 Clara Boulanger1,2,3,4*
Clara Boulanger1,2,3,4* Stuart Hawkins4
Stuart Hawkins4 Ceri Shipton5
Ceri Shipton5 Thomas Ingicco3
Thomas Ingicco3 Anne-Marie Sémah3
Anne-Marie Sémah3 Sofia Samper Carro4,6
Sofia Samper Carro4,6 Sue O'Connor4,6
Sue O'Connor4,6- 1International Research Fellow, Japan Society for the Promotion of Science, Tokyo, Japan
- 2Department of Modern Society and Civilization, National Museum of Ethnology, Osaka, Japan
- 3UMR 7194 Histoire Naturelle de l'Homme Préhistorique, Muséum National d'Histoire Naturelle, Paris, France
- 4Archaeology and Natural History, School of Culture, History and Language, College of Asia and the Pacific, Australian National University, Canberra, ACT, Australia
- 5Institute of Archaeology, University College London, London, United Kingdom
- 6ARC Centre of Excellence for Australian Biodiversity and Heritage, Australian National University, Canberra, ACT, Australia
Homo sapiens were adept at fishing in a range of aquatic habitats by the time they left Africa and reached Southeast Asia ca. 73 kya. In the insular region of Wallacea, humans adapted to a significant maritime environment with sophisticated marine fishing methods and technology by at least 42 kya. However, despite a growing array of evidence suggesting an early inland terrestrial adaptation on large islands in this tropical region, there was previously no evidence of fishing in inland wetlands habitats on the depauperate islands of Wallacea. Here we present new evidence of both marine and freshwater fishing recovered from different occupation phases from the cave sites Laili (ca. 44.6–11.7 kya) and Matja Kuru 2 (ca. 40 kya to Late Holocene) on the island of Timor (Timor-Leste), located near significant riverine and lake environments respectively. This indicates that humans adapted to a wider range of aquatic habitats over time and space in Wallacea than previously thought and moved freely between inland and coastal habitats. Diversification of fishing strategies likely improved chances of survival in an island landscape with an impoverished suite of terrestrial vertebrates under changing climatic conditions.
Introduction
Aquatic environments were important in the evolution of Homo where abundant shellfish and fish resources were available to provide essential protein and fatty acid nutrients to support hominin brain development (Mellars et al., 2013; Archer et al., 2014; Joordens et al., 2014; Marean, 2014). Archaic species of Homo were clearly able to exploit sessile and easy to gather shellfish from riverine environments since at least 600 kya in Southeast Asia, as seen at Trinil, Java, Indonesia by H. erectus (Joordens et al., 2015). However, efficient exploitation of fish requires complex technologies and sophisticated fishing strategies designed in relation to the life habits of much more mobile and reactive prey than sessile invertebrates (Kirch and Dye, 1979; Masse, 1986; Butler, 1994; Ono and Addison, 2009; Boulanger et al., 2019; Ono et al., 2019; Boulanger, 2021).
The oldest evidence of fishing and fish consumption could have taken place in freshwater habitats of Africa, during the Lower Pleistocene, 1.95 Mya, in the Turkana Basin (Archer et al., 2014). At the site of Olduvai, Tanzania, early hominins may have regularly captured some freshwater catfish species that are easy to catch using very little technology (Stewart, 1994). In the Middle East, a large assemblage dominated by two species of large Cyprinidae (carp) showing evidence of cooking by 780,000 years ago has been recovered at the site of Gesher Benot Ya'aqov, Israel (Zohar et al., 2022). In Europe, it is possible that H. neanderthalensis occasionally exploited freshwater ecosystems and the fish and shellfish species that inhabit them. However, the origins of fish bone accumulations in cave site assemblages associated with this hominin species cannot be unequivocally demonstrated since birds of prey and carnivorous mammals significantly contributed to the accumulation of bones (Kettle et al., 2008; Russ, 2010; Guillaud et al., 2018, 2021). Only the Iberian Neandertals of Figueira Brava seem to have regularly exploited marine aquatic food between 106 and 86 kya (Zilhão et al., 2020). In summary, there is therefore no firm evidence for Neanderthals freshwater fishing (Guillaud et al., 2021).
The advanced cognitive capabilities of H. sapiens significantly facilitated their adaptability to a wider range of aquatic resources and environments before and after their dispersal out of Africa during the late Middle Pleistocene and early Late Pleistocene (Groucutt et al., 2015; Roberts and Stewart, 2018). The earliest unequivocal evidence for hominin fishing is in South Africa ca. 70,000 BP by H. sapiens, both in marine and freshwater habitats (Brooks et al., 1995; Feathers and Migliorini, 2001; Plug, 2006; van Niekerk, 2011; Mitchell, 2015). Yet it is in Island Southeast Asia (ISEA) that the origins of dedicated maritime foraging in the Indo-Pacific probably lie (Kirch and Dye, 1979). The current earliest discoveries, but disputed (see Wood, 2017 for Madjedbebe), of H. sapiens remains in Sunda by 73 kya (Westaway et al., 2017) and in Sahul by 65 kya (Clarkson et al., 2017), indicate the colonization of the insular Wallacean archipelago, situated between these two great continental shelves, was earlier than the oldest known H. sapiens sites (ca. 45 kya) thus far found in the region (Hawkins et al., 2017; Shipton et al., 2019). Zooarchaeological and stable isotope data suggest that early H. sapiens in Wallacea adapted their foraging behavior to marine shellfish and fish in extensive coral reef environments significantly earlier than it was previously thought (O'Connor et al., 2011; Kealy et al., 2020; Roberts et al., 2020, 2022; Boulanger, 2021). Central to this adaptation was a sophisticated and diverse array of marine fishing practices, such as trolling, line fishing, netting or gathering by the use of traps, following lunar cycles and seasons and using a variety of fishing gears such as bone fishing gorges, shell hooks, net sinkers and probably baskets and nets made of vegetal fibers, highlighting the ecological plasticity of our species (O'Connor et al., 2011, 2019; Samper Carro et al., 2016; Boulanger et al., 2019, 2023; Kealy et al., 2020; Boulanger, 2021).
The tropical forests of ISEA were once considered hostile and relatively depauperate in fauna by comparison with coastal habitats (Roberts and Stewart, 2018). However, the presence of early H. sapiens in inland Sumatra, Borneo, and Sulawesi suggests an early adaptation of behavioral and cultural strategies to best meet the challenges of the tropical rainforest environment (Glover, 1981; Barker et al., 2007; Amano et al., 2016; Westaway et al., 2017; Brumm et al., 2018; Ingicco et al., 2020; Ono et al., 2020). There is evidence for exploitation of freshwater shellfish at inland Sunda and ISEA sites as H. sapiens moved into riverine environments on Borneo (Piper and Rabett, 2009), Sulawesi (Glover, 1981; Brumm et al., 2018), and Timor (Glover, 1986; Hawkins et al., 2017) during the early settlement of this region in the Pleistocene. Isotopic evidence indicates an increasing reliance by H. sapiens on a more diverse array of terrestrial resources in Wallacea by the terminal Pleistocene (Roberts et al., 2020).
Freshwater ecosystems may have been crucial to the sustenance and development of foraging cultures moving inland along the waterways on the larger insular islands such as Timor and Sulawesi in Wallacea, as they were in Sunda and Sahul (Barker et al., 2007; Habgood and Franklin, 2008). Freshwater resources were likely more important at inland sites in Wallacea, where terrestrial fauna were relatively depauperate compared to the continental landmasses of Sunda and Sahul (O'Connor and Aplin, 2007). In Sahul, although the anthropogenic origin of sun perch (Macquaria ambigua) otolith accumulations has not been indubitably demonstrated, it is very likely that H. sapiens at Lake Mungo, Australia, consumed them as soon as they arrived on the site 46 kya (Bowler et al., 2003; Long et al., 2018). Early fishing adaptations to inland freshwater habitats at Lake Mungo, speaks to the full modernity of humans throughout their migration from Sunda to Sahul (Langley et al., 2011).
Globally, it appears that fishing using a variety of nets, spears, traps, and lines in these terrestrial aquatic habitats became more widespread during the Pleistocene-Holocene transition (Gorman, 1971; Gobalet and Fenenga, 1993; Piper and Rabett, 2009; Pargeter et al., 2017; Boulanger et al., 2019; Álvarez-Fernández et al., 2020). This coincided with H. sapiens settlement of riverine valleys to engage in sedentary pastoralism, where fishing appears as a secondary activity (Hongo, 1989; Boschian, 1993; Losey et al., 2012; Weber et al., 2013). Eventually carp aquaculture (Cyprinidae) developed around 8,000 years ago in China (Nakajima et al., 2019), but there is no evidence of this spreading to ISEA during prehistory. Only a few sites in the remote islands of the Pacific witness the systemization of inland fishing during the late Holocene (Butler, 2001; Bouffandeau et al., 2019). For a variety of reasons that likely include identification issues, there is little evidence for freshwater fishing practices in ISEA during the Pleistocene (Piper and Rabett, 2009; Hawkins et al., 2017), despite the availability of several endemic edible freshwater fish taxa in wetlands (Zakaria-Ismail, 1994; Göltenboth et al., 2006).
In this paper we investigate H. sapiens fishing practices in Wallacea after inland settlement beside riverine and lakeside terrestrial habitats. To this end we review the scarce evidence for freshwater fish remains from archaeological sites in tropical ISEA while incorporating new evidence from ichthyofaunal bone assemblages from two inland sites in Timor-Leste, Matja Kuru 2 (MK2) and Laili Cave. Laili (4.7 km inland), where human occupation dates between 44.6 and 11.7 kya (MIS 3 and Last Glacial Maximum (LGM)/early Holocene), is situated overlooking a large valley through which the Laleia River runs (Hawkins et al., 2017). MK2 (8 km inland), is dated from ca. 40 kya until the late Holocene but appears to be unoccupied from the onset of the LGM, 28-15 kya (Lambeck and Chappell, 2001) until the beginning of the Holocene. It is situated a few hundred meters from the edge of Lake Ira Lalaro at its full position, the largest freshwater lake in the island of Timor (O'Connor et al., 2014; Roberts et al., 2020) (Figures 1, 2).
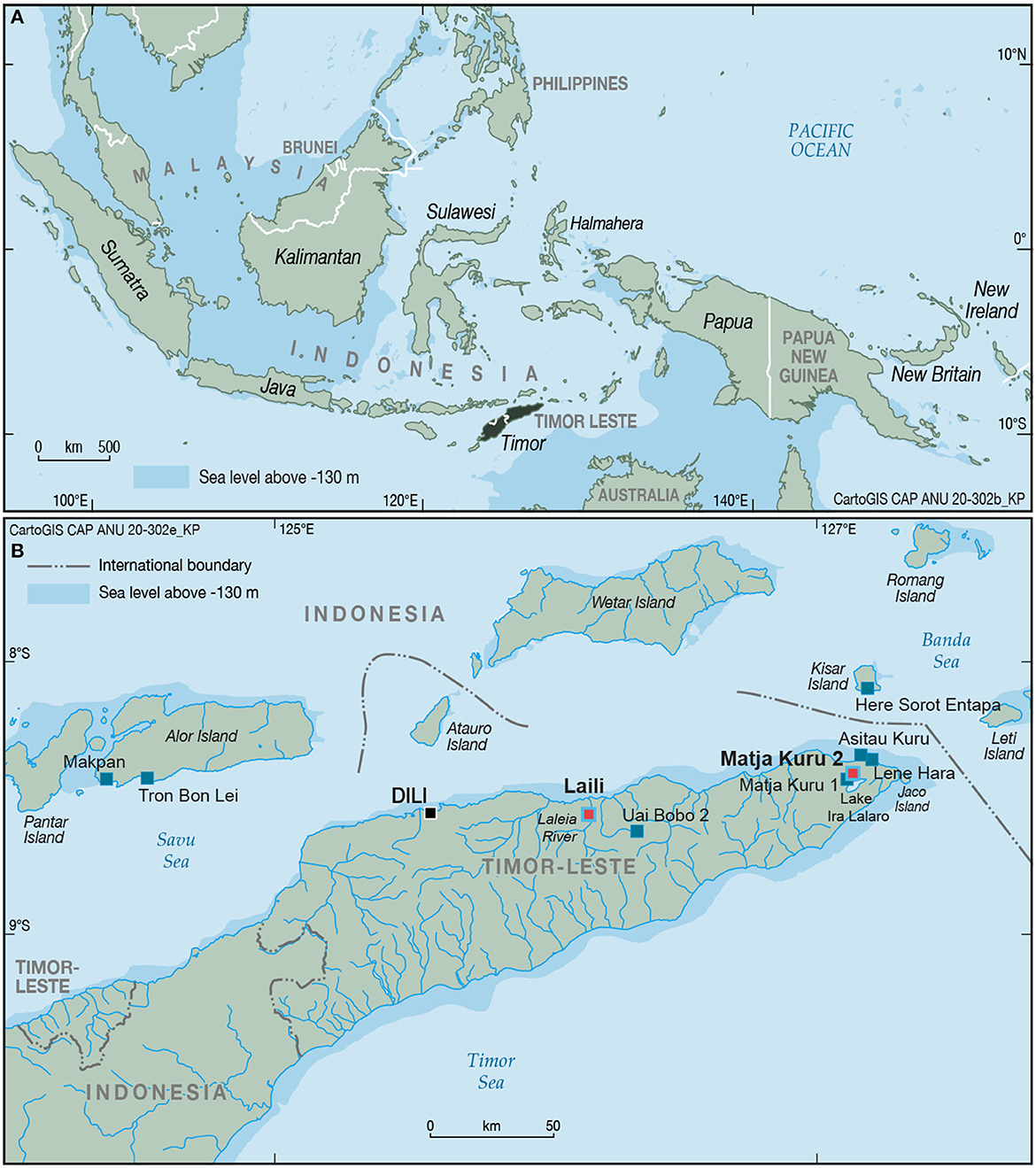
Figure 1. (A) Map of Island Southeast Asia (ISEA) showing the position of Timor Island. (B) Map of Timor-Leste with main freshwater ecosystems and nearby archaeological sites.

Figure 2. (A) Lake Ira Lalaro near Matja Kuru 2, Timor-Leste, June 2019 (Photo: S. Connor). (B) Laleia river near Laili, Timor-Leste, June 2019.
Materials and methods
Matja Kuru 2
MK2 is an uplifted limestone ridge located at a grid reference 8°24.88” S and 127°07.642” E north-east of the modern village of Poros and a few hundred meters from the shoreline of Lake Ira Lalaro during lake high stands. The site is located at an elevation of 370 m above mean sea level and around 8 km in straight-line distance from the north coast of Timor-Leste (Figure 1). MK2 is close to the site of Matja Kuru 1 located along the same ridge line (Langley et al., 2016). The surrounding vegetation is dominated by mesophyll vine and evergreen forests at the ridgeline, with an ecotone transition to savannahs and grasslands toward the lake and village (Roberts et al., 2020). In 2001, a 1 × 1 m test pit (Square D) was excavated by the Australian National University, but due to time constraints excavation was discontinued without reaching bedrock. In 2014, MK2 was reopened in order to extend the depth of the original excavation to bedrock and enlarge the excavation area. A 3 × 2 m area was excavated, taking in the original Square D, and including five new squares, DD, C, B, BB and AA (Roberts et al., 2020). The site deposit comprised a sandy silt sediment with likely a small clay component stratigraphic filling covering the end of the Pleistocene from ca. 40 kya until ca. 30 kya followed by a hiatus and a second phase of occupation from the early Holocene through to the late Holocene (Figure 3).
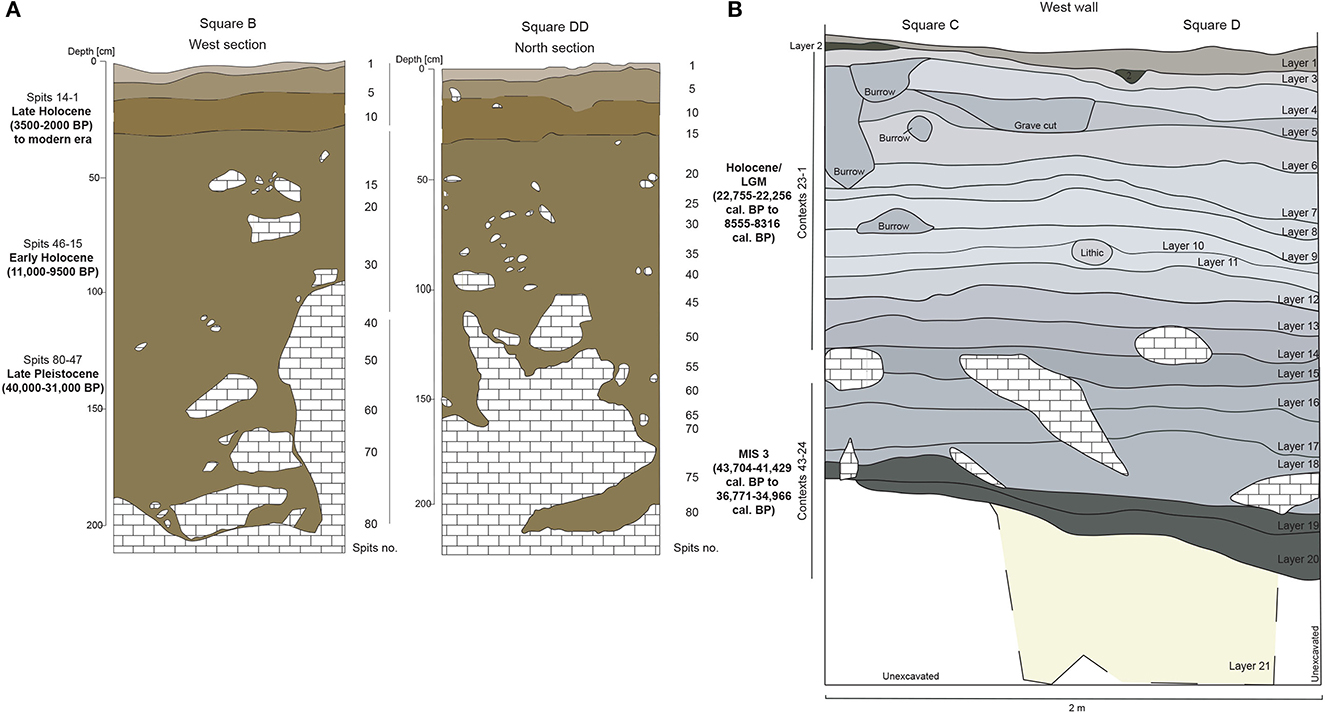
Figure 3. Stratigraphic profile of (A) Matja Kuru 2 Square B and DD using the Munsell Color Charts, according to Roberts et al. (2020), (B) Laili East wall (Shipton et al., 2021).
The first use of MK2 has an age estimate of 36,866–35,282 cal. BP (Langley et al., 2016). Phasing produced for MK2 based on archaeological and chronometric information is as follows: Spits 47–78 of Square B, Spits 47–53 of Square DD, and Spits 36–60 of Square D cover a Late Pleistocene period (ca. 40,000–31,000 years ago); Spits 15–46 of Square B and DD, and Spits 15–35 of Square D represent the Pleistocene/early Holocene transition (13,000–9,500 years ago); and Spits 1–14 in squares B, D and DD date to the late Holocene (3500–1000 years ago) (Roberts et al., 2020).
Based on analysis of the material remains from the original 2001 and the 2014 excavation undertaken thus far, stone artifacts and invertebrate marine fauna are most abundant in the Pleistocene levels, whereas vertebrate faunal remains are most abundant in the Holocene occupation (Veth et al., 2005; Samper Carro, 2023). Analysis of all the finds from both excavation seasons is nearing completion and will improve our understanding of the pattern of occupation at MK2. Terrestrial fauna includes introduced species such as Phalanger sp., Macaca sp., Sus sp., Cervus sp., Crocidura sp., but also giant rats of the genus Coryphomys, large and small rats of the genera Komodomys, Melomys and Rattus (Roberts et al., 2020). The presence of marine shellfish shows evidence for collection and transport over 8 km and discard of marine resources from initial occupation (Veth et al., 2005). The cave was therefore probably an attractive campsite around ca. 35,000 years ago, before abandonment from about 30,000 years and during the LGM, followed by reoccupation across the Pleistocene-Holocene transition and through into modern times (Veth et al., 2005; O'Connor and Aplin, 2007; Langley et al., 2016). Four human teeth dated to late and early Holocene were also recovered at MK2 (Roberts et al., 2020).
MK2, as well as the nearby archaeological sites of Matja Kuru 1, Asitau Kuru (Jerimalai), and Lene Hara, have delivered rare examples of symbolic material culture in ISEA such as shell beads made of Oliva spp. and Nautilus sp. dated back to 38,246–36,136 cal. BP (O'Connor, 2010; Langley and O'Connor, 2015, 2016; Langley et al., in press), and utilized ochre pieces (Langley and O'Connor, 2019). Evidence for fishing technology is sparse in these inland sites. At MK2 a modified sting ray barb (spit 27) (Boulanger et al., 2023) and the haft end of a large projectile dated to 36,500 to 34,500 cal. BP (O'Connor et al., 2014) that would have formed part of a composite spear have been suggested as utilized in fishing (O'Connor et al., 2014; Boulanger et al., 2023), and further examples of fishing technology may come to light when the bone and shell assemblage are fully analyzed. A stone-lined oven dated to around 10,000 years BP (Spits 25 to 19), and a dog burial found in Spit 25 and directly dated to 3075–2921 cal. BP, have also been reported from the site (Gonzalez et al., 2013; O'Connor, 2015).
Laili Cave
Laili is a partially collapsed cave located adjacent to the modern village of Laleia at a grid reference 8°32.25” S and 126°09.50” E, about 4.3 km from the north coast of Timor-Leste (Figure 1). It is formed in a limestone outcrop ca. 86 m above mean sea level and overlooks the braided plain of the Laleia River about 350 m to the east. The local surrounding environment around the site consists of open savannah woodlands and grasslands in a broad floodplain valley. Two 1 × 1 m squares (Square A and B) have been excavated by spits in the center of the cave platform by the Australian National University. While Square B was heavily disturbed, Square A produced a deep and undisturbed stratigraphic sequence ca. 2.2 m deep (Hawkins et al., 2017). In 2011 forty spits were excavated grouped into eleven stratigraphic units (Hawkins et al., 2017). Nineteen charcoal samples were collected and subjected to radiocarbon dating (Hawkins et al., 2017). Breccia deposits containing cultural remains such as marine shell and stone artifacts were cemented to the walls of the cave at heights well above the current floor of the deposit. Marine shell samples recovered from these breccia deposits have been dated to between 8,600 and 2,000 cal. BP, demonstrating that the cave was occupied during the Holocene. Three main chronostratigraphic periods were defined on the basis of these units and the radiocarbon dates: a terminal Pleistocene period dated between 17,468 and 11,161 cal. BP, another one representing the LGM dated between 22,500 and 18,500 cal. BP and an earliest period including pre-LGM deposits dates between 44,631 and 27,000 cal. BP (Hawkins et al., 2017). During the time of initial site use, the coast was ca. 4.7 km distant from Laili, that is, about 450 m further than it is today, while the coast was probably 5 km distant from Laili during the peak of the LGM ca. 18 kya ago (Hawkins et al., 2017).
The 2019 excavation at Laili expanded the excavation adjacent to Square A by 3 square meters: Square C west toward the rockshelter wall and Squares D and E to the north to give a total excavation size of 2 × 2 m. Two main occupation phases were differentiated in this excavation, Contexts 43 to 24 (Layers 20–16) spanning the middle part of MIS 3 (43,704–41,429 cal. BP to 36,771–34966 cal. BP, both dates on marine shells), and Contexts 23 to 3 (Layers 15–3) spanning the LGM (22,755–22,256 cal. BP) to the initial Holocene (8,555–8,316 cal. BP), while Contexts 2–1 are modern disturbance (Figure 3) (Shipton et al., 2021). Vertebrate NISP is most abundant during the early phase of site use and appears to decline over time. At least sixteen distinct mammal taxa were identified across the sequence, including human (H. sapiens), juvenile dog (Canis familiaris) in a disturbed layer, four extinct small rat species and four extinct giant rats (Muridae), at least one fruit bat (Pteropodidae), and at least six insectivorous bat taxa (Vespertilionidae). Seven bird taxa (Anseriformes, Charadriiformes, Columbiformes, Galliformes and Passeriformes) were identified and at least one species of frog/toad (Anura). Four reptile taxa were represented by small lizard (Lacertilia), sea turtle (Chelonioidea), freshwater turtle (Testudines), and snake (Serpentes). Molluscs (Mollusca) occur throughout the occupation sequence. At least 41 species of them were identified from a range of habitats including marine, mudflat/mangrove and freshwater environments. Crustaceans (Decapoda) were also identified from habitats including terrestrial, marine, and freshwater environments. Fish (Osteichthyes) from the first field season were represented by two taxa, freshwater eels (Anguillidae), and parrotfishes (Scaridae) in small quantities and from mostly unidentified fragments of vertebrae, spines, and ribs. They are predominantly found in the earlier phase and show a sharp decline during the LGM (Hawkins et al., 2017). Laili Square A produced 28,324 flaked stone artifacts, making this the largest excavated collection of stone artifacts per unit volume from Pleistocene ISEA. Artifacts were identified throughout the sequence, with the highest frequencies found from the end of the LGM to the onset of the Holocene. The assemblage is dominated by high quality, locally abundant chert in a variety of colors. Chert nodules can be found in eroding terraces within 0.5 km of the site and make up a sizable portion of the Laleia River gravels (Hawkins et al., 2017). At Laili no fishing technology has been identified.
Ichthyoarchaeological analysis
Fish remains from MK2 Square B and DD were analyzed, as well as the fish remains from Laili Squares C-E. All excavated sediment from the sites were dry sieved and then wet sieved through a 1.5 mm mesh screen, ensuring good recovery of small bones. The MK2 and Laili assemblages have been identified using the 450 Indo-Pacific specimens modern osteological reference collection housed at the Muséum national d'Histoire naturelle (UMR 7209), Paris (France). All recovered fish bone specimens were anatomically and taxonomically identified by C. Boulanger whenever it was possible.
Once identified, the different skeletal elements were counted to estimate taxon frequencies using common zooarchaeological quantifiers. The Number of Identified Specimens (NISP) was quantified by counting the number of remains identified to the Teleostei infraclass in the assemblage. Minimum Number of Elements (MNE) was calculated by counting the most frequent portion of each skeletal element. This allowed the calculation of the Minimum Number of Individuals (MNI) by counting the most frequent skeletal element, taking into account side. MNI is thus a secondary unit compared with the NISP, as it corresponds to the smallest number of individual animals necessary to account for all the NISP of a specific taxon in an assemblage (White, 1953). At MK2, MNI was calculated for Squares B and DD.
We have recorded several types of anthropic and natural modifications to bone specimens in these assemblages, before and after deposition; the traces of combustion linked to the action of fire and cooking of food, cut marks caused by tools during the preparation or consumption of food, and concretions due to the precipitation of dissolved carbonates from the limestone wall of the shelters. The fragmentation ratio was calculated in order to assess the relative degree of element destruction, by dividing the total Number of Identified Specimens (NISP) by the total Minimum Number of Elements (NISP/MNE) (“the more anatomically complete specimens, the less the difference between NISP and MNE”), values close to 1 indicating low fragmentation, and even higher values indicating even higher fragmentation and therefore smaller fragments (Lyman, 2008).
Results
Taxonomy
A greater proportion of the MK2 vertebrates was identifiable to taxa and to the Teleostei infraclass than those from Laili. The number of MK2 fish bones identified to order level numbered 89 (42.79%), followed by 88 (42.30%) to the family level (Table 1). This compared to the Laili assemblage NISP identified to order level of 56 (11.5%), while only a few have been identified to genus or species level (Table 1).
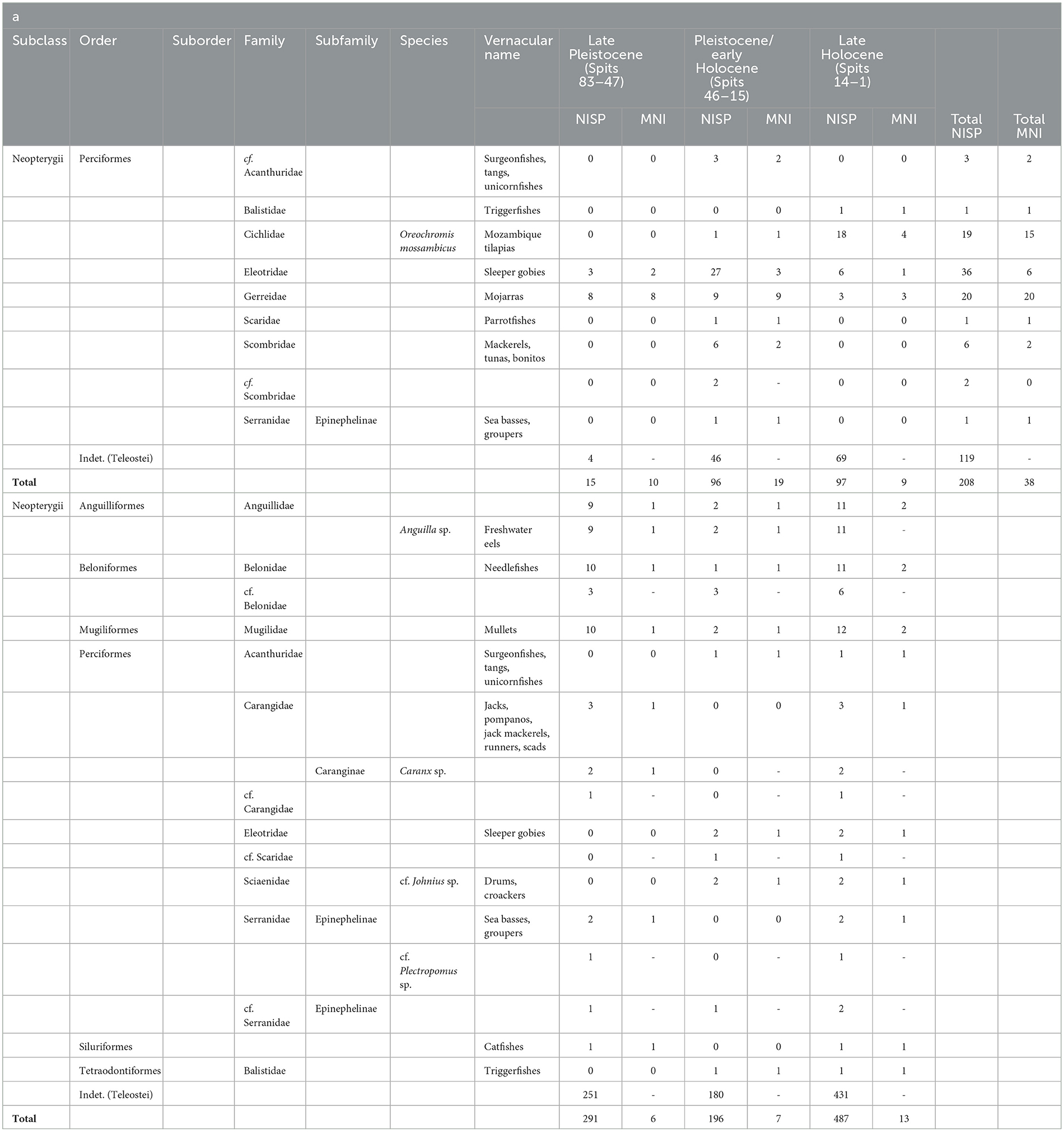
Table 1. Number of Identified Specimens (NISP) and Minimum Number of Individual (MNI) at (a) Matja Kuru 2 (Square B and Square DD combined) (b) Laili (Contexts 25B, 31B and 31C are pit cuts and have been added to the upper contexts; Context 43 is likely intrusive so has been removed from this table).
From MK2, at least eight different families and one suborder were identified belonging to the Perciformes and Tetraodontiformes orders. These include Eleotridae (sleeper gobies), Cichlidae (here, Mozambique tilapia), cf. Acanthuridae (surgeonfishes, tangs, unicornfishes), Scaridae (parrotfishes), Scombridae (mackerels, tunas, bonitos), Serranidae (sea basses, groupers), and Balistidae (triggerfishes). Gerreidae (mojarras) represented by a series of lower pharyngeal plates has also been identified (Figure 4; Tables 1, 2). At MK2 the dominant taxa are Gerreidae (mojarras) in the late Pleistocene phase, Eleotridae (sleeper gobies) in the Pleistocene/early Holocene phase, and Oreochromis mossambicus (Mozambique tilapia) in the late Holocene phase (Figure 4; Tables 1, 2).

Figure 4. Fish bones recovered at (A) Matja Kuru 2 and (B) Laili. The bones being damaged, each of the views shows the most preserved and diagnostic part of each specimen. 1. Caudal vertebra of Balistidae (external view) recovered from Square DD, Spit 9. 2. Lower pharyngeals (dorsal view) of Gerreidae recovered from Square DD, Spit 10 (top) and Square DD, Spit 26. 3. Vertebra of Anguilla sp. (external view) recovered from context CDE43. 4. Precaudal vertebra of Belonidae (ventral view) recovered from context D31 showing burning coloration. 5. Precaudal vertebra of Eleotridae (ventral view) recovered from Context E34 at Laili.
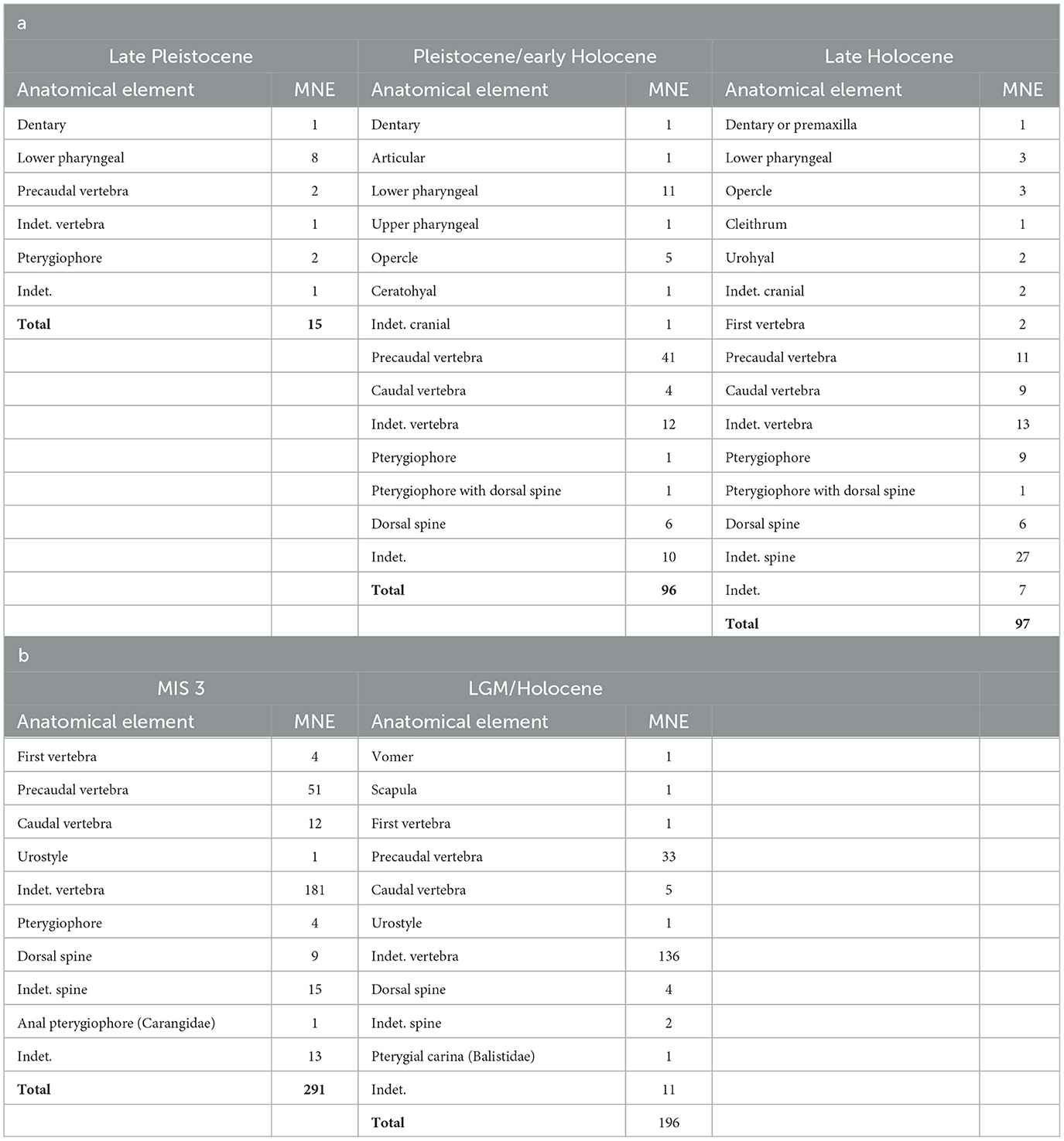
Table 2. Minimum Number of Elements (MNE) at (a) Matja Kuru 2, (b) Laili.
The Laili fish assemblage was far more diverse with at least ten different taxa identified, belonging to six different orders: the Anguilliformes, the Beloniformes, the Mugiliformes, the Siluriformes, the Tetraodontiformes, and the Perciformes, this last order represented by six families. The Anguillidae and the genus Anguilla (freshwater eels) are represented in the assemblage only by postcranial remains (Figure 4). Eleotridae (sleeper gobies) (Figure 4) and Sciaenidae (drums, croakers) have also been identified at the site. Siluriformes (catfishes) are represented only by one specimen (one spine) that could not be further identified. The remaining seven taxa are Belonidae (needlefishes) (Figure 4); Mugilidae (mullets); Acanthuridae (surgeonfishes, tangs, unicornfishes); Carangidae (jacks, pompanos, jack mackerels, runners, scads) including the genus Caranx; cf. Scaridae (parrotfishes); Serranidae including the genus Plectropomus (sea basses, groupers); and Balistidae (triggerfishes) (Table 1). The most abundant taxa at Laili Cave are the Anguillidae (freshwater eels), Mugilidae (mullets) and Belonidae (needlefishes) present mostly in the early Pleistocene phase (MIS 3).
Cultural and taphonomic disturbances
There was no evidence of cut marks indicating stone-tool butchery of the fish from either assemblage, which is unsurprising since fish were probably roasted on coals or steamed in ovens whole with little processing, as is still the case today, and even if knives were used, cut marks disappear over time after burial (Willis and Boehm, 2014). Possible indirect evidence for cooking was observed. A significant proportion of the fish bones from both sites appear to have suffered from combustion consistent with fire exposure (Figure 4; Table 3), although the burning of the bones is unlikely to have occurred during the cooking process, but rather after consumption when the bones were discarded. This was more pronounced for the Laili fish bones. At MK2 many of the bones in the upper deposits (Spits 20 to 1) appear “modern” or relatively recently deposited based on their color and condition, suggesting that they have been minimally impacted by burial processes (Table 3) (Samper Carro et al., 2023). This may be the result of deposition by an unidentifed avian predator; however, the site is still used by local villagers from the nearby settlement of Poros for ceremonies and during hunting trips and this may account for the fresh condition of the bones. Both assemblages appear to have been affected by post-depositional modifications. Most bones in each assemblage and occupation phase (with the exception of MK2 late Holocene phase) are partially fragmented (Table 3). However, while fragmentation was relatively extensive, the fragmentation intensity in these assemblages as measured by the fragmentation ratio is low (Table 3) and we were able to identify most bones to anatomical element. Both site assemblages and all chronological phases are dominated by small postcranial elements, predominantly vertebrae, which are more difficult to identify and often limit identification to family level only, compared to the denser and more distinctive cranial bones, although this is more pronounced at Laili (Tables 2, 3). Small quantities of the bones from the sites are covered in calcium carbonate concretions consistent with moisture exposure, further compounding identification (Table 3).
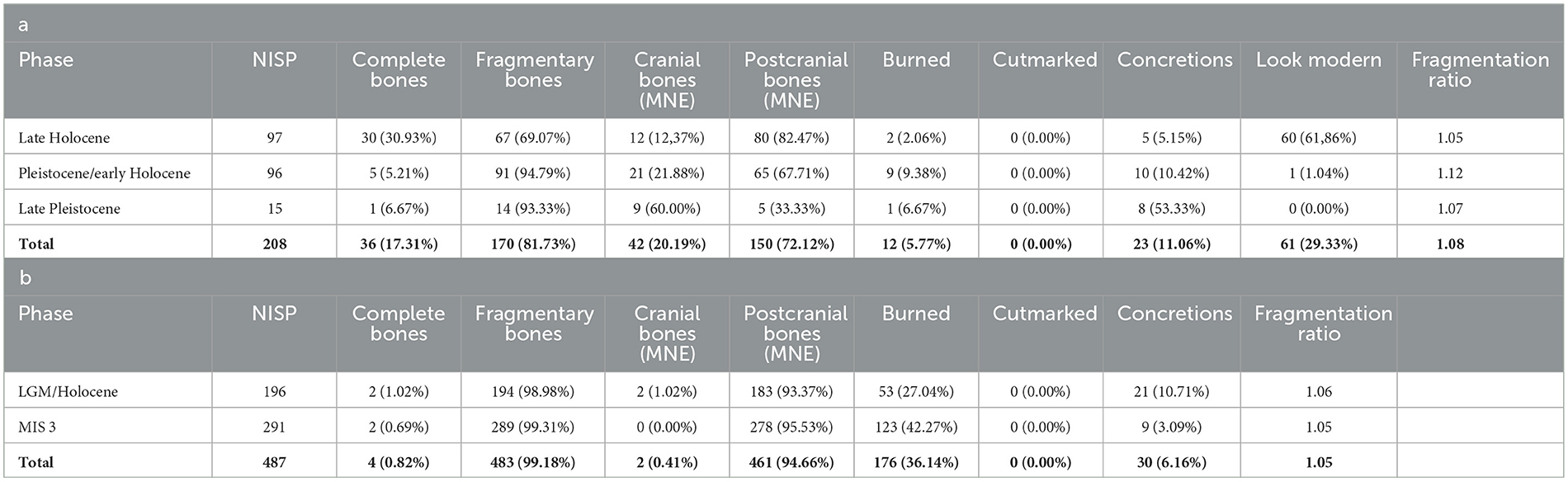
Table 3. Number and percentage of cultural and taphonomic disturbances on fish remains at (a) Matja Kuru 2 (Square B and Square DD combined), (b) Laili.
Discussion and conclusions
Near-shore reef fishing was practiced on the Timor coast both at MK2 and Laili, as shown by the presence of marine taxa such as Belonidae (needlefishes) (Figure 4), Acanthuridae (surgeonfishes, tangs, unicornfishes), Carangidae (jacks, pompanos, jack mackerels, runners, scads), Scaridae (parrotfishes), Scombridae (mackerels, tunas, bonitos), Serranidae (sea basses, groupers) and Balistidae (triggerfishes) (Figure 4; Table 1) presumably incorporating a range of methods that would have perhaps included spearing, netting, and angling to capture fish with different feeding behaviors and habitats (Butler, 1994), thus requiring a good knowledge of local marine environments and appropriate fishing techniques.
Freshwater catadromous and sometimes brackish water taxa, probably adults based on the average size of the bones, are represented at both sites. Gerreidae (mojarras) and Eleotridae (sleeper gobies) dominate the MK2 assemblage. Currently, three species belonging to the genus Gerres (G. filamentosus, G. oblongus and G. oyena) have been recorded on the Timorese coasts and rivers. However, because of the lack of reference material, we were unable to firmly identify the specimens to species or genus level. Most of mojarras are marine and occur in sandy or muddy bottoms bordered by mangroves. They occasionnaly enter rivers and lakes (Froese and Pauly, 2022). Eleotridae could be one of four species (Belobranchus belobranchus, Bunuka gyrinoides, Eleotris fusca and Giuris margaritacea) that have been reported from the south coast of Timor-Leste (De Beaufort and Weber, 1912; Larson et al., 2007). Eleotridae also occur in the Irasiquero River system near MK2, on which Lake Ira Lalaro is located (Figure 1) (Larson et al., 2007). The specimens in MK2 cannot be further identified because of the absence of those taxa in reference collection, added to the fact that we only found postcranial remains. The ecology of Eleotridae is a little more complicated as fish from this family can be found in a variety of environments with a preference for brackish and freshwater habitats, depending on the species and the lifestage of the fish (Bouffandeau et al., 2019; Froese and Pauly, 2022). MK2 and Laili are in close proximity to a freshwater lake and a river where these fish would be readily available and probably caught, as we think it is unlikely that the site's inhabitants would have walked to the distant coast to fish them, albeit we cannot exclude this possibility. A few modern bone fragments have been identified as Cichlidae in the upper more recent spits of MK2. These are consistent with the Mozambique tilapia, Oreochromis mossambicus (Cichlidae), which is endemic to Southeastern Africa. This species was introduced throughout ISEA and Timor-Leste in the 1940's to sustain food production for Timorese populations (Trewavas, 1983). Indeed, the introduction of carnivorous exotic African species such as Clarias gariepinus (Clariidae, north African catfish) and opportunistic omnivores such as O. mossambicus, perhaps modified the Timorese freshwater ecosystems causing endemic species demise (Larson et al., 2007).
Eels, the dominant freshwater taxa at Laili, are present throughout the sequence and four species have been currently recorded in Timor-Leste (Froese and Pauly, 2022). While acknowledging that some taxa in the family Anguillidae may be found in a variety of environments including marine (Froese and Pauly, 2022), we have characterized the Anguillidae in Laili as freshwater, as Anguilla sp. is the only genus that occurs in Timor (A. bicolor, A. celebesensis, A. marmorata, and A. reinhardtii) which only goes to the sea for spawning; this migration pattern remaining locally largely unknown (Arai, 2020). In addition, mullets (Mugilidae), represented by six genera and eight species (Cestraeus goldiei, Crenimugil buchanani, C. seheli, Ellochelon vaigiensis, Osteomugil cunnesius, Planiliza ordensis, P. subviridis, Plicomugil labiosus) today in Timor-Leste have been identified predominantly in the earliest phase of occupation of the site. Mullets can be found in a variety of habitats including coastal marine environments, to brackish and freshwater ecosystems (Froese and Pauly, 2022). Drums or croakers (Sciaenidae), probably of the genus Johnius (as it is the only genus today represented in Timor-Leste with two representatives, J. amblycephalus and J. borneensis) can also sometimes be found in brackish and estuarine environments (Froese and Pauly, 2022).
Today local villagers fish in Lake Ira Lalaro using long flexible bamboo rods equipped with metal hooks. Additionally, invasive species such as tilapia have been introduced to the inland waterways of Timor-Leste. These changes make it difficult to reconstruct traditional fishing techniques. The eels exploited at Laili were probably caught in the adjacent Laleia River using eel pots or nets (Froese and Pauly, 2022). De Almeida (1963) mentions that eels are usually found in springs and streams and, during his anthropological work, recorded that some of them were regarded as sacred by the locals. Sleepers, at MK2 and Laili, could have been fished using traps, weirs or nets (Bouffandeau et al., 2019) made of vegetal fibers in the Irasiquero River system (Larson et al., 2007) or in the Laleia River, while mojarras are usually caught using nets and sometimes line fishing in freshwater/brackish/marine environments (Carpenter and Niem, 1998). Drums or croakers were perhaps caught near the Laleia river mouth by angling or netting.
Different wetland habitats and distance from marine environments have therefore played some role in differences in ichthyofaunal assemblages between MK2 and Laili, with a higher diversity of fished environments at MK2 which is further from the coast and closer to Lake Ira Lalaro (Figures 1, 2). Previously, the most inland prehistoric site on Timor was found at the cave site Uai Bobo 2, located some 20 km from the coast when first occupied ca. 17 kya, but no fish bones were identified there (Kealy et al., 2016) (Figure 1). The new ichthyofaunal assemblages from the inland Timor-Leste sites, Laili and MK2, include small quantities of both marine, brackish and freshwater fish taxa. This indicates exploitation by H. sapiens of a variety of aquatic habitats and some degree of mobility between the coast and hinterland requiring the use of a variety of subsistence strategies culturally adapted to the different environments, as well as planning in terms of resources management. MK2 and Laili are not directly coastal; a return trip from the shelters to the coast on foot can today be made within a day. The Laili fish assemblage was much larger (NISP = 491, MNI = 14) than that of MK2 (NISP = 208, MNI = 38) (Table 1), despite the comparable size of the excavation areas at both sites. This discrepancy may be due to Laili's greater proximity to the coast and river. Although the MK2 fish assemblage is smaller than in Laili, the MNI is higher due to the larger number of Gerreidae pharyngeal plates identified (Figure 4).
Fishing persisted over time at MK2 and Laili, attesting to a detailed knowledge of the aquatic environments passed on over generations through social learning. Nonetheless, as might be expected, fishing at the inland Timor-Leste sites MK2 and Laili was not as intensive as at coastal sites throughout Wallacea such as Asitau Kuru (O'Connor et al., 2011), Makpan (Kealy et al., 2020), Tron Bon Lei (Samper Carro et al., 2016) and Here Sorot Entapa (O'Connor et al., 2019) (Figure 1). Inland fishing therefore, would have been a complementary component of terrestrial subsistence strategies that may have focused on gathered plant foods and even arboriculture until the spread of neolithic farming societies (O'Connor, 2015). This is a pattern reflected at inland sites elsewhere in Southeast Asia such as Niah cave on Borneo west of Wallace's line, where small amounts of freshwater fish complement freshwater shellfish gathering and hunting of medium to large forest mammals (Piper and Rabett, 2009). East of Wallace's line on the large Wallacean island of Sulawesi, medium to large endemic mammals were also hunted and freshwater shellfish gathered (Glover, 1981; Simons and Bulbeck, 2004). Freshwater fish and eels are rare in archaeological assemblages on the island. They have been identified at the inland shelter Gua Mo'o hono, but are confined to the late Holocene levels (O'Connor et al., 2018).
There are differences in fishing behavior over time between MK2 and Laili. Fishing activity was concentrated during different periods at each site. At Laili the majority of the fish remains were located in the lower part of the stratigraphy in the initial Pleistocene period of settlement ~44.6 kya (63.1%), although fishing continued into the terminal Pleistocene period with the same diversity of foraged environments (36.9%) after the LGM (Table 1). At MK2 most of the fishing was conducted during the Holocene period, possibly reflecting warmer wetter conditions and elevated freshwater levels in the lake, as well as more intensive human occupation in Timor-Leste after the LGM (Hawkins et al., 2017; Shipton et al., 2019, 2021). There were very few fish bones in the MK2 pleistocene deposits, with 92.79% of the fishing being identified in the early Holocene and late Holocene deposit from 11,000 years ago to the modern era, which could be a function of increased distance from the coastline between 40,000 and 31,000 BP (Table 1). These spatio-temporal differences in fishing practices could also be due to differences in regional culture as well as the effects of climate change on different habitats. Laili was perhaps occupied while procuring lithic resources from the river bed (Hawkins et al., 2017), while MK2 was likely occupied as a basecamp from which a variety of activities were undertaken during periods of high lake levels in Ira Lalaro (Veth et al., 2005; Langley and O'Connor, 2015, 2019; Langley et al., 2016), which probably requiring scheduling. Coral reef associated taxa at Laili indicate that reef platforms were present near the north coast of the site despite the steep coastline (Hawkins et al., 2017). MK2 shares similarities with Asitau Kuru, which are both situated near the eastern tip of Timor. Both sites had limited occupation during the LGM, likely due to sharp reduction in rainfall during the LGM which made settlement less favorable (O'Connor et al., 2011; Hawkins et al., 2017; Shipton et al., 2019; Roberts et al., 2020). MK2 and Asitau Kuru also had a shared material culture history that included shell beads and ochre pieces, also found at the neighboring sites of Matja Kuru 1 and Lene Hara respectively (Langley and O'Connor, 2015; Langley et al., 2016; O'Connor et al., 2019). Further, the presence of Scombridae (mackerels, tunas, bonitos) recorded at MK2 has resonances with Asitau Kuru, highlighting a possible connection between the two sites of a highly specialized fishing tradition targeting this fast-moving predator off the edge of the reef.
These findings demonstrate that with access to more adequate modern reference specimens it is possible to identify the different habitats of archaeological fish specimens by comparative morphological analyses and thus reveal a greater range of H. sapiens adaptations to the tropical islands of Wallacea. Our results allow us to hypothetize that H. sapiens in Timor-Leste were able to utilize a flexible and sophisticated set of fishing strategies across a diverse range of inland and coastal aquatic habitats from at least 45 kya. Such foraging adaptations, requiring the scheduling of subsistence activities between the coast and hinterland wetland habitats, would have required a high degree of planning as well as diverse capture technology. This mobility is likely a precursor to the development of trade and exchange networks by the terminal Pleistocene in the Wallacea (Reepmeyer et al., 2011, 2019).
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
CB: conceptualization, formal analysis, investigation, methodology, writing—original draft, and writing—review and editing. SH: conceptualization, writing—original draft, and writing—review and editing. CS: investigation, project administration, supervision, and writing—review and editing. TI and A-MS: conceptualization and writing—review and editing. SSC: formal analysis and writing—review and editing. SO'C: funding acquisition, investigation, project administration, supervision, and writing—original draft. All authors contributed to the article and approved the submitted version.
Funding
This project was undertaken with the assistance of the Ministerio da Educacao, Cultura, Juventude e Desporto de Timor-Leste and the villagers of Laleia. This work was supported by the Australian Research Council (CE170100015 and FL120100156), the Australian National University and the ARC Laureate Project FL120100156.
Acknowledgments
We thank the UMR7209 laboratory (Muséum national d'Histoire naturelle) and Philippe Béarez for giving us access to the fish osteological reference collection they patiently built year after year. We acknowledge the CartoGIS Services (ANU College of Asia and the Pacific, The Australian National University) for providing all base maps (Creative Commons Attribution-ShareAlike 4.0 international license (CC BY SA).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Álvarez-Fernández, E., Bécares, J., Jordá Pardo, J.F., Agirre-Uribesalgo, A., and Álvarez-Alonso, D. (2020). Palaeoenvironmental and chronological context of human occupations at El Cierro cave (Northern Spain) during the transition from the late Upper Pleistocene to the early Holocene. J. Archaeol. Sci. Rep. 29, 102138. doi: 10.1016/j.jasrep.2019.102138
Amano, N., Moigne, A. M., Ingicco, T., Sémah, F., Awe, R. D., Simanjuntak, T., et al. (2016). Subsistence strategies and environment in Late Pleistocene–Early Holocene Eastern Java: Evidence from Braholo Cave. Quat. Int. 416, 46–63. doi: 10.1016/j.quaint.2015.09.072
Arai, T. (2020). Ecology and evolution of migration in the freshwater eels of the genus Anguilla schrank. J. Heliyon. 6, e05176. doi: 10.1016/j.heliyon.2020.e05176
Archer, W., Braun, D. R., Harris, J. W. K., McCoy, J. T., and Richmond, B. G. (2014). Early Pleistocene aquatic resource use in the Turkana Basin. J. Hum. Evol. 77, 74–87. doi: 10.1016/j.jhevol.2014.02.012
Barker, G., Barton, H., Bird, M., Daly, P., Datan, I., and Dykes, A. (2007). The ‘human revolution' in lowland tropical Southeast Asia: the antiquity and behavior of anatomically modern humans at Niah Cave (Sarawak, Borneo). J. Hum. E52, 243–261. doi: 10.1016/j.jhevol.2006.08.011
Boschian, G. (1993). Continental deposits and archaeological data in the Trieste Karst area (north-east Italy: evidence of sea-level changes and possible tectonic activity in the Late Pleistocene and Early Holocene. Geol. Rundsch. 82, 828. doi: 10.1007/BF00191828
Bouffandeau, L., Béarez, P., Keith, P., Bedford, S., and Spriggs, M. (2019). Freshwater fishing among Lapita people: the sleepers (Teleostei: Eleotridae) of Teouma, Vanuatu. J. Archaeol. Sci. Rep. 26, 101894. doi: 10.1016/j.jasrep.2019.101894
Boulanger, C. (2021). Aquatic resources exploitation and adaptation of Anatomically Modern Human in Island Southeast Asia: Palaeoenvironmental and Cultural Implications (PhD dissertation). Muséum national d'Histoire naturelle, Australian National University, Paris, Canberra.
Boulanger, C., Hawkins, S., Ono, R., and O'Connor, S. (2023). Continuity and variability in prehistoric fishing practices by Homo sapiens in Island Southeast Asia: new ichthyofaunal data from Asitau Kuru, Timor-Leste. World Archaeol. 54, 288–316. doi: 10.1080/00438243.2023.2192518
Boulanger, C., Ingicco, T., Piper, P. J., Amano, N., Grouard, S., Ono, R., et al. (2019). Coastal subsistence strategies and mangrove swamp evolution at Bubog I rockshelter (Ilin Island, Mindoro, Philippines) from the Late Pleistocene to the mid-Holocene. J. Isl. Coast. Archaeol. 8, 1–21. doi: 10.1080/15564894.2018.1531957
Bowler, J. M., Johnston, H., Olley, J. M., Prescott, J. R., Roberts, R. G., Shawcross, W., et al. (2003). New ages for human occupation and climatic change at Lake Mungo, Australia. Nature 421, 837. doi: 10.1038/nature01383
Brooks, A. S., Helgren, D. M., Cramer, J. S., Franklin, A., Hornyak, W., Keating, J. M., et al. (1995). Dating and context of three middle stone age sites with bone points in the upper Semliki Valley, Zaire. Science 268, 548–553. doi: 10.1126/science.7725099
Brumm, A., Hakim, B., Ramli, M., Aubert, M., and Bergh, G. D. (2018). A reassessment of the early archaeological record at Leang Burung 2, a Late Pleistocene rock-shelter site on the Indonesian island of Sulawesi. PLoS ONE 13, e0193025. doi: 10.1371/journal.pone.0193025
Butler, V. L. (1994). Fish feeding behaviour and fish capture: the case for variation in lapita fishing strategies. Archaeol. Ocean. 29, 81–90. doi: 10.1002/arco.1994.29.2.81
Butler, V. L. (2001). Changing Fish Use on Mangaia, Southern Cook Islands: Resource Depression and the Prey Choice Model. Int. J. Osteoarchaeol. 88–100. doi: 10.1002/oa.548
Carpenter, K. E., and Niem, V. H. (1998). The Living Marine Resources of the Western Central Pacific. Rome: FAO.
Clarkson, C., Jacobs, Z., Marwick, B., Fullagar, R., Wallis, L., and Smith, M. (2017). Human occupation of northern Australia by 65000 years ago. Nature 547, 306. doi: 10.1038/nature22968
De Beaufort, D., and Weber, L. F. M. (1912). On the freshwater fishes of Timor and Babber in KNAW, Proceedings, 235–240.
Feathers, J. K., and Migliorini, E. (2001). Luminescence dating at Katanda — a reassessment. Quat. Sci. Rev. 20, 961–966. doi: 10.1016/S0277-3791(00)00041-X
Froese, R., and Pauly, D. (2022). FishBase [WWW Document]. Available online at: http://www.fishbase.org (accessed November 25, 2022).
Glover, I. (1981). Leang Burung 2 : an upper Palaeolithic rock shelter in South Sulawesi, Indonesia. Mod. Quat. Res. Southeast Asia 6, 1–38.
Gobalet, K. W., and Fenenga, J. L. (1993). Terminal Pleistocene-early holocene fishes from Tulare Lake, San Joaquin Valley, California, with comments on the evolution of sacramento squawfish (Ptychocheilus grandis: Cyprinidae). PaleoBios 15, 1–8.
Göltenboth, F., Timotius, K. H., Milan, P. P., and Margraf, J. (2006). Ecology of Insular Southeast Asia: The Indonesian Archipelago. Amsterdam: Elsevier.
Gonzalez, A., Clark, G., O'Connor, S., and Matisoo-Smith, L. (2013). A 3000 year old dog burial in Timor-Leste. Aust. Archaeol. 76, 13–20. doi: 10.1080/03122417.2013.11681961
Gorman, C. (1971). The Hoabinhian and after: subsistence patterns in Southeast Asia during the late pleistocene and early recent periods. World Archaeol. 2, 300–320. doi: 10.1080/00438243.1971.9979482
Groucutt, H. S., Petraglia, M. D., Bailey, G., Scerri, E. M. L., Parton, A., and Clark-Balzan, L. (2015). Rethinking the dispersal of Homo sapiens out of Africa. Evol. Anthropol. 24, 149–164. doi: 10.1002/evan.21455
Guillaud, E., Béarez, P., Daujeard, C., Defleur, A. R., Desclaux, E., Roselló-Izquierdo, E., et al. (2021). Neanderthal foraging in freshwater ecosystems: a reappraisal of the Middle Paleolithic archaeological fish record from continental Western Europe. Quat. Sci. Rev. 252, 106731. doi: 10.1016/j.quascirev.2020.106731
Guillaud, E., Lebreton, L., and Béarez, P. (2018). Taphonomic signature of Eurasian eagle owl (Bubo bubo) on fish remains. Folia Zool. 67, 143–153. doi: 10.25225/fozo.v67.i3-4.a1.2018
Habgood, P. J., and Franklin, N. R. (2008). The revolution that didn't arrive: a review of Pleistocene Sahul. J. Hum. E55, 187–222. doi: 10.1016/j.jhevol.2007.11.006
Hawkins, S., O'Connor, S., Maloney, T. R., Litster, M., Kealy, S., Fenner, J. N., et al. (2017). Oldest human occupation of Wallacea at Laili Cave, Timor-Leste, shows broad-spectrum foraging responses to late Pleistocene environments. Quat. Sci. Rev. 171, 58–72. doi: 10.1016/j.quascirev.2017.07.008
Hongo, H. (1989). Freshwater fishing in the Early Jomon period (Japan): An analysis of fish remains from the Torihama shell-mound. J. Archaeol. Sci. 16, 333–354. doi: 10.1016/0305-4403(89)90010-1
Ingicco, T., Amano, N., Setiagama, K., and Moigne, A. M. (2020). From Food to Grave Good: Nonhuman Primate Exploitation in Early to Mid-Holocene Eastern Java (Indonesia). Curr. Anthropol. 61, 264–277. doi: 10.1086/708186
Joordens, J. C. A., d'Errico, F., Wesselingh, F. P., Munro, S., de Vos, J., and Wallinga, J. (2015). Homo erectus at Trinil on Java used shells for tool production and engraving. Nature 518, 228–231. doi: 10.1038/nature13962
Joordens, J. C. A., Kuipers, R. S., Wanink, J. H., and Muskiet, F. A. J. (2014). A fish is not a fish: patterns in fatty acid composition of aquatic food may have had implications for hominin evolution. J. Hum. Evol. The role of freshwater and marine resources in the evolution of the human diet. Brain Behav. 77, 107–116. doi: 10.1016/j.jhevol.2014.04.004
Kealy, S., Louys, J., and O'Connor, S. (2016). Islands under the sea: a review of early modern human dispersal routes and migration hypotheses through Wallacea. J. Isl. Coast. Archaeol. 11, 364–384. doi: 10.1080/15564894.2015.1119218
Kealy, S., O'Connor, S., Mahirta, Sari, D.M., Shipton, C., and Langley, M.C. (2020). Forty-thousand years of maritime subsistence near a changing shoreline on Alor Island (Indonesia). Quat. Sci. Rev. 249, 106599. doi: 10.1016/j.quascirev.2020.106599
Kettle, A. J., Heinrich, D., Barrett, J. H., Benecke, N., and Locker, A. (2008). Past distributions of the European freshwater eel from archaeological and palaeontological evidence. Quat. Sci. Rev. 27, 1309–1334. doi: 10.1016/j.quascirev.2008.03.005
Kirch, P. V., and Dye, T. S. (1979). Ethno-archaeology and the development of Polynesian fishing strategies. J. Polyn. Soc. 88, 53–76.
Lambeck, K., and Chappell, J. (2001). Sea level change through the last glacial cycle. Science 292, 679–686. doi: 10.1126/science.1059549
Langley, M. C., Clarkson, C., and Ulm, S. (2011). From small holes to grand narratives: the impact of taphonomy and sample size on the modernity debate in Australia and New Guinea. J. Hum. E61, 197–208. doi: 10.1016/j.jhevol.2011.03.002
Langley, M. C., Kealy, S., and Mahirta O'Connor, S. (in press). Sequins from the Sea: The Nautilus Shell Bead Technology of Makpan, Alor Island, Indonesia. Antiquity.
Langley, M. C., and O'Connor, S. (2015). 6500-Year-old Nassarius shell appliqués in Timor-Leste: technological and use wear analyses. J. Archaeol. Sci. 62, 175–192. doi: 10.1016/j.jas.2015.06.012
Langley, M. C., and O'Connor, S. (2016). An Enduring Shell Artefact Tradition from Timor-Leste: Oliva Bead Production from the Pleistocene to Late Holocene at Jerimalai, Lene Hara, and Matja Kuru 1 and 2. PLoS ONE 11, e0161071. doi: 10.1371/journal.pone.0161071
Langley, M. C., and O'Connor, S. (2019). 40, 000 years of ochre utilization in Timor-Leste: powders, prehensile traces, and body painting. PaleoAnthropol. Early Pers. Ornaments 27, 82−104. doi: 10.4207/PA.2019.ART125
Langley, M. C., O'Connor, S., and Piotto, E. (2016). 42, 000.-year-old worked and pigment-stained Nautilus shell from Jerimalai (Timor-Leste): evidence for an early coastal adaptation in ISEA. J. Hum. Evol. 97, 1–16. doi: 10.1016/j.jhevol.2016.04.005
Larson, H. K., Buckle, D., Lynas, J., Storey, A., and Humphrey, C. (2007). Additional records of freshwater fishes from Timor-Leste, with notes on the fish fauna of the unique land-locked Irasiquero River system. Beagle Rec. Mus. Art Galleries North. Territ. 23, 131–135. doi: 10.5962/p.320166
Long, K., Wood, R., Williams, I. S., Kalish, J., Shawcross, W., Stern, N., et al. (2018). Fish otolith microchemistry: Snapshots of lake conditions during early human occupation of Lake Mungo, Australia. Quat. Int. Adv. Geoarchaeol. Res. 463, 29–43. doi: 10.1016/j.quaint.2016.10.026
Losey, R. J., Nomokonova, T., and White, D. (2012). Fish and Fishing in Holocene Cis-Baikal, Siberia: A Review. J. Isl. Coast. Archaeol. 7, 126–145. doi: 10.1080/15564894.2011.611854
Marean, C. W. (2014). The origins and significance of coastal resource use in Africa and Western Eurasia. J. Hum. Evol. 77, 17–40. doi: 10.1016/j.jhevol.2014.02.025
Masse, W. B. (1986). A Millennium of Fishing in the Palau Islands, Micronesia, Pacific Anthropological Records. Presented at the Traditional Fishing in the Pacific: Ethnographical and Archaeological Papers from the 15th Pacific Science Congress. Honolulu: Bishop Museum Press, 85–119.
Mellars, P., Gori, K. C., Carr, M., Soares, P. A., and Richards, M. B. (2013). Genetic and archaeological perspectives on the initial modern human colonization of Southern Asia. Proc. Natl. Acad. Sci. 110, 10699–10704. doi: 10.1073/pnas.1306043110
Mitchell, P. (2015). “Discourse on Rivers, and Fish and Fishing': Freshwater Aquatic Resources and Hunter-gatherers in Southern African Prehistory,” in The Oxford Handbook of the Archaeology of Diet, eds Lee-Thorp, J., Katzenberg, M.A. Oxford: Oxford University Press.
Nakajima, T., Hudson, M. J., Uchiyama, J., Makibayashi, K., and Zhang, J. (2019). Common carp aquaculture in Neolithic China dates back 8, 000. years. Nat. Ecol. Evol. 1–7. doi: 10.1038/s41559-019-0974-3
O'Connor, S. (2010). Continuity in shell artefact production in Holocene East Timor, in 50 Years of Archaeology in Southeast Asia: Essays in Honour of Ian Glover, eds B. Bellina, E. A. Bacus, T. O. Pryce (Bangkok: River Books), 218–233.
O'Connor, S. (2015). Rethinking the Neolithic in Island Southeast Asia, with Particular Reference to the Archaeology of Timor-Leste and Sulawesi. Archipel. Études interdisciplinaires sur le monde insulindien. 45, 15–47. doi: 10.4000/archipel.362
O'Connor, S., and Aplin, K. (2007). A Matter of Balance: An overview of Pleistocene occupation history and the impact of the Last Glacial Phase in East Timor and the Aru Islands, eastern Indonesia. Archaeol. Ocean. 42, 82–90. doi: 10.1002/j.1834-4453.2007.tb00021.x
O'Connor, S., Bulbeck, D., Piper, P., Aziz, F., Marwick, B., Campos, F., et al. (2018). The human occupation record of Gua Mo'o hono shelter, Towuti-Routa region of Southeastern Sulawesi. Archaeol. Res. Pleistocene 11, 117–151. doi: 10.22459/TA48.11.2018.09
O'Connor, S., Ono, R., and Clarkson, C. (2011). Pelagic fishing at 42,000 years before the present and the maritime skills of modern humans. Science 334, 1117–1121. doi: 10.1126/science.1207703
O'Connor, S., Robertson, G., and Aplin, K. P. (2014). Are osseous artefacts a window to perishable material culture? Implications of an unusually complex bone tool from the Late Pleistocene of East Timor. J. Hum. E67, 108–119. doi: 10.1016/j.jhevol.2013.12.002
O'Connor, S., Shimona kaely, M., Boulanger, C., Maloney, T. R., Hawkins, S., and Langley, M. C. (2019). Kisar and the archaeology of small islands in the Wallacean archipelago. J. Isl. Coast. Archaeol. 14, 198–225. doi: 10.1080/15564894.2018.1443171
Ono, R., and Addison, D. J. (2009). Ethnoecology and Tokelauan fishing lore from Atafu Atoll, Tokelau. SPC Tradit. Mar. Resour. Manag. Knowl. Inf. Bull. 26, 3–22.
Ono, R., Fuentes, R., Pawlik, A., and Sofian, H. O. (2020). Island migration and foraging behaviour by anatomically modern humans during the late Pleistocene to Holocene in Wallacea: new evidence from Central Sulawesi, Indonesia. Quat. Int. 27, 1040618220301610. doi: 10.1016/j.quaint.2020.03.054
Ono, R., Hawkins, S., and Bedford, S. (2019). “Lapita maritime adaptations and the development of fishing technology: A view from Vanuatu,” in Debating Lapita: Distribution, Chronology, Society and Subsistence, Terra Australis, eds S. Bedford and M. Spriggs (Canberra: ANU Press), 415–438.
Pargeter, J., Loftus, E., and Mitchell, P. (2017). New ages from Sehonghong rock shelter: implications for the late Pleistocene occupation of highland Lesotho. J. Archaeol. Sci. Rep. 12, 307–315. doi: 10.1016/j.jasrep.2017.01.027
Piper, P. J., and Rabett, R. J. (2009). Hunting in a tropical rainforest: evidence from the Terminal Pleistocene at Lobang Hangus, Niah Caves, Sarawak. Int. J. Osteoarchaeol. 19, 551–565. doi: 10.1002/oa.1046
Plug, I. (2006). Aquatic animals and their associates from the Middle Stone Age levels at Sibudu. South. Afr. Humanit. 18, 289–299.
Reepmeyer, C., O'Connor, S., and Brockwell, S. (2011). Long-term obsidian use at the Jerimalai rock shelter in East Timor. Archaeol. Ocean. 46, 85–90. doi: 10.1002/j.1834-4453.2011.tb00102.x
Reepmeyer, C., O'Connor, S., Mahirta, S., and Maloney, T. (2019). Kisar, a small island participant in an extensive maritime obsidian network in the Wallacean Archipelago. Archaeol. Res. Asia 19, 100–139. doi: 10.1016/j.ara.2019.100139
Roberts, P., Douka, K., Tromp, M., Bedford, S., Hawkins, S., Bouffandeau, L., et al. (2022). Fossils, fish and tropical forests: prehistoric human adaptations on the island frontiers of Oceania. Philos. Trans. R. Soc. B Biol. Sci. 377, 20200495. doi: 10.1098/rstb.2020.0495
Roberts, P., Louys, J., Zech, J., Shipton, C., Kealy, S., Carro, S. S., et al. (2020). Isotopic evidence for initial coastal colonization and subsequent diversification in the human occupation of Wallacea. Nat. Commun. 11, 1–11. doi: 10.1038/s41467-020-15969-4
Roberts, P., and Stewart, B. A. (2018). Defining the ‘generalist specialist' niche for Pleistocene Homo sapiens. Nat. Hum. Behav. 2, 542. doi: 10.1038/s41562-018-0394-4
Russ, H. (2010). The Eurasian eagle owl (Bubo bubo): a fish bone accumulator on Pleistocene cave sites? J. Taphon. 8, 281–290.
Samper Carro, S. C. (2023). Hominin adaptations in the Lesser Sunda Islands: exploring the vertebrate record to investigate fauna diversity before, during and after the Last Glacial Maximum. World Archaeol. 3, 1–24. doi: 10.1080/00438243.2023.2172073
Samper Carro, S. C., O'Connor, S., Louys, J., Hawkins, S., and Mahirta, M. (2016). Human maritime subsistence strategies in the Lesser Sunda Islands during the terminal Pleistocene–early Holocene: New evidence from Alor, Indonesia. Quat. Int. 416, 64–79. doi: 10.1016/j.quaint.2015.07.068
Samper Carro, S. C., Raymond, C., Weisbecker, V., and Sue O'Connor. (2023) Big questions for small animals. Taphonomic analysis of small vertebrates in Matja Kuru 2 (Timor-Leste) during the Late Pleistocene. Quaternary Sci. Adv.
Shipton, C., O'Connor, S., Jankowski, N., O'Connor-Veth, J., Maloney, T., Kealy, S., et al. (2019). A new 44,000-year sequence from Asitau Kuru (Jerimalai), Timor-Leste, indicates long-term continuity in human behaviour. Archaeol. Anthropol. Sci. 11, 5717–5741. doi: 10.1007/s12520-019-00840-5
Shipton, C., O'Connor, S., and Kealy, S. (2021). The biogeographic threshold of Wallacea in human evolution. Quat. Int. 574, 1–12. doi: 10.1016/j.quaint.2020.07.028
Simons, A., and Bulbeck, D. (2004). “Late Quaternary faunal successions in South Sulawesi, Indonesia,” in Modern Quaternary Research in Southeast Asia, eds S. G. Keates, and J. M. Pasveer (London: CRC Press). 167–190.
Stewart, K. M. (1994). Early hominid utilisation of fish resources and implications for seasonality and behaviour. J. Hum. E27, 229–245. doi: 10.1006/jhev.1994.1044
Trewavas, E. (1983). Tilapiine fishes of the genera Sarotherodon, Oreochromis, and Danakilia. London: British Museum.
van Niekerk, K. L. (2011). Marine fish exploitation during the middle and later Stone Age of South Africa (PhD dissertation). Cape Town: University of Cape Town.
Veth, P., Spriggs, M., and O'Connor, S. (2005). Continuity in Tropical Cave Use: Examples from East Timor and the Aru Islands, Maluku. Asian Perspect. 44, 180–192. doi: 10.1353/asi.2005.0015
Weber, A., Peter, J., and Hirofumi, K. (2013). Environmental change and cultural dynamics of Holocene hunter-gatherers in Northeast Asia : Comparative analyses and research potentials in Cis-Baikal (Siberia, Russia) and Hokkaido (Japan). Quat. Int. 290, 3–20. doi: 10.1016/j.quaint.2012.07.021
Westaway, K. E., Louys, J., Awe, R. D., Morwood, M. J., Price, G. J., Zhao, J., et al. (2017). An early modern human presence in Sumatra 73,000–63,000 years ago. Nature 548, 322–325. doi: 10.1038/nature23452
White, T. E. (1953). A method of calculating the dietary percentage of various food animals utilized by aboriginal peoples. Am. Antiq. 18, 396–398. doi: 10.2307/277116
Willis, L. M., and Boehm, A. R. (2014). Fish bones, cut marks, and burial: implications for taphonomy and faunal analysis. J. Archaeol. Sci. 45, 20–25. doi: 10.1016/j.jas.2014.01.026
Wood, R. (2017). Comments on the chronology of Madjedbebe. Aust. Archaeol. 83, 172–174. doi: 10.1080/03122417.2017.1408545
Zakaria-Ismail, M. (1994). Zoogeography and biodiversity of the freshwater fishes of Southeast Asia, in Ecology and Conservation of Southeast Asian Marine and Freshwater Environments Including Wetlands, Developments in Hydrobiology, eds A. Sasekumar, N. Marshall, D. J. Macintosh (Dordrecht: Springer), 41–48.
Zilhão, J., Angelucci, D. E., Igreja, M. A., Arnold, L. J., Badal, E., Callapez, P., et al. (2020). Last Interglacial Iberian Neandertals as fisher-hunter-gatherers. Science 367, aaz7943. doi: 10.1126/science.aaz7943
Keywords: ichthyoarchaeology, palaeoecology, Paleolithic, Timor-Leste, island environments, fish bones
Citation: Boulanger C, Hawkins S, Shipton C, Ingicco T, Sémah A-M, Samper Carro S and O'Connor S (2023) Inland fishing by Homo sapiens during early settlement of Wallacea. Front. Environ. Archaeol. 2:1201351. doi: 10.3389/fearc.2023.1201351
Received: 06 April 2023; Accepted: 15 June 2023;
Published: 06 July 2023.
Edited by:
Christina Giovas, Simon Fraser University, CanadaReviewed by:
Marshall Weisler, The University of Queensland, AustraliaCharles Higham, University of Otago, New Zealand
Reno Nims, Portland State University, United States
Copyright © 2023 Boulanger, Hawkins, Shipton, Ingicco, Sémah, Samper Carro and O'Connor. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Clara Boulanger, Y2xhcmEuYm91bGFuZ2VyQG1uaG4uZnI=