 Mariana Nabais
Mariana Nabais Catherine Dupont
Catherine Dupont João Zilhão
João Zilhão- 1IPHES-CERCA - Institut Català de Paleoecologia Humana i Evolució Social, Zona Educacional 4, Campus Sescelades URV (Edifici W3), Tarragona, Spain
- 2Department d'Història i Història de l'Art, Universitat Rovira i Virgili, Tarragona, Spain
- 3UNIARQ - Centro de Arqueologia da Universidade de Lisboa, Faculdade de Letras, Universidade de Lisboa, Lisbon, Portugal
- 4CNRS, CReAAH, University of Rennes, Rennes, France
Hominin consumption of small prey has been much discussed over the past decades. Such resources are often considered to be unproductive in the Middle Paleolithic due to their limited meat yield and, hence, low energy return. However, ethnographic studies suggest that small prey—including shellfish—are a reliable, predictable and by no means marginal resource, and there is increasing evidence for their inclusion in hominin diets during the Middle Paleolithic and even earlier. Gruta da Figueira Brava features a MIS 5c-5b Neanderthal occupation that left behind substantial, human-accumulated terrestrial and marine faunal remains, capped by reworked levels that contain some naturally accumulated, recent Holocene material, namely the remains of small crab species and echinoderms. The brown crab Cancer pagurus (Linnaeus, 1758) predominates in the intact Middle Paleolithic deposit, and reconstruction of its carapace width, based on regression from claw size, shows a preference for relatively large individuals. The detailed analysis of the Cancer pagurus remains reveals that complete animals were brought to the site, where they were roasted on coals and then cracked open to access the flesh.
1. Introduction
There is growing evidence for the Neanderthals' subsistence exploitation of small terrestrial animals. This has been demonstrated by several studies like Morin et al. (2019) for leporids, Blasco et al. (2016) for birds, Nabais and Zilhão (2019) for tortoises, to name just a few examples from southern France and the Iberian Peninsula. This evidence accords with the argument that Neanderthals preferentially consumed terrestrial animals (Naito et al., 2016; Jaouen et al., 2019), consistent with the scarcity of marine species in Middle Paleolithic faunal assemblages. Yet, we now know that fish and shellfish harvesting played a significant role in the subsistence economy of Last Interglacial Iberian Neanderthals (Zilhão et al., 2020).
Shellfish and other small size marine resources have been frequently considered to be a rather unproductive source of food in the Middle Paleolithic. According to Optimal Foraging Theory (OFT), such resources rank low because of their limited raw meat weight and energy return (e.g., Winterhalder and Smith, 2000; Dusseldorp, 2010). However, the Nutritional Ecology approach outlined by Hockett and Haws (2003, 2004, 2005) ranks marine resources much higher because of their protein and vitamin intake, fat content, reliability, and low-risk collection, and a number of studies (e.g., Hardy and Moncel, 2011; Langejans et al., 2012) have been able to demonstrate their dietary significance. This change of perspective has boosted the study of the archaeology of molluscs, while crustaceans and echinoderms still get relatively little attention.
An important contribution to the zooarchaeology of crabs is a study of the Lower Paleolithic occupation at Gesher Benot Ya'aqov, Israel (Ashkenazi et al., 2005). There is also brief mention of crab remains in faunal assemblages from the Spanish Upper Paleolithic levels of Altamira (Álvarez Fernández, 2010), and Fuente del Salín (Gutiérrez Zugasti et al., 2013). Crustacean and echinoderm remains become increasingly visible in the Late Pleistocene and Early Holocene, as apparent at several sites from northern Spain (e.g., Gutiérrez Zugasti, 2009, 2011; Gutiérrez Zugasti et al., 2016; García-Escárzaga et al., 2017; Dupont and Gruet, 2022). In Portugal, crustaceans are briefly mentioned in the Mesolithic sites of Toledo (Dupont, 2011), Fiais (Lubell et al., 2007), Cabeço da Amoreira (Pinto, 1986; Dupont and Bicho, 2015), Moita do Sebastião (Roche, 1958), Montes de Baixo (Silva and Soares, 1997), Poças de São Bento (Arnaud, 2000), Quinta da Sardinha (Ribeiro, 1884), Samouqueira I (Lubell et al., 2007), Cabeço da Arruda (Lentacker, 1986), Arapouco (Arnaud, 2000), Cabeço do Rebolador (Arnaud, 2000), Vale Frade (Araújo et al., 2014), Barranco das Quebradas 1, Barranco das Quebradas 4 and Rocha das Gaivotas (Carvalho, 2001; Valente, 2008; Dupont, 2011). However, it is only at the French Mesolithic sites of Beg-er-Vil (Gruet, 2002; Dupont and Gruet, 2005), Beg-an-Dorchenn (Dupont and Gruet, 2005; Dupont et al., 2010), Hoëdic and Téviec (Dupont et al., 2007), and the Neolithic site of Ponthezières (Gruet and Laport, 1996; Gruet, 2009) that in-depth studies of the prehistoric remains of crustaceans have been carried out (Dupont and Gruet, 2022). Otherwise, the archaeological remains of crustaceans and echinoderms are mostly underreported; more often than not, their presence is simply referred to in species lists.
Echinoderms and crustaceans are easily found in coastal areas. Echinoderms (sea urchins), e.g., Paracentrotus lividus [Lamarck, 1816; the scientific names of marine species used in this paper follow the WoRMS Editorial Board (2023) nomenclature], are distributed from Scotland to southern Morocco, including the Mediterranean Sea. P. lividus inhabits several substrates from rocks and boulders to sea meadows, living from the low-water limit down to −20 m. It is found at low densities in meadows of Posidonia oceanica (Linnaeus) (Delile, 1813), an endemic Mediterranean species, but it is more frequent in shallow, hard substrate algal communities, like the red alga Lithophyllum incrustans Philippi, 1837. It is commonly found in Portugal, in intertidal rock pools and shallow subtidal reefs. P. lividus thrives under highly variable environmental conditions (e.g., temperature and wave action) and can switch from grazing-feeding behaviors to drift-feeding when in its self-built burrow (Saldanha, 1995; Boudouresque and Verlaque, 2001; Tomas et al., 2004; Bayed et al., 2005; Jacinto et al., 2013). P. lividus is a “regular echinoid” featuring the typical spherical shape flattened at the top (with the anus at its center) and at the base (where the mouth is located). Its surface is made of test plates that are covered by needle-like spines. The edible part is the roe, which is only large enough for a few weeks in the year, thus making P. lividus a good seasonality indicator. It is easily harvested by hand at low tide, and is often found in large groups (Hayward and Ryland, 1995; Campbell, 2008; Martin, 2012). According to Campbell (2008), the most frequent processing technique is to break out the base and the mouth structures, and eat the roe.
Nowadays, crustaceans—barnacles from the Maxillopoda class, and crabs, shrimp, and lobsters from the Malacostraca class—are some of the most appreciated seafood. Barnacles are found attached to hard substrates in all marine habitats, and on all zones of the shore. They are divided into two orders: the Sessilia (acorn barnacles) that grow their shell directly onto the substrate, and the Pedunculata (goose barnacles and others) that attach themselves by means of a stalk (Lohse and Raimondi, 2007). In the studied assemblage, only the acorn barnacle Perforatus perforatus (Bruguière, 1789) was found. It has a volcano-like shape composed of six test plates that attach to the substrate at the base, whereas the top forms an opening (the operculum) that is covered by two pairs of movable plates, the terga and the scuta (Southward, 2008). P. perforatus occurs naturally from SW Wales to West Africa, and in the Mediterranean Sea, in semi-exposed and shady rock surfaces, crevices, cave entrances or encrusted onto shells and the skin of marine mammals. The breeding season of this species begins earlier and lasts longer in the Portuguese Atlantic coast essentially due to favorable water temperatures and food availability provided by the Canary/Iberian upwelling (Cunha et al., 2018). P. perforatus is one of the larger barnacles of European coasts and, although not eaten, can be used as fishing bait (Claassen, 2013).
Crabs are easily identifiable through their outer shell, or exoskeleton. The exoskeleton needs to shed several times in a lifetime to accomodate the animal's growth. This process (known as molting) happens in hiding, since the animals get very soft and defenseless. It is also frequently associated with reproduction, e.g., in the Cancer genus: molting female crabs attach themselves to hard-shelled males with copulation occurring in the process (see a summary in Oesterling, 2012). The other notable feature of crabs is their pincers, which function in defense and feeding. They yield substantial amounts of meat and are the most taxonomically diagnostic elements. In all crab species, two types of pincers exist: crusher (shorter and stouter; generally on the right), and cutter (longer and thinner; generally on the left) claws. Each pincer has two fingers—a dactylopodus (flexible finger) and a propodus (unmovable finger) (Crothers and Crothers, 1988; Ingle, 1996; Gruet, 2002)—from which carapace body size estimations can be calculated based on measurements described below. On rocky seashores, crabs will shelter under stones or in nearly any hole, cave or crevice. Therefore, most seawater crabs can be found in the intertidal zone, and even subtidal species (like Maja squinado Herbst, 1788) come to shore for reproduction (Ingle, 1996; Gruet, 2002).
Here, we report on the first known Middle Paleolithic marine crustacean assemblage, adding to the growing body of evidence mentioned above. We present a detailed study of the crab component of the Neanderthal diet as revealed by the evidence from the Portuguese cave site of Gruta da Figueira Brava (38° 28′ 14″ N, 08° 59′ 10″ W) (Figure 1A).

Figure 1. Gruta da Figueira Brava. (A) Site location. (B) Plan of the cavities (elevations are in m above sea level). (C) The cave's three entrances seen from the sea. (D) Stratigraphic profile of Area F; units IL2 and IL3 contain occupation remains from Phase FB3, units IH2-IH3 to IH8 contain occupation remains from Phase FB4; IT0 denotes the reworked deposit. [(B) and (D) are reproduced from Zilhão et al. (2020), published under a CC BY license, with permission from The American Association for the Advancement Studies, original copyright 2020].
2. Gruta da Figueira Brava
The cave is located about 30 km south of Lisbon, on the southern slope of the Serra da Arrábida, just 5 meters above modern sea level. The area features thermo-mediterranean climatic conditions and a vegetation cover dominated by evergreen trees and shrubs, as well as several Mediterranean perenial herbs (e.g., thyme, lavender) (Ribeiro, 1945).
Currently, Gruta da Figueira Brava features three main entrances (Figure 1B). The interior can only be accessed via Entrance 1, which connects to the areas behind the other two, speleothem-cluttered, entrances. In Area C, the archaeological deposit is sealed by a stalagmitic crust and was excavated in the 1980s. An extremely narrow passage connects this area with a small chamber, Area F (Figure 1C). The archaeological deposit in this chamber, also protected by a stalagmitic crust, was excavated in 2010–2013 (Figure 1D). In Entrance 3, exteriorly of Area F, the cave remains partially protected by an overhang. Here, the Last Interglacial cobble beach underlies brecciated Upper Pleistocene deposits rich in charcoal, bone, shells and stone tools. When the cave was in use, the now-unroofed Gruta da Figueira Brava marine abrasion platform formed the bedrock floor of a large cave whose original morphology was lost due to littoral erosion caused by Late Glacial and Holocene sea level rise (Figures 1B, C; Zilhão, 2012; Zilhão et al., 2020).
U-series and OSL dating place the accumulation of the archaeological deposit, which spans four human occupation phases, in the ca. 86–106 ka interval (Zilhão et al., 2020). Except for two crab remains from Phase FB3 and one barnacle remain from Phase FB2, the crustacean material discussed here comes from the most recent phase, FB4, which dates to MIS 5b. Climatic conditions were very similar to present, as indicated by the plant cover reconstructed from the paleobotanical evidence. However, the outline of the coast was significantly different due to the lowered sea level (Badal et al., 2011; Zilhão et al., 2020).
Table 1 provides a list and counts for the mammals, birds and reptiles recovered in the 1980s excavations. Due to the non-separation of deposits affected by burrowing, this assemblage contains an undetermined quantity of Holocene intrusions. In the case of the larger size vertebrate taxa, it was possible to identify such intrusions based on bone surface appearance (i.e., degree of fossilization, patina, concretion coating) (Cardoso, 1993; Antunes, 2000). However, this approach is inapplicable in the case of the remains of marine invertebrates (molluscs, crustaceans and echinoderms), which the 1980s researchers analyzed in bulk. They provided no specimen counts, only a list of taxa, which includes Paracentrotus lividus, three species of barnacles (Chthamalus stellatus Poli, 1791; Chthamalus montagui Southward, 1976; and Perforatus perforatus Bruguière, 1789), and three species of crabs (Cancer pagurus Linnaeus, 1758; Maja squinado Herbst, 1788; and Portumnus sp. Leach, 1814) (Callapez, 2000).
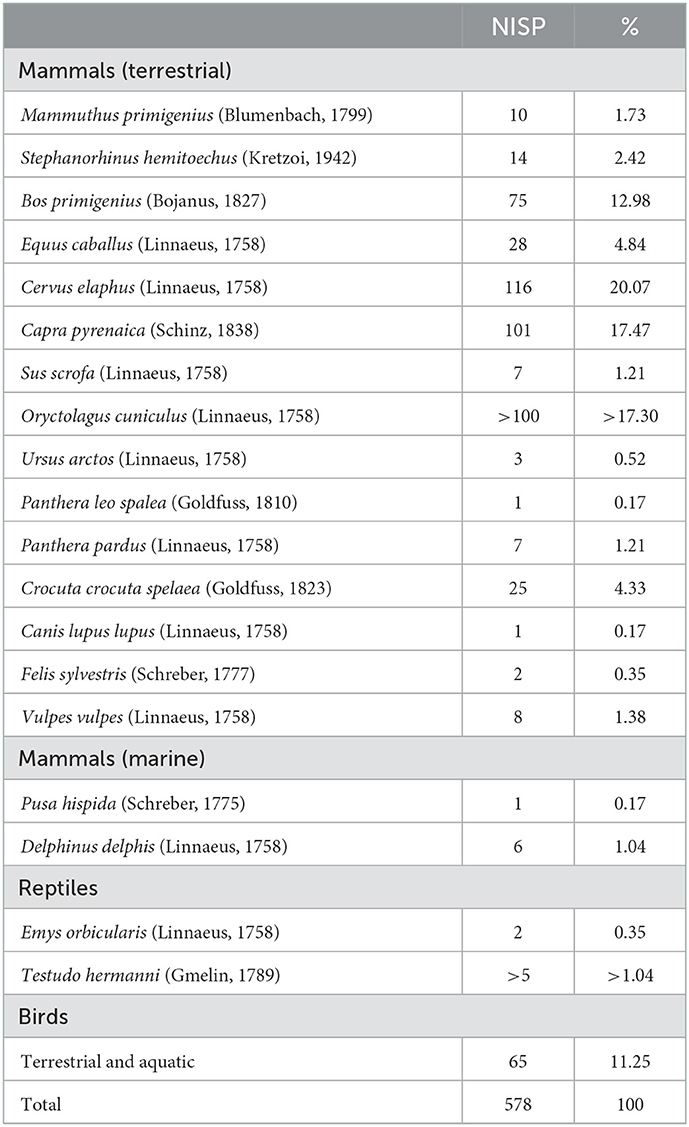
Table 1. Faunal list from the 1980s excavation of Area C of Gruta da Figueira Brava (after Cardoso, 1993, p. 531 and Antunes, 2000, p. 259).
In the 2010–2013 excavations, the reworked parts of the deposit were processed separately. The faunal analysis from the Last Interglacial levels reveals a mammal species composition like that described for the 1980s assemblage, with minor differences, namely the absence of the larger taxa (e.g., rhinoceros, elephant, marine mammals), and a narrower range of carnivores. Similarly, the 2010–2013 stone tool assemblage compares well with that from the 1980s excavation. Both are typically Mousterian assemblages dominated by quartz and in which the limited number of imported flints document use of the Levallois method (Zilhão et al., 2020).
3. Materials and methods
The faunal assemblage studied here was archaeologically excavated between 2010 and 2013. Most remains were hand-collected in the excavation trench, and otherwise retrieved from dry-screening with 3 mm meshes, followed by wet-screening or floatation of the residue in the field laboratory. All fragments identified to species during excavation were triangulated on site (X, Y, Z) and given a unique identifier number. The remainder were gathered in appropriately spit/quadrant-specific general finds bags.
The study of the crustacean and echinoderm remains was carried out in the Archaeomalacofauna Laboratory of the Center de Recherche en Archéologie, Archéosciences, Histoire (CReAAH) of the University of Rennes 1 (Rennes, France). Identifications were based on the reference collection and aided by handbooks, such as, Southward (2008) for barnacles, Southward and Campbell (2006) and Mougne (2015) for echinoderms, Crothers and Crothers (1988), Ingle (1996), and Gruet (2002) for crabs (Figure 2).
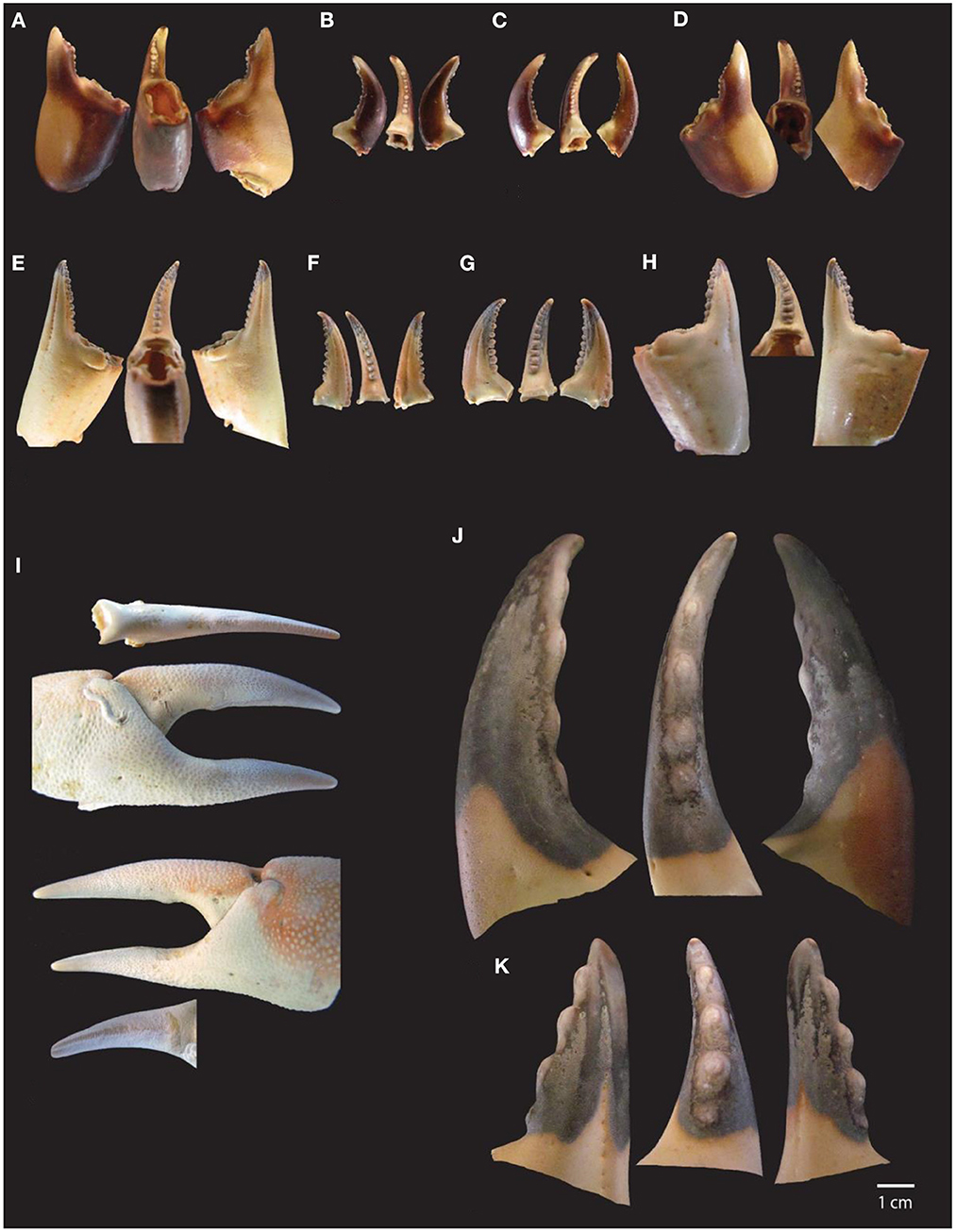
Figure 2. Distinctions between crab species based on claw morphology. In some species, left and right claws show some differences, but in other species they are similar. (A–D) Pachygrapsus marmoratus left propodus (A), left dactylopodus (B), right dactylopodus (C) and right propodus (D). (E–H) Carcinus maenas left propodus (E), left dactylopodus (F), right dactylopodus (G) and right propodus (H). (I) Maja squinado right claw right dactylopodus, right claw on anterior side, right claw on dorsal side and right propodus. (J, K) Cancer pagurus right dactylopodus (J) and right propodus (K).
Every fragment was studied in detail. The portion of the skeletal element present was recorded along with the anatomical part identified and, whenever possible, sided (left, right, indeterminate). Echinoderms were quantified based on their inner skeletal structure, the Aristotle's lantern, whereas for barnacles the test plates were used for such calculations. The Minimum Number of Individuals (MNI) for crabs was based on the highest figure of left or right propodus (unmovable finger) or dactylopodus (flexible finger) for each stratigraphic unit. The Minimum Animal Units (MAU) were computed, following Binford (1978), by dividing the number of propodus or dactylopodus by the number of times they occur in the body of the animal. After quantification by stratigraphic unit, both the MNI and the MAU, were agglomerated according to the site's occupation phasing (FB4, FB3, and FB2). The finds from the reworked deposit were considered, as their naturally accumulated Holocene components provided a standard against which to assess the taphonomy of the in situ Pleistocene assemblages: mode of accumulation, and agents (natural, carnivore/raptor or human) thereof.
As mentioned above, claw size and carapace width are correlated, and so crab pincers can be used for body size estimations (Figure 3). Based on a group of 50 crabs from the reference collection, with both sexes and several sizes represented (from small to large individuals), an allometric equation predicting carapace width from the length of dactylopodus and propodus fragments was determined (r = 0.98). Carapace width estimations were only calculated for Carcinus maenas (Dupont, 2017) and Cancer pagurus (Dupont et al., in press).
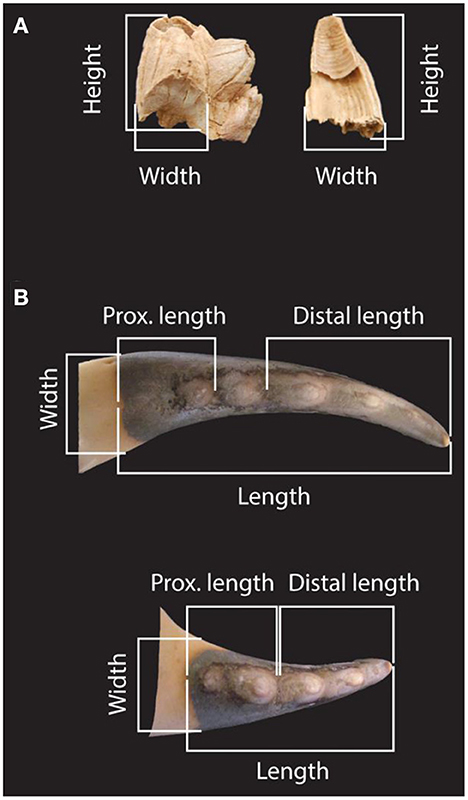
Figure 3. Crustacean biometry. (A) From left to right: Measurements taken on complete barnacles, and complete plate. (B) From top to bottom: Measurements taken on crab dactylopodus (flexible finger) and propodus (unmovable finger); note that proximal and distal lengths should be measured from the interior side of claws, where the protuberances are located, whereas width and length should be measured from the exterior side, where the claw protuberances are absent.
In order to quickly assess fragmentation, every remain was assigned to a size category in centimeters (e.g., 0–1 cm, 1–2 cm). Shell breakage was recorded according to Villa and Mahieu (1991): fracture outline was recorded as transverse, curved/V-shaped or longitudinal; fracture angle as oblique, right or mixed; surface edge as jagged or smooth; and the time of fracture was described as old or new based on appearance, in order to assess the damage caused by excavation.
Due to the lack of a referential corpus for the interpretation of archaeological crab taphonomy, we relied upon evidence derived from the study of mammals and molluscs (an experimental study on Cancer pagurus has recently been finalized1). Shell surface modification was assessed based on the presence/absence of percussion marks (such as pits, notches, impact scars, and adhering flakes) and butchery marks (like cuts, scrapes, and chops) (Binford, 1981; White, 1992; Capaldo and Blumenschine, 1994; Fisher, 1995; Díez et al., 1999; Pickering and Egeland, 2006; Blasco and Fernández Peris, 2012a,b). Additional information is given on the number of striations, and their distribution (isolated, clustered, crossed), orientation (transverse, longitudinal, oblique), delineation (straight or curved), location, and side of the skeletal element they are found on (posterior, anterior, medial, lateral). Burning was also recorded based on the experimental work conducted on mammals (Shipman et al., 1984; Nicholson, 1993) and molluscs (Villagran, 2014; Milano et al., 2016). Thermo-alterations were assessed based on the color scheme defined by such experiments (not burnt, brown, black, gray and white), which correlates with different temperatures of heat exposure. Unburnt crab claws can present a natural dark color on the external surface but not on the interior one, which is naturally very light colored, whereas manganese stains are patchy and tend not to show on claw sections: in crabs, therefore, burning was identified by surface observation of shell fragments' sections.
Carnivore mark classification used the categories defined by Binford (1981), Andrews (1990), and Fisher (1995): punctures, pits, scores, crenulation, and digestion. The number of marks, their location on the anatomical element and their distribution (isolated, clustered, crossed) were recorded. The maximum length of carnivore punctures was recorded in millimeters. Other modifications—abrasion, dissolution, incrustation, sediment filling, and concretion coating—were also observed and their presence or absence was recorded.
4. Results
4.1. Taxonomic and body part frequencies
The sea urchin sample comprises five remains: one hemipyramid, and four test plates (Table 2). Species identification is based on one test plate that is large enough to allow the diagnostic count of pore-pairs, indicating the presence of Paracentrotus lividus (Lamarck, 1816). All specimens come from the reworked parts of the deposit that contain Holocene intrusions.
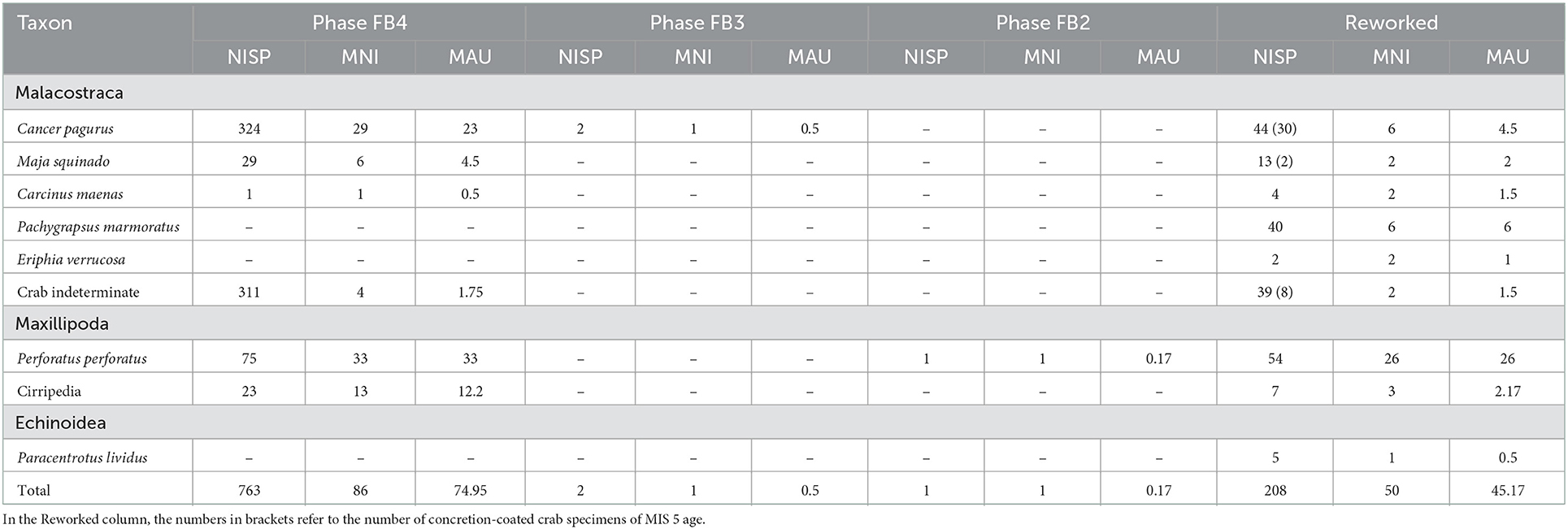
Table 2. Number of identified specimens (NISP), Minimum number of individuals (MNI), and minimum animal units (MAU) per taxon and occupation phase of Gruta da Figueira Brava.
Based on test remains (Figure 4A; no terga or scuta plates were recovered) 81.25% (NISP = 130) of the barnacle assemblage could be identified as Perforatus perforatus (Bruguière, 1789). About a quarter of the remains (38.13% or NISP = 61) come from the reworked deposit, but the great majority was found in the MIS 5 deposit; most are from Phase FB4, and a single remain is from Phase FB2 (Table 2).

Figure 4. Crustaceans from Gruta da Figueira Brava. (A) Perforatus perforatus recovered from reworked sediment; note their good preservation and absence of calcareous concretion. (B) Cancer pagurus carapace and pincer fragments with black burns; recovered from the MIS-5 deposit. (C) Cancer pagurus dactylopoda showing impact flakes on their ventral proximal side, reflecting pincer disarticulation; recovered from the MIS-5 deposit. (D) Cancer pagurus pincers with longitudinal breaks; recovered from the MIS-5 deposit [Images (B) and (D) are reproduced from Zilhão et al. (2020), published under a CC BY license, with permission from The American Association for the Advancement Studies, original copyright 2020].
With regards to crabs, a total of 809 remains were recovered and examined; more than half (or 56.7%) were identified to species (Tables 2, 3). The taxonomic identification was based on pincers (propodus and dactylopodus) and mandibles, allowing the identification of five different species. The assemblage from the MIS 5 deposit is overwhelmingly dominated by Cancer pagurus (Linnaeus, 1758; the brown crab), which is also represented in the reworked deposit, where most remains of this taxon feature the diagnostic concretion coating of the in situ material and therefore must correspond to Pleistocene-age finds. Conversely, Pachygrapsus marmoratus (Fabricius, 1787; the marbled rock crab) is absent from the in situ Pleistocene assemblage, despite being the dominant taxon amongst the non-concretion-coated crabs of the reworked deposit. Maja squinado (Herbst, 1788; the spider crab) occurs more frequently in the MIS 5 deposit than in the reworked deposit. A few non-concretion-coated remains of Eriphia verrucosa (Forsskål, 1775; the yellow crab) and Carcinus maenas (Linnaeus, 1758; the green crab) were found in the reworked deposit and, like P. marmoratus, must correspond to Holocene intrusions.
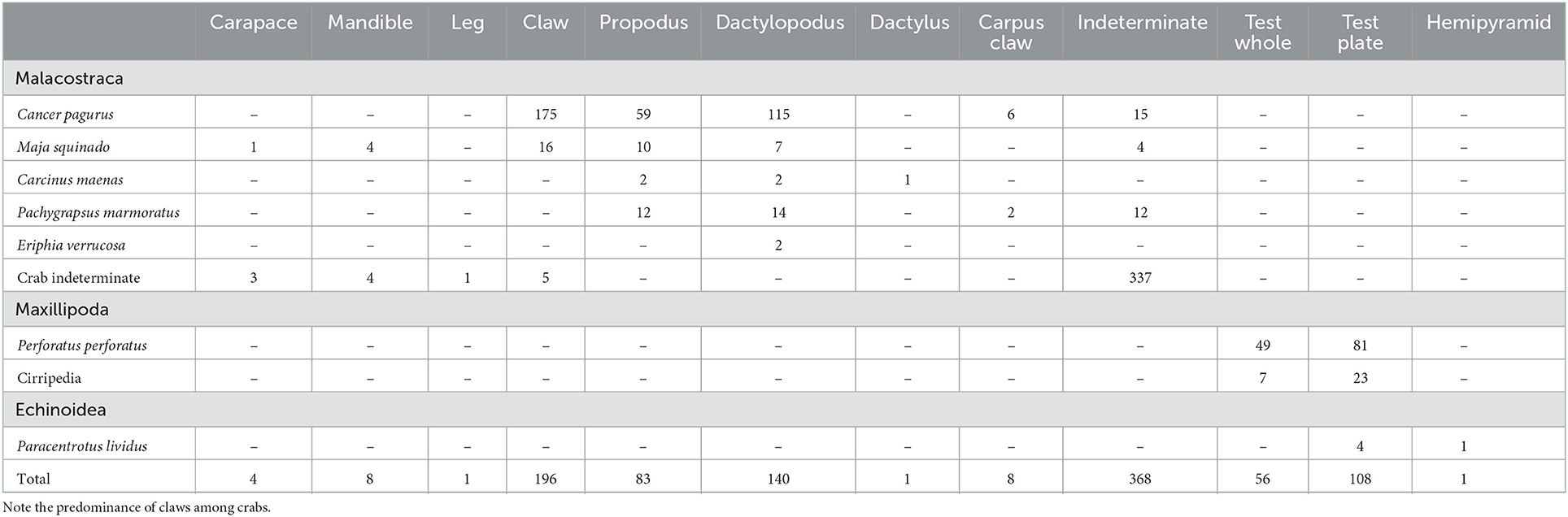
Table 3. Body part representation per taxon among the Crustaceans from Gruta da Figueira Brava.
4.2. Fragmentation and type of fracture
The remains from the reworked deposit tend to show less fragmentation when compared to those found in the in situ MIS 5 deposit. This is clear for barnacles, which, in the reworked deposit, are complete in 75.40% (NISP = 46) of the cases, as opposed to only 53.54% (NISP = 53) in the in situ MIS 5 deposit; their robust calcite tests explain the high degree of completeness seen in both cases. The sea urchin remains all show recent fractures.
Overall, crab remains are mostly fractured (90.98%; NISP = 736). However, it is notable that 67.5% (NISP = 27) of the P. marmoratus assemblage corresponds to complete or nearly complete claws, confirming good shell preservation in a species that is naturally fragile and prone to breakage. The opposite happens to the robust shell of Cancer pagurus, for which only 7.03% (or NISP = 26) of the claw remains are nearly complete (Tables 3, 4). The use of power tools to excavate the heavily brecciated parts of the deposit does not seem to have had much impact on the crab remains, since most show no excavation-induced breakage; most recent fractures were observed in the reworked deposit (NISP = 83; cf. NISP = 5 in the MIS 5 deposit). Finally, when broken, most crab claws identified to species present transversal fractures, except for Cancer pagurus, among which longitudinal fractures predominate and complete pincers are absent (Table 4). Longitudinal fractures are regularly found on the outer edge of the pincers of Cancer pagurus, i.e., on the edge opposite to the claw's protuberances (Figure 4D). However, whenever the propodus and the dactylopodus are identified based on the presence of their distal ends, fractures tend to be transverse. Both longitudinal and transverse fractures on Cancer pagurus claws are clean and smooth edged with a right angle.
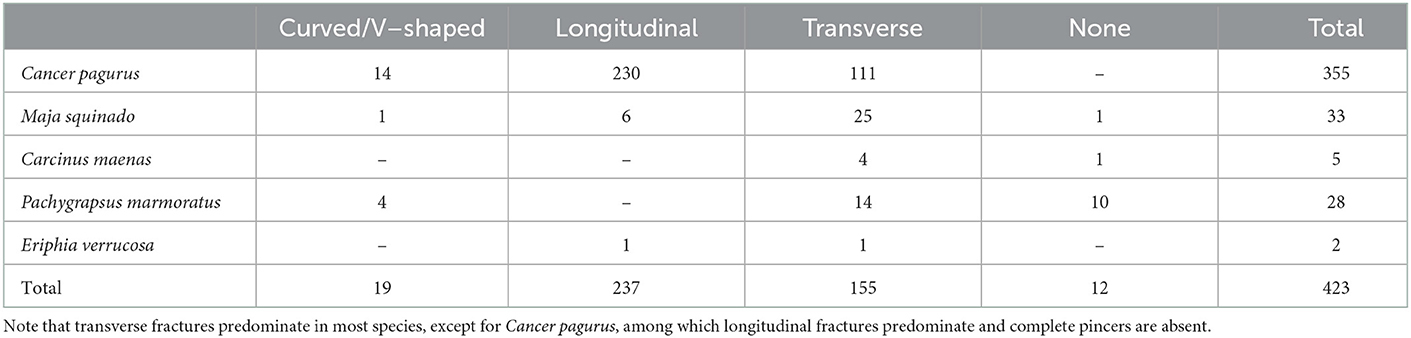
Table 4. Type of fracture on all crab claws identifiable to species from Gruta da Figueira Brava.
4.3. Burning
Thermo-alterations were identified on two barnacle plate fragments, but were mostly found among the crab remains; none were seen in the sea urchin assemblage. In the crab assemblage, 8.41% (NISP = 68) of the remains are burnt (Table 5); black burns are the most frequent (Figure 4B), burning colors reflecting very high temperatures (gray and white) are nearly absent and, except for six carapace elements, it is the claws that are burnt, and almost exclusively Cancer pagurus ones. No burning was observed on remains of the taxa that only occur in the reworked deposit containing Holocene intrusions.
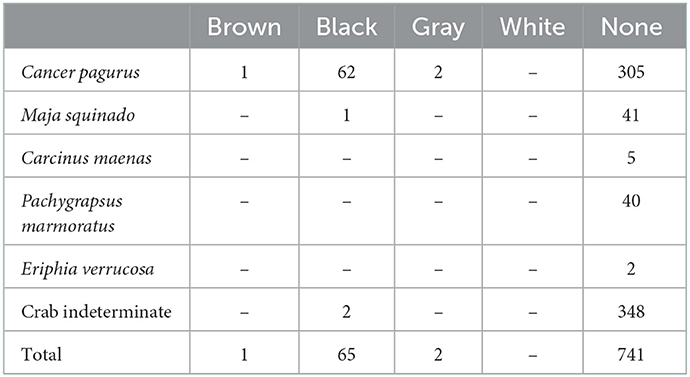
Table 5. Burning on the crab remains recovered from Gruta da Figueira Brava.
4.4. Surface modification
No surface modifications were observed on sea urchin remains. Half of the barnacle assemblage (51.3% or NISP = 82) shows calcareous coatings, but only two remains feature sediment fillings and another two have dissolution marks on their dorsal side. Abrasion and incrustation are absent. There is evidence of one perforation, but due to its size and shape, it is probably due to gastropod predation.
Crab remains do not feature incrustation or abrasion marks, the latter implying the lack of exposure to strong tide activity. Focusing on the MIS 5 remains, all show coating with calcareous concretions, 18.2% (or NISP = 122) have sediment fillings, and dissolution was detected on ten specimens. No carnivore or rodent gnawing marks were detected, and anthropogenic incisions are also absent. A total of ten impact scars were found on the ventral side of the proximal end of Cancer pagurus dactylopoda (Figure 4C).
4.5. Body size
Carapace width could be measured in two Carcinus maenas remains from the reworked deposit, revealing a population ranging in size between 53 and 73 mm. The in situ deposit is overwhelmingly dominated by Cancer pagurus, whose carapace width was estimated from the length of the pincers to range between 111 and 223 mm, averaging 162 mm (Figure 5).
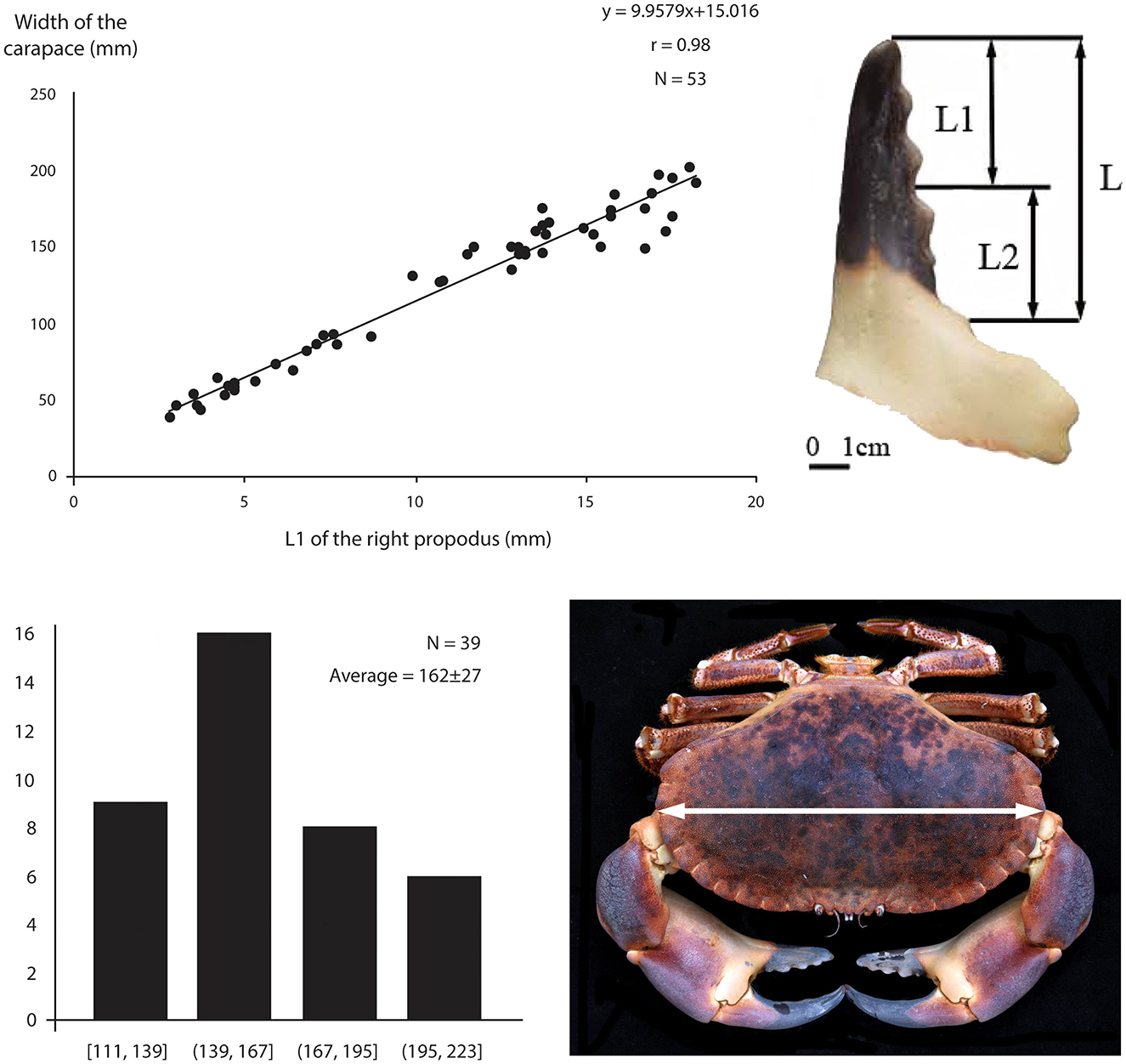
Figure 5. (Top left) Cancer pagurus carapace width (mm); the regression line reveals the biometric relationship between the size of the right propodus and the size of the carapace. (Bottom left) Distribution of carapace width estimates for Gruta da Figueira Brava's MIS 5 brown crabs. (Top right) Propodus measurements. (Bottom right) Width measurement.
5. Discussion
5.1. Agent of accumulation
Considering the scarce literature available for crustacean and echinoderm archaeological remains, most of the assumptions underpinning our identification of the agents of accumulation involved derive from criteria used in studies of other shellfish and small animal remains.
Natural accumulations are expected to show individuals in anatomical connection or in close association, with a relatively good state of preservation and low fragmentation (Bishop, 1986). As summarized by Erlandson and Moss (2001), several carnivores—e.g., canids, felids, bears, and otters—are opportunistic shellfish predators, and there are also several non-human primates that eat crabs (Russon et al., 2014; Koops et al., 2019). These animals can transport crabs to archaeological sites, and their participation in the accumulation can be inferred from the presence of their own skeletons and the identification of digestion, gnawing, and bite marks. A variety of birds can also be responsible for the accumulation of crab remains, e.g., cormorants, condors, sea eagles, corvids, and gulls. Such accumulations are often related with nesting and show high levels of fragmentation with random types of fracture, as those birds use gravity (dropping of the shellfish from high above onto rocks below) to break the shell open and expose the edible flesh (Erlandson and Moss, 2001).
An anthropogenic accumulation is expected to be found in stratigraphic association with stone tools and other features, such as hearths; to show surface modifications (like burning), patterns of mechanical fracture and, potentially, some cuts and percussion marks. Based on such criteria, Neanderthal mollusc accumulations have been identified in Gibraltar caves (Finlayson, 2008; Stringer et al., 2008), as well as in Bajondillo Cave, Spain (Córtez-Sánchez et al., 2011). Similar criteria have been used for the South African Middle Stone Age shellfish accumulations recovered from Sea Harvest (Volman, 1978), Hoedjiespunt (Will et al., 2013; Kyriacou et al., 2015), Klasies River (Thackeray, 1988; Langejans et al., 2012), Ysterfontein 1 (Klein et al., 2004), Pinnacle Point 13B (Marean et al., 2007; Jerardino and Marean, 2010) and Blombos Cave (Langejans et al., 2012). Moreover, an anthropogenic accumulation is expected to be size-selective, thus showing preference for larger animals with higher meat content.
At Gruta da Figueira Brava, two different agents seem to have been involved in the accumulation of the crab remains. The reworked deposit shows high species diversity and is whence all specimens of Pachygrapsus marmoratus and Eriphia verrucosa come; most Carcinus maenas remains were also recovered therein. Even though the remains of these species are fragile and prone to breakage, preservation tends to be good and there is a high degree of completeness; there are several complete pincers, and most fractures are recent, ruling out accumulation by carnivores, which would typically feature a high degree of fragmentation. The same applies to the fragile test fragments of sea urchins. We can therefore conclude that these remains were not significantly exposed to damage, either from the dynamics of sedimentary accumulation or from the impact of marine dynamics (e.g., shell abrasion, or incrustation by barnacles or parasites). Likely, the remains represent recent Holocene natural deaths, whether in the cave itself or in the adjacent rocky beach, that eventually made their way into the sedimentary fill via natural agency, namely burrowing by small mammals. Indeed, nowadays Pachygrapsus marmoratus is common in the intertidal pools around Gruta da Figueira Brava. As is also the case with Carcinus maenas and Eriphia verrucosa, these crabs move along the creeks and crevices of the rocks, and they inhabit seashore caves whose wet environment provides welcoming shelter (Ingle, 1996). Moreover, no carnivore marks were identified in any crustacean or urchin remains, and overall carnivore presence in the cave is scant (Zilhão et al., 2020).
With regards to the in situ deposit, which is dominated by Cancer pagurus, the crab remains bear no evidence of carnivore or raptor modification. Moreover, brown crabs were selected for large carapace width (on average, 160 mm), which reflects a preference for adult males weighing around 800 g (Woll, 2006; Haigh et al., 2015). Apart from humans, the other agents capable of transporting such large crabs over the distances involved—during Phase FB4, where 99% of Cancer pagurus and Maja squinado remains belong, the site-to-shore distance was ~2 km (Table 2; Zilhão et al., 2020)—are the aquatic eagle and other raptors of similar size (Erlandson and Moss, 2001). However, such birds do not nest in caves, and even less so when human occupation is frequent and regular. In the adult age indicated by the body size estimations, the brown crab can inhabit the low waters close to the shoreline and has a preference for living in the sea floor, which argues against the archaeological specimens representing predation by land-based animals, as does the estimated site-to-shore distance. Finally, the type of fracture observed on crab pincers, the fact that many remains are burnt, and the impact scars suggesting claw disarticulation further corroborate the anthropogenic nature of the accumulation of the MIS 5 crabs.
5.2. Crab harvesting and processing
Archaeological crab remains are mostly identified by their pincers, which are the most diagnostic body parts. Unsurprisingly, all five crab species represented at Gruta da Figueira Brava were identified that way; however, the presence of diverse skeletal parts, even if some are poorly represented, bespeaks of the introduction of complete animals. Experimental work on the crab cuticles confirms that pincers are the body part most likely to preserve due to its low porosity (Plotnick et al., 1988; Mutel et al., 2008; Waugh et al., 2009). Krause et al. (2011) have also shown that pincers have a high calcium carbonate content and, hence, preserve better than the other parts of the exoskeleton.
The taxa identified at Figueira Brava can all be found in the Portuguese Atlantic coast, and they are all edible, even though some (e.g., Pachygrapsus marmoratus) are currently ignored due to their small size. All species are intertidal and can easily be found in low tide pools among algae, beach boulders, and rock creeks. However, large size adults of Cancer pagurus and Maja squinado are most commonly reported to be found in such locations only during the summer months (sometimes until October), when seawater warms up and the crabs migrate to the shore for reproduction. During the rest of the year, large size indivuals of these species remain confined to the subtidal zone (Ingle, 1996; Gruet, 2002; Woll, 2006).
Nowadays, several commercially important species of crabs are harvested using entrapment gear, like basket traps or pots, baited with herring, squid, or shad. Only sessile or slow-moving animals, like molluscs, gooseneck barnacles, and sea urchins are harvested by hand (Flick, 2012). Nonetheless, and despite the crabs' quick movement and escape strategies, there are several ethnographic examples of hand-collection from low tide pools, frequently aided by spears. The Nehalem Tillamook men of the northern Oregon Coast of America hunted crabs using long poles to stab them (Losey et al., 2004). This practice is also mentioned by von Brandt (1984) for coastal countries in Western Europe. Such a technique can be easily applied on shallow waters, as described by Swan (1972) for the Chinook of Willapa Bay in Washingston State (USA), who waded for crabs of large size in low tide pools. The Squamish people (Canada) are known to gather crabs from the beach in the summer (Conner and Bethune-Johnson, 1986). This is an activity described for women from several indigenous peoples from the Pacific Northwest Coast (e.g., the Haida, Murdock, 1963; the Kwakiutl, Wolcott, 1967; the Squamish, Conner and Bethune-Johnson, 1986); whereas men tend to be associated with crab hunting by spearing, as observed among the Makah (Renker and Gunther, 1990), the Eastern Abenaki speaking people (Snow, 1978), or the Wampanoag Indians of Massachusetts (Speck and Dexter, 1948). Batdorf (1990) notes that Dungeness crabs (from the Cancer genus) were hit by spears on the area behind the eyes, in order to daze the crab and to facilitate hand collection. The Cancer pagurus assemblage from Gruta da Figueira Brava shows clear selection for large size animals, which reflects individual hunting and not mass collection. Large individuals are easier to spot than smaller ones and, if crabs had been mass harvested, one would expect to find specimens of all sizes, as described by Losey et al. (2004) for the Oregon coast of America, which is not the case.
How crabs were processed can be inferred from the burning and breakage patterns. Even though most burning is found on the pincers, it can be seen on carapace fragments too (Figure 4B), further supporting the notion that complete animals were processed at the site. Ethnographic studies indicate that crabs are not consumed raw, and that they are commonly boiled, steamed or roasted (Suttles, 1974; Batdorf, 1990; Emmons, 1991). Boiling and steaming are not likely to result in the burning of crab remains, but roasting on the coals does leave thermo-alteration marks. At Gruta da Figueira Brava, the most common are black burns. Microstructurally, the exoskeleton of crabs, the bones of mammals, and the shells of molluscs are different, but the lessons garnered from the experimental work done on molluscs by Villagran (2014) and Milano et al. (2016) can nonetheless be used to interpret crustaceans. Based on such studies, black burns on crab shell reflect exposure to fire temperatures between 300 and 500°C (Villagran, 2014; Milano et al., 2016), which are generally accepted as typical for cooking (e.g., Pearce and Luff, 1994; Montón-Subías, 2002). Ethnography demonstrates that crab meat is considered to be cooked when it no longer sticks to the shell (Batdorf, 1990). Exposure to temperatures starting at 300°C results in dramatic changes in shell surface color, texture, and microstructure: shells become more fragile and easier to break (Milano et al., 2016), and, when processed, they will shatter because of their lower density (Rick et al., 2015). The flaking marks found on the ventral proximal side of dactylopoda suggest manual disarticulation. The predominance of longitudinal fractures on Cancer pagurus pincers bespeaks of the intention to access the flesh within the fingers, while the carapace was targeted for its large brown meat content (Table 6), as is the case today.
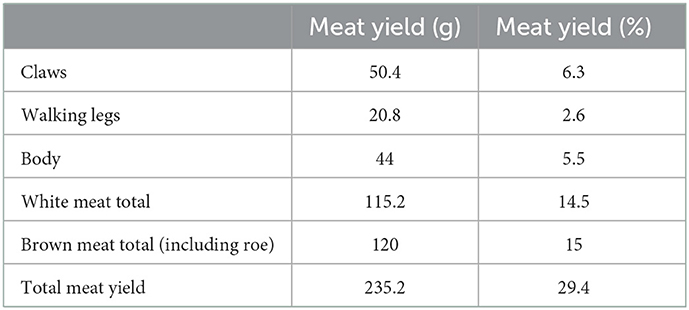
Table 6. Meat yield for boiled and hand-picked Cancer pagurus crabs, which on average were of 800 g live weight, adapting from the values given by Woll (2006) for female crabs caught in October.
Cancer pagurus is one of the crustaceans mostly consumed in Southern Europe—Portugal, Spain, France, and Italy—during the summer holidays and over Christmas (Barrento et al., 2008). The marks seen on the archaeological material are very similar to those empirically produced when eating them today. The disarticulation of the claw fingers is done by manually breaking the joint backwards, which is sometimes aided by a small hammer. Such hammers, however, are most valuable for breaking the fingers themselves to access the white meat inside, which recurrently produces the same kinds of longitudinal fractures observed on the archaeological material.
5.3. Dietary importance
Currently, crabs are seen as an important food resource, as they are considered to enhance good healthy living due to their high-quality and low-calorie protein, associated with a wide range of vitamins and minerals, but essentially due to the presence of long-chain polyunsaturated fatty acids (also known as PUFAs) including docosahexaenoic (DHA) and eicosapentaenoic (EPA) acids (Tsape et al., 2010; Hicks, 2012; Ovat et al., 2018). Cancer pagurus, in particular, provides ample doses of elements such as potassium, calcium, copper, zinc, selenium, and PUFAs. Overall, brown meat (i.e., the tissue in the body cavity, mostly gonads and the hepatopancreas) contains higher fat, cadmium, calcium, iron, magnesium, and copper, whereas muscle (i.e., the white meat in the legs and claws) has a higher zinc content. Therefore, such crabs are among the seafoods with a higher potential to contribute to the improvement of cardiovascular and neurological aspects of human health (Maulvault et al., 2012).
Such nutritional properties are even more relevant if the crabs in question are adult Cancer pagurus, as is the case at Gruta da Figueira Brava, and they can be enhanced through culinary treatment. Crab cooking results in a decrease of the moisture content and EPA but in an increase in mineral, protein, carbohydrate, carbon, zinc, bromine, and chlorine content (Maulvault et al., 2012). Therefore, despite the low ranking of crabs in diet-breadth models due to their small size and apparent limited caloric intake (e.g., Winterhalder and Smith, 2000; Bird and O'Connell, 2006), they offer the benefit of nutrient diversity (Hockett and Haws, 2003, 2005) and are an appealing protein, fat and PUFA package easily acquired from the nearby shore, within minimal processing and transport efforts. In terms of meat yield, the selection for 800 g crabs would guarantee about 200 g of edible crab meat, since about 70% of the weight corresponds to the exoskeleton (Table 6)—i.e., in terms of edible meat, a single crab is the weight-equivalent of up to 250 limpet shells.
With regards to current seasonal crab availability, opinions diverge, as noted by Gutiérrez Zugasti et al. (2016). For instance, Brown and Bennett (1980) report that Cancer pagurus juveniles can remain in intertidal waters year-round and that adults are rare on the shore during the winter months, but Silva et al. (2014) report year-round presence with little variation in specimen density. This could be explained by the optimal conditions of food availability provided by upwelling, which, for Portuguese coast Cancer pagurus, results in an earlier breeding season (Cunha et al., 2018). Even though seasonality is, therefore, difficult to assess, it is clear that, in Phase FB4, crabs were consistently consumed throughout. Food is not only consumed for the satisfaction of nutritional needs, but also to experience esteemed flavors and to fulfill specific roles in social life. As demonstrated by ethnographic studies in Australia (Bailey, 1975; Meehan, 1977, 1983), resources that appear as minor in quantitative or nutritional terms may in fact be of significant importance in social, as well as, in subsistence terms. For the Middle Paleolithic, however, whether such foods were perceived as tasteful, reflected some sort of festivity, added social value to whoever harvested them, or had other consumption-associated meanings is beyond our grasp.
6. Conclusion
The marine crustacean assemblage recovered from Gruta da Figueira Brava is the first known from the Middle Paleolithic. The anthropogenic nature of the accumulation found in the MIS 5 deposit, dominated by large size specimens of Cancer pagurus, indicating a harvest that targeted mature individuals with a high meat content, is unquestionable. Harvesting may have been done during the summer, when these animals come to shore for reproduction, but it is also possible that such resources were available and consumed year-round. It is likely that foraging was performed at low tide, by hand or with long poles. The catch was brought to the cave, cooked on coals, and consumed on site. Whether meanings other than the satisfaction of alimentary needs were involved must remain a matter for speculation; what is certain is that this resource provided coastal populations of Neanderthals with an ample array of nutrients rich in protein, DHA, calcium and sodium, which are important to a healthy diet.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
MN: conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, resources, visualization, and writing—original draft preparation. CD: formal analysis, methodology, resources, supervision, visualization, and writing—review and editing. JZ: funding acquisition, investigation, project administration, resources, supervision, visualization, and writing—review and editing. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the London Arts and Humanities Partnership (project 526079 - “Neanderthal subsistence in Portugal: Small and large prey consumption during the Marine Isotope Stage 5”), the Institute of Archaeology, University College London, IoA Awards (travel grant), Fundação para a Ciência e Tecnologia, Portugal (projects PTDC/HIS-ARQ/098164/2008 - “Middle Paleolithic Archaeology of the Almonda Karstic System,” PTDC/HAR-ARQ/30413/2017 - “Archaeology and Evolution of Early Humans in the Western Façade of Iberia,” UIDB/00698/2020, and UIDP/00698/2020). Additional financial support has been provided by MN's two-year postdoc contract for project “Neanderthal and Anatomically Modern Human interactions with small prey in Atlantic Iberia throughout the changing environments of the Pleistocene”, as part of the European Union's Horizon 2020 research and innovation programme under the Marie Skodowska-Curie grant agreement 101034349, and from the State Research Agency of the Spanish Ministry of Science and Innovation through the Program Maria de Maeztu Unit of Excellence (CEX2019-000945-M).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
1. ^Nabais, M., Portero, R., and Zilhão, J. (submitted). Neanderthal Brown Crab Recipes: A Combined Approach Using Experimental, Archaeological and Ethnographic Evidence. Paper submitted to “Historical Biology.”
References
Álvarez Fernández, E. (2010). Moluscos, crustáceos y equinodermos: restos de origen marino en los yacimientos prehistóricos. El Futuro Pasado 1, 83–95. doi: 10.14201/fdp.24497
Andrews, P. (1990). Owls, Caves and Fossils. Predation, Preservation, and Accumulation of Small Mammal Bones in Caves, with an Analysis of the Pleistocene Cazve faunas from Westbury-sub-Mendip, Somerset, UK. Chicago, IL: University of Chicago Press.
Antunes, M. (2000). Last Neanderthals in Portugal. Odontologic and Other Evidence. Classe das Ciências. Tomo XXXVIII. Lisboa: Academia das Ciências de Lisboa.
Araújo, A. C., Moreno García, M., Gabriel, S. (2014). Para além de Toledo: outros dados, novas revisões e algumas reflexões sobre o Mesolítico Antigo do litoral da Estremadura. Rev. Portuguesa Arqueol. 17, 5–34.
Arnaud, J. M. (2000). Os concheiros mesolíticos do vale do Sado e a exploração dos recursos estuarinos (nos tempos pré-históricos e na actualidade). Actas do Encontro sobre Arqueologia da Arrábida. Trabalhos Arqueol. 14, 21–44.
Ashkenazi, A., Motro, U., Goren-Inbar, N., Biton, R., Rabinovich, R. (2005). New morphometric parameters for assessment of body size in the fossil freshwater crab assemblage from the Acheulian site of Gesher Benot Ya'aqov, Israel. J. Archaeol. Sci. 32, 675–689. doi: 10.1016/j.jas.2004.12.003
Badal, E., Villaverde, V., Zilhão, J. (2011). The fire of Iberian Neanderthals. Wood charcoal from three new Mousterian sites in the Iberian Peninsula. Sagvntvm Extra 11, 77–78.
Bailey, G. (1975). The role of molluscs in coastal economies: the results of midden analysis in Australia. J. Archaeol. Sci. 2, 45–62. doi: 10.1016/0305-4403(75)90045-X
Barrento, S., Marques, A., Pedro, S., Vaz-Pires, P., Nunes, M. L. (2008). The trade of live crustaceans in Portugal: space for technological improvements. ICES J. Mar. Sci. 65, 551–559. doi: 10.1093/icesjms/fsn037
Bayed, A., Quiniou, F., Benrha, A., Guillou, M. (2005). The Paracentrotus lividus populations from the northern Moroccan Atlantic coast: growth, reproduction and health condition. J. Mar. Biol. Assoc. 85, 999–1007. doi: 10.1017/S0025315405012026
Bird, D. W., O'Connell, J. F. (2006). Behavioral ecology and archaeology. J. Atchaeol. Res. 14, 143–188. doi: 10.1007/s10814-006-9003-6
Bishop, G. (1986). Taphonomy of the North American Decapods. J. Crustacean Biol. 6, 326–355. doi: 10.2307/1548174
Blasco, R., Fernández Peris, J. (2012a). Small and large game: human use of diverse faunal resources at Level IV of Bolomor Cave (Valencia, Spain). Comptes Rendus Palevol 11, 265–282. doi: 10.1016/j.crpv.2012.01.003
Blasco, R., Fernández Peris, J. (2012b). A uniquely broad spectrum diet during the Middle Pleistocene at Bolomor Cave (Valencia, Spain). Quat. Int. 252, 16–31. doi: 10.1016/j.quaint.2011.03.019
Blasco, R., Rosell, J., Rufà, A., Sánchez Marco, A., Finlayson, C. (2016). Pigeons and choughs, a usual resource for the Neanderthals in Gibraltar. Quat. Int. 421, 62–77. doi: 10.1016/j.quaint.2015.10.040
Boudouresque, C. F., Verlaque, M. (2001). “Ecology of Paracentrotus lividus,” in Edible Sea Urchins, Biology and Ecology, ed J. M. Lawrence (Amsterdam: Elsevier Sciences B.V.), 177–216. doi: 10.1016/S0167-9309(01)80013-2
Brown, C. G., Bennett, D. B. (1980). Population and catch structure of the edible crab (Cancer pagurus) in the English Channel. J. Conseil 39, 88–100. doi: 10.1093/icesjms/39.1.88
Callapez, P. (2000). “Upper Pleistocene marine invertebrates from Gruta da Figueira Brava (Arrábida, Portugal),” in Last Neanderthals in Portugal. Odontologic and Other Evidence. Classe das Ciências. Tomo XXXVIII, ed M. Antunes (Lisboa: Academia das Ciências de Lisboa), 83–103.
Campbell, G. (2008). Sorry, wrong phylum: a neophyte archaeomalacologist's experiences in analyzing a European Atlantic Sea Urchin assemblage. Archaeofauna 17, 77–90.
Capaldo, S., Blumenschine, R. (1994). A quantitative diagnosis of notches made by hammerstone percussion and carnivore gnawing on bovid long bones. Am. Antiq. 59, 724–748. doi: 10.2307/282345
Cardoso, J. (1993). Contribuição para o conhecimento dos grandes mamíferos do Plistocénico Superior de Portugal. Oeiras: Câmara Municipal de Oeiras.
Carvalho, A. F. (2001). A Neolitização do Portugal Meridional. Os exemplos de Maciço Calcário Estremenho e do Algarve Ocidental. Promontoria Monografica, 12. Faro: Universidade do Algarve.
Claassen, C. (2013). “Invertebrates on San Salvador island, Bahamas: the use of shellfish as bait,” in Shell Energy: Mollusc Shells as Coastal Resources, eds G. N. Bailey, K. Hardy, and A. Camara (Oxford: Oxbow Books), 59–67.
Conner, D. C. G., Bethune-Johnson, D. (1986). Our Coast Salish Way of Life - The Squamish. Scarborough, ON: Prentice-Hall Canada Inc.
Córtez-Sánchez, M., Morales-Muñiz, A., Simón-Vallejo, M., Lozano-Francisco, M., Vera-Peláez, J., Finlayson, C., et al. (2011). earliest known use of marine resources by Neanderthals. PLoS ONE 6, e24026. doi: 10.1371/journal.pone.0024026
Crothers, J., Crothers, M. (1988). A key to the crabs and crab-like animals of british inshore waters. Field Stud. 5, 753–806.
Cunha, I., Azevedo, T., Vasconcelos, V., Almeida, J. R. (2018). Distribution ranges of the acorn barnacle Perforatus (=Balanus) perforates (Bruguière, 1789) in the NE Atlantic are influenced by reproductive parameters. Hydrobiologia 806, 227–235. doi: 10.1007/s10750-017-3362-5
Díez, J., Fernández-Jalvo, Y., Rosell, J., Cáceres, I. (1999). Zooarchaeology and taphonomy of Urora Stratum (Gran Dolina, Sierra de Atapuerca, Spain). J. Hum. Evol. 37, 623–652. doi: 10.1006/jhev.1999.0346
Dupont, C. (2011). “Les invertébrés marins du ≪ concheiro ≫ de Toledo (Lourinhã, Portugal),” in O Concheiro de Toledo no contexto do Mesolítico Inicial do Litoral da Estremadura, ed A. C. Araújo (Lisboa: Igespar), 185–227.
Dupont, C. (2017). Connaitre l'exploitation du littoral par l'Homme à partir des invertébrés marins découverts en contexte archéologique. Nouvelles Archéol. 148, 28–33. doi: 10.4000/nda.3714
Dupont, C., Bicho, N. (2015). “Marine invertebrates and models of economic organization of the coastal zone during the Mesolithic: French and Portuguese examples. Chapter seven,” in Muge 150th, The 150th Anniversary of the Discovery of Mesolithic Shellmiddens, vol. 1, eds N. Bicho, C. Detry, T. D. Price, and E. Cunha (Newcastle: Cambridge Scholars Publishing), 89–103.
Dupont, C., Gruet, Y. (2005). Malacofaune et crustacés marins des amas coquilliers mésolithiques de Beg-an-Dorchenn (Plomeur, Finistère) et de Beg-er-Vil (Quiberon, Morbihan). Mémoire Soc. Préhistorique Française 36, 139–161.
Dupont, C., Gruet, Y. (2022). Why mesolithic populations only started eating crabs on the European atlantic façade over the past 15 years? Open Archaeol. 8, 670–695. doi: 10.1515/opar-2022-0255
Dupont C. Gruet Y. Arthur M. Digard O. (in press). “A question of size! The importance of marine crabs in food remains from Mesolithic fisher-hunter-gatherers at Beg-er-Vil (Quiberon, Morbihan, France),” in Investigate the Shore, Sound the Past: New Methods Practices of Maritime Prehistory. Actes de la séance de la Société préhistorique française de Brest 2-4 décembre 2020, eds P. Stephan, G. Marchand, Y. Pailler (Paris: Société préhistorique française).
Dupont, C., Marchand, G., Carrión Marco, Y., Desse-Berset, N., Gaudin, L., Gruet, Y., et al. (2010). Beg-an-Dorchenn (Plomeur, Finistère): une fenêtre ouverte sur l'exploitation du littoral par les peuples mésolithiques du VIe millenaire dans l'Ouest de la France. Bull. Soc. Préhist. Française 107, 227–290. doi: 10.3406/bspf.2010.13929
Dupont, C., Schulting, R., Tresset, A. (2007). “Prehistoric shell middens along the French Atlantic façade: the use of marine and terrestrial resources in the diets of coastal human populations,” in Shell Middens in Atlantic Europe, eds N. Milner, O. Craig, and G. Bailey (Oxford: Oxbow Books), 123–135.
Dusseldorp, G. (2010). Prey choice during the South African middle stone age: avoiding Dangerous prey or maximising returns? Afr. Archaeol. Rev. 27, 107–133. doi: 10.1007/s10437-010-9072-2
Emmons, G. T. (1991). “Food and its preparation,” in The Tlingit Indians, ed F. de Laguna (New York, NY: American Museum of Natural History), 140–153.
Erlandson, J., Moss, M. (2001). Shellfish feeders, carrion eaters, and the archaeology of aquatic adaptations. Am. Antiquity 66, 413–432. doi: 10.2307/2694242
Finlayson, C. (2008). On the importance of coastal areas in the survival of Neanderthal populations during the Late Pleistocene. Quat. Sci. Rev. 27, 2246–2252. doi: 10.1016/j.quascirev.2008.08.033
Fisher, J. (1995). Bone surface modifications in zooarchaeology. J. Archaeol. Method Theory 2, 7–68. doi: 10.1007/BF02228434
Flick, G. J. (2012). “Harvesting techniques,” in The Seafood Industry: Species, Products, Processing, and Safety, eds L. Ankenman Granata, G. J. Flick, R. E. Martin (Chichester: Wiley-Blackwell), 14–26. doi: 10.1002/9781118229491.ch2
García-Escárzaga, A., Gutiérrez Zugasti, I., González-Morales, M., Cobo-García, A. (2017). Shells and Humans: molluscs and other coastal resources from the earliest human occupations at the mesolithic shell midden of El Mazo (Asturias, Northern Spain). Papers Inst. Archaeol. 27, 1–17. doi: 10.5334/pia-481
Gruet, Y. (2002). Reconnaissance de quelques espèces communes de crustacés (balanes et crabes): application au site Mésolithique de Beg-er-Vil (Morbihan, France). Rev. Archéomét. 26, 125–139. doi: 10.3406/arsci.2002.1028
Gruet, Y. (2009). “Les Crustacés: crabes pêchés,” in Des premiers paysans aux premiers métallurgistes sur les côtes charentaises, Chapitre 6 - Approches stylistiques, technologiques et fonctionnelles du mobilier: la parure. Mémoire XXXIII, ed L. Laporte (Chauvigny: Association des Publications Chauvinoise), 582–585.
Gruet, Y., Laport, L. (1996). Crabes pêchés au Néolithique Final à Ponthezières: identification des modes de pêche et application de la mètrique. Rev. Archéométr. 1, 197–201.
Gutiérrez Zugasti, I. (2009). La Exploitación de Moluscos y Otros Recursos Litorales en la Región Cantábrica Durante el Pleistoceno Final. Santander: Universidad de Cantábria.
Gutiérrez Zugasti, I. (2011). The use of echinoids and crustaceans as food during the pleistocene-holocene transition in northern Spain: methodological contribution and dietary assessment. J. Island Coast. Archaeol. 6, 115–133 doi: 10.1080/15564894.2010.487421
Gutiérrez Zugasti, I., Cuenca-Solana, D., González-Morales, M., García-Moreno, A. (2013). “Exploitation of molluscs as food during the Gravettian at Fuente del Salín Cave (Cantabria, Northern Spain),” in Ancient Maritime Communities and the Relationship between People and Environment along the European Atlantic Coasts, eds M. Daire, C. Dupont, A. Baudry, C. Billard, J. Large, L. Lespez, et al. (BAR International Series), 2570.
Gutiérrez Zugasti, I., Tong, E., García-Escárzaga, A., Cuenca-Solana, D., Beiley, G. N., González-Morales, M. R. (2016). Collection and consumption of echinoderms and crustaceans at the Mesolithic shell midden site of El Mazo (northern Iberia): opportunistic behaviour or social strategy? Quat. Int. 407, 118–130. doi: 10.1016/j.quaint.2015.11.149
Haigh, J. A., Pantin, J. R., Salomonsen, H., Kaiser, M. J. (2015). Size at maturity of the edible crab (Cancer pagurus) in Welsh waters. Fisheries Conservation Science report No 51. Bangor University, 26
Hardy, B., Moncel, M. H. (2011). Neanderthal use of fish, mammals, birds, starchy plants and wood 125-250,000 years ago. PLoS ONE 6, e23768. doi: 10.1371/journal.pone.0023768
Hayward, P. J., Ryland, J. S. (1995). Handbook of the Marine Fauna of North-West Europe. Oxford: Oxford University Press.
Hicks, D. T. (2012). “Nutrition and preparation,” in The Seafood Industry: Species, Products, Processing, and Safety, eds L. Ankenman Granata, G. J. Flick, R. E. Martin (Chichester: Wiley-Blackwell), 172–192 doi: 10.1002/9781118229491.ch15
Hockett, B., Haws, J. (2003). Nutritional ecology and diachronic trends in Paleolithic diet and health. Evol. Anthropol. 12, 211–216. doi: 10.1002/evan.10116
Hockett, B., Haws, J. (2004). “Theoretical perspectives on the dietary role of small animals in human evolution,” in Petits animaux et sociétés humaines. Du complément alimentaire aux ressources utilitaires, eds J. P. Brugal and J. Desse (Antibes: Éditions APDCA).
Hockett, B., Haws, J. (2005). Nutritional ecology and the human demography of Neandertal extinction. Quat. Int. 137, 21–34. doi: 10.1016/j.quaint.2004.11.017
Ingle, R. (1996). “Shallow-water crabs. Keys and notes for identification of the species,” in Synopses of the British Fauna (New Series), eds R. Barnes and J. Crothers (Shrewsbury: Field Studies Council), 25.
Jacinto, D., Bulleri, F., Benedetti-Cecchi, L., Cruz, T. (2013). Patterns of abundance, population size structure and microhabitat usage of Paracentrotus lividus (Echinodermata: Echinoidea) in SW Portugal and NW Italy. Mar. Biol. 160, 1135–1146. doi: 10.1007/s00227-013-2166-z
Jaouen, K., Richards, M. P., Le Cabec, A., Welker, F., Rendu, W., Hublin, J. J., et al. (2019). Exceptionally high δ15N values in collagen single amino acids confirm Neandertals as high-trophic levels carnivores. Proc. Natl. Acad. Sci. U. S. A. 116, 4928–4933. doi: 10.1073/pnas.1814087116
Jerardino, A., Marean, C. (2010). Shellfish gathering, marine palaeoecology and modern human behavior: perspectives from cave PP13B, Pinnacle Point, South Africa. J. Hum. Evol. 59, 412–424. doi: 10.1016/j.jhevol.2010.07.003
Klein, R., Avery, G., Cruz-Uribe, K., Halkett, D., Parkington, J., Steele, T., et al. (2004). The Ysterfontein 1 Middle Stone Age site, South Arfica, and early human exploitation of coastal resources. Proc. Natl. Acad. Sci. U. S. A. 101, 5708–5715. doi: 10.1073/pnas.0400528101
Koops, K., Wrangham, R. W., Cumberlidge, N., Fitzgerald, M. A., van Leeuwen, K. L., Rothman, J. M., et al. (2019). Crab-fishing by chimpanzees in the Nimba Mountains, Guinea. J. Hum. Evol. 133, 230–241. doi: 10.1016/j.jhevol.2019.05.002
Krause, R., Parsons-Hubbard, K., Walker, S. (2011). Experimental taphonomy of a decapod crustacean: long-term data and their implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 312, 350–362. doi: 10.1016/j.palaeo.2011.03.020
Kyriacou, K., Parkington, J., Will, M., Kandel, A., Conrad, N. (2015). Middle and Later Stone Age shellfish exploitation strategies and coastal foraging at Hoedjiespunt and Lynch Point, Saldanha Bay, South Africa. J. Archaeol. Sci. 57, 197–206. doi: 10.1016/j.jas.2015.01.018
Langejans, G., Niekerk, K., Dusseldorp, G., Thackeray, J. (2012). Middle Stone Age shellfish exploitation: potential indications for mass collecting and resource intensification at Blombos Cave and Klasies River, South Africa. Quat. Int. 270, 80–94. doi: 10.1016/j.quaint.2011.09.003
Lentacker, A. (1986). Preliminary results of the fauna of cabeço de Amoreira and Cabeço de Arruda (Muge, Portugal). Trabalhos Antropol. Etnol. 1–4, 9–26.
Lohse, D. P., Raimondi, P. T. (2007). “Barnacles,” in Encyclopedia of Tidepools and Rocky Shores, eds M. W. Denny and S. D. Gaines (Berkeley, CA; Los Angeles, CA: University of California Press), 61–64.
Losey, R., Yamada, S., Largaespada, L. (2004). Late-Holocene Dungeness crab (Cancer magister) harvest at an Oregon coast estuary. J. Archaeol. Sci. 31, 1603–1612. doi: 10.1016/j.jas.2004.04.002
Lubell, D., Jackes, M., Sheppard, P., Rowley-Conwy, P. (2007). “The Mesolithic-Neolithic in the Alentejo: archaeological investigations, 1984-1986,” in From the Mediterranean basin to the Portuguese Atlantic Shore: Papers in Honor of Anthony Marks, ed N. Bicho (Faro: Universidade do Algarve), 119–139.
Marean, C., Bar-Matthews, M., Bernatchez, J., Fisher, E., Goldberg, P., Herries, A., et al. (2007). Early human use of marine resources and pigment in South Africa during the Middle Pleistocene. Nature 449, 905–908. doi: 10.1038/nature06204
Martin, R. E. (2012). “A history of the seafood industry,” in The Seafood Industry: Species, Products, Processing, and Safety, eds L. Ankenman Granata, G. J. Flick and R. E. Martin (Chichester: Wiley-Blackwell), 1–13. doi: 10.1002/9781118229491.ch1
Maulvault, A. L., Anacleto, P., Lourenço, H. M., Carvalho, M. L., Nunes, M. L., Marques, A. (2012). Nutritional quality and safety of cooked edible crab (Cancer pagurus). Food Chem. 133, 277–283. doi: 10.1016/j.foodchem.2012.01.023
Meehan, B. (1977). “Man does not live by calories alone: the role of shellfish in coastal cuisine,” in Sunda and Sahul: Prehistoric Studies in Southeast Asia, Melanesia, and Australia, eds J. Allen, J. Golson, and R. Jones, R. (London: Academic Press), 493–531.
Meehan, B. (1983). “A matter of choice? Some thoughts on shell gathering strategies in northern Australia,” in Animals and Archaeology 2: Shell Middens, Fishes and Birds, Vol. 183, eds C. Grigson and J. Clutton-Block (British Archaeological Reports, International Series), 3–17.
Milano, S., Prendergast, A. L., Schöne, B. R. (2016). Effects of cooking on mollusk shell structure and chemistry: implications for archeology and paleoenvironmental reconstruction. J. Archaeol. Sci. Rep. 7, 14–26. doi: 10.1016/j.jasrep.2016.03.045
Montón-Subías, S. (2002). “Cooking in zooarchaeology: is this issue still raw?” inConsuming Passions and Patterns of Consumption, eds P. Miracle and N. Milner (Cambridge: McDonald Institute of Archaeological Research), 7–15.
Morin, E., Meier, J., El Guennouni, K., Moigne, A. M., Lebreton, L., Rusch, L., et al. (2019). New evidence of broader diets from archaic Homo populations in the northwestern Mediterranean. Sci. Adv. 5, eaav9106. doi: 10.1126/sciadv.aav9106
Mougne, C. (2015). Exploitation et utilisation des invertébrés marins durant la Protohistoire sur le territoire continental et littoral Manche-Atlantique français (Unpublished Ph.D dissertation). Université de Rennes 1, France. doi: 10.4000/archeopages.900
Murdock, G. P. (1963). The Haida of British Colombia: Our Primitive Contemporaries. New York, NY: MacMillan Co.
Mutel, M. H. E., Waugh, D. A., Feldmann, R. M., Parsons-Hubbard, K. (2008). Experimental taphonomy of Callinectes sapidus and cuticular controls on preservation. Palaios 23, 615–623. doi: 10.2110/palo.2008.p08-024r
Nabais, M., Zilhão, J. (2019). The consumption of tortoise among Last Interglacial Iberian Neanderthals. Quat. Sci. Rev. 217, 225–246. doi: 10.1016/j.quascirev.2019.03.024
Naito, Y. I., Chikaraishi, Y., Drucker, D. G., Ohkouchi, N., Semal, P., Wiβing, C., et al. (2016). Ecological niche of Neanderthals from Spy Cave revealed by nitrogen isotopes of individual amino acids in collagen. J. Hum. Evol. 93, 82–90. doi: 10.1016/j.jhevol.2016.01.009
Nicholson, R. (1993). A morphological investigation of burnt animal bone and an evaluation of its utility in archaeology. J. Archaeol. Sci. 20, 411–428. doi: 10.1006/jasc.1993.1025
Oesterling, M. J. (2012). “Shellfish – crustaceans,” in The Seafood Industry: Species, Products, Processing, and Safety, eds L. Ankenman Granata, G. J. Flick and R. E. Martin (Chichester: Wiley-Blackwell), 83–94. doi: 10.1002/9781118229491.ch7
Ovat, O. I., Edet, D. I., Suleiman, I. D., Agabi, E. D. (2018). Assessement of edible crab harvesting and consumption in Southern Cross River State, Nigeria. J. Res. Forestry Wildl. Environ. 12, 24–32.
Pearce, J., Luff, R. (1994). “The taphonomy of cooked bone,” in Whither Environmental Archaeology?, eds R. Luff and P. Rowley-Conwy (Oxford: Oxbow Books), 51–56.
Pickering, T., Egeland, C. (2006). Experimental patterns of hammerstone percussion damage on bones: implications for inferences of carcass processing by humans. J. Archaeol. Sci. 33, 459–469. doi: 10.1016/j.jas.2005.09.001
Pinto, R. S. (1986). Les fouilles du Kjoekkenmoedding de Cabeço de Amoreira à Muge (Potugal). Trabalhos Antropol. Etnol. 26, 220–229.
Plotnick, R. E., Baumiller, T., Wetmore, K. L. (1988). Fossilization potential of the mud crab, Panopeus (Brachyura, Xanthidae) and temporal variability in crustacean taphonomy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 63, 27–43. doi: 10.1016/0031-0182(88)90089-2
Renker, A. M., Gunther, E. (1990). “Makah,” in Handbook of North American Indians, volume 7: Northwest Coast, ed W. Suttles (Washington, DC: Smithsonian Institution), 422–426.
Ribeiro, C. (1884). Les kjoekkenmoedings de la vallée du Tage. Comptes rendus de la IXème session du congrès International d'Anthropologie et d'Archéologie Préhistorique (Lisbonne. 1880). Paris: Typographie de L'académie Royale des Sciences, 279–290
Rick, T., Ogburn, M., Kramer, M., McCanty, S., Reeder-Myers, L., Miller, H., et al. (2015). Archaeology, taphonomy, and historical ecology of Chesapeake Bay blue crabs (Callinectes sapidus). J. Archaeol. Sci.55, 42–54. doi: 10.1016/j.jas.2014.12.016
Roche, J. (1958). Le gisement mésolithique de Moita do Sebastião (Muge, Portugal). Archeologie, MCMLX. Lisboa: Instituto de Alta Cultura.
Russon, A., Compost, A., Kuncoro, P., Ferisa, A. (2014). Orangutan fish eating, primate aquatic fauna eating, and their implications for the origins of ancestral hominin fish eating. J. Hum. Evol. 77, 50–63. doi: 10.1016/j.jhevol.2014.06.007
Shipman, P., Foster, G., Schoeninger, M. (1984). Burnt bones and teeth: an experimental study of color, morphology, crystal structure and shrinkage. J. Archaeol. Sci. 11, 307–325. doi: 10.1016/0305-4403(84)90013-X
Silva, A. C. F., Boaventura, D. M., Thompson, R. C., Hawkins, S. J. (2014). Spatial and temporal patterns of subtidal and intertidal crabs excursions. J. Sea Res. 85, 343–348. doi: 10.1016/j.seares.2013.06.006
Silva, C. T., Soares, J. (1997). Economias costeiras na Pré-História do Sudoeste portugês. O concheiro de Montes de Baixo. Setúbal Arqueol. 11–12, 69–108.
Snow, D. R. (1978). “Eastern Abenaki,” in Handbook of North American Indians, vol. 5, ed D. Damas (Washington, DC: Smithsonian Institution), 285–298.
Southward, A. (2008). Barnacles. Keys and Notes for the Identification of British Species. Synopses of the British Fauna (New Series). Dorchester: Field Studies Council, 57.
Southward, E., Campbell, A. (2006). Echinoderms. Keys and Notes for the Identification of British Species. Synopses of the British Fauna (New Series). Dorchester: Field Studies Council, 56.
Speck, F. G., Dexter, R. W. (1948). Utilization of marine life by the Wampanoag Indians of Massachussetts. J. Washington Acad. Sci. 38, 257–265.
Stringer, C., Finlayson, J., Barton, R., Fernández-Jalvo, Y., Cáceres, I., Sabin, R., et al. (2008). Neanderthal exploitation of marine mammals in Gibraltar. Proc. Natl. Acad. Sci. U. S. A. 105, 14319–14324. doi: 10.1073/pnas.0805474105
Suttles, W. P. (1974). Coast Salish and Western Washington Indians The Economic Life of the Coast Salish of Haro and Rosario Straits. New York, NY; London: Garland Publishing Inc.
Swan, J. G. (1972). The Northwest Coast, Or, Three Years Residence in Washington Territory. Seattle, WA: University of Washington Press.
Thackeray, J. (1988). Molluscan Fauna From Klasies River, South Africa. South Afr. Archaeol. Bull. 43, 27–32. doi: 10.2307/3887610
Tomas, F., Romero, J., Turon, X. (2004). Settlement and recruitment of the sea urchin Paracentrotus lividus in two contrasting habitats in the Mediterranean. Mar. Ecol. Progress Series 282, 173–184. doi: 10.3354/meps282173
Tsape, K., Sinanoglu, V. J., Miniadis-Meimaroglou, S. (2010). Comparative analysis of the fatty acids and sterol profiles of widely consumed Mediterranean crustacean species. Food Chem. 122, 292–299. doi: 10.1016/j.foodchem.2010.02.019
Valente, M. J. (2008). As últimas sociedades de caçadores-recolectores no Centro e Sul de Portugal (10.000 – 6.000 anos BP): aproveitamento dos recursos animais (Unpublished Ph.D. thesis from the Universidade do Algarve).
Villa, P., Mahieu, E. (1991). Breakage patterns of human long bones. J. Hum. Evol. 21, 27–48. doi: 10.1016/0047-2484(91)90034-S
Villagran, X. (2014). Experimental micromorphology on burnt shells of Anomalocardia brasiliana (Gmelin 1791) (Bivalvia, Veneridae) and its potential for identification of combustion features on shell-matrix sites. Geoarchaeol. Int. J. 29, 389–396. doi: 10.1002/gea.21486
Volman, T. (1978). Early Archaeological Evidence for Shellfish Collecting. Science 201:911–913. doi: 10.1126/science.201.4359.911
von Brandt, A. (1984). Fish Catching Methods of the World. 3rd Edition. Stratford-upon-Avon, Warwickshire: Avon Litho Ltd.
Waugh, D. A., Feldmann, R. M., Burrell, J. L., Hull, A. L., Hein, K., Schweitzer, C. E. (2009). Ontogenic variations in cuticle morphology in the blue crab Callinected sapidus Rathbun, 1896. J. Crustacean Biol. 29, 141–156. doi: 10.1651/08-3105.1
White, T. (1992). Prehistoric Cannibalism at Mancos 5MTURM-2346. Princeton, NJ: Princeton University Press. doi: 10.1515/9781400852925
Will, M., Parkington, J., Kandel, A., Conrad, N. (2013). Coastal adaptations and the Middle Stone Age lithic assemblages from Hoedjiespunt 1 in the Western Cape, South Africa. J. Hum. Evol. 64, 518–537. doi: 10.1016/j.jhevol.2013.02.012
Winterhalder, B., Smith, E. (2000). Analyzing adaptive strategies: human behavioural ecology at twenty-five. Evol. Antropol. 9, 51–72. doi: 10.1002/(SICI)1520-6505(2000)9:2andlt;51::AID-EVAN1andgt;3.0.CO;2-7
Woll, A. K. (2006). The Edible Crab: Biology, Grading and Handling Live Crabs. Ålesund: Møresforsking Ålesund.
WoRMS Editorial Board (2023). World Register of Marine Species. Available online at: https://www.marinespecies.org at VLIZ (accessed January 18, 2023).
Zilhão, J. (2012). “Neandertals from World's End: results of recent research,” in Biodiversidad Humana y Evolución, eds D. Turbón, L. Fañanás, C. Rissech, A. Rosa (Barcelona: Universidad de Barcelona, Sociedad Española de Antropología Física), 68–77.
Keywords: crab, marine resources, diet breadth, Middle Paleolithic, Pleistocene, Cancer pagurus
Citation: Nabais M, Dupont C and Zilhão J (2023) The exploitation of crabs by Last Interglacial Iberian Neanderthals: The evidence from Gruta da Figueira Brava (Portugal). Front. Environ. Archaeol. 2:1097815. doi: 10.3389/fearc.2023.1097815
Received: 14 November 2022; Accepted: 04 January 2023;
Published: 07 February 2023.
Edited by:
Cheryl Makarewicz, University of Kiel, GermanyReviewed by:
Marshall Weisler, The University of Queensland, AustraliaMarcello A. Mannino, Aarhus University, Denmark
Copyright © 2023 Nabais, Dupont and Zilhão. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariana Nabais,  bWFyaWFuYW5hYmFpc0BnbWFpbC5jb20=
bWFyaWFuYW5hYmFpc0BnbWFpbC5jb20=