
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Sci. , 23 September 2022
Sec. Freshwater Science
Volume 10 - 2022 | https://doi.org/10.3389/fenvs.2022.1003207
This article is part of the Research Topic Freshwater Science in the Tropical Anthropocene View all 8 articles
 Carlos Carrasco-Badajoz1,2*
Carlos Carrasco-Badajoz1,2* Carolina Rayme-Chalco1,2
Carolina Rayme-Chalco1,2 Jerry Arana-Maestre3*Daniel Álvarez-Tolentino4,5
Jerry Arana-Maestre3*Daniel Álvarez-Tolentino4,5 Yuri Ayala-Sulca6Marco Sanchez-Peña7,8
Yuri Ayala-Sulca6Marco Sanchez-Peña7,8In the Anthropocene, man has imposed his activities above the natural processes of ecosystems. Rivers have undergone major changes associated with urban centers, so monitoring them is important to determine their status and suitability to provide environmental services. Studies applying the community ecology approach, such as energy flow in aquatic macroinvertebrate assemblages, may be appropriate to determine their status and functioning. This study characterized an Andean river that runs across the city of Ayacucho (Peru). Water physicochemical parameters and biotic quality indices were used and compared with the abundance of trophic guild components and functional feeding groups to demonstrate that the latter are at least equally suitable to estimate water quality. The river section studied ranged from 2640 to 2900 m a.s.l. and was monitored in the wet and dry seasons. Six monitoring stations were established according to the extent of the influence of the urban center. Water physicochemical variables and macroinvertebrate abundances were determined. A Surber net was used for multihabitat sampling; macroinvertebrates were identified to genus level (in most cases) and counted to estimate relative abundances. Trophic guilds (detritivore, predator, herbivore) were formed using the genus data from identification according to the information available in Argentina, Colombia, Ecuador, Bolivia, and Peru. Functional feeding groups were set based on the classification of families. The sensitivity of trophic metrics was assessed by relating them to environmental variables and the Andean Biotic Index (ABI), the Biological Monitoring Working Party adapted for Colombia (BMWP/Col.), and the Ephemeroptera, Plecoptera, Trichoptera (EPT) indices. Environmental variables and biotic indices increased significantly in the most impacted stations, where the abundance of most taxa decreased. The components of trophic guilds and functional groups varied, and those feeding on fine particles increased in sites with a higher urban impact. The metrics based on trophic characteristics were highly correlated with the BMWP/Col and ABI indices. Therefore, these indices can be used to estimate river water quality and provide information on its functioning.
Freshwater ecosystems provide essential environmental goods and services. The ecological characteristics, biodiversity, and productivity of these ecosystems, which influence human well-being, depend primarily on the volume and quality of water in these systems (Albert et al., 2021). However, anthropogenic activities and the prioritization of human needs jeopardize the resources and services of freshwater bodies to the detriment of the present and future generations (Francis, 2014; Piano et al., 2020). Water, building materials, and multiple organisms are extracted from rivers. Rivers are also commonly used to dispose of waste (Rios-Touma et al., 2022). Also, microbasins running through urban centers have been waterproofed, and the flow channel is bordered by barriers that insulate it from its surroundings, influencing water quality and biodiversity (Carpenter et al., 2011). These microbasins are therefore considered hybrid or artificial ecosystems (Francis, 2014). Information on urban rivers is limited, mainly in Latin America, despite the recent increase in studies in other regions (Walteros and Chestano, 2020).
The quality of rivers is a central aspect to monitor. Historically, this has been carried out using physicochemical attributes, although biological indicators have been incorporated more frequently in recent decades (Palottini et al., 2017). Macroinvertebrates are among the biological communities frequently used (Li et al., 2010; Roldan-Pérez, 2016; Pascual et al., 2019), based on taxonomic information, mainly at the family level. The presence (and abundance, in some cases) of macroinvertebrate taxa sensitive to alterations is used as a criterion to assign a water quality category (Rios-Touma et al., 2014). Indices such as BMWP, ABI, EPT, and FBI, among others, are frequently used in Neotropical rivers. However, the information generated from them is inadequate to determine the functioning of these ecosystems from an energy standpoint, which is an important parameter in altered systems.
The characteristics of aquatic macroinvertebrate assemblages are strongly influenced by the local environmental variables. Extreme alterations in the environment from water pollution change the taxonomic composition, species richness, and abundances (Acosta et al., 2009; Roldán-Pérez, 2016; Kengne et al., 2022). They also alter the guilds that are classified into trophic and functional feeding groups according to their feeding type and behavior (Merritt and Cummins, 1996; Ramirez and Gutiérrez-Fonseca, 2014; Merritt et al., 2019; Jiménez et al., 2021). The introduction of food resources, such as organic matter in wastewater either untreated or subjected to basic treatment only, promotes the presence or dominance of groups that are able to use these resources at the expense of others.
The feeding habits of aquatic macroinvertebrates are variable and adaptations are based on the type of food available, time of the year, hydrological regime, and the microhabitat they inhabit (Tomanova et al., 2006; Chara-Serna et al., 2012; Bae and Park, 2016). The feeding strategies of macroinvertebrates allow them to be grouped into trophic guilds; when the behavior and morphological characteristics associated with this activity are considered, macroinvertebrates can be grouped into functional feeding groups (Ramírez and Gutiérrez-Fonseca, 2014; Serna et al., 2022). In urban rivers, the research approach based on the presence and abundance of specific trophic guilds and functional feeding groups can be a valuable tool for estimating the quality and status of these ecosystems (Bae and Park, 2016; Walteros and Castaño, 2020; Jiménez et al., 2021), in addition to providing information on its functioning at the energy level.
Studies on trophic guilds and functional feeding groups of macroinvertebrates are available in temperate regions (Merritt et al., 2019). However, these are scarce in South America; of the few that exist, these mainly address insects (Guzmán-Soto and Tamarís-Turizo, 2014; Motta, 2018; Tamarís-Turizo et al., 2018). It is worth mentioning that the assignment of taxa to guild categories or functional groups is at the species level (Tomanova et al., 2006). However, it is not necessary for studies of functional diversity (Doledec et al., 2000; Gayraud et al., 2003), which facilitates classification when the available taxonomic information is limited. Lists are available of genera and families sorted into trophic levels and functional groups based on the composition of the diet (gastric content) and their morphology associated with the diet for Bolivia (Tomanova et al., 2006); feeding groups for Colombia (Chara-Serna et al., 2012); functional feeding groups and trophic guilds for Colombia (only for Ephemeroptera, Plecoptera, and Trichoptera; Guzman-Soto and Tamaris-Turizo, 2014); and trophic guilds for Peru (Silva and Huamantinco, 2022). In addition, there are publications on the same subject that take as reference the results of the studies described above regarding trophic guilds and functional groups for Latin America (Ramírez and Gutiérrez-Fonseca, 2014), trophic guilds for Colombia (Jiménez et al., 2021), and functional groups for Peru (de Souza and Aguilar, 2022). Physiographic, climatic, and ecosystem heterogeneity contribute to the lack of detailed information for the Neotropical region.
Many Andean rivers are being disturbed by urban centers. This is the case of the Alameda River, which flows across the city of Ayacucho from south to north, showing a marked gradient of human population density settled on its banks. This has led to modifications in channel morphology, the physicochemical quality of water, and macroinvertebrate assemblages. Given this scenario, our working hypothesis is that the assessment of the status of an urban river based on trophic guilds and functional feeding groups yields a similar or better diagnosis than quality indices based on a taxonomic approach at the family level, besides providing information on its trophic functioning.
The Alameda River is located in the department of Ayacucho (southern Peru). It flows across approximately 12.46 km from its source in a system of high-Andean wetlands (at over 4000 m a.s.l.) to its junction with the Huatatas River (Figure 1). It is part of the Mantaro basin on the Amazon slope. The study area is located in the Subtropical Low Montane thorny steppe life zone (ee-MBS, for its acronym in Spanish) (Holdridge, 1987). The mean maximum annual temperature ranges between 25.7°C (November) and 22.8°C (February), and the mean minimum temperature ranges from 5.1°C (July) to 10.7°C (January). The total annual rainfall is 632 mm, peaking in summer when it reaches 109.2 mm (January) (Castro et al., 2020).
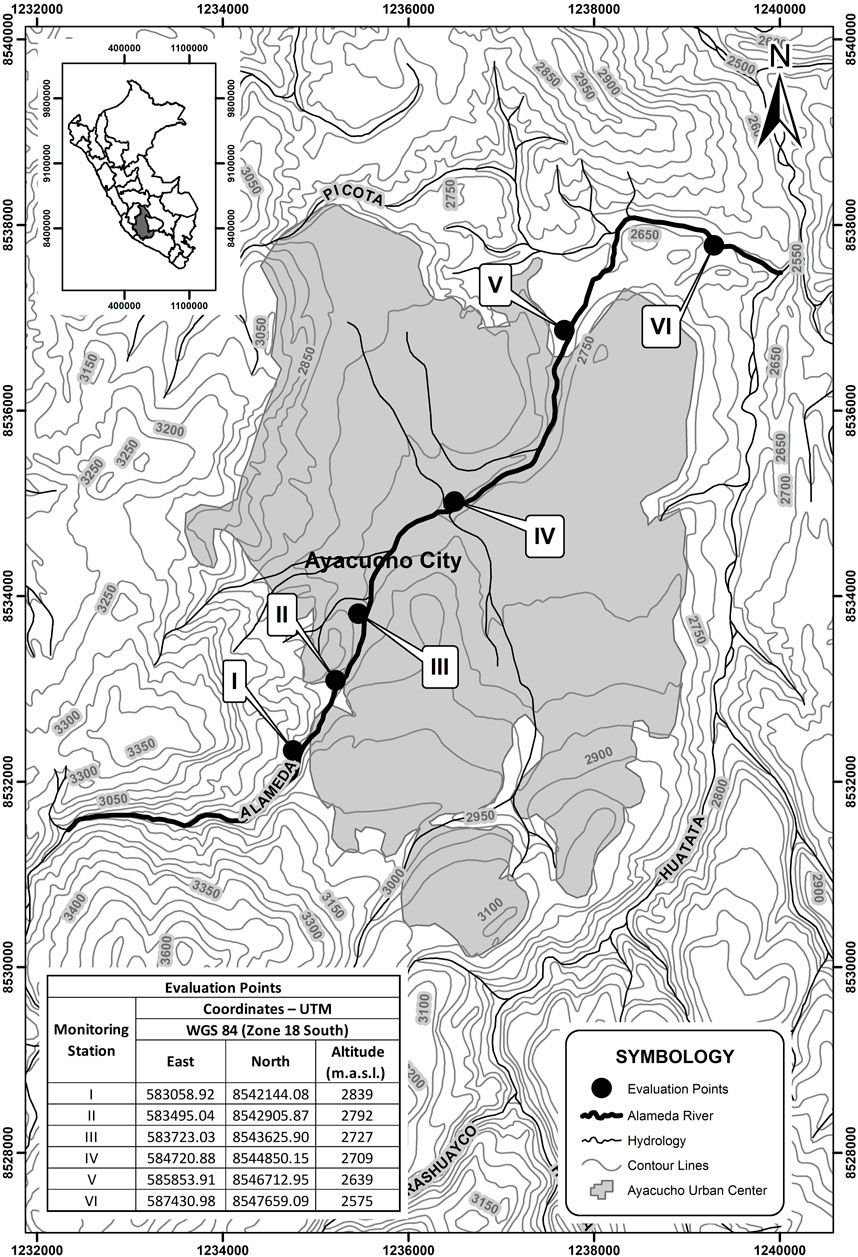
FIGURE 1. Location of the study area in the Alameda River microbasin, Ayacucho (Peru).
The river crosses the urban area of Ayacucho over approximately 5 km, descending 300 m in altitude. The river flows across steep slopes with riparian vegetation dominated by permanent xeric vegetation constituted by Cedrela lilloi, Agave americana, Schinus molle, Opuntia ficus-indica, Baccharis sp., Dodonaea viscosa, Caesalpinia spinosa, Arundo donay, Eucalyptus globulus, Spartium junceum, and Salix chilensis, with the fruit trees Prunus persica, Prunus serotina, Mespilus germanica, Inga feuillei, Persea americana, among others, growing along riverbanks further down. Pennisetum clandestinum is common on downstream banks, as well as vegetable crops of Apium graveolens, Lactuca sativa, Daucus carota, Coriandrum sativum, and Medicago sativa.
The natural river channel is frequently invaded by urban constructions as a result of unrestrained urban growth. Ayacucho shows an annual population growth rate of 3.7%, one of the highest in Peru; it was home to 216 444 inhabitants in 2017 (INEI, 2017). The river running across the city is channeled, even covered below motorways. It is also the final recipient of an artificial drainage system encompassing the historic city center and natural gorges whose microbasins cover a large part of the urban area. In the middle and lower sectors, the river receives wastewater from households with no basic services and effluents from a treatment plant, including physical and biological wastewater treatment.
Six monitoring stations (I, II, III, IV, V, and VI) were established considering aspects of accessibility and location relative to the urban center (Figure 1). Samples were collected in duplicate from November 2019 to February 2020 (wet season) and from July to October 2021 (dry season). Stations I and II were located on the periphery of the city, considered reference sites with no direct urban influence; stations III and IV were located within the city, and stations V and VI downstream (outside the city). Station VI was directly affected by the effluents of a wastewater treatment plant and human settlements.
The environmental variables of water recorded in situ were pH and electrical conductivity, using a Hanna HI 9829 portable multi-parameter analyzer. In the laboratory, total hardness, chlorides, turbidity, nitrates, and phosphates were determined using the methodology recommended by the American Public Health Association (APHA, 2017); the biochemical oxygen demand (BOD5) was determined with a BOD BD-600 (Lovibond) measurement system.
Aquatic macroinvertebrates were sampled employing the multihabitat methodology (Rios-Touma et al., 2014; Carrasco et al., 2020) using a 3.0 mm mesh Surber net (40 cm × 30 cm). Each sample was composed of five collections. Samples were fixed with 96% ethanol (Carrasco et al., 2020); and transported to the laboratory for processing. Macroinvertebrates were identified to genus level whenever possible, using the keys by Dominguez and Fernandez (2009), Prat et al. (2011), and Merrit et al. (2008). Once the specimens were identified and counted, they were placed in 70% ethanol test tubes and deposited in the Limnology Department of the Natural History Museum at the Universidad Nacional de San Cristobal de Huamanga.
The trophic guilds categories considered in this study were predator, herbivore, and detritivore, based on information available for South America (Guzmán-Soto and Tamarís-Turizo, 2014; Reynaga et al., 2017; Silva and Huamantinco, 2022). The “omnivore” category was not considered despite its inclusion in the classification by Ramirez and Gutiérrez-Fonseca (2014), to facilitate the allocation of the identified taxa in any of the above categories. This procedure was not possible in the case of 10 taxa, either for lack of information (Cailloma, Dicrotendipes, Itaura, Leptohyphodes, Limnophora, Lispe, Ochrotrichia, Onconeura, Planariidae) or because the available information is contradictory (Tipulidae).
The functional feeding group categories considered were predator, filter-feeder, perforator, scraper, and collector (Ramírez and Gutiérrez-Fonseca, 2014), and 23 of the 24 families reported were assigned (Planariidae was excluded for lack of information). It is recommended that the allocation to a given functional group or trophic guild be made at the species or genus level (Merrit et al., 2008); however, in the present study, it was done at the family level because of the lack of detailed information, to produce preliminary data on the behavior of functional groups in an urban Andean river.
Water quality estimates were obtained through the indices BMWP/Col (Biological Monitoring Working Party adapted for Colombia) (Roldán, 2003), ABI (Andean Biotic Index) (Acosta et al., 2009), and EPT (Ephemeroptera, Plecoptera, Trichoptera) (Klemm et al., 2002). The first two were estimated according to the presence of families with tolerances ranging from 1 to 10 (lower values indicate a higher tolerance to disturbance); the third is based on the relative abundance of the specimens classified in those three orders. The estimated scores were used for statistical analyses, rather than the quality categories usually derived from the transformed scores.
Comparisons of physicochemical variables between stations and sampling periods were made using the Kruskal Wallis test (p < 0.05), because the data were not normally distributed (p < 0.05). Data on physicochemical and macroinvertebrate variables were square root-transformed for the multivariate classification analysis (dendrogram) and clustering (principal components). The dendrogram was constructed based on the Euclidian distance supplemented by the Simproff test (p < 0.05) (Oyanedel et al., 2008), and the principal component analysis was run with the Bray-Curtis index. The hypothesis of differences between monitoring zones was tested using the multivariate non-parametric analysis of variance NPMANOVA (9999 permutations); Euclidean distance was used for environmental variables and the Bray-Curtis index for biological variables, using the software PAST version 3.02 (Anderson, 2001). This procedure was used for comparisons according to water quality indices, trophic guilds, and functional feeding groups. The representativeness of the biological sampling was estimated using the accumulation curve (Jiménez-Valverde and Hortal, 2003). Finally, non-parametric correlations (Spearman, p < 0.05) were performed between the percentages of trophic groups, functional groups, and water quality indices.
These analyses were performed using the packages Vegan (Oksanen et al., 2017) and FactoMineR (Lê et al., 2008) in R version 3.5.1 (R Core Team, 2015) and the software PAST version 3.02.
The values of the physicochemical parameters measured at each sampling station are shown in Table 1. The variables generally displayed a wide range of values over a short section of the river. An increasing trend was observed from station I to station VI, except for pH: Stations V and VI recorded the highest values; Stations I and II, the lowest values. All variables showed significant differences (p < 0.05) between sampling stations, except for pH. The variables related to organic pollution by wastewater showed 4- to 8-fold increases from station I to station VI for chlorides and conductivity, while nitrates and BOD5 showed increases of 19.5 and 28.7 times, respectively; phosphates was the parameter with the highest increase (90.6 times).

TABLE 1. Physicochemical characteristics (mean and standard deviation) of water at the Alameda River sampling stations, Ayacucho (Peru)
With regard to seasonal variations, total hardness, chlorides, pH, and phosphates were significantly higher in the dry season (p < 0.05), while turbidity was higher in the wet season (p < 0,001); conductivity, nitrates, and BOD5 showed no significant differences (p > 0.05).
The cluster analysis defined five groups, with stations I and II forming a single group (p > 0.05) and the rest stations as separate groups (Figure 2A). In the product biplot of the principal component analysis (Figure 2B) of the relationship between physicochemical variables and sampling stations, the first two components explain more than 98% of the variability; these results are consistent with the cluster analysis. Stations V and VI are associated with total hardness, chlorides, electrical conductivity, turbidity, phosphates, nitrates, and BOD5 and recorded the highest values. Stations I, II, and III are located in an opposite plane for recording the lowest values.
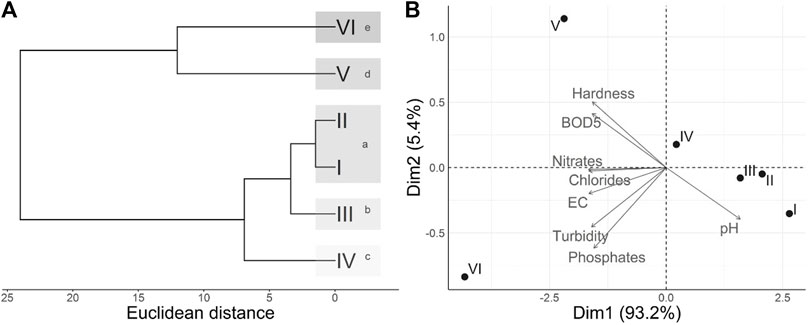
FIGURE 2. (A) Cluster dendrogram based on physicochemical variables of the sampling stations. (B) Principal Component Analysis (PCA) biplot based on physicochemical variables and sampling stations.
The NPMANOVA showed significant differences (F = 76.44; p = 0.0001) when sampling stations were compared, where stations III, IV, V, and VI were different, but stations I and II were not (p = 0.513), confirming the findings with the cluster and PCA analyses.
The rarefaction curve (Figure 3) showed that sample sizes above 30 show no significant increases in taxa richness at any of the six sampling stations, indicating that most species have been collected.
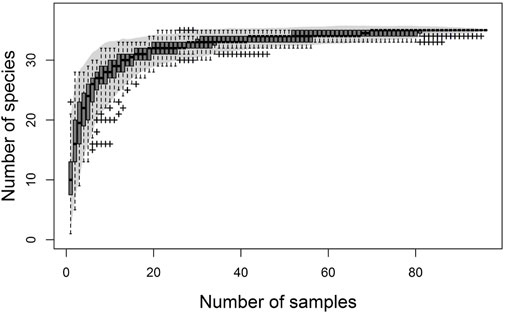
FIGURE 3. Accumulation curve according to the number of samples collected in the Alameda River microbasin, Ayacucho (Peru).
A total of 23 765 individuals were collected, comprising 35 taxa distributed in 11 orders and 24 families (Table 2). The highest diversity and abundance corresponded to the class Insecta, orders Coleoptera, Diptera, Ephemeroptera, Odonata, Plecoptera, Trichoptera, and Megaloptera. The number of taxa decreased gradually by 13 taxa (approximately 56%) as the river flowed through the city from station I to station VI. This same trend is also observed in total abundance, from 9 405 individuals at station I to 155 at station VI. The most abundant taxa were Simulium (Diptera: Simuliidae) and Macrelmis (Coleoptera: Elmidae), with 9 947 and 6 718 individuals, respectively, corresponding to 70.12% of the total number of organisms collected. A total of 11 taxa were recorded at all stations. The following taxa (number of individuals in parentheses) decreased from station I to station VI: Macrelmis (6 718), Dicrotendipes (2 383), Onconeura (607), Penteura (37), Cricotopus (641), Simulium (9 947), Tipulidae (18), Andesiops (703), Aeshnidae (265), and Lumbriculidae (100); the opposite trend was recorded for Alotanypus (333), Psychodidae (316), and Syrphidae (40 individuals), which showed higher abundances at stations V and VI. In terms of orders, Diptera attained the highest number of individuals (14 448) grouped into nine families and 12 genera, followed by Coleoptera (6813 individuals in one family and one genus), and Trichoptera (1 126 individuals, four families, and six genera). The orders Coleoptera, Diptera, Ephemeroptera, Odonata, and Lumbriculida were recorded at all stations, while Plecoptera and Megaloptera were recorded only at stations I, II, and III.

TABLE 2. Taxonomic composition and total abundance of macroinvertebrates at the sampling stations in the Alameda River microbasin, Ayacucho, Peru.
Finally, the abundance of orders recorded over the entire sampling period differed between monitoring stations, with Diptera increasing downstream (p < 0.05), whereas the orders Coleoptera and Ephemeroptera showed the opposite trend (Figure 4).
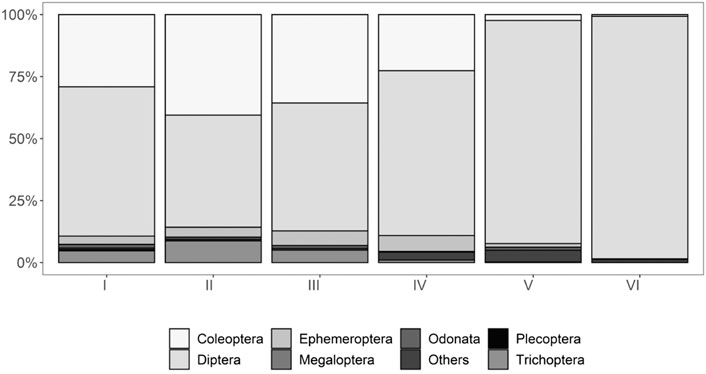
FIGURE 4. Percentage of aquatic macroinvertebrate orders by sampling stations in the Alameda River, Ayacucho (Peru).
According to the season, the most abundant taxa in the wet season were Simulium (7 608 individuals) and Macrelmis (2 304 individuals), while Corydalus (Megaloptera: Corydalidae) and Planariidae (Tricladida) recorded no individuals. By contrast, the most abundant taxa in the dry season were Macrelmis (4 414 individuals), followed by Simulium (2 239 individuals), with no individuals recorded for Tabanus (Diptera: Tabanidae) and Leptohypes (Ephemeroptera: Leptohypidae).
According to the alpha diversity between the sampling stations (Table 3), the species richness (S) was higher at stations I and III and lower at station VI (p > 0.05). The Shannon Wiener diversity index (H) ranged between 1.53 (station I) and 1.96 (station IV) and dominance (D) varied from 0.66 (station I) to 0.79 (station IV); in both cases, the variation was not significant (p > 0.05).
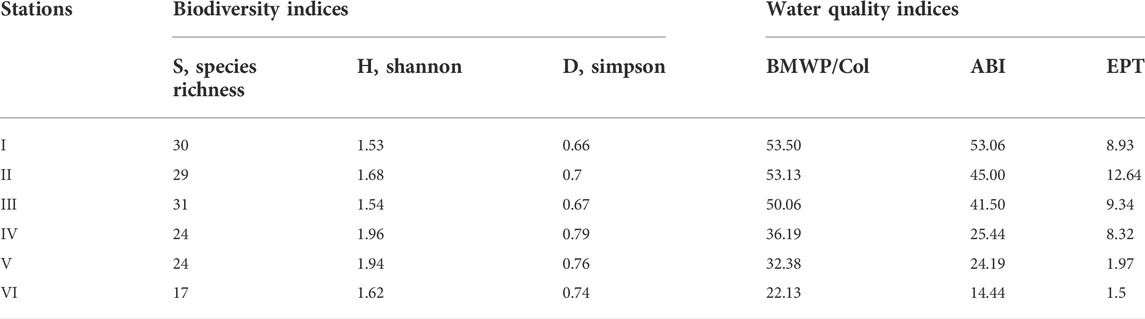
TABLE 3. Mean alpha diversity and water quality indices for the Alameda River sampling stations.
With regard to the water quality indices, BMWP/Col, ABI, and EPT decreased gradually from station I to station VI. According to the quality categories, BMWP/Col lists stations I, II, and III as questionable and stations IV, V, and VI as critical. According to the ABI index, stations I, II and III show moderate water quality, and stations IV, V, and VI have poor quality. Finally, according to the EPT index, all stations had poor quality (Table 3). The NPMANOVA analysis determined that stations I, II, and III are similar to each other (p > 0.05), whereas stations V and VI differ from all other stations (p < 0.01) but are similar to each other (p = 0.001).
The groups forming the trophic guilds showed variable relative abundances between the wet and dry seasons (Figure 5A). The abundance of detritivores and herbivores amounts to more than 98% and 95% for the wet and dry seasons, respectively, with clear trends in abundance observed in the sampling stations. Detritivores showed a 4.9-fold increase in abundance from station I to station VI in the wet season and a 10.6-increase in the dry season. Herbivores showed a declining trend by just over one-half between stations I and VI. The abundance of predators was higher at stations I, II, and III and then decreased in the downstream seasons.
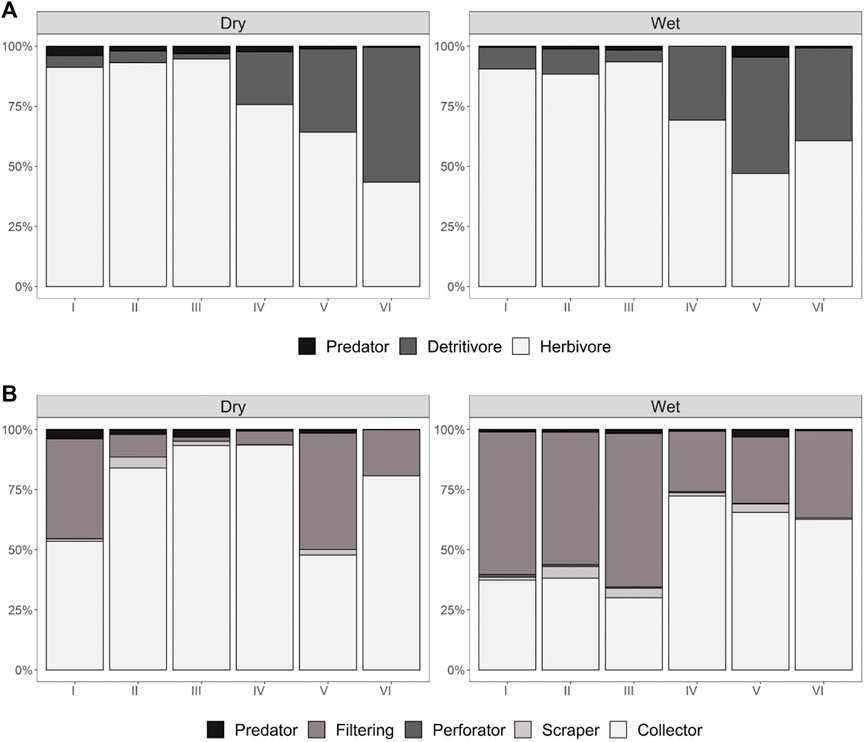
FIGURE 5. Relative abundance by sampling stations and season of the year in the Alameda River. (A) Trophic guilds; (B) Functional groups.
In the functional feeding groups (Figure 5B), filter-feeders and collectors accumulate the highest densities (>95%). Filter-feeders are generally more abundant at stations I and II, decreasing at stations V and VI; the opposite trend was observed in collectors as they increased at stations V and VI (mainly in the wet season). Predators and perforators are most abundant at the upstream sampling stations (I, II, and III), and the latter group is not recorded at station VI. Scrapers are also more abundant at stations I, II, and III and decrease at station VI. The Kruskal-Wallis test confirmed the above trend for both trophic guilds and functional feeding groups, showing a significant difference (p < 0.005). In addition, the NPMANOVA for trophic guilds showed that stations I, V, and VI differ from the rest (p < 0.05), while stations II and III (p = 0.8284) and stations V and VI (p = 0,394) showed no differences. The NPMANOVA results for the functional feeding groups were similar to those for the trophic guilds.
For trophic guilds, the first two PCA components explain 94.6% of the variance (Figure 6A). Detritivores are related to total hardness, chlorides, electrical conductivity, turbidity, phosphates, nitrates, and BOD5, which reached peak values at stations V and VI; on the other hand, herbivores and predators are related to pH, which was higher at stations I and II. Also, in Figure 6B, the first two components explain 98.1% of the variance; herbivores and predators are positively and significantly correlated with the indices BMWP/Col, ABI, and EPT (p < 0.05), whereas detritivores showed a significant negative correlation (p < 0.05).
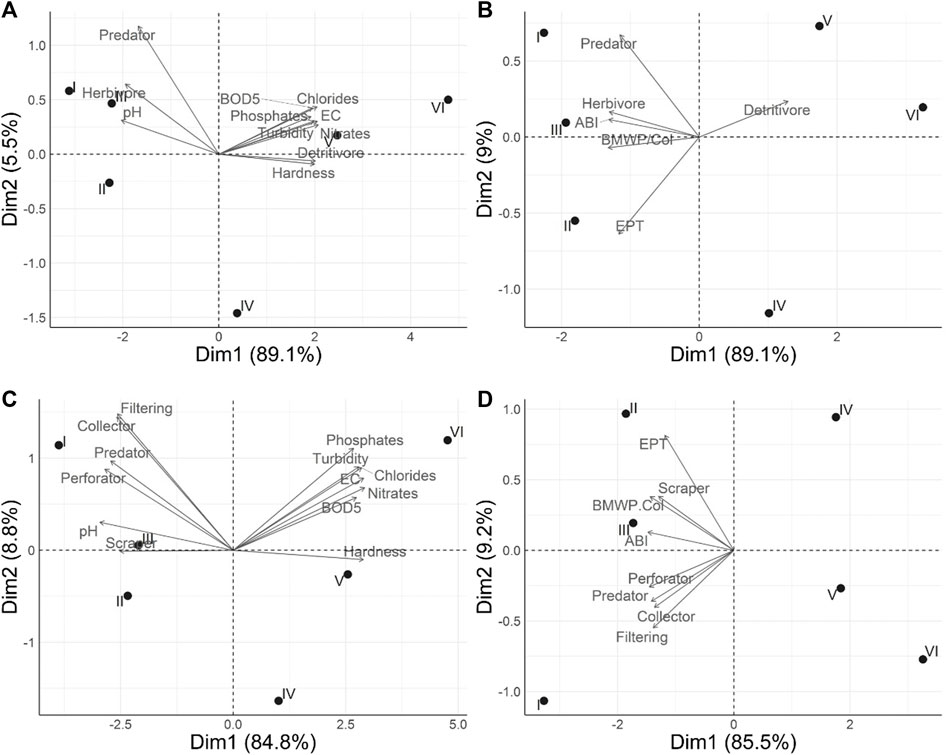
FIGURE 6. Principal Component Analysis (PCA) biplot based on (A) trophic guilds and physicochemical variables, (B) trophic guilds and water quality indices, (C) functional groups and physicochemical variables, and (D) functional groups and water quality indices.
For the functional feeding groups, in Figure 6C (93.6% of the variance explained by the first two components), collectors are associated with total hardness, chlorides, electrical conductivity, total dissolved solids, turbidity, phosphates, nitrates, and BOD5 (reaching peak values at stations V and VI), whereas predators, filter-feeders, perforators, and scrapers were located on the opposite side of the plot, associated with the lowest values of the physicochemical parameters. Figure 6D (94.7% of the variance explained) shows that collectors are associated with the indices BMWP/Col, ABI, and EPT, which reached minimum values at stations V and VI, while predators, filters, perforators, and scrapers exhibit the opposite behavior and are located on the opposite side of the figure.
The indices BMWP/Col and ABI showed a highly significant correlation (<0.01) with all the components of the trophic and functional groups; the EFA was not significantly correlated (p > 0.05) with predators within trophic groups and filter-feeders within functional groups (Table 4). Considering the indices BMWP/Col and ABI, Spearman’s Rho coefficients are positive for predators and herbivores and negative for detritivores within trophic guilds.
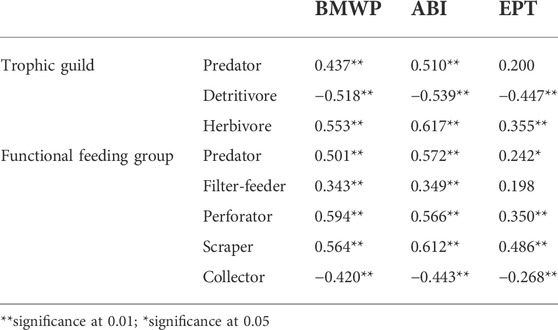
TABLE 4. Spearman’s Rho correlation coefficients of water quality indices with trophic guilds and functional feeding groups
The results obtained in the present study showed an increase in all physicochemical parameters along the river path from station I to station VI (Table 1), with the latter attaining the highest values in almost all the parameters; the exception was pH, which remained relatively stable throughout the sampling period, being slightly alkaline. This remarkable variability between stations is due to the frequent wastewater discharges and excess rainwater flowing through the city and carrying various compounds into the river water, as observed in the Cunas River, another Andean stream located in Peru (Custodio and Chanamé, 2016). The total hardness values increased between stations and seasons (dry and rainy seasons), and the river water can be classified as hard (>120 mg/L) according to the mean values recorded throughout the study. This parameter may affect the production of calcareous exoskeletons and the growth and reproduction of macroinvertebrate communities (Fernández and Volpedo, 2020). This parameter is associated not only with the presence of calcium and magnesium salts but also with chlorides, which is consistent with the increased mean Cl concentrations recorded (Soto, 2010). The presence of chloride is an indicator of wastewater pollution in rivers that limits the distribution of species in aquatic ecosystems; chloride may be highly harmful if it reacts with the cyanide produced by some microorganisms (US Department of Health and Human Services, 1999; Biancucci et al., 2017). Water turbidity in Andean rivers is associated with suspended, colloidal, and dissolved particles in water, which increase markedly in the rainy season. This factor limits the development of microalgae such as chlorophytes, cyanophytes, diatoms, and other photosynthetic organisms that serve as food for the communities studied, and their increase is related to the wastewater discharges in the sections between stations IV and VI (Gómez, 2007). The high water conductivity from stations III to VI is a clear indicator of the entrance of salts from local wastewater discharges; although it is not a water quality parameter, it is an indicator of dissolved salts.
Given the lack of appropriate technologies for removing mineral salts from wastewater, their levels are maintained when discharged into rivers, directly affecting the populations and diversity of local aquatic species. This is consistent with the principal component analysis biplot results shown in Figure 2 (Gómez, 2007; Fernández and Volpedo, 2020). The high values of nitrates, phosphates, and BOD5 in water, as shown in the averages for the seasons sampled in Table 1, are associated with compounds such as detergents, the decomposition of organic compounds, and other nutrients abundant in wastewater (Guarín, 2011; Bolaños-Alfaro et al., 2017). These high values indicate a high organic load at stations V and VI that can serve as food for detritivorous macroinvertebrate groups such as Dicrotendipes and Simulium, which had the largest populations at both stations. In contrast, the stations with the lowest concentrations showed genera that are more sensitive to organic pollutants, such as Claudioperla, Leptohyphodes, and Leptohypes, among others (Mauad et al., 2015).
Aquatic macroinvertebrate assemblages of river ecosystems flowing through urban areas are generally characterized by few taxa, and taxa of pollution-sensitive insects are either poorly represented or absent in such environments (Correa-Araneda et al., 2010; Ríos-Touma et al., 2022). This was also observed in the Alameda River, with a decreasing trend in the abundance of macroinvertebrates belonging to the EPT group (Ephemeroptera, Plecoptera, and Trichoptera) from the upstream stations of Ayacucho (I and II) to those located downstream (V and VI). Some sensitive macroinvertebrates taxa even disappear, such as Claudioperla (Plecoptera: Gripopterygidae) and Cailloma (Trichoptera: Hydrobiosidae), while tolerant taxa such as Psychodidae (Diptera) and Tubificidae (Haplotaxida) showed higher abundances (Roldan, 2003). A similar pattern was described for the urban periphery of the Peruvian Amazon city of Puerto Maldonado—an area also subjected to anthropic pressures (Araujo-Flores, 2014; Ibana-López et al., 2021).
As regards the abundance of macroinvertebrate orders, the Diptera increased downstream, contrasting with the trend of decreasing abundance observed for Coleoptera and Ephemeroptera. The prevalence of preimaginal stages of dipterans is likely due to the wide variety of aquatic habitats where they thrive, which is greater than for other insect orders (Castillo et al., 2018; Omar et al., 2020).
The family Chironomidae (Diptera), besides being present in all the sampling stations, showed the highest richness with six genera, with Dicrotendipes as the most abundant genus. Previous studies showed a similar number of families and the dominance of Chironomidae. This is the case of the Ccollpahuaycco and Chicucha rivers located in the district of Acocro, Huamanga, Ayacucho (Silva and Huamantinco, 2022) and of streams associated with two Ayacuchanan high-Andean wetlands called Guitarrachayocc and Pichccahuasi (Carrasco et al., 2020). Aquatic insects were also the most representative taxa in the Alto Madre de Dios River basin (Arana-Maestre et al., 2021) and in high-Andean rivers that flow across the Camisea Project gas pipeline (Pimentel, 2014) and in the area of influence of the gas pipeline in two departments of southern Peru (Maestre and Carranza, 2017).
Our results differ in the composition and abundance of aquatic macroinvertebrates from other studies on South American rivers with urban influence (Restrepo-Manrique and Salcedo-Reyes, 2018; Castro et al., 2020; Rios-Touma et al., 2022). This is probably due to differences in latitude, altitude, substrates, and ecosystems, as well as to the collection effort employed.
Rivers influenced by urban centers undergo changes in their physicochemical characteristics and biodiversity (Francis, 2014). This behavior is observed in the Alameda River due to the influence of the city of Ayacucho, with a marked increase in the characteristics associated with the input of organic matter from treated and untreated wastewater. Nitrates, phosphates, organic matter, and other characteristics such as electrical conductivity and turbidity show marked increases in urban rivers (Rios-Touma et al., 2022).
Changes in environmental variables affect organism assemblages, as is the case of macroinvertebrates in the present study, reducing the diversity, richness, and abundance of most taxa (except for the most tolerant taxa) (Correa-Araneda et al., 2010; Piano et al., 2020; Walteros and Ramírez, 2020). This effect was also reflected in the values of the indices estimated from the information obtained. Diversity indices tend to increase in assemblages with higher richness and evenness (Shannon), which generally occur in undisturbed environments (Nuñez and Fragoso-Castilla, 2019); however, this effect was not marked in the Alameda River, except for richness.
Indices such as BMWP/Col, ABI, and EPT are useful for estimating the quality of rivers, as they attain lower values associated with more altered environments (Gustavson et al., 2013; Damanik-Ambarita et al., 2016; Trama et al., 2020). These indexes rate the presence of families, who are assigned a score according to their tolerance to changes in their environment (BMWP/Col and ABI) (Ríos-Touma et al., 2014; Roldán-Pérez, 2016) or estimate the relative abundance of the orders Ephemeroptera, Plecoptera, and Trichoptera (Masese and Raburu, 2017). In the river studied, index values decreased at stations with greater urban impact since they showed tolerant families such as Psychodidae, Tubificidae, Syrphidae (Custodio and Chanamé, 2016; Roldán, 2020; Magallón et al., 2021). However, only BMWP/Col and ABI showed quality categories that differentiate one station from another, which is not the case with EPT due to the infrequent presence of organisms in such orders.
The presence and abundance of trophic guild components (predator, detritivore, herbivore) and functional feeding groups (predator, filter-feeder, perforator, scraper, and collector) is in response to the availability of specific food resources (Cummins et al., 2022). Neotropical macroinvertebrates adapt easily because of their high food flexibility (Chará-Serna et al., 2010), and many taxa vary their diet according to their development stage. The availability of a given food type is determined by the presence of microhabitats, the hydrological regime (Merritt and Cummins, 1996; Chará-Serna et al., 2010), microbasin characteristics, and anthropic activities.
Fine debris, such as finely particulate organic matter, is the main food resource in tropical streams (Tomanova et al., 2006), so it is frequently found in the gut contents of macroinvertebrates (Villada-Bedoya et al., 2017; Silva and Huamantinco, 2022) and is directly related to the abundance of collectors (Chara-Serna et al., 2012). The Alameda River—a recipient of treated and untreated wastewater—increases the availability of finely particulate organic matter; this promotes the population growth of organisms that use this food resource and withstand the environmental limitations derived from the change in physicochemical characteristics (pollution). Detritivores (trophic guilds) and collectors (functional feeding groups) increase their abundance in areas impacted by urban activities, while predators and herbivores (trophic guilds), or predators, filter-feeders, perforators, and scrapers (functional groups) decrease.
The presence of detritivores and scrapers is reported as an indicator of sites with better water quality, while predators indicate poorer water quality due to anthropogenic activities (Jiménez et al., 2021). This was not the case in the present study, and this may be due to environmental differences (climatic, geological, and geomorphological) that affect the river systems studied. Therefore, general statements in this regard are unsupported (Boulton et al., 2008), and further research is needed (Tomanova et al., 2006; Chará-Serna et al., 2012; Ramírez and Gutiérrez-Fonseca, 2014). In the stream studied, we observed an association between environmental variables indicating organic pollution (nutrients, BOD5) and detritivores (trophic guilds); however, it is less clear for the functional feeding groups, probably because the taxonomic level used in this case was family.
On the other hand, we observed an unequivocal relationship between quality index scores (ABI, BMWP/Col) and all the categories of trophic guilds and functional feeding groups because there is a marked gradient of human influence in the river studied. This relationship is positive, with the more abundant groups in less impacted areas indicating better river water conditions. By contrast, it is negative with the most abundant groups in the most impacted stations. In the case of the EPT index, no relationship was observed with all of the categories in any of the two groups; besides, this index did not differentiate monitoring stations into quality categories. This difference can be explained because the other two indices have been validated and calibrated for South America, especially ABI for Andean rivers.
These reasons lead us to believe that research on aquatic macroinvertebrates based on trophic guilds and functional feeding groups is an important approach for determining water quality and understanding the effects of human activities on the functioning of aquatic ecosystems from a community ecology perspective. In addition, it is necessary to conduct further research at different latitudes and altitudes in the Neotropics, covering different types and gradients of human impact, as this will allow recording the current variability with the resulting generalization of the results.
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding authors.
Writing—original draft preparation; Review and editing. CC-B, CR-C, JA-M, DA-T, YA-S, and MS-P.
To the Vice-rectorate of Research at the Universidad Nacional de San Cristobal de Huamanga and the General Office of Research and Innovation, for the financing provided. Also, to the Natural History Museum of the Universidad Nacional Mayor de San Marcos and the Research Office of the Universidad Privada del Norte. Likewise, to the Universidad Peruana Los Andes and the Universidad Nacional Intercultural de la Selva Central Juan Santos Atahualpa. Thanks also to Maria Elena Sanchez-Salazar for the English translation of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acosta, R., Ríos, B., Rieradevall, M., and Prat, N. (2009). Propuesta de un protocolo de evaluación de la calidad ecológica de ríos andinos (CERA) y su aplicación a dos cuencas en Ecuador y Perú. Limnética 28 (1), 035–064. doi:10.23818/limn.28.04
Albert, J. S., Destouni, G., Duke-Sylvester, S. M., Magurran, A. E., Oberdorff, T., Reis, R. E., et al. (2021). Scientists’ warning to humanity on the freshwater biodiversity crisis. Ambio 50 (1), 85–94. Scopus. doi:10.1007/s13280-020-01318-8
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26 (1), 32–46. doi:10.1111/j.1442-9993.2001.01070.pp.x
APHA (2017). Standard Methods for the Examination of Water and Wastewater. 23rd Ed. Water Environment Federation, American Public Health Association, American Water Works Association.
Arana-Maestre, J., Álvarez-Tolentino, D., Miranda, R., Tobes, I., Araujo-Flores, J., Carrasco-Badajoz, C., et al. (2021). Distribución altitudinal de macroinvertebrados acuáticos y su relación con las variables ambientales en un sistema fluvial amazónico (Perú). Rev. Acad. Colomb. Cienc. Exactas Fis. Nat. 45 (177), 1097–1112. doi:10.18257/raccefyn.1436
Araújo-Flores, J. (2014). Biodiversidad de Masas de Agua Sometidas a Diferente Presión Antrópica en el Entorno de un Área Urbana de la Amazonía Peruana (Puerto Maldonado. Madre de Dios). Biodivers. Amaz. 4 (4), 17–33. doi:10.55873/rba.v1i1
Bae, M. J., and Park, Y. S. (2016). Responses of the functional diversity of benthic macroinvertebrates to floods and droughts in small streams with different flow permanence. Inland Waters 6 (3), 461–475. doi:10.1080/IW-6.3.891
Biancucci, G. F., González, D., Pérez, A., Ridolfi, A., and Strobl, A. (2017). Manual de procedimiento analíticos toxicológicos para laboratorios de baja complejidad. Argentina: International programme on chemical safety, 186.
Bolaños-Alfaro, J. D., Cordero-Castro, G., and Segura-Araya, G. (2017). Determinación de nitritos, nitratos, sulfatos y fosfatos en agua potable como indicadores de contaminación ocasionada por el hombre, en dos cantones de Alajuela (Costa Rica). Rev. Tecnol. Marcha 30 (4), 15–27. doi:10.18845/tm.v30i4.3408
Boulton, A. J., Boyero, L., Covich, A. P., Dobson, M., Lake, S., and Pearson, R. (2008). “9—are tropical streams ecologically different from temperate streams?,” in Tropical stream ecology. Editor En D. Dudgeon (Cambridge, Massachusetts, United States: Academic Press), 257–284. doi:10.1016/B978-012088449-0.50011-X
Carpenter, S. R., Stanley, E. H., and Vander, M. J. (2011). State of the world’s freshwater ecosystems: Physical, chemical, and biological changes. Annu. Rev. Environ. Resour. 36, 75–99. Scopus. doi:10.1146/annurev-environ-021810-094524
Carrasco, C., Rayme, C., Alarcón, R., Ayala, Y., Arana, J., and Aponte, H. (2020). Macroinvertebrados acuáticos en arroyos asociados con bofedales altoandinos, Ayacucho Perú. Rev. Biol. Trop. 68, 116–S161. doi:10.15517/rbt.v68is2.44344
Castillo, M. M., Barba-Álvarez, R., and Mayorga, A. (2018). Riqueza y diversidad de insectos acuáticos en la cuenca del río Usumacinta en México. Rev. Mex. Biodivers. 89, 45–64. doi:10.22201/ib.20078706e.2018.0.2177
Castro, M., Capurro Leites, L. F., Chalar Marquisá, G., and Arocena, R. (2020). Macroinvertebrados bentónicos indican empeoramiento de la calidad del agua en una zona de influencia urbana en el Río Negro. Bol. La Soc. Zoológica Del Urug. 29 (2), 116–125. doi:10.26462/29.2.8
Chará-Serna, A. M., Chará, J. D., Zúiga, M. C., Pedraza, G. X., and Giraldo, L. P. (2010). Clasificación trófica de insectos acuáticos en ocho quebradas protegidas de la ecorregión cafetera colombiana. Univ. Sci. (Bogota). 15 (1), 27–36. Scopus. doi:10.11144/javeriana.sc15-1.tcoa
Chará-Serna, A. M., Chará, J. D., Zúñiga, M. C., Pearson, R. G., and Boyero, L. (2012). Diets of leaf litter-associated invertebrates in three tropical streams. Ann. Limnol. - Int. J. Lim. 48 (2), 139–144. doi:10.1051/limn/2012013
Correa-Araneda, F., Rivera, R., Urrutia, J., de los Ríos, P., Contreras, Á., and Montoya, F. E. (2010). Efectos de una zona urbana sobre la comunidad de macroinvertebrados bentónicos de un ecosistema fuvial del sur de Chile. Limnetica 29 (2), 183–194. doi:10.23818/limn.29.15
Cummins, K. W., Wilzbach, M., Kolouch, B., and Merritt, R. (2022). Estimating macroinvertebrate biomass for stream ecosystem Assessments. Int. J. Environ. Res. Public Health 19 (6), 3240. doi:10.3390/ijerph19063240
Custodio, M., and Chanamé, F. C. (2016). Analysis of benthic macroinvertebrates biodiversity of Cunas river by means of environmental indicators, Junin-Peru. Sci. Agropecu. 7 (1), 33–44. doi:10.17268/sci.agropecu.2016.01.04
Damanik-Ambarita, M. N., Lock, K., Boets, P., Everaert, G., Nguyen, T. H. T., Forio, M. A. E., et al. (2016). Ecological water quality analysis of the Guayas river basin (Ecuador) based on macroinvertebrates índices. Limnologica 57, 27–59. Scopus. doi:10.1016/j.limno.2016.01.001
De Souza, N. M., and Aguilar, E. (2022). Influencia de factores ambientales en la composición de la comunidad de macroinvertebrados bentónicos y grupos funcionales alimenticios en tres sectores de la cuenca del río Chillón (Lima). Lima, Perú: Universidad Peruana Cayetano Heredia.
Dolédec, S., Olivier, J. M., and Statzner, B. (2000). Accurate description of the abundance of taxa and their biological traits in stream invertebrate communities: Effects of taxonomic and spatial resolution. Fundam. Appl. Limnol. 148, 25–43. doi:10.1127/archiv-hydrobiol/148/2000/25
Domínguez, E., and Fernández, H. R. (2009). Macroinvertebrados bentónicos sudamericanos. Sistemática y biología. Tucumán, Argentina: Fundación Miguel Lillo.
Fernandez, A., and Volpedo, A. (2020). Indicadores físico-químicos: ¿qué, cómo y cuánto reflejan la calidad del agua? En La bioindicación en el monitoreo y evaluación de los sistemas fluviales de la Argentina: Bases para el análisis de la integridad ecológica. Argentina: Eudeba. https://ri.conicet.gov.ar/handle/11336/142626
Francis, R. A. (2014). Urban rivers: Novel ecosystems, new challenges. WIREs Water 1 (1), 19–29. Scopus. doi:10.1002/wat2.1007
Gayraud, S., Statzner, B., Bady, P., Haybachp, A., Schöll, F., Usseglio-Polatera, P., et al. (2003). Invertebrate traits for the biomonitoring of large European rivers: An initial assessment of alternative metrics. Freshw. Biol. 48 (11), 2045–2064. doi:10.1046/j.1365-2427.2003.01139.x
Gómez, L. M. (2007). Microalgas: Aspectos ecológicos y biotecnológicos. Rev. Cubana Quím. 19 (2), 3–21.
Guarín, L. J. (2011). Estandarización de las técnicas de fosfatos y cloruros en aguas crudas y tratadas para el laboratorio de la Asociación Municipal de Acueductos Comunitarios (AMAC) en el municipio de Dosquebradas. Colombia: Universidad Tecnológica de Pereira. https://hdl.handle.net/11059/2337.
Gustavson, S. S., Cosme, L. A., and Trama, F. A. (2013). Macroinvertebrados bentónicos como indicadores de la calidad de agua en la microcuenca San Alberto, Oxapampa, Perú. Apunt. Cienc. Soc. 3 (2), 124–139. Article 2. doi:10.18259/acs.2013016
Guzmán-Soto, C. J., and Tamarís-Turizo, C. E. (2014). Hábitos alimentarios de individuos inmaduros de Ephemeroptera, Plecoptera y Trichoptera en la parte media de un río tropical de montaña. Rev. Biol. Trop. 62, 169–178. doi:10.15517/rbt.v62i0.15786
Ibana-Lopez, K., Sihuay-Perales, M., Garate-Quispe, J., Araújo-Flores, J., Herrera-Machaca, M., Alarcón, G., et al. (2021). Contaminación de agua superficial de la periferia urbana de Puerto Maldonado, al sureste de la amazonia peruana. Rev. Investig. Vet. Peru. 32 (6), e20365–14. doi:10.15381/rivep.v32i6.20365
INEI (2017). Características de la población. Censos Nacionales de Población y Vivienda. Perú: Instituto Nacional de Estadística e Informática.
Jiménez, R., Barnuevo, E., Timbe, B., and Astudillo, P. X. (2021). El uso de gremios tróficos en macroinvertebrados acuáticos como herramienta de monitoreo en los Altos Andes del Sur de Ecuador. Neotropical Biodivers. 7 (1), 310–317. doi:10.1080/23766808.2021.1953891
Jiménez-Valverde, A., and Hortal, J. (2003). Las curvas de acumulación de especies y la necesidad de evaluar los inventarios biológicos. Rev. Iber. Aracnol. 8, 151–161. https://dialnet.unirioja.es/servlet/articulo?codigo=787222
Kengne, J., Foto, S., Meyer, A., Leprêtre, A., and Usseglio-Polatera, P. (2022). Relationships between physico-chemical parameters and taxonomic structure of benthic macroinvertebrate assemblages in streams of west Cameroon. Water 14 (9), 1490. doi:10.3390/w14091490
Klemm, D. J., Lewis, P., Fulk, F., and Lazorchak, J. (2002). Macroinvertebrate field and laboratory methods for evaluating the biological integrity of surface waters. Washington, D.C. U.S. Environmental Protection Agency. EPA/600/4-90/030 (NTIS PB91171363).
Lê, S., Josse, J., and Husson, F. (2008). FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 25 (1), 1–18. doi:10.18637/jss.v025.i01
Li, L., Zheng, B., and Liu, L. (2010). Biomonitoring and bioindicators used for river ecosystems: Definitions, approaches and trends. Procedia Environ. Sci. 2, 1510–1524. doi:10.1016/j.proenv.2010.10.164
Maestre, J. A., and Carranza, C. C. (2017). Macroinvertebrados acuáticos y caracterización ecológica de los ambientes dulceacuícolas del área de influencia del gasoducto PERÚ LNG en los departamentos de Ica y Huancavelica. Rev. del Inst. Investig. Fac. Minas, Metal. ciencias geográficas 20 (40), 86–93. doi:10.15381/iigeo.v20i40.14394
Magallón, G., Escalera, C., López-López, E., Sedeño-Díaz, J. E., López Hernández, M., Arroyo-Damián, M., et al. (2021). Water quality analysis in a subtropical river with an adapted biomonitoring working party (BMWP) index. Diversity 13 (11), 606. doi:10.3390/d13110606
Masese, F. O., and Raburu, P. O. (2017). Improving the performance of the EPT Index to accommodate multiple stressors in Afrotropical streams. Afr. J. Aquatic Sci. 42 (3), 219–233. Scopus. doi:10.2989/16085914.2017.1392282
Mauad, M., Miserendino, M. L., Risso, M. A., and Massaferro, J. (2015). Assessing the performance of macroinvertebrate metrics in the challhuaco-ñireco system (northern patagonia, Argentina). Iheringia. Ser. Zool. 105 (3), 348–358. doi:10.1590/1678-476620151053348358
Merrit, R. W., Cummins, K. W., and Berg, M. B. (2008). An introduction to the aquatic insects of North America. 2nd ed. Dubuque, Iowa, United States: Kendall/Hunt Publishing Company.
Merritt, R. W., Cummins, K. W., and Berg, M. B. (2019). An introduction to the aquatic insects of North America. Dubuque, Iowa, United States: Kendall Hunt Publishing Company.
Merritt, R. W., and Cummins, K. W. (1996). Trophic relationships of macroinvertebrates. Dubuque, Iowa, United States: Kendall/Hunt.
Motta, A. J. (2018). Modelación de idoneidad de hábitat de la comunidad y grupos funcionales de macroinvertebrados. Available at: http://repository.javeriana.edu.co/handle/10554/43325 (Accessed July 9, 2022).
Nuñez, J. C., and Fragoso-Castilla, P. J. (2019). Uso de Macroinvertebrados Acuáticos como Bioindicadores de Contaminación del Agua de la Ciénaga Mata de Palma (Colombia). Inf. Tecnol. 30 (5), 319–330. Scopus. doi:10.4067/S0718-07642019000500319
Oksanen, J., Kindt, R., and O’Hara, B. (2017). Vegan: Community ecology package. R. Package Version 1.6-10 2, 88. https://cran.r-project.org/web/packages/vegan/vegan.pdf
Omar, S. A. S., Rak, A. E., Sanusi, A. F. A., Yusoff, A. M., Hajisamae, S., and Shukor, A. M. (2020). Aquatic insect (larvae) distribution and assemblages at river intakes in pergau lakes. IOP Conf. Ser. Earth Environ. Sci. 549 (1), 012044–012048. doi:10.1088/1755-1315/549/1/012044
Oyanedel, A., Valdovinos, C., Azocar, M., Moya, C., Mancilla, G., Pedreros, P., et al. (2008). Patrones de distribución espacial de los macroinvertebrados bentónicos de la cuenca del río Aysen (Patagonia Chilena). Gayana (Concepción) 72 (2), 241–257. doi:10.4067/S0717-65382008000200011
Pallottini, M., Cappelletti, D., Fabrizi, A., Gaino, E., Goretti, E., Selvaggi, R., et al. (2017). Macroinvertebrate functional trait responses to chemical pollution in agricultural–industrial landscapes. River Res. Appl. 33 (4), 505–513. Scopus. doi:10.1002/rra.3101
Pascual, G., Iannacone, J., and Alvariño, L. (2019). Macroinvertebrados bentónicos y ensayos toxicológicos para evaluar la calidad del agua y del sedimento del río Rímac, Lima, Perú. Rev. Investig. Vet. Peru. 30 (4), 1421–1442. doi:10.15381/rivep.v30i4.17164
Piano, E., Souffreau, C., Merckx, T., Baardsen, L. F., Backeljau, T., Bonte, D., et al. (2020). Urbanization drives cross-taxon declines in abundance and diversity at multiple spatial scales. Glob. Change Biol. 26 (3), 1196–1211. Scopus. doi:10.1111/gcb.14934
Pimentel, H. F. (2014). Análisis desde la perspectiva de los índices bióticos, EVCA-agua y manejo adaptativo; usando macroinvertebrados bentónicos en ríos altoandinos—Camisea 2009-2012. Lima, Perú: Universidad Nacional Agraria La Molina. http://repositorio.lamolina.edu.pe/handle/20.500.12996/1906.
Prat, N., Rieradevall, M., Acosta, R., and Villamarín, C. (2011). Guía para el reconocimiento de las larvas de Chironomidae (Diptera) de los ríos altoandinos de Ecuador y Perú. Grupo de Investigación FEM Departamento de Ecología. Barcelona, España: Universidad de Barcelona.
R Core Team (2015). A language and environment for statistical computing. R. Foundati 2 (3.5.1), 1–135. Computer software. Available at: https://www.r-project.org/.
Ramírez, A., and Gutiérrez-Fonseca, P. E. (2014). Functional feeding groups of aquatic insect families in Latin America: A critical analysis and review of existing literature. Rev. Biol. Trop. 62, 155–167. doi:10.15517/rbt.v62i0.15785
Restrepo-Manrique, R., and Salcedo-Reyes, J. C. (2018). Application of physico-chemical indexes and BMWP/Col. To establish the impact of urbanization on Río Frío (Bucaramanga, Colombia). Univ. Sci. (Bogota). 23 (1), 109–128. doi:10.11144/javeriana.sc23-1.aopc
Reynaga, M. C., Dávalos, N., and Molineri, C. (2017). Variaciones espaciales y ontogenéticas en la dieta de un plecóptero de amplia distribución Claudioperla tigrina Klapálek (Plecoptera: Gripopterygidae). Rev. Biol. Trop. 65 (3), 1174–1184. doi:10.15517/rbt.v65i3.23865
Rios-Touma, B., Acosta, R., and Prat, N. (2014). The Andean Biotic Index (ABI): Revised tolerance to pollution values for macroinvertebrate families and index performance evaluation. Rev. Biol. Trop. 62 (0), 249–273. doi:10.15517/rbt.v62i0.15791
Ríos-Touma, B., Villamarín, C., Jijón, G., Checa, J., Granda-Albuja, G., Bonifaz, E., et al. (2022). Aquatic biodiversity loss in Andean urban streams. Urban Ecosyst., 1–11. doi:10.1007/s11252-022-01248-1
Roldán, G. (2003). Bioindicación de la calidad del agua en Colombia: Uso del método BMWP/Col. Antioquía, Colombia: Universidad de Antioquia.
Roldán, G. (2020). Revisión histórica de la limnología en Colombia. Rev. Acad. Colomb. Cienc. Exactas Fis. Nat. 44 (171), 303–328. doi:10.18257/raccefyn.1056
Roldán-Pérez, G. (2016). Los macroinvertebrados como bioindicadores de la calidad del agua: Cuatro décadas de desarrollo en Colombia y Latinoamérica. Rev. Acad. Colomb. Cienc. Exactas Fis. Nat. 40 (155), 254–274. doi:10.18257/raccefyn.335
Serna, J. P., Fernandez, D., Velez, F., Ruiz, J., Breckling, B., and Aguirre, N. (2022). Altitudinal distribution of the functional feeding groups of aquatic macroinvertebrates using an ecological network in Andean streams. Rev. Biol. Trop. 70, 82–95. doi:10.15517/rev.biol.trop.v70i1.46904
Silva, M. A., and Huamantinco, A. A. (2022). Preferencia de microhábitat y gremios tróficos de macroinvertebrados acuáticos en ríos altoandinos, Ayacucho, Perú. Limnetica 41 (1), 1–16. doi:10.23818/limn.41.01
Soto, F. (2010). La dureza del agua como indicador básico de la presencia de incrustaciones en instalaciones domésticas sanitarias. Ing. Investig. Tecnol. 11 (2), 167–177. doi:10.22201/fi.25940732e.2010.11n2.014
Tamaris-Turizo, C. E., Pinilla, G. A., and Muñoz, I. (2018). Trophic network of aquatic macroinvertebrates along an altitudinal gradient in a Neotropical mountain river. Rev. Bras. Entomol. 62 (3), 180–187. doi:10.1016/j.rbe.2018.07.003
Tomanova, S., Goitia, E., and Helešic, J. (2006). Trophic levels and functional feeding groups of macroinvertebrates in neotropical streams. Hydrobiologia 556 (1), 251–264. Scopus. doi:10.1007/s10750-005-1255-5
Trama, F. A., Salcedo, S. A., Demarcy, L., Erbure, L., Jara, B. A., Muñoz, F. E., et al. (2020). Índices de calidad de habitat y macroinvertebrados en siete Cuencas del Parque Nacional Yanachaga Chemillén y su Zona de Amortiguamiento:. Rev. Peru. Biol. 27 (2), 149–168. doi:10.15381/rpb.v27i2.16730
US Department of Health and Human Services (1999). Agency for toxic substances and disease registry—Atsdr. United States: ATSDR. http://www.atsdr.cdc.gov/atsdrhome.html.
Villada-Bedoya, S., Triana-Moreno, L. A., and Dias, L. G. (2017). Grupos funcionales alimentarios de insectos acuáticos en quebradas andinas afectadas por agricultura y minería: Functional feeding groups of aquatic insects in Andean streams affected by agriculture and mining. Caldasia 39 (2), 370–387. doi:10.15446/caldasia.v39n2.62800
Walteros, J. M., and Castaño, J. M. (2020). Composición y aspectos funcionales de los macroinvertebrados acuáticos presentes en una microcuenca de cabecera en los Andes de Risaralda, Colombia. Rev. Acad. Colomb. Ciencias Exactas, Físicas Nat. 44 (171), 581–592. doi:10.18257/raccefyn.1085
Keywords: biomonitoring, biotic indices, anthropic pressure, benthos, trophic web
Citation: Carrasco-Badajoz C, Rayme-Chalco C, Arana-Maestre J, Álvarez-Tolentino D, Ayala-Sulca Y and Sanchez-Peña M (2022) Aquatic macroinvertebrate trophic guilds, functional feeding groups, and water quality of an andean urban river. Front. Environ. Sci. 10:1003207. doi: 10.3389/fenvs.2022.1003207
Received: 26 July 2022; Accepted: 23 August 2022;
Published: 23 September 2022.
Edited by:
Tatenda Dalu, University of Mpumalanga, South AfricaReviewed by:
Linton Munyai, University of Venda, South AfricaCopyright © 2022 Carrasco-Badajoz, Rayme-Chalco, Arana-Maestre, Álvarez-Tolentino, Ayala-Sulca and Sanchez-Peña. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carlos Carrasco-Badajoz, Y2FybG9zLmNhcnJhc2NvQHVuc2NoLmVkdS5wZQ==; Jerry Arana-Maestre, amFyYW5hbUB1bm1zbS5lZHUucGU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.