 Malicha Loje Hare
Malicha Loje Hare Xin Wen Xu2,3
Xin Wen Xu2,3
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Sci. , 27 July 2021
Sec. Conservation and Restoration Ecology
Volume 9 - 2021 | https://doi.org/10.3389/fenvs.2021.692239
This article is part of the Research Topic Restoration of Degraded Dryland Ecosystems: Enhancing Land/Livestock Productivity and Sustainable Livelihoods View all 6 articles
Encroachment of woody plants has negative effects on grass species and seriously challenging livestock production and pastoral livelihood in semi-arid savanna. The effect of thinning and season on grass species composition and biomass was investigated in lowland grazing area for three seasons. The study consisted of 15 plots (each 50 m × 50 m), thinned to differing intensities. The plots were located next to each other on a homogeneous area of 3.8 ha. Five thinning intensities (0, 25, 50, 75, and 100%) were allocated randomly to the plots and replicated three times. The 0% plot was left unthinned referred to as the control plot. The remaining plots were thinned to the equivalents of 25, 50, 75, and 100% of the tree density of that of the control plot. A two-way analysis of variance and Tukey Honest Significant Differences tests were used to compare responses of grass species. Ordination of variables was carried out by multivariate technique using correspondence analysis to explore the similarity relationship of plots, seasons and species. The differences in species composition and biomass in the control and the thinned plots were significant (p < 0.05). Higher grass species composition and biomass were documented in thinned plots than in the control plot. Species abundance in thinned plots ranged from 11 to 12, while that of the 0% plot was six species. Grass biomass (Kg ha-1) varied from 2,688.4 ± 736.8 to 5,035.8 ± 743.9 across thinned plots, whereas in the control plot the yield was 885.8 ± 369.1. The result showed that grass species composition and biomass increased with the increasing intensity of thinning. Grass values appeared to be optimum at the heavy intensity of thinning woody plants. Seasons were also significant with the highest values recorded in the rainy season. Rainfall played an important role by interacting with thinning and influenced total grass values. Overall, thinning that combined improved soil moisture negatively influenced woody cover and enhanced grass species composition and biomass production. Thinning of bushes to the 75% of tree density would sustain the original savanna and maximize forage production for grazers.
Savanna vegetation consists of diverse tree-grass mixtures with continuous herbaceous cover and a discontinuous cover of woody plants (Scholes and Archer, 1997). Over the past century, however, there has been a shift toward increased density and cover of woody plant species referred to as “woody plant encroachment” worldwide (Sala and Maestre, 2014; Archer et al., 2017). Encroachment of woody species is a type of range degradation where an excessive and undesirable increase in woody plant abundance results in the suppression of palatable grasses by encroaching woody species and herbaceous forbs often unpalatable to domestic livestock (Ward, 2005; Angassa and Oba, 2008). The encroaching woody plants can be non-native species that were introduced or native species that have increased in density within their historic geographic ranges (Van Auken, 2009; Archer et al., 2017).
Woody encroachment is a global phenomenon reported in many geographically different savanna ecosystems. It has been discovered that grassland and savanna communities in North America experience a decrease in plant species richness with an increase in tree densities (Ratajczak et al., 2012). In Australia, the encroachment of woody plant species tends to spread out over extensive areas of semi-arid savanna as a result of the European settlement (Noble, 1997). The expansion and encroachment of woody plants have been a challenging problem in Africa since the mid-1900 (Moleele et al., 2002). About 7.5 million km2 in sub-Saharan Africa has been under woody plant cover over the past 3 decades (Venter et al., 2018). In the savanna of the study area, the spread of woody plant species was noticed after the 1960s and 83% of the communal grazing lands are threatened by a combination of thorny woody species and unpalatable forbs (Angassa, 2007). Fifteen encroacher woody species were identified throughout the southern Ethiopian savanna (Coppock, 1994) where Acacia drepanolobium (Wiersema, 2019) is among the major encroaching woody species. It has been stated that a woody cover of 40% which is approximately equal to a density of 2,400 plants per hectare (ha−1) is considered to be at equilibrium between encroached and non-encroached (Roques et al., 2001).
The robust generalization of the causes of woody plant encroachment has been challenging worldwide (Eldridge et al., 2011). Several factors such as fire and grazing regimes, climate change and concentrations of atmospheric CO2 are widely reported to promote increases in woody density and cover (Sankaran et al., 2005; Angassa and Oba, 2008; D'Odorico et al., 2010).
Livestock forage productivity in savannas is, largely attributed to rainfall which determines the amount of water and soil moisture available for plant growth (Sankaran et al., 2004; Lehmann et al., 2011). However, the temporal or seasonal variation of rainfall in savanna is high and increases with aridity with many areas experiencing regular droughts which can be a primary cause of low forage production (Ellis and Swift, 1988; Angassa and Oba, 2007).
Woody plant encroachment is threatening the productivity of the world’s savanna grasslands with a negative effect on ecology and the economic status of more than two billion people worldwide (Eldridge et al., 2011; Buitenwerf et al., 2012) owing to the loss of ecosystem goods and services (Van Auken, 2009). Dense woody plants have negative effects on herbaceous species due to competition for available soil water, nutrients, and decreased light intensity for under canopy species (Kahi et al., 2009; Teshome et al., 2012; Ward et al., 2013). Such suppression of grasses by woody species leads to a decline in grazing capacity in grazed savannas (Kellner, 2008; Kangalawe 2009).
Savanna grassland is the main feed source for the livestock in southern Ethiopia but has lately been under threat from woody encroachment which is seriously challenging livestock production and pastoral livelihood. Bush clearing was considered as a solution to improve livestock forage supply, especially for grass-dependent cattle (Angassa, 2002). However, bush clearing efforts did not have a positive impact on rangeland conditions except temporary forage improvement for cattle (Smit, 2004; Gemedo et al., 2006; Angassa and Oba, 2009). The assumption is that the establishment of a woody plant is normally a continuous process in savanna and so control cannot be achieved with single cutting operations.
Maintenance of a desirable combination of grass and woody plants might be a sound and practical approach to sustainable vegetation management in savannas. Thinning of trees to some predetermined density, after which post-thinning management will be needed to implement desired composition rather than eliminating woody plants. Through selective thinning, the development of structured savanna with large trees is encouraged, and these large trees can suppress the establishment of new seedlings which is particularly noticeable with Acacia species (Smit et al., 1999; Smit, 2014). Tree thinning employing mechanical method results in immediate changes in composition between woody and herbaceous plants. And it will normally result in an increase in grass production and thus also the grazing capacity (Smit, 2005). In general, the productivity levels of savannas are on a broad scale related to the relative proportion of trees to grasses (Smit, 2004; Solomon and Mblambo, 2010). Understanding the effects of thinning and seasonality on grass species is important for the management of savanna ecosystems.
So far, the effect of thinning intensity and seasonality on grass species has not yet been investigated in the area. Therefore; the objectives of this study were: 1) to examine the effect of thinning intensity on grass species composition and biomass production, 2) to evaluate the effect of season on grass values of the selectively thinned savanna, and 3) to establish relations between thinning intensity, season and total grass values (i.e., species abundance and biomass). The following hypothesis was tested:
Ho: Woody tree thinning and seasonality does not affect grass species composition and biomass production.
The study was undertaken in the Borana lowland of southern Ethiopia. The Borana savanna lies approximately between 4o 36′ to 6o 38′ N latitude and 36o 43′ to 42o 30′ E longitude, and the landscape is characterized by slightly undulating peaks up to 2000 m above sea level (m.a.s.l) in some areas (Coppock, 1994). The weather is characterized by erratic and variable rainfall with most areas receiving between 238 and 896 mm (mm) annually (Angassa and Oba, 2007). The rainfall pattern is distinctly bimodal, 59% of annual precipitation occurs from March to May, while 27% occurs from September to November (Coppock, 1994). The region is dominated by savannah type of vegetation containing a mixture of perennial grasses and woody plants (Desta and Coppock, 2004). The typical African savanna vegetation consists of a diverse range of tree-grass mixtures, different species of perennial grasses and sedges, trees, woody plants and shrubs with the grass cover relatively continuous and woody cover discontinuous (Scholes and Archer, 1997; Archer et al., 2017). The dominant vegetation-type of the study site is acacia grassland and consists mostly of perennial grasses and sparsely distributed woody plants. The grass cover is dominated by Cenchrus ciliaris and Chrysopogon aucheri, while the woody cover consists mainly of Acacia drepanolobium trees on black cotton soil sites (Coppock, 1994). However, a shift from the open savanna grassland to closed woodland has taken place in the area. Encroachment of woody plants is the major form of rangeland characterized by invasion of undesirable woody species, shrubs and unpalatable forbs, and losses of grasses (Oba, 1998; Angassa, 2007).
The main agricultural system in the Borana rangelands is livestock production and the main economic value is provision of grazing for livestock. Communal grazing is the main land use system in the area (Helland, 1997). The Borana savanna had good ecological potential for livestock production and was considered until the early 1980s as one of the few remaining productive pastoral systems in East Africa (Cossins and Upton, 1987). The Borana pastoralists traditionally practiced strategic grazing management to avoid local overstocking around the scarce dry season water sources. They used different grazing areas in the dry and wet seasons; lactating cows with their calves were herded separately from dry cows, bulls and young stock (Coppock, 1994). This management regime in combination with episodic climatic events and the planned use of fire maintained a relatively stable tree-grass balance (Angassa, 2002). However, in the last few decades, increasing grazing pressure and the development of water ponds has resulted in more sedentary settlements and enhanced crop cultivation which minimized grazing land. Overgrazing, changes in land use and suppression of fire have resulted in the proliferation of bush encroachment and a general decline in forage production (Oba and Kaitira, 2006; Angassa and Oba, 2007).
In African savannas, fires are ecological process that impact on the savannas ecosystem processes and also influence functions and service delivery of savannas (Ansley and Castellano, 2006; Angassa and Oba, 2008). Fire suppresses woody plants growth by killing and reducing their density (Angassa and Oba, 2008). Frequent fires reduce woody cover and maintain woody vegetation in a juvenile state by “top-killing” seedlings and saplings, retarding transition to adulthood in tree species which can re-sprout from rootstocks after damage of aboveground structures (Connor, 1995; Kaiser, 1999; Sankaran et al., 2008). If implemented properly, and used in conjunction with other appropriate range management practices, prescribed fire can be used to reduce bush encroachment and increase the forage production and quality for grazing animals (Oba, 1998; Sankaran et al., 2008; Joubert et al., 2012; O'Connor et al., 2014). In fact, fire can produce physical, chemical and biological alterations in soil properties. The changes to the soil produced by controlled fire are in most cases only transient (Úbeda and Outeiro, 2009). A study report from the semi-arid savanna also indicated that fire cannot effectively be used as a management technique for bush thinning due to a lack of enough herbaceous material caused by increased grazing pressure and bush encroachment (Joubert et al., 2012).
In the study site, the pastoral community periodically undertook range burning in the dry season before the site was transformed to closed woodland (Angassa and Oba, 2009). According to the opinion of the community members, because of the lack of enough herbaceous material, fire cannot be used as it was in the past. The encroachment of woody plants particularly acacia species, resulted in severe degradation of the area and critical feed shortage particularly during dry periods, threatening the livelihoods of the local community.
Prior to site selection, discussion was held at the district level to create awareness on the purpose of the study and also to identify the representative site for this purpose. Selection criteria was developed and included: 1) areas representative of bush encroachment with the most widely distributed species; 2) sites characterized by a dense stand of woody plants with poor grass cover; 3) representative rangeland-livestock based pastoral production systems of Borana pastoralists’ land use type, and 4) easily accessible. Based on the above criteria, representative major grazing areas encroached by different woody plant species were assessed through participatory reconnaissance survey by a team comprising community members, elders (7) who have enough knowledge about the history of rangeland vegetation, experts from pastoral offices (3) and the researcher. Accordingly, the Dida Yabalo communal grazing site with abundant mature trees of A. drepanolobium was selected as the study site and the coordinates were taken (i.e., 04o 12′ 0.67′′ N and 05o 47′ 8.76′′ E, 04o 12′ 4.07′′ N and 05o 47′ 8.02′′ E, 04o 12′ 4.12′′ N and 05o 47′ 9.01′′ E, 04o 11′ 2.73′′ N and 05o 47′ 9.12′′ E, 04o 12′ 0.76′′ N and 05o 47′ 9.22′′ E.). The altitude of the experimental site is 1,518 m.a.s.l. with a valley-plain type of landscape.
The study site consisted of 15 plots (each 50 m × 50 m), thinned to differing intensities. The plots were located next to each other on a homogeneous area of 3.8 ha. Five thinning intensities (i.e., 0, 25, 50, 75, and 100%) were allocated randomly to the plots and replicated three times. Thinning intensities hereafter called treatments and categorized as un-thinned (0%), light thinning (25%), moderate thinning (50%), heavy thinning (75%), and total clearing (100%). Trees were randomly marked for removal during the thinning process. The 0% plot was left unthinned referred to as the control plot, representing the current bush invaded range sites against which the other treatments could be evaluated. The remaining plots were thinned to the equivalents of 25, 50, 75, and 100% (total clearing) of the tree density of that of the 0% plot (Smit, 2005). This ensured a fairly even spread of the remaining trees without favoring a particular tree size. The resultant thinned plots resembled the structure of naturally occurring open stands of woody plants in savanna. During thinning, trees were sawn off at ground level and removed from the plot. The cut stumps were treated with fire using dry cattle dung as a fuel load, thus ensuring that the sawn trees were killed without affecting the remaining plants. Thinning was completed in February during the dry season of 2017. Borders of 10 m and 1 m distance between treatment plots were left to minimize edge effects. The study site was fenced using removed stems to exclude disturbances. The response of the grass species was studied during the two growing and one dry seasons following tree thinning.
The number of A. drepanolobium trees in the experimental plot was counted during the pretreatment and post-treatment periods. Woody densities were calculated as the number of trees per plot (2,500 m2) and averaged to obtain the mean density per plot. The value obtained was then multiplied by 10,000/2,500 to derive the density ha−1. To determine the total area of coverage by individual woody canopies, we measured the canopy diameter (d) of 12 randomly selected trees within each treatment plot by spreading measuring tape either beneath or above the canopy as appropriate. The canopy diameter then gave a canopy radius (r) measurement (r = ½ d). The A. drepanolobium trees have a round canopy (Okello et al., 2001) and we modeled the shape covered by the canopy as circular and calculated the area covered by each canopy as Area = πr2. The total area covered by all tree canopies within the plots is defined as the ratio of 1-ha area covered by woody canopies (Innocent and Munyati, 2014).
Rainfall data was taken from the nearby meteorological station installed by the Meteorological Service Agency of Ethiopia. Also, the soil moisture content was determined by taking 200 g of soil in the five treatment plots to a depth of 200 mm approximately close to the root zone of herbaceous species (Ward et al., 2013) using auger after a rainfall event in April, August, and October, and moist weight was taken immediately. Two samples were prepared per plot and oven-dried for 24 h at 105°C and weighed to determine the moisture content of the soil by subtracting the oven-dry weight from the moist weight (Dirksen, 1999). Moisture content is expressed as a percentage of the oven-dry weight of the soil.
Grass biomass was measured by mowing fresh biomass using sickle in quadrates (0.5 m × 0.5 m). Each grass species in a 0.25 m2 area was identified and recorded in the field with the help of herdsmen and technical experts. The name of the species was cross-checked with Gemedo et al. (2005). A total of 60 quadrates per plot were sampled by randomly placing in each of the thinning plot (Smit, 2005). Individual grass species were counted in each quadrat to determine species abundance. The frequency of grass species was calculated as the number of quadrates in which the species occurred over a total number of quadrates per plot times 100 representing each thinning plot. It is expressed in terms of percentage. Grass species were categorized into different desirability classes namely: highly desirable, desirable, and less desirable as preferred by grazers (cattle and sheep) based on local ecological knowledge of herdsmen derived from focus group discussion. The group was organized at the grazing area level and composed of range scouts/herdsmen (5) and technical expert (1) all having better experience and knowledge about forage grasses. All grass samples were clipped per species at ground level and then placed in a plastic bag separately for dry matter (DM) analysis. Samples were oven-dried at 100°C for 24 h and then weighed (ILCA, 1990; Rau et al., 2009). Sampling was carried out for three seasons (two rainy and one dry season) hereafter called Season 1 (March–May), Season 2 (June–August), and Season 3 (September–November) of the study period. Control grazing was applied following sampling of fresh grass biomass using cattle at light stocking rate (Smit et al., 1999; Smit, 2005) to minimize carryover of standing dry biomass to the coming season. In order to achieve intended impact on standing dry hay, 20 heads of cattle ha−1 were herded in the plots at the end of the sample season for 7 days after the grass was fully dry and enriched the soil seed bank. This stocking rate was not based on objective assessment of the available forage, but was intended to impact the standing dry biomass moderately which might complicate the fresh growth and sampling in the coming season if left undisturbed.
A two-way ANOVA and Tukey (HSD) tests were used to test for differences in species composition and biomass for all plots and seasons. This was done to verify any effect of thinning and season, and their interaction on the grass layer in the area. The treatment plots, seasons, and species recorded in each of the five plots and three seasons were ordinated using correspondence analysis (Greenacre, 2007). The CA analysis determines similarity relationships among plots, seasons, and species by mapping the grass data in two-dimensional space in which ecologically similar plots, seasons, and species are located near each other and dissimilar plots, seasons and species far apart. Natural groups of species, if present, appear as clusters and distinct groups as a cluster of species separated by gaps (Abule et al., 2005). All computations were performed in R statistical packages (R Development Core Team, 2018), making use of the ca package for CA developed by Nenadi and Greenacre (2007). Regression analysis was performed to examine the relationships between thinning intensity and grass values (i.e., species abundance and biomass). Significant differences were considered at p < 0.05.
Table 1 provides calculated densities ha−1 of trees within the different plots. The pre-treatment density of A. drepanolobium trees in the individual treatment plots was more or less similar (2,677.8 ± 25.80 ha−1). However, the post-treatment densities differed significantly (p < 0.05) with the lower densities recorded in the thinned plots (25, 50, 75, and 100%) compared to the control plot. The total area covered by woody canopies in the various plots is computed from the respective mean canopy area of trees. The result showed that a higher (p < 0.05) woody canopy cover was recorded in the un-thinned plot which had higher tree density than the other four thinned plots. In thinned plots, the canopy area decreased over the gradient of increasing intensity of thinning, up to the point of no representation in the total cleared (100%) plot.
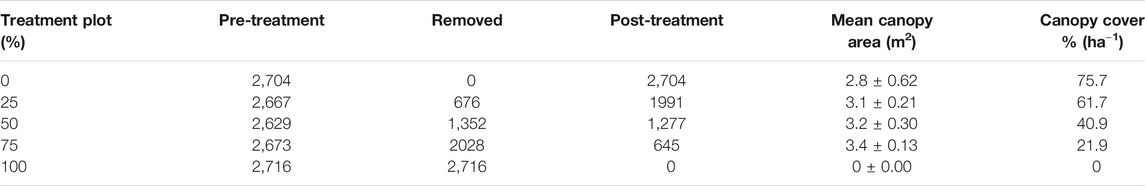
TABLE 1. The pre and post-treatment densities (ha−1), mean canopy area and cover of woody plants in the various thinning plots in the savanna of southern Ethiopia.
According to the rainfall data collected in 2018, total monthly rainfall ranged from 0.0 mm (millimeter) in January to 379.1 mm in April (Figure 1). The total monthly rainfall was significant (p < 0.05) with the peaks in April and October which demonstrates the bimodal type of rainfall of the study site. The amounts of seasonal rains also varied significantly (p < 0.05) with the higher rain recorded in season 1 followed by season 3. The total seasonal rainfalls were 642.1, 8.6, and 257.3 mm in seasons 1, 2, and 3, respectively. The total annual rainfall of the study year was 960.3 mm including 52.3 mm received during the dry months of December, November, and January 2018.
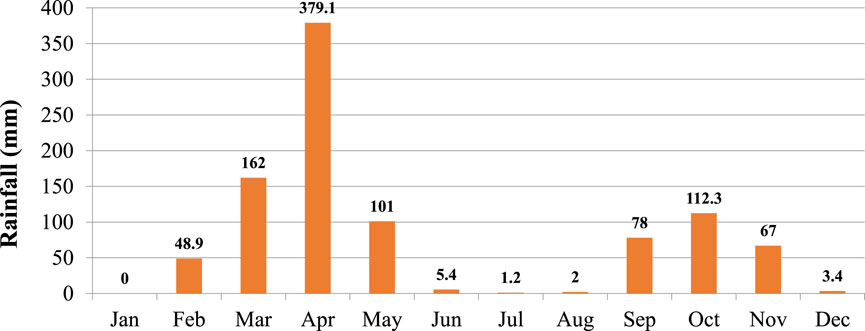
FIGURE 1. Total monthly rainfall of the study site in Borana area of Southern Ethiopia. A 5% significant level was used to analyze the means (n = 12).
Table 2 provides the seasonal soil moisture contents of the various treatment plots in the experimental site. The result showed insignificant (p > 0.05) difference among the five treatment plots. Over the seasons, the average soil moisture contents were 21.7, 22.3, 22.1, 22.1, and 21.8% in the 0, 25, 50, 75, and 100% plots, respectively. There was a significant (p < 0.05) difference among seasons in terms of soil moisture contents with the highest moisture level recorded during the main rainy season (season 1). The average soil moisture levels were 34.6% in season 1, 4.9% in season 2, and 26.5% in season 3.
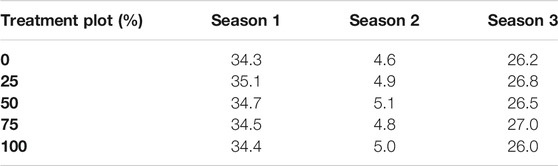
TABLE 2. Soil moisture contents (%) within the various thinning plots and season in the savanna of southern Ethiopia.
A total of 15 grass species were recorded in the five treatment plots sampled for three seasons (Supplementary Appendix Table A1). Out of the 15 identified types of grass, 4 (%) and 11 (%) species were annuals and perennials, respectively. Of these species recorded 53.3, 26.7, and 20.0% were highly desirable, desirable, and less desirable grasses, respectively.
Derived from Supplementary Appendix A, Table 1, the proportion of the individual classes of grass species differed significantly (p < 0.05) among the five treatment plots. The percentage composition of the highly desirable species included 62.5% (five species) and 100% (eight species) in the 0 and 50% plots, respectively while the value in the 25, 75, and 100% plots was 87.5% (seven species). Thinned plots had the larger proportion of highly desirable class of grasses where the greater value was recorded in the 50% plot. There was no significant (p > 0.05) difference among the 25, 75, and 100% plots evidencing the decline of this particular class of species after the moderate (i.e., 50%) thinning of woody plants. Similarly, the composition of desirable class of grasses was significant (p < 0.05) among the five plots. Their abundance was higher in thinned plots than in the control (0%) with the higher value obtained in the 50, 75, and 100% plots. Of the desirable class of grass recorded, 25% (one species) and 50% (two species) were encountered in the 0 and 25% plots, respectively although the percentage composition recorded for the other three plots (50, 75, and 100%) was 75% (three species) which were indifferent to each other. Likewise, there was a significant difference (p < 0.05) among the five plots in terms of the proportion of the less desirable class of species. Higher proportion of the less desirable grasses was observed in the four thinned plots compared to the unthinned one. However, significant difference was not obtained among the 25, 75, and 100% plots. The composition in the above mentioned three plots was 66.7% (two species), and 33.3% (one species) of this particular class was observed in the 50% plot. The less desirable class of species was almost absent in composition in the control plot. According to the current study, thinned plots had the larger proportion of highly desirable, desirable and less desirable classes of grass species than the unthinned plot indicating that thinning increased the composition of different classes of grasses in the study savanna. On comparing the individual classes, the highly desirable class had the largest (p < 0.05) proportion throughout the gradient showing an increasing trend with the highest value obtained in the 50% plot area. The highly desirable species dominated (84.9%) the bulk of the desirability classes of grasses followed by the desirable class of species (60%). According to pastoralists’ assessments, the existence of highly desirable and desirable grasses indicates good rangeland quality, while the existence of less desirable species indicates poor rangeland quality.
The composition of grass species differs along the thinning gradients. The thinned plots had proportionally more (p < 0.05) individuals of grasses with the highest value recorded in the 50, 75,, and 100% plots which did not differ from each other. Percentage species composition was lower in the control than in the other four thinned plots. The coverage of species composition was 40% (six species) in the control, 73.3% (11 species) in the 25%, and 80% (12 species) in each of the 50, 75,, and 100% plots.
The rate of occurrence of grass species (Supplementary Appendix Table A1) was dissimilar across the various plots as indicated by ordination analysis (Figure 2). The highly desirable species such as C. ciliaris, Digitaria milanjiana and Eragrostis papposa were most frequent in the 25% plot, while C. aucheri occurred abundantly in the 50% plot. Similarly, Bothriochloa insculpta, Chloris roxburghiana, Cynodon dactylon, Sporobolus pellucidus, and Themeda triandra were most frequent in the 75% plot. Setaria verticillata, the desirable annual grass, and the less desirable grasses including Eleusine Jaegeri and Pennisetum mezianum were the most abundant species in the 100% plot. The abundance of Aristida adscensionis revealed high in the 100% plot, whereas that of Dactyloctenium species was in the 0% plot. The greater frequency of Panicum maximum (the highly desirable perennial grass) was evident in the dense (0%) plot than in the thinned plots.
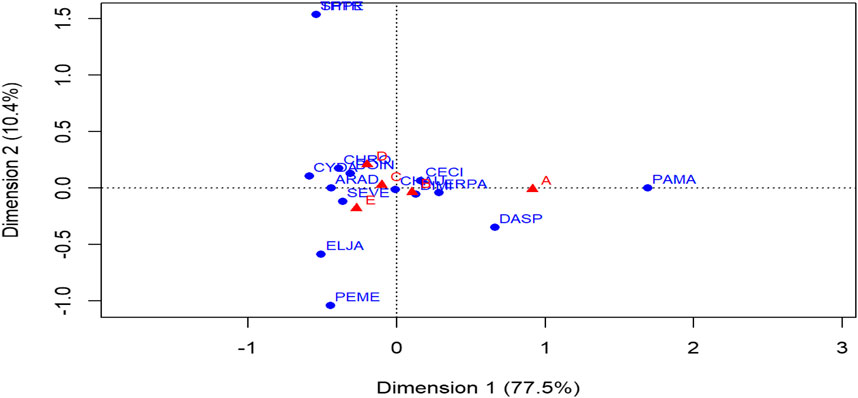
FIGURE 2. CA ordination of grass species abundance by thinning plots in Borana rangelands of Ethiopia. “Dimension 1” represents ‘Columns,’ “Dimension 2” represent ‘Rows.’ Red diamond denotes plots: (A) = 0%, (B) = 25%, (C) = 50%, (D) = 75%, and (E) = 100%. Blue dot specifies species. Species abbreviations are: ARAD, Aristida adscensionis; BOIN, Bothriochloa insculpta; CECI, Cenchrus ciliaris; CHRO, Chloris roxburghiana; CHAU, Chrysopogon aucheri; CYDA, Cynodon dactylon; DASP, Dactyloctenium species; DIMI, Digitaria milanjiana; ELJA, Eleusine jaegeri; ERPA, Eragrostis papposa; PAMA, Panicum maximum; EME, Pennisetum mezianum; SEVE, Setaria verticillata; SPPE, Sporobolus pellucidus, and THTR, Themeda triandra.
Correspondence analysis based on species frequency separated the species into recognizable patterns representing the five thinning intensities although there was some overlap (Figure 2). The two CA dimensions, cumulatively explained 87.9% of the total variance in species abundance along the gradient owing to differences among grass species preference to thinning intensity.
The average (± SE) total grass biomass (kg DMY ha−1) was 885.8 ± 369.1, 2,688.4 ± 736.8, 3,733.2 ± 791.5, 4,163.4 ± 811.3, and 5,035.8 ± 743.9 in the 0, 25, 50, 75, and 100% plots, respectively. The result showed that there was a highly significant (p < 0.001) difference in grass biomass among the five gradients. Mean comparison test also showed significant (p < 0.05) difference (Figure 3). The grass biomass was generally low in the un-thinned plot (0%), while in the thinned plots the biomass were substantially higher, with the yield where trees have been totally removed (100% plot) being the highest. Total grass biomass showed an increasing trend over the gradient possibly due to reduced competition from A. drepanolobium trees.
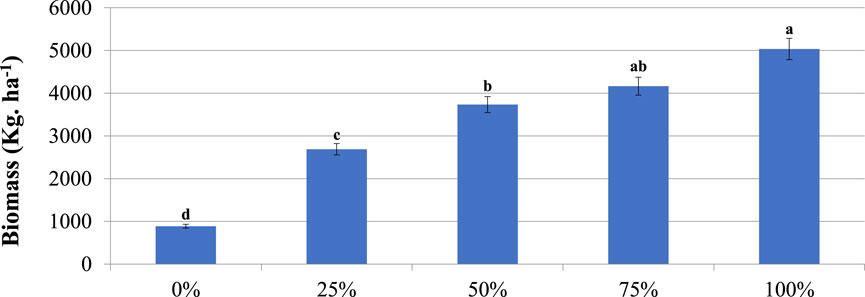
FIGURE 3. The effect of thinning on grass biomass in the communal grazing lands of southern Ethiopia. % = thinning plot. Significance differences between plots are indicated by different letters. Vertical bars mean a 0.95 confidence interval.
The biomass of the individual grass species within the five treatment plots is provided in Table 3. The productivity of the most dominant key forage species was significant (p < 0.001) within the various plots. Higher biomass was recorded by C. aucheri across the thinning gradient followed by C. ciliaris, however both with a declining trend after the moderate (50%) thinning intensity. The other desirable grasses such as A. adscensionis, B. insculpta, C. roxburghiana, and D. milanjiana also showed similar trend with a better performance in the total cleared (100%) plot. The trend of others was not apparent due to low and fluctuating yields. Overall, C. aucheri (960.60 ± 50.30 kg ha−1) and C. ciliaris (786.81 ± 49.04 kg ha−1) were the most yielding perennial grasses in this order.
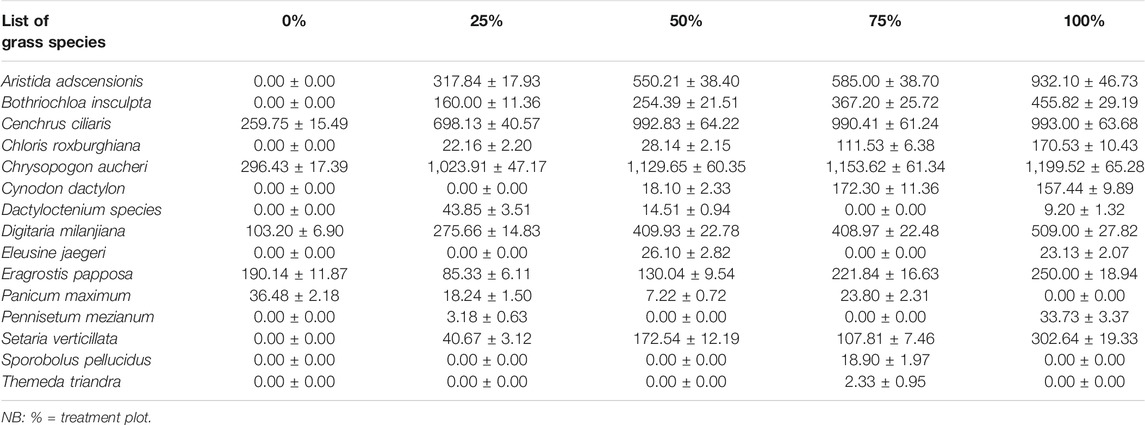
TABLE 3. Mean (± SE) biomass productivity (Kg. ha−1) of the individual grass species along the thinning gradient in the savanna of southern Ethiopia.
Seasonal composition of the individual grass species and different desirability classes is provided in Supplementary Appendix Table A2. Percentage composition of grass species differed significantly (p < 0.05) across seasons. The results showed the highest total species composition in season 3 (93.3% or 14 species) followed by season 1 (80% or 12 species) indicates that moisture played an essential role and initiated the establishment of grass species. Lowest species composition (33.3% or five species) was recorded in season 2 which received only a shower of rainfall.
The composition of the different classes of grass was significant (p < 0.05) across the three seasons. Higher percentage composition of the highly desirable grass (100% or eight species) was observed in season 1 followed by the desirable (75% or three species) class, while the lowest value was recorded for the less desirable (33.3% or one species) class of species. Similarly, in season 2 percent composition of the highly desirable class (50% or four species) was higher than those for the desirable (almost absent) and less desirable (33.3% or one species) classes of species. Also in season 3, the composition of the highly desirable class is higher than the desirable class but had comparable value with the less desirable one. Significant difference was not obtained between season 1 and 3 in the percentage composition of the highly desirable class of grass species. Likewise, there was no significant (p > 0.05) difference between season 1 and 3 in terms of percentage composition of the desirable class of grasses.
Overall, season 1 and 3 (i.e., the two rainy seasons) were comparable and greatly increased the composition of the highly desirable class of grass species in relation to season 2 (the dry season). The results indicate that effective rainfall greatly enhanced the composition of the highly desirable class of species. Next to the highly desirable class, the composition of the desirable and less desirable classes was also increased in the main and small rainy seasons than in the dry season. Dry period has negative effect on the abundance of different desirability classes of grass.
The frequency of the individual grass species was dissimilar among the three seasons (Figure 4; Supplementary Appendix Table A2). C. ciliaris, C. aucheri, and D. milanjiana were abundantly distributed across season 1, 2 and 3. Also, A. adscensionis showed stability in the dry season (season 2) following its establishment during the first rainy season. Some of the grass species such as E. papposa, P. maximum and S. pellucidus were abundant in season 1. Similarly, C. dactylon and S. verticillata were commonly frequent in season 1 and season 3, while others like B. insculpta, C. roxburghiana, E. jaegeri, P. mezianum, T. triandra and Dactyloctenium species were abundantly found in season 3. The CA dimension 1 explained the most variance among seasons with 78.3%, followed by dimension 2 with 21.7%. Cumulatively, dimensions 1 and 2 indicated 100% of the total variance in species composition and abundance owing to differences among grass species in response to seasonal soil moisture.
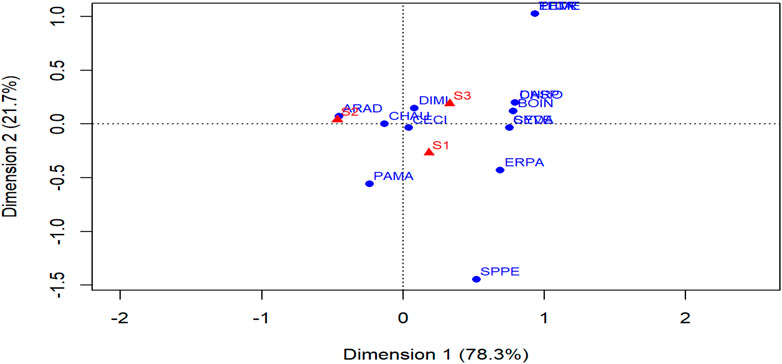
FIGURE 4. CA ordination of grass species abundance by season in the woody thinned Borana rangelands of Southern Ethiopia. “Dimension 1” and “Dimension 2” represents ‘Columns’ and ‘Rows,’ respectively. Red diamond represents season. S1, season 1; S2, season 2, and S3, season 3. Blue dot means species. Species codes are: ARAD, Aristida adscensionis; BOIN, Bothriochloa insculpta; CECI =Cenchrus ciliaris; CHRO, Chloris roxburghiana; CHAU, Chrysopogon aucheri; CYDA, Cynodon dactylon; DASP, Dactyloctenium species; DIMI, Digitaria milanjiana; ELJA, Eleusine jaegeri; ERPA, Eragrostis papposa; PAMA, Panicum maximum; EME, Pennisetum mezianum; SEVE, Setaria verticillata; SPPE, Sporobolus pellucidus, and THTR, Themeda triandra.
Supplementary Appendix Tables B1–B3 provide the seasonal biomass of grass species within the different treatment plots. The seasonal grass biomass of the thinned plots differed significantly (p < 0.05) largely following rainfall patterns (Figure 5). Highest grass biomass in the treatment plots was achieved during season 1 which received higher amount of rainfall compared to the other two seasons. In season 1 the total grass biomass increased following the thinning gradient where the highest value was recorded in the totally cleared (100%) plot. There was a significant difference (p < 0.05) in grass biomass among the different plots during season 3 of the study year. Thinned plots had higher grass biomass than the unthinned plot. The grass biomass was comparable and highest in the 75 and 100% plots. However, the difference between the 25 and 50% plots was insignificant (p > 0.05) during this period. In contrast, significant difference was not seen in terms of kg ha−1 of grass biomass among the different treatment plots during season 2 which received lowest amount of rainfall. On the other end, higher (p < 0.05) biomass was recorded in the 100% plot in season 1; however this condition was changed to an insignificant (p < 0.05) difference during season 3 between the total cleared (100%) and heavily thinned (75%) plot. The high biomass where trees were removed may be only temporary probably due to the declined positive tree-grass interaction which is more disadvantageous for grasses. The biomass of the control (0%) plot was similar in season 1 and season 3 both received normal amount of rainfalls displaying that, soil moisture had little effect on grass biomass in high tree density grazing lands possibly attributed to increased competition by encroaching woody plants.
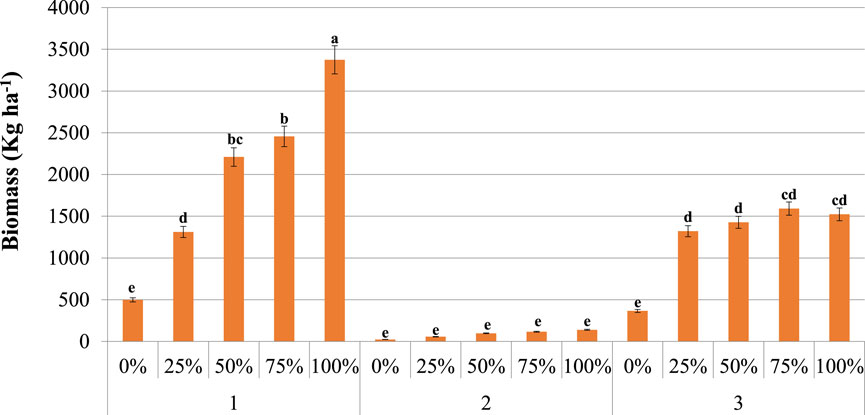
FIGURE 5. The combined effects of season and thinning on grass biomass in the savanna of southern Ethiopia. % = treatment plot. #1 = Season 1, #2 = Season 2, and #3 = Season 3. Significant differences between plots based on Tukey HSD pairwise tests are indicated by different letters. Vertical bars denote a 0.95 confidence interval.
According to the current study, the mean (± SE) seasonal grass biomass of the selectively thinned range site (kg DMY ha−1) was 1970 ± 519, 86.2 ± 18.3, and 1,245.3 ± 359.7 in season 1, 2, and 3, respectively with the total annual biomass of 3,301.5 ± 897 kg. ha−1 (3.3 ± 0.9 ton ha−1). The results showed highly significant (p < 0.05) difference (Figure 6) across seasons with varying amounts of rainfalls. The grass biomass was higher during the first main season (season 1) indicating that increased rainfall which in turn enhanced soil moisture status played a significant role in improving grass biomass production in the woody thinned savanna rangelands.
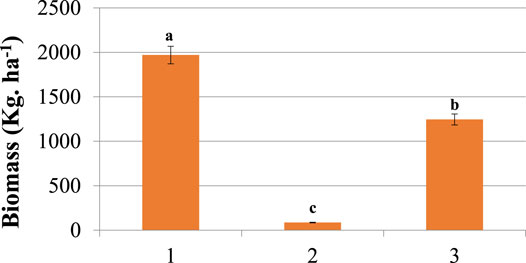
FIGURE 6. Comparison of seasonal grass biomass production of the selectively thinned savanna of southern Ethiopia. Season code: #1 = Season 1; #2 = Season 2, and #3 = Season 3. The presence of significant difference among seasons is indicated by different letters. Vertical bars mean a 0.95 confidence interval.
The study showed that there was an interaction (p < 0.001) between thinning intensity and season on grass biomass (p < 0.05) indicating the variability of biomass production among different thinning plots across seasons of varying amounts of rainfall.
Thinning intensities and grass variables were incorporated in assessing their relationship. There was a significant (p < 0.05) linear relationship among thinning intensity, species composition, and biomass (Table 4) which indicates that species abundance increase with increasing intensity of thinning which in turn increased grass biomass. Interestingly, grass biomass appeared to be ideal at between the moderate and (50%) and heavy (75%) thinning gradients possibly attributed to the prominent abundances of the highly desirable grasses such as C. aucheri and C. ciliaris in these two gradients. Despite the higher biomass achieved at the 100% plot, the total species abundance in the 50, 75, and 100% plots was similar and stable across seasons. Besides, a decline in biomass that was observed in the 100% plot during the third season further validated that optimum grass values could be achieved at the 75% of thinning woody plants. In this gradient (i.e., 75%) maximum species abundance was 12 (ha−1), whereas peak biomass was 4,163.4 ± 811.3 kg ha−1.
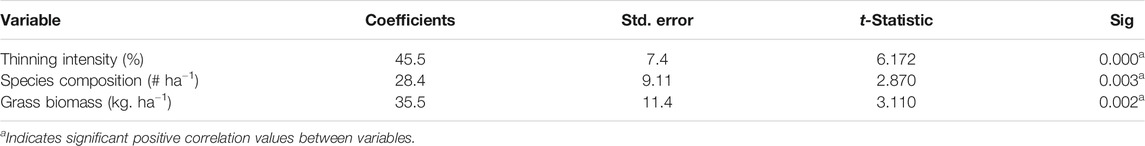
TABLE 4. Correlation coefficient values between thinning, grass species composition and biomass production.
Total grass species composition was highly significantly (p< 0.001) positively correlated with rainy seasons (main and small rainy seasons) but negatively associated with the dry season (Table 5). Similarly, the biomass of grass species was positively correlated with rainy seasons, while it was negatively related to dry season. The finding showed that, total grass values particularly the biomass increases with the increase in the amount of rainfall which in turn increase the availability of soil moisture for grass growth following thinning of woody plants.
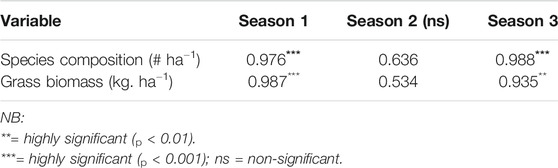
TABLE 5. Correlation analyses (n = 5) of relations between season, grass species composition and biomass production.
The negative effect of densely populated A. drepanolobium trees was evident in the 0% plot which had more (60%) rare species indicating that this limited abundance of desirable forage grasses in the dense plot may be one indicator of the deteriorating condition of savanna rangelands. Liao and Fei (2017) discussed shifts in grass species composition toward woody plant encroachment in the savannas of Southern Ethiopia with a general decline in desirable forage species. The consequence of such a shift in species composition might result in a high risk of livestock feed shortage.
Grass species composition will have an effect on animal grazing behavior, abundance and distribution of the species in savanna rangelands. Species composition determines forage production of rangelands which is important to livestock production system (Smit et al., 1999; Smit, 2005). The results of this study showed that thinning increased grass species composition. The observed increase in species composition mainly ascribed to reduced woody density and cover whereby sunlight interceptions were improved for grasses. The establishment of grass species was initiated by the reduced competition from A. drepanolobium following thinning. Similar findings (Burton, 2000; Ludwig et al., 2004; Sang-Hyun et al., 2018) have been reported globally that thinning reduces canopy cover and increases the utilization of sunlight use by remaining understory plants. Because woody species removed by mechanical thinning are typically sparsely distributed throughout the system, thinning may increase understory light variability by creating light patches in areas where the trees are thinned, while leaving light levels relatively unaffected at un-thinned sites (Chiang et al., 2012). Sun-light is particularly important for photosynthetic plants as it is the main source of energy to carry on the physiological functions working and, consequently has an enormous influence on plant development (Thomas, 2006).
Forage production of rangelands is important to pastoral livestock systems and animal production is in most cases directly related to the productivity, perenniality, desirability and acceptability of the plants to the animals (Oba et al., 2000; Smit, 2005). According to the study results, thinning enhanced the composition/abundance of the highly desirable and desirable classes of the perennial grass species. The treated rangeland site at the end of the study in general, resembled the original savanna covered mostly by highly desirable class of grasses that dominated the bulk (56.9%) of the desirability classes of grass species. According Abule et al. (2005), perennial grasses add to the nutritive value of rangeland and improve livestock productivity which in turn enhances people’s livelihood.
The most abundant highly desirable perennial grass was C. aucheri (19.2%) which mostly confined to the light thinning area. According to Ahmad et al. (2009)C. aucheri is the perennial C4 bunchgrass and the most widespread species found in the semi-arid grasslands and bush lands of the Horn of Africa (Herlocker, 1999; Solomon and Mblambo, 2010) and is preferably grazed by cattle (Gemedo et al., 2005). Herlocker (1999) indicated that C. aucheri is very sensitive to grazing whereby heavy grazing results in the elimination of the species.
The second most abundant perennial grass C. ciliaris (15.7%), showed preference to light thinning (25%). The species is commonly known as “Buffel grass,” is grown widely in tropical and subtropical arid savannas around the globe because of its high drought tolerance and capacity to withstand heavy grazing (Marshall et al., 2012). The species has higher palatability and better adaptation to hot dry areas with high evapotranspiration (Herlocker, 1999; Guevara et al., 2009). Lazarides et al. (1997) stated that Buffel grass is uniquely suited to the pastoral system because it has high nutritional value for sheep and cattle, and the ability to withstand heavy grazing, a deep stabilizing root system and responds quickly to rainfall events. More recently, Buffel grass is highly valued for site rehabilitation and erosion control (Solomon et al., 2010).
Among the most abundant species in the cleared plot (100%) were C. roxburghiana and A. adscensionis. The species were widespread in drylands areas and is usually found growing in association with C. aucheri and C. ciliaris in Commiphora and Acacia woodland. Chloris roxburghiana is an important species for livestock and wildlife. This species contributes up to 50 percent of the diet of wild herbivores in eastern Kenya (IBPGR, 1984; Herlocker, 1999) but is in danger of disappearing due to land degradation.
The only highly desirable perennial grass that did not appear in the 100% plot was P. maximum however relatively abundant in the un-thinned plot. This finding confirmed the study by Abule et al. (2005) in the mid-rift valley area of Ethiopia showing that Panicum maximum grows better underneath the tree canopy. Panicum maximum is a tall, fast-growing, and nutritious grass species preferred by herbivores (Smit, 2005). The species is often found associated with Cenchrus and Bothriochloa in Acacia woodland in the dry savannah areas. Herders in the study area also reported that P. maximum is a highly preferred grass species, particularly by cattle. The species usually grows under canopy area, stay greener for a longer period and serve as standing hay to feed calves and weak animals during the dry period. It is, therefore, important to note that P. maximum would probably be lost with the complete elimination of trees.
The preference of grass species to various tree densities under the current study confirms the observation from Tunisia (Abdallah and Chaieb, 2013) that show a positive interrelationship between the woody plants and grass layer. The finding of the current study, therefore, suggests that total grass species composition and biomass production may be favored by the presence of woody plants to a certain density. Clearing of woody plants in areas dominated by acacia trees may result in the loss of desirable nutritious grasses and a shift in total biomass yields of the most desired species. Such losses in species composition may affect negatively grazing livestock and the livelihood of the community.
In general, changes in understory species composition following thinning of woody plants have been recorded worldwide. For example; in South Korea, an increase in herbaceous vegetation density was recorded where Chamaecyparis obtusa stands have been thinned around Mt Moonsu (Sang-Hyun et al., 2018). Thinning of Colophospermum mopane trees in South Africa increased the grass species (Smit and Rethman, 1999). In Taiwan, thinning of the Cryptomeria japonica forest between 25 and 50% has increased plant diversity and richness (Tsai et al., 2018). Distinct changes in the grass species composition were also observed during this study and the pattern of establishment of the various grass species demonstrates some indication of the successional order of establishment.
The grass biomass recorded in the 0% was lower compared to thinned plots possibly attributed to the negative effect of A. drepanolobium trees which out-competed grass species for light, soil moisture, and nutrients. It was noted during the study period that, the grass yield differed substantially (677.5%) between the two ends (0 and 100%) of the thinning gradient. This indicates that the negative effect of bush encroachment on grass biomass is severe at high woody density and cover. The negative effect of dense woody species on the production of grass species has already been reported globally to savanna ecologies (Ward, 2005; Eldridge et al., 2011; Keba, 2013; Archer et al., 2017). Encroachment of woody plants jeopardizes grassland biodiversity, threatens the sustainability of pastoral subsistence and commercial livestock grazing (Eldridge et al., 2011). Richter et al. (2001) found that bush encroachment had an adverse influence on grass biomass production and decreased potential grazing capacity. And the decrease in grass biomass production directly affects livestock production, on which the livelihoods of pastoralists depend (Solomon and Mblambo, 2010).
The result of the current study showed that thinning increased the biomass of the grass component. The grass biomasses recorded in the thinned plots were substantially higher compared to the 0% plot showing the apparent existence of the positive effect of scattered woody plants on grass production. The higher biomass recorded in the thinned plots might be attributed to the reduced competition for light, soil moisture, and nutrients following thinning of A. drepanolobium. Ducherer et al. (2013) and Smit and Rethman (2000) all indicated that reducing or eliminating competition from overstory trees can increase soil moisture and mineral nutrients for understory plants, and allow colonization of bare soil by herbaceous plants. The higher dry matter yield recorded in the thinned plots and in a plot where trees have been removed corresponds to Smit (2005) who reported that in semi-arid savannas higher grass biomass is recorded at low tree densities and the complete removal of the woody layer results in a substantial increase in grass production. And the increase in grass biomass improves carrying capacity and livestock production.
The highest species composition recorded during the end of the main rainy season was mainly attributed to higher rainfall and soil moisture content. It can easily be understood from the ordination map of species and season (Figure 4) when the grasses are abundantly distributed. Sankaran et al. (2005) have also indicated that the availability of soil water is an important determinant of a tree: grass ratios in arid and semi-arid savannas. Similarly, Smit and Rethman (2000) emphasized the significant positive effect that thinning has on the soil water status of an area, which enhances the establishment of highly desirable and desirable grass species. In this study, the highly desirable class of grass species was dominant in composition during the main and small rainy seasons probably attributed to the effective rainfall which increased soil moisture available for grasses following thinning suggesting that, thinning of woody plants should be planned before the onset of rainy season to improve the availability of soil moisture which would support the establishment, growth and production of desirable grass species in the semi-arid savannas.
The grass biomass yield obtained during the main rainy season was significantly higher than the yields for the other two seasons. The lower soil moisture recorded in season 2 is responsible for the lower biomass in this regard particularly during July and August, resulting in lower vegetative production of the grass layer. Therefore, climatic variability is the major contributing factor for the low biomass production in savanna ecosystems (Keba, 2013). The interactive effect of thinning and season on grass biomass was evident as indicated by the variability of biomass among thinning plots across seasons. In general, when the biomass is high as in season 1 and season 3 the thinning had a big effect, but the same effect could not be seen in season 2 due to low grass vegetative production which resulted in low biomass yield.
The total annual biomass yield of 3,301.5 ± 897 kg. ha−1 (3.3 ± 0.9 ton ha−1) documented by the current study is higher than a study survey estimate of 1.21 t. DMY ha−1 by Keba (2013) in severely bush encroached and seasonally grazed areas of the region. Similarly, Amsalu and Baars (2002) also reported a lower DM yield of 1.3 t ha−1 for all the communally grazed zones in bush encroached Mid Rift valley area of Ethiopia. In general, the higher annual grass biomass under the current study is owing to the effect of selectively thinning woody plants that combines rainfall.
Savanna is a curious vegetation state characterized by the coexistence of grasses and trees. For grasses to persist, they would have to be the superior competitor during periods of the simultaneous tree and grass growth (Scholes and Archer 1997). The same authors recognized the importance of frequent disturbances that affect competitive advantages between trees and grasses. Disturbances that limit tree dominance such as mechanical thinning and post-thinning management hypothesized to provide the mechanism that prevents trees from negatively influencing the grassy layer (Smit, 2004). In this paradigm, with the absence of these measures, savannas are expected to be unstable. This means that without disturbances, savannas will generally tend toward a wooded scenario in arid systems (Scholes and Archer 1997). The current study result showed that thinning reduced woody cover and increased species composition which in turn responsible for increased biomass production suggesting that thinning intensity, grass species composition, and biomass are strongly positively related. This means grass species composition and biomass increased with the increasing intensity of thinning woody plants. Such a positive relationship between thinning intensity and grass species has been widely reported in semi-arid savanna ecosystems (Smit, 2005; Gemedo et al., 2006; Kahumba, 2010).
To answer the question at which thinning level the ideal productivity can be reached; CA ordination illuminated the richness of the majority of the highly desirable grass species at the 50 and 75% intensities suggesting that, maximum species richness and biomass may be realized at the heavy intensity (75%) of thinning woody plants in savanna rangelands. This is further validated by the indifferent species composition and a decline in total biomass in the cleared (100%) plot during the third season. The decline might be attributed to the removed positive tree-grass interaction as a result of clearing existing woody plants suggesting that, the high grass yield recorded in the 100% plot area during the first season may be only temporary.
According to the study findings, there was a positive linear relationship between rainy seasons and total grass values showing that effective rainfall increases grass species abundance and biomass production in the natural savanna. In contrast, grass species abundance and biomass production was negatively associated with dry season indicates that climatic variability is the major contributing factor for the decline in grass species composition and low biomass production in the semi-arid savanna. The relationships between grass species composition, biomass production and precipitation in the savanna of southern Ethiopia have been found to be linear. Periods or seasons of high rainfall favor growth of grass species, while drought periods limit plants establishment and growth. Ecologists (Scholes et al., 2002; Sankaran et al., 2004) also reported that the relationships between biomass and rainfall in savannas are almost linear just as that between productivity and days of water stress (House and Hall, 2001). The relationship between season and grass species reported by the current study could therefore make the decision and program to develop the most important forage grass species much easier and effective in the savanna system.
The study of the influence of thinning and season on grass species composition and biomass was carried out in semi-arid savanna where bush encroachment is the major threat to livestock production and people’s livelihood. Thinning negatively influenced woody cover and enhanced grass species composition and biomass production. Higher grass species composition and biomass were recorded in thinned plots compared to un-thinned plots. Gaps in the woody canopy were created due to thinning affecting the availability of light and potentially increased growth of grass species and production. The relationship among thinning, species composition, and biomass was linear showing that total grass values increase with increasing intensity of thinning encroaching woody plant species. Rainfall and soil moisture also played an important role by interacting with thinning and influenced species composition and grass yields. Higher species composition and biomass were recorded in wet seasons than in the dry season. The relationships between grass species composition, biomass and precipitation in the study savanna have been found to be linear. Aridity increases rainfall variability with rainfall remains the main limiting factor of forage production for livestock in savannas.
Based on some observed qualitative benefits of thinning differentiation, with certain desirable grass species that showed preference to the thinning intensity, desirable grass species would probably be lost with the complete removal of trees. The current study is, therefore, suggests thinning of bushes to the 75% of tree density in heavily encroached grazing lands if the objective is to maximize forage production for grazers. The sustainability of woody tree thinning technology may possibly challenging given the normal grazing systems in the area, because grass is more sensitive to overgrazing than trees. Appropriate rangeland management practices must be implemented usually involving the protection of thinned areas from grazing for a period of time and provision of adequate rests are usually required during the post-thinning period to ensure sustainability. Any mismanagement practices that promoted woody plant invasion in the first place must be eliminated from the management system. Sound grazing management practices, especially during wet season, which will ensure the vigorous and competitive grasses is of critical importance. After bush thinning has been implemented, maintaining a productive grass layer through sound rangeland management should prevent renewed bush encroachment. Early treatment will minimize efforts and costs that come with treating well established plants or full-blown infestations. The best way of preventing bush encroachment, therefore, is to maintain a high cover of grasses through good management at all times.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
MH conducted the fieldwork, analyzed the data, interpreted the findings and wrote the manuscript. YW and AG initiated the idea and wrote the manuscript. XX and YY critically commented the draft. All authors fully participated in processing the manuscript. All authors have read and agreed to the final version of the manuscript.
This study received financial support from African Great Green Wall Adaptation Technical Cooperation Research and Demonstration (2018YFE0106000), Science and Technology Partnership Program, Ministry of Science and Technology of China (Grant No. KY 201702010), and International cooperation and Exchanges NSFC (Grant No. 41861144020).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors wish to thank all funding agencies for providing financial support for the success of this particular study. Authors are also very grateful to the Dida Tuyura ranch management and area experts participating in this study by providing access to their lands and sharing their knowledge.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2021.692239/full#supplementary-material
Abdallah, F., and Chaieb, M. (2013). Interactions of Acacia Tortilis with Herbaceous Vegetation in Relation with Tree Size under North African Pre Saharan Region. Pakistan. J. Bot. 45, 1715–1720.
Abule, E., Smit, G. N., and Snyman, H. A. (2005). The Influence of Woody Plants and Livestock Grazing on Grass Species Composition, Yield and Soil Nutrients in the Middle Awash valley of Ethiopia. J. Arid Environments 60, 343–358. doi:10.1016/j.jaridenv.2004.04.006
Ahmad, S., Islam, M., Bano, G., Aslam, S., and Koukab, S. (2009). Seasonal Variation in Current Season and Dead Biomass of Chrysopogon Aucheri (Boiss) STAF. And Cymbopogon Jwarancusa (JONES) SCHULT. In highland Balochistan, Pakistan. Pakistan J. Bot. 41, 519–527.
Amsalu, S., and Baars, R. M. T. (2002). Grass Composition and Rangeland Condition of the Major Grazing Areas in the Mid Rift Valley, Ethiopia. Afr. J. Range Forage Sci. 19, 161–166. doi:10.2989/1022011020948578
Angassa, A., and Oba, G. (2009). Bush Encroachment Control Demonstrations in Southern Ethiopia: 1. Woody Species Survival Strategies with Implications for Herder Land Management. Afr. J. Ecol. 47, 63–76. doi:10.1111/j.1365-2028.2007.00919.x
Angassa, A., and Oba, G. (2008). Effects of Management and Time on Mechanisms of bush Encroachment in Southern Ethiopia. Afr. J Ecol 46, 186–196. doi:10.1111/j.1365-2028.2007.00832.x
Angassa, A., and Oba, G. (2007). Relating Long-Term Rainfall Variability to Cattle Population Dynamics in Communal Rangelands and a Government Ranch in Southern Ethiopia. Agric. Syst. 94, 715–725. doi:10.1016/j.agsy.2007.02.012
Angassa, A. (2007). The Dynamics of savanna Ecosystem and Management in Borana, South Ethiopia. Norway: Norwegian University of Life Sciences, 183. Ph.D. Thesis.
Angassa, A. (2002). The Effect of Clearing Bushes and Shrubs on Range Condition in Borana, Ethiopia. Trop. Grass 36, 69–76.
Ansley, R. J., and Castellano, M. J. (2006). Strategies for savanna Restoration in the Southern Great Plains: Effects of Fire and Herbicides. Restor Ecol. 14, 420–428. doi:10.1111/j.1526-100x.2006.00150.x
Archer, S. R., Andersen, E. M., Predick, K. I., Schwinning, S., Steidl, R. J., and Woods, S. R. (2017). “Woody Plant Encroachment: Causes and Consequences,” in Rangeland Systems. Editor D. D. Briske (Springer), 25–84. doi:10.1007/978-3-319-46709-2_2
Buitenwerf, R., Bond, W. J., Stevens, N., and Trollope, W. S. W. (2012). Increased Tree Densities in South African Savannas: >50 Years of Data Suggests CO2 as a Driver. Glob. Change Biol. 18, 675–684. doi:10.1111/j.1365-2486.2011.02561.x
Chiang, J. M., Lin, K. C., Hwong, J. L., Wang, H. C., and Lin, T. C. (2012). Immediate Effects of Thinning with a Small Patch Clearcut on Understory Light Environments in a Cryptomeria Japonica Plantation in central Taiwan. Taiwan J. For. Sci. 27, 319–331. doi:10.1097/ncq.0b013e31825ba89e
Connor, O. T. G. (1995). Acacia Karoo Invasion of Grassland: Environmental and Biotic Effects Influencing Seedling Emergence and Establishment. Oecologia 103, 214–223. doi:10.1007/BF00329083
Coppock, D. (1994). “The Borana Plateau of Southern Ethiopia: Synthesis of Pastoral Research, Development and Changes 1980-1990,” in System Study No.5 (ILCA), 374.
Cossins, N. J., and Upton, M. (1987). The Borana Pastoral System of Southern Ethiopia. Agric. Syst. 25, 199–218. doi:10.1016/0308-521x(87)90020-5
D'Odorico, P., Fuentes, J. D., Pockman, W. T., Collins, S. L., He, Y., Medeiros, J. S., et al. (2010). Positive Feedback between Microclimate and Shrub Encroachment in the Northern Chihuahuan Desert. Ecosphere 1, 1–11. doi:10.1890/es10-00073.1
Desta, S., and Coppock, D. (2004). Pastoralism under Pressure: Tracking System Change in Southern Ethiopia. J. Hum. Ecol. 32, 4. doi:10.1023/b:huec.0000043516.56037.6b
Ducherer, K., Bai, Y., Thompson, D., and Broersma, K. (2013). Thinning of a Ponderosa pine/Douglas-fir forest in South-central BC: Impacts on Understory Vegetation. J. Ecosystem Manag. 14, 1, 2013. Available at: http://jem.forrex.org/index.php/jem/article/viewFile/187/486.
Eldridge, D. J., Bowker, M. A., Maestre, F. T., Roger, E., Reynolds, J. F., and Whitford, W. G. (2011). Impacts of Shrub Encroachment on Ecosystem Structure and Functioning: towards a Global Synthesis. Ecol. Lett. 14, 709–722. doi:10.1111/j.1461-0248.2011.01630.x
Ellis, J. E., and Swift, D. M. (1988). Stability of African Pastoral Ecosystems: Alternate Paradigms and Implications for Development. J. Range Manag. 41, 450–459. doi:10.2307/3899515
Gemedo, D., Brigitte, L. M., and Johannes, I. (2005). Plant Biodiversity and Ethnobotany of Borana Pastoralists in Southern Oromia, Ethiopia. Econ. Bot. 59, 43–65. doi:10.1663/0013-0001(2005)059[0043:PBAEOB]2.0.CO;2
Gemedo, D., Maass, B. L., and Isselstein, J. (2006). Encroachment of Woody Plants and its Impact on Pastoral Livestock Production in the Borana Lowlands Southern Oromia, Ethiopia. Afr. J. Ecol. 44, 237–246. doi:10.1111/j.1365-2028.2006.00638.x
Guevara, J. C., Grünwaldt, E. G., Estevez, O. R., Bisigato, A. J., Blanco, L. J., Biurrun, F. N., et al. (2009). Range and Livestock Production in the Monte Desert, Argentina. J. Arid Environments 73, 228–237. doi:10.1016/j.jaridenv.2008.02.001
Helland, J. (1997). “Social Organization and Water Control Among the Borana of Southern Ethiopia,” in Working Document 16 (Nairobi, Kenya: ILCA (International Livestock Center for Africa)), 30.
Herlocker, D. (1999). Rangeland Ecology and Resource Development in Eastern Africa. Nairobi, Kenya: GTZ.
House, J. I., and Hall, D. O. (2001). “Productivity of Tropical Savannas and Grasslands,” in Terrestrial Global Productivity: Past.
Innocent, N. S., and Munyati, C. (2014). Quantifying Savanna Woody Cover in the Field and on Historical Imagery: A Methodological Analysis. South Afr. J. Geomatics. 3, 2. doi:10.4314/sajg.v3i2.1
Joubert, D. F., Smit, G. N., and Hoffman, M. T. (2012). The Role of Fire in Preventing Transitions from a Grass Dominated State to a bush Thickened State in Arid Savannas. J. Arid Environments 87, 1–7. doi:10.1016/j.jaridenv.2012.06.012
Kahi, C. H., Ngugi, R. K., Mureithi, S. M., and Ng’ethe, J. C. (2009). The Canopy Effects of Prosopis Juliflora and Acacia Tortilis Trees on Herbaceous Plants Species and Soil Physio-Chemical Properties in Njemps Flats, Kenya. Trop. Subtrop. Agroecosystems. 10, 441–449.
Kahumba, A. (2010). Comparison of the Rehabilitative Effects of Mechanical and Chemical Methods of bush Control on Degraded highland and savanna Rangelands in Namibia. Namibia: University of Namibia, 69. M.Sc. Thesis.
Kaiser, J. (1999). FIGHTING BACK:Stemming the Tide of Invading Species. Science 285, 1836–1841. doi:10.1126/science.285.5435.1836
Kangalawe, R. Y. M. (2009). Ecosystems Changes and Implications on Livelihoods of Rural Communities in Africa. Afr. J. Ecololgy 47, 1–2. doi:10.1111/j.1365-2028.2008.01042.x
Keba, H. T. (2013). “The Impact of Changes in Land-Use and Rainfall Variability on Range Condition and Pastoral Livelihoods in the Borana Rangelands of Southern Oromia, Ethiopia,” in Thesis Submitted in the Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in Pasture Science (South Africa: University of Pretoria). Available at: https://repository.up.ac.za/bitstream/handle/2263/32981 (Accessed February 16, 2021).
Kellner, K. (2008). Studies in Land Degradation, Desertification and Restoration Ecology: Challenges and Opportunities. Potchefstroom: North-West University. 978-1-86822-566-8.
Lazarides, M., Cowley, K., and Hohnen, P. (1997). CSIRO Handbook of Australian Weeds. Australia: CSIRO.
Lehmann, C. E. R., Archibald, S. A., Hoffmann, W. A., and Bond, W. J. (2011). Deciphering the Distribution of the savanna Biome. New Phytol. 191, 197–209. doi:10.1111/j.1469-8137.2011.03689.x
Liao, C., and Fei, D. (2017). Pastoralist Adaptation Practices under Non-governmental Development Interventions in Southern Ethiopia. Rangel. J. 39, 189–200. doi:10.1071/rj16015
Ludwig, F., de Kroon, H., Berendse, F., and Prins, H. H. T. (2004). The Influence of savanna Trees on Nutrient, Water and Light Availability and the Understorey Vegetation. Plant Ecol. 170, 93–105. doi:10.1023/b:vege.0000019023.29636.92
Marshall, V. M., Lewis, M. M., and Ostendorf, B. (2012). Buffel Grass (Cenchrus Ciliaris) as an Invader and Threat to Biodiversity in Arid Environments: A Review. J. Arid Environments 78, 1–12. doi:10.1016/j.jaridenv.2011.11.005
Moleele, N. M., Ringrose, S., Matheson, W., and Vanderpost, C. (2002). More Woody Plants? the Status of bush Encroachment in Botswana's Grazing Areas. J. Environ. Manage. 64, 3–11. doi:10.1006/jema.2001.0486
Nenadi, O., and Greenacre, M. J. (2007). Correspondence Analysis in R, with Two- and Three-Dimensional Graphics: the Ca Package. J. Stat. Softw. 20, 3. doi:10.18637/jss.v020.i03
Noble, J. C. (1997). The Delicate and Noxious Scrub: Studies on Native Tree and Shrub Proliferation in Semi-arid Woodlands of Australia. Canberra: CSIRO Division of Wildlife and Ecology.
Oba, G. (1998). Assessment of Indigenous Range Management of the Borana Pastoralists of Southern Ethiopia. GTZ: Borana Lowland Pastoral Development Program. Available at: https://pdfs.semanticscholar.org/50e1/75f0378501f8da98a5a8f0dc568acc65f652 (Accessed March 5, 2020).
Oba, G., and Kaitira, L. M. (2006). Herder Knowledge of Landscape Assessments in Arid Rangelands in Northern Tanzania. J. Arid Environments 66, 168–186. doi:10.1016/j.jaridenv.2005.10.020
Oba, G., Post, E., Syvertsen, P. O., and Stenseth, N. C. (2000). Bush Cover and Range Condition Assessments in Relation to Landscape and Grazing in Southern Ethiopia. Landscape Ecol. 15, 535–546. doi:10.1023/a:1008106625096
O’Connor, T. G., Puttick, J. R., and Hoffman, M. T. (2014). Bush Encroachment in Southern Africa: Changes and Causes. Afr. J. Range Forage 31, 67–88. doi:10.2989/10220119.2014.939996
Okello, B. D., O’Connor, T. G., and Young, T. P. (2001). Growth, Biomass Estimates, and Charcoal Production of Acacia Drepanolobium in Laikipia, Kenya. For. Ecol. Manag. 142, 143–153. doi:10.1016/s0378-1127(00)00346-7
R Development Core Team (2018). A Language and enviRonment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing.
Ratajczak, Z., Nippert, J. B., and Collins, S. L. (2012). Woody Encroachment Decreases Diversity across North American Grasslands and Savannas. Ecology 93, 697–703. doi:10.1890/11-1199.1
Rau, B. M., Tausch, R., Reiner, V., Johnson, D. W., Chambers, J. C., Blank, R. R., et al. (2009). Influence of Prescribed Fire on Ecosystem Biomass, Carbon and Nitrogen in a Pinyon Juniper woodland. Rangeland Ecol. Manag. 63, 197–202. doi:10.2111/rem-d-09-00088.1
Richter, C., Snyman, H., and Smit, G. (2001). The Influence of Tree Density on the Grass Layer of Three Semi-arid savanna Types of Southern Africa. Afr. J. Range Forage Sci. 18, 103–109. doi:10.2989/10220110109485762
Roques, K. G., O'Connor, T. G., and Watkinson, A. R. (2001). Dynamics of Shrub Encroachment in an African Savanna: Relative Influences of Fire, Herbivory, Rainfall and Density Dependence. J. Appl. Ecol. 38, 268–280. doi:10.1046/j.1365-2664.2001.00567.x
Sala, O. E., and Maestre, F. T. (2014). Grass-woodland Transitions: Determinants and Consequences for Ecosystem Functioning and Provisioning of Services. J. Ecol. 102, 1357–1362. doi:10.1111/1365-2745.12326
Sang-Hyun, L., Seongjun, K., and Hyun-Jun, K. (2018). Effects of Thinning Intensity on Understory Vegetation in Chamaecyparis Obtusa Stands in South Korea. For. Sci. Tech. 14, 7–15. doi:10.1080/21580103.2017.1409661
Sankaran, M., Hanan, N. P., Scholes, R. J., Ratnam, J., Augustine, D. J., Cade, B. S., et al. (2005). Determinants of Woody Cover in African Savannas. Nature 438, 846–849. doi:10.1038/nature04070
Sankaran, M., Ratnam, J., and Hanan, N. P. (2004). Tree-grass Coexistence in Savannas Revisited - Insights from an Examination of Assumptions and Mechanisms Invoked in Existing Models. Ecol. Lett. 7, 480–490. doi:10.1111/j.1461-0248.2004.00596.x
Sankaran, M., Ratnam, J., and Hanan, N. (2008). Woody Cover in African Savannas: The Role of Resources, Fire and Herbivory. Glob. Ecol Biogeogr. 17, 236–245. doi:10.1111/j.1466-8238.2007.00360.x
Scholes, R. J., and Archer, S. R. (1997). Tree-grass Interactions in Savannas. Annu. Rev. Ecol. Syst. 28, 517–544. doi:10.1146/annurev.ecolsys.28.1.517
Scholes, R. J., Dowty, P. R., Caylor, K., Parsons, D. A. B., Frost, P. G. H., and Shugart, H. H. (2002). Trends in savanna Structure and Composition along an Aridity Gradient in the Kalahari. J. Vegetation Sci. 13, 419–428. doi:10.1111/j.1654-1103.2002.tb02066.x
Smit, G. N. (2004). An Approach to Tree Thinning to Structure Southern African Savannas for Long-Term Restoration from bush Encroachment. J. Environ. Manage. 71, 179–191. doi:10.1016/j.jenvman.2004.02.005
Smit, G. N. (2014). An Expansion of the Aboveground Biomass Quantification Model for Trees and Shrubs to Include the wood Component. Afr. J. Range Forage. Sci. 25, 199–218. doi:10.2989/10220119.2013.866161
Smit, G. N., and Rethman, F. G. (1999). The Influence of Tree Thinning on the Establishment of Herbaceous Plants in a Semi-arid savanna of Southern Africa. Afr. J. Range Forage Sci. 16, 9–18. doi:10.2989/10220119909485713
Smit, G. N., and Rethman, N. F. G. (2000). The Influence of Tree Thinning on the Soil Water in a Semi-arid savanna of Southern Africa. J. Arid Environments 44, 41–59. doi:10.1006/jare.1999.0576
Smit, G. N., Ritcher, C. G. F., and Aucamp, A. J. (1999). “Bush Encroachment: an Approach to Understanding and Managing the Problem,” in Veld Management in South Africa Chapter 10. Editor N. M Tainton (Pietermaritzburg, South Africa: University of Natal Press).
Smit, G. N. (2005). Tree Thinning as an Option to Increase Herbaceous Yield of an Encroached Semi-arid savanna in South Africa. BMC Ecol. 5, 4. doi:10.1186/1472-6785-5-4
Solomon, B. T., and Mblambo, V. (2010). Encroachment of Acacia Brevispica and Acacia Drepanolobium in Semi-arid Rangelands of Ethiopia and Their Influence on Sub Canopy Grasses. Restoration J. Bot. 5, 1–3. doi:10.3923/rjb.2010.1.13
Solomon, T., Dlamini, B. J., and Dlamini, A. M. (2010). Changes in Soil Characteristics and Grass Layer Condition in Relation to Land Management Systems in the Semi-arid Savannas of Swaziland. J. Arid Environ. 74, 675–684. doi:10.1016/j.jaridenv.2009.10.016
Teshome, A., Abule, E., and Lisanework, N. (2012). Evaluation of Woody Vegetation in the Rangeland of Southeast Ethiopia. Int. Res. J. Agri. Sci. Soil Sci. 2, 113–126. Available online: http://www.interesjournals.org/IRJAS.
Tsai, H.-C., Chiang, J.-M., McEwan, R. W., and Lin, T.-C. (2018). Decadal Effects of Thinning on Understory Light Environments and Plant Community Structure in a Subtropical forest. Ecosphere 9, e02464. doi:10.1002/ecs2.2464
Úbeda, X., and Outeiro, L. (2009). “Physical and Chemical Effects of Fire on Soil,” in Fire Effect on Soils and Restoration Strategies. Editors A. Cerdà, and P. R. Robichaud (Enfield, NH: Science Publishers), 105–132. doi:10.1201/9781439843338-c4
Van Auken, O. W. (2009). Causes and Consequences of Woody Plant Encroachment into Western North American Grasslands. J. Environ. Manage. 90, 2931–2942. doi:10.1016/j.jenvman.2009.04.023
Venter, Z. S., Cramer, M. D., and Hawkins, H.-J. (2018). Drivers of Woody Plant Encroachment over Africa. Nat. Commun. 9, 2272. doi:10.1038/s41467-018-04616-8
Ward, D., Wiegand, K., and Getzin, S. (2013). Walter's Two-Layer Hypothesis Revisited: Back to the Roots!. Oecologia 172, 617–630. doi:10.1007/s00442-012-2538-y
Ward, D. (2005). Do we Understand the Causes of bush Encroachment in African Savannas?. Afr. J. Range Forage Sci. 22, 101–105. doi:10.2989/10220110509485867
Keywords: thinning, Acacia drepanobium, grass, species composition, biomass
Citation: Hare ML, Xu XW, Wang YD, Yuan Y and Gedda AE (2021) Do Woody Tree Thinning and Season Have Effect on Grass Species’ Composition and Biomass in a Semi-Arid Savanna? The Case of a Semi-Arid Savanna, Southern Ethiopia. Front. Environ. Sci. 9:692239. doi: 10.3389/fenvs.2021.692239
Received: 07 April 2021; Accepted: 09 July 2021;
Published: 27 July 2021.
Edited by:
Stephen M. Mureithi, University of Nairobi, KenyaReviewed by:
Kevin Mganga, South Eastern Kenya University, KenyaCopyright © 2021 Hare, Xu, Wang, Yuan and Gedda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Malicha Loje Hare, bWFsaWNoYWxvamVAeWFob28uY29t; Yong Dong Wang, d2FuZ3lkQG1zLnhqYi5hYy5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.