 Sékou F. M. Coulibaly1*
Sékou F. M. Coulibaly1* Bruna R. Winck2
Bruna R. Winck2 Marthe Akpa-Vinceslas1
Marthe Akpa-Vinceslas1 Laurent Mignot1
Laurent Mignot1 Marc Legras3
Marc Legras3 Estelle Forey1
Estelle Forey1 Matthieu Chauvat1
Matthieu Chauvat1- 1Normandie Univ, UNIROUEN, IRSTEA, ECODIV, FR SCALE CNRS 3730, Rouen, France
- 2Departamento de Ecologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
- 3UniLaSalle, Aghyle Unit, FED NORVEGE 4277, Mont-Saint-Aignan, France
Soil processes such as decomposition are mainly performed by soil biota. Although soils worldwide are extremely biodiverse, the relationship between decomposers (fauna and microorganisms), and ecosystem function is poorly understood. Collembola are abundant and ubiquitous microarthropods that are found in terrestrial ecosystems. They can affect the amount of biomass and the activity of microbial communities, either directly through selectively feeding on fungi and bacteria, or indirectly by dissemination of microbial propagules, and the alteration of nutrient availability. However, despite the functional role they play in belowground food webs, the interactions between natural assemblages of soil microbes and Collembola receive little attention. This study, conducted in microcosm conditions, examines the effects of two distinct natural assemblages of functional groups of Collembola (ep- and euedaphic) upon microbial communities using PLFA markers and their associated soil functions (e.g., enzymatic activities and C mineralization rate) over a 2-month period. Our principal objective was to determine whether different functional groups of Collembola had varying effects on microbial soil community abundance, structure and activity, resulting in potentially important effects on ecosystem processes. Our findings show that the interactions of the functional groups of Collembola with microbial communities vary significantly whether they are alone or combined. A distinct response in the composition of the microbial communities was found at the end of the 2-month period. The communities were significantly different from each other in terms of PLFA marker composition. We found that the epedaphic species were related to and promoted Gram+ bacteria whereas euedaphic species were related to Gram- bacterial markers. This had further repercussions on soil function, such as nutrient recycling. Combining both functional groups did not lead to a complementary effect on soil microbial properties, with a drastically different outcome between the first and the second month of the experiment. Additional research dealing with the interactions between decomposers using natural assemblages will help to predict the functional outcomes of soil biota structure and composition.
Highlights
- Functional groups of Collembola shape soil microbial communities.
- Each functional group has a specific effect on microbial communities and associated roles.
- No complementarity was observed between epedaphic and euedaphic collembolan species
- Response of microbial communities to the presence of Collembola is time dependent.
Introduction
Decomposition, with primary production, is one of the most important ecosystem functions found in soil. Indeed, ~90% of the terrestrial net primary production (NPP) enters the soil food web to be consumed and then decomposed into mineral forms and eventually reabsorbed by plants. Decomposition processes are mainly regulated by environmental drivers including abiotic factors (e.g., water content, temperature), litter quantity and quality, and by the activity of the decomposer biota (Seastedt, 1984; Anderson, 1991; Wardle et al., 2004).
Decomposition in terrestrial ecosystems and nutrient cycling are primarily regulated by soil micro-organisms (fungi and bacteria). Although, most nutrient mineralization is governed directly by the activities of bacteria and fungi, this mineralization process is regulated by interactions with soil fauna (Bardgett and Cook, 1998; Kaneko et al., 1998; Bardgett and Shine, 1999; Mikola et al., 2002; Tiunov and Scheu, 2005; Lenoir et al., 2007; Chauvat et al., 2014). For example, several studies showed that soil food web properties strongly impact organic matter recycling and, thus, affect the quality and quantity of the nutrients available to plants (Heemsbergen et al., 2004), see Whalen et al. (2013) for a complete review on N cycling. Other studies also highlighted links between the structure and composition of soil fauna and several enzymatic activities (Sauvadet et al., 2017).
Within terrestrial soil fauna, Collembola are abundant and ubiquitous microarthropods, which feed predominantly on fungi, but also bacteria, actinomycetes, and algae (Chen, 1995). Soil Collembola can affect the biomass and activity of the microbial community, either directly through selectively feeding on fungi and bacteria, or indirectly by comminution of organic matter, dissemination of microbial propagules, and the alteration of nutrient availability (Moore et al., 1988; Verhoef and Brussaard, 1990; Lussenhop, 1992; Griffiths and Bardgett, 1997). Grazing pressure exerted by Collembola depends on invertebrate body size, population density as well as feeding preferences (Hedlund and Augustsson, 1995; Kaneko et al., 1998; Crowther and A'Bear, 2012). The interaction between soil microbes and Collembola is important because of their trophic and functional significance within belowground food webs. However, within this framework, most mechanistic studies have focused on a limited number of Collembola species, while none to our knowledge have considered natural assemblages of Collembola. Though, these microarthropods belong to a very heterogeneous group with contrasting life-forms, e.g., litter-dwelling or soil-dwelling, they occupy different soil sub-horizons. Basically, epedaphic collembolan species are large-bodied, have a high metabolic activity, consume a food substrate of high quality and are surface-dwellers. Conversely, euedaphic species are small deep-living species that consume low-quality food and have low metabolic activity. Euedaphic species are colorless with reduced appendices (e.g., furca, antennae, leg). Finally, the hemiedaphic group includes species sharing intermediate attributes (Petersen, 2002). Considering their functional traits, it is common to consider these groups as functional groups, even if we lack knowledge on how they perform or drive different functions. For example, Caravaca and Ruess (2014) showed that varied and specific grazing intensities were associated with each life-form (one species per life-form) of arbuscular mycorrhizal fungi. Other studies highlighted clear positive relationships between euedaphic collembolans and microbial biomass (Perez et al., 2013), while less obvious relationships were depicted for epedaphic ones. Furthermore, different feeding preferences of epedaphic and euedaphic species upon fungi have been highlighted (Thimm and Larink, 1995; Ponge, 2000; Nakano et al., 2017). Finally, these functional groups have been shown to express different foraging patterns toward microbial food resources (Chauvat et al., 2014). For example, intermediate levels of fungal grazing by Collembola can stimulate fungal growth and promote soil respiration, whereas overgrazing can depress fungal populations, causing a decline in rates of carbon mineralization (Anderson et al., 1981; Hedlund and Öhrn, 2000; Cortet et al., 2003; Cole et al., 2004).
To gain further insights on how different functional groups within a single decomposer taxa (i.e., Collembola) impact soil processes, we performed a microcosm experiment investigating the response of microbial communities (structurally and functionally) to different Collembola functional groups, alone or in combination. We hypothesized that (i) each Collembola functional group, due to the differences in life-history traits, has a specific impact on soil microflora, (ii) the presence of euedaphic species will generate a stronger response in soil microflora (biomass or activities) than would the presence of epedaphic species compared to a control without Collembola. Furthermore, Eisenhauer et al. (2010) showed that functionally dissimilar decomposer groups could synergistically impact soil processes. We, thus, hypothesized that (iii) complementarity may occur between different Collembola functional groups using different resources along the soil profile. Finally, as time is a determinant aspect of the outcome of biotic interactions, we investigated the response of microflora to the different Collembola treatments over a 2-month period.
Materials and Methods
Experimental Setup
The experiment was conducted under microcosm conditions in closed glass jars (9 cm diameter and 9.5 cm height). The soil, microorganisms, and Collembola come from a low intensity cow-grazing area established since 1968 and managed by the Lycée Agricole d'Enseignement Générale et Technique Agricole of Yvetot (north-west France, 49°37′04.00″N, 0°45′18.76″E). The climate of the region is temperate oceanic, with an average annual temperature and rainfall of 10°C and 800 mm, respectively. The original soil was classified as Neoluvisol-Luvisol (pH water = 6.1, clay = 15%, silt = 65%, sand = 20%, total carbon = 10.40 g kg−1, total nitrogen = 1.04 g kg−1; IUSS, 2006) and supported a vegetation dominated by Agrostis capillaris (L.), Lolium perenne (L.), and Ranunculus acris (L.).
Substrate
The substrate in the microcosm consisted of a mixture of 1 part sand to 5 parts soil. The soil was collected at a depth of 0–15 cm and sieved through a 5 mm mesh. To eliminate the original fauna and microflora, the substrate was autoclaved with two cycles of 105°C at 48 h intervals. Subsequently, the substrate was dried at 105°C and aliquots of the substrate were sampled to determine the soil water holding capacity. Finally, each microcosm was filled with 150 g of dry soil and soil suspension was used to adjust the soil moisture to 70% of the soil water-holding capacity.
Extraction and Inoculation of Microorganisms
Microorganisms were extracted from soil broth filtrates, prepared by weighing 500 g of sieved fresh soil and dissolved with 2 L of physiological water (0.85% NaCl), according to the protocol of Eisenhauer et al. (2009). A volume of 37 ml of microbial filtrate (i.e., microbial suspension) was inoculated into each microcosm. In order to establish the microbial community, the microcosms were then incubated at a temperature of 25°C for 10 days.
Collembola Extraction and Composition of Treatments
Collembola were extracted from soil monoliths using the Berlese-Tullgren device (Tullgren, 1918). In order to select and sample the two functional groups of Collembola (epedaphic or euedaphic), either the top 2 cm or the bottom 4 cm of the monoliths (10 cm depth) were placed in the Berlese-Tullgren device. The Collembola were placed into pots filled with moist plaster, and were then transferred 10 days after microbial inoculation with pooters into the microcosms to establish four different treatments: a control without Collembola, a treatment with only epedaphic Collembola (“Ep”), a treatment with only eudedaphic species (“Eu”), and a treatment with both epedaphic and euedaphic species (“Ep + Eu”). To efficiently set up the different treatments, Collembola were sorted out under binocular before being transferred with the pooters into the different microcosms. During this phase, Collembola were believed to be epedaphic based on three morphological criteria (Petersen, 2000): presence of pigmentation, presence of a large patch of ocelli, and presence of a well-developed jump organ: the furca (i.e., the mucro of the furca ending beyond the end of the abdomen). Alternatively, individuals were considered as euedaphic if they were not pigmented, had no ocelli and no furca observable at the binocular. Each treatment was replicated 16 times resulting in a total of 64 microcosms. Collembola addition to the microcosms resulted in different numbers of epedaphic or euedaphic individuals, reflecting differences occurring in natural conditions. We were more interested in the functional importance of each group (relying on natural abundances or either epedaphic or euedaphic collembolans) rather than the functional identity of each group of Collembola. The microcosms were closed and stored in a climate chamber at 18°C, with a daily light/dark cycle of 12/12 h for 60 days. The microcosms were opened every week to aerate and adjust soil moisture. The influence of Collembola on microbial variables was assessed 1 and 2 months after Collembola inoculation. At the end of the experiment, Collembola were extracted from 4 of the microcoms to check functional collembolan assemblages in each treatment (see Table 1).

Table 1. Mean (and standard deviation) densities of Collembola (number of individuals) in each treatment after 2 months.
At each sampling period of 1 and 2 months, 32 microcosms (eight replicates per treatment) were dismantled.
Microbial Community Structure
At each sampling period, the microbial community structure was determined by Phospholipid fatty acid analysis (PFLA) and microbial activity by potential C mineralization rate and by enzymatic activities.
PLFA extraction and analysis was performed using a modified protocol from Frostegård and Bååth (1996) (see details in Supplementary Material). The results were expressed as nmol PLFA g−1 dry soil. We used the bacterial acid methyl ester (BAME) 26 Mix of Sigma-Aldritch as a reference and further in the analyses only considered those identified markers and did not take into account un-recognized peaks, as they accounted for < 1% in area in our chromatograms compared to the whole set of the BAME 26 mix reference. We retained the following 16 PLFAs as indicators of the microbial community structure: branched and saturated PLFAs i15:0, a15:0, i16:0, and i17:0 (Gram+ positive bacteria); mono-unsaturated and cyclopropyl PLFAs 16:1ω7c (16:1ω9), cy17:0, (Gram-negative bacteria) 18:1ω9c, 18:1ω9t, 18:2ω6,9 (fungi), 10me-16:0 and 10me-18:0 (Actinobacteria), and, lastly, 14:0, 15:0, 16:0, 17:0, 18:0 (general indicators).
We assessed the potential Carbon mineralization rate of the microorganisms by measuring CO2 evolution at both 30 and 60 days after the introduction of Collembola (Anderson and Domsch, 1978). Twenty grams of soil under controlled conditions (28°C and initial sample humidity) for a period of 10 days in hermetic pots. CO2 released was captured by NaOH (0.2 M) and measured using a conductivity meter (ThermoScientific, Orion 011007; see Perez et al., 2013). We also measured three enzymatic activities related to C and N cycles. Beta-glucosidase activity (C cycle), Urease (N cycle), and Fluorescein DiAcetate (FDA) hydrolysis (measures a wide spectrum of enzymes, such as proteases, lipases, and esterases). FDA is a method of measuring overall activity potential. Beta-glucosidase was evaluated by Eivazi and Tabatabai method 1988, using the p-nitrophenyl-beta-glucopyranoside (pNPG) as a substrate (see Supplementary Material for details). Urease activity, an enzyme linked to the conversion of the amine (NH2) to ammonium (NH4+), was measured by determining the amount of ammonium released during incubation (2 h at 37°C) following Kandeler and Gerber (1988) (see Supplementary Material for details). The FDA activity was determined according to the method of Schnürer and Rosswall (1982) (see Supplementary Material).
Statistical Analyses
Statistical analyses were performed on the data collected for each time period (Month 1 and 2). Normality and homoscedasticity of the data were tested (Shapiro-Wilk and Bartlett, α = 0.05) to decide whether to use parametric tests (the data met both assumptions) or non-parametric tests (at least one of the assumptions was not met). Analysis of Variance (ANOVA) was used to examine the effect of treatments on measured variables over the whole course of the experiment. Single groups (“Ep” & “Eu”), their combination (“Ep + Eu”), and the control treatment were taken as single treatments in these analyses, as performed in Cragg and Bardgett (2001). Therefore, we used the following model y~Treatment and not y~EP*EU. Regarding biological factors, in our case Collembola introduced in microcosms, it is almost impossible to account for initial numbers and trace their survival during the experiment. Therefore, the combination of the treatments could have not been properly controlled in order to assess interactive effects. Tukey's HSD (honestly significant difference) was used to identify means that were significantly different at the 5% level. We performed analyses to quantify changes between the two sampling periods for a single treatment. Means between the 2 months were compared with Pairwise t-tests or Wilcoxon. Significance was tested with α = 0.05. Furthermore, to evaluate how the temporal aspect might or might not influence our results, we performed a second analysis with “treatments” as a fixed factor and “time” as a random factor. However, as trajectories of biological communities in closed and simplified environment are surely biased compared to natural system, we interpret them with caution, and rather focused on differences between treatments and the control at each sampling period.
In order to summarize/visualize the effect of Collembola life forms on the 20 PLFA markers, we performed a principal component analysis (PCA) for each sampling period (1 and 2 months). Prior to analysis data were z-transformed and submitted to Hellinger transformation (Legendre and Gallagher, 2001). We added microbial activity (CO2) and the three enzymatic activities as supplementary passive variables.
Finally, the hypothesis of no difference in PLFA marker assemblages between treatments was tested using one-way ANOSIM based on the Bray Curtis dissimilarity distance configuration (Clarke et al., 2006). If two groups of treatments are different in their PLFA marker assemblages, then compositional dissimilarities between the groups ought to be greater than those within the groups. ANOSIM was performed with 10,000 permutations and Bonferroni's correction was applied a posteriori. In the case of significant results, SIMPER analysis based on the Bray-Curtis dissimilarity distance was run to determine the PLFAs that contribute most in differentiating the two groups tested by ANOSIM.
Except ANOSIM and SIMPER analyses performed with the free PAST 3.14 software, all statistics were performed in R 3.1.2 (R Core Team, 2013) using the Rstudio (RStudio Team, 2015). The following libraries were used: ade4, lme4, MuMIn (Barton, 2013), multcomp (Hothorn et al., 2008), car (Fox and Weisberg, 2011), and vegan (Oksanen et al., 2013).
Results
PLFA
Total PLFAs differed significantly between treatments at each sampling period (Figure 1). After the first month, “Ep” treatment positively influenced total PLFA with higher values (1,240 nmol g−1 of dry soil, on average) compared to the control (1,052 nmol of g−1 of dry soil, on average). In contrast, we found lower values of total PLFAs in the “Ep + Eu” treatment (350 nmol g−1 dry soil, on average) compared to all other treatments. Finally, the “Eu” treatment had intermediate values of total PLFAs between the “Ep” and the control treatments. After 2 months, all treatments with Collembola (“Ep,” “Eu,” and “Ep + Eu”) significantly and positively influenced total PLFAs (mean value of 892, 874, and 988 nmol g−1 dry soil, respectively) compared to the control treatment without Collembola (627 nmol g−1 dry soil, on average). No difference was noticed between the different functional group treatments. Finally, among all the experimental treatments, only the “Ep + Eu” treatment showed higher values of total PLFA at the second sampling period compared to the first one, with an almost 3-fold increase (Figure 1).
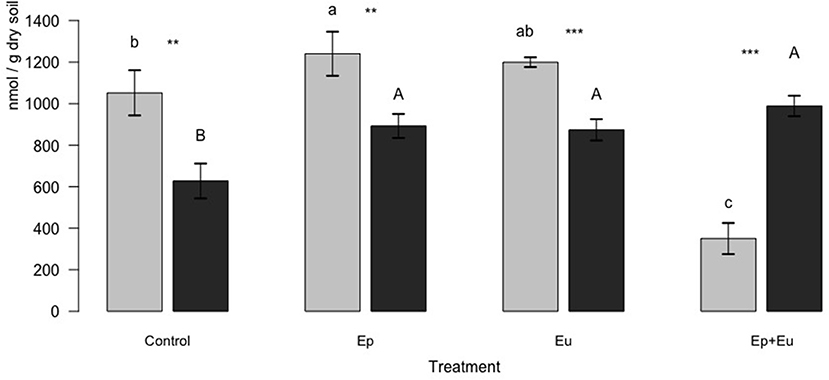
Figure 1. Total PLFA (mean ± standard deviation) of four experimental treatments 1 and 2 months after Collembola re-inoculation. Gray bar-plot = first month, black bar-plot = second month. Different minuscule letters (i.e., “a”, “b”, “c”) indicate significant differences (α = 0.05) between treatments for first month. Different capital letters (i.e., “A,” “B”) indicate significant differences (α = 0.05) between treatments for second month. The asterisks indicate significant differences between the months for a given treatment (***p < 0.001, **p < 0.01). Control, without Collembola; Ep, Epedaphic Collembola; Eu, Euedaphic Collembola; Ep+Eu, mixed functional groups (n = 4).
Regarding the ratio of bacterial to fungal PLFA markers, no difference was observed between all the treatments at both sampling periods, except for “Ep + Eu” that significantly increased the bacterial/fungal ratio during the 2 months of experimentation (+23.5%; Table 2).
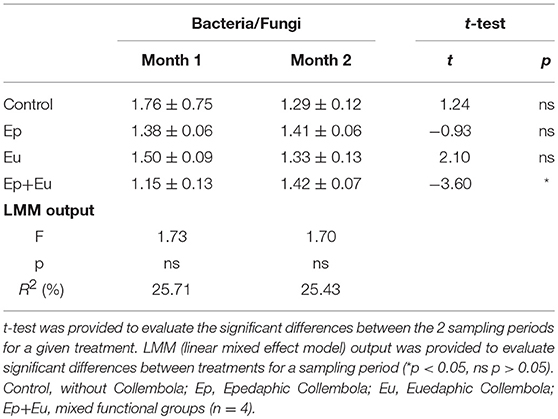
Table 2. Ratio of bacterial to fungal PLFA (mean ± standard deviation) of four experimental treatments 1 and 2 months after Collembola re-inoculation.
The Gram+ and Gram– bacterial PLFA markers and their ratios (Gram+/Gram–) differed significantly between treatments on the two sampling periods (Table 3). During the first month, only the “Ep + Eu” treatment had a negative influence on the PLFA markers. After 2 months, the pattern changed, the “Ep” and “Ep + Eu” treatments had higher concentrations of Gram+ bacteria compared to both the “Eu” and control treatments. In parallel, after 2 months, “Eu” had a higher concentration of Gram- bacteria and a higher ratio of Gram+/Gram– compared to both “Ep” and “Ep + Eu,” the control being intermediate. Between the two sampling periods, the amount of PLFA Gram+ significantly decreased in all treatments (by 1.2 to 1.6 times) except in “Ep + Eu” where it increased by 3.2 times. The amount of PLFA Gram– significantly decreased by more than 2-fold in the control and in “Ep” between the first and second month. Finally, the ratio Gram+/Gram– decreased significantly in “Ep” and in “Ep + Eu” between the two dates.
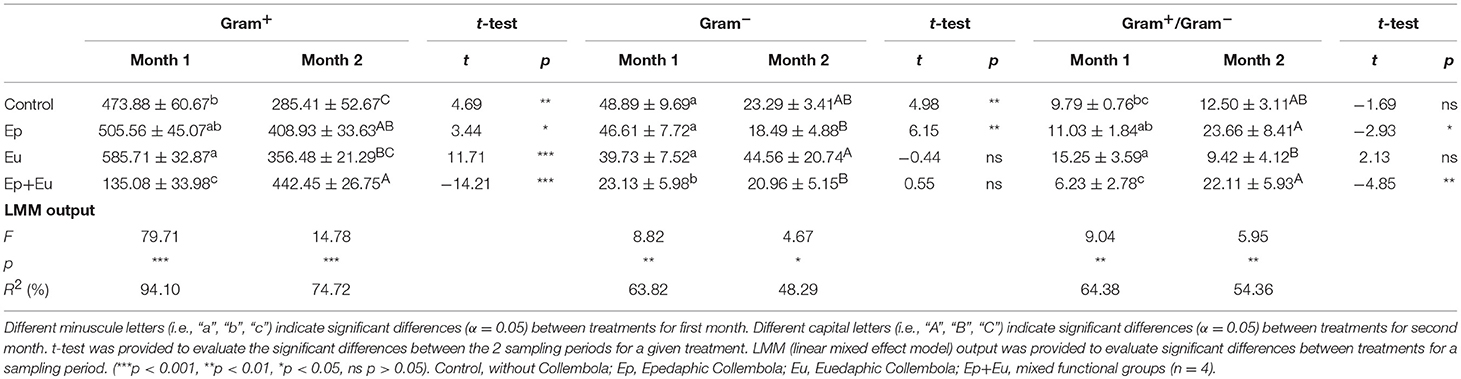
Table 3. Gram+ and Gram− bacterial PLFA and their ratios (mean ± standard deviation) for four experimental treatments 1 and 2 months after Collembola re-inoculation.
When considering the temporal aspect as a random factor, besides Gram+ bacterial PLFA markers and the ratio of bacterial to fungal PLFA markers, all other PLFA variables significantly differed between the treatments (Appendix A Table A.1). The total PLFA, bacterial PLFA, fungal PLFA, the Gram+ bacterial PLFA markers were all significantly higher in the “Ep” and the “Eu” treatments compared to the “Ep + Eu” treatment; the control showing intermediates values. The amount of PLFA Gram– had almost the same pattern being significantly higher in “Eu” compared to “Ep + Eu,” the two other treatments showing intermediates values. Including time as a random factor drastically increased the explained variance compared to a model only including the fixed factor “treatment” for two variables: PLFA gram– and the ratio of bacterial to fungal PLFA markers (Appendix A Table A.1).
Microbial Activities
Significant differences in released CO2 were observed between treatments for both sampling periods. During the first sampling period, almost twice the amount of CO2 was released in the control and “Ep” treatments compared to the “Eu” and “Ep + Eu” treatments. During the second sampling period, released CO2 was much lower for all treatments. However, it was still significantly higher in all treatments containing Collembola than in the control (Figure 2).

Figure 2. CO2 release (mean ± standard deviation) of four experimental treatments 1 and 2 months after Collembola re-inoculation. Gray bar-plot = first month, black bar-plot = second month. Different minuscule letters (i.e., “a,” “b”) indicate significant differences (α = 0.05) between treatments for first month; Different capital letters (i.e., “A,” “B”) indicate significant differences (α = 0.05) between treatments for second month. The asterisks indicate significant differences between the months for a given treatment (***p < 0.001, ns p > 0.05). Control, without Collembola; Ep, Epedaphic Collembola; Eu, Euedaphic Collembola; Ep+Eu, mixed functional groups (n = 4).
In terms of enzymatic activity, for the first sampling period, the FDA activity was significantly higher (55–92.5%) in all treatments containing Collembola, either alone or in combination, than in the control (Figure 3). This pattern changed radically after 2 months. Significantly higher FDA activity was found in the control than in the “Eu” and the “Ep + Eu” treatments (Figure 3). The “Ep + Eu” treatment was also significantly different from single functional group treatments with a lower value of FDA. Overall, the FDA activity decreased from −14% in “Ep” to −70% in “Ep + Eu” during the course of the experiment except for the control treatment where it increased by 55%.

Figure 3. FDA activity (mean ± standard deviation) of four experimental treatments 1 and 2 months after Collembola re-inoculation. Gray bar-plot = first month, black bar-plot = second month. Different minuscule letters (i.e., “a,” “b”) indicate significant differences (α = 0.05) between treatments for first month; Different capital letters (i.e., “A”, “B”, “C”) indicate significant differences (α = 0.05) between treatments for second month. The asterisks indicate significant differences between the months for a given treatment (***p < 0.001, **p < 0.01, ns p > 0.05). Control, without Collembola; Ep, Epedaphic Collembola; Eu, Euedaphic Collembola; Ep+Eu, mixed functional groups (n = 4).
The urease activity only differed between treatments during the first sampling period, with a higher mean value in the “Ep” treatment compared to all other treatments (Table 4). In opposite, the beta-glucosidase activity only differed between treatments during the second sampling period, with a 2-fold higher activity in the control and the “Ep” treatments compared to the “Eu” and the “Ep + Eu” treatments (Table 4).
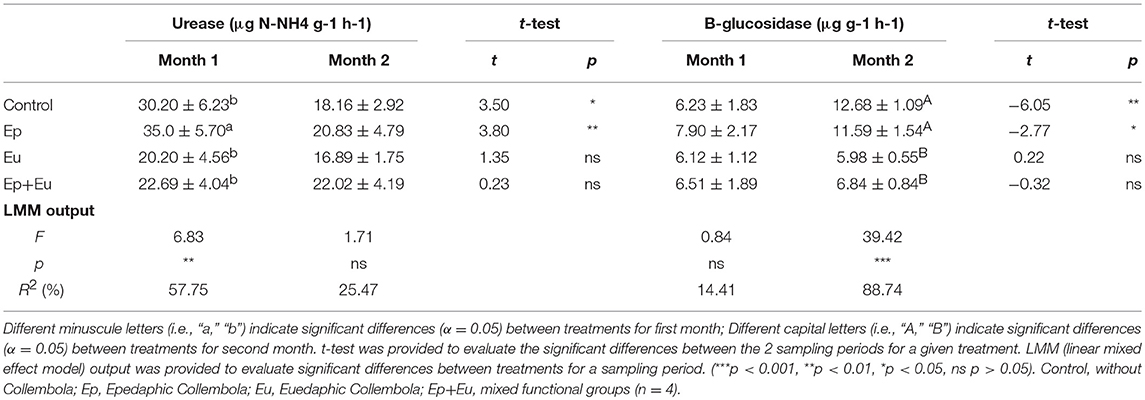
Table 4. Urease and beta-glucosidase activities (means ± standard deviation) of four experimental treatments in 1 and 2 months after Collembola re-inoculation.
When considering the temporal aspect as a random factor, all variables but FDA activity significantly differed between the treatments (Appendix A Table A.1). The released CO2 was higher in “Ep” compared to both “Eu” and “Ep + Eu.” The beta-glucosidase activities were higher in the Control and in “Ep” compared to “Eu” and “Ep + Eu.” Finally, Urease activity was higher in “Ep” compared to “Eu.” Including the temporal aspect as a random factor led to obtain a much higher explained variance for all the variables related to microbial activity (Appendix A Table A.1).
Interdependence of Collembola and Microbial Community
For the first sampling period, the proportion of the total variance in the PCA (Figure 4), which is explained by all PLFA markers, is 60%. The first-third axes accounted for 49, 18, and 10% of the variance, respectively. The first axis clearly separated “Ep + Eu” from all other treatments, while the second axis separated the two single functional groups “Ep” and “Eu.” “Ep + Eu” was related to higher values of C18.2, C11, C17D, and high values of FDA activity. “Ep” was related to higher values of CO2 and urease activity and high concentrations of C16.1.9, C18. Lastly, “Eu” was related to C15a and C15i.
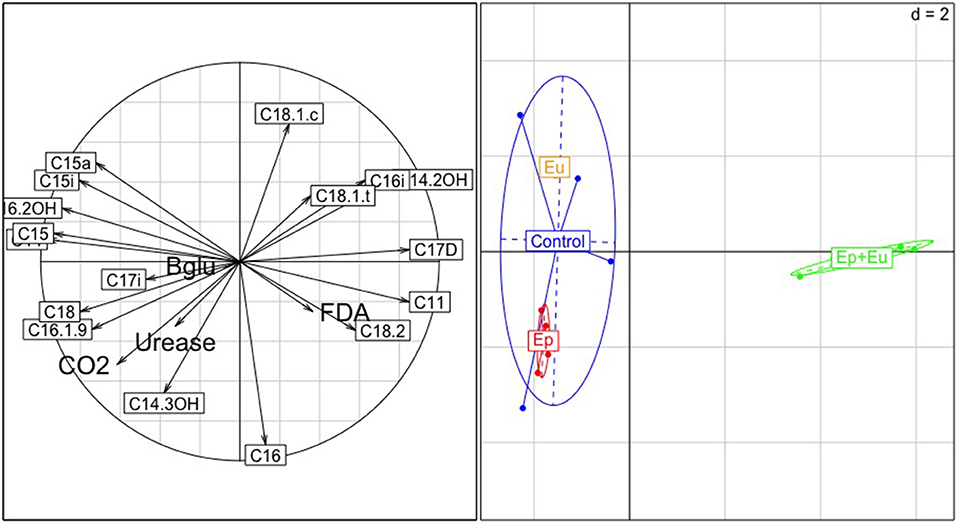
Figure 4. Principal Component Analysis (PCA) of soil microbial parameters 1 month after Collembola re-inoculation. Correlation circle with the variables is on the left; and projection of the treatments on the right panel. The centroids are the barycenters of each treatment and each ellipse represents a 95% confidence interval. Control, without Collembola; Ep, Epedaphic Collembola; Eu, Euedaphic Collembola; Ep+Eu, mixed functional groups; FDA, FDA activity; Urease, Urease activity; CO2, CO2 activity.
For the first sampling period, the ANOSIM analyses (Table 5) showed, that PLFA assemblages found in the “Ep +Eu” were significantly different from the communities found in the control or “Ep,” or “Eu” treatments. Furthermore, “Eu” and “Ep” were also significantly different from each other. This pattern was due to the presence of five PLFA markers, C16, C15a, C16i, C15i, C18.1.c (cf. Appendix A Table A.2).
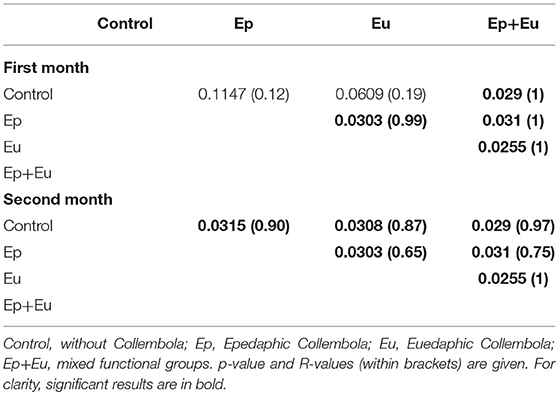
Table 5. Results of ANOSIM analyses between PLFA compositions 2 months after Collembola re-inoculation of four treatments.
For the second sampling period, the total variance in the PCA (Figure 5), which is explained by all PLFA markers, is 75%. The first-third axes accounted for 35, 24, and 12% of the variance, respectively. The first axis separated the control from all other treatments. The second axis separated the “Eu” treatment from the treatments containing the epedaphic species “Ep” and “Ep + Eu.” “Eu” was related to higher values of C18 and C16.1.9 (General indicators and bacterial Gram–, rather r-strategists). “Ep” and “Ep + Eu” were correlated, with higher values of C15i, C17i (bacterial Gram+ that are rather k-strategists). The control was variable and was correlated to higher values of C17D (bacterial Gram-), C15a, C16i (bacterial Gram+), C18.1.c, C18.2, and C18.1.t (fungal indicators). Differences between assemblages of PLFA markers were stronger during the second sampling period. The assemblages in all of the experimental treatments differed significantly from each other. Lastly, a different set of PLFA markers contributed to dissimilarity (cf. Appendix A Table A.3).
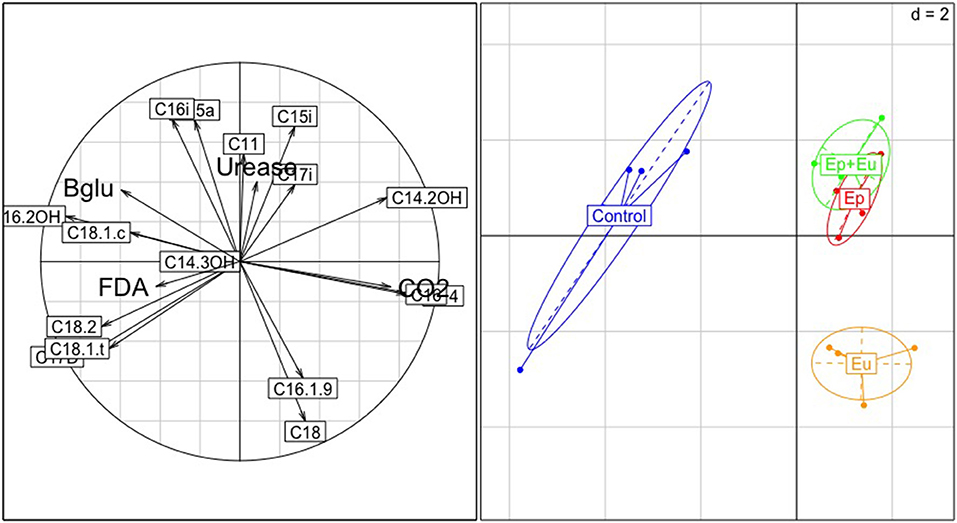
Figure 5. Principal Component Analysis (PCA) of soil microbial parameters during the second month after Collembola re-inoculation. Correlation circle with the variables is on the left; and projection of the treatments on the right. Control, without Collembola; Ep, Epedaphic Collembola; Eu, Euedaphic Collembola; Ep+Eu, mixed functional groups; FDA, FDA activity; Urease, Urease activity; CO2, CO2 activity.
Finally, the treatments containing Collembola (“Ep,” “Eu,” and “Ep + Eu”) were more correlated with higher values of CO2 and urease activity, while the control treatment was correlated with beta-glucosidase and FDA activities.
Discussion
Our results clearly demonstrate that the presence of Collembola drives the trajectories of soil microbial communities over time. Furthermore, categorizing the functional identity of Collembola assemblages is an important key to explaining the nature and intensity of microflora responses.
At the conclusion of the experiment, all treatments with Collembola promoted microorganism biomass (all PLFAs) to the same extent as the control. However, a clear difference in abundance of PLFA markers in the various treatments with Collembola, demonstrated that each functional group of Collembola did impact the structure of the microbial assemblages, but not necessarily the biomass. Microbial communities and activities were both affected by Collembola, often in different ways. Overall a coarse value like respiration indicates overall higher metabolic activity with the presence of Collembola. Our design does not allow us to disentangle the role of the abundance and identity of each functional groups upon microbial community. However, our study dealt with natural assemblages of either epedaphic or euedaphic Collembola, and therefore gives insights on how microbial communities respond to these groups representatives of natural conditions in terms of both species and abundances.
Our second hypothesis was that the presence of euedaphic species would generate a stronger response in soil microflora than would the presence of epedaphic species due to their closer link to the soil microbial compartment (Perez et al., 2013). In general, our results showed that euedaphic species (“Eu”) did not induce a stronger response in microbial communities, but rather a different response from the epedaphic species (“Ep”) compared to the control. This differentiation of response clearly appeared in the composition of the microbial communities at the end of the 60-day experiment with a significant difference in PLFA marker composition. In a recent study, a single euedaphic Collembola species (Protaphorura armata) was shown to reduce the Gram+/Gram– ratio after 20 days (Maboreke et al., 2017). Although Collembola are depicted as fungivores, with many studies highlighting strong linkages between fungi and Collembola species, especially euedaphic (Gange, 2000; Jørgensen et al., 2005; A'Bear et al., 2012), in our study, the bacterial/fungal PLFA marker ratio did not vary across treatments. Using PLFA markers, Kutáková et al. (2018) found a similar pattern that showed a stronger impact of three sympatric species of Collembola upon bacterial communities than upon fungal communities. This indicates a stronger interaction between all collembolan functional groups and soil bacteria than originally thought. The fact that epedaphic and euedaphic species led to different assemblages of PLFA markers supports the differentiation of niches between those functional groups. Furthermore, the differentiation in the composition of microbial communities observed in presence of Collembola and, also, between the “Ep” and “Eu” treatments impacted soil functions such as nutrient recycling (Kaneko et al., 1998; Chauvat et al., 2014). Unlike FDA and beta-glucosidase activities, at the end of the experiment, microbial and urease activity (linked to the N cycle) were the highest in Collembola treatments. This is consistent with the results found by Cragg and Bardgett (2001) with a positive effect of three distinct Collembola species on both microbial activity and leaching of nitrate. These authors showed that after 70 days, microbial activity and nitrate release were significantly higher in microcosms containing Collembola compared to a defaunated control. As suggested earlier, positive effects of Collembola on ecosystem processes are likely to be indirect. Though, a positive effect of their feeding on the activity of microorganisms is an increase in enzymatic activities and excretion of nutrients (Visser et al., 1981; Bardgett et al., 1993). Furthermore, our study revealed a temporal change in soil processes associated with microbial communities under the influence of Collembola. After 1 month, potential C mineralization was strongly reduced in the “Eu” treatment compared to the control or the “Ep” treatment. This fits our initial hypothesis, with a strong link between euedaphic species and microbial assemblages. We do not know the mechanism causing the difference in C mineralization. Other than urease, none of the other parameters (i.e., PLFA, enzymatic activities) showed a difference between “Ep” and “Eu” treatments after 1 month. We have shown that there is a temporal impact on community differences, demonstrating strong driving forces exerted by the different Collembola assemblages, even if the microcosms do not have resources (e.g., root exudates) coming from primary producers. As exemplified in other studies, positive effects of Collembola on belowground processes after 2 months of experimentation may have consequences on aboveground systems through plant growth or phenology (Forey et al., 2015) or on plant soil feedbacks (Kutáková et al., 2018).
We also aimed to explore the functional complementarity effect by combining collembolan functional groups. Surprisingly, the combination of functional groups (“Ep + Eu”) did not always further promote the microbial community structure, composition, or activities. One month after the beginning of the experiment, treatments with a single Collembola functional group significantly promoted the total PLFAs compared to the defaunated treatment, the “Ep + Eu” treatment strongly inhibited it. Previous studies demonstrated that Collembola trigger compensatory growth of the fungi on which they graze (Hanlon, 1981; Hedlund et al., 1991), but the outcome of this interaction is largely dependent on the species composition and population density of the fungivores, with high Collembola densities hampering microbial biomass (Ek et al., 1994; Mikola and Setälä, 1998). We may thus hypothesize that adding natural assemblages of both “Ep” and “Eu” together led to an important top-down regulation on microbial communities. This pauperization of the microbial communities led in parallel to a decrease of CO2 release and urease activity. In the same way, PCA ordination of the experimental treatments over the first month on each PLFAs marker revealed a clear differentiation between Control, “Ep” and “Eu” on one side and “Ep + Eu” on the other side of the second axis.
As previously mentioned, differentiation between treatments varies according to the time elapsed since the beginning of the experiment. The “Ep + Eu” treatment was more similar to “Ep” treatment than to the “Eu” treatment, suggesting a dominant role of epedaphic species in the combined treatment. This does not support our hypothesis of a complementarity between the functional groups of Collembola. However, this study was conducted under controlled microcosm conditions with all the limitations resulting from this experimental design. for example, microcosms may artificially increase the interactions between epedaphic and euedaphic individuals, probably slightly modifying our complementarity results. However, our study still showed how both functional groups may directly or not interact to drive the microbial community. Overall, we need to be very cautious when extrapolating results from microcosm experiments to field situation, especially regarding the temporal aspect (Carpenter, 1996). Despite these limitations of not adequately reproduce environmental ecosystems, our microcosms offered the opportunity, based on a simplified system, to focus on processes or mechanisms at fine spatial and temporal scale to better understand relationships between soil organisms.
The fact that the influence of Collembola can vary depending on their ecological traits is an important finding for soil food web and interaction web research, as was the virtual lack of complementarity observed between the two functional groups investigated. Furthermore, although Collembola are thought to be primarily fungivores, they largely influenced composition of bacterial-related PLFAs, supporting the emerging view of strong indirect or non-trophic interactions between Collembola and soil bacterial communities.
Author Contributions
MC and SC formulated the initial idea. SC, BW, MC, and EF designed the experiment. SC and BW conceived and performed the experiment. SC and BW collected the data including microbial analyzes. SC, MA-V, and LM conducted the PLFA analyzes. SC, BW, and ML conducted the enzymatic activities analyzes. SC performed the statistical analyzes. SC, MC, BW, and EF wrote the manuscript. Additionally, MC supervised the study.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was funded through the Agrobiof project (GRR VASI – Région Haute-Normandie, France). SC obtained a grant from the Région Haute-Normandie, France through the GRR TERA – SCALE. As authorized by the University of Rouen-Normandie, several parts of this manuscript were extracted from the thesis manuscript of SC (Coulibaly, 2018). The authors are grateful to Philippe Delporte, Caroline Dubois, and Lisa Castel for technical assistance and Benoit Richard for fruitful discussions on statistical analyses.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2019.00052/full#supplementary-material
References
A'Bear, A. D., Boddy, L., and Hefin Jones, T. (2012). Impacts of elevated temperature on the growth and functioning of decomposer fungi are influenced by grazing Collembola. Glob. Chang. Biol. 18, 1823–1832. doi: 10.1111/j.1365-2486.2012.02637.x
Anderson, J. M. (1991). The effects of climate change on decomposition processes in grassland and coniferous forests. Ecol. Appl. 1, 326–347. doi: 10.2307/1941761
Anderson, J. P. E., and Domsch, K. H. (1978). A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 10, 215–221. doi: 10.1016/0038-0717(78)90099-8
Anderson, R. V., Coleman, D. C., and Cole, C. V. (1981). “Effects of saprotrophic grazing on net mineralization,” in Terrestrial Nitrogen Cycles, Vol. 33, eds F. E. Clark and T. Rosswall (Stockholm: Ecological Bulletins), 201–215.
Bardgett, R. D., and Cook, R. (1998). Functional aspects of soil animal diversity in agricultural grasslands. Appl. Soil Ecol. 10, 263–276. doi: 10.1016/S0929-1393(98)00125-5
Bardgett, R. D., and Shine, A. (1999). Linkages between plant litter diversity, soil microbial biomass and ecosystem function in temperate grasslands. Soil Biol. Biochem. 31, 317–321. doi: 10.1016/S0038-0717(98)00121-7
Bardgett, R. D., Whittaker, J. B., and Frankland, J. C. (1993). The effect of collembolan grazing on fungal activity in differently managed upland pastures: a microcosm study. Biol. Fertil. Soils 16, 255–262. doi: 10.1007/BF00369301
Barton, K. (2013). MuMIn: Multi-Model Inference. R package version 1.9. 13. Vienna: The Comprehensive R Archive Network (CRAN).
Caravaca, F., and Ruess, L. (2014). Arbuscular mycorrhizal fungi and their associated microbial community modulated by Collembola grazers in host plant free substrate. Soil Biol. Biochem. 69, 25–33. doi: 10.1016/j.soilbio.2013.10.032
Carpenter, S. R. (1996). Microcosm experiments have limited relevance for community and ecosystem ecology. Ecology 77, 677–680. doi: 10.2307/2265490
Chauvat, M., Perez, G., and Ponge, J.-F. (2014). Foraging patterns of soil springtails are impacted by food resources. Appl. Soil Ecol. 82, 72–77. doi: 10.1016/j.apsoil.2014.05.012
Chen, B. (1995). Food preference and effects of food type on the life history of some soil Collembola. Pedobiologia 39, 496–505.
Clarke, K. R., Somerfield, P. J., and Chapman, M. G. (2006). On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray–Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 330, 55–80. doi: 10.1016/j.jembe.2005.12.017
Cole, L., Dromph, K. M., Boaglio, V., and Bardgett, R. D. (2004). Effect of density and species richness of soil mesofauna on nutrient mineralisation and plant growth. Biol. Fertil. Soils 39, 337–343. doi: 10.1007/s00374-003-0702-6
Cortet, J., Joffre, R., Elmholt, S., and Krogh, P. H. (2003). Increasing species and trophic diversity of mesofauna affects fungal biomass, mesofauna community structure and organic matter decomposition processes. Biol. Fertil. Soils 37, 302–312. doi: 10.1007/s00374-003-0597-2
Coulibaly, S. F. M. (2018). Effets des Changements de Pratiques Agronomiques sur la Diversité des vers de Terre et Collemboles-Conséquences sur Leurs Fonctions Associées. Doctoral dissertation, Normandie Université.
Cragg, R. G., and Bardgett, R. D. (2001). How changes in soil faunal diversity and composition within a trophic group influence decomposition processes. Soil Biol. Biochem. 33, 2073–2081. doi: 10.1016/S0038-0717(01)00138-9
Crowther, T. W., and A'Bear, A. D. (2012). Impacts of grazing soil fauna on decomposer fungi are species-specific and density-dependent. Fungal Ecol. 5, 277–281. doi: 10.1016/j.funeco.2011.07.006
Eisenhauer, N., Hörsch, V., Moeser, J., and Scheu, S. (2010). Synergistic effects of microbial and animal decomposers on plant and herbivore performance. Basic Appl. Ecol. 11, 23–34. doi: 10.1016/j.baae.2009.11.001
Eisenhauer, N., König, S., Sabais, A. C. W., Renker, C., Buscot, F., and Scheu, S. (2009). Impacts of earthworms and arbuscular mycorrhizal fungi (Glomus intraradices) on plant performance are not interrelated. Soil Biol. Biochem. 41, 561–567. doi: 10.1016/j.soilbio.2008.12.017
Eivazi, F., and Tabatabai, M. A. (1988). Glucosidases and galactosidases in soils. Soil Biol. Biochem. 20, 601–606. doi: 10.1016/0038-0717(88)90141-1
Ek, H., Sjögren, M., Arnebrant, K., and Söderström, B. (1994). Extramatrical mycelial growth, biomass allocation and nitrogen uptake in ectomycorrhizal systems in response to collembolan grazing. Appl. Soil Ecol. 1, 155–169. doi: 10.1016/0929-1393(94)90035-3
Forey, E., Coulibaly, S. F. M., and Chauvat, M. (2015). Flowering phenology of a herbaceous species (Poa annua) is regulated by soil Collembola. Soil Biol. Biochem. 90, 30–33. doi: 10.1016/j.soilbio.2015.07.024
Fox, J., and Weisberg, S. (2011). An R companion to applied regression. Los Angeles, CA: Sage Publications.
Frostegård, Å., and Bååth, E. (1996). The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fertil. Soils 22, 59–65. doi: 10.1007/BF00384433
Gange, A. (2000). Arbuscular mycorrhizal fungi, Collembola and plant growth. Trends Ecol. Evol. 15, 369–372. doi: 10.1016/S0169-5347(00)01940-6
Griffiths, B. S., and Bardgett, R. D. (1997). “Interactions between microbe-feeding invertebrates and soil microorganisms,” in Modern Soil Microbiology (New York, NY: Marcel Dekker), 165–182.
Hanlon, R. D. G. (1981). Influence of grazing by Collembola on the activity of senescent fungal colonies grown on media of different nutrient concentration. Oikos 36, 362–367. doi: 10.2307/3544634
Hedlund, K., and Augustsson, A. (1995). Effects of enchytraeid grazing on fungal growth and respiration. Soil Biol. Biochem. 27, 905–909. doi: 10.1016/0038-0717(95)00016-8
Hedlund, K., Boddy, L., and Preston, C. M. (1991). Mycelial responses of the soil fungus, Mortierella isabellina, to grazing by Onychiurus armatus (Collembola). Soil Biol. Biochem. 23, 361–366. doi: 10.1016/0038-0717(91)90192-M
Hedlund, K., and Öhrn, M. S. (2000). Tritrophic interactions in a soil community enhance decomposition rates. Oikos 88, 585–591. doi: 10.1034/j.1600-0706.2000.880315.x
Heemsbergen, D. A., Berg, M. P., Loreau, M., Van Hal, J. R., Faber, J. H., and Verhoef, H. A. (2004). Biodiversity effects on soil processes explained by interspecific functional dissimilarity. Science 306, 1019–1020. doi: 10.1126/science.1101865
Hothorn, T., Bretz, F., and Westfall, P. (2008). Simultaneous inference in general parametric models. Biometr. J. 50, 346–363. doi: 10.1002/bimj.200810425
Jørgensen, H. B., Johansson, T., Canbäck, B., Hedlund, K., and Tunlid, A. (2005). Selective foraging of fungi by collembolans in soil. Biol. Lett. 1, 243–246. doi: 10.1098/rsbl.2004.0286
Kandeler, E., and Gerber, H. (1988). Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 6, 68–72. doi: 10.1007/BF00257924
Kaneko, N., McLean, M. A., and Parkinson, D. (1998). Do mites and Collembola affect pine litter fungal biomass and microbial respiration? Appl. Soil Ecol. 9, 209–213. doi: 10.1016/S0929-1393(98)00077-8
Kutáková, E., Cesarz, S., Münzbergová, Z., and Eisenhauer, N. (2018). Soil microarthropods alter the outcome of plant-soil feedback experiments. Sci. Rep. 8:11898. doi: 10.1038/s41598-018-30340-w
Legendre, P., and Gallagher, E. D. (2001). Ecologically meaningful transformations for ordination of species data. Oecologia 129, 271–280. doi: 10.1007/s004420100716
Lenoir, L., Persson, T., Bengtsson, J., Wallander, H., and Wiren, A. (2007). Bottom–up or top–down control in forest soil microcosms? Effects of soil fauna on fungal biomass and C/N mineralisation. Biol. Fertil. Soils 43, 281–294. doi: 10.1007/s00374-006-0103-8
Lussenhop, J. (1992). Mechanisms of microarthropod-microbial interactions in soil. Adv. Ecol. Res. 23, 1–33. doi: 10.1016/S0065-2504(08)60145-2
Maboreke, H. R., Graf, M., Grams, T. E. E., Herrmann, S., Scheu, S., and Ruess, L. (2017). Multitrophic interactions in the rhizosphere of a temperate forest tree affect plant carbon flow into the belowground food web. Soil Biol. Biochem. 115, 526–536. doi: 10.1016/j.soilbio.2017.09.002
Mikola, J., Bardgett, R. D., and Hedlund, K. (2002). “Biodiversity, ecosystem functioning and soil decomposer food webs,” in Biodiversity and Ecosystem Functioning: Synthesis and Perspectives, eds M. Loreau, S. Naeem, and P. Inchausti (Oxford: Oxford University Press),169–180.
Mikola, J., and Setälä, H. (1998). No evidence of trophic cascades in an experimental microbial-based soil food web. Ecology 79, 153–164.
Moore, J. C., Walter, D. E., and Hunt, H. W. (1988). Arthropod regulation of micro-and mesobiota in below-ground detrital food webs. Annu. Rev. Entomol. 33, 419–435.
Nakano, M., Ochiai, A., Kamata, K., and Nakamori, T. (2017). The preference of Morulina alata (Collembola: Neanuridae) feeding on some fungal sporocarps and the effects of passage through the gut on spores. Eur. J. Soil Biol. 81, 116–119. doi: 10.1016/j.ejsobi.2017.06.005
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O'Hara, R. B., et al. (2013). Package ‘vegan.' Community Ecology Package, version 2. Available online at: http://cran.rproject.org/web/packages/vegan/index.html
Perez, G., Decaëns, T., Dujardin, G., Akpa-Vinceslas, M., Langlois, E., and Chauvat, M. (2013). Response of collembolan assemblages to plant species successional gradient. Pedobiologia 56, 169–177. doi: 10.1016/j.pedobi.2013.04.001
Petersen, H. (2000). Collembola populations in an organic crop rotation: population dynamics and metabolism after conversion from clover-grass ley to spring barley. Pedobiologia 44, 502–515. doi: 10.1078/S0031-4056(04)70067-0
Petersen, H. (2002). Effects of non-inverting deep tillage vs. conventional ploughing on collembolan populations in an organic wheat field. Eur. J. Soil Biol. 38, 177–180. doi: 10.1016/S1164-5563(02)01145-7
Ponge, J.-F. (2000). Vertical distribution of Collembola (Hexapoda) and their food resources in organic horizons of beech forests. Biol. Fertil. Soils 32, 508–522. doi: 10.1007/s003740000285
R Core Team (2013). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Sauvadet, M., Chauvat, M., Brunet, N., and Bertrand, I. (2017). Can changes in litter quality drive soil fauna structure and functions? Soil Biol. Biochem. 107, 94–103. doi: 10.1016/j.soilbio.2016.12.018
Schnürer, J., and Rosswall, T. (1982). Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl. Environ. Microbiol. 43, 1256–1261.
Seastedt, T. R. (1984). The role of microarthropods in decomposition and mineralization processes. Annu. Rev. Entomol. 29, 25–46.
Thimm, T., and Larink, O. (1995). Grazing preferences of some collembola for endomycorrhizal fungi. Biol. Fertil. Soils 19, 266–268.
Tiunov, A. V., and Scheu, S. (2005). Facilitative interactions rather than resource partitioning drive diversity-functioning relationships in laboratory fungal communities. Ecol. Lett. 8, 618–625. doi: 10.1111/j.1461-0248.2005.00757.x
Tullgren, A. (1918). Ein sehr einfacher Ausleseapparat fur terricole Tierformen. Z. Angew. Entomol. 4, 149–150.
Verhoef, H. A., and Brussaard, L. (1990). Decomposition and nitrogen mineralization in natural and agroecosystems: the contribution of soil animals. Biogeochemistry 11:175.
Visser, S., Whittaker, J. B., and Parkinson, D. (1981). Effects of collembolan grazing on nutrient release and respiration of a leaf litter inhabiting fungus. Soil Biol. Biochem. 13, 215–218.
Wardle, D. A., Bardgett, R. D., Klironomos, J. N., Setälä, H., Van Der Putten, W. H., and Wall, D. H. (2004). Ecological linkages between aboveground and belowground biota. Science 304, 1629–1633. doi: 10.1126/science.1094875
Keywords: PLFAs, enzymatic activities, springtails, life-forms, belowground interactions
Citation: Coulibaly SFM, Winck BR, Akpa-Vinceslas M, Mignot L, Legras M, Forey E and Chauvat M (2019) Functional Assemblages of Collembola Determine Soil Microbial Communities and Associated Functions. Front. Environ. Sci. 7:52. doi: 10.3389/fenvs.2019.00052
Received: 01 February 2019; Accepted: 02 April 2019;
Published: 02 May 2019.
Edited by:
Philippe C. Baveye, AgroParisTech Institut des Sciences et Industries du Vivant et de L'environnement, FranceReviewed by:
Andrey S. Zaitsev, University of Giessen, GermanyAlberto Canarini, University of Vienna, Austria
Copyright © 2019 Coulibaly, Winck, Akpa-Vinceslas, Mignot, Legras, Forey and Chauvat. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sékou F. M. Coulibaly, c2Vrb3UtZmFudGEtbWFkLmNvdWxpYmFseUB1bml2LXJvdWVuLmZy; c2ZhbnRhbWFkeS5jQGdtYWlsLmNvbQ==