 Quentin Goffette1,2*†
Quentin Goffette1,2*† Veerle Rots2,3
Veerle Rots2,3 Grégory Abrams4,5
Grégory Abrams4,5 Stéphane Pirson6,7Kévin Di Modica5
Stéphane Pirson6,7Kévin Di Modica5 Fabrice Bray8Dries Cnuts2Dominique Bonjean5
Fabrice Bray8Dries Cnuts2Dominique Bonjean5 Linda Amos9*†
Linda Amos9*†- 1Quaternary Environments and Humans, OD Earth and History of Life, Royal Belgian Institute of Natural Sciences, Brussels, Belgium
- 2TraceoLab/Prehistory, University of Liège, Liège, Belgium
- 3F.R.S.-FNRS, Fund for Scientific Research, Brussels, Belgium
- 4ArcheOs Research Laboratory for Biological Anthropology, Department of Archaeology, Ghent University, Ghent, Belgium
- 5Scladina Cave Archaeological Centre, Espace muséal d'Andenne, Andenne, Belgium
- 6Direction scientifique et technique, Agence wallonne du Patrimoine, Namur, Belgium
- 7Department of Geology (RU Geology & Astrobiology) and European Archaeometry Centre (RU Art, Archaeology, Heritage), University of Liège, Liège, Belgium
- 8Unité d'appui et de recherche (UAR) 3290 Miniaturisation pour la Synthèse, l'Analyse et la Protéomique (MSAP), Centre national de la recherche scientifique (CNRS), University of Lille, Villeneuve D'ascq, France
- 9School of Biological and Environmental Sciences, Liverpool John Moores University, Liverpool, United Kingdom
For a long time, Neanderthals were considered hunters of large mammals, whereas the diversification of the exploited faunal spectrum to include smaller taxa, including birds, was assumed to be specific to anatomically modern humans. In recent decades, archeozoological analyses of faunal remains from layers associated with Middle Paleolithic lithic industries have revealed traces of human manipulation of small taxa, indicating the exploitation of a wider range of animals than previously thought. These new data have challenged the view that Neanderthals did not exploit small animals, thereby narrowing the behavioral gap with anatomically modern humans. Nevertheless, the information currently available comes almost exclusively from southern Europe and the nature of Neanderthal small fauna exploitation in northern Europe remains largely unknown. The present study aims to fill this gap by applying archeozoological methods, including detailed taphonomic and traceological analyses, to 119 bird remains recovered from layers containing Middle Paleolithic industries at Scladina Cave, Belgium. Analyses of proteomics were applied to clarify the taxonomic identity of two morphologically non-diagnostic elements. Modifications made by non-human predators or scavengers, suggest that mammalian carnivores are responsible for accumulating a considerable portion of the avian assemblage. In total, seven bird bones exhibit anthropogenic marks, and one element presents questionable marks. Various Galliformes taxa and a great cormorant were exploited likely for their meat. The talon of a likely lesser spotted eagle displays intense polishing possibly linked to human manipulation of this element, although this remains hypothetical. On the radius of a Western capercaillie, two deep incisions may indicate bone working, and intense use-wear indicates that the bone has been utilized, potentially on soft organic material. This study provides the first evidence of the exploitation of birds by Neanderthal in Belgium and constitutes the only detailed zooarchaeological analysis of Middle Paleolithic bird material in northwestern Europe. The likely modification and subsequent utilization of a bird bone is only the second example known from Neanderthal occupations in Eurasia. The novel taxa identified as Neanderthal prey highlight the plasticity of Neanderthal ecological behavior, adapting to different landscapes and climates and exploiting a large spectrum of locally available prey.
1 Introduction
1.1 Historical background and archaeological perspective
Traditionally, Neanderthals were considered hunters of large mammals (Rabinovich and Tchernov, 1995; Hoffecker and Cleghorn, 2000; Speth and Tchernov, 2007; Gaudzinski-Windheuser and Niven, 2009; Delagnes and Rendu, 2011), whereas the diversification of the exploited faunal spectrum to include smaller, faster taxa was assumed to be specific to anatomically modern humans. Building on Flannery (1969)'s post-Paleolithic broad spectrum revolution (BSR), researchers suggested that the true diversification of dietary exploitation took place during the transition between the Middle to Upper Paleolithic (Stiner, 2001; Hockett and Haws, 2002; Stiner and Munro, 2002) driven by the alleged greater cognitive capacities of the anatomically modern humans moving out of Africa and into the Eurasian Neanderthal homelands.
However, in recent decades, multiple lines of evidence indicating complex cultural behavior in Neanderthals have emerged, demonstrating that they manufactured bone tools (Soressi et al., 2013; Abrams et al., 2014; Hutson et al., 2018), practiced hafting and hunted with stone projectiles (Rots, 2013, 2015), collected marine shells, stones, and pigment for aesthetic purposes (Soressi and d'Errico, 2007; Zilhão et al., 2010; Radovčić et al., 2016), produced abstract signs (Rodríguez-Vidal et al., 2014; Hoffmann et al., 2018), and engaged in mortuary practices (Maureille and Van Peer, 1998; Rendu et al., 2014; Maureille et al., 2016) at several sites across Europe predating the arrival of anatomically modern humans. Collectively, these studies have led to a shift in our perception of Neanderthals from archaic distant relatives to cognitively equal contemporaries of anatomically modern humans in the Pleistocene landscapes (Villa and Roebroeks, 2014; Romagnoli et al., 2022).
Archeozoological analyses of faunal assemblages from Middle Paleolithic sites, especially those focusing on the small mammal and avian components, have shed further light on the necessity for sophisticated hunting, and on dietary practices of Neanderthals. At several sites in southern Europe, traces of Neanderthal manipulation of small taxa are now relatively frequent, indicating the exploitation of a wider range of animals than previously thought, including small or fast-moving animals such as mollusks, leporids, marine mammals, and birds, some of which present challenges in their collection (Stiner et al., 2000; Speth and Tchernov, 2002; Stringer et al., 2008; Bonjean et al., 2012; Blasco et al., 2014, 2016, 2022; Morin et al., 2019; Zilhão et al., 2020).
Rock doves, corvids and Galliformes are the most commonly documented food bird species exploited by Neanderthals. Cutmarks and thermal modifications identified on bones from Middle Paleolithic contexts across Iberia and southern Europe, demonstrate butchery and cooking practices (Cova Negra IIIb, Spain—Martínez Valle et al., 2016; Gorham's, Vanguard, and Ibex Cave, Gibraltar, UK—Finlayson et al., 2012; Blasco et al., 2014, 2016; Arbreda Cave, Spain—Lloveras et al., 2018; Gruta da Oliveira and Gruta da Figueira Brava, Portugal—Nabais et al., 2023; La Crouzade, France—Garcia-Fermet et al., 2023; Pié Lombard, France—Romero et al., 2017; Fumane A9, Italy—Fiore et al., 2016). Ducks were a regular part of the Neanderthal diet during successive occupations at Bolomor Cave (Spain, 350–120 ka; Blasco and Fernández Peris, 2009; Blasco et al., 2013), while one swan bone has been recovered at the same location (Bolomor Cave XII, Spain—Blasco et al., 2008). Other species appear to also bear evidence of dietary exploitation, including Charadriiformes and diurnal raptors (Axlor, Spain—Gómez-Olivencia et al., 2018; La Crouzade Cave, France—Garcia-Fermet et al., 2023; Riparo del Broion, Italy—Romandini et al., 2023) while feather barbules reported on lithic tools at Payre and Abri du Maras in southern France suggest bird exploitation without direct evidence visible on any recovered bird remains (Hardy and Moncel, 2011; Hardy et al., 2013; Rufà et al., 2016). One feather barbule has also been recovered on a stone tool from the Middle Paleolithic layer A11 at Fumane (Cnuts et al., 2022).
A growing number of sites also report marks related to the harvest of feathers and talons, sometimes interpreted as having been used for cultural purposes such as personal decoration and pendants. A collection of cut-marked and polished white-tailed eagle pedal phalanxes from Krapina, Croatia (130 ka) have been interpreted as a part of a necklace (Radovčić et al., 2015), while cutmarks on the ulnae of raptors and dark-feathered species and/or cutmarks on talons were observed in Gibraltar (Finlayson et al., 2012), southern France (Combe Grenal and Les Fieux—Morin and Laroulandie, 2012; Laroulandie et al., 2016; Pech de L'Azé I and IV and Mandrin—Soressi et al., 2008; Dibble et al., 2009), and Italy (Fumane—Peresani et al., 2011; Romandini et al., 2016; Rio Secco—Romandini et al., 2014). Consequently, multiple lines of evidence now show that Neanderthals in southern Europe hunted a range of bird species for both utilitarian and non-utilitarian purposes. However, other than cutmarks on two Anseriformes elements from Salzgitter-Lebenstedt, Germany (Gaudzinski-Windheuser and Niven, 2009), cutmarks on a hare pelvis from Unit 5 at Scladina Cave, Belgium (Bonjean et al., 2012), and possible evidence of fishing at Walou Cave, Belgium (Van Neer and Wouters, 2011), the purpose of Neanderthal small fauna exploitation in northern Europe remains largely unknown.
Belgium is a particularly favorable ground for the study of the Paleolithic due to its ideal location in the plains of northwestern Europe and favorable geological parameters. In southern Belgium, the presence of deep valleys cutting through the limestone substrate creates conditions conducive to the formation of rock shelters or underground cavities, from small passages to complex cave networks. While most of these caves are of geological and ecological interest only, they regularly offer archaeological significance where, most of the time, excellent preservation of artifacts and osseous material provides information about past human societies (Di Modica et al., 2016). As part of the faunal record, bird bones also frequently preserve well, despite their fragile nature (Goffette et al., 2020, 2023; López-García et al., 2024). Over more than 150 years, thousands of bird bones have been recovered from rich archaeological sites in Belgium, including Middle Paleolithic occupations, but these have not yet been studied from a zooarchaeological point of view. As a result, the exploitation of birds by Neanderthals in Belgium is hitherto unknown.
The present study aims to fill in this gap by applying archeozoological and taphonomic methods to bird remains recovered from the archaeological site of Scladina Cave in southern Belgium. Scladina is an exceptional site because it is one of only two archaeological excavations from the end of the 20th century in Belgium that have yielded an extensive stratigraphic sequence covering at least the end of the Middle Pleistocene up to the Holocene, delivering Middle and Upper Paleolithic industries as well as Neanderthal remains (Pirson, 2007, 2011; Pirson et al., 2008; Toussaint et al., 2017). Scladina also contains several Neanderthal occupations (Di Modica and Bonjean, 2004), allowing us to investigate Neanderthal behavior at the same geographical location at two different intervals during the Middle Paleolithic (MIS 6 and MIS 3).
1.2 Scladina Cave
Scladina Cave was discovered by amateur archaeologists in 1971 and has been the subject of scientific excavations since 1978, initially under the leadership of the University of Liège and now under the direction of the Espace muséal d'Andenne with the support of various institutions such as the Walloon Heritage Agency, the Wallonia-Brussels Federation, and the City of Andenne. Since the mid-1980s, the excavations have become permanent and are now conducted in an interdisciplinary manner.
The cave is located between Andenne and Namur, on the right bank of the Meuse, in a secondary valley where the Ri de Pontainne, a small water stream, intermittently flows. Scladina is part of a complex network of caves that was partially excavated starting from the late 1940s (Dewez, 1981). These excavations have notably uncovered numerous artifacts and abundant fauna in the adjacent Saint-Paul Cave, as well as in the underlying Sous-Saint-Paul cave.
Initially, Scladina Cave was recognized for yielding archaeological remains from two major Middle Paleolithic occupations (Units 1A and 5), then later also from many smaller occupations (Loodts, 1998; Moncel, 1998; Otte, 1998a; Di Modica and Bonjean, 2004; Di Modica, 2010). Unit 5 yielded abundant and well-preserved fauna bearing anthropogenic modifications (Patou-Mathis, 1998) and bones used as retouchers, including cave bear remains (Abrams et al., 2014; Abrams, 2018). Unit 1A, although more recent, does not display the same state of preservation. The faunal remains are heavily altered, making it difficult to observe potential anthropogenic modifications (Bourdillat, 2008). However, nearly 200 burned bone fragments attest to their use as fuel (Abrams et al., 2010). Fragments of black coloring rocks, interpreted as pigment, have also been unearthed within this assemblage (Bonjean et al., 2015).
Starting from 1990, the discovery of dental and facial remains belonging to a Neanderthal juvenile individual bestowed Scladina with additional significance (Toussaint and Pirson, 2006; Toussaint et al., 2014). In Belgium, apart from a tooth unearthed at the Trou de l'Abîme in Couvin a few years earlier (Toussaint et al., 2010) and another tooth from Walou Cave (Toussaint et al., 2017), no Neanderthal remains had been found in stratigraphic context since the late 19th Century (Toussaint and Pirson, 2006; Toussaint et al., 2011).
Although the site is primarily known for its occupations by Neanderthal populations, more anecdotal evidence related to the Upper Paleolithic complements the cultural sequence (Otte, 1998b). Modified bones associated with an Upper Paleolithic end scraper on a blade have contributed to dating the emergence of the Upper Paleolithic in Belgium (Abrams et al., 2024).
1.3 Stratigraphy, sedimentary dynamics, paleoenvironment and chronostratigraphic framework
In the last 20 years, major interdisciplinary studies focusing on stratigraphy, sedimentary dynamics, paleoenvironmental reconstructions, and chronostratigraphy were conducted in Scladina. These recent studies complement and refine the numerous datasets already available for the site (Otte, 1992; Otte et al., 1998). With its sedimentary infill covering a long period from the Middle Pleistocene to the Holocene, Scladina appears as one of the most complete cave entrance sequences in Belgium (Pirson, 2007, 2014; Pirson et al., 2018, 2014). It is a reference site for cave entrance sedimentology and a key sequence for the study of paleoenvironments in northwestern Europe.
Since Scladina Cave is still under excavation, the stratigraphy meets the quality criteria of a modern excavation. Dozens of sedimentary profiles were studied in detail throughout the whole site (e.g., Gullentops and Deblaere, 1992; Haesaerts, 1992; Otte, 1992; Otte et al., 1998; Pirson, 2007, 2014), illustrating the complexity of the stratigraphy. Important lateral sedimentary variations were highlighted as well as complex geometries and varied lithologies. The 2007 stratigraphic reappraisal eventually led to the definition of the actual 15-m-thick Scladina stratigraphic sequence encompassing ca. 120 layers grouped in 30 major units (Pirson, 2007) (Figure 1). These field studies revealed the diversity of the sedimentary sources and a large variety of sedimentary dynamics dominated by slope processes, including debris flow, run-off, rock fall, solifluction, settling, torrential flow, and speleothem formation (Pirson, 2007, 2014). The stratigraphy of sediments accumulated under an aven, or sinkhole, opening in the cave ceiling was recorded separately. Complex diagenetic processes have also been recognized.

Figure 1. Scladina Cave stratigraphic sequence and chronostratigraphic interpretation, showing the stratigraphic position of bird remains with anthropogenic marks. (?) = uncertain anthropogenic marks. Black braces indicate the stratigraphic attribution. The blue bar indicates the potential stratigraphic origin of the bones discovered in sedimentary Complex 4, taking into account possible attribution errors and rearrangements (see text). Dates: synthesis of all reliable dates available for the Scladina Cave sequence (see Bonjean, 1998; Pirson, 2007; Bonjean et al., 2012; Pirson et al., 2014; Abrams et al., 2024). All the 14C dates on this figure are uncalibrated. When an age interval is given, the number of dates concerned is indicated in a circle. Black = 14C date on bone; italic black = 14C date on calcite (in situ and reworked speleothems); bold black = 14C date on teeth accurately positioned in the new stratigraphic system; * + bold black = 14C date (hydroxyproline) on bone retoucher accurately positioned in the new stratigraphic system; = U/Th date on in situ speleothem; = U/Th date on reworked fragments of speleothems; = U/Th date (gamma spectrometry) obtained on the Neanderthal mandible; = thermoluminescence date on sediment; italic red = thermoluminescence date on in-situ speleothem; = luminescence date on burned flint (TL) and on sediment (IRSL); = coupled U/Th–ESR dates on bone; circled numbers = number of dates for CC4 interval. () = problematic date.
Scladina yielded one of the best interdisciplinary-studied cave entrance sequences in Belgium, together with Walou Cave, east of Liège (Draily et al., 2011; Pirson et al., 2011). The paleoenvironments have been documented through large and small mammals, amphibians, reptiles, palynological, anthracological, sedimentological, magnetic susceptibility, and pedological studies (e.g., Otte, 1992; Otte et al., 1998; Ellwood et al., 2004; Pirson, 2007; Pirson et al., 2008, 2014; Blain et al., 2014). More than twenty alternating climatic phases have been documented, including cold stadials, short interstadials, and longer temperate phases, either interglacial or early glacial. Scladina is also the most densely dated prehistoric site in Belgium, with more than 90 available dates (TL, IRSL, combined U/Th-ESR, U/Th, and radiocarbon). Recently obtained U/Th dates (Vonhof et al., 2024) led to refine the existing chronostratigraphic framework (see Pirson et al., 2014, 2008). The current chronostratigraphic interpretation of this complex sequence is based on climatostratigraphic, biostratigraphic, and mineralogical data, numerical dates, and comparison with the reference loess sequence from Central Belgium (Figure 1).
In this paper, we will focus on stratigraphic Unit 1A (MIS 3), the Sedimentary Complexes 3 and 4A (MIS 5), Unit 4B (MIS 6), and Unit 5 (MIS 6) encompassing in total more than 30 layers (Pirson, 2007, 2014). Unit 1A was mainly deposited by two sedimentary processes (debris flow and run-off). Complex of Units 3 is dominated by debris flow and run-off (locally torrential flow), with also rock fall and speleothem formation. Unit 4B mainly results from settling and run-off, while Unit 5 consists of a diamicton affected by solifluxion. The situation of 4A is special, as the stratigraphic reappraisal revealed a much more important complexity in this part of the stratigraphy than previously thought. Four distinct Units, each encompassing several layers, have been described in this sedimentary complex. Unit 4A-AP results from debris flow and wash processes. Palynological data suggests it records the first stage of a strong climatic improvement. Unit 4A-IP mainly consists of a thick stalagmitic floor (CC4) pointing to temperate conditions. Unit 4A-CHE is the result of an important erosional phase with cut-and-filled layers developed in a deep gully structure, resulting from a cold episode. Finally, Unit 4A-POC consists of fine-grained sediments mainly deposited by run-off and covering both underlying Units 4A-IP and 4A-CHE. This complexity was not fully understood before the stratigraphic reappraisal, therefore sometimes implying some uncertainty in the stratigraphic attribution of the material unearthed before 2007. This is especially the case for Unit 4B, which was supposed to lie below the stalagmitic floor CC4 in the original stratigraphy. Following the reappraisal, the recognition of the 4A-CHE gully, opening downwards from the surface of CC4, implies that in some situations Unit 4B corresponds in fact to 4A-CHE. It is also worth mentioning here that the gully erodes underlying Units 4A-IP, 4A-AP, and 4B, and locally also part of Unit 5.
The new U/Th dates recently obtained on speleothems led to reinterpret Sedimentary Complex 4 (Vonhof et al., 2024), as they indicate that the thick CC4 speleothem records the Eemian. As a result, underlying units 4B and 5 are now placed at the end of the Middle Pleistocene, in MIS 6.
Several modified bird bones have been recovered from the upper part of the Scladina sequence, but as they were unearthed before the stratigraphic reappraisal (Pirson, 2007), some uncertainties remain about their detailed stratigraphic position. This is mainly the case for Unit T, where early Upper Paleolithic material (Abrams et al., 2024) has been found alongside artifacts reworked from Middle Palaeolithic layer 1A. In this context, anthropologically modified bones excavated from Unit T before the stratigraphic reappraisal cannot be precisely attributed, therefore requiring radiocarbon dating. These are in progress and will be presented in another paper.
2 Materials and methods
Our study consisted of several steps. First, we selected the stratigraphic layers that had yielded testimonies of human activities, targeting the assemblages with the highest potential for traces of bird exploitation. We thus targeted 22 sedimentary assemblages from the Upper Paleolithic and Middle Paleolithic, from which we sorted all the bone remains to isolate the bird bones. The only exception was the avian remains from the layers of Unit 1A, that had been isolated by researchers from the Institut de Paléontologie Humaine (Paris) following the sorting of the faunal material from this assemblage. A new sorting of this assemblage was therefore not undertaken. Some bird bones had been isolated by the excavation team. This explains the presence of bones from units 3-SUP, 3-INF, and 6A to 6C, which we did not sort in the course of this work. This selection means that we have not studied certain units (2A, 2B, 6D, and 7A to 7C), while others are only partially studied (3-SUP, 3-INF, and 6A to 6C). Further bird bones may still be identified among the faunal remains of these layers, but the potential for finding remains with anthropogenic modifications is low given the low frequency of other archaeological material. All sediment was sieved to a maximum mesh size of 5 mm, except in the case of some specific research efforts (e.g., small mammals) when a 2.5 mm mesh was used.
For all the specimens, the attribution to a stratigraphic unit determined at the time of discovery has been retained in this manuscript, unless a stratigraphic reattribution in relation with the 2007 stratigraphic reappraisal could be carried out with certainty (e.g., based on the examination of existing neighboring stratigraphic sections). Then, the new stratigraphic attribution was used. This point is particularly important for unit 4B, to which bones were attributed before the stratigraphic reappraisal, although their attribution to unit 4B is questionable (see Discussion). However, due to the lack of possible reattribution, we retained the original attribution.
The sorting led to the recovery of 159 bird remains (Middle Paleolithic, n = 119; Upper Paleolithic, n = 40). All were subjected to an archeozoological study but here, the focus is on material recovered from Middle Paleolithic-related sedimentary units. Elements with uncertain stratigraphic attribution that potentially come from layers including Middle Paleolithic material were also retained. The stratigraphic details are given in Figure 1 and Supplementary Tables 1, 2. Bones associated with the Upper Paleolithic have been analyzed but will not be presented in this article.
The zooarchaeological study includes detailed taphonomic and traceological analyses. First, we determined the anatomical part and the taxon present using reference collections held at the Institute of Natural Sciences (Brussels, Belgium), as well as reference manuals dedicated to the identification of certain taxonomic groups (Woolfenden, 1961; Bacher, 1967; Woelfle, 1967; Erbersdobler, 1968; Kraft, 1972; Langer, 1980; Bocheński et al., 2022; Wertz et al., 2022). The taxonomy used follows that of Gill et al. (2022). The nomenclature used to describe anatomic features of bones and soft tissues follows Baumel (1993). The side (left or right) of the bone was recorded. Where possible, measurements were taken according to von den Driesch (1976) or dedicated manuals. The calculation of the minimum number of individuals (MNI) was based on the minimum number of elements (MNE) completed by the laterality and the age of the individuals. Age estimation was made by examining the ossification stage to attribute each bone to one of the four age classes summarized in Serjeantson (2009). The sex of certain taxa was assessed based on measurements. The completeness or fragmentation of each element was recorded, as were the parts preserved and the morphology of the fracture edges (Fernández-Jalvo and Andrews, 2016; Romero et al., 2016). In addition, each bone was examined using a binocular microscope (magnification 6.5–50 × ) with oblique cold light, to record surface alterations, including color, root etching, erosion, thermal alterations, weathering, and gnawing or beak marks (Behrensmeyer, 1978; Lyman, 1994; Behrensmeyer et al., 2003; López-González et al., 2006; Bocheński et al., 2009, 2018; Fernández-Jalvo and Andrews, 2016). To create a workable dataset, while still preserving the integrity of the stratigraphy, we grouped the Middle Paleolithic remains into five samples according to sedimentary units and MIS (Supplementary Table 3).
To observe and illustrate some of the modifications in detail, two scanning electron microscopes were used and operated under low vacuum conditions to eliminate the need for a conductive coating, a FEI Quanta 200 (23 kV; Mineralogical Laboratory of the Geological Survey of Belgium) and a JEOL-IT300 (20 kV; TraceoLab of ULiège). In addition, a digital microscope Hirox HRX-01 equipped with an H-2500E attachment and an LED ring light was used (acquisition mode multi-focus or 3D; TraceoLab of ULiège). The works of Barisic (2006) and Domínguez-Rodrigo et al. (2009) have been used to help distinguish between cut and trampling marks, bearing in mind that it can be very difficult to differentiate between intentional cutmarks and trampling marks, particularly when bones are mixed with lithic pieces in the sediment (Barisic, 2006).
Analyses of proteomics (MALDI) were applied to clarify the taxonomic identity of two bird bones with anthropogenic modifications that did not preserve sufficient morphological characteristics to allow any taxonomic identification beyond the class level. The digestion of bones was described in Bray et al. (2023). Briefly, 1–5 mg of bones were deposited in 96 wells plate MultiScreenHTS-IP, 0.45 μm (MSIPS4510, Millipore, Billerica, MA, USA). The bone powder was demineralized, washed, gelatinized and digested in the 96 wells plate MultiScreenHTS-IP. The demineralization solution was digested in classical 96 well plates. The peptides form bones and demineralized solution were purified on C18 96 well plates (Affinisep, Petit-Couronne, France). Purification of peptides was performed by washing the plates once with 500 μL of acetonitrile (ACN) followed by a washing step with 80% ACN, H2O 0.5% acetic acid repeated 3 times and a second washing repeated 3 times with H2O alone 0.5% acetic acid. Tryptic peptides from bone powder were resuspended in 200 μL of a H2O, 0.5% acetic acid solution. Tryptic peptides form both solutions were transferred to C18 96-well plate and eluted with a vacuum manifold. The plate was washed 3 times with 200 μL of H2O, 0.5% acetic acid. Peptides were recovered in a V-bottom well collecting plate using 100 μL of a 80% ACN, 0.1% acetic acid solution followed by 100 μL of ACN. The plate was evaporated on TurboVap 96 Evaporator (Caliper LifeScience, Hopkinton, USA). For mass spectrometry analysis, the sample was dissolved again in 10 μl of H2O 0.1% formic acid. Mass spectrometry analysis was performed by depositing desalted peptides (1 μL) on 384 massive MALDI plates (Bruker Daltonics, Bremen, Germany), then 1 μL of HCCA matrix at 10 mg/mL in ACN/H2O 70:30 v/v 0.1% TFA was added for each sample. The spots were dried at room temperature. MALDI FTICR experiments were carried out on a Bruker 9.4 Tesla SolariX XR FTICR mass spectrometer (Bruker Daltonics, Bremen, Germany). The frequency of the laser was set to 1,000 Hz using the “Minimum” predefined shot pattern. MALDI FTICR spectra were generated in the m/z range from 693.01 to 5,000 by accumulating 10 scans. Two M data points were used per spectrum which corresponds to a transient duration of 5.0332 s. The transfer time to the ICR cell was set to 1.2 ms and the quadrupole mass filter set at m/z 600 was operated in RF-only mode. MS raw data from MALDI FTICR were processed using DataAnalysis 5.0. The identification is supported by all peptide markers presented in previous reports (Eda et al., 2020; Codlin et al., 2022). The mass spectrometry proteomics data have been deposited on the ProteomeXchange Consortium (https://proteomecentral.proteomexchange.org/) via the PRIDE partner repository with the data set identifier PXD051309.
3 Results
3.1 Taxonomic composition
The bird bone assemblage from Scladina Cave recovered from Middle Paleolithic stratigraphic units comprises a total of 119 remains representing at least 32 individuals from 16 taxa (Table 1; Supplementary Tables 1, 2). The assemblage does not appear highly taxonomically diverse. Most of the remains have been identified to genus or specific level, so taxonomic diversity is probably not underestimated due to identification issues. The bones present come mainly from medium- (such as ducks and ptarmigans), to large-sized taxa (such as goose and capercaillie). It is likely that conservation conditions within the sediment favored the preservation of the largest, most resistant bones, to the detriment of the smaller elements. If we consider the proportions of the different orders present, Galliformes largely dominate the assemblage both in terms of NISP and MNI, ahead of Anseriformes, while the other orders are less abundant.
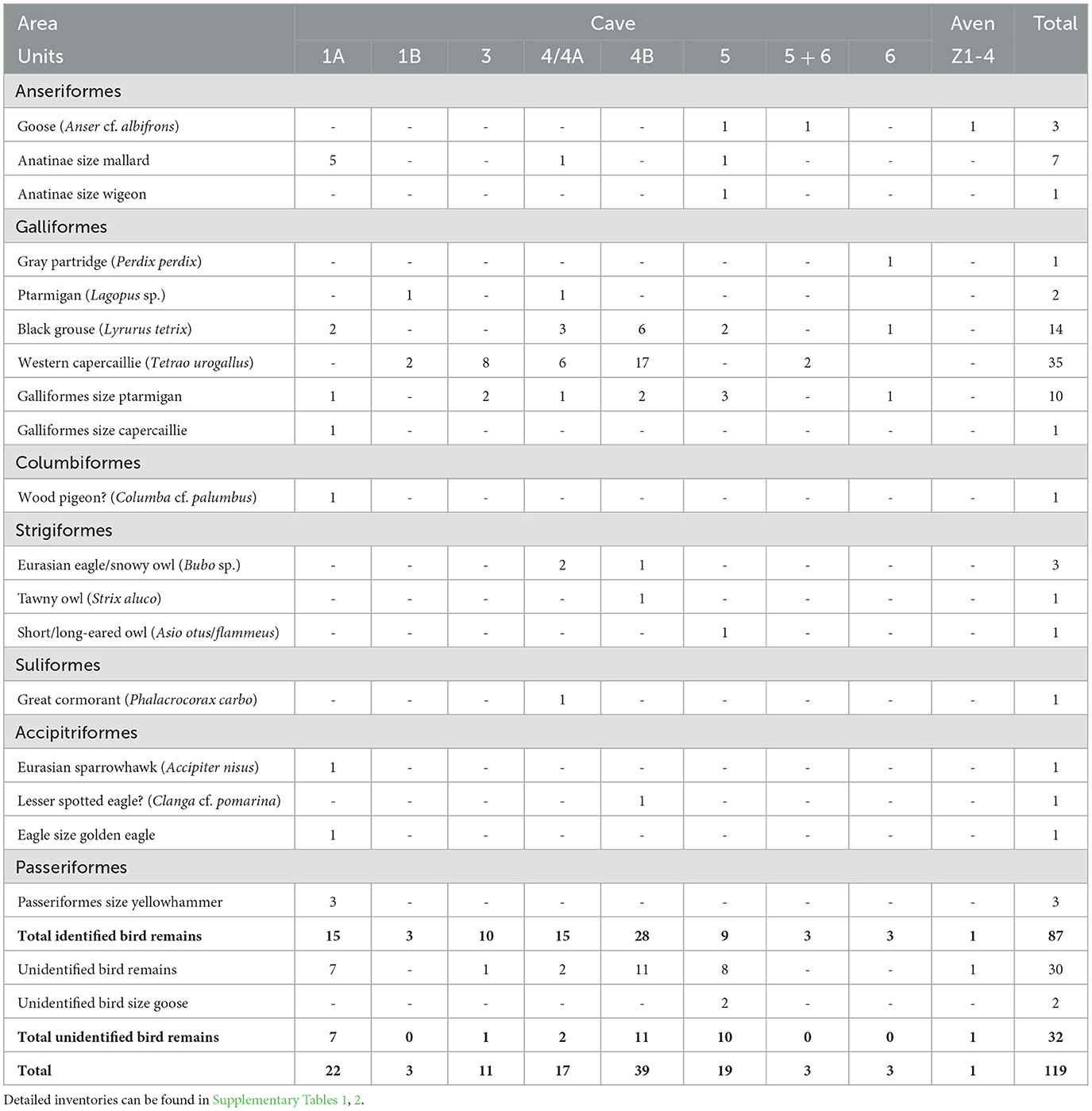
Table 1. Inventory of the number of identified specimens (NISP) of bird remains from Scladina Cave, grouped by Units.
Within the order Anseriformes, remains of geese were present alongside duck bones. The order Galliformes included almost exclusively remains belonging to the subfamily Tetraoninae and was chiefly dominated by the Western capercaillie. The black grouse and willow/rock ptarmigan were also present. Furthermore, one bone of gray partridge was identified. The order Strigiformes was represented by the tawny owl (Figure 2A), the long-eared/short-eared owl, and a large owl, possibly the Eurasian eagle owl in the case of a premaxillary of very large size (Figure 2C). In the order Accipitriformes, we noted the presence of a small species of raptor, the European sparrowhawk, and two species of eagle; one the size of a golden eagle, the other likely a lesser spotted eagle. In the order Passeriformes, three remains were from a passerine the size of the yellowhammer. Additionally, one bone of a great cormorant (Suliformes) and one of a pigeon, probably the wood pigeon (Columbiformes) were identified in the assemblage.

Figure 2. Scladina Cave. (A) Tawny owl, left coracoid (Sc1999-64-5; Unit 4B). (B) Western capercaillie, proximal right humerus with gnawing marks from carnivore (Sc1997-52-258-1; Unit 4B). (C) Eurasian eagle or snowy owl, premaxilla (Sc1993-192-194-6 and Sc1993-192-194-7; Unit 4A). (D) Western capercaillie, partial skull (Sc1995-397-94-1; Unit 4B).
One radius fragment from a large bird with no taxonomically diagnostic morphological features (Sc1985-910-309) was identified through proteomics as Tetraoninae (Supplementary Figure 1 and Supplementary Table 4). The only Tetraoninae known to have occurred during the Middle Paleolithic in Western Europe large enough to match the size of this element is the Western capercaillie. This taxonomic identification has been retained on this basis. No morphometric criteria contradict the identification obtained through proteomic analyses. Proteomic analyses were also applied to another unidentified bone fragment (Sc1985-783-196), but taxonomic identification was not successful (Supplementary Figure 2). Results showed that the specimen has been affected by unknown contaminants.
3.2 Age and sex
All bird bones collected are of adult individuals, with the exception of the European sparrowhawk which was a sub-adult.
The sparrowhawk bone came from a female individual. The sex ratio in the black grouse (NISP = 3 male; 1 female; 3 likely female) is balanced but in the capercaillie, practically all the bones identified come from males since only 3 bones out of 30 which could be sexed come from female birds. However, this is not significant because a detailed examination of the remains of capercaillie from layer 4B, which delivered most of the bones from this species, revealed the presence of two female individuals to one male individual.
3.3 Taphonomic analysis
The taphonomic sample totaled 99 specimens from 15 layers; 16 from Unit 1A, 10 from complex of Units 3, 18 from complex of Units 4A, 36 from Unit 4B, and 19 from Unit 5. Remains from other Middle Paleolithic contexts were also examined, but are excluded from this study due to uncertain provenance.
3.3.1 Post-depositional processes
Concreted sediments were observed on 75% of the specimens in Unit 1A (Figure 3; Supplementary Table 5), 60% of specimens in the complex of Units 3, and 52.7% of specimens in Unit 5. In the Sedimentary Complex 4A and in Unit 4B concretions were observed on 17.7% and 22.3% respectively. The modifications appeared as localized attached sediments covering between 0–75% of the bone surfaces. Modifications related to weathering (cracks, flaking, etc.) affected 100% of the bones in complex of Units 3, 75% of elements in Unit 1A, ca. 70% of remains in Unit 5 and complex of Units 4A, and 54.3% in Unit 4B. In line with observations made by Bocheński and Tomek (1997), only three stages of weathering were apparent on the material, not including stage 0 (no weathering).
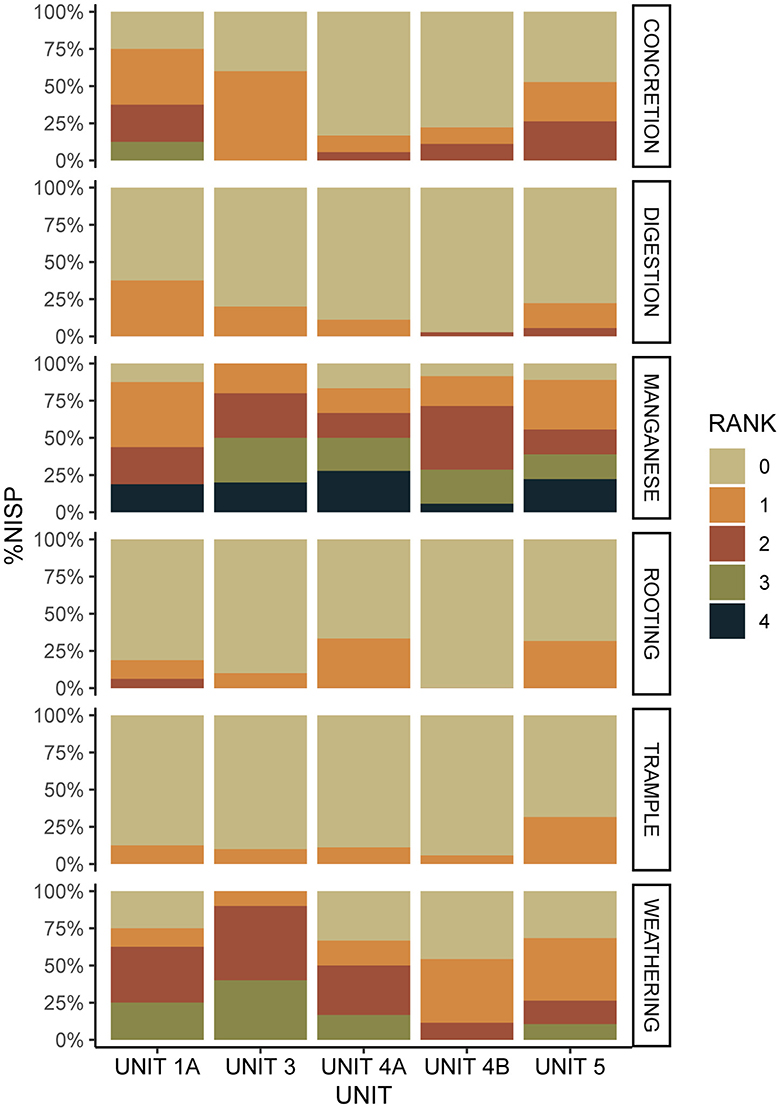
Figure 3. Bar charts showing the dispersal of natural taphonomic modifications and digestion modifications observed in the taphonomic samples ranked on a scale from 0 (= none) to 4 (= very high).
Manganese dioxide deposits were observed at high frequencies in all five samples (Unit 1A = 87.5%, complex of Units 3 = 100%, complex of Units 4A = 88.3%, Unit 4B = 91.5%, Unit 5 = 89.5%), which may indicate high bacterial activities and/or high humidity levels within the cave. These processes are not mutually exclusive. Modifications ranged from small dark stains on the bone surface (grade 1) to covering the entire bone surface (grade 4). Irregular u-shaped striations from rooting were absent in Unit 4B and appeared infrequently in complex of Units 3 (10%) and Unit 1A (18.8%). Rooting marks were of a similar frequency in complex of Units 4A (35.2%) and Unit 5 (31.5%). Striations related to trampling were infrequent throughout the five samples, but most prominent in the occupation sediments of Unit 5 where 31.5% of the sample was affected. However, sample sizes were too small to interpret trampling frequencies beyond presence and absence.
3.3.2 Fragmentation
Fragmentation is significant throughout the entire Middle Paleolithic sequence: only 9 out of 100 long bones are complete (9%). The proportion of complete bones per taxon reflects the abundance of their bones within the site. The Western capercaillie, a species with large and robust bones, provides only one complete element. There does not seem to be any preferential conservation of a particular skeletal element. Strikingly, Unit 4B and Unit 5 yield no complete long bone despite being the richest in bird bones. This testifies to the intense fragmentation in these layers. On the contrary, complex of Units 4 delivered parts of two bird skulls, one of an owl of the genus Bubo (Figure 2C) and another of a Western capercaillie (Figure 2D). Although incomplete and fragmented, these fragile elements are quite well preserved, highlighting the complexity of the sedimentary dynamics within Scladina Cave.
3.3.3 Corrosive and mechanical predator modifications
Marks concurrent with digestion were identified in all five samples (Figure 3; Supplementary Table 5). In Unit 1A (37.5%), complex of Units 3 (20%), and complex of Units 4A (11.8%) all corrosion was of low level (grade 1). Two levels of corrosion were identified in Unit 5 (grade 1 = 16.7%, grade 2 = 5.6%), and a single element in Unit 4B exhibited grade 2 corrosion (2.8%). Along with low occurrences of polish (Unit 1A = 6.3%, complex of Units 3 = 10%, Unit 5 = 10.5%), these modifications could indicate bird of prey agency and mammalian carnivore agency.
Mechanical marks in the form of pitting, punctures, and crenulated edges add further information to the predator profile. 57.6% of the mechanical marks were concentrated on the articular ends of the bones. In Unit 1A, 37.5% of the remains bore modifications caused by mammalian carnivores (Figure 4; Supplementary Table 5). These included remains of Galliformes, ducks, and a rock dove-sized bird. A further 12.5% displayed marks that were likely produced by carnivores. Seven of 10 Galliformes bird remains from complex of Units 3 were predated by mammalian carnivores as attested by punctures, crenulated edges and tooth grooves. All but one of these remains were concentrated at the junction between Scladina and Saint-Paul caves, and may indicate carnivore denning activity. In the complex of Units 4A, carnivore agency (29.4%) was observed in the remains of two species of Galliformes (black grouse and Western capercaillie) and one large owl of the genus Bubo. Two further specimens (11.8%), one great cormorant and one Western capercaillie, displayed pitting and puncture patterns that could be of human origin. Carnivore modifications affected 33.3% of the remains in Unit 4B (33.3%), all of which present on Galliformes. Unit 5 contained the highest frequency of mechanical modifications, with pitting and punctures from small carnivores visible on 47.4% of the remains of waterfowl and Galliformes. Rodent gnawing was observed on a single element (5.3%). No conclusive evidence of mechanical bird of prey modifications was observed in the samples.
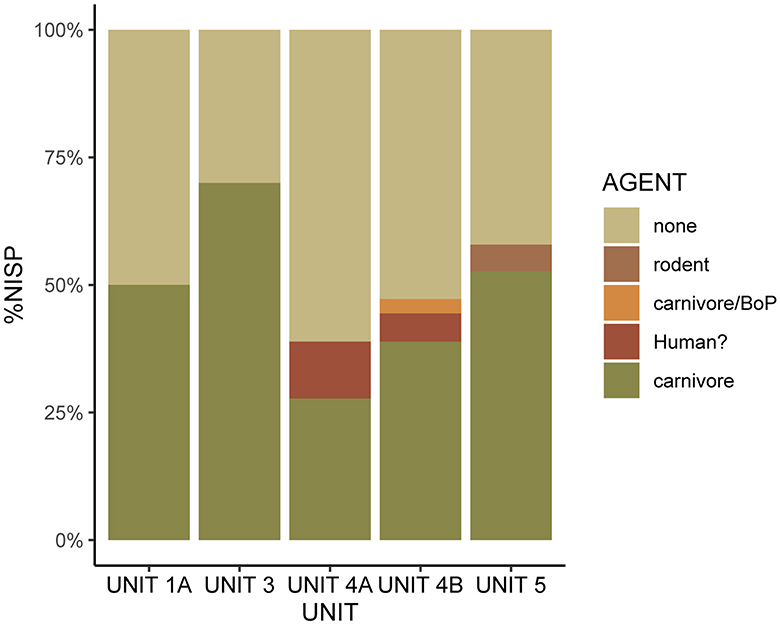
Figure 4. Bar chart showing the proportions of mechanical agency visible on the avian remains in the five taphonomic samples from Scladina Cave. Carnivore marks appear most frequently throughout the sample. BoP, bird of prey.
3.3.4 Anthropogenic modifications
Marks interpreted as anthropogenic were recorded on 7 remains (6.7% of the Middle Paleolithic bird material from Scladina Cave) recovered from the complex of Units 4 (MIS 5 or 6; n = 6; 8.9%) and in Unit 5 (MIS 5 or 6; n = 6; 10.5%) while another specimen from the complex of Units 4 displays uncertain anthropogenic marks (Table 2). The stratigraphic position of these bones appears on the stratigraphic log (Figure 1).
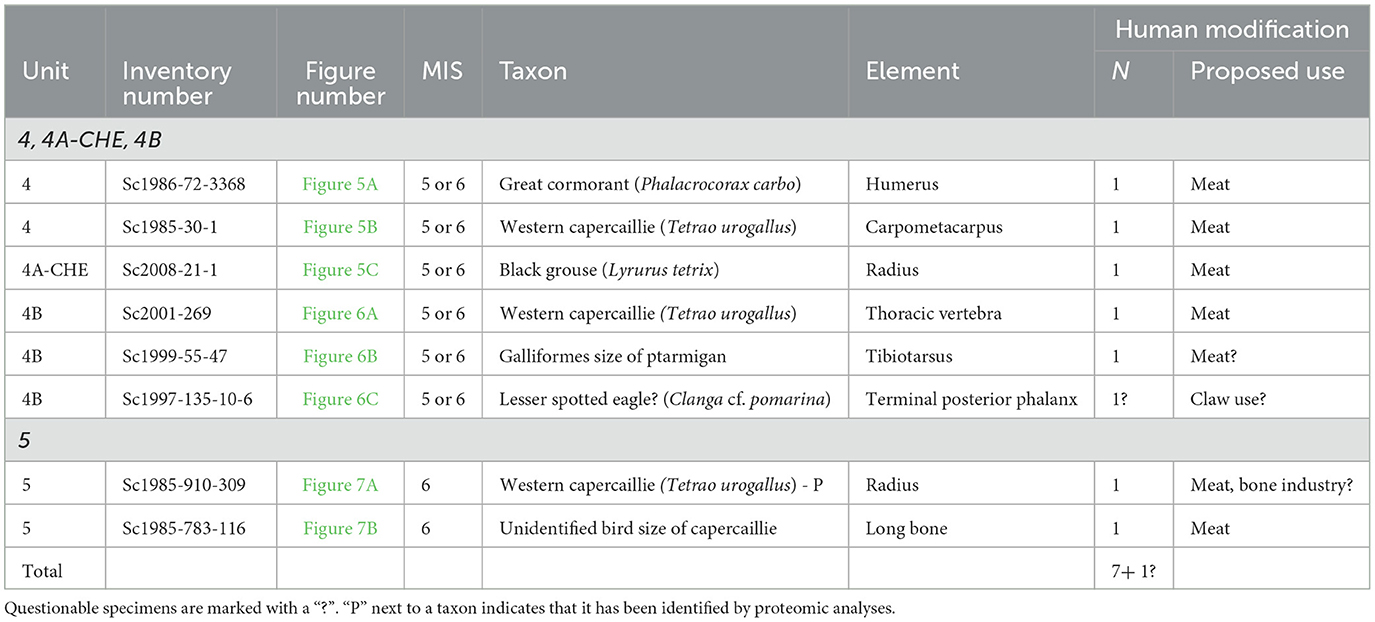
Table 2. Inventory of bird remains showing marks interpreted as anthropogenic in origin.
3.3.4.1 Cutmarks
3.3.4.1.1 Complex of units 4 (MIS 5 or 6)
In Unit 4 sensu lato, the left humerus of a great cormorant displayed two double incisions on the anterior face as well as an area of scraping on the posterior face; these marks were located on the body of the bone, near the distal end (Figure 5A). The two double incisions were located at the level of the brachialis muscle insertion fossa, and penetrate into it (Figure 5A', arrow). This location is consistent with butchering activities (Laroulandie, 2001) and does not support interpretation as trampling. The scraping area on the posterior surface was likely linked to the removal of meat. Furthermore, a group of fairly superficial transverse striations was located near the fracture, on the proximal part of the body of the bone. These could be marks linked to the removal of meat but trampling cannot be excluded.
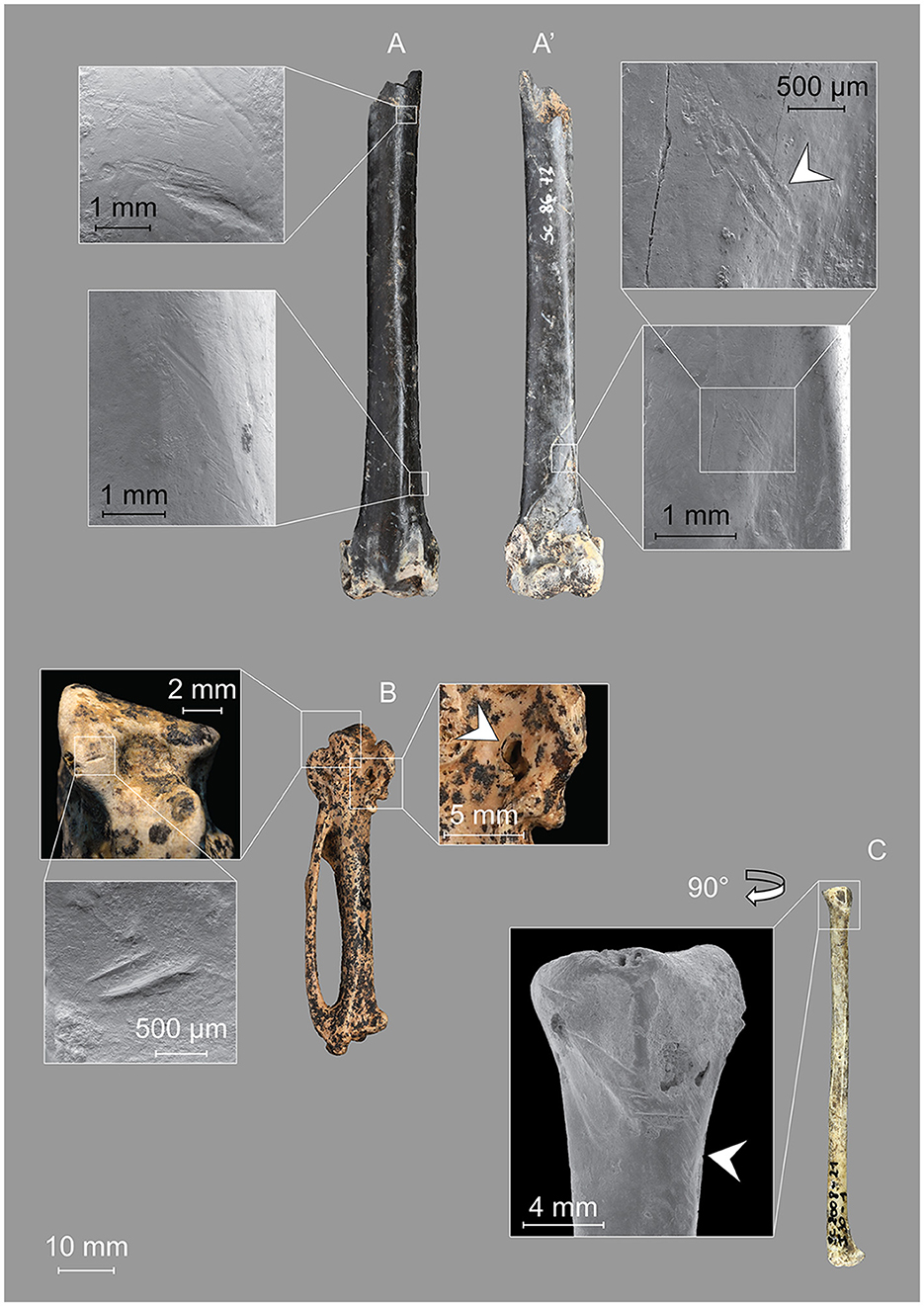
Figure 5. Units 4 and 4A-CHE. (A) Great cormorant, left distal humerus showing incisions and scraped areas (Sc1986-72-3368; Unit 4). Two double incisions enter the brachialis muscle insertion fossa (arrow). (B) Western capercaillie, right carpometacarpus with two short incisions at the proximal joint (Sc1985-30-1; Unit 4). Two tooth impressions, or one with a bilobed appearance, are present (arrow). (C) Black grouse, right radius with several deep incisions and a scraped area (arrow) in the proximal part (Sc2008-21-1; Unit 4A-CHE).
A right carpometacarpus of a Western capercaillie bears two incisions on the ventral side of the articular surface of the proximal condyle (Figure 5B), probably left during the disarticulation of the wrist (Laroulandie, 2001). Imprints of teeth with a bilobed appearance on the processus extensorius suggest the action of a small carnivore (Figure 5B, arrow).
3.3.4.1.2 Unit 4A-CHE (MIS 5 or 6)
The complete right radius of a black grouse found in Unit 4A-CHE exhibited several deep, oblique incisions on the posterior side at the level of the proximal joint, as well as a scraped area on the ventral side (Figure 5C). These tool marks were concentrated near the facies articularis ulnaris and were probably produced during the disarticulation of the radius and ulna during meat recovery (Laroulandie, 2001).
3.3.4.1.3 Unit 4B (MIS 5 or 6)
Unit 4B provided two Galliformes bones with anthropogenic marks. The first was a thoracic vertebra of a Western capercaillie bearing a deep incision on one of the lateral faces accompanied by a second, more superficial incision, both aligned and most likely resulting from the same movement (Figure 6A). A left distal tibiotarsus of a ptarmigan-sized taxon showed longitudinal and oblique scraping marks on the posterior and medial surfaces of the body of the bone (Figure 6B). They are partly under the varnish applied to accommodate the archaeological marking. Their purpose is unclear, and may have been produced during the harvesting of the meat.

Figure 6. Unit 4B. (A) Western capercaillie, thoracic vertebra bearing a deep incision accompanied by a second, more superficial incision, in the same alignment (Sc2001-269; Unit 4B). (B) Ptarmigan-sized Galliformes, left distal tibiotarsus with fine longitudinal and oblique incisions on the body of the bone probably resulting from the removal of meat (Sc1999-55-47; Unit 4B). (C) Lesser spotted eagle (?), terminal posterior phalanx of the second toe showing areas of intense polish (Sc1997-135-10-6; Unit 4B).
3.3.4.1.2 Unit 5 (MIS 6)
In Unit 5, anthropogenic marks were identified on two bones of large birds (10.5% of the bones from Unit 5). The first was analyzed by proteomics and identified as a Western capercaillie (see above). It is a fragment of the body of a radius fractured at both ends (Figure 7A). One of these fractures displays a modern aspect and is associated with residues left by a metal tool. This fracture was likely produced during the excavation. The fracture on the other side of this element is strongly polished and associated with two deep incisions. The absence of metal particles close to the incisions and the presence of black manganese deposits on the inside exclude that they were produced recently, during the excavation or later. Particular attention was paid to this bone, especially to reconstruct the sequence of actions responsible for the marks observed.

Figure 7. Unit 5. (A) Western capercaillie, right radius shaft presenting a very deep incision accompanied by a second parallel, more superficial incision (Sc 1985-910-309). One of the fractures shows a mark of peeling (arrow). The fracture edge is strongly rounded and shows a very developed polish (detail view). (B) Large bird, unidentified bone fragment with longitudinal incisions and superficial scraped areas (Sc1985-783-116).
First, the two deep incisions were produced. These marks could bear witness to butchery practices, but their location, far from a joint, is inconsistent with the disarticulation of the carcass, and their depth is unusual in the context of meat removal. Instead, they could be associated with a form of bone work and may have been intended to facilitate the fracture of this radius. The depth and breadth of the largest incision, in particular, suggests a repeated “sawing” type movement, which supports the hypothesis that these incisions were produced to help fracture the bone. Second, the bone has been broken transversally by flexion, which produced a flaking of the bone surface. The fracture progression was stopped by the incisions. Third, the bone has been used for a prolonged time, which resulted in the development of a strong polish on the broken edge (see Section 3.3.4.2.2.).
The second specimen is a small fragment of the body of a long bone from an unidentified large bird the size of a Western capercaillie. The specimen displays fine longitudinal incisions and several scraped areas in the form of groups of oblique striae (Figure 7B). The marks observed suggest different finalities. The longitudinal incisions probably relate to the action of cutting, and oblique striae are likely scraping marks left during butchering activities. Trampling is probably responsible for the fragmentation of the original bone into this tiny fragment and for the production of faint, randomly oriented striae observed at the surface of this specimen.
3.3.4.2 Polish
Two bones show strong polish.
3.3.4.2.1 Unit 4B (MIS 5 or 6)
A claw (phalanx 3) of the second digit of a likely lesser spotted eagle has very fine polish on the reliefs of the articular surface as well as on the area underlying the keratinous sheath of the phalanx (Figure 6C). These polishes are associated with micro-striations (visible at 800 × ), which could suggest human intervention. The absence of polish in the hollow parts of the affected areas tends to exclude digestion or post-depositional processes such as polishing by movements in the sediments, but further research on talon polish is required to ascertain this assumption. The fractured distal end does not show rounding nor polish. No tool marks were observed. The attribution of these modifications to human activities remains uncertain.
3.3.4.2.2 Unit 5 (MIS 6)
The incised Western capercaillie radius fragment described above shows a highly developed polish (Figure 7A). This polish is located at the apical part of one of the two fractured ends and does not affect the entire surface of the bone. The polish is spread over the area where the peeling is observed, which excludes that the fracture and the flaking of the bone surface occurred after the development of the polish. At high magnification (800 × ), a few fine streaks are visible. It was not possible to determine if the polish resulted from the use of this element as a tool dedicated to a specific action, or if the polish was produced unintentionally during the prolonged transport of the object. Anyway, the use of this specimen produced very fine micro-wear associated with short and multiple orientations micro-striations located close to the edge. These characteristics exclude that it was used on strongly abrasive materials such as stone or dry skin, but instead suggest a use on soft material such as fresh skin or plants. However, more comparative research is needed to determine the kind of material which generated the polish observed.
3.4 Spatial distribution
The bird bones bearing anthropogenic modifications were plotted onto a map of the cave (Supplementary Figures 3–6). Their location was based on the square of origin only, as the exact location within the square is not always known. We also examined the spatial distribution for Unit 1A because it constitutes one of the two main archaeological assemblages from the Middle Paleolithic, although it delivered no bird bones with anthropogenic modifications. Until now, evaluations of spatial distribution have hardly been attempted for the Scladina Cave, all types of remains combined. This limited the possibility of comparisons with other remains.
The spatial distribution of all avian remains should be interpreted with caution because the sediments were all reworked and none of the remains are in primary position. This is especially the case in 4A-CHE (very dynamic erosion and transport). Therefore, the spatial distribution of avian remains in no way represents the pattern of deposition of bird bones within the cave, and cannot inform us about behavior, be it Neanderthal or non-human predators.
3.4.1 Unit 1A (MIS 3)
In Unit 1A, the provenance square of 15 out of 16 avian remains is known. The remains were distributed evenly throughout the cave and do not appear to be deposited in any organized concentration (Supplementary Figure 3). Some fragments were found close to, and even within, a spread of burned mammal bones described by Abrams et al. (2010). However, there is no greater evidence of a concentration of avian remains here than elsewhere in the cave. Mechanical predator marks were identified throughout the cave, while digestion modifications were only identified in specimens located toward the back of the cave.
3.4.2 Complex of Units 4, Unit 4A-CHE and Unit 4B (MIS 5 and 6)
The bird bones from these units are presented on the same map, despite each sedimentary unit originating from a different set of processes (Supplementary Figure 4). The map shows the spatial position of the remains of the Neanderthal child within Unit 4A-CHE. There is a relative proximity between the bird bones with anthropogenic modifications and the human remains but they are not from the same grid squares and the sedimentary unit itself represents an erosion gully.
3.4.3 Unit 5 (MIS 6)
The bird bones from Unit 5 are mainly present within the first 25 meters from the cave opening (Supplementary Figures 5, 6). Specimens found in this area also bear marks made by small carnivores. Two concentrations can be distinguished in the distribution of avian remains: one closer to the entrance of the cave (10–13 m), and a second further into the cave (18–23 m). Both concentrations contain a specimen bearing anthropogenic modifications and are located close to numerous lithic artifacts and anthropogenically modified mammal remains. Analysis of the spatial distribution of lithics and faunal remains did not reveal any conclusive concentrations, and indicates instead that the archaeological remains in this unit were widely dispersed, perhaps due to the post-depositional disturbance by mammals frequenting the cave when Neanderthals were not present (Bonjean, 1998; Patou-Mathis and Lopez-Bayon, 1998). However, the main sedimentary process responsible for the deposition of Unit 5, i.e., solifluction (Pirson, 2007, 2014), is also at least partly responsible for the spatial distribution of the material. Two bird bones bearing evidence of non-human predators were recovered from a natural alcove within the rock where the Scladina Cave joins with Saint-Paul Cave (30–33 m). A further specimen bearing digestion marks also appeared at this depth, isolated from other avian remains.
4 Discussion
4.1 Exploitation of birds by Neanderthals
The analysis of the avifauna of Scladina Cave provides the first testimonies of bird exploitation by Neanderthals in Belgium, extending the geographic area of dietary bird exploitation more than 600 km to the north. In total, 6 bird bones from MIS 5 or 6 (including 1 uncertain), and 2 from MIS 6 bear anthropogenic modifications.
The initial stratigraphic position of the bones unearthed from Unit 4A-CHE is uncertain, because they were reworked by the gully from other layers. In some situations, the stratigraphic position of the bones excavated from former Units 4 (4A and 4B) can also be uncertain, because of the complexity of this part of the sequence deciphered during the 2007 stratigraphic reappraisal (in former layers 4A and 4B, the new system implies 21 layers grouped into 5 Units). This is particularly true for bones initially attributed to Unit 4B. Given (1) the scarcity of bone remains in the whole Unit 4B, (2) the sedimentary dynamic of this unit (dominated by decantation in a pool covering a large surface inside the cave), and (3) the discrepancy between the environmental signal of certain bird taxa identified and that reconstructed by palynology for Unit 4B (see Section 4.3), it is unlikely that these objects truly came from Unit 4B. It seems plausible that they were mistakenly assigned to sedimentary unit 4B, as they were unearthed before the stratigraphic reappraisal which pointed to the existence of a deep gully (Unit 4A-CHE) eroding Unit 4B, and therefore lying at the same altitude. This hypothesis is all the more plausible because a bird bone initially attributed to Unit 4B has been reattributed to Unit 4A-CHE, thanks to careful re-examination of the available section drawings and pictures of the area, and thanks to the coordinates of the piece. This proves that at least some of the elements originally attributed to Unit 4B actually come from the gully, which seems much more consistent with the environmental significance of forest taxa identified as 4A-CHE reworked the temperate Sub-Units 4A-AP and 4A-IP, which correspond to MIS 5e (Eemian). However, we cannot exclude that some of these bones truly came from Unit 4B, or that some specimens unearthed from 4A-CHE come from Unit 5 (MIS 6), which delivered one of the two main Middle Paleolithic assemblages, as it was sporadically eroded by the 4A-CHE gully. This latter hypothesis nevertheless appears unlikely for the tawny owl bone, since this species favors temperate climate while palynology reconstructs cold environmental conditions in Unit 5. For all these reasons, for some of the studied specimens, we will only mention former Units 4, 4A or 4B, implying a chronostratigraphic attribution to the end of MIS 6 and/or the first part of MIS 5.
In Units 4, 4A-CHE, and 4B (MIS 5 or 6) the exploited taxa are the Western capercaillie, the great cormorant, the black grouse, another species of small grouse, and possibly the likely lesser spotted eagle. The exploitation of the great cormorant indicates predation at the edge of a waterbody or a watercourse of a given breadth, perhaps located at the level of the current Meuse Valley. The two grouse species may have been captured in woodlands, or an open environment in the case of the black grouse. The striae observed denote butchery activities, except for the lesser spotted eagle talon which exhibits a strong polish possibly related to manipulation by hominins. Two bones of large birds from Unit 5 (MIS 6) bear marks of tools also compatible with the removal of meat. However, the two deep incisions observed on the radius of a Western capercaillie suggest a sawing action which implies possible working of the bone. In addition, this element presents a highly developed polish on a broken edge which indicates that the bone was utilized for an unknown purpose.
Several Middle Paleolithic sites from southern Europe have revealed features of disarticulation and/or flesh removal on meat-bearing avian elements, suggesting consumption of the flesh. It is possible that some of these marks were produced during the recovery of other products such as tendons (Blasco et al., 2019), in particular when bones bearing little meat are affected. In general, remains of birds bearing marks of butchery activities are rare within the same site (Blasco et al., 2022), and this is confirmed in the case of Scladina. However, the sites of Bolomor, Cova Negra, Gorham's cave, Pié Lombard, and Fumane yielded more substantial assemblages (Peresani et al., 2011; Blasco and Fernández Peris, 2012; Finlayson et al., 2012; Blasco et al., 2013, 2014, 2016; Fiore et al., 2016; Romero et al., 2017). From a geospatial point of view, Blasco et al. (2022) indicated regional variations in the taxa exploited: the use of pigeons and partridges of the genus Alectoris seems to be specific to the Mediterranean Region, while diurnal birds of prey, corvids, and birds from the Anatidae family are exploited elsewhere in Europe. The consumption of the Western capercaillie by Neanderthals at Scladina proves to be the first direct evidence of exploitation of this species of the Galliformes order, unlike the black grouse that has been identified in Neanderthal contexts previously.
The assemblage from the Scladina Cave mainly includes birds from the order Galliformes. This does not stand out in comparison with other Middle Paleolithic sites where species of the Galliformes genus Alectoris are frequently exploited (Martínez Valle et al., 2016; Romero et al., 2017). Rather it suggests that Alectoris exploitation in the Mediterranean region is part of an overall dietary trend toward the exploitation of Galliformes in general, and that the exploitation of certain genera was constrained only by species distribution. Neanderthals exploited whichever species were available in the Pleistocene landscape whatever the latitude band.
The exploitation of the riparian great cormorant emphasizes the Neanderthals' ability to exploit a large range of game available to them in the vicinity of Scladina Cave. Indeed, despite being less useful as a proxy for paleoclimatic reconstruction, the ecological information from exploited taxa still informs us about the presence of a variety of habitats close to Scladina Cave at the time of original deposition. This aligns well with established preferences in human habitat choice (Finlayson et al., 2011). The presence of cormorants, among several other aquatic bird species, has also been reported as the result of Neanderthal activities at Gruta da Figueira Brava (Nabais et al., 2023). The avian contribution to the human diet at Scladina Cave indicates that Neanderthals in MIS 5 and MIS 6 exploited woodland and wetland environments close to the cave. While some wetland species identified amongst the assemblage, like ducks, might be found in the small river valley close to the cave, others such as the great cormorant were more likely to frequent the larger waters running at the location of today's river Meuse. Extensive forested areas with large openings in the surroundings hosted Western capercaillie and black grouse. Knowledge and understanding of prey availability in this diverse ecotonal habitat testifies to Neanderthal cognitive complexity in regard to hunting and subsistence strategies.
Some sites have yielded phalanges of birds bearing marks of tools. The phalanges, especially the distal phalanx carrying the keratinous sheath of the talon, are devoid of nutritional value, implying an objective other than food (Morin and Laroulandie, 2012; Radovčić et al., 2015; Rodríguez-Hidalgo et al., 2019; Romandini et al., 2014). All phalanges modified during the Middle Paleolithic or Middle-to-Upper Paleolithic transition are of large diurnal birds of prey, at the exception of an incised swan phalanx unearthed from a Mousterian context at Baume de Gigny (Mourer-Chauviré, 1975) and an eagle owl phalanx found in the Châtelperronian layers of the Grotte du Renne (Vanhaeren et al., 2019). The possible manipulation of a talon from a likely lesser spotted eagle at Scladina Cave fits well into this pattern, although it remains hypothetical at this stage.
Examples of the use of hard materials from mammals by Neanderthals in tool-making are multiplying, with examples intended for working with skin, wood or lithic materials (for example, Gaudzinski, 1999; Burke and d'Errico, 2008; Mallye et al., 2012; Soressi et al., 2013; Tartar and Costamagno, 2016; Baumann et al., 2020, 2022). On the other hand, the shaping and use of bird bones has only been very rarely demonstrated for the Middle Paleolithic. The deep incision on the radius of a Western capercaillie from Scladina Cave presents a morphology reminiscent of incisions observed on the radius of a common raven discovered at the site of Zaskalnaya, Crimea (Majkić et al., 2017). The Zaskalnaya specimen shows marks of cutting on the anterior surface and seven parallel notches on the posterior surface of the bone, distributed over a length of approximately 1 cm. These notches are considered to have decorative value, although their production may have initially been utilitarian in purpose, such as ensuring a good grip on the bone. Indeed, the notches were produced in a series of events, some having been added to fill a void, which is interpreted as aesthetic desire (Majkić et al., 2017). Currently, there is no argument to suggest any decorative intention in the incisions on the Scladina Cave specimen, but it is interesting to note the morphological similarities. The Scladina radius also shows strong polish associated with micro-striations that resembles those identified on the fracture edge of an incised black vulture carpometacarpus at Fumane Cave and interpreted as possible use-wear produced during the utilization of the bone (Peresani et al., 2011).
4.2 Non-human accumulating agents
All bird remains in the occupationally scant complex of Units 3 displayed some indication of carnivore agency. It is also the only context group where all types of carnivore modification were visible on the bone surfaces. The remains were of Galliformes (MNI = 2; a Western capercaillie and a black grouse-sized bird) that were likely predated close to the cave. The remains were limited to an area with an alcove at the point where Scladina and Saint-Paul caves merge. The bird remains from complex of Units 3 were accumulated by a small to medium carnivore and indicated denning behavior through their distribution. The mammalian fauna identified in Scladina Cave is rich in carnivores, including large mammals such as cave bears (Ursus spelaeus), wolves (Canis lupus), and cave hyenas (Crocuta crocuta spelea) (Simonet, 1992; Patou-Mathis, 1998; Daujeard et al., 2016; Table 6.3). However, taphonomy indicates that the carnivores responsible for the partial accumulation of bird remains in Scladina were smaller. Smaller carnivores such as dhole (Cuon sp.), red fox (Vulpes vulpes), badger (Meles meles), wild cat (Felis silvestris), and mustelid remains in the sequence are prime candidates for the agent behind the mechanical and digestive marks identified on bone surfaces in complex of Units 3 (Daujeard et al., 2016; Marin-Monfort et al., 2019; Royer et al., 2021). However, this is inconsistent with the majority of digestion recorded in the avian assemblage of Scladina, which is more indicative of bird of prey action.
Though small carnivores hunted birds, they may also have scavenged the discarded remains of human meals. This behavior may be surmised from the presence of both cutmarks and subsequent carnivore tooth marks on the bone of a Western capercaillie in Sedimentary Complex 4A. In Unit 5, avian remains with carnivore marks accounted for 52.6% of the sample, but there was no evidence of carnivores scavenging Neanderthal leftover bird carcasses. Instead, rodent gnawing was present on the discarded bone of a grouse-sized bird. In the natural alcove utilized by carnivores in complex of Units 3, a co-occurrence of remains suggests its use as a den at some point during the deposition of Unit 5. The mammalian record shows the presence of canids and bears during this phase (Patou-Mathis, 1998; Daujeard et al., 2016), but the bird bones do not reflect predation by large mammalian carnivores. This is not unexpected as larger carnivores would eat the whole carcass and digest all the bones (Andrews, 1990).
The spatial proximity of certain owl (Strigiformes) remains with the main arch of the cave in Unit 5 (MIS 6) might suggest that these were the result of attritional accumulation in connection to roosts and nests. They were unlikely to have roosted when the cave was occupied by Neanderthals (Bocheński, 2005; Bocheński et al., 2018). There is no mechanical evidence of raptors catching birds in the recovered material, but a study of digestion marks on the molars of micromammals from MIS 5 and MIS 3 determined that two classes of Strigiform predator were responsible for their accumulation (López-García et al., 2017). Digestion profiles found on the avian remains, along with the presence of three species of owl remains (Bubo sp., Strix aluco, Asio sp.) in the assemblage align well with their findings, however digestion traces are infrequent in the avian sample, suggesting that avian predators were not the main accumulation agent at Scladina. There are also clear taphonomic traces pointing toward small carnivore agency in complex of Units 3. Diurnal raptor remains in Unit 1A (n = 2) were found in an area between 35 and 40 meters from the cave entrance. Neither the large eagle (Aquila sp.) nor the sparrowhawk (Accipiter nisus) specimen exhibited identifiable marks of human agency, and sedimentary dynamics could easily explain their position in the cave and they are therefore not considered likely agents of accumulation.
4.3 Environment and climate
The bird remains reflect the presence of both open and wooded environments throughout the deposition of the Middle Paleolithic sediments (MIS 6–MIS 3) with intermittent wetland signals in the form of Suliformes and Anseriformes. This could indicate the presence of an ecotonal, mosaic type landscape in the vicinity of Scladina Cave during the Middle Paleolithic, and/or a dynamic shifting of habitats throughout time. Western capercaillie and black grouse would have frequented boreal forest environments around the cave, while the less arboreal ptarmigans would have occupied a more open, tundra-like environment (Cramp and Simmons, 1977-1994). Waterfowl such as ducks and geese could have exploited the nearby water bodies. Depending on the species, ducks can be highly adaptable and exploit any water body available. Geese however may have been drawn to the islands on the River Meuse for breeding and inland water bodies. The islands along the Meuse also provide a nesting habitat for the great cormorant (Phalacrocorax carbo). This insular nesting behavior is seen in the species today, which is still nesting on islands in the Meuse Valley nowadays.
These observations are supported by pollen analyses (Pirson, 2007; Pirson et al., 2008, 2014) and studies of various authors on the small and large mammals (Cordy, 1992; Simonet, 1992; Moncel, 1998; Patou-Mathis, 1998; Daujeard et al., 2016; López-García et al., 2017). Herbs and forbs are most frequent, in both Unit 1A (MIS 3) and Unit 5 (MIS 6), signaling open areas. However, these signals are highly variable between sedimentary units. The presence of boreal conifers in both stages also alludes to patches of forest (Pirson, 2007; Pirson et al., 2008). Micromammals from MIS 3 (Unit 1A) indicate cold and dry conditions (Lagurus lagurus, Lemmus lemmus, Dicrostonyx torquatus, Microtus arvalis, and Chionomys nivalis). In MIS 6 the species representation indicated sporadic peaks in aquatic local environments (Microtus agrestis, Talpa europaea, Sorex gr. araneus, and Microtus gregalis) (López-García et al., 2017).
However, despite the seeming correlation between proxies, it is important to note that some of the avian remains may have been reworked into their current position by sedimentary and/or post-depositional processes. In Unit 1A, the presence of a European sparrowhawk and a probable wood pigeon, both forest-dwelling taxa, could be due to more recent intrusions given that converging signals document the deposition of this unit in a cold steppe environment (Pirson et al., 2008). However, as mentioned above, pollen signals do attest to the presence of boreal conifers in this period, and both the wood pigeon and Eurasian sparrowhawk are multi-latitudinal semi-generalist to generalist species that can adapt to dynamic bioclimatic parameters (Finlayson et al., 2011).
The numerous remains of Western capercaillie (n = 23) and a single bone of a tawny owl (Figure 2A) attributed to Unit 4B are also forest-dwelling taxa. However, this unit records one of the harshest climate conditions in the entire sequence (Pirson et al., 2008, 2014). Therefore, as stated before, these remains were most likely erroneously attributed to Unit 4B and come from the gully, which reworked temperate layers characterized by a forest environment more compatible with these woodland taxa, especially the tawny owl (see Section 4.1).
5 Conclusion
Despite complex sedimentogenesis along with small sample size, the bird assemblage from Scladina Cave has proven highly informative as a proxy for Neanderthal behavior. Results demonstrate that Neanderthals navigating the dynamic Pleistocene landscapes of northwestern Europe in MIS 6 and 5 exploited birds as part of their diet. Additionally, they utilized bird bones as tools and may have collected talons for symbolic purposes. The avian prey spectrum at Scladina indicates a Neanderthal population with a profound understanding of available habitats and the avian game available to them with prey-species representing woodland and semi-open habitats, as well as wetlands. Our results from Scladina Cave contribute two novel species to the spectrum of known prey species during the Middle Paleolithic, and provide new geospatial data from northwestern Europe to the growing number of sites reporting bird exploitation by Neanderthals.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Author contributions
QG: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Visualization, Writing – original draft, Writing – review & editing. VR: Conceptualization, Investigation, Resources, Validation, Visualization, Writing – original draft, Writing – review & editing. GA: Conceptualization, Data curation, Investigation, Methodology, Resources, Writing – original draft, Writing – review & editing. SP: Conceptualization, Data curation, Investigation, Methodology, Resources, Visualization, Writing – original draft, Writing – review & editing. KD: Conceptualization, Data curation, Investigation, Methodology, Resources, Writing – original draft, Writing – review & editing. FB: Formal analysis, Funding acquisition, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. DC: Formal analysis, Investigation, Visualization, Writing – original draft, Writing – review & editing. DB: Data curation, Investigation, Resources, Writing – original draft, Writing – review & editing. LA: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The work of Q. Goffette partly took place within the framework of a partnership between the Royal Belgian Institute of Natural Sciences and the Agence Wallonne du Patrimoine. The work of L. Amos was financed thanks to a PhD Scholarship from John Moores University Faculty of Science (2017 to 2021). Access to the CNRS research infrastructure Infranalytics (FR2054) is gratefully acknowledged. V. Rots acknowledges support from the F.R.S.-FNRS.
Acknowledgments
We are grateful to C. Vandenbulcke (Espace muséal d'Andenne) for his help with the sorting of the bone material.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fearc.2024.1441926/full#supplementary-material
References
Abrams, G. (2018). “Palaeolithic bone retouchers from Belgium: a preliminary overview of the recent research through historic and modern bone collections”, in The Origins of Bone Tool Technologies, eds. J.M. Hutson, A. García-Moreno, E.S. Noack, E. Turner, A. Villaluenga and S. Gaudzinski-Windheuser (Mainz: Verlag des Römisch-Germanisches Zentralmuseums - Leibniz-Forschungsinstitut für Archäologie), 197–214.
Abrams, G., Bello, S. M., Di Modica, K., Pirson, S., and Bonjean, D. (2014). When Neanderthals used cave bear (Ursus spelaeus) remains: Bone retouchers from unit 5 of Scladina Cave (Belgium). Quarter. Int. 326, 274–287. doi: 10.1016/j.quaint.2013.10.022
Abrams, G., Bonjean, D., Di Modica, K., Pirson, S., Otte, M., and Patou-Mathis, M. (2010). Les os brûlés de l'ensemble sédimentaire 1A de Scladina (Andenne, Belgique): apports naturels ou restes de foyer(s) néandertalien(s)? Notae Praehistor. 30, 5–13.
Abrams, G., Devièse, T., Pirson, S., De Groote, I., Flas, D., Jungels, C., et al. (2024). Investigating the co-occurrence of Neanderthals and modern humans in Belgium through direct radiocarbon dating of bone implements. J. Hum. Evol. 186:103471. doi: 10.1016/j.jhevol.2023.103471
Bacher, A. (1967). Vergleichend morphologische Untersuchungen an Einzelknochen des postkranialen Skeletts in Mitteleuropa vorkommender Schwäne und Gänse. PhD thesis, München, Universität München.
Barisic, M. (2006). Étude de l'effet du piétinement humain sur des ossements de faune mélangés à un amas de taille. PhD thesis, Bordeaux, Université Bordeaux 1, UMR PACEA 5199.
Baumann, M., Plisson, H., Rendu, W., Maury, S., Kolobova, K., and Krivoshapkin, A. (2020). The Neandertal bone industry at Chagyrskaya cave, Altai Region, Russia. Quater. Int. 559, 68–88. doi: 10.1016/j.quaint.2020.06.019
Baumann, M., Ready, E., Plisson, H., Maury, S., Vanderesse, N., Coqueugniot, H., et al. (2022). Not so unusual Neanderthal bone tools: new examples from Abri Lartet, France. Archaeol. Anthropol. Sci. 14:200. doi: 10.1007/s12520-022-01674-4
Baumel, J. J. (1993). Handbook of Avian Anatomy: Nomina Anatomica Avium. Cambridge: Publications of the Nuttall Ornithological Club 23.
Behrensmeyer, A. K. (1978). Taphonomic and ecologic information from bone weathering. Paleobiology 4, 150–162. doi: 10.1017/S0094837300005820
Behrensmeyer, A. K., Stayton, C. T., and Chapman, R. E. (2003). Taphonomy and ecology of modern avifaunal remains from Amboseli Park, Kenya. Paleobiology 29, 52–70. doi: 10.1666/0094-8373(2003)029<0052:TAEOMA>2.0.CO;2
Blain, H.-A., López-García, J. M., Cordy, J.-M., Pirson, S., Abrams, G., Di Modica, K., et al. (2014). Middle to late Pleistocene herpetofauna from Scladina and Sous-Saint-Paul caves (Namur, Belgium). Comptes Rendus Palevol. 13, 681–390. doi: 10.1016/j.crpv.2014.03.006
Blasco, R., Cochard, D., Colonese, A. C., Laroulandie, V., Meier, J., Morin, E., et al. (2022). “Small animal use by Neanderthals,” in Updating Neanderthals: understanding behavioural complexity in the Late Middle Palaeolithic, eds F. Romagnoli, F. Rivals and S. Benazzi. (London: Academic Press, Elsevier), 123–143. doi: 10.1016/B978-0-12-821428-2.00010-X
Blasco, R., and Fernández Peris, J. (2009). Middle Pleistocene bird consumption at Level XI of Bolomor Cave (Valencia, Spain). J. Archaeol. Sci. 36, 2213–2223. doi: 10.1016/j.jas.2009.06.006
Blasco, R., and Fernández Peris, J. (2012). Small and large game: Human use of diverse faunal resources at Level IV of Bolomor Cave (Valencia, Spain). Comptes Rendus Palevol. 11, 265–282. doi: 10.1016/j.crpv.2012.01.003
Blasco, R., Fernández Peris, J., and Rosell, J. (2008). Estrategias de subsistencia en los momentos finales del Pleistoceno medio: el nivel XII de la Cova del Bolomor (La Valldigna, Valencia). Zephyrvs 62, 63–80.
Blasco, R., Finlayson, C., Rosell, J., Marco, A. S., Finlayson, S., Finlayson, G., et al. (2014). The earliest pigeon fanciers. Sci. Rep. 4, 1–7. doi: 10.1038/srep05971
Blasco, R., Rosell, J., Domínguez-Rodrigo, M., Lozano, S., Past,ó, I., Riba, D., et al. (2013). Learning by heart: cultural patterns in the faunal processing sequence during the middle pleistocene. PLoS ONE 8:e55863. doi: 10.1371/journal.pone.0055863
Blasco, R., Rosell, J., Rufà, A., Sánchez Marco, A., and Finlayson, C. (2016). Pigeons and choughs, a usual resource for the Neanderthals in Gibraltar. Quater. Int. 421, 62–77. doi: 10.1016/j.quaint.2015.10.040
Blasco, R., Rosell, J., Sánchez-Marco, A., Gopher, A., and Barkai, R. (2019). Feathers and food: Human-bird interactions at Middle Pleistocene Qesem Cave, Israel. J. Hum. Evol. 136:102653. doi: 10.1016/j.jhevol.2019.102653
Bocheński, Z. M. (2005). “Owls, diurnal raptors and humans: signatures on avian bones,” in Biosphere to Lithosphere: new studies in vertebrate taphonomy, ed. (Oxford: Oxbow Books), 31–45. doi: 10.2307/j.ctt1kw294s.8
Bocheński, Z. M., and Tomek, T. (1997). Preservation of bird bones: erosion versus digestion by owls. Int. J. Osteoarchaeol. 7, 372–387. doi: 10.1002/(SICI)1099-1212(199707/08)7:4<372::AID-OA355>3.0.CO;2-W
Bocheński, Z. M., Tomek, T., and Wertz, K. (2009). The state of preservation of bird remains in a cave environment: a case study from Ciemna Cave (Poland). Acta Zool. Cracov. 52, 1–16.
Bocheński, Z. M., Tomek, T., and Wertz, K. (2022). Whose talon is this? A manual for the identification of ungual phalanges of European accipitrid birds of prey. Int. J. Osteoarchaeol. 33, 1–17. doi: 10.1002/oa.3165
Bocheński, Z. M., Wertz, K., and Tomek, T. (2018). What happens to dead birds left in a cave and its vicinity for a quarter of a century? A taphonomic experiment. Histor. Biol. 30, 855–867. doi: 10.1080/08912963.2017.1335721
Bonjean, D. (1998). “Répartition spatiale de l'industrie lithique,” in Recherches aux grottes de Sclayn. Volume 2. L'archéologie, eds. M. Otte, M. Patou-Mathis, and D. Bonjean (Liège : Études et Recherches archéologiques de l'Université de Liège 79), 340–376.
Bonjean, D., Abrams, G., and Di Modica, K. (2012). Grotte Scladina, une preuve de la chasse au lièvre au Paléolithique moyen. Chronique de l'Archéologie Wallonne 19, 209–211.
Bonjean, D., Vanbrabant, Y., Abrams, G., Pirson, S., Burlet, C., Di Modica, K., et al. (2015). A new Cambrian black pigment used during the late Middle Palaeolithic discovered at Scladina Cave (Andenne, Belgium). J. Archaeol. Sci. 55, 253–265. doi: 10.1016/j.jas.2014.11.040
Bourdillat, V. (2008). Hommes, carnivores? Caractériser l'action de l'hyène des cavernes: de l'utilisation des données fossiles pour l'interprétation des sites mixtes. PhD thesis, Paris, Muséum national d'Histoire naturelle.
Bray, F., Fabrizi, I., Flament, S., Locht, J. L., Antoine, P., Auguste, P., et al. (2023). Robust high-throughput proteomics identification and deamidation quantitation of extinct species up to pleistocene with ultrahigh-resolution MALDI-FTICR mass spectrometry. Anal. Chem. 95, 7422–7432. doi: 10.1021/acs.analchem.2c03301
Burke, A., and d'Errico, F. (2008). A middle palaeolithic bone tool from crimea (Ukraine). Antiquity 82, 843–852. doi: 10.1017/S0003598X00097611
Cnuts, D., Peresani, M., and Rots, V. (2022). The contribution of stone tool residues in reconstructing Late Pleistocene hominin stone tool behaviour at Grotta di Fumane, Italy. Quat. Sci. Rev. 297:107829. doi: 10.1016/j.quascirev.2022.107829
Codlin, M. C., Douka, K., and Richter, K. K. (2022). An application of zooms to identify archaeological avian fauna from Teotihuacan, Mexico. J. Archaeol. Sci. 148:105692. doi: 10.1016/j.jas.2022.105692
Cordy, J.-M. (1992). “Bio- et chronostratigraphie des dépôts quaternaires de la grotte Scladina (province de Namur, Belgique) à partir des micromammifères,” in Recherches aux grottes de Sclayn, Volume 1. Le contexte, ed M. Otte (Liège: Études et Recherches archéologiques de l'Université de Liège 27), 79–125.
Cramp, S., and Simmons, K.E.L. (1977-1994). The Birds of the Western Palearctic. Oxford, New York: Oxford University Press.
Daujeard, C., Abrams, G., Germonpré, M., Le Pape, J. M., Wampach, A., Di Modica, K., et al. (2016). Neanderthal and animal karstic occupations from southern Belgium and South-eastern France: Regional or common features? Quater. Int. 411, 179–197. doi: 10.1016/j.quaint.2016.02.009
Delagnes, A., and Rendu, W. (2011). Shifts in Neandertal mobility, technology and subsistence strategies in western France. J. Archaeol. Sci. 38, 1771–1783. doi: 10.1016/j.jas.2011.04.007
Dewez, M. (1981). Les grottes de Sclayn: ensemble rituel moustérien ou repaire d'ours? Activités SOS Fouilles 2, 79–83.
Di Modica, K. (2010). Les productions lithiques du Paléolithique moyen de Belgique: variabilité des systèmes d'acquisition et des technologies en réponse à une mosaïque d'environnements contrastés. PhD thesis, Liège, Université de Liège and Muséum d'Histoire Naturelle de Paris.
Di Modica, K., and Bonjean, D. (2004). Scladina (Sclayn, province de Namur): ensembles lithiques moustériens méconnus. Notae Praehistoricae 24, 5–8.
Di Modica, K., Toussaint, M., Abrams, G., and Pirson, S. (2016). The Middle Palaeolithic from Belgium: Chronostratigraphy, territorial management and culture on a mosaic of contrasting environments. Quater. Int. 411, 77–106. doi: 10.1016/j.quaint.2015.12.072
Dibble, H. L., Berna, F., Goldberg, P., Mcpherron, S.-P., Mentzer, S., Niven, L., et al. (2009). A preliminary report on Pech de l'Azé IV, Layer 8 (Middle Paleolithic, France). Paleoanthropology 2009, 182–219. doi: 10.4207/PA.2008.ART30
Domínguez-Rodrigo, M., de Juana, S., Galán, A. B., and Rodríguez, M. (2009). A new protocol to differentiate trampling marks from butchery cut marks. J. Archaeol. Sci. 36, 2643–2654. doi: 10.1016/j.jas.2009.07.017
Draily, C., Toussaint, M., and Pirson, S. (2011). “La grotte Walou à Trooz (Belgique),“ in Fouilles de 1996 à 2004. Volume 2. Les sciences de la vie et les datations. Namur, Service public de Wallonie (Études et documents – Archéologie 21).
Eda, M., Morimoto, M., Mizuta, T., and Inoué, T. (2020). ZooMS for birds: Discrimination of Japanese archaeological chickens and indigenous pheasants using collagen peptide fingerprinting. J. Archaeol. Sci. 34:102635. doi: 10.1016/j.jasrep.2020.102635
Ellwood, B. B., Harrold, F. B., Benoist, S. L., Thacker, P. T., Otte, M., Bonjean, D., et al. (2004). Magnetic susceptibility applied as an age-depth-climate relative dating technique using sediments from Scladina Cave, a Late Pleistocene cave site in Belgium. J. Archaeol. Sci. 31, 283–293. doi: 10.1016/j.jas.2003.08.009
Erbersdobler, K. (1968). Vergleichend morphologische Untersuchungen an Einzelknochen des postkranialen Skeletts in Mitteleuropa vorkommender mittelgroßer Hühnervögel. PhD thesis, München, Universität München.
Fernández-Jalvo, Y., and Andrews, P. (2016). Atlas of Taphonomic Identifications. 1001+ Images of Fossil and Recent Mammal Bone Modification. The Netherlands: Springer. doi: 10.1007/978-94-017-7432-1
Finlayson, C., Brown, K., Blasco, R., Rosell, J., Negro, J. J., Bortolotti, G. R., et al. (2012). Birds of a feather: neanderthal exploitation of raptors and corvids. PLoS ONE 7:e45927. doi: 10.1371/journal.pone.0045927
Finlayson, C., Carrión, J., Brown, K., Finlayson, G., Sánchez-Marco, A., Fa, D., et al. (2011). The Homo habitat niche: using the avian fossil record to depict ecological characteristics of Palaeolithic Eurasian hominins. Quat. Sci. Rev. 30, 1525–1532. doi: 10.1016/j.quascirev.2011.01.010
Fiore, I., Gala, M., Romandini, M., Cocca, E., Tagliacozzo, A., and Peresani, M. (2016). From feathers to food: reconstructing the complete exploitation of avifaunal resources by Neanderthals at Fumane cave, unit A9. Quater. Int. 421, 134–153. doi: 10.1016/j.quaint.2015.11.142
Flannery, K. V. (1969). “Origins and ecological effects of early domestication in Iran and the Near East,” in The Domestication and Exploitation of Plants and Animals, eds. P. J. Ucko and G. W. Dimbleby (Chicago: Aldine), 73–100. doi: 10.4324/9781315131825-7
Garcia-Fermet, T., Testu, A., Moigne, A., Saos, T., and Grégoire, S. (2023). The bird remains from La Crouzade Cave (Gruissan, Aude): the mixed origin of a Middle Palaeolithic bone accumulation. Quarter. Environ. Hum. 1:100001. doi: 10.1016/j.qeh.2023.100001
Gaudzinski, S. (1999). Middle Palaeolithic bone tools from the open-air site Salzgitter-Lebenstedt (Germany). J. Archaeol. Sci. 26, 125–141. doi: 10.1006/jasc.1998.0311
Gaudzinski-Windheuser, S., and Niven, L. (2009). “Hominin subsistence patterns during the middle and late Paleolithic in Northwestern Europe,” in The Evolution of Hominin Diets. Vertebrate Paleobiology and Paleoanthropology, eds J. J. Hublin, M. P. Richards (Dordrecht: Springer). doi: 10.1007/978-1-4020-9699-0_7
Gill, F., Donsker, D., and Rasmussen, P. (2022). IOC World bird list (v 12.1). Available at: https://www.worldbirdnames.org/new/ioc-lists/master-list-2/ (accessed March 23, 2022).
Goffette, Q., Germonpré, M., Lefèvre, C., Brecko, J., Goemaere, E., and Rots, V. (2020). Bird bones from Trou de Chaleux and the human exploitation of birds during the late Magdalenian in Belgium. J. Archaeol. Sci. 29:102096. doi: 10.1016/j.jasrep.2019.102096
Goffette, Q., Lepers, C., Jadin, I., and Rots, V. (2023). A handful of duck radiuses: Peculiarities of the avifaunal exploitation at the Gravettian site of Maisières-Canal (Belgium). Int. J. Osteoarchaeol. 33, 668–682. doi: 10.1002/oa.3194
Gómez-Olivencia, A., Sala, N., Sanchis, A., and Arlegi, M. (2018). First data of Neandertal bird and carnivore exploitation in the Cantabrian Region (Axlor; Barandiaran excavations; Dima, Biscay, Northern Iberian Peninsula). Sci. Rep. 8, 1–14. doi: 10.1038/s41598-018-28377-y
Gullentops, F., and Deblaere, C. (1992). “Érosion et remplissage de la grotte Scladina,” in Recherches aux grottes de Sclayn, Volume 1. Le contexte, ed. M. Otte (Liège, Études et Recherches archéologiques de l'Université de Liège 27), 9–31.
Haesaerts, P. (1992). “Les dépôts pléistocènes de la terrasse de la grotte Scladina à Sclayn (province de Namur, Belgique),” in Recherches aux grottes de Sclayn, Volume 1. Le contexte, ed. M. Otte (Liège, Études et Recherches archéologiques de l'Université de Liège 27),33–55.
Hardy, B. L., and Moncel, H. (2011). Neanderthal use of fish, mammals, birds, starchy plants and wood 125-250,000 years ago. PLoS ONE 6:e23768. doi: 10.1371/journal.pone.0023768
Hardy, B. L., Moncel, M., Daujeard, C., Fernandes, P., Béarez, P., Desclaux, E., et al. (2013). Impossible Neanderthals? Making string, throwing projectiles and catching small game during Marine Isotope Stage 4 (Abri du Maras, France). Quat. Sci. Rev. 82, 23–40. doi: 10.1016/j.quascirev.2013.09.028
Hockett, B., and Haws, J. A. (2002). Taphonomic and methodological perspectives of leporid hunting during the Upper Paleolithic of the Western Mediterranean Basin. J. Archaeol. Method Theory 9, 269–302. doi: 10.1023/A:1019503030246
Hoffecker, J. F., and Cleghorn, N. E. (2000). Mousterian hunting patterns in the Northwestern Caucasus and the ecology of the Neanderthals. Int. J. Osteoarchaeol. 10, 368–378. doi: 10.1002/1099-1212(200009/10)10:5<368::AID-OA555>3.0.CO
Hoffmann, D. L., Angelucci, D. E., Villaverde, V., Zapata, J., and Zilhão, J. (2018). Symbolic use of marine shells and mineral pigments by Iberian Neandertals 115,000 years ago. Sci. Adv. 107, 1023–1028. doi: 10.1126/sciadv.aar5255
Hutson, J. M., García-Moreno, A., Noack, E. S., Turner, E., Villaluenga, A., and Gaudzinski-Windheuser, S. (2018). The Origins of Bone Tool Technologies. Mainz: Verlag des Römisch-Germanisches Zentralmuseum - Leibniz-Forschungsinstitut für Archäologie.
Kraft, E. (1972). Vergleichend morphologische Untersuchungen an Einzelknochen nord- und mitteleuropäischer kleinerer Hünhervögel. PhD thesis, München, Universität München.
Langer, G. (1980). Vergleichend morphologische Untersuchungen an Einzelknochen in Mitteleuropa vorkommender mittelgrosser Eulenarten. PhD thesis, München, Universität München.
Laroulandie, V. (2001). ≪ Les traces liées à la boucherie, à la cuisson et à la consommation d'oiseaux. Apport de l'expérimentation, ≫ in Préhistoire et approche expérimentale, eds L. Bourguignon, I. Ortega, and M.-C. Frère-Sautot, (Montagnac: Éditions Monique Mergoil), 97–108.
Laroulandie, V., Faivre, J., Gerbe, M., and Mourre, V. (2016). Who brought the bird remains to the Middle Palaeolithic site of Les Fieux (Southwestern, France)? Direct evidence of a complex taphonomic story. Quater. Int. 421, 116–133. doi: 10.1016/j.quaint.2015.06.042
Lloveras, L., Garcia, L., Maroto, J., Soler, J., and Soler, N. (2018). The bird assemblage from the Middle Palaeolithic level I of Arbreda Cave: a taphonomic story. J. Archaeol. Sci. 21, 758–770. doi: 10.1016/j.jasrep.2018.08.040
Loodts, I. (1998). “Une approche comportementale de l'Homme de Neandertal. L'industrie lithique de la couche 1A de la grotte Scladina, économie des matières premières et coexistence des chaînes opératoires au Paléolithique moyen récent,” in Recherches aux grottes de Sclayn. Volume 2. L'archéologie, eds M. Otte, M. Patou-Mathis, and D. Bonjean (Liège : Études et Recherches archéologiques de l'Université de Liège 79), 69–71.
López-García, J. M., Blain, H., Cordy, J.-M., Pirson, S., Abrams, G., Di Modica, K., et al. (2017). Palaeoenvironmental and palaeoclimatic reconstruction of the Middle to Late Pleistocene sequence of Scladina Cave (Namur, Belgium) using the small-mammal assemblages. Histor. Biol. 29, 1125–1142. doi: 10.1080/08912963.2017.1288229
López-García, J. M., Blain, H.-A., Goffette, Q., Cousin, C., and Folie, A. (2024). Environmental and climatic inferences for Marine Isotope Stage 2 of southern Belgium (Meuse valley, Namur Province) based on rodent assemblages. Palaeogeogr. Palaeoclimatol. Palaeoecol. 636:111976. doi: 10.1016/j.palaeo.2023.111976
López-González, F., Grandal-d'Anglade, A., and Vidal-Romaní, J. R. (2006). Deciphering bone depositional sequences in caves through the study of manganese coatings. J. Archaeol. Sci. 33, 707–717. doi: 10.1016/j.jas.2005.10.006
Lyman, R. L. (1994). Vertebrate Taphonomy. Cambridge: Cambridge University Press. doi: 10.1017/CBO9781139878302
Majkić, A., Evans, S., Stepanchuk, V., Tsvelykh, A., and d'Errico, F. (2017). A decorated raven bone from the Zaskalnaya VI (Kolosovskaya) Neanderthal site, Crimea. PLoS ONE 12:e0173435. doi: 10.1371/journal.pone.0173435
Mallye, J.-B., Thiébaut, C., Mourre, V., Costamagno, S., Claud, É., and Weisbecker, P. (2012). The Mousterian bone retouchers of Noisetier Cave: experimentation and identification of marks. J. Archaeol. Sci. 39, 1131–1142. doi: 10.1016/j.jas.2011.12.018
Marin-Monfort, M. D., García-Morato, S., Olucha, R., Yravedra, J., Piñeiro, A., Barja, I., et al. (2019). Wildcat scats: taphonomy of the predator and its micromamal prey. Quat. Sci. Rev. 225:106024. doi: 10.1016/j.quascirev.2019.106024
Martínez Valle, R., Guillem Calatayud, P. M., and Villaverde Bonilla, V. (2016). Bird consumption in the final stage of Cova Negra (Xátiva, Valencia). Quater. Int. 421, 85–102. doi: 10.1016/j.quaint.2016.01.068
Maureille, B., Bayle, P., and Balzeau, A. (2016). Néandertal et ses morts. Néandertal à la loupe. Les Eyzies-de-Tayac: Musée National de Préhistoire 120–130.
Maureille, B., and Van Peer, P. (1998). A little known element concerning the burial of the first adult at La Ferrassie (Savignac-de-Miremont, Dordogne). Paléo 10, 291–301. doi: 10.3406/pal.1998.1141
Moncel, M.-H. (1998). “L'industrie lithique de la grotte Scladina (Sclayn). La couche moustérienne 1A,” in Recherches aux grottes de Sclayn. Volume 2. L'Archéologie, eds. M. Otte, M. Patou-Mathis, and D. Bonjean (Liège: Université de Liège) 103–112.
Morin, E., and Laroulandie, V. (2012). Presumed symbolic use of diurnal raptors by Neanderthals. PLoS ONE 7:e32856. doi: 10.1371/journal.pone.0032856
Morin, E., Meier, J., El Guennouni, K., Moigne, A. M., Lebreton, L., Rusch, L., et al. (2019). New evidence of broader diets for archaic Homo populations in the northwestern Mediterranean. Sci. Adv. 5 :eaav9106. doi: 10.1126/sciadv.aav9106
Mourer-Chauviré, C. (1975). Faunes d'oiseaux du pléistocène de France: Systématique, évolution et adaptation, interprétation paléoclimatique. Geobios 8, 333–352. doi: 10.1016/S0016-6995(75)80048-9
Nabais, M., Pimenta, C., and Zilhão, J. (2023). Human-bird interaction in last Interglacial Iberia: A combined approach using skeletal part analysis, bone surface modification, bird ethology and ethnography. J. Archaeol. Sci. 49:104023. doi: 10.1016/j.jasrep.2023.104023
Otte, M. (1992). Recherches aux grottes de Sclayn. Volume 1: Le contexte. Liège: Études et Recherches Archéologiques de l'Université de Liège.
Otte, M. (1998a). “Industrie lithique de la couche 5,” in Recherches aux grottes de Sclayn. Volume 2. L'archéologie, eds. M. Otte, M.Patou-Mathis, and D. Bonjean (Liège : Études et Recherches archéologiques de l'Université de Liège 79), 277–278.
Otte, M. (1998b). “Le Paléolithique supérieur,” in Recherches aux grottes de Sclayn. Volume 2. L'archéologie, eds M. Otte, M.Patou-Mathis, and D. Bonjean (Liège : Études et Recherches archéologiques de l'Université de Liège 79), 63–68.
Otte, M., Patou-Mathis, M., and Bonjean, D. (1998). Recherches aux grottes de Sclayn. Volume 2. L'archéologie. Liège: Études et Recherches Archéologiques de l'Université de Liège 79.
Patou-Mathis, M. (1998). “Les espèces chassées et consommées par l'homme en couche 5,” in Recherches aux grottes de Sclayn, Volume 2. L'archéologie, eds. M. Otte, M. Patou-Mathis, and D. Bonjean (Études et Recherches archéologiques de l'Université de Liège 79), 297–310.
Patou-Mathis, M., and Lopez-Bayon, I. (1998). “Analyse spatiale des ossements de la couche 5,” in Recherches aux grottes de Sclayn. Volume 2. L”archéologie, eds. M. Otte, M. Patou-Mathis, and D. Bonjean (Études et Recherches archéologiques de l'Université de Liège 79), 377-395.
Peresani, M., Fiore, I., Gala, M., Romandini, M., and Tagliacozzo, A. (2011). Late Neandertals and the intentional removal of feathers as evidenced from bird bone taphonomy at Fumane Cave 44 ky B.P., Italy. Proc. Nat. Acad. Sci. 108, 3888–3893. doi: 10.1073/pnas.1016212108
Pirson, S. (2007). Contribution à l'étude des dépôts d'entrée de grotte en Belgique au Pléistocène supérieur. Stratigraphie, sédimentogenèse et paléoenvironnement. PhD thesis, Liège, Université de Liège.
Pirson, S. (2011). “Contextes paléoenvironnemental et chronostratigraphique du remplissage de la grotte Walou: apport de la géologie et comparaison avec les autres disciplines,” in La grotte Walou à Trooz (Belgique). Fouilles de 1996-2004. Volume 1. Les sciences de la terre, eds S. Pirson, C. Draily, and M. Toussaint (Namur: Institut du Patrimoine Wallon), 170–206.
Pirson, S. (2014). “The stratigraphic sequence of Scladina Cave,” in The Scladina I-4A Juvenile Neandertal (Andenne, Belgium). Palaeoanthropology and Context, eds. M. Toussaint, and D. Bonjean (Liège: Études et Recherches archéologiques de l'Université de Liège 134), 49–68.
Pirson, S., Court-Picon, M., Damblon, F., Balescu, S., Bonjean, D., and Haesaerts, P. (2014). “The palaeoenvironmental context and chronostratigraphic framework of the Scladina Cave sedimentary sequence (units 5 to 3-SUP),” in The Scladina I-4A Juvenile Neandertal (Andenne, Belgium). Palaeoanthropology and Context, eds. M. Toussaint, and D. Bonjean (Liège: Études et Recherches archéologiques de l'Université de Liège 134), 69–92.
Pirson, S., Court-Picon, M., Haesaerts, P., Bonjean, D., and Damblon, F. (2008). New data on geology, anthracology and palynology from the Scladina Cave pleistocene sequence: preliminary results. Mem. Geol. Surv. Belgium 55, 71–93.
Pirson, S., Draily, C., and Toussaint, M. (2011). La grotte Walou à Trooz (Belgique). Fouilles de 1996 à 2004. Volume 1. Les sciences de la terre. Namur, Service public de Wallonie (Études et documents – Archéologie 20).
Pirson, S., Toussaint, M., Bonjean, D., and Di Modica, K. (2018). “Spy and Scladina caves: A Neandertal's story,” in Landscapes and landforms of Belgium and Luxemburg, ed. A. Demoulin (Cham: Springer, World Geomorphological Landscapes), 357–383. doi: 10.1007/978-3-319-58239-9_21
Rabinovich, R., and Tchernov, E. (1995). Chronological, paleoecological and taphonomical aspects of the Middle Paleolithic site of Qafzeh, Israel. Archaeozool. Near East 2, 5–44.
Radovčić, D., JapundŽić, D., Sršen, A. O., Radovčić, J., and Frayer, D. W. (2016). An interesting rock from Krapina. Comptes Rendus Palevol. 15, 988–993. doi: 10.1016/j.crpv.2016.04.013
Radovčić, D., Sršen, A. O., Radovčić, J., and Frayer, D. W. (2015). Evidence for neandertal jewelry: modified white-tailed eagle claws at krapina. PLoS ONE 10:e0119802. doi: 10.1371/journal.pone.0119802
Rendu, W., Beauval, C., Crèvecœur, I., Bayle, P., Balzeau, A., Bismuth, et al. (2014). Evidence supporting an intentional Neandertal burial at La Chapelle-aux-Saints. Proc. Nat. Acad. Sci. 111, 81–86. doi: 10.1073/pnas.1316780110
Rodríguez-Hidalgo, A., Morales, J. I., Cebrià, A., Courtenay, L. A., Fernández-Marchena, J. L., García-Argudo, G., et al. (2019). The Châtelperronian Neanderthals of Cova Foradada (Calafell, Spain) used imperial eagle phalanges for symbolic purposes. Sci. Adv. 5:eaax1984. doi: 10.1126/sciadv.aax1984
Rodríguez-Vidal, J., d'Errico, F., Pacheco, F. G., Blasco, R., Rosell, J., Jennings, R. P., et al. (2014). A rock engraving made by Neanderthals in Gibraltar. Proc. Nat. Acad. Sci. 111, 13301–13306. doi: 10.1073/pnas.1411529111
Romagnoli, F., Rivals, F., and Benazzi, S. (2022). Updating Neanderthals: understanding behavioural complexity in the Late Middle Palaeolithic. London: Academic Press, Elsevier.
Romandini, M., Fiore, I., Gala, M., Cestari, M., Guida, G., Tagliacozzo, A., et al. (2016). Neanderthal scraping and manual handling of raptors wing bones: evidence from Fumane Cave. Experimental activities and comparison. Quater. Int. 421, 154–172. doi: 10.1016/j.quaint.2015.12.078
Romandini, M., Peresani, M., Laroulandie, V., Metz, L., Pastoors, A., Vaquero, M., et al. (2014). Convergent evidence of eagle talons used by late neanderthals in europe: a further assessment on symbolism. PLoS ONE 9:e101278. doi: 10.1371/journal.pone.0101278
Romandini, M., Silvestrini, S., Real, C., Lugli, F., Tassoni, L., Carrera, et al. (2023). Late Neanderthal “menu” from northern to southern Italy: freshwater and terrestrial animal resources. Quat. Sci. Rev. 315:108233. doi: 10.1016/j.quascirev.2023.108233
Romero, A. J., Díez, J. C., and Brugal, J. P. (2017).. Aves de caza. Estudio tafonómico y zooarqueológico de los restos avianos de los niveles musterienses de Pié Lombard (Alpes-Maritimes, Francia). Munibe 68, 73–84. doi: 10.21630/maa.2017.68.14
Romero, A. J., Díez, J. C., Rodríguez, L., and Arceredillo, D. (2016). Anthropic fractures and human tooth marks: an experimental approach to non-technological human action on avian long bones. Quater. Int. 421, 219–227. doi: 10.1016/j.quaint.2015.10.005
Rots, V. (2013). Insights into early Middle Palaeolithic tool use and hafting in Western Europe. The functional analysis of level IIa of the early Middle Palaeolithic site of Biache-Saint-Vaast (France). J. Archaeol. Sci. 40, 497–506. doi: 10.1016/j.jas.2012.06.042
Rots, V. (2015). “Hafting and site function in the European Middle Paleolithic,” in Settlement Dynamics of the Middle Palaeolithic and Middle Stone Age (Vol. 4), eds. N.J. Conard and A. Delagnes (Tübingen: Kerns Verlag), 383–410.
Royer, A., Mallye, J.-B., Pelletier, M., and Griselin, S. (2021). Who killed the small mammals of ittenheim? An integrative approach and new taphonomic data for investigating bone assemblages accumulated by small carnivores. Quaternary 4:41. doi: 10.3390/quat4040041
Rufà, A., Blasco, R., Roger, T., and Moncel, M. (2016). What is the taphonomic agent responsible for the avian accumulation? An approach from the Middle and early Late Pleistocene assemblages from Payre and Abri des Pêcheurs (Ardèche, France). Quater. Int. 421, 46–61. doi: 10.1016/j.quaint.2015.05.016
Simonet, P. (1992). “Les associations de grands mammifères du gisement de la grotte Scladina à Sclayn (Namur, Belgique),” in Recherches aux grottes de Sclayn, Volume 1. Le contexte, ed. M. Otte (Liège: Études et Recherches archéologiques de l'Université de Liège 27), 9–31.
Soressi, M., and d'Errico, F. (2007). “Pigments, gravures, parures: les comportements symboliques controversés des Néandertaliens,” in Néandertaliens: biologie et cultures, eds. B. Vandermeersch, B. Maureille, and Y. Coppens (Paris: CTH, Documents Préhistoriques 23), 297–309.
Soressi, M., McPherron, S. P., Lenoir, M., DogandŽić, T., Goldberg, P., Jacobs, Z., et al. (2013). Neandertals made the first specialized bone tools in Europe. Proc. Nat. Acad. Sci. 110, 14186–14190. doi: 10.1073/pnas.1302730110
Soressi, M., Rendu, W., Texier, P.-J., Claud, É., D'Errico, F., Laroulandie, et al. (2008). “Pech-de-l'Azé I (Dordogne, France): nouveau regard sur un gisement moustérien de tradition acheuléenne connu depuis le XIXe siècle,” in Les sociétés Paléolithiques dans un Grand Sud-Ouest: nouveaux gisements, nouvelles méthodes, nouveaux résultats - Actes des journées décentralisées de la SPF des 24-25 novembre 2006, eds. J. Jaubert, J. G. Bordes and I. Ortega (Paris: Société Préhistorique française), 95–132.
Speth, J. D., and Tchernov, E. (2002). Middle Paleolithic tortoise use at Kebara cave (Israel). J. Archaeol. Sci. 29, 471–483. doi: 10.1006/jasc.2001.0740
Speth, J. D., and Tchernov, E. (2007). The Middle Paleolithic occupations at Kebara Cave: a faunal perspective. Kebara Cave, Mt. Carmel, Israel. Middle Upper Palaeol. Archaeol. 165:e260.
Stiner, M. C. (2001). Thirty years on the “Broad Spectrum Revolution” and paleolithic demography. Proc. Nat. Acad. Sci. 98, 6993–6996. doi: 10.1073/pnas.121176198
Stiner, M. C., and Munro, N. D. (2002). Approaches to prehistoric diet breadth, demography, and prey ranking systems in time and space. J. Archaeol. Method Theory 9, 181–214. doi: 10.1023/A:1016530308865
Stiner, M. C., Munro, N. D., and Surovell, T. A. (2000). The tortoise and the hare: small-game use, the broad-spectrum revolution, and Paleolithic demography. Curr. Anthropol. 41, 39–79. doi: 10.1086/300102
Stringer, C. B., Finlayson, J. C., Barton, R. N. E., Fernández-Jalvo, Y., Cáceres, I., Sabin, R. C., et al. (2008). Neanderthal exploitation of marine mammals in Gibraltar. Proc. Nat. Acad. Sci. 105, 14319–14324. doi: 10.1073/pnas.0805474105
Tartar, E., and Costamagno, S. (2016). “L'utilisation des matières osseuses au Moustérien,” in Néandertal à la loupe (Les Eyzies-de-Tayac, Musée national de Préhistoire).
Toussaint, M., Bonjean, D., and Pirson, S. (2014). “The Scladina I-4A juvenile Neandertal: a synthesis,” in The Scladina I-4A Juvenile Neandertal (Andenne, Belgium). Palaeoanthropology and Context, eds. M. Toussaint, and D. Bonjean (Liège: Études et Recherches archéologiques de l'Université de Liège 134), 409–418.
Toussaint, M., Olejniczak, A. J., El Zaatari, S., Cattelain, P., Flas, D., Letourneux, C., et al. (2010). The Neandertal lower right deciduous second molar from Trou de l'Abîme at Couvin, Belgium. J. Hum. Evol. 58, 56–67. doi: 10.1016/j.jhevol.2009.09.006
Toussaint, M., and Pirson, S. (2006). Neandertal Studies in Belgium: 2000–2005. Period. Biol. 108, 373–387.
Toussaint, M., Semal, P., and Pirson, S. (2011). “Les Néandertaliens du bassin mosan belge: bilan 2006-2011,” in Le Paléolithique moyen en Belgique. Mélanges Marguerite Ulrix-Closset, eds. M. Toussaint, K. Di Modica, and S. Pirson (Liège : Études et Recherches Archéologiques de l'Université de Liège 128 - Les Chercheurs de la Wallonie Hors-série 4), 149–196.
Toussaint, M., Verna, C., Le Cabec, A., Gómez-Robles, A., Draily, C., Richards, M. P., et al. (2017). The Late Neandertal permanent lower left third premolar from Walou Cave (Trooz, Belgium) and its context. Am. J. Phys. Anthropol. 164, 193–202. doi: 10.1002/ajpa.23252
Van Neer, W., and Wouters, W. (2011). “Les restes de poissons des niveaux du Paléolithique moyen et supérieur de la grotte Walou,” in La Grotte Walou à Trooz (Belgique). Fouilles de 1996 à 2004. Volume 2, eds. C. Draily, S., Pirson, M. Toussaint (Namur, Département du Patrimoine et Institut du Patrimoine wallon), 68–82.
Vanhaeren, M., d'Errico, F., Julien, M., Mourer-Chauviré, C., and Lozouet, P. (2019). “Les objets de parure,” in Le Châtelperronien de la grotte du Renne (Arcy-sur-Cure, Yonne, France). Les fouilles d'André Leroi-Gourhan (1949-1963), ed. M. Julien (Les Eyzies-de-Tayac: Musée National de Préhistoire), 259–285.
Villa, P., and Roebroeks, W. (2014). Neandertal demise: an archaeological analysis of the modern human superiority complex. PLoS ONE 9:e96424. doi: 10.1371/journal.pone.0096424
von den Driesch, A. (1976). A Guide to the Measurement of Animal Bones From Archaeological Sites. New York: Harvard.
Vonhof, H., Verheyden, S., Bonjean, D., Pirson, S., Weber, M., Scholz, D., et al. (2024). Improving the age constraints on the archeological record in Scladina Cave (Belgium): new speleothem U-Th ages and paleoclimatological data. Clim. Past Discuss. 2024, 1–30. doi: 10.5194/cp-2024-27
Wertz, K., Tomek, T., and Bocheński, Z. M. (2022). Whose talon is this? A manual for the identification of ungual phalanges of European birds of prey: falcons and owls. Int. J. Osteoarchaeol. 33, 562–576. doi: 10.1002/oa.3144
Woelfle, E. (1967). Vergleichend morphologische Untersuchungen an Einzelknochen des postcranialen Skelettes in Mitteleuropa vorkommender Enten, Halbgänse und Säger. PhD thesis, München, Universität München.
Woolfenden, G. E. (1961). Postcranial osteology of the waterfowl. Bull. State FL. State Museum, Biol. Sci. 6, 1–129. doi: 10.58782/flmnh.amoc3264
Zilhão, J., Angelucci, D. E., Daniel, F., Dayet, L., Douka, K., Higham, et al. (2010). Symbolic use of marine shells and mineral pigments by Iberian Neandertals. Proc. Nat. Acad. Sci. 107, 1023–1028. doi: 10.1073/pnas.0914088107
Keywords: archaeozoology, middle paleolithic, neanderthal-bird relationships, taphonomy, tool marks, traceology, use-wear analysis
Citation: Goffette Q, Rots V, Abrams G, Pirson S, Di Modica K, Bray F, Cnuts D, Bonjean D and Amos L (2024) Neanderthal exploitation of birds in north-western Europe: Avian remains from Scladina Cave (Belgium). Front. Environ. Archaeol. 3:1441926. doi: 10.3389/fearc.2024.1441926
Received: 31 May 2024; Accepted: 21 August 2024;
Published: 20 September 2024.
Edited by:
Mariana Nabais, Institut Català de Paleoecologia Humana i Evolució Social (IPHES), SpainReviewed by:
Anna Rufà, University of Algarve, PortugalVéronique Laroulandie, Centre National de la Recherche Scientifique (CNRS), France
Copyright © 2024 Goffette, Rots, Abrams, Pirson, Di Modica, Bray, Cnuts, Bonjean and Amos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Quentin Goffette, cWdvZmZldHRlQG5hdHVyYWxzY2llbmNlcy5iZQ==; Linda Amos, bGFtb3NAdWZnLnVuaS1raWVsLmRl
†These authors have contributed equally to this work and share first authorship