 Tingting Deng1†
Tingting Deng1† Xixue Lu2†
Xixue Lu2† Xuemin Jia2
Xuemin Jia2 Jinxin Du1
Jinxin Du1 Lijuan Wang1,2
Lijuan Wang1,2 Baorui Cao1
Baorui Cao1 Meina Yang3,4*
Meina Yang3,4* Ying Yin5*
Ying Yin5* Fanjie Liu2*
Fanjie Liu2*- 1College of Traditional Chinese Medicine, Shandong University of Traditional Chinese Medicine, Jinan, China
- 2Bone Biomechanics Engineering Laboratory of Shandong Province, Shandong Medicinal Biotechnology Center (School of Biomedical Sciences), Neck-Shoulder and Lumbocrural Pain Hospital of Shandong First Medical University, Shandong First Medical University & Shandong Academy of Medical Sciences, Jinan, China
- 3National Health Commission (NHC) Key Laboratory of Biotechnology Drugs (Shandong Academy of Medical Sciences), Biomedical Sciences College, Shandong First Medical University, Jinan, China
- 4Department of Endocrinology, The First Affiliated Hospital of Shandong First Medical University, Jinan, China
- 5Department of Acupuncture, Affiliated Hospital of Shandong University of Traditional Chinese Medicine, Jinan, China
Background: Previous observational epidemiological studies reported an association between cathepsins and cancer, however, a causal relationship is uncertain. This study evaluated the causal relationship between cathepsins and cancer using Mendelian randomization (MR) analysis.
Methods: We used publicly available genome-wide association study (GWAS) data for bidirectional MR analysis. Inverse variance weighting (IVW) was used as the primary MR method of MR analysis.
Results: After correction for the False Discovery Rate (FDR), two cathepsins were found to be significantly associated with cancer risk: cathepsin H (CTSH) levels increased the risk of lung cancer (OR = 1.070, 95% CI = 1.027–1.114, P = 0.001, PFDR= 0.009), and CTSH levels decreased the risk of basal cell carcinoma (OR = 0.947, 95% CI = 0.919–0.975, P = 0.0002, PFDR= 0.002). In addition, there was no statistically significant effect of the 20 cancers on the nine cathepsins. Some unadjusted low P-value phenotypes are worth mentioning, including a positive correlation between cathepsin O (CTSO) and breast cancer (OR = 1.012, 95% CI = 1.001–1.025, P = 0.041), cathepsin S (CTSS) and pharyngeal cancer (OR = 1.017, 95% CI = 1.001–1.034, P = 0.043), and CTSS and endometrial cancer (OR = 1.055, 95% CI = 1.012–1.101, P = 0.012); and there was a negative correlation between cathepsin Z and ovarian cancer (CTSZ) (OR = 0.970, 95% CI = 0.949–0.991, P = 0.006), CTSS and prostate cancer (OR = 0.947, 95% CI = 0.902–0.944, P = 0.028), and cathepsin E (CTSE) and pancreatic cancer (OR = 0.963, 95% CI = 0.938–0.990, P = 0.006).
Conclusion: Our MR analyses showed a causal relationship between cathepsins and cancers and may help provide new insights for further mechanistic and clinical studies of cathepsin-mediated cancer.
1 Introduction
Cathepsins are a class of proteases found in various animal tissues intracellular (particularly in the lysosomal fraction). They finely regulate biological processes, such as proteolysis, metabolite storage, foreign body removal, immune response, and apoptosis, through efficient, highly selective, and limited specific substrate cleavage, thereby maintaining normal body homeostasis. However, irregularities in protein hydrolysis activity or “imbalances” of insufficient protease activity or excessive protein hydrolysis or dysregulation of signaling pathways are causative factors in diseases (1), including cancer, cardiovascular diseases, inflammatory and autoimmune diseases (2). A variety of catalytically active cathepsins act as potent effectors that alter the tumour microenvironment by remodeling the extracellular matrix (ECM) (at neutral pH), as well as the activation, processing, or degradation of chemokines, cytokines, and growth factors (3, 4). They also promote tissue invasion and metastasis by releasing cell adhesion molecules (5, 6) and are part of a dynamic response to anticancer therapy in the tumour microenvironment (7–9).
Recent studies have revealed the role of several cathepsins in promoting or inhibiting various cancers (e.g., lung (10), ovarian (11), thyroid (12), and colorectal (13)), including cathepsin B (CTSB) (14), cathepsin L (CTSL) (15), cathepsin G (CTSG) (16), and cathepsin S (CTSS) (17). However, few observational studies and clinical trials have investigated the relationship between cathepsins and cancer. Previous studies reported the high CTSB expression in pancreatic ductal adenocarcinoma (PDAC) cells in serum samples from patients with PDAC (18). One study found that the serum cystatin/CTSB ratio was a prognostic indicator of survival in patients with esophageal cancer (19). CTSS levels are significantly elevated in the sera of patients with gastric, esophageal, liver, colorectal, nasopharyngeal, and lung cancers (20). Despite extensive research, no uniform or conclusive study has been conducted on the correlation between cathepsins and cancer. Therefore, there is a need for further research on the causal relationship between the different types of cathepsins and cancer risk.
Mendelian randomization (MR) uses exposure-related genetic variants as instrumental variables (IVs) to robustly assess causality between exposure and outcome (21, 22). As alleles are randomly assigned and do not change in response to disease onset, MR analyses effectively reduce the influence of confounding factors, avoid reverse causation bias, and yield more reliable causal effects than observational studies (23, 24). MR analysis is now widely used to explore causal associations between exposure factors and cancer (25, 26). In oncology, MR analysis can provide insight into the complex relationship between exposure factors and cancer development, providing a basis for prevention and treatment in clinical research (27). Therefore, this study collected data on nine cathepsins and cancers from a large-scale genome-wide association study (GWAS), performed two-sample MR, followed by inverse MR to adjust for the pleiotropic effects of genetic tools and potential confounders, and assessed potential genetic-causal associations between cathepsins and cancers to provide a basis for future prevention and treatment strategies.
2 Materials and methods
2.1 Study design
A GWAS was performed for nine cathepsins and 20 cancers from the IEU GWAS database (https://gwas.mrcieu.uk/) at the University of Bristol, UK. Cathepsin data were obtained from an INTERVAL study, which included 3,301 Europeans (28). All donors completed a trial consent form, and the INTERVAL study was approved by the US National Research Ethics Committee (11/EE/0538). Considering the effect of linkage disequilibrium (LD) among SNPs, we screened for SNPs that were independent of each other and had genome-wide significance in the strength of association with cathepsin from the pooled GWAS data of cathepsin using the following screening criteria (29): (1)P < 5×10–6 of the correlation effect between cathepsin and IVs; (2) the physical distance between every two genes > 10,000 kb; and (3) R2 < 0.001 for LD between genes.
2.2 Data source
The GWAS summary statistics for a wide range of cancers were obtained from publicly available databases from the MRC IEU OpenGWAS (MR-base) database. We identified 20 cancer outcomes: bladder, lung, anal, testicular, thyroid, colorectal, ovarian, prostate, breast, esophageal, pharyngeal, endometrial, pancreatic, cecum, sialadenitis, hepatocellular, vulvar, gastric, basal cell, and bronchogenic carcinomas. The number of cases ranged from 105 to 122,188 (Supplementary Table S1).
2.3 Selection of IVs
We refer to the three core assumptions of association, independence, and exclusivity, which must be fulfilled in MR analyses. Single nucleotide polymorphisms (SNPs) with the genome-wide significance of association strength with cathepsins were selected as IVs. Weak IV bias was determined using the F-test statistic, and no weak IV bias was considered to exist if F > 10. The F statistic was calculated as F = [(N-K-1)/K]×[R2/(1-R2)], where N is the sample size, K is the number of IVs, and R2 denotes the variance of the exposure explained by each IV alone (30). A flowchart of the study is shown in Figure 1.
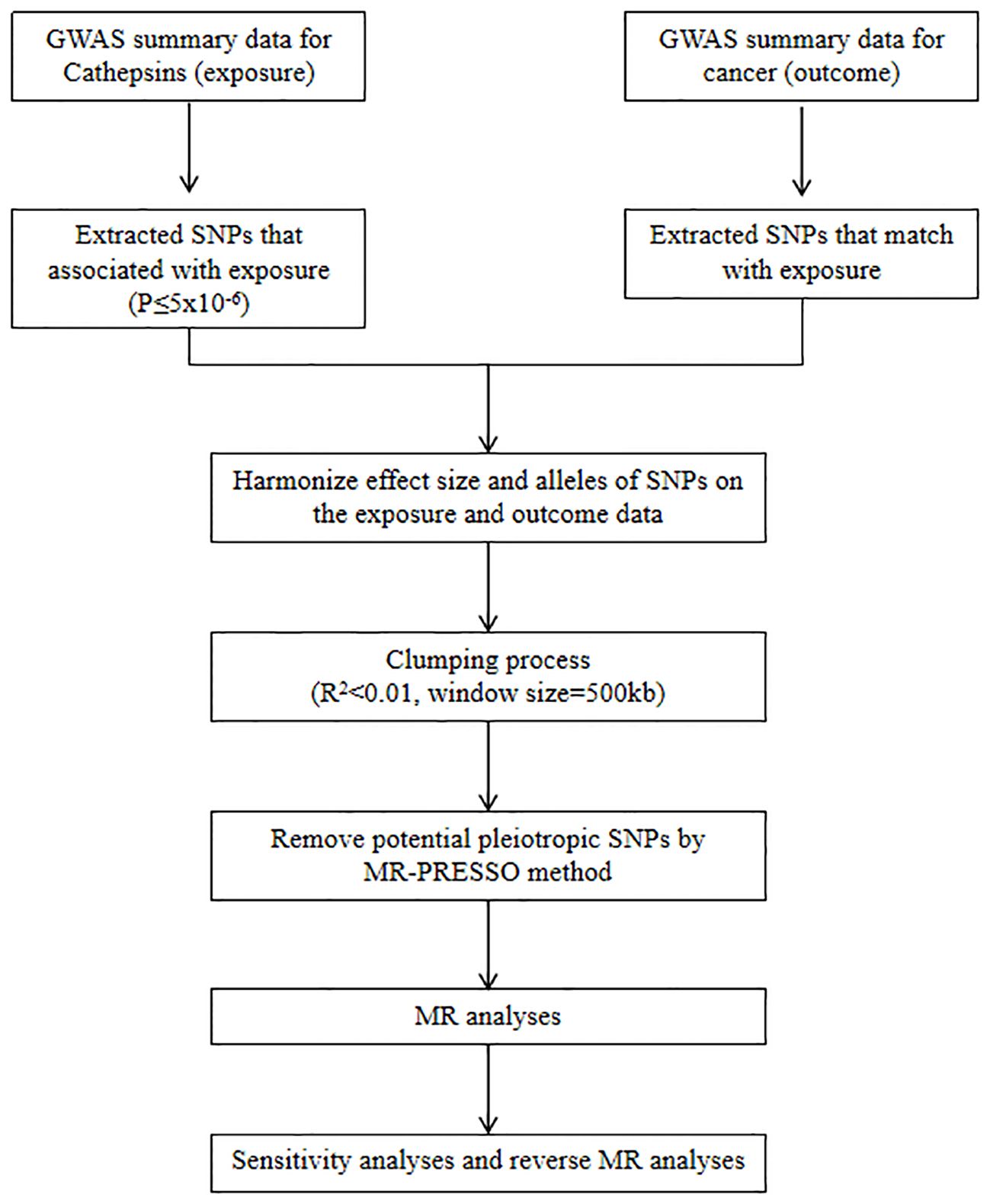
Figure 1 Study design and workflow.
2.4 Statistical analysis
Determination of the causal relationship between cathepsins and cancer risk was carried out in two-sample Mendelian randomization using five methods: Inverse variance weighting (IVW) (31), MR-Egger (32), Weighted Median (33), Simple Mode (34), and Weighted Mode (35). Odds ratios (OR) and 95% confidence intervals (CI) were used to determine whether a causal relationship existed between cathepsins and cancer risk. According to previous studies, the IVW method is superior to other tests (36, 37), and is used as the main MR analysis method (38–40). Among these, the IVW was used as the primary method of analysis. Because of the multiple exposures and outcomes in this study, multiple test corrections were performed using the FDR method (41), and it was necessary to report whether the P-values tested by the IVW method reached nominal significance (P < 0.05) and statistical significance (PFDR < 0.05). The MR-Egger intercept was used to assess the relationship between IVs and other potential confounders and to ensure that the selected IVs did not influence the outcome variables through pathways other than exposure factors. Horizontal pleiotropy (27) is indicated if the MR-Egger intercept analysis shows a statistically significant relationship (P < 0.05). At P < 0.05, an outlier test was used to eliminate horizontal pleiotropy using the MR-PRESSO global test (42). An OR less than 1 indicates that exposure plays a protective role in predicting the occurrence of an outcome event. In other words, exposure played a positive role in preventing or reducing the occurrence of outcome events. Conversely, if the OR is greater than 1, the exposure is categorized as a risk factor for the outcome, and exposure can promote the occurrence of the outcome. Cochran’s Q statistic was used to perform the heterogeneity test. Statistically significant (P < 0.05) Cochran’s Q test proves that the analyses were significantly heterogeneous (43).
We performed a reverse MR analysis (20 cancers as exposures and cathepsins as outcomes) to explore whether cancer has a causal effect on cathepsins identified in the forward MR analysis. The analysis procedure was consistent with that of the forward MR analysis.
MR analyses were performed using “TwoSampleMR” (version 0.5.6) in R (version 4.2.3), Mendelian Randomization (0.7.0), and TwoSample MR (0.5.6). P < 0.05 indicates that the results are statistically significant.
3 Results
3.1 IVs selection
Based on the screening criteria, nine IVs for cathepsin were included in this study. The F-statistic for each IV was > 10, indicating low evidence of weak IV bias (Supplementary Data 1).
3.2 MR main analysis results
The IVW approach revealed significant evidence of a causal relationship between cathepsins and different cancer risks. Our pooled analysis identified nine cathepsins that exhibited potential causal associations with 20 cancers (Figure 2). Of the 180 associations included (9 exposures x 20 outcomes), six were statistically significant in the IVW analysis (Figure 2). Cathepsin H (CTSE) levels reduced the risk of vulvar carcinoma (OR = 0.483, 95% CI = 0.241–0.966, P = 0.039), and cathepsin H (CTSH) levels reduced basal cell carcinoma risk (OR = 0.947, 95% CI = 0.919–0.975, P = 0.0002); CTSF levels increased the risk of vulvar carcinoma (OR = 1.736, 95% CI = 1.026–2.937, P = 0.040), CTSS levels increased the risk of colorectal cancer (OR = 1.051, 95% CI = 1.008–1.097, P = 0.02), CTSZ levels increased the risk of thyroid cancer (OR = 1.157, 95% CI = 1.017–1.317, P = 0.026), CTSH levels increased the risk of lung cancer (OR = 1.070, 95% CI = 1.027–1.114, P = 0.001).
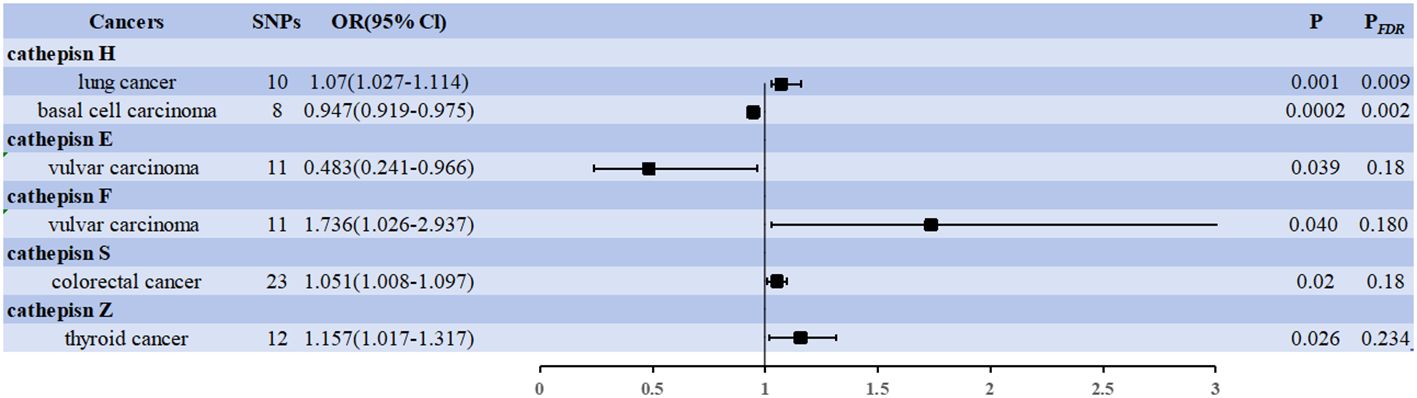
Figure 2 Forest plots showed the causal associations between cathepsins and cancers. IVW, inverse variance weighting; CI, confidence interval; FDR, false discovery rate.
Two associations were based on the number of exposure-outcome pairs showing FDR-corrected significance (P < 0.05). CTSH levels increased the risk of lung cancer (OR = 1.070, 95% CI = 1.027–1.114, P = 0.001, PFDR = 0.009), and CTSH levels decreased the risk of basal cell carcinoma (OR = 0.947, 95% CI = 0.919–0.975, P = 0.0002, PFDR = 0.002). These two associations had 10 and 8 IVs, respectively, and the robustness of these causal relationships was further supported by combined data from multiple sensitivity analyses (Supplementary Data 2). Specifically, our analyses by Cochran’s Q did not reveal any signs of heterogeneity (P = 0.729 > 0.05, P = 0.065 > 0.05). The MR-Egger intercept assessment did not provide evidence of horizontal pleiotropy (P = 0.236 > 0.05, P = 0.969 > 0.05).
3.3 Reverse MR analysis results
We used cancer as the exposure factor, cathepsins as the outcome, and cancer-associated SNPs (P < 5 × 10–5) as the IVs to explore whether there was reverse causality for the significant results obtained. Figure 3 shows the six cathepsin immunophenotypes potentially affected by cancer. After reverse analysis, six were statistically significant: a positive correlation between CTSO and breast cancer (OR = 1.012, 95% CI = 1.001–1.025, P = 0.041), CTSS (OR = 1.017, 95% CI = 1.001–1.034, P = 0.043) and pharyngeal cancer, and CTSS (OR = 1.055, 95% CI = 1.012–1.101, P = 0.012) and endometrial cancer; There was a negative correlation between CTSZ and ovarian cancer (OR = 0.970, 95% CI = 0.949–0.991, P = 0.006), CTSS and prostate cancer (OR = 0.947, 95% CI = 0.902–0.944, P = 0.028), CTSE and pancreatic cancer (OR = 0.963, 95% CI = 0.938–0.990, P = 0.006). The Cochrane Q-test provided no evidence of heterogeneity (P = 0.388 > 0.05, P = 0.837 > 0.05; P = 0.909 > 0.05, P = 0.221 > 0.05, P = 0.667 > 0.05, P = 0.667 > 0.05, P = 0.832 > 0.05). SNP pleiotropy was not detected for the MR-Egger test intercept (P = 0.872 > 0.05, P = 0.393 > 0.05, P = 0.695 > 0.05; P = 0.200 > 0.05, P = 0.558 > 0.05, P = 0.290 > 0.05). These associations, based on the number of exposure-outcome pairs, did not show FDR-corrected significance (P > 0.05) (Figure 3). The results of the heterogeneity and pleiotropy tests are presented in Supplementary Data 3.
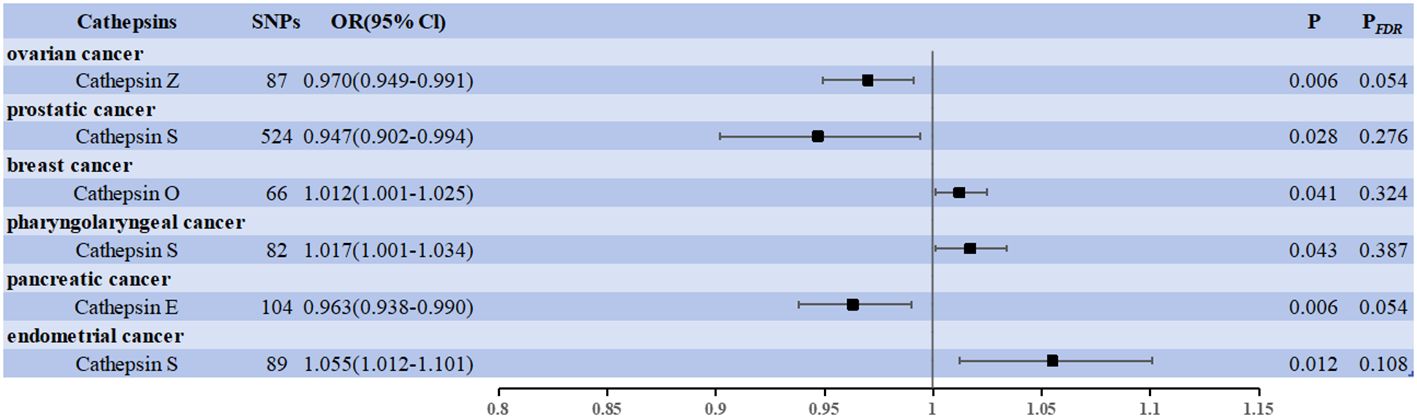
Figure 3 Forest plots showed the causal associations between cancers and cathepsins. IVW, inverse variance weighting; CI, confidence interval; FDR, false discovery rate.
4 Discussion
This study investigated the causal association between cathepsin levels and cancer. The causal effects of nine cathepsins on 20 cancers were comprehensively evaluated by MR analysis. The results showed a causal association between certain cathepsins and cancers, suggesting that cathepsins may have an essential influence on cancer and play an important role in cancer development.
In recent decades, the incidence of various types of cancer has increased; cancer has become a significant public health problem worldwide. It is the second leading cause of death in humans, after cardiovascular diseases (44). CTSH acts as an aminopeptidase and endopeptidase with endo protein hydrolytic activity and can hydrolyze a wide range of proteins (45). CTSH has been detected in type II pneumocytes and alveolar macrophages in the lung (46, 47). It is located in lamellipodia, dense multivesicular vesicles, and type II complex vesicle pneumocytes, which constitute sites of surfactant maturation (48, 49). Microarray analysis studies have shown that CTSH expression is lower in non-small cell lung cancer than in normal lung tissue (50) and that CTSH is involved in SP-B maturation by cleaving the peptide bond between pro-SP-B residues 279 and 280 (51, 52). Some studies have also found that silencing of CTSH significantly reduces SP-B maturation and subsequently reduces SP-B secretion (53). CTSH progression in lung cancer may regulate the sPLA2-PKCδ-MAPKs-cPLA2α pathway by modulating SP-B maturation, thereby regulating lipid metabolism in the lungs (54, 55). CTSH is highly expressed in small cells and in adenocarcinomas (56, 57). Luyapan et al. (58) conducted a transcriptome-wide association study using expression weights from a quantitative trait locus study of lung expression and found that the gene most strongly associated with lung cancer was CTSH.
The epidermis of the skin constantly undergoes cell renewal and differentiation to maintain its normal structure and function. However, when the balance between renewal and differentiation is disrupted, uncontrolled cell proliferation and cancer can result (59). Basal cell carcinoma, the most common form of skin cancer, originates in the basal layer of the epidermis and appendages. The tumor grows slowly, rarely metastasizes, and generally infiltrates the surrounding tissues slowly (60). The interplay between various environmental, genetic, phenotypic, and genetic risk factors contributes to the development of basal cell carcinomas. Cathepsin is an essential protease required for invasion. It has been found that CTSH is mainly localized in the lowermost basal cell layer (61). Basal cells are undifferentiated and can grow and divide. CTSH is a lysosomal cysteine protease involved in the degradation of extracellular matrix components and has been found to be more active in basal cell carcinoma tumors than in normal skin tissue (62). The mechanism underlying the involvement of CTSH in the development of basal cell carcinoma has not yet been investigated. However, CTSH activity is dysregulated in tissues surrounding basal cell carcinoma tumors, leading to its overexpression and secretion into the extracellular space to degrade structural proteins such as collagen and fibronectin (8, 63–67), thereby regulating the structure and stability of the extracellular matrix and promoting tumor cell invasion (68, 69).
It is also worth noting that breast cancer was associated with elevated CTSO, pharyngeal and endometrial cancers with elevated CTSS, ovarian cancer with decreased CTSZ, prostate cancer with decreased CTSS, and pancreatic cancer with decreased CTSE. CTSO was found to be significantly overexpressed in T47D, CAMA-1, and ZR75–1 cells, reducing BRCA1 levels and promoting cell proliferation by promoting the cysteine protease-mediated degradation of metadherin, polyadenylate-binding protein 4-like, recombinant lamin A/C, and recombinant eukaryotic translation elongation factor 1 alpha 1 protein levels (70–72). However, CTSS and CTSE are overexpressed in prostate cancer (73) and pancreatic cancer (74), respectively; this is contrary to the results of the present study and needs to be verified by more clinical and experimental studies in the future.
Previous studies did not comprehensively analyze the causal relationship between cathepsins and cancer. This study used two samples of MR studies and obtained reliable results: firstly, MR analysis has the advantage of avoiding reverse causal associations and confounders and saving time and resources compared to observational studies; secondly, according to our analysis, multiple cathepsins are risk and protective factors for cancers, and this study did not reveal potential horizontal pleiotropy, thus confirming the reliability of the conclusions. However, there are some limitations to this study. First, this study only observed a causal effect of cathepsins on the risk of multiple cancers at the gene level. Future MR studies with larger sample sizes and randomized controlled trials are required to validate these results. Second, the study was limited to the European population, and it is not possible to demonstrate whether the findings can be extended to other populations. Furthermore, as with all published MR studies, the possibility that unobserved pleiotropy affects the results cannot be ruled out, even if measures are taken to identify and eliminate aberrant variants (21); the study was unable to infer a non-linear correlation between cathepsins and cancers. Lastly, the cathepsins and cancer GWAS data were obtained from publicly available databases, and subgroup analyses were not possible due to the lack of detailed clinical patient information. In summary, the results of this study, using two-sample and inverse MR methods, suggest a causal relationship between cathepsins and various cancers. The results of this study should be interpreted with caution. More investigative studies should be conducted to validate the results and consider their application in clinical trials.
5 Conclusion
In conclusion, these results suggest a potential causal relationship between cathepsins and cancer. These findings provide new insights for further mechanistic studies on cathepsin-mediated cancers, potential targets, and new biomarkers for the early diagnosis and interventional therapy of cancers.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
Ethical approval was not required for the study involving humans in accordance with the local legislation and institutional requirements. Written informed consent to participate in this study was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and the institutional requirements.
Author contributions
TD: Conceptualization, Methodology, Writing – original draft. XL: Visualization, Writing – original draft. XJ: Data curation, Project administration, Writing – original draft. JD: Data curation, Project administration, Writing – original draft. LW: Software, Writing – original draft. BC: Formal Analysis, Investigation, Writing – original draft. MY: Funding acquisition, Writing – review & editing. YY: Resources, Supervision, Writing – review & editing. FL: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was financially supported by the National Natural Science Foundation of China (No. 82004212), the State Administration of Traditional Chinese Medicine Science and technology department co-build traditional Chinese medicine science and technology project (No.GZY-KJS-SD-2023-084), the Shandong Province Medical Health Science and Technology Development Plan Project (No. 202204070951), the TCM Science and Technology Project of Shandong Province (No. 2021M175), and the Shandong Traditional Chinese medicine classic famous collaborative innovation center open subject (No.2019KFY07).
Acknowledgments
We would like to express our gratitude to all the researchers and contributors to the GWASs. We would also like to express our gratitude to all the GWASs for opening up their summary data to the public.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2024.1428433/full#supplementary-material
References
1. Ruiz-Blázquez P, Pistorio V, Fernández-Fernández M, et al. The multifaceted role of cathepsins in liver disease. J Hepatol. (2021) 75:1192–202. doi: 10.1016/j.jhep.2021.06.031
2. Smyth P, Sasiwachirangkul J, Williams R, Cathepsin S. (CTSS) activity in health and disease - A treasure trove of untapped clinical potential. Mol Aspects Med. (2022) 88:101106. doi: 10.1016/j.mam.2022.101106
3. Soond SM, Kozhevnikova MV, Zamyatnin AA Jr. 'Patchiness' and basic cancer research: unravelling the proteases. Cell Cycle. (2019) 18:1687–701. doi: 10.1080/15384101.2019.1632639
4. Gocheva V, Wang HW, Gadea BB, et al. IL-4 induces cathepsin protease activity in tumor-associated macrophages to promote cancer growth and invasion. Genes Dev. (2010) 24:241–55. doi: 10.1101/gad.1874010
5. Sevenich L, Bowman RL, Mason SD, et al. Analysis of tumour- and stroma-supplied proteolytic networks reveals a brain-metastasis-promoting role for cathepsin S. Nat Cell Biol. (2014) 16:876–88. doi: 10.1038/ncb3011
6. Joyce JA, Baruch A, Chehade K, et al. Cathepsin cysteine proteases are effectors of invasive growth and angiogenesis during multistage tumorigenesis. Cancer Cell. (2004) 5:443–53. doi: 10.1016/S1535-6108(04)00111-4
7. Verbovšek U, Van Noorden CJ, Lah TT. Complexity of cancer protease biology: Cathepsin K expression and function in cancer progression. Semin Cancer Biol. (2015) 35:71–84. doi: 10.1016/j.semcancer.2015.08.010
8. Palermo C, Joyce JA. Cysteine cathepsin proteases as pharmacological targets in cancer. Trends Pharmacol Sci. (2008) 29:22–8. doi: 10.1016/j.tips.2007.10.011
9. Soond SM, Kozhevnikova MV, Frolova AS, et al. Lost or Forgotten: The nuclear cathepsin protein isoforms in cancer. Cancer Lett. (2019) 462:43–50. doi: 10.1016/j.canlet.2019.07.020
10. Xiao Y, Cong M, Li J, et al. Cathepsin C promotes breast cancer lung metastasis by modulating neutrophil infiltration and neutrophil extracellular trap formation. Cancer Cell. (2021) 39:423–437.e7. doi: 10.1016/j.ccell.2020.12.012
11. Pranjol MZ, Gutowski N, Hannemann M, Whatmore J. The potential role of the proteases cathepsin D and cathepsin L in the progression and metastasis of epithelial ovarian cancer. Biomolecules. (2015) 5:3260–79. doi: 10.3390/biom5043260
12. Kim EK, Song MJ, Jang HH, Chung YS. Clinicopathologic analysis of cathepsin B as a prognostic marker of thyroid cancer. Int J Mol Sci. (2020) 21:9537. doi: 10.3390/ijms21249537
13. Wang Z, Chen K, Zhang K, He K, Zhang D, Guo X, et al. Agrocybe cylindracea fucoglucogalactan induced lysosome-mediated apoptosis of colorectal cancer cell through H3K27ac-regulated cathepsin D. Carbohydr Polym. (2023) 319:121208. doi: 10.1016/j.carbpol.2023.121208
14. Xu LB, Qin YF, Su L, Huang C, Xu Q, Zhang R, et al. Cathepsin-facilitated invasion of BMI1-high hepatocellular carcinoma cells drives bile duct tumor thrombi formation. Nat Commun. (2023) 14:7033. doi: 10.1038/s41467-023-42930-y
15. Liu P, Ju M, Zheng X, Jiang Y, Yu X, Pan B, et al. Methyltransferase-like 3 promotes cervical cancer metastasis by enhancing cathepsin L mRNA stability in an N6-methyladenosine-dependent manner. Cancer Sci. (2023) 114:837–54. doi: 10.1111/cas.15658
16. Tanigawa K, Kiriya M, Hayashi Y, Shinden Y, Kijima Y, Natsugoe S, et al. Cathepsin G-induced Malignant progression of MCF-7 cells involves suppression of PAF signaling through induced expression of PAFAH1B2. Biochim Biophys Acta Mol Cell Biol Lipids. (2022) 1867:159164. doi: 10.1016/j.bbalip.2022.159164
17. Small DM, Burden RE, Jaworski J, Hegarty SM, Spence S, Burrows JF, et al. Cathepsin S from both tumor and tumor-associated cells promote cancer growth and neovascularization. Int J Cancer. (2013) 133:2102–12. doi: 10.1002/ijc.28238
18. Jiang Y, Han L, Xue M, Wang T, Zhu Y, Xiong C, et al. Cystatin B increases autophagic flux by sustaining proteolytic activity of cathepsin B and fuels glycolysis in pancreatic cancer: CSTB orchestrates autophagy and glycolysis in PDAC. Clin Transl Med. (2022) 12:e1126. doi: 10.1002/ctm2.1126
19. Yan Y, Zhou K, Wang L, Wang F, Chen X, Fan Q. Clinical significance of serum cathepsin B and cystatin C levels and their ratio in the prognosis of patients with esophageal cancer. Onco Targets Ther. (2017) 10:1947–54. doi: 10.2147/OTT
20. Liu WL, Liu D, Cheng K, Liu YJ, Xing S, Chi PD, et al. Evaluating the diagnostic and prognostic value of circulating cathepsin S in gastric cancer. Oncotarget. (2016) 7:28124–38. doi: 10.18632/oncotarget.v7i19
21. Birney E. Mendelian randomization. Cold Spring Harb Perspect Med. (2022) 12:a041302. doi: 10.1101/cshperspect.a041302
22. Sekula P, Del Greco MF, Pattaro C, Köttgen A. Mendelian randomization as an approach to assess causality using observational data. J Am Soc Nephrol. (2016) 27:3253–65. doi: 10.1681/ASN.2016010098
23. Carter AR, Sanderson E, Hammerton G, Richmond RC, Davey Smith G, Heron , et al. Mendelian randomisation for mediation analysis: current methods and challenges for implementation. Eur J Epidemiol. (2021) 36:465–478. doi: 10.1007/s10654-021-00757-1
24. Pang Y, Kartsonaki C, Lv J, Fairhurst-Hunter Z, Millwood IY, Yu C, et al. Associations of Adiposity, Circulating Protein Biomarkers, and Risk of Major Vascular Diseases [published correction appears in JAMA Cardiol. 2021 Feb 1;6(2):246]. JAMA Cardiol. (2021) 6:276–86. doi: 10.1001/jamacardio.2020.6041
25. Yu N, Qi H, Guo Y, Wu L, Su J, Huang K, et al. Associations between rheumatoid arthritis and skin cancer: A bidirectional two-sample Mendelian randomization study. J Am Acad Dermatol. (2024) 90:198–200. doi: 10.1016/j.jaad.2023.09.046
26. Chen Y, Xie Y, Ci H, Cheng Z, Kuang Y, Li S, et al. Plasma metabolites and risk of seven cancers: a two-sample Mendelian randomization study among European descendants. BMC Med. (2024) 22:90. doi: 10.1186/s12916-024-03272-8
27. Gala H, Tomlinson I. The use of Mendelian randomisation to identify causal cancer risk factors: promise and limitations. J Pathol. (2020) 250:541–54. doi: 10.1002/path.5421
28. Sun BB, Maranville JC, Peters JE, Stacey D, Staley JR, Blackshaw J, et al. Genomic atlas of the human plasma proteome. Nature. (2018) 558:73–9. doi: 10.1038/s41586-018-0175-2
29. Noyce AJ, Kia DA, Hemani G, Nicolas A, Price TR, De Pablo-Fernandez E, et al. Estimating the causal influence of body mass index on risk of Parkinson disease: A Mendelian randomisation study. PloS Med. (2017) 14:e1002314. doi: 10.1371/journal.pmed.1002314
30. Levin MG, Judy R, Gill D, Vujkovic M, Verma SS, Bradford Y, et al. Genetics of height and risk of atrial fibrillation: A Mendelian randomization study. PloS Med. (2020) 17:e1003288. doi: 10.1371/journal.pmed.1003288
31. Mounier N, Kutalik Z. Bias correction for inverse variance weighting Mendelian randomization. Genet Epidemiol. (2023) 47:314–31. doi: 10.1002/gepi.22522
32. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger. Eur J Epidemiol. (2017) 32:377–89. doi: 10.1007/s10654-017-0255-x
33. Li P, Wang H, Guo L, Gou X, Chen G, Lin D, et al. Association between gut microbiota and preeclampsia-eclampsia: a two-sample Mendelian randomization study. BMC Med. (2022) 20:443. doi: 10.1186/s12916-022-02657-x
34. Huang D, Lin S, He J, Wang Q, Zhan Y. Association between COVID-19 and telomere length: A bidirectional Mendelian randomization study. J Med Virol. (2022) 94:5345–53. doi: 10.1002/jmv.28008
35. Wu F, Huang Y, Hu J, Shao Z. Mendelian randomization study of inflammatory bowel disease and bone mineral density. BMC Med. (2020) 18:312. doi: 10.1186/s12916-020-01778-5
36. Lin Z, Deng Y, Pan W. Combining the strengths of inverse-variance weighting and Egger regression in Mendelian randomization using a mixture of regressions model. PloS Genet. (2021) 17:e1009922. doi: 10.1371/journal.pgen.1009922
37. Bowden J, Davey Smith G, Haycock PC, Haycock PC, Burgess S. Consistent estimation in mendelian randomization with some invalid instruments using a weighted median estimator. Genet Epidemiol. (2016) 40:304–14. doi: 10.1002/gepi.21965
38. Cui K, Song N, Fan Y, Zeng L, Shi P, Wang Z, et al. A two-sample Mendelian randomization analysis: causal association between chemokines and pan-carcinoma. Front Genet. (2023) 14:1285274. doi: 10.3389/fgene.2023.1285274
39. Tang Y, Zhang L, Ye D, Zhao A, Liu Y, Zhang M, et al. Causal relationship between Type 1 diabetes and osteoporosis and fracture occurrence: a two-sample Mendelian randomization analysis. Osteoporos Int. (2023) 34:1111–7. doi: 10.1007/s00198-023-06734-6
40. Yang H, Shi P, Li M, Kong L, Liu S, Jiang L, et al. Mendelian-randomization study reveals causal relationships between nitrogen dioxide and gut microbiota. Ecotoxicol Environ Saf. (2023) 267:115660. doi: 10.1016/j.ecoenv.2023.115660
41. Glickman ME, Rao SR, Schultz MR. False discovery rate control is a recommended alternative to Bonferroni-type adjustments in health studies. J Clin Epidemiol. (2014) 67:850–7. doi: 10.1016/j.jclinepi.2014.03.012
42. Verbanck M, Chen CY, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases [published correction appears. Nat Genet. (2018) 50:693–8. doi: 10.1038/s41588-018-0099-7
43. Bowden J, Del Greco MF, Minelli C, Davey Smith G, Sheehan N, Thompson J, et al. A framework for the investigation of pleiotropy in two-sample summary data Mendelian randomization. Stat Med. (2017) 36:1783–802. doi: 10.1002/sim.7221
44. Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. (2023) 73:17–48. doi: 10.3322/caac.21763
45. Wang Y, Zhao J, Gu Y, Wang H, Jiang M, Zhao S, et al. Cathepsin H: Molecular characteristics and clues to function and mechanism. Biochem Pharmacol. (2023) 212:115585. doi: 10.1016/j.bcp.2023.115585
46. Ishii Y, Hashizume Y, Kominami E, Uchiyama Y. Changes in immunoreactivity for cathepsin H in rat type II alveolar epithelial cells and its proteolytic activity in bronchoalveolar lavage fluid over 24 hours. Anat Rec. (1991) 230:519–23. doi: 10.1002/ar.1092300411
47. Brasch F, Ten Brinke A, Johnen G, Ochs M, Kapp N, Müller KM, et al. Involvement of cathepsin H in the processing of the hydrophobic surfactant-associated protein C in type II pneumocytes. Am J Respir Cell Mol Biol. (2002) 26:659–70. doi: 10.1165/ajrcmb.26.6.4744
48. Ishii Y, Hashizume Y, Watanabe T, Waguri S, Sato N, Yamamoto M, et al. Cysteine proteinases in bronchoalveolar epithelial cells and lavage fluid of rat lung. J Histochem Cytochem. (1991) 39:461–8. doi: 10.1177/39.4.2005374
49. Woischnik M, Bauer A, Aboutaam R, Pamir A, Stanzel F, de Blic J, et al. Cathepsin H and napsin A are active in the alveoli and increased in alveolar proteinosis. Eur Respir J. (2008) 31:1197–204. doi: 10.1183/09031936.00081207
50. Ueno T, Linder S, Na CL, Rice WR, Johansson J, Weaver TE, et al. Processing of pulmonary surfactant protein B by napsin and cathepsin H. J Biol Chem. (2004) 279:16178–84. doi: 10.1074/jbc.M312029200
51. Guttentag S, Robinson L, Zhang P, Brasch F, Bühling F, Beers M, et al. Cysteine protease activity is required for surfactant protein B processing and lamellar body genesis. Am J Respir Cell Mol Biol. (2003) 28:69–79. doi: 10.1165/rcmb.2002-0111OC
52. Bühling F, Kouadio M, Chwieralski CE, Kern U, Hohlfeld JM, Klemm N, et al. Gene targeting of the cysteine peptidase cathepsin H impairs lung surfactant in mice. PloS One. (2011) 6:e26247. doi: 10.1371/journal.pone.0026247
53. Lee S, Kim D, Kang J, Kim E, Kim W, Youn H, et al. Surfactant protein B suppresses lung cancer progression by inhibiting secretory phospholipase A2 activity and arachidonic acid production. Cell Physiol Biochem. (2017) 42:1684–700. doi: 10.1159/000479418
54. Han WK, Sapirstein A, Hung CC, Alessandrini A, Bonventre JV. Cross-talk between cytosolic phospholipase A2 alpha (cPLA2 alpha) and secretory phospholipase A2 (sPLA2) in hydrogen peroxide-induced arachidonic acid release in murine mesangial cells: sPLA2 regulates cPLA2 alpha activity that is responsible for arachidonic acid release. J Biol Chem. (2003) 278:24153–63. doi: 10.1074/jbc.M300424200
55. Hite RD, Grier BL, Waite BM, Veldhuizen RA, Possmayer F, Yao LJ, et al. Surfactant protein B inhibits secretory phospholipase A2 hydrolysis of surfactant phospholipids. Am J Physiol Lung Cell Mol Physiol. (2012) 302:L257–65. doi: 10.1152/ajplung.00054.2011
56. Schweiger A, Staib A, Werle B, Krasovec M, Lah TT, Ebert W, et al. Cysteine proteinase cathepsin H in tumours and sera of lung cancer patients: relation to prognosis and cigarette smoking. Br J Cancer. (2000) 82:782–8. doi: 10.1054/bjoc.1999.0999
57. Linnerth NM, Sirbovan K, Moorehead RA. Use of a transgenic mouse model to identify markers of human lung tumors. Int J Cancer. (2005) 114:977–82. doi: 10.1002/ijc.20814
58. Luyapan J, Bossé Y, Li Z, Xiao X, Rosenberger A, Hung RJ. Candidate pathway analysis of surfactant proteins identifies CTSH and SFTA2 that influences lung cancer risk. Hum Mol Genet. (2023) 32:2842–55. doi: 10.1093/hmg/ddad095
59. Hsu YC, Fuchs E. Building and maintaining the skin. Cold Spring Harb Perspect Biol. (2022) 14:a040840. doi: 10.1101/cshperspect.a040840
60. Heath MS, Bar A. Basal cell carcinoma. Dermatol Clin. (2023) 41:13–21. doi: 10.1016/j.det.2022.07.005
61. Rinne A, Kirschke H, Järvinen M, Hopsu-Havu VK, Wieranders B, Bohley P. Localization of cathepsin H and its inhibitor in the skin and other stratified epithelia. Arch Dermatol Res. (1985) 277:190–4. doi: 10.1007/BF00404315
62. Fröhlich E, Möhrle M, Klessen C. Cathepsins in basal cell carcinomas: activity, immunoreactivity and mRNA staining of cathepsins B, D, H and L [published correction appears. Arch Dermatol Res. (2004) 295:411–21. doi: 10.1007/s00403-003-0449-9
63. Villalobo E, Moch C, Fryd-Versavel G, Fleury-Aubusson A, Morin L, et al. Cysteine proteases and cell differentiation: excystment of the ciliated protist Sterkiella histriomuscorum. Eukaryot Cell. (2003) 2:1234–45. doi: 10.1128/EC.2.6.1234-1245.2003
64. Jokimaa V, Oksjoki S, Kujari H, Anttila L. Expression patterns of cathepsins B, H, K, L and S in the human endometrium. Mol Hum Reprod. (2001) 7:73–8. doi: 10.1093/molehr/7.1.73
65. Sevenich L, Joyce JA. Pericellular proteolysis in cancer. Genes Dev. (2014) 28:2331–47. doi: 10.1101/gad.250647.114
66. Gocheva V, Joyce JA. Cysteine cathepsins and the cutting edge of cancer invasion. Cell Cycle. (2007) 6:60–4. doi: 10.4161/cc.6.1.3669
67. Tsushima H, Ueki A, Matsuoka Y, Mihara H, Hopsu-Havu VK. Characterization of a cathepsin-H-like enzyme from a human melanoma cell line. Int J Cancer. (1991) 48:726–32. doi: 10.1002/ijc.2910480516
68. Pickup MW, Mouw JK, Weaver VM. The extracellular matrix modulates the hallmarks of cancer. EMBO Rep. (2014) 15:1243–53. doi: 10.15252/embr.201439246
69. Wu SM, Huang YH, Yeh CT, Tsai M-M, Liao C-H, Cheng W-L, et al. Cathepsin H regulated by the thyroid hormone receptors associate with tumor invasion in human hepatoma cells. Oncogene. (2011) 30:2057–69. doi: 10.1038/onc.2010.585
70. Cairns J, Ingle JN, Wickerham LD, Weinshilboum R, Liu M, Wang L, et al. SNPs near the cysteine proteinase cathepsin O gene (CTSO) determine tamoxifen sensitivity in ERα-positive breast cancer through regulation of BRCA1. PloS Genet. (2017) 13:e1007031. doi: 10.1371/journal.pgen.1007031
71. Somasundaram K, Zhang H, Zeng YX, Houvras Y, Peng Y, Zhang H, et al. Arrest of the cell cycle by the tumour-suppressor BRCA1 requires the CDK-inhibitor p21WAF1/CiP1. Nature. (1997) 389:187–90. doi: 10.1038/38291
72. Ouchi T, Monteiro AN, August A, Aaronson SA, Hanafusa H. BRCA1 regulates p53-dependent gene expression. Proc Natl Acad Sci U.S.A. (1998) 95:2302–6. doi: 10.1073/pnas.95.5.2302
73. Fernández PL, Farré X, Nadal A, Fernández E, Peiró N, Sloane BF, et al. Expression of cathepsins B and S in the progression of prostate carcinoma. Int J Cancer. (2001) 95:51–5. doi: 10.1002/(ISSN)1097-0215
Keywords: cathepsins, cancers, Mendelian randomization, causality, single nucleotide polymorphisms (SNPs)
Citation: Deng T, Lu X, Jia X, Du J, Wang L, Cao B, Yang M, Yin Y and Liu F (2024) Cathepsins and cancer risk: a Mendelian randomization study. Front. Endocrinol. 15:1428433. doi: 10.3389/fendo.2024.1428433
Received: 06 May 2024; Accepted: 17 May 2024;
Published: 31 May 2024.
Edited by:
Min Tu, Nanjing Medical University, ChinaReviewed by:
Yang Wu, Nanjing Medical University, ChinaTong Zhu, LMU Munich University Hospital, Germany
Copyright © 2024 Deng, Lu, Jia, Du, Wang, Cao, Yang, Yin and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fanjie Liu, bGl1ZmoxOTgyMTFAMTI2LmNvbQ==; Ying Yin, NTYzMjk4MDk4QHFxLmNvbQ==; Meina Yang, bWVpbmE4NjEwMTBAMTYzLmNvbQ==
†These authors have contributed equally to this work and share first authorship