 Malicha L. Hare
Malicha L. Hare Yong D. Wang1,3*
Yong D. Wang1,3* Abule E. Gedda
Abule E. Gedda- 1Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Ürümqi, China
- 2University of Chinese Academy of Sciences, Beijing, China
- 3National Engineering Technology Research Center for Desert-Oasis Ecological Construction, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Ürümqi, China
- 4The Federal Democratic Republic of Ethiopia, Pastoral Area Development Commission of the Oromia Regional State, Finfine, Ethiopia
- 5Wageningen University and Research (SNV-Ethiopia), Research and Innovation Department, Finfine, Ethiopia
High bush density and cover have negative effects on the environment in general and on livestock production, in particular in savanna rangelands. The study examined the effects of bush control techniques on woody sapling recruitment in the low land savanna region. The study consisted of 21 plots located next to each other on an area of 5.3 ha encroached by Acacia mellifera and Acacia reficiens trees. Seven bush control treatments were assigned randomly to the plots and replicated three times. In the bush clearing plot, woody species were removed at ground level using a handsaw. In the tree thinning; thinning and grazing; thinning and browsing; thinning and fire, and thinning with fire-browsing treatments, trees were thinned to the equivalents of 60% of the density of that of the control plot. Positive changes in sapling densities were observed in the clearing, thinning, and thinning and grazing treatments. In contrast, the control, thinning and browsing, and thinning and fire treatments presented negative changes in densities. A change in sapling density was highest in the cleared plot, whereas it was lowest in the thinning, fire, and browsing treatment. The correlation between changes in sapling densities and new recruitments was highly (p < 0.001) significant in the clearing, thinning, and thinning and grazing treatments thus increased sapling recruitment with the proportion greater for the clearing treatment. The significant correlation induced average sapling recruitment in the control, and low in the thinning with browsing and thinning with fire treatments. The relationship was insignificant in the thinning, fire, and browsing combination treatment and resulted in the lowest rate of saplings’ recruitment. Postthinning techniques reduced woody tree recruitment and would sustain open savanna under the continuous management plan.
1 Introduction
Savannas are described by a continuous herbaceous layer with a widely spaced woody plant species (Scholes and Archer, 1997; Desta and Coppock, 2004). Study reports (Smit, 2004; Vadigi, 2013) have indicated that density and distribution of trees in savannas maintain ecosystem structure. Woody plants in savannas create favorable microenvironments through deposition of leaf litter and shading and habitats that can support a great diversity of flora and fauna (Smit, 2004). Furthermore, trees increase plant species richness and structural complexity by providing a distinct microclimate under subcanopy areas with increased soil nutrients (Manning et al., 2006). African savannas are an important wildlife habitat, provide forage to livestock, and are also a major source of food, fuel wood, and other products (Dalle et al., 2006a; Solomon et al., 2006b; Mworia et al., 2008a). However, an increase in woody plants density is considered a serious problem to the quality, productivity, economic viability of rangelands, and ecological integrity of savanna ecosystem (Ward 2005; Archer et al., 2017; Belayneh and Tessema, 2017). Ecologists are concerned about the loss of biodiversity as savanna grassland is lost to bush encroachment (Solomon and Mblambo, 2010; Smit, 2014). In African savannas, the problem of bush encroachment has also raised concerns among pastoralists whose livelihood depends on livestock production.
The competition for different resources between grass and woody plants has been widely documented as an important driver of savanna dynamics (Riginos, 2009; Cramer et al., 2010; Ward and Esler, 2010). The survival and growth of tree seedlings/saplings most often are suppressed by competition within the herbaceous layer (Riginos and Young, 2007) and are mainly influenced by the availability of light, water, and soil nutrients (Kraaij and Ward, 2006). Riginos (2009) found that grass competition strongly suppressed all demographic stages of tree growth. When grass was removed, tree growth rates doubled for all tree age classes. A healthy grass sward and sound grazing management may limit the ability of woody plants to germinate and establish, therefore controlling the density of woody plants (Hagenah et al., 2009; Cramer et al., 2010). Grasses may also provide moist microclimatic conditions (Grellier et al., 2012) and cover from browsers, facilitating tree sapling establishment (Riginos and Young, 2007; Seymour, 2008).
Solar energy is the principal resource requirement for plant life, and plants compete for it as they struggle to establish (Holmgren et al., 1997). The intertree competition has been noted to have an important effect on woody plant density and structure in savanna (Wiegand et al., 2006). Wiegand et al. (2008) indicated that self-thinning will take place in woody layer as competition for light and water increases. During this process, the different plant species that are adapted to specific microenvironment conditions will colonize the gaps in the canopy and the various subcanopies (Scholes and Archer 1997; Reynolds and Ford, 2005). As shading becomes more intense, seedling and grass establishments decrease, and the vegetation physiognomy will progress toward larger, more widely spaced woody individuals (Reynolds and Ford 2005; Wiegand et al., 2006).
The amount of water may also variably affect all growth stages of vegetation in savannas (Scholes et al., 2002; Bond, 2008). High and frequent rainfall has been shown to increase the establishment of Acacia seedlings (Ward 2005; Kraaij and Ward 2006). Grasses on the other side can establish themselves in the presence of low rainfall (Menaut, 1983). The length and severity of the dry period limit seed germination and establishment, resulting in decline of woody plants in arid savannas (Wiegand et al., 2006). A shortage of water has been recorded to result in the death of seedlings and mature trees (Mueller et al., 2005; Dalle et al., 2006a). The favorable rainfall events for woody plant establishment are thus required in domination of the savanna vegetation (Wiegand et al., 2006).
Factors such as fire and herbivory generally cause high mortality of establishing woody plants (Sankaran et al., 2005). Fire plays an important role in affecting woody plant community structure in African savannas, controlling tree density and improving fodder quality (Angassa and Oba, 2009; Midgley et al., 2010; Devine et al., 2015). Frequent and hot fires limit woody density and cover and maintain trees in a juvenile state by killing seedlings/saplings, retarding the transition of savanna to closed woodlands system (Wiegand et al., 2006; Devine et al., 2015). Herbivores are the major drivers of vegetation dynamics through directly reducing the abundance of the plants they consume and altering the competitive interactions between woody and herbaceous layers (Wiseman et al., 2004; Riginos and Young, 2007). The effects of herbivores on savanna ecosystems will vary, depending on the vegetation type, the herbivore, and the environment. Bond (2008) found herbivory together with fire to be key determinants of vegetation structure and other ecosystem functions. Browsers prevent woody plant recruitment to matured individuals attributed to their browsing-induced mortality of woody seedlings and saplings (Augustine and McNaughton, 2004; Sankaran et al., 2008). On the contrary, intensive grazing is normally associated with an increase in woody density (Riginos and Young, 2007). Overgrazing mainly by cattle has been found to establish woody seedling, reduce competition from grazed grasses, and reduce fire frequency and intensity due to lowered grass-fuel loads (Oba et al., 2000a; Roques et al., 2001).
The Boran savanna of southern Ethiopia supports a great diversity of livestock. The livestock are sources of food, income, prestige, and security in times of hardship. They depend on native vegetation for forage (Coppock, 1994; Angassa, 2005). The Borana pastoralists traditionally practiced strategic grazing management to avoid local overstocking around the scarce dry season water sources. They used different grazing areas in the dry and wet seasons (Coppock, 1994). This management regimen, variable climatic events, and the planned use of range burning maintained a relatively stable tree–grass balance (Angassa, 2002). However, the conditions in the Borana rangelands were changed, and the communal grazing land was encroached by trees of different species, which in turn reduced the herbage composition and production of the rangelands (Coppock, 1994; Dalle et al., 2006a). Increased grazing pressure, suppression of fire, exclusion of browsing ungulates, and stochastic rainfall events have been reported to increase bush density and resulted in a general decline in rangeland production (Oba et al., 2000b; Desta and Coppock, 2002; Angassa and Oba, 2008) and negatively affected pastoral production in the area (Dalle et al., 2006a). Manual clearing of woody plants was initiated as an option to reduce the adverse effects of bush encroachment on rangeland production and people’s livelihood (Forrest et al., 2015). However, removing of woody plants through clearing had no long-term positive effects on bush density and livestock forage production (Dalle et al., 2006a; Angassa and Oba, 2009). Bush clearing (BC) appears to be critically important in the beginning of the new encroachment process because it creates opening for tree seeds to germinate and seedlings/saplings/woody plants to establish and reoccupy the cleared areas, particularly when the herbaceous seed bank is depleted (Masters and Sheley 2001; Smit, 2004).
Smit (2014) suggested the option of tree thinning rather than tree clearing to implement a sustainable woody–grass ratio. This can be achieved by tree thinning to a predetermined density, after which postthinning operations, including low-level stocking rates with the control of regeneration in woody vegetation (Higgins et al., 2007), follow to keep the woody–grass ratio in equilibrium (Smit, 2004; Archer et al., 2017). Tree thinning, by means of mechanical or some other method, results in immediate changes in composition between woody and herbaceous plants (Smit, 2005; Smit 2014). However, Smit (2004) described that the results of woody plant removal may differ between vegetation types, with the outcome determined by both negative and positive responses to tree removal. Therefore, there is a need for selective thinning to be applied. Selective thinning has the advantage that individual species or plants can be removed from the system, and others are allowed to remain. Through selective tree thinning, the development of structured savanna with large trees is encouraged, and these large trees are able to suppress the establishment of new seedlings. This is particularly noticeable with Acacia species.
Understanding the effect of bush control techniques on woody sapling establishment is important for the management of tree density in grazing lands. Studies (Angassa and Oba, 2009; Bikila et al., 2014) have reported the effect of manual clearing on woody plants in terms of stump mortality and coppicing in the area. Nevertheless, the effects of BC and thinning practices on tree density, cover, and recruitment (by sapling) have yet been not well documented. Therefore, the objectives of this study were (i) to evaluate the influence of bush control treatments on tree density and cover, (ii) to investigate the effects on woody sapling recruitment, and (iii) to evaluate responses of individual woody species in terms of sapling establishment and the relationship between changes in sapling population and new recruitments concerning the individual treatments’ effects and species (i.e., damage due to treatments). The hypothesis tested was:
Ho: Bush control technique does not have effect on the density, cover, and recruitment of woody plants.
2 Materials and Methods
2.1 Study Area
The Borana rangelands are located in the southern part of Ethiopia between 4°36′ and 6°38′ N latitude and 36°43′–42°30′ E longitude (Figure 1). The altitude ranges from 900 to 1,650 m above sea level (m.a.s.l.) (Coppock, 1994). The weather is characterized by erratic and variable rainfall, with most areas receiving between 238 and 896 mm annually. Rainfall is bimodal with 60% of the total rainfall expected during March to May and the remaining expected during September to November (Angassa and Oba, 2007). The geology of the study area is dominated by 40% quaternary deposits, 38% basement complex formations, and 20% volcanic (Coppock, 1994). The soil types of the region comprise 53% sand, 30% clay, and 17% silt, which is sandy clay loam. The region is dominated by savannah types of vegetation containing a mixture of perennial grasses and woody plants (Desta and Coppock, 2004). The typical African savanna vegetation consists of a diverse range of tree–grass mixtures with the grass cover relatively continuous and woody cover discontinuous (Scholes and Archer, 1997).
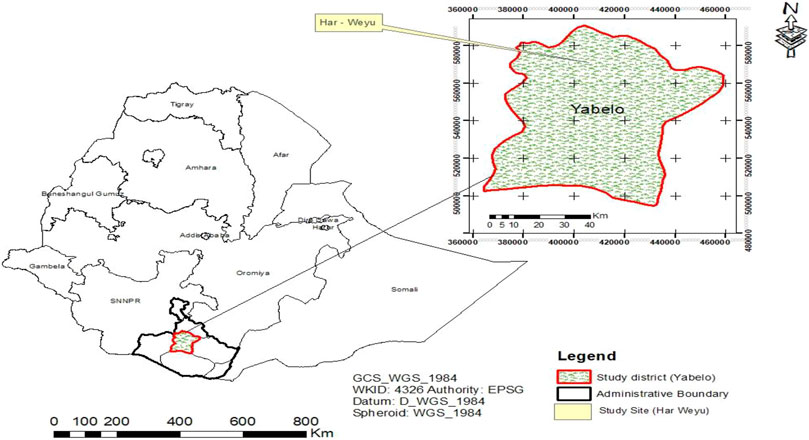
FIGURE 1. Map of the Federal Democratic Republic of Ethiopia showing the administrative boundary of the study zone, district, and site within the Oromiya region.
2.2 Study Procedures
2.2.1 Site Selection and Description
Site selection criteria were developed, and representative grazing areas invaded by woody species were assessed through a participatory reconnaissance survey by a team comprising community elders who have enough knowledge about the history of rangeland vegetation and experts. Accordingly, the Har Weyu communal grazing area was selected as the study site. The coordinate of the experimental site was recorded (Supplementary Appendix Table A1) with a geographical positioning system and plotted on an Ethiopian map, using GIS techniques (Figure 1). The site has an altitude of 1,265 m.a.s.l. The vegetation consisted mainly of closed savanna dominated by Acacia mellifera and Acacia reficiens trees with herbaceous species almost absent. According to the elder’s opinion, no burning of the grazing areas took place for decades due to the limited use of range fire. Bush density greatly reduced rangeland production and hence livestock carrying capacity, especially for grass-dependent cattle.
2.2.2 Sampling Design
The study site consisted of 21 plots (each 50 × 50 m) located next to each other on an area of 5.3 ha encroached by A. mellifera and A. reficiens trees. Seven bush encroachment control treatments (i.e., CO, BC, T, T + G, T + B, T + F, and T + F + B) were assigned randomly to the plots and replicated three times. The control (CO) plot represented the current bush thickened rangeland and was left untreated. In the BC plots, existing woody species (matured and saplings) were removed (i.e., cutting of all trees) at ground level using a handsaw. The other control techniques were tree thinning (T), which refers to the removal of some of the woody plant species from the population to a predetermined density; thinning and grazing (T + G), which refer to the grazing of a thinned plot; thinning and browsing (T + B), which refer to browsing of a thinned plot by browsers; thinning and fire (T + F), which refer to the burning of thinned plot; and thinning that combined burning and browsing (T + F + B) treatments. In the five treatment plots (i.e., T, T + G, T + B, T + F, and T + F + B), woody plant species (≥2 m) were thinned to the equivalents of 60% of the tree density of that of the control (CO) plot. Tree saplings (≥0.5 m < 2 m) were also removed manually from all plots except in the CO to ensure the existence of large tree structures particularly in the thinning plots (Smit et al., 1999).
The experimental plots were fenced to realize the implementation of activities specific to the individual treatments. Accordingly, the CO, BC, and T plots were excluded from grazing by all types of livestock. In the T + G plot, cattle were allowed to graze the area at the low-stocking rate to impact tree seedlings (Smit et al., 1999). During the practice, 10 heads of cattle ha−1 were allowed to graze for 7 days in the plot area. Similarly, in the browsing plots (T + B and T + F + B) range goats were allowed to browse at a high-stocking rate (Smit et al., 1999; Allen, 2000). During the browsing period, 30 heads of goat ha−1 were allowed to browse in the plots for 14 days to achieve a greater impact on woody seedlings. This stocking rate was not based on the objective assessment of the available browse but was intended to impact tree saplings as severely as possible. Burning plots (i.e., T + F and T + F + B) were allowed to build up the fuel load for two rainy periods (i.e., main and small rainy seasons in 2018) and were enclosed with a fire-line to ensure safety. Before burning, the herbaceous biomass was harvested using quadrats to determine the fuel load for burning. A total of five quadrats were harvested randomly from the individual plots along with replications. The fresh weight of samples was taken immediately and oven-dried for 24 h at a temperature of 105°C and fuel load estimated. The moisture content of the fuel load was determined by subtracting the fresh weight from dry weight × 100. A fuel load of 2,500 to 3,000 kg dry matter ha−1 was reported sufficient to ignite fire (Angassa and Oba, 2009; Kahumba, 2010). Moreover, grass should be dryer than 40% moisture content, and the air should also be dry and warm (Kahumba, 2010). The average fuel load in the treatment plots ranged from 2,754 in the T + F plot to 2,836 kg dry matter ha−1 in the T + F + B plot. The moisture content of herbaceous biomass in the plots was 17% (83% drier) and 21% (79% drier) in the T + F and T + F + B plots, respectively. Plot burning was conducted between 11:00 and 15:00 h local time. The air temperature recorded at the nearest meteorology station (Dambala Wachu located at 22 km southeast of Har Weyu) ranged from 27.7°C to 30.9°C. The direction of the wind was determined using textile material tied to the upper tree stem erected outside the area of the plots (Angassa and Oba, 2009). The wind direction was westward with low speed at the site.
2.2.3 Survey and Treatment Application
Pretreatment data collection was conducted in the middle of the small rainy season in October 2017, when tree species and previously established saplings were visible and easy to identify. The number of individual trees in the treatment plots was counted, and their species recorded. Trees with ≥2-m height were recorded as mature individuals, but plants ≥0.5-m but <2-m height were noted as saplings (Chauhan et al., 2008; Bharali et al., 2012). To determine the total area of coverage by the individual woody cover, we measured the canopy diameter (d) of 24 randomly selected trees within each treatment plot by spreading measuring tape either beneath or above the canopy as appropriate. The canopy diameter then gave a canopy radius (r) measurement (r = ½ d). We modeled the shape covered by the canopy as circular and calculated the area covered by each canopy as area = πr2 (Innocent and Munyati, 2014). Tree clearing and thinning practices were completed in February 2018 during the dry season. Cut stumps were treated with fire using dry woody branches as a fuel load, thus ensuring that the removed individuals were killed not to affect other plants. Grazing and browsing were applied three times between 2018 (April and October) and 2019 (April) after the onset of seasonal rains. Burning was carried out in February 2019 (i.e., the hot dry period) before the onset of the main rain normally expected in March. Posttreatment establishment of woody species was monitored at the end of the main rain in April 2019. The identification between species was done together with technical persons and herders.
2.2.4 Rainfall and Soil Moisture
Rainfall data were recorded at the Yabalo town, which is the nearest meteorological station. In addition, the soil moisture content was estimated by taking 200 g of soil in the seven treatment plots to a depth of 200 mm (Ward et al., 2013) using auger after a rainfall event in April and October 2018 and in April 2019. The moist weight of the soil sample was taken immediately. Two samples were prepared per plot and oven-dried for 24 h at 105°C and weighed again.
2.2.5 Analysis
Density is a count of the number of individuals of a species within the sample plot (Kent and Coker, 1992). The sum of individuals per species was calculated and analyzed in terms of species/woody/tree/sapling density ha−1 (Shankar, 2001; Tiwari et al., 2010). The total area covered by woody canopy within the plots is defined as the ratio of 1-ha area covered by woody canopies (Innocent and Munyati, 2014). The average monthly and annual rainfalls were calculated from the 2 years’ amounts. The moisture content of the soil was determined by subtracting the oven-dry weight from the moist weight and expressed as a percentage of the oven-dry weight of the soil (Dirksen, 1999). Two-way analysis of variance and Tukey (honestly significant difference) test method was performed for comparing mean values in the treatment plots. Posttreatment changes in sapling densities were calculated for each treatment and species to determine the change in the total populations (λ) as the proportion of established individuals plus mean change in sapling densities relative to pretreatment sapling density (Otterstrom et al., 2006). A χ2 test was used to analyze species’ response (i.e., saplings ha−1) to the bush control techniques. The statistical software R (R Development Core Team 2018) was used to analyze the data. The Pearson correlation was applied to test the association between changes in sapling densities and new recruitment in the individual treatments and tree species. Significant differences were considered at p < 0.05.
3 Results
3.1 Woody Density and Cover
Figure 2 provides tree densities and canopy covers within the various plots derived from the Supplementary Appendix Table B1. The average density of woody plants was almost similar in all treatments during the pretreatment period. However, posttreatment density and canopy cover differed significantly (p < 0.001) among the seven treatments (Figure 2). Tree density and cover were higher in the CO plot than in the other six plots. The lowest density and cover were recorded in the BC treatment to the point of no representation in the plot area. Thinning treatments reduced (p < 0.001) bush density and cover compared with the CO and BC treatments. Although there was no significant difference (p > 0.05) in the density and cover of woody plants among the five (T, T + G, T + B, T + F, and T + F + B) thinning treatments. The average density of the remained plants in the five thinning plots was 795 ± 27.37 trees ha−1 retaining a mean canopy cover of 31.0% ha−1. On average, thinning reduced woody density by 39.3% relative to the CO plot and conserved 40.4% of tree population ha−1 when compared with the BC plot. Similarly, thinning reduced 41.9% tree canopy cover compared with the control plot and sustained 31.0% canopy cover ha−1 in relation to the cleared plot. The trend suggests that tree thinning has the advantage of preserving woody individuals important for proper ecosystem functioning in the area. On the contrary, clearing appeared to be removing the important positive effects that woody plants have on the system, which might have negative consequences on composition and structure of savanna vegetation.
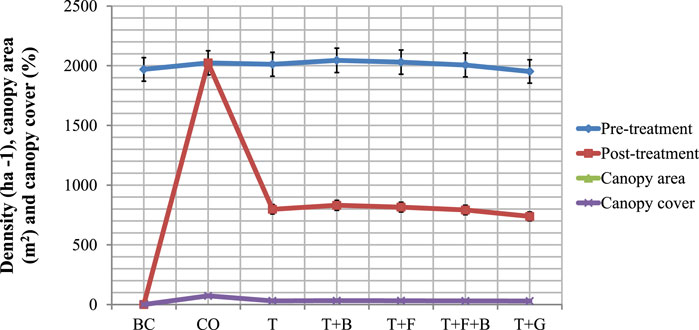
FIGURE 2. The pretreatment and posttreatment mean density, canopy area, and cover of woody plants (ha−1) in the different bush control treatment plots in Borana, southern Ethiopia. Vertical bars mean a 0.95 confidence interval.
3.2 Rainfall and Soil Moisture Status
According to the rainfall data collected, mean monthly rainfall ranged from 0.0 mm in January to 291.2 mm in October (Figure 3). The average monthly rainfall was significant (p < 0.05) with the peaks in October and April demonstrating the bimodal type of rainfall in the study area. Total annual rainfalls recorded were 960.3 and 1,349.9 mm in 2018 and 2019, respectively. The variation in annual amounts was significant (p < 0.05) and also greater (p < 0.05) than the long-term average of 643.6 mm (NMASE, 2018).
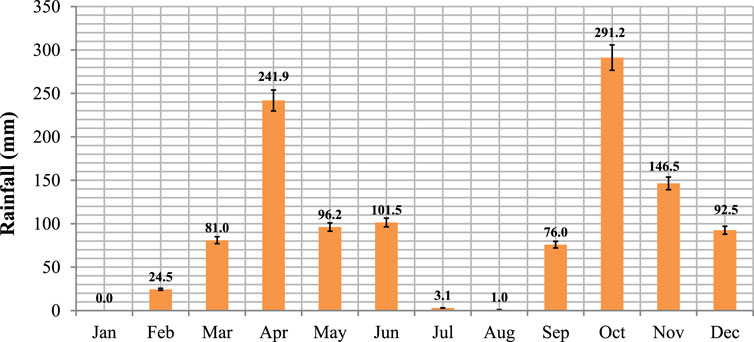
FIGURE 3. Average monthly rainfalls (2018–2019) in the study district at the Yabalo meteorological station. A 5% significant level was used to analyze the means (n = 12).
The soil moisture contents of the individual treatment plots during the rainfall events were presented (Figure 4). There was no significant (p > 0.05) difference in average soil moisture contents among the seven treatments. Over the period, mean soil moisture contents were 32.0%, 32.2%, 32.5%, 30.9%, 33.9%, 31.4%, and 33.0% in the CO, BC, T, T + G, T + B, T + F, and T + F + B plots, respectively, with the overall average of 32.3% in the experimental site. The frequent and higher rains received during the study years may improve the soil moisture contents within the various plots and support plants growth in a similar way.
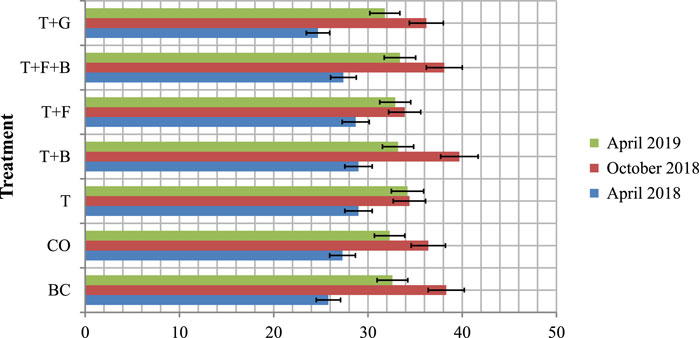
FIGURE 4. Soil moisture contents (%) in the various bush control plots during the main rainfall periods in Borana, southern Ethiopia. Vertical bars mean a 0.95 confidence interval.
3.3 Effects on Tree Sapling Recruitment
The pretreatment and posttreatment sapling densities of the individual woody species in the different plots are provided (Figure 5) as computed from the Supplementary Appendix Table C1. During the pretreatment period, cumulative sapling density was almost similar across the study treatments. However, significant (p < 0.05) difference was observed during the postcontrol period. In the control plot, sapling recruitment amounted to 109 trees ha−1, which was greater compared with the postthinning treatments (T + B, T + F, and T + F + B). In the BC treatment, total sapling density was superior (267 plants ha−1) to the other six treatments. For the T plot, sapling density was 175 plants ha−1, which was higher compared with the other postthinning treatments (T + G, T + F, T + B, and T + F + B). The T + G treatment induced cumulative recruitment of 146 plants ha−1, which was greater than the densities in the similarly designed thinning treatments (T + F, T + B, and T + F + B). The T + F treatment experienced low sapling recruitment of 82 plants ha−1, compared with CO, BC, T, and T + G treatments. The T + B treatment exhibited lower sapling regeneration of 63 plants ha−1 relative to other treatments, but the T + F + B treatment. The T + F + B treatment experienced the least sapling establishment (34 plants ha−1) compared with the other six treatments.
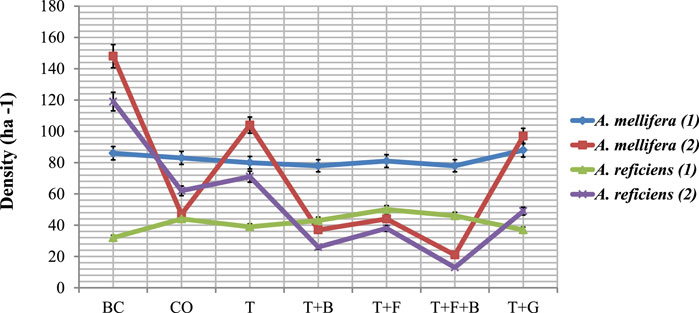
FIGURE 5. The pretreatment (1) and posttreatment (2) mean sapling densities ha−1 of the two woody species in the bush control treatment plots in Borana, southern Ethiopia. Vertical bars mean a 0.95 confidence interval.
Table 1 presents all the seven treatments and their posttreatment sapling recruitments. Among the seven bush control treatments, three of them (BC, T, T + G) showed positive changes in sapling population, whereas the remaining four (CO, T + B, T + F, and T + F + B) treatments presented negative changes. A change in sapling population was higher in the cleared plot than in the other plots, whereas it was recorded lowest in the T + F + B treatment.
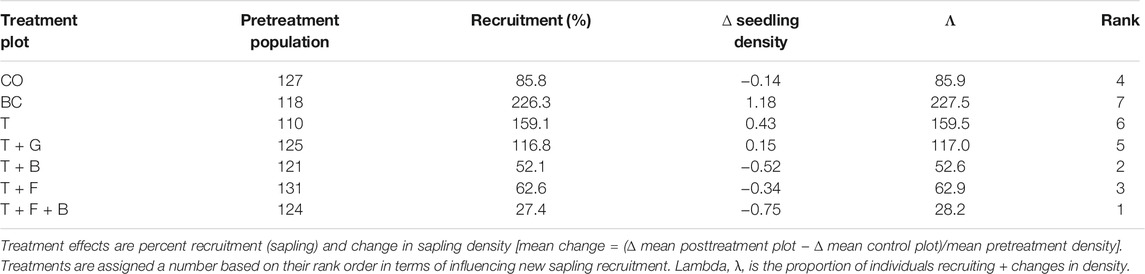
TABLE 1. Effects of the seven bush control treatments on woody sapling recruitment and change in sapling densities in Borana area, southern Ethiopia.
Table 2 also presents the correlation between changes in the sapling population and postrecruitment within the various treatment plots. There was a significant positive correlation (p < 0.5) between changes in sapling density and new sapling recruitment in the CO, BC, T, and T + G treatments. The correlation was highly significant and strong (p < 0.001) in the bush cleared treatment, resulting in the highest recruitment proportion (λ), demonstrating that clearing caused new invasion through aiding sapling establishment. The change in sapling density and new recruitment was highly significantly (p < 0.01) correlated in the thinning plot prompting tree recruitment relative to the more intensified thinning treatments. However, the thinning alone treatment reduced (p < 0.05) recruitment compared with the BC treatment, displaying the significant potential role of thinning in stabilizing the problem of increasing tree density. Similarly, the association between changes in sapling density and recruitment in the thinning and grazing plot was highly significant (p < 0.01), showing that thinning, mainly grazing, enhanced the recruitment of tree saplings compared with the similarly designed other postthinning treatments (i.e., T + B, T + B, and T + F + B). The trend in the thinning and grazing treatment suggests that immediate grazing may encourage the establishment of woody saplings. The decline in sapling establishment was minor (14.1%) in the control plot; hence, significant correlation existed between the change in sapling density and total regeneration, which promoted new recruitment evidence that tree density continues to increase in savanna through sapling recruitment. Less significant (p < 0.05) association occurred between the change in density and sapling recruitment and has resulted in declined tree regeneration in the thinning and fire treatment. However, the thinning and fire treatment was less effective in preventing re-encroachment compared with T + B and T + F + B treatments, suggesting that postcontrol fire may initiate germination in woody plants from the soil seed-bank seedlings. In the thinning and browsing treatment, the correlation between the change in plants density and regeneration was less significant (p < 0.05), leading to reduced plant recruitment, indicating the greater effectiveness of this combination in controlling sapling recruitment in woody plants. The relationship between the change in density and sapling regeneration was weak and statistically insignificant in the T + F + B treatment and produced the lowest tree sapling recruitment, evidencing that the combination of thinning, burning, and browsing methods was prominently efficient in suppressing bush encroachment in the study savanna.

TABLE 2. Correlation analyses (n = 10) of relations between changes in woody sapling population and new recruitment for the bush control techniques in Borana, Ethiopia.
3.4 Responses of Woody Species to Treatments
Supplementary Appendix Table C1 provides sapling density ha−1 of the individual tree species during the pretreatment and posttreatment periods. During the postperiod, the total recruitment through saplings disclosed significant (p < 0.05) changes between A. mellifera and A. reficiens across the seven treatments (Table 3). In the control plot, A. reficiens experienced greater (p < 0.01) sapling recruitment, indicating that the performance of species recruitment might vary under the established bush density environment. In BC and T treatments, sapling recruitment was highly increased (p < 0.001) in A. mellifera, accounting for the greater regeneration following the removal of existing tree species. Again, A. mellifera accounted for the greater proportion (p < 0.01) of the recruitment in the T + G, T + F, T + B, and T + F + B treatments.

TABLE 3. χ2 Tests for the two woody species in terms of cumulative saplings recruitment in bush control treatments in Borana, Ethiopia.
The two woody species showed positive changes in sapling density in BC, T, and T + G treatments, with the values higher for A. reficiens except in the T + G, which is indifferent to the two species (Table 4). The changes were negative and positive for A. reficiens and A. mellifera, respectively, in the control plot. On the other hand, tree species similarly experienced negative changes in sapling populations in the other three (T + F, T + B, and T + F + B) treatments (Table 4). The negative values are higher and lower for A. mellifera and A. reficiens, respectively, across those plots. Overall, a change in sapling density was positive (greater) in A. reficiens, whereas it was negative (lower) in A. mellifera (Table 7). There was a strong significant correlation (r = 0.86, p = 0.01) between change in total sapling population and new recruitment in A. reficiens, indicating the greater recruitment of A. reficiens through sapling establishment, further suggesting the resistance of A. reficiens to the applied treatments relative to A. mellifera. On the other hand, there was a weak but significant correlation (r = 0.79, p = 0.05) between changes in density and new sapling recruitment in A. mellifera trees. The lower negative value in A. mellifera might be an indication of the susceptibility of this species to bush control techniques, particularly to the postthinning (T + F, T + B and T + F + B) treatments.
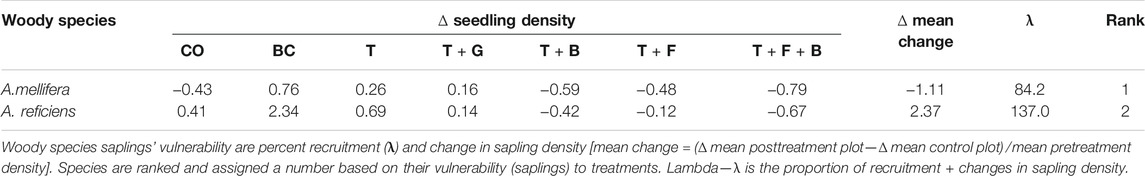
TABLE 4. Changes in sapling densities of the two woody species in the bush control plots and their percent recruitment in Borana, Ethiopia.
4 Discussion
4.1 Effects of Bush Control Techniques on Woody Density, Cover, and Recruitment
The success of woody plants is related to the competition for soil moisture and nutrient between the woody and herbaceous layers in arid savannas, which will ultimately determine the success of woody and grass species establishment (Ward, 2005; Salazar et al., 2012). Water availability alters seedling regeneration (Van der Waal et al., 2011; Sardans and Pen˜uelas, 2013). The woody vegetation layer mostly has a much better-adapted root system occurring in a wide range of soil layers, ensuring their survival at the expense of the herbaceous layer (Joubert et al., 2013). According to the current study, the high woody density and cover in the control plot might be related to the competition between plants for soil moisture and light. Higgins et al. (2000) stated that rainfall and moisture are assumed to enhance bush encroachment in savanna system. With adequate rainfall in savannas, trees would dominate the landscape, provided that fire and browsing were suppressed. In principle, this unstable system may tend toward closed woodland, depending on rainfall disturbance regimes influencing subcanopy species regeneration. In addition, it has been reported that, in dense vegetation stands, self-thinning will normally take place as competition for soil moisture and nutrient increases (Wiegand et al., 2006). During the process, the different plant species that are adapted to specific microenvironment conditions will colonize the gaps in the canopy and the various subcanopies (Reynolds and Ford 2005; Berhane et al., 2015). Amelioration of solar radiation and stressful environments under canopy shade is considered favorable for tree seedling establishment (Rolhauser et al., 2011; Salazar et al., 2012).
The evidence recorded about the decline (14.1%) in sapling recruitment in the control plot suggests that high woody density and cover suppress the establishment of new saplings. This finding is in line with the findings of Beckage et al. (2000) and Nilsen et al. (2001), who all reported that canopied vegetation delays and modifies woody regeneration through competing for soil resources and light. As canopy cover increases, seedling establishment decreases, and the vegetation physiognomy will progress toward larger, more widely spaced individuals with closed canopies (Wiegand et al., 2006). Hence, bush density remains increasing in savannas through seedling/sapling establishment under stressful environmental conditions, leading to a stable state, if left aside without considering any practical management technique.
BC practice disrupts the growth of the above-ground woody biomass without necessarily reducing tree density ha−1 in rangelands. Clearing of matured individuals and keystone woody species, responsible for the regulation of fundamental ecosystem processes, could have severe effects on ecosystem stability, causing an imbalance in the woody–grass ratio (Solomon and Mblambo, 2010; Berhane et al., 2015). An increase in the rate of sapling recruitment under the current study is consistent with the findings of Mathew and Brian (2006) that BC accelerated understory recruitment of tree species that regenerated from seed and germinated from a seedling bank without follow-up treatment. Ludwig et al. (2004) also indicated in their study report that wide gaps in the savanna are created by the removal of woody vegetation affecting the availability of light, nutrients, and soil moisture, which can potentially further alter species composition. If the herbaceous seed bank is depleted, gaps created by bush-clearing practices could lead to the establishment of new woody individuals (Smit, 2004). Therefore, tree-removing practice would worsen postcontrol encroachment conditions by aiding seedling establishments in woody plants, thus reducing the effective time span of restoration measures in rangelands.
Thinning negatively influenced bush density and sapling recruitment compared with the clearing practice. This finding is according to Smit (2004), who reported that, with thinning techniques implemented, the remaining trees will benefit from the reduced competition, which will lead to an increased growth combined with a suppressing effect, eventually outcompeting the newly established seedlings. Tree thinning contributes to structural changes of the woody layer, which in turn influence the distribution of resources, for example, soil moisture and nutrients, and the microclimate (Ludwig et al., 2004). These changes have cascading effects on biodiversity, with some organisms responding directly to changes in woody structure, whereas others are influenced by resource availability and microclimate (Williams et al., 2002; Ripple and Beschta, 2004; Berhane et al., 2015). According to the current findings, the relative increase in woody sapling recruitment following thinning suggests that thinning alone would not be adequate to control bush encroachment unless followed by postthinning treatments to prevent new plant invasion.
In savanna area, grazing alters vegetation composition through selectively feeding on grasses (Wiseman et al., 2004; Dalle et al., 2006a; Riginos, 2009). Leaves defoliation by grazers decreases grass vigor (Ferraro and Oesterheld, 2002) and increases the competitive ability of woody plants. This corresponds to the current negative effect of cattle on grass species following thinning treatment, because grazers prefer consuming grasses to the freshly growing tree seedlings. According to Cramer et al. (2007) and Riginos (2009), a decrease in grass vigor increases the relative growth rates of woody plants. Trampling can also destroy the roots or rhizomes of grasses and therefore reduce their vigor and create gaps for woody plant seed germination (Danell et al., 2006). By proving niche gaps for woody plants, both grazing and trampling may have positive effects on woody plant germination (Savadogo et al., 2007). In addition, grazing increases water availability for woody plants as a result of reduced uptake by grasses (Gordijn et al., 2012). It is therefore understood by the current study that grazing has a positive relationship with woody plant recruitment in a grazing savanna. As a result, grazing cannot be used as an effective postmanagement to regulate problem of increasing tree density in savanna rangelands. Immediate grazing without adequate resting (i.e., allowing vigorous grass growth) of thinned areas may prematurely remove grasses, reduce grass competition, and favor the establishment of woody plants.
According to the current study, postthinning fire reduced woody recruitment by 36.1% mainly attributed to the abundance and spatial connectivity of combustible herbaceous biomass, which supported intense fire and resulted in reduced occurrence of tree saplings. This finding agrees with Wiegand et al. (2006) and Werner (2012) all reported that intense fire reduces woody density by top-killing seedlings and saplings, influencing the recruitment to adults in tree species. Gordijn et al. (2012) also showed that fires are capable of reducing tree density through sapling mortality in savannas. This is because seedlings of many woody plant species are vulnerable to intense fires (Werner, 2012). Fire also noted to promote seedling establishment following tree thinning. The current evidence about the increase (62.9%) in sapling recruitment during the postfire period might be an indication that fire initiated seedling germination in woody plants. Past studies (Angassa and Oba, 2009; Werner, 2012) have reported that fire triggers germination from the seed bank by breaking seed dormancy. Kahumba (2010) also indicated that fire stimulates seedling establishments in woody plants, and therefore, postburning aftercare of savanna is critical. In general, fire frequency and intensity determine the recruitment of seedlings to matured-large tree classes (Govender et al., 2006). Therefore, it is suggested from the study results that tree thinning together with frequent and intense fires would probably be efficient to slow down the problem of bush encroachment in rangelands.
The current decline (47.4%) in the recruitment of tree saplings in the thinning and browsing treatment corresponds with Smit et al. (1999) and De Klerk (2004) all reported that goats can be introduced successfully to utilize and control regrowth as a follow-up to other methods such as mowing and can be effective in preventing the establishment of tree seedlings. This is possibly attributed to the effect of browse preference of freshly establishing seedlings by goats. Browsing is then the most important follow-up tool to control bush density/encroachment because goats as browsers effectively impacted the recruitment of encroaching woody plant species. Sankaran et al. (2008) reported higher woody cover in sites without elephants compared with those with high elephant biomass. And the exclusion of the browsing ungulates is one of the drivers of bush encroachment in Borana rangelands of southern Ethiopia (Angassa and Oba, 2010). Thus, browsing herbivore in savannas is important in complementing other techniques to maintain the tree–grass balance (Van der Waal et al., 2011). The decline in woody recruitment as a result of browsing suggests the introduction of browsing animals (goats and camels) as an effective management practice in controlling the thickening of woody plants in the Borana savanna.
Thinning, fire, and browsing combinations prominently reduced (71.8%) recruitments in woody plants. The lowest sapling population in this treatments combination was largely due to the synergetic effects of fire and browsing following a thinning practice. The results further support the greater efficacy of fire and browsing as a postthinning management measure to suppress tree regenerations. The negative effects of fire and herbivory on woody plants have been widely reported elsewhere in savannas (Angassa and Oba, 2009; Midgley et al., 2010; Vadigi and Ward, 2014). Allred et al. (2011), suggesting that the interaction between methods, for example, fires and herbivores, is ecologically more significant than the independent effect and that the combination should be viewed as a single disturbance responsible for markedly different effects on vegetation compared with a single practice alone. In African savanna, the interactive effects of browsers and fire have led to a decline of some woodlands and their subsequent replacement by grasslands or open savanna ecosystems (Bond, 2008). Repeated herbivory exposes woody plants to the fire by preventing their escape from the fire-prone lower height strata (Sankaran et al., 2008). Tree thinning that combined fire and herbivore is therefore ranked as the best alternative option in limiting bush density and cover in the semiarid Borana region that was, without this management combination, transformed to a bush-dominated savanna land.
4.2 Responses of Woody Species to Bush Control Techniques
In the dense control plot, tree sapling population ha−1 varied between species, indicating that the rate and speed of woody species regeneration might vary in the savanna system. Grundy et al. (1994) discovered that certain species respond differently to competition from neighboring woody species, where some species use the canopy of other species as a “safe site” for the establishment; others tend to appear only in the opening between canopy environments.
In the current study, woody species were cleared/thinned with imposed postthinning treatments, which differently promoted sapling regenerations in woody species. Past studies (Bond et al., 2001; Vesk, 2006; Angassa and Oba, 2009) indicated that woody species respond differently to different control treatments. Woody species uses alternative strategies in response to control methods, for example, by investing in seedling recruitment (Smit, 2004; Angassa and Oba, 2009). The proportion of the sapling population in the individual species is related to the ability of saplings to withstand the adverse effects of treatments by aggressively competing for available soil resources to replace the removed trees. On evaluating at the individual species level, A. mellifera was the least regenerating species, justifying the vulnerability of this particular species to the combinations of bush control treatments. A. reficiens was found to be the most recruiting one by sapling, indicating the relative resistance to the control techniques, suggesting that the most intensive and frequent treatment is recommendable for the aggressively regenerating tree species.
5 Conclusion
The study was carried out in the semi-arid Borana rangelands of southern Ethiopia. The objective of the study was to investigate the suppressive effects of bush encroachment control techniques on the density, cover, and recruitment of invasive acacia species for achieving successful control methods in rangelands ecosystem. At the study site, woody density and cover were significantly higher in the control than in the BC and thinning treatments. In the BC treatment, large woody individuals were almost absent, however, with greater sapling regeneration compared with the control and postthinning treatments. The evidence suggests that the control and BC methods promoted bush encroachment and were not effective in alleviating the problem of increasing woody density in grazing lands. On the contrary, tree density and recruitment were declined in the postthinning treatments when compared with the control and clearing practices. The current study concludes that thinning postthinning techniques were effective in terms of reducing tree density and also in the killing of saplings. Responses of individual woody species to the thinning and postthinning measures have important implications for bush encroachment management and conservation of biodiversity in savanna systems.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Author Contributions
MH designed the study, conducted the fieldwork, analyzed the data, interpreted the findings and wrote the manuscript. YW and AG designed the study and wrote the manuscript. XX, YY, and ZN critically commented the draft. All authors fully participated in processing the manuscript. All authors have read and agreed to the final version of the manuscript.
Funding
This study received financial support from Integration and application of appropriate technologies for desertification control in Africa (Grant No. SAJC 202108), African Great Green Wall adaptation technical cooperation research and demonstration (Grant No. 2018YFE0106000) and this research was proposed by the “Tianchi doctor program” of Xinjiang Uygur Autonomous Region in 2020.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors wish to thank all funding agencies for providing financial support for the success of this particular study. Authors are also very grateful to the Har Weyu community participating in this study by providing access to their lands and sharing their knowledge.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2021.777146/full#supplementary-material
References
Allen, G. R. (2000). Restoration of Grazing Lands in the Liben Woreda, Borana. Utah State University. American Grasslands. J. Environ. Mgt. 90, 2931–2942.
Allred, B. W., Fuhlendorf, S. D., Engle, D. M., and Elmore, R. D. (2011). Ungulate Preference for Burned Patches Reveals Strength of Fire-Grazing Interaction. Ecol. Evol. 1, 132–144. doi:10.1002/ece3.12
Angassa, A., and Oba, G. (2009). Bush Encroachment Control Demonstrations in Southern Ethiopia: 1. Woody Species Survival Strategies with Implications for Herder Land Management. Afr. J. Ecol. 47, 63–76. doi:10.1111/j.1365-2028.2007.00919.x
Angassa, A., and Oba, G. (2010). Effects of Grazing Pressure, Age of Enclosures and Seasonality on bush Cover Dynamics and Vegetation Composition in Southern Ethiopia. J. Arid Environments 74, 111–120. doi:10.1016/j.jaridenv.2009.07.015
Angassa, A., and Oba, G. (2008). Effects of Management and Time on Mechanisms of bush Encroachment in Southern Ethiopia. Afr. J Ecol 46, 186–196. doi:10.1111/j.1365-2028.2007.00832.x
Angassa, A., and Oba, G. (2007). Relating Long-Term Rainfall Variability to Cattle Population Dynamics in Communal Rangelands and a Government Ranch in Southern Ethiopia. Agric. Syst. 94, 715–725. doi:10.1016/j.agsy.2007.02.012
Angassa, A. (2005). The Ecological Impact of bush Encroachment on the Yield of Grasses in Borana Rangeland Ecosystem. Afr. J. Ecol. 43, 14–20. doi:10.1111/j.1365-2028.2005.00429.x
Angassa, A. (2002). The Effect of Clearing Bushes and Shrubs on Range Condition in Borana, Ethiopia. Trop. Grass 36, 69–76.
Archer, S. R., Andersen, E. M., Predick, K. I., Schwinning, S., Steidl, R. J., and Woods, S. R. (2017). “Woody Plant Encroachment: Causes and Consequences,” in “Woody Plant Encroachment: Causes and Consequences,” in Rangeland Systems. Editor Briske D. D. (Springer), 25–84. doi:10.1007/978-3-319-46709-2_2
Augustine, D. J., and McNaughton, S. J. (2004). Regulation of Shrub Dynamics by Native Browsing Ungulates on East African Rangeland. J. Appl. Ecol. 41, 45–58. doi:10.1111/j.1365-2664.2004.00864.x10.1111/j.1365-2664.2004.00864.x
Beckage, B., Clark, J. S., Clinton, B. D., and Haines, B. L. (2000). A Long-Term Study of Tree Seedling Recruitment in Southern Appalachian Forests: the Effects of Canopy Gaps and Shrub Understories. Can. J. For. Res. 30, 1617–1631. doi:10.1139/x00-075
Belayneh, A., and Tessema, Z. K. (2017). Mechanisms of bush Encroachment and its Inter-connection with Rangeland Degradation in Semi-arid African Ecosystems: a Review. J. Arid Land 9 (2), 299–312. doi:10.1007/s40333-016-0023-x
Berhane, A., Totland, Ø., Haile, M., and Moe, S. R. (2015). Intense Use of Woody Plants in a Semiarid Environment of Northern Ethiopia: Effects on Species Composition, Richness and Diversity. J. Arid Environments 114, 14–21. doi:10.1016/j.jaridenv.2014.11.001
Bharali, S., Paul, A., Latif Khan, M., and Bihari Singha, L. (2012). Impact of Altitude on Population Structure and Regeneration Status of Two Rhododendron Species in a Temperate Broad Leaved Forest of Arunachal Pradesh, India. Ije 2, 19–27. doi:10.5923/j.ije.20120201.04
Bikila, N., Bedasa, E., Samuel, T., Barecha, B., Jaldesa, D., and Nizam, H. (2014). Control of bush Encroachment in Borana Zone of Southern Ethiopia: Effects of Different Control Techniques on Rangeland Vegetation and Tick Populations. Pastoralism 4, 18.
Bond, W. J., Smythe, K. A., and Balfour, D. A. (2001). Acacia Species Turnover in Space and Time in an African savanna. J. Biogeogr. 28, 117–128. doi:10.1046/j.1365-2699.2001.00506.x
Bond, W. J. (2008). What Limits Trees in C4 Grasslands and Savannas? Annu. Rev. Ecol. Evol. Syst. 39, 641–659. doi:10.1146/annurev.ecolsys.39.110707.173411
Chauhan, D. S., Dhanai, C. S., Singh, B., Chauhan, S., Todaria, N. P., and Khalid, M. A. (2008). Regeneration and Tree Diversity in Natural and Planted Forests in a Terai- Bhabhar Forest in Katarniaghat Wildlife Sanctuary, India. Trop. Ecol 49, 53–67.
Coppock, D. L. (1994). The Borana Plateau of Southern Ethiopia: Synthesis of Pastoral Research Development and Changes, 1980-90. Addis Ababa: ILCA (International Livestock Centre for Africa, 374.
Cramer, M. D., Chimphango, S. B. M., van Cauter, A., Waldram, M. S., and Bond, W. J. (2007). Grass Competition Induces N2-Fixation in Some Species of African Acacias. J. Ecol. 95, 1123–1133. doi:10.1111/j.1365-2745.2007.01285.x
Cramer, M. D., van Cauter, A., and Bond, W. J. (2010). Growth of N2-Fixing African savanna Acacia Species Is Constrained by Below-Ground Competition with Grass. J. Ecol. 98, 156–167. doi:10.1111/j.1365-2745.2009.01594.x
Dalle, G., Maass, B. L., and Isselstein, J. (2006a). Encroachment of Woody Plants and its Impact on Pastoral Livestock Production in the Borana Lowlands, Southern Oromia, Ethiopia. Afr. J Ecol 44, 237–246. doi:10.1111/j.1365-2028.2006.00638.x
Danell, K., Duncan, P., Bergström, R., and Pastor, J. (Editors) (2006). Large Herbivore Ecology, Ecosystem Dynamics and Conservation (Cambridge, UK: Cambridge University Press).
De Klerk, J. N. (2004). Report on Phase 1 of the Bush Encroachment Research, Monitoring and Management Project. Windhoek, Namibia: Ministry of environment and tourism.Bush Encroachment in Namibia
Desta, S., and Coppock, D. L. (2002). Cattle Population Dynamics in the Southern Ethiopian Rangelands, 1980-97. J. Range Manage. 55, 439–451. doi:10.2307/4003221
Desta, S., and Coppock, D. L. (2004). Pastoralism under Pressure: Tracking System Change in Southern Ethiopia. Hum. Ecol. 32, 465–486. doi:10.1023/B:HUEC.0000043516.56037.6b
Devine, A. P., Stott, I., McDonald, R. A., and Maclean, I. M. D. (2015). Woody Cover in Wet and Dry African Savannas after Six Decades of Experimental Fires. J. Ecol. 103, 473–478. doi:10.1111/1365-2745.12367
Ferraro, D. O., and Oesterheld, M. (2002). Effect of Defoliation on Grass Growth. A Quantitative Review. Oikos 98, 125–133. doi:10.1034/j.1600-0706.2002.980113.x
Forrest, B. W., Coppock, D. L., Bailey, D., and Ward, R. A. (2015). Economic Analysis of Land and Livestock Management Interventions to Improve Resilience of a Pastoral Community in Southern Ethiopia. J. Afr. Econ. 25, 233–266. doi:10.1093/jae/ejv021
Gordijn, P. J., Rice, E., and Ward, D. (2012). The Effects of Fire on Woody Plant Encroachment Are Exacerbated by Succession of Trees of Decreased Palatability. Perspect. Plant Ecol. Evol. Syst. 14, 411–422. doi:10.1016/j.ppees.2012.09.005
Govender, N., Trollope, W. S. W., and van Wilgen, B. W. (2006). The Effect of Fire Season, Fire Frequency, Rainfall and Management on Fire Intensity in savanna Vegetation in South Africa. J. Appl. Ecol. 43, 748–758. doi:10.1111/j.1365-2664.2006.01184.x
Grellier, S., Barot, S., Janeau, J.-L., and Ward, D. (2012). Grass Competition Is More Important Than Seed Ingestion by Livestock for Acacia Recruitment in South Africa. Plant Ecol. 213, 899–908. doi:10.1007/s11258-012-0051-3
Grundy, I. M., Campbell, B. M., and Frost, P. G. (1994). Spatial Pattern, Regeneration and Growth Rates of Brachystegia Spiciformis and Julbenardia Globiflora. Vegetation 115, 101–107. Available at: https://www.jstor.org/stable/20046510.
Hagenah, N., Munkert, H., Gerhardt, K., and Olff, H. (2009). Interacting Effects of Grass Height and Herbivores on the Establishment of an Encroaching savanna Shrub. Plant Ecol. 201, 553–566. doi:10.1007/s11258-008-9466-2
Higgins, S. I., Bond, W. J., February, E. C., Bronn, A., Euston-Brown, D. I. W., and Enslin, B. (2007). Effects of Four Decades of Fire Manipulation on Woody Vegetation Structure in savanna. Ecology 88, 1119–1125. doi:10.1890/06-1664
Higgins, S. I., Bond, W. J., and Trollope, W. S. W. (2000). Fire, Resprouting and Variability: a Recipe for Grass-Tree Coexistence in savanna. J. Ecol. 88, 213–229. doi:10.1046/j.1365-2745.2000.00435.x
Holmgren, M., Scheffer, M., and Huston, M. A. (1997). The Interplay of Facilitation and Competition in Plant Communities. Ecology 78, 1966–1975. doi:10.1890/0012-9658(1997)078[1966:tiofac]2.0.co;2
Innocent, N. S., and Munyati, C. (2014). Quantifying Savanna Woody Cover in the Field and on Historical Imagery: A Methodological Analysis. S. Afr. J. Geomatics 3, 2. doi:10.4314/sajg.v3i2.1
Joubert, D. F., Smit, G. N., and Hoffman, M. T. (2013). The Influence of Rainfall, Competition and Predation on Seed Production, Germination and Establishment of an Encroaching Acacia in an Arid Namibian savanna. J. Arid Environments 91, 7–13. doi:10.1016/j.jaridenv.2012.11.001
Kahumba, A. (2010). Masters of Science Thesis. Namibia: University of Namibia. Available at: http://repository.unam.edu.na/handle/11070/507 (Accessed March 23, 2021).Comparison of the Rehabilitative Effects of Mechanical and Chemical Methods of bush Control on Degraded highland and savanna Rangelands in Namibia
Kent, M., and Coker, R. (1992). Vegetation Description and Analysis: A Practical Approach. New York, NY: John Wiley & Sons, 363.
Kraaij, T., and Ward, D. (2006). Effects of Rain, Nitrogen, Fire and Grazing on Tree Recruitment and Early Survival in bush-encroached savanna, South Africa. Plant Ecol. 186, 235–246. doi:10.1007/s11258-006-9125-4
Ludwig, F., de Kroon, H., Berendse, F., and Prins, H. H. T. (2004). The Influence of savanna Trees on Nutrient, Water and Light Availability and the Understorey Vegetation. Plant Ecol. 170, 93–105. doi:10.1023/b:vege.0000019023.29636.92
Manning, A. D., Fischer, J., and Lindenmayer, D. B. (2006). Scattered Trees Are keystone Structures - Implications for Conservation. Biol. Conservation 132, 311–321. doi:10.1016/j.biocon.2006.04.023
Masters, R. A., and Sheley, R. L. (2001). Principles and Practices for Managing Rangeland Invasive Plants. J. Range Manage. 54, 502–517. doi:10.2307/4003579
Mathew, A., and Brian, C., M. (2006). Effects of Prescribed Fire and Thinning on Tree Recruitment Patterns in central Hardwood Forests. For. Ecol Mgt 226, 88–103.
Menaut, J. C. (1983).The Vegetation of African Savannas. Ecosystems Of the World 13: Tropical Savannas (ed. by Bourlière F.), pp.109–145. Elsevier Scientific Publishing Company, Amsterdam.
Midgley, J. J., Lawes, M. J., and Chamaillé-Jammes, S. (2010). Savanna Woody Plant Dynamics: the Role of Fire and Herbivory, Separately and Synergistically. Aust. J. Bot. 58, 1–11. doi:10.1071/bt09034
Mueller, R. C., Scudder, C. M., Porter, M. E., Talbot Trotter, R., Gehring, C. A., and Whitham, T. G. (2005). Differential Tree Mortality in Response to Severe Drought: Evidence for Long-Term Vegetation Shifts. J. Ecol. 93, 1085–1093. doi:10.1111/j.1365-2745.2005.01042.x
Mworia, J. K., Kinyamario, J. I., and Githaiga, J. M. (2008a). Influence of Cultivation, Settlements and Water Sources on Wildlife Distribution and Habitat Selection in South-East Kajiado, Kenya. Envir. Conserv. 35, 117–124. doi:10.1017/s0376892908004670
Nilsen, E. T., Clinton, B. D., Lei, T. T., Miller, O. K., Semones, S. W., and Walker, J. F. (2001). Does Rhododendron Maximum L. (Ericaceae) Reduce the Availability of Resources above and Belowground for Canopy Tree Seedlings? The Am. Midland Naturalist 145, 325–343. doi:10.1674/0003-0031(2001)145[0325:drmler]2.0.co;2
Oba, G., Post, E., Syvertsen, P. O., and Stenseth, N. C. (2000a). Bush Cover and Range Condition Assessments in Relation to Landscape and Grazing in Southern Ethiopia. Landscape Ecol. 15, 535–546. doi:10.1023/a:1008106625096
Oba, G., Stenseth, N. C., and Lusigi, W. J. (2000b). New Perspectives on Sustainable Grazing Management in Arid Zones of Sub-saharan Africa. Bioscience 50, 35–51. doi:10.1641/0006-3568(2000)050[0035:nposgm]2.3.co;2
Otterstrom, S. M., Schwartz, M. W., and Velazquez-Rocha, I. (2006). Responses to Fire in Selected Tropical Dry Forest Trees1. Biotropica 38, 592–598. doi:10.1111/j.1744-7429.2006.00188.x
R Development Core Team (2018). Revisited - Insights from an Examination of Assumptions and Mechanisms Invoked in Existing Models. A Language and Environment for Statistical Computing. Ecol. Lett., 7. Vienna, Austria: Foundation for Statistical Computing, 480–490.
Reynolds, J. H., and Ford, E. D. (2005). Improving Competition Representation in Theoretical Models of Self-Thinning: a Critical Review. J. Ecol. 93, 362–372. doi:10.1111/j.1365-2745.2005.00976.x
Riginos, C. (2009). Grass Competition Suppresses savanna Tree Growth across Multiple Demographic Stages. Ecology 90, 335–340. doi:10.1890/08-0462.1
Riginos, C., and Young, T. P. (2007). Positive and Negative Effects of Grass, Cattle, and Wild Herbivores on Acacia Saplings in an East African savanna. Oecologia 153, 985–995. doi:10.1007/s00442-007-0799-7
Ripple, W. J., and Beschta, R. L. (2004). Wolves and the Ecology of Fear: Can Predation Risk Structure Ecosystems? BioScience 54, 755–766. doi:10.1641/0006-3568(2004)054[0755:wateof]2.0.co;2
Rolhauser, A. G., Chaneton, E. J., and Batista, W. B. (2011). Influence of Conspecific and Heterospecific Adults on Riparian Tree Species Establishment during Encroachment of a Humid palm savanna. Oecologia 167, 141–148. doi:10.1007/s00442-011-1957-5
Roques, K. G., O'Connor, T. G., and Watkinson, A. R. (2001). Dynamics of Shrub Encroachment in an African savanna: Relative Influences of Fire, Herbivory, Rainfall and Density Dependence. J. Appl. Ecol. 38, 268–280. doi:10.1046/j.1365-2664.2001.00567.x
Salazar, A., Goldstein, G., Franco, A. C., and Miralles-Wilhelm, F. (2012). Differential Seedling Establishment of Woody Plants along a Tree Density Gradient in Neotropical Savannas. J. Ecol. 100, 1411–1421. doi:10.1111/j.1365-2745.2012.02028.x
Sankaran, M., Hanan, N. P., Scholes, R. J., Ratnam, J., Augustine, D. J., and Cade, B. S. (2005). Determinants of Woody Cover in African Savannas. Nature 438, 846–849. doi:10.1038/nature04070
Sankaran, M., Ratnam, J., and Hanan, N. (2008). Woody Cover in African Savannas: the Role of Resources, Fire and Herbivory. Glob. Ecol Biogeogr. 17, 236–245. doi:10.1111/j.1466-8238.2007.00360.x
Sardans, J., and Peñuelas, J. (2013). Plant-soil Interactions in Mediterranean forest and Shrublands: Impacts of Climatic Change. Plant Soil 365, 1–33. doi:10.1007/s11104-013-1591-6
Savadogo, P., Sawadogo, L., and Tiveau, D. (2007). Effects of Grazing Intensity and Prescribed Fire on Soil Physical and Hydrological Properties and Pasture Yield in the savanna Woodlands of Burkina Faso. Agric. Ecosyst. Environ. 118, 80–92. doi:10.1016/j.agee.2006.05.002
Scholes, R. J., and Archer, S. R. (1997). Tree-grass Interactions in Savannas. Annu. Rev. Ecol. Syst. 28, 517–544. doi:10.1146/annurev.ecolsys.28.1.517
Scholes, R. J., Dowty, P. R., Caylor, K., Parsons, D. A. B., Frost, P. G. H., and Shugart, H. H. (2002). Trends in savanna Structure and Composition along an Aridity Gradient in the Kalahari. J. Vegetation Sci. 13, 419–428. doi:10.1111/j.1654-1103.2002.tb02066.x
Seymour, C. L. (2008). Grass, Rainfall and Herbivores as Determinants of Acacia Erioloba (Meyer) Recruitment in an African savanna. Plant Ecol. 197, 131–138. doi:10.1007/s11258-007-9366-x
Shankar, U. (2001). A Case of High Tree Diversity Ina Sal (Shorea Robusta ) Dominated Lowland Forest of Eastern Himalaya: Floristic Composition, Regeneration and Conservation. Curr. Sci 81, 776–786.
Smit, G. N. (2004). An Approach to Tree Thinning to Structure Southern African Savannas for Long-Term Restoration from bush Encroachment. J. Environ. Manage. 71, 179–191. doi:10.1016/j.jenvman.2004.02.005
Smit, G. N., Ritcher, C. G. F., and Aucamp, A. J. (1999). “Bush Encroachment: an Approach to Understanding and Managing the Problem,” in Veld Management in South Africa Chapter 10. Editor Tainton N. M. (Pietermaritzburg, South Africa: University of Natal Press).
Smit, G. N. (2005). Tree Thinning as an Option to Increase Herbaceous Yield of an Encroached Semi-arid savanna in South Africa. BMC Ecol. 5, 4. doi:10.1186/1472-6785-5-4
Smit, N. (2014). BECVOL 3: an Expansion of the Aboveground Biomass Quantification Model for Trees and Shrubs to Include the wood Component. Afr. J. Range Forage Sci. 31, 179–186. doi:10.2989/10220119.2013.866161
Solomon, B. T., and Mblambo, V. (2010). Encroachment of Acacia Brevispica and Acacia Drepanolobium in Semi-arid Rangelands of Ethiopia and Their Influence on Sub Canopy Grasses. Restoration J. Bot. 5, 1–3. doi:10.3923/rjb.2010.1.13
Solomon, T. B., Snyman, H. A., and Smit, G. N. (2006b). Rangeland Dynamics of Southern Ethiopia: (2) Assessment of Woody Vegetation Structure in Relation to Land Use and Distance from Water in Semi-arid Borana Rangelands. J. Environ. Mgt. doi:10.1016/j.jenvman.10.008
Tiwari, K. P. G., Tadele, K., Aramde, F., and Tiwari, S. C. (2010). Community Structure and Regeneration Potential of Shorea Robusta Forest in Subtropical Submontane Zone of Garhwal Himalaya, India. Nat. Sci 8, 70–74.
Vadigi, S. (2013). Factors Affecting savanna Tree Sapling Recruitment. A Thesis Submitted in Partial Fulfillment of the Academic Requirements for the Degree of Doctor of Philosophy School of Life Sciences. Pietermaritzburg: College of Agriculture, Engineering and Science University of KwaZulu-Natal Pietermaritzburg.
Vadigi, S., and Ward, D. (2014). Herbivory Effects on Saplings Are Influenced by Nutrients and Grass Competition in a Humid South African savanna. Perspect. Plant Ecol. Evol. Syst. 16, 11–20. doi:10.1016/j.ppees.10.00210.1016/j.ppees.2013.10.002
Van der Waal, C., Kool, A., Meijer, S. S., Kohi, E., Heitkönig, I. M. A., and De Boer, W. F. (2011). Large Herbivores May Alter Vegetation Structure of Semi-arid Savannas through Soil Nutrient Mediation. Oecologia 165, 1095–1107. doi:10.1007/s00442-010-1899-3
Vesk, P. A. (2006). Plant Size and Resprouting Ability: Trading Tolerance and Avoidance of Damage? J. Ecol. 94, 1027–1034. doi:10.1111/j.1365-2745.2006.01154.x
Ward, D. (2005). Do we Understand the Causes of bush Encroachment in African Savannas? Afr. J. Range Forage Sci. 22, 101–105. doi:10.2989/10220110509485867
Ward, D., and Esler, K. J. (2010). What Are the Effects of Substrate and Grass Removal on Recruitment of Acacia Mellifera Seedlings in a Semi-arid Environment? Plant Ecol. 212, 245–250. doi:10.1007/s11258-010-9818-6
Ward, D., Wiegand, K., and Getzin, S. (2013). Walter's Two-Layer Hypothesis Revisited: Back to the Roots!. Oecologia 172, 617–630. doi:10.1007/s00442-012-2538-y
Werner, P. A. (2012). Growth of Juvenile and Sapling Trees Differs with Both Fire Season and Understorey Type: Trade-Offs and Transitions Out of the Fire Trap in an Australian savanna. Austral Ecol. 37, 644–657. doi:10.1111/j.1442-9993.2011.02333.x
Wiegand, K., Saltz, D., and Ward, D. (2006). A Patch-Dynamics Approach to savanna Dynamics and Woody Plant Encroachment - Insights from an Arid savanna. Perspect. Plant Ecol. Evol. Syst. 7, 229–242. doi:10.1016/j.ppees.2005.10.001
Wiegand, K., Saltz, D., Ward, D., and Levin, S. A. (2008). The Role of Size Inequality in Self-Thinning: a Pattern-Oriented Simulation Model for Arid Savannas. Ecol. Model. 210, 431–445. doi:10.1016/j.ecolmodel.2007.08.027
Williams, S. E., Marsh, H., and Winter, J. (2002). Spatial Scale, Species Diversity, and Habitat Structure: Small Mammals in Australian Tropical Rain forest. Ecology 83, 1317–1329. doi:10.2307/307194610.1890/0012-9658(2002)083[1317:sssdah]2.0.co;2
Keywords: bush control method, woody plants, density, cover, recruitment
Citation: Hare ML, Wang YD, Xu XW, Yuan Y, Na Z and Gedda AE (2021) Do Bush Control Techniques Have an Effect on the Density, Cover and Recruitment of Woody Plants in a Semi-Arid Savanna? The Case of a Semi-Arid Savanna, Southern Ethiopia. Front. Environ. Sci. 9:777146. doi: 10.3389/fenvs.2021.777146
Received: 14 September 2021; Accepted: 28 October 2021;
Published: 14 December 2021.
Edited by:
Stephen M. Mureithi, University of Nairobi, KenyaReviewed by:
Wolde Mekuria, International Water Management Institute, EthiopiaRodolfo C. R. Abreu, Universidade Federal Rural do Rio de Janeiro, Brazil
Margeret Nyaga, University of Nairobi, Kenya
Copyright © 2021 Hare, Wang, Xu, Yuan, Na and Gedda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Malicha L. Hare, bWFsaWNoYWxvamVAeWFob28uY29t; Yong D. Wang, d2FuZ3lkQG1zLnhqYi5hYy5jbg==