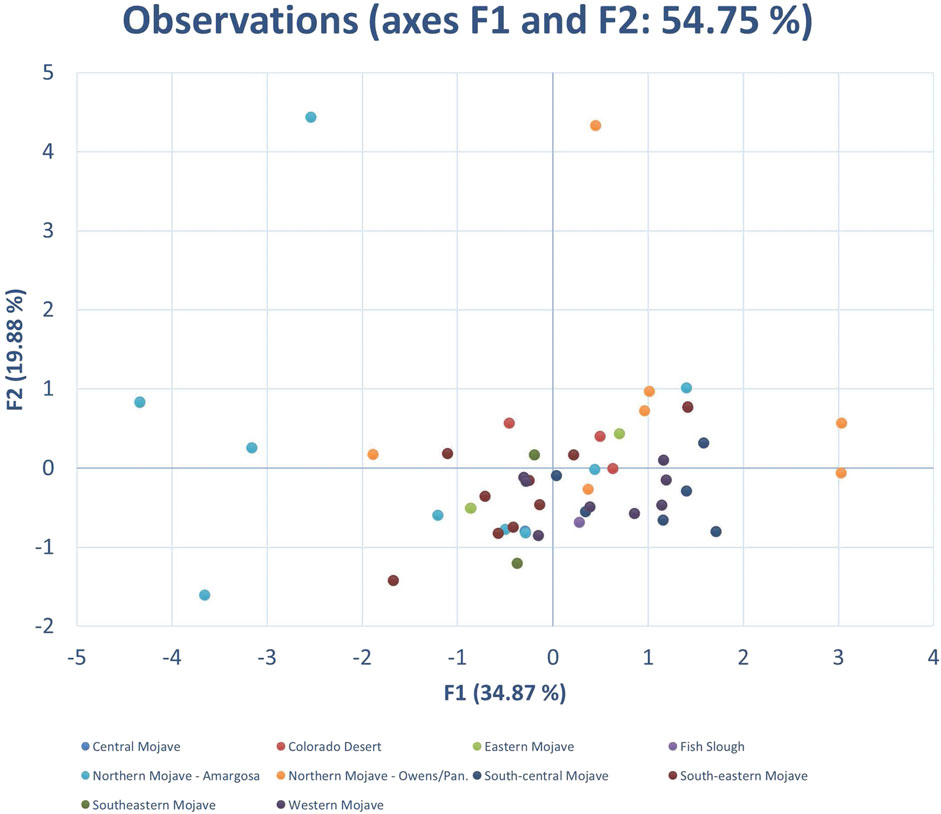
FIGURE 3. PCA biplot of F1 and F2 for ecohydrological dataset of California Desert springs with designated ecoregion color coded as shown.
To try to prevent extreme geographic diversity from obscuring meaningful relationships between hydrologic and ecological parameters, we also evaluated the overall ecohydrological database based on quadrants reflecting differences in the stable isotope abundances in California Desert spring water (Zdon, 2018) as shown in Figure 4. The difference in stable isotope abundance, divided into northwest, northeast, southwest, and southeast quadrants that separate groups of springs as they might reflect difference in the influence of summer monsoonal versus winter maritime precipitation sources. The quadrants presented are based on field measured stable isotope values from California Desert springs and from precipitation patterns (Zdon and Love 2020).
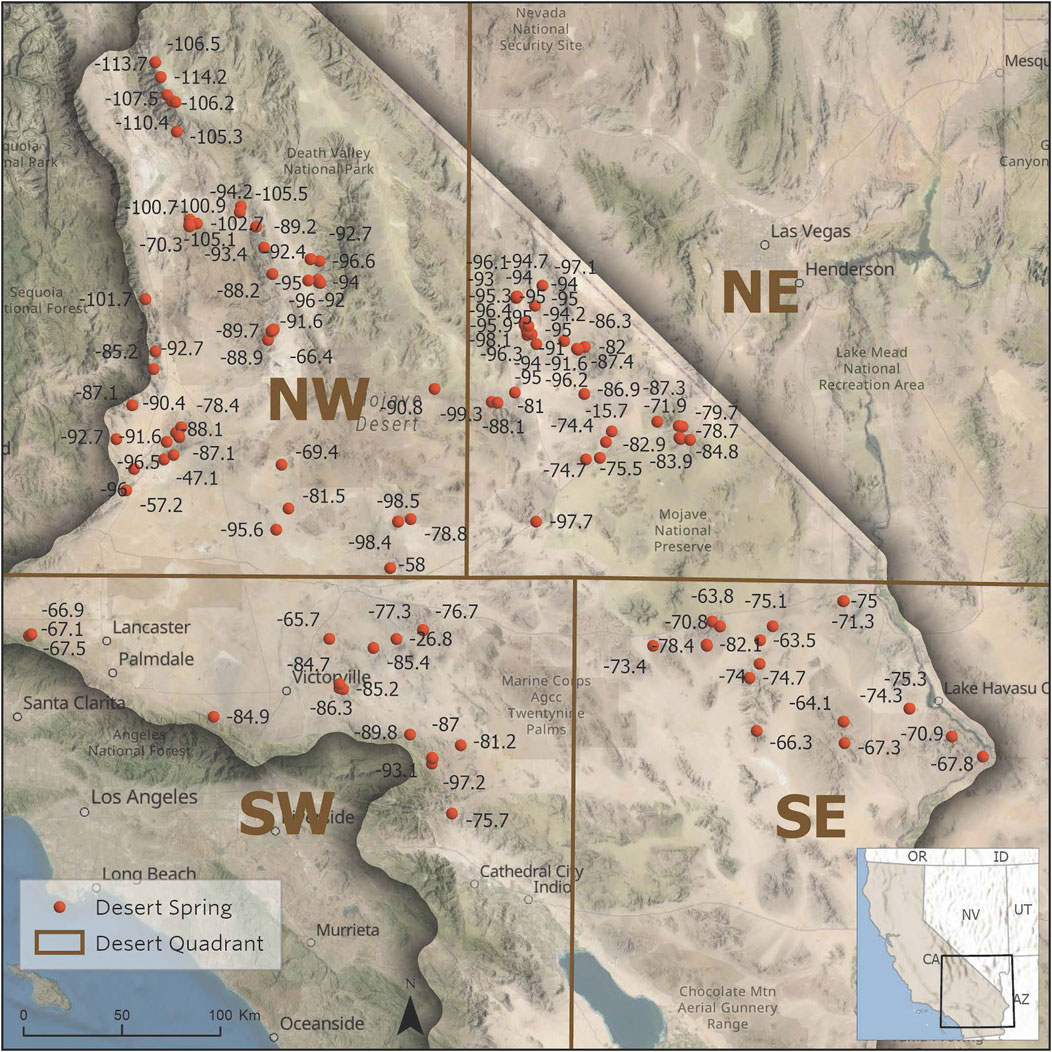
FIGURE 4. Quadrants of the California Desert region based on stable isotope abundances in desert springs (from Zdon, 2018).
As with the overall database, pairwise correlations of the ecohydrologic parameters from each quadrant show no meaningful pairwise correlations between hydrogeologic and ecological parameters. Table 4 shows the linear correlation among ecohydrological parameters for the quadrants of the California Desert region.
Northern Desert Quadrant: None of the pairwise correlations between hydrogeologic and ecological parameters are different from 0 with a significance level alpha = 0.05.
Western Desert Quadrant: None of the pairwise correlations between hydrogeologic and ecological parameters are different from 0 with a significance level alpha = 0.05.
South Desert Central Quadrant: None of the pairwise correlations between hydrogeologic and ecological parameters are different from 0 with a significance level alpha = 0.05.
Southeast Desert Quadrant: Only the pairwise positive correlation between electrical conductivity (salinity) and hectares is different from 0 with a significance level alpha = 0.05. No specific ecohydrological mechanistic which explains this correlation in the southeast regions has been identified.
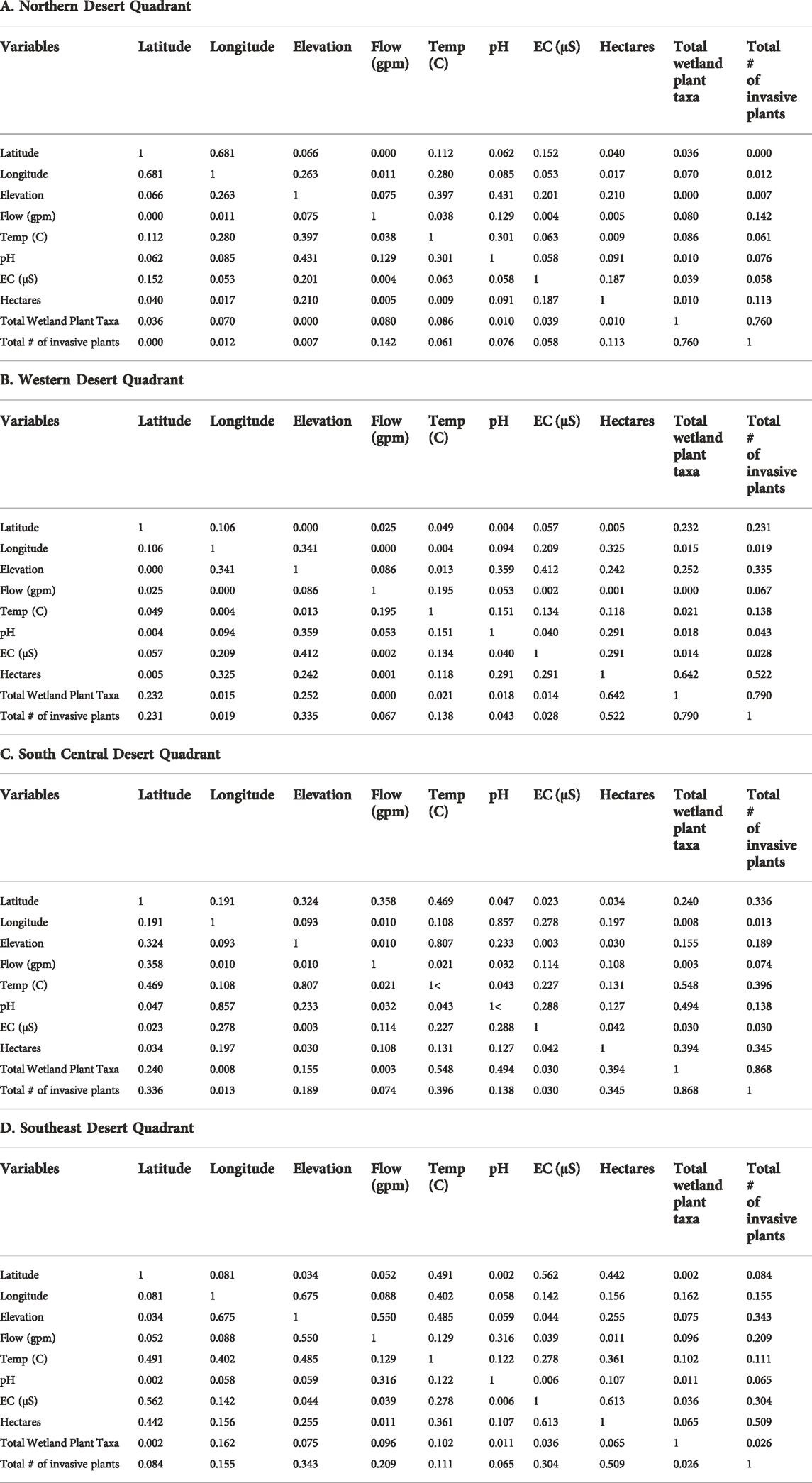
TABLE 4. Linear correlation among ecohydrological parameters for the quadrants of the California Desert region based on stable isotope abundances in desert springs (from Zdon, 2018). Values shown are Pearson coefficients of determination (r2).
Similarly, multivariate PCA did not reveal any more complex multiparameter correlation among hydrologic and ecologic parameters for desert springs in each quadrant. As with the evaluation of the ecohydrological dataset as a whole, the first few PCA factors for each quadrant have the strongest correlations to either primarily ecological parameters or primarily hydrologic parameters. There appears to be little cross dependence in the spring characteristics among the ecological and hydrologic parameters.
Because Morongo Canyon Spring and Bonanza Spring were each anomalous in their quadrant and therefore skewed the distribution of factors in the PCA evaluations, biplots are shown with and without the inclusion of the data for each of those springs. Figures 5–8 show the PCA biplot of F1 and F2 for ecohydrological dataset of each quandrant. While most of the springs in each quadrant appear fully distinct in ecohydrologic characteristics from each another, there appears to be only a small cluster of springs in the Southeast quadrant with somewhat similar ecohydrologic attributes (when Bonanza Spring is removed – Figure 8B): Tan-Tan spring, Tan-Tan Well, and Arrowweed Spring A. These springs are each within 350 m of each other and may reflect both similarly in hydrologic conditions and overlap in ecological footprint.

FIGURE 5. PCA biplot of F1 and F2 for ecohydrological dataset for the Northern Desert region.
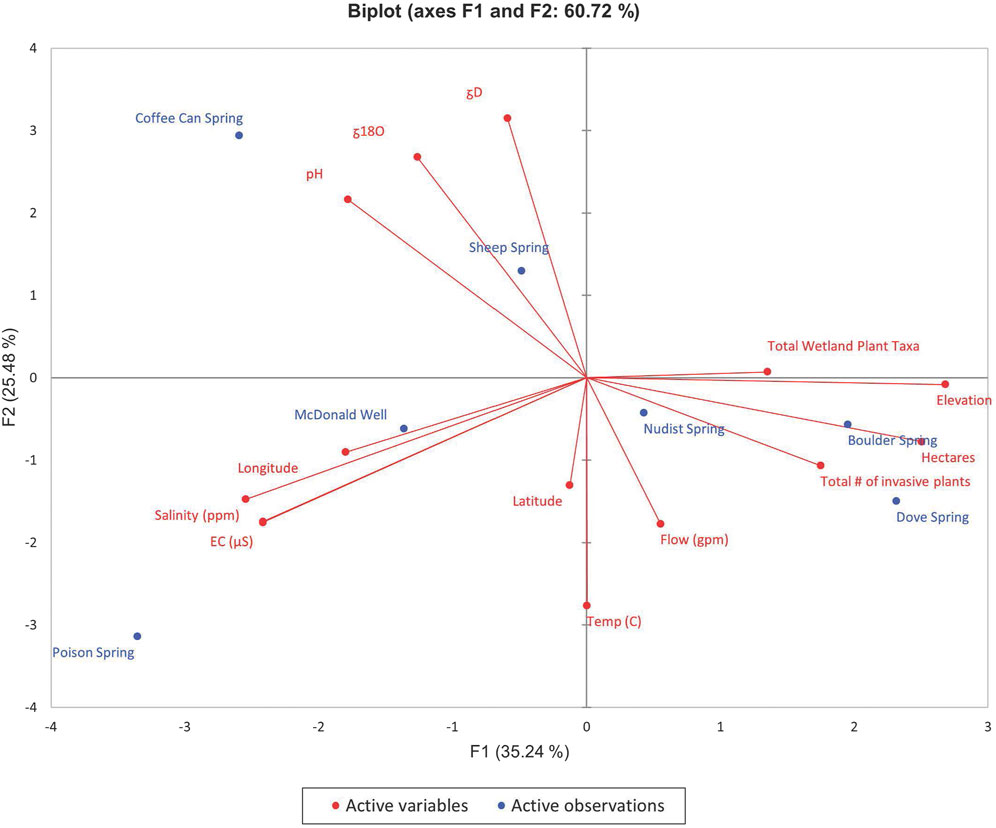
FIGURE 6. PCA biplot of F1 and F2 for ecohydrological dataset for the Western Desert region.
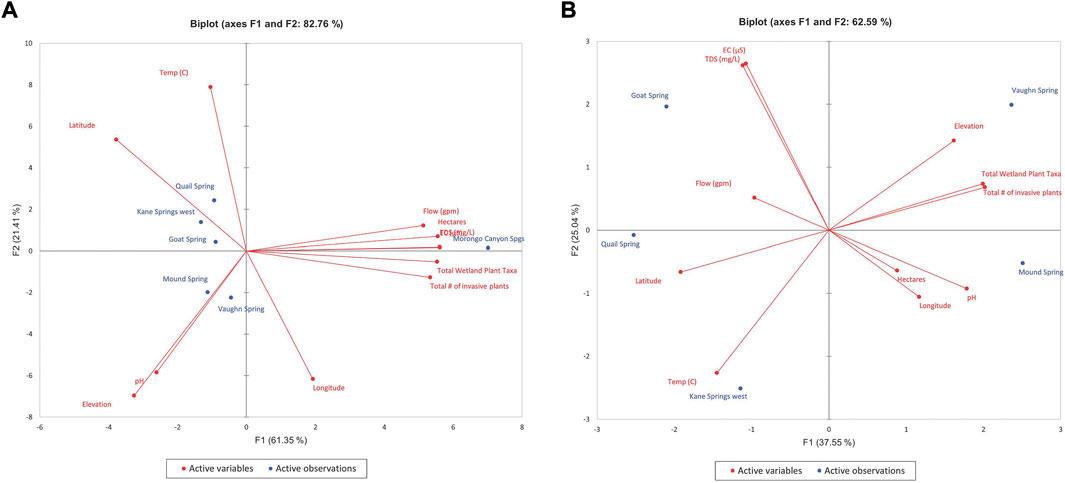
FIGURE 7. PCA biplot of F1 and F2 for ecohydrological dataset for the South Central Desert region (A) Moronga Canyon Spring included, (B) Moronga Canyon Spring excluded.
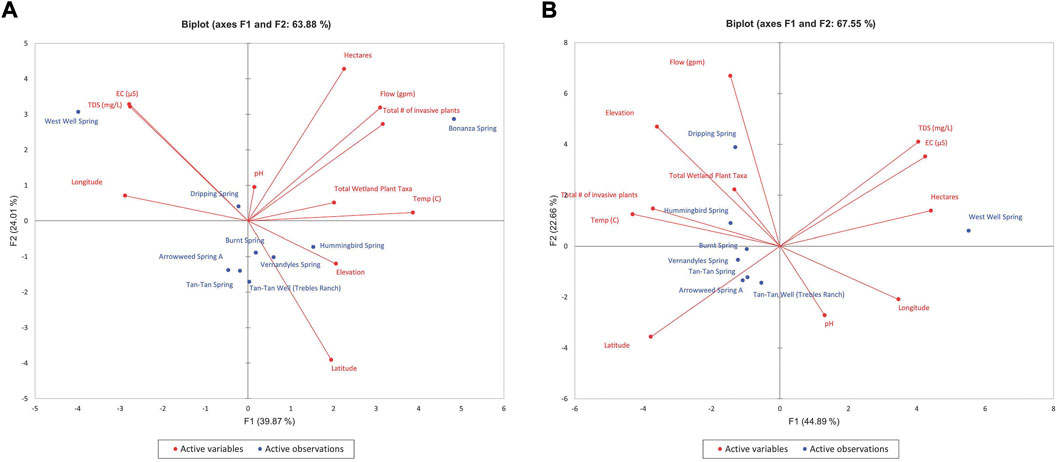
FIGURE 8. PCA biplot of F1 and F2 for ecohydrological dataset for the Southeastern Desert region (A) Bonanza Spring included, (B) Bonanza Spring excluded.
In general, both pairwise and multivariate statistical evaluation of the ecohydrological characteristics of the springs in each quadrant generally reflect a high degree of uniqueness across each area, with no clear hydrogeologic or geographic factors that control the ecological diversity observed in each habitat.
The statistical ecohydrologic uniqueness of each spring based on this analysis is consistent with observations of eDNA from Palacios Mejia et al. (2020), which found that each of four California Desert springs also supports a unique biological community with low overlap in animal species. While the eDNA study had some limitation in eDNA primers that resulted in significant species data gaps, the observed biodiversity via both visual observations and eDNA reflect a comparable uniqueness among the small dataset.
Similarly, Fraga (2022) concluded that while desert springs collectively support a large proportion of plant diversity, or nearly 22% of the total vascular plant diversity known within the California Desert in a tiny percentage of the total land area, this 22% represented a low degree of species richness - as nearly 50% of the springs had fewer than 20 total plant taxa present. Among the plant species observed at the springs, the botanical composition was highly dissimilar. Nearly 40% (185 of 479) of the documented plant taxa were recorded at only one spring site. While wetland and non-native plant taxa were the most frequently encountered, there was a relatively high proportion of ephemeral upland taxa reflecting local conditions.
As the results show above, including of the various hydrogeological characteristics of the springs into the ecohydrological database ultimately did not provide a means to explain the overall botanical diversity differences observed among springs. While the wide range in hydrogeological source, quantity, and quality allowed for an addition dimension of data evaluation, none of these was shown to be an overarching factor influencing overall ecological diversity.
Instead, there appears to be a sufficient variety of observed plant taxa and other biodiversity elements to allow for the development of unique ecosystems that are each reflective of the wide array of influencing factors – including water source, quantity, and quality; geographic position and meteorology; regional seed dispersal; local upland conditions; human disturbance – without any factor in this region dominating the development of each habitat. Certainly, the relative geographic isolation of desert springs facilitates the development of these unique habitats without the added influence of other nearby habitats readily expanding its ecological footprint to an adjacent spring location. The resulting uniqueness of each desert spring’s ecohydrological condition suggests that each habitat defies generalizations and instead is its own “snowflake” that requires specific study to truly understand the details of the habitat elements that have been established. Without an ability to make generalization about desert springs, these springs require the continuation and expansion of hydrogeological and botanical spring surveys to identify the unique ecohydrological characteristics of each of these important habitats and to effectively document baseline spring conditions that enable identification of when these fragile systems are adversely impacted. Expansion of botanical field surveys to cover a greater overall percent of the identified California Desert spring wetlands is critical for developing more meaningful conservation assessments and providing the information necessary to guide management of these natural biodiversity resources.
These results enforce the premise that spring ecosystems are not fungible, and therefore plans for management and restoration should be uniquely tailored to each spring location
The California Environmental Quality Act (CEQA) and the National Environmental Policy Act (NEPA) both require that the potential impacts on species, habitat and farmland from development be considered. Measures are taken to balance those negative impacts through a process known as “mitigation.” Mitigation is frequently required when significant impacts are identified by the environmental review process. Mitigation is defined (California Council of Land Trusts 2022) in Section 15370 of the California Code of Regulations (CEQA Guidelines) as:
1. Avoiding the impact altogether by not taking a certain action or parts of an action.
2. Minimizing impacts by limiting the degree or magnitude of the action and its implementation.
3. Rectifying the impact by repairing, rehabilitating, or restoring the impacted environment.
4. Reducing or eliminating the impact over time by preservation and maintenance operations during the life of the action.
5. Compensating for the impact by replacing or providing substitute resources or environment”
a. Biodiversity offsets are the last step in the mitigation hierarchy. They constitute measurable conservation gains, deliberately achieved to balance any significant biodiversity losses that cannot be countered by avoiding or minimizing impacts from the start, or addressing the damage done through restoration (Forest Trends, 2022).
These results imply that because of the uniqueness of these desert ecosystems the idea of mitigation compensation or mitigation offset via replacement or substitution can never truly be achieved if/when these spring areas are impacted. Any desert spring ecosystem used as a replacement or substitution could not reasonably be expected to be similar in terms of the hydrologic and associated ecologic conditions to the ecosystem lost. While such offsets can represent a coarser view of replacements or substitution, the more specific conditions of the biodiversity cannot easily be replaced or substituted.
The original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding author.
Conception or design of the work (AL, AZ, and SP). Acquisition, analysis, or interpretation of the data (AL, AZ, NF, BC, and MP). Drafting the work or revising it critically (AL, AZ, NF, BC, MP, RM, and SP). As such, all authors agree to be accountable for the content of the work.
The authors would like to acknowledge the funding provided that supported the data collection for this study from The Nature Conservancy, U.S. Bureau of Land Management, California Department of Water Resources, County of Inyo, Mojave Desert Land Trust, Amargosa Conservancy, Transition Habitat Conservancy, and California Botanic Garden. Both Roux and UCLA provided in-kind support.
AL, AZ, and RM were employed by the company Roux.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2022.1020243/full#supplementary-material
Baldwin, Bruce G. (2012). The Jepson manual: Vascular plants of California. Editors D Goldman, D J. Keil, R Patterson, J. Thomas, and Rosatti (University of California Press).
Bogan, M. T., Noriega-Felix, N., Vidal-Aguilar, S. L., Findley, L. T., Lytle, D. A., Gutiérrez-Ruacho, O. G., et al. (2014). Biogeography and conservation of aquatic fauna in spring-fed tropical canyons of the southern Sonoran Desert, Mexico. Biodivers. Conserv. 23, 2705–2748. doi:10.1007/s10531-014-0745-z
Bureau of Land Management (2016). Executive summary for the record of decision.” in Desert Renewable Energy conservation plan. Available at: https://eplanning.blm.gov/public_projects/lup/66459/133459/163123/DRECP_BLM_ROD_Executive_Summary.pdf.
California Council of Land Trusts (2022). Mitigation. Available at: https://www.calandtrusts.org/conservation-basics/conservation-tools/mitigation/ (Accessed July 15, 2022)
Cantonati, M., Füreder, L., Gerecke, R., Jüttner, I., and Cox, E. J. (2012). Crenic habitats, hotspots for freshwater biodiversity conservation: Toward an understanding of their ecology. Freshw. Sci. 31 (2), 463–480. doi:10.1899/11-111.1
Cartwright, J. M. (2020). Oases of the future? Springs as potential hydrologic refugia in drying climates. Front. Ecol. Environ. 18 (5), 245. doi:10.1002/fee.2191
Consortium of California Herbaria portal (Cch2), (2019). CCH2. Available at: https://www.cch2.org/portal/index.php.
Davis, J. A., Kerezsy, A., and Nicol, S. (2017). Springs: Conserving perennial water is critical in arid landscapes. Biol. Conserv. 211, 30–35. doi:10.1016/j.biocon.2016.12.036
Davis, J., Pavlova, A., Thompson, R., and Sunnucks, P. (2013). Evolutionary refugia and ecological refuges: Key concepts for conserving Australian arid zone freshwater biodiversity under climate change. Glob. Chang. Biol. 19 (7), 1970–1984.
Department of Water Resources, (2022). California groundwater basin boundary assessment tool. Available at: https://gis.water.ca.gov/app/bbat/ (Accessed July 15, 2022).
Driscoll, Fletcher G. (1989). Groundwater and wells. Second Edition, 1. St. Paul: Johnson Filtration Systems, Inc., 089p.
Fensham, R. J., Silcock, J. L., Kerezsy, A., and Ponder, W. (2011). Four desert waters: Setting arid zone wetland conservation priorities through understanding patterns of endemism. Biol. Conserv. 144, 2459–2467. doi:10.1016/j.biocon.2011.06.024
Fensham, R. J., and Fairfax, R. J. (2003). Spring wetlands of the great artesian basin, Queensland, Australia. Wetl. Ecol. Manag. 11 (5), 343–362. doi:10.1023/b:wetl.0000005532.95598.e4
Flora of North America Editorial Committee (1993). Flora of North America North of Mexico. New York and Oxford. 22+ vols Available at: http://beta.floranorthamerica.org.
Forest Trends (2022). The mitigation hierarchy. Available at: https://www.forest-trends.org/bbop/bbop-key-concepts/mitigation-hierarchy/.
Fraga, N. S. (2022). October.Floristic patterns and conservation values of Mojave and Sonoran desert springs in CaliforniaNat. Areas J.
Gordon, N. D. (2004). Stream hydrology: An introduction for ecologists. Wiley. Available at: https://www.wiley.com/en-us/Stream+Hydrology%3A+An+Introduction+for+Ecologists%2C+2nd+Edition-p-9780470843574
Harris, C. R. (1992). Mound springs: South Australian conservation initiatives. Rangel. J. 14 (2), 157
Hunter, M. L., Acuña, V., Bauer, D. M., Bell, K. P., Calhoun, A. J., Felipe-Lucia, M. R., et al. (2017). Conserving small natural features with large ecological roles: A synthetic overview. Biol. Conserv. 211, 88–95. doi:10.1016/j.biocon.2016.12.020
Jepson Flora Project (2022). Jepson eFlora. Available at: https://ucjeps.berkeley.edu/eflora/.
Kodric-Brown, A., and Brown, J. H. (1993). Highly structured fish communities in Australian desert springs. Ecology 74 (6), 1847–1855. doi:10.2307/1939942
Murphy, N. P., Guzik, M. T., Cooper, S. J., and Austin, A. D. (2015). desert Spring refugia: Museums of diversity or evolutionary cradles? Zool. Screen. 44 (6), 693–701. doi:10.1111/zsc.12129
Omernik, J. M. (1987). Ecoregions of the conterminous United States. Annals of the Association of American Geographers 77 (1), 118–125. doi:10.1111/j.1467-8306.1987.tb00149.x
Palacios Mejia, M., Curd, E., Edalati, K., Renshaw, M. A., Dunn, R., Potter, D., et al. (2021). The utility of environmental DNA from sediment and water samples for recovery of observed plant and animal species from four Mojave Desert springs. Environ. DNA 3, 214–230. doi:10.1002/edn3.161
Parker, S. S., Zdon, A., Christian, W. T., Cohen, B. S., Palacios Mejia, M., Fraga, N. S., et al. (2021). Conservation of Mojave Desert springs and associated biota: Status, threats, and policy opportunities. Biodivers. Conserv. 30, 311–327. doi:10.1007/s10531-020-02090-7
Partner Engineering and Science, Inc (2020). California desert springs and waterholes: Spring survey update, mono, Inyo, san bernardino, kern, los angeles. California: Riverside and Imperial Counties.
Randall, J. M. (2010). Mojave Desert ecoregional assessment. Version 1.1 Available at: https://www.scienceforconservation.org/assets/downloads/Mojave_Desert_Ecoregional_Assessment_2010.pdf (Accessed July 15, 2022).
Rossini, R. A., Fensham, R. J., Stewart‐Koster, B., Gotch, T., and Kennard, M. J. (2018). Biogeographical patterns of endemic diversity and its conservation in Australia's artesian desert springs. Divers. Distrib. 24 (9), 1199. doi:10.1111/ddi.12757
Sada, D. W. (1990). Recovery plan for the endangered and threantened species of Ash Meadows, Nevada. Available at: https://www.fws.gov/carlsbad/SpeciesStatusList/RP/19900928_RP_Ash Meadows Plants and Fish.pdf (Accessed Nov 18, 2019).
Sada, D. W., and Pohlman, K. F. (2006). U.S. National park service Mojave inventory and monitoring network spring survey protocols: Level I, 19 november 2003. Reno: Desert Research Institute, Inc. Available at: https://www.nps.gov/moja/learn/nature/upload/LevelI&IISurveyProtocols.pdf.
Seinet, (2019). SEINet data portal. Available at: https://swbiodiversity.org/seinet/ (Accessed July 15, 2022).
Sky Island Alliance (2012). Springs inventory and assessment training manual. Spring ecosystem inventory and assessment protocols version 2.0. Available at: https://skyislandalliance.org/wp-content/uploads/2020/07/Misztal_DLCC_FInalReport_AppendixA.pdf (Accessed July 15, 2022).
K. Smith (2012). Aridland Springs in North America: Ecology and Conservation. Ethnobiol. Lett. 3, 14–15. doi:10.14237/ebl.3.2012.51
Suhling, F., Sahlén, G., Martens, A., Marais, E., and Schütte, C. (2006). Dragonfly assemblages in arid tropical environments: A case study from Western Namibia. Biodivers. Conserv. 15 (1), 311–332. doi:10.1007/s10531-005-2007-6
The Nature Conservancy (2021). GDE Pulse version 2. Available at: https://gde.codefornature.org/#/home.
Weiss, A. D. (2001). Topographic position and landform analysis, 200. San Diego, CA: Poster presentation, ESRI user conference. Available at: http://www.jennessent.com/downloads/TPI-poster-TNC_18x22.pdf.
Zdon, A., and Love, A. H. (2020). Groundwater forensics approach for differentiating local and regional springs in arid Eastern California. USA: Environmental Forensics. Available at: https://www.tandfonline.com/doi/abs/10.1080/15275922.2020.1836075?journalCode=uenf20.
Zdon, A. (2018). Understanding the source of water for selected springs within Mojave trails national monument, California. Environmental Forensics 19 (2), 99–111. Available at: https://www.tandfonline.com/doi/abs/10.1080/15275922.2018.1448909. doi:10.1080/15275922.2018.1448909
Keywords: wetland, ecology, hydrology, discharge, area, temperature, salinity, plant taxa
Citation: Love AH, Zdon A, Fraga NS, Cohen B, Palacios Mejia M, Maxwell R and Parker SS (2022) Statistical evaluation of the similarity of characteristics in springs of the California Desert, United States. Front. Environ. Sci. 10:1020243. doi: 10.3389/fenvs.2022.1020243
Received: 16 August 2022; Accepted: 10 October 2022;
Published: 25 October 2022.
Edited by:
Jenny Davis, Charles Darwin University, AustraliaReviewed by:
Jennifer Lesley Silcock, The University of Queensland, AustraliaCopyright © 2022 Love, Zdon, Fraga, Cohen, Palacios Mejia, Maxwell and Parker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Adam H. Love, YWxvdmVAcm91eGluYy5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.