
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Endocrinol. , 18 January 2024
Sec. Reproduction
Volume 14 - 2023 | https://doi.org/10.3389/fendo.2023.1293780
This article is part of the Research Topic Analyzing Male Reproductive Risk, Understanding Molecular Targets, and Developing Treatments View all 10 articles
 Layla Simón*
Layla Simón* María Salomé Mariotti-Celis*
María Salomé Mariotti-Celis*About 8-12% of couples experience infertility, with male infertility being the cause in 50% of cases. Several congenital and acquired conditions, including chronic diseases and their treatments, can contribute to male infertility. Prostate cancer incidence increases annually by roughly 3%, leading to an increment in cancer treatments that have adverse effects on male fertility. To preserve male fertility post-cancer survival, conventional cancer treatments use sperm cryopreservation and hormone stimulation. However, these techniques are invasive, expensive, and unsuitable in prepubertal patients lacking mature sperm cells. Alternatively, nutritional therapies enriched with bioactive compounds are highlighted as non-invasive approaches to prevent male infertility that are easily implementable and cost-effective. In fact, curcumin and resveratrol are two examples of bioactive compounds with chemo-preventive effects at the testicular level. In this article, we summarize and discuss the literature regarding bioactive compounds and their mechanisms in preventing cancer treatment-induced male infertility. This information may lead to novel opportunities for future interventions.
Infertility is a major health problem that affects approximately 8% to 12% of couples worldwide. Male infertility stands as the primary cause in 20% to 30% of cases and contributes to infertility in an additional 20% of couples. Collectively, male factors account for 50% of infertility cases. Various congenital, acquired and idiopathic factors contribute to male infertility. In terms of acquired factors, chronic diseases and their treatments, such as cancer, chemotherapy and radiation, respectively, play a role in male infertility (1).
Cancer is a major health problem worldwide, with incidence and death rates historically higher in men than women. In fact, the probability of developing invasive cancer within lifetime was 1 in 2 men compared to 1 in 3 women in the United States between 2017-2019. Indeed, men are more exposed to carcinogenic factors, such as endogenous hormones, smoking, height, and immune response. In this sense, it is expected that around one-half of cancers in men will be concentrated in prostate (29%), lung (12%) and colorectal (8%) cancers in 2023. In the United States, prostate cancer incidence has increased by roughly 3% annually, similar to lung cancer (2%) (2). Therefore, the sustaining increase in cancer incidence leads to an increment in cancer treatments that can have adverse effects on male fertility.
Hopefully, the 5-year cancer survival rate has increased from 49% in the mid-1970s to 68% in the last decade. Moreover, prostate cancer (97%) has the highest survival rate after thyroid cancer (98%) (2), thereby increasing the number of patients surviving after cancer treatment.
Male infertility after cancer treatment is caused by: (a) decreased gonadotropin secretion from the pituitary gland caused by immune inhibitors, cranial irradiation and central nervous system tumors surgeries; (b) spermatogenic dysfunction due to chemotherapy or irradiation; (c) obstruction of seminal tracts caused by intrapelvic surgeries; (d) sexual dysfunction due to intrapelvic or retroperitoneal surgeries or irradiation. In fact, almost 46% of young cancer survivors overcome male infertility, and 30% of patients have testicular dysfunction due to chemo- or radio-therapy (3). Unfortunately, male survivors of childhood cancer have a higher desire for children compared with their siblings (25% vs. 7%) (4), thereby developing interventions that preserve male fertility is a necessity for cancer patients.
Adult cancer patients use sperm cryopreservation and hormone stimulation to preserve male fertility post-cancer treatments. However, these techniques are invasive, expensive, and unsuitable in prepubertal patients lacking mature sperm cells. Using less gonadotoxic chemo- and radio-therapies, organ-sparing surgeries and cryopreserving testicular tissue are some methods to preserve or restore fertility in prepuberal males undergoing cancer therapies. Unfortunately, these approaches are even more expensive, invasive, possible only for some patients and available in only few medical centers (5).
On the other hand, nutritional therapies enriched with bioactive compounds seem to be cost-effective, easily-implementable, and non-invasive approaches to prevent male infertility. In a metanalysis review, L-Carnitine administrated with micronutrients, antioxidants and herbal supplements increases pregnancy rates (6). In fact, antioxidants such as L-Carnitine, Coenzyme-Q10, ω3 fatty acid and selenium improves sperm quality parameters (7).
In this article, we summarize and discuss the literature concerning bioactive compounds and their mechanisms of action within preventing cancer treatment-induced male infertility. This information may contribute to develop novel opportunities for future interventions.
Male infertility is classified into four categories: (a) hypothalamic-pituitary axis disturbances, (b) spermatogenic qualitative and (c) quantitative defects, and (d) ductal obstruction or dysfunction (8). Independently of the category, about 30% of infertility cases are due to idiopathic conditions, and 70% are caused by genetic mutations or acquired conditions (9).
Genetic mutations affect almost 15% of males with infertility and 25% of men with azoospermia (no spermatozoa in the ejaculate). Some genetic alterations are chromosomal numerical or structural abnormalities, Y chromosomal deletions, azoospermia factor (AZF) deletions, androgen receptor (AR) gene mutations, cystic fibrosis transmembrane conductance regulator (CFTR) gene mutations (8).
Some acquired conditions that affect male fertility are obesity, pesticide exposure, smoking and medications (9). For instance, obesity is a health problem that course with concomitant diseases such as cardiovascular disease, type 2 diabetes and cancer. Moreover, obesity affects the hypothalamic-pituitary-gonadal axis, disrupts testicular steroidogenesis, and induces erectile dysfunction, poor semen quality and prostatitis. Some mediators of infertility-induced obesity are hyperinsulinemia, hyperleptinemia, chronic inflammation and oxidative stress (10). In the case of pesticide exposure, workers and exposed populations have deleterious semen quality (volume and sperm count, motility and morphology), DNA fragmentation and chromosome aneuploidy (11). Males with smoking habits have a high probability of being infertile, which increases with long smoking duration (>10 years) (12). Also, medications have adverse effects on reproductive achievement. For example, males receiving major antidepressant drugs have reduced conceptional rates (13).
Similarly, other environmental pollutants that have shown to affect male fertility are heavy metals, microplastics and endocrine-disrupting chemicals such as bisphenols, phthalates, and parabens. Heavy metals such as Zn, Se, Pb, and Cd are known to increase lipid peroxidation, reduce antioxidant capacity, and thereby impair sperm function (14). Also, high blood and seminal levels of Pb, Cd, Ba and U are associated with low sperm viability (15). In the case of microplastics, it has been proved in animals that microplastics impair semen quality in an equivalent human dose of 0.016 mg/kg/d, which is nearly achieved in Japan and South Korea (16). In addition, bisphenols, phthalates, and parabens increase ROS production, DNA damage and apoptosis, leading to abnormal sperm count and semen quality (17).
Testicular tumors represent almost 2% of all cancers in men. Testicular tumors are classified into two main groups: germ cell and sex cord tumors. Germ cell tumors include germ cell neoplasia in situ, seminoma, teratoma, non-seminomatous germ cell tumors, embryonal carcinoma, Yolk sac tumors, trophoblastic and regressed germ cell tumors. Sex cord stromal tumors cover Sertoli and Leydig cells tumors, and myoid gonadal stromal tumors (18). Although testicular cancer is a disease that reduces male fertility by disturbing spermatogenesis, vasculogenesis and the secretion of paracrine factors, the reduction in fertility is more a consequence of cancer treatment than the primary testicular tumor effects. In the case of surgery, 50% of patients after orchiectomy have a decrease in sperm number. Radiotherapy effects on spermatogenesis depend on doses. In fact, doses less than 1 Gy allow spermatogenesis recovery after 18 months, doses up to 3 Gy after 30 months, and up to 4 Gy after 5 years. Moreover, the chemo- and radiotherapy combination increases their gonadotoxic effects that when are administrated alone. The main adverse effect of chemotherapy on fertility is a consequence of targeting proliferating cells as germ cells (19). Spermatogonia are sensitive to the cytotoxic effects of cancer treatments, with increased susceptibility during the spermatogenesis undergoing differentiation (3). In addition, chemotherapy induces mutations in stem cell spermatogonia, thereby causing permanent damage to spermatogenesis. Some alkylating agents such as vincristine, prednisolone and cyclophosphamide induce permanent germ cell depletion and azoospermia in 80% of patients. Etoposide and doxorubicin promote azoospermia in 90% of patients (19).
In addition, other types of tumors dismiss male fertility by endocrine, nutritional, metabolic and immune alterations (19). In fact, germ cells are sensitive to the toxic effects of cancer therapy. Due to spermatogonia are more sensitive than highly differentiated germ cells, initially cancer treatment reduces the number of spermatogonia but does not affect the number of spermatocytes, spermatids and sperm cells. So, the sperm count is maintained at the beginning of the treatment but is reduced dramatically after 1 or 2 months. The azoospermia occurs after 12 weeks of cancer treatment, and in the case of low cytotoxic drugs (vinblastine, bleomycin, methotrexate, 5-fluorouracil), sperm count is recovered after 12 weeks of discontinuation chemotherapy. But, the highly cytotoxic drugs (cyclophosphamide, cisplatin, busulfan) induce more prolonged and even permanent azoospermia. Furthermore, cranial surgery or irradiation affect the hypothalamic-pituitary-gonadal axis, reduce gonadotrophin secretion and thereby spermatogenesis (3).
Treatments for male fertility are selected in function of the etiology. For instance, male hypogonadotropic hypogonadism is secondary to gonadotropin deficiency and is treated with hormone stimulation to replace the missing hormones. Therapeutic GnRH stimulates pituitary gonadotropin secretion. Gonadotropins imitate LH and FSH. Selective Oestrogen Receptor Modulators (SERMs) inhibit Oestrogen Receptors in the hypothalamus and pituitary and suppress the oestrogen-mediated negative feedback on the hypothalamic-pituitary-gonadal axis. Aromatase inhibitors inhibit the conversion of testosterone to oestradiol, thereby decreasing the oestradiol-mediated negative feedback (20). Although hormone stimulation has promising effects on animals, it is ineffective in human cancer patients (5).
In the case of male infertility due to cancer treatment in reproductive age, an alternative to preserve fertility is sperm cryopreservation followed by in vitro fertilization or intracytoplasmic sperm injection. These approaches are effective when sperm is collected for 2-3 ejaculates and before the cancer treatment. But, in some cases starting cancer treatment is an urgency that no contemplate time for sperm cryopreservation. Moreover, this option is expensive and difficult to develop in some centers (19). Furthermore, sperm cryopreservation and assisted-reproductive technology have 30% pregnancy and 25% live birth rates, respectively. Although being compared to general fertility treatment, is not effective in all the patients (3).
Cryopreserving sperm is impossible in prepuberal patients. The cryopreservation of testicular tissue obtained through a biopsy is possible in these patients (19). Moreover, cryopreserving testicular tissue is the only option in men unable to ejaculate and those with azoospermia (3). However, testicular biopsy has potential risks in leukemia patients. Furthermore, fertility restoration is still a problem in young cancer survivors for clinical reasons (19). In fact, the alternatives for using cryopreserving spermatogonia stem cells are under experimentation. For instance, testicular tissue auto- or xeno-grafting, spermatogonial stem cells transplantation and in vitro spermatogenesis are being studied in animals (5). However, culturing sperm and germ cells has been unsuccessful in humans. In addition, these approaches have some disadvantages: the small number of spermatogonia stem cells, contamination with cancer cells, surgical procedures, intracytoplasmic sperm injection and virus infection (3).
Regarding nutritional aspects, abnormal sperm parameters and hormone levels are associated with a high intake of alcohol, processed starchy and meat foods and foods rich in trans and saturated fatty acids (21–27). On the other hand, nutritional healthy habits such as high consumption of vegetables, fruits and seafood products have shown a positive association with the prevention of male infertility (28).
The Western diet is rich in saturated fatty acids, carbohydrates and proteins found in processed foods. It is also reduced in polyunsaturated fatty acids, dietary fibers and antioxidants, consequently having a negative effect on sperm quality. In this sense, the Western diet is associated with obesity, dyslipidemia, insulin resistance, oxidative stress, and aromatase activity. As a result, the Western diet reduces testosterone levels leading to a decrease in sperm count, motility and morphology. Moreover, the Western diet modifies the metabolism of sperm cells by decreasing glycolysis and mitochondrial respiration, reducing ATP content and sperm motility. Contrary, vegetarian diets are rich in plant-based antioxidants such as polyphenols with beneficial effects on sperm quality. For instance, quercetin is a flavonoid that interacts with mitochondrial membranes at the coenzyme Q-binding site, suppresses superoxide generation and promotes ATP synthesis (29). Similarly, the Mediterranean diet is characterized by a high intake of polyphenols, monounsaturated fatty acids, fiber, ω3 fatty acids, and vitamins, while being reduced in saturated and trans-fatty acids. Therefore, the Mediterranean diet is associated with a positive impact on male fertility, high sperm count and motility (28). Furthermore, the Mediterranean diet reduces oxidative stress and increases ATP generated through energetic metabolism, thus improving sperm quality (29).
As we previously described, there exists an intricate connection between nutritional habits and fertility. In vivo models have confirmed that administrating several bioactive compounds found in plant sources can help to mitigate the detrimental effects caused by unhealthy habits and environmental pollutants, thereby aiding in the recovery of fertility. Table 1 summarizes the nutritional habits known to prevent male infertility, classifying the sources and bioactive compounds that control the detrimental effects.
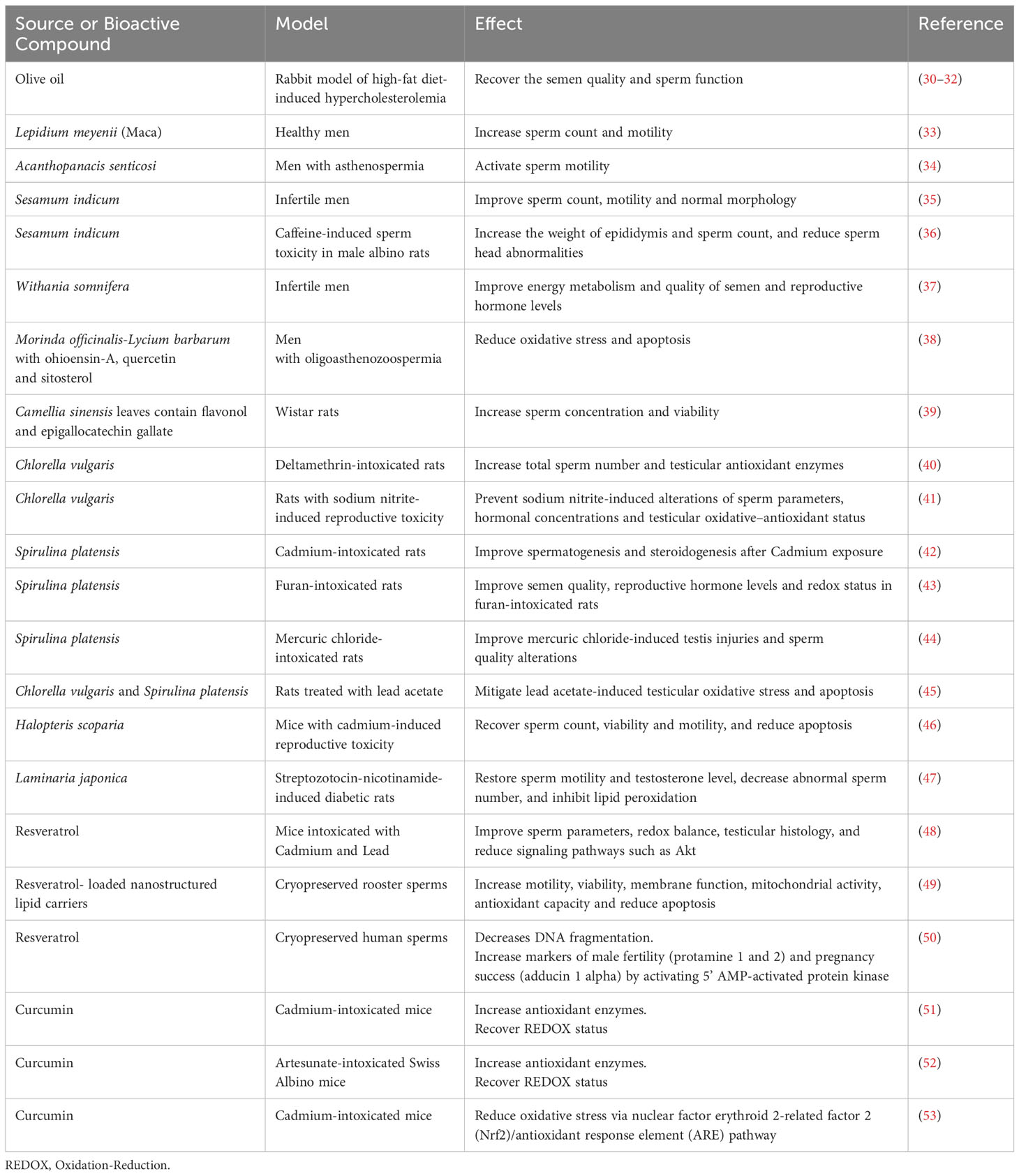
Table 1 Bioactive compounds preventing male infertility.
Mice fed with a high-fat diet develop a metabolic syndrome-like condition (increased body weight, hypercholesterolemia, hyperglycemia and glucose intolerance) associated with deleterious reproductive status. For instance, high-fat diet-fed mice have an increment in gonadal fat, associated with a reduction in epididymis weight and sperm count (54). However, switching from a high-fat diet to a normal diet recovers the fertility potential in obese male mice. In this sense, obese mice changing to a normal diet have a reduction in gonadal fat content, and an increment in FSH serum levels and fertility potential (55). In addition, the impact of a high-fat diet on semen parameters has been studied in a rabbit model of diet-induced hypercholesterolemia. In fact, a high-fat diet is associated with a reduction of semen volume, sperm count and motility, but an increment in sperm cholesterol content, lipid droplets, functional (acrosomal reaction) and morphological abnormalities. The testicular inefficiency is associated with reduced testosterone levels, decreased differentiation from spermatogonia to sperm cells, and increased apoptosis of germ cells. On the other hand, the addition of olive oil to the diet recovered the semen quality and sperm function dismissed by hypercholesterolemic diet in rabbits Table 1) (30–32).
Plants-based diets are alternative and sustainable approaches managing male infertility. Active principles and crude extracts of medicinal plants are used because of their antioxidant, anti-inflammatory, and positive effects on the testis. They have bioactive compounds such as polyphenols (anthocyanins, proanthocyanidins), phyto-oestrogens, diosgenin and thymoquinone (56). For instance, Lepidium meyenii (Maca), administrated to healthy men at 1.75 g/day for 3 months, increases sperm count and motility (33). Furthermore, Acanthopanacis senticosi activates sperm motility when administrated in humans (34). Moreover, Sesamum indicum, administrated at 0.5 mg/kg during 3 months, improves sperm count, motility and normal morphology (35, 36). In addition, roots of Withania somnifera administrated 5 g/day during 3 months improve energy metabolism and quality of semen and reproductive hormone levels in infertile men (37). Zingiber officinale (Ginger) powder or root are used because of their antioxidant, anti-inflammatory, anti-tumorigenic and androgenic activity. In fact, ginger contains gingerdiol, gingerol, shogaols,zingerone, zingibrene, folic acid, sesquiterpenes and vitamin C (57).
Morinda officinalis-Lycium barbarum coupled herbs are traditional Chinese medicines that reduce oxidative stress and apoptosis, thereby improving male fertility. These herbs contain ohioensin-A, quercetin and sitosterol that target androgen and estrogen receptors, MAPK, PI3K/Akt and glyceraldehyde-3-phosphate dehydrogenase (38). Camellia sinensis (green tea) leaves contain flavonol and epigallocatechin gallate and increase sperm concentration and viability when administrated for 52 days in rats (39, 57). Microalgae such as Chlorella vulgaris and Spirulina platensis improve spermatogenesis and steroidogenesis and protect against oxidative stress in rats (40–45). Algae such as Halopteris scorapia and Laminaria japonica increase sperm count, motility and testosterone levels, meanwhile decreasing sperm abnormalities, inflammation and oxidative stress, in mice and rats, respectively (46, 47, 57).
Vitis vinífera (grape) contains resveratrol and flavonoids (catechin, quercetin, anthocyanin and pro-anthocyanidins) with protective effects on testicles. In this sense, grape seed extracts reduce oxidative stress and apoptosis, meanwhile improve testicular histology, hormone levels, and sperm count and morphology (57). In mice, resveratrol reduces the toxic effects of cadmium and lead at the testicular level. Moreover, resveratrol prevents the development of testicular germ cell neoplasia in situ promoted by heavy metals. In this sense, resveratrol improves sperm parameters, redox balance, testicular histology, and reduces signaling pathways such as Akt (48). Moreover, resveratrol supplementation in a cryopreservation medium improves the post-thawed sperm quality and fertility of roosters. Indeed, rooster semen cryopreserved with 40 µM resveratrol-loaded nanostructured lipid carriers has higher motility, viability, membrane function, mitochondrial activity, antioxidant capacity and lower apoptosis than non-treated frozen sperm (49). In human cryopreserved semen samples, 15 µM resveratrol decreases DNA fragmentation mean increases markers of male fertility (protamine 1 and 2) and pregnancy success (adducin 1 alpha) by activating 5’ AMP-activated protein kinase (50).
Antioxidants lead the list of natural products that are protective agents for male infertility. In this regard, oxidative stress damages sperm membranes and DNA, thereby promoting infertility. Curcumin is a bioactive compound present in the turmeric plant Curcuma longa that reduce oxidative stress, lipid peroxidation and oxidative DNA damage. Curcumin increases the levels of GSH, glutathione peroxidase, superoxide dismutase and catalase. In addition, curcumin increases testosterone, FSH and LH levels in mice (51–53). Ellagic acid is a polyphenol with similar effects to curcumin. Vitamin C protects spermatogenesis, prevents sperm agglutination, and increases testosterone, FSH and LH levels. Moreover, vitamin C induces antioxidant enzymes, and reduces LDL cholesterol and lipid peroxidation. Vitamin E is an antioxidant that protects cell membrane components from oxidative damage. At reproductive level, vitamin E also protects spermatogenesis and testosterone levels (57).
Because of their availability, safety and low cost, bioactive compounds contained in fruits, vegetables and spices are potential agents for the prevention and treatment of cancer. Even though they have limitations (low bioavailability, high metabolism, poor water solubility), these molecules have antitumorigenic effects against a wide range of cancers: colon, lung, prostate, breast, gastric, liver, and brain cancer (58). Moreover, contrary to current cancer treatments, bioactive compounds are potential chemo-preventive and -therapeutic agents with low side effects on the health of patients (59, 60).
Bioactive compounds target cancer cells, macrophages and adipocytes in the tumor microenvironment, thereby preventing cancer development and progression (61). In this way, curcumin, myricetin, geraniin, genistein, tocotrienol, fucoxanthin, anthocyanin, epigallocatechin gallate, gallic and ellagic acids have anti-proliferative, pro-apoptotic and anti-metastatic effects. For instance, curcumin, in nanoparticles, piperine, phospholipid complexes and liposomes, inhibits PI3K/Akt and NF-κB pathways, but upregulates p53 and Bax expression, thereby promoting apoptosis of cancer cells. In addition, curcumin downregulates MMP-9 expression and reduce the metastatic potential of cancer cells (59). Furthermore, Tripterygium wilfordii used in the Chinese medicine contains bioactive compounds such as triptolide, celastrol and tripchlorolide that also inhibit PI3K/Akt and NF-κB pathways thereby exerting anticancer and anti-inflammatory effects (62).
As previously mentioned, several plant-derived bioactive compounds are effective in improving male infertility. In addition, these compounds have been shown to reverse cancer-induced infertility. Table 2 summarizes the sources and bioactive compounds known to prevent cancer-associated male infertility by controlling the detrimental effects of cancer therapy.
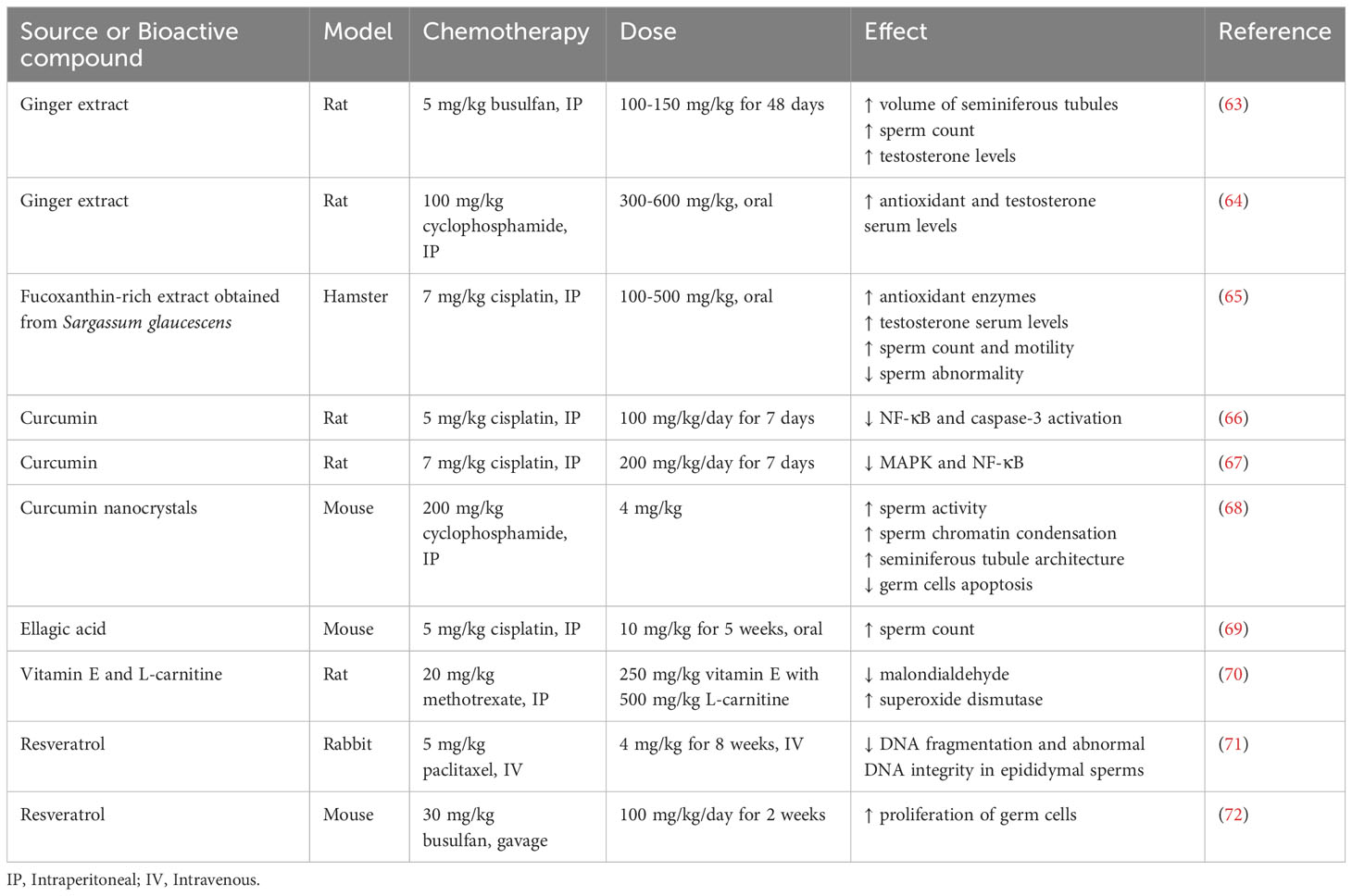
Table 2 Bioactive compounds preventing cancer therapy-induced male infertility.
For instance, ginger and algae extracts have beneficial reproductive effects post cancer-therapy (57). Rats treated with a single intraperitoneal (IP) injection of 5mg/kg busulfan (chronic myelogenous leukemia treatment) and with 100-150 mg/kg ginger extract for 48 days have increased volume of seminiferous tubules, sperm count and testosterone levels, previously impair by busulfan (Table 2) (63). Another group demonstrated that oral administration of 300-600 mg/kg ginger extract recovers the epithelium thickness and germ cell count of rat testis affected after a single IP dose of 100 mg/kg cyclophosphamide treatment by increasing antioxidant and testosterone serum levels (64). In addition, oral administration of fucoxanthin-rich brown algae Sargassum glaucescens extract ameliorates cisplatin-induced testicular damage in hamsters. Hamsters were intraperitoneal injected with 7 mg/kg cisplatin and treated with 100, 200 and 500 mg/kg fucoxanthin extract orally administrated. In fact, fucoxanthin-enriched extract recovers the testosterone level, seminiferous tubules morphology and sperm number, motility and morphology affected by cisplatin treatment (65).
Additionally, several bioactive compounds such as curcumin, ellagic acid and vitamin E are involved in reverting cancer-induced male infertility (57). For instance, rats treated with a single IP dose of 5 mg/kg cisplatin and 100 mg/kg/day curcumin during 7 days reverse testicular damage by reducing NF-κB and caspase-3 activated by cisplatin (66). Rats treated with a single IP dose of 7 mg/kg cisplatin and orally administrated 200 mg/kg/day curcumin for 10 days recover oxidative stress and testicular damage induced by cisplatin via mitogen-activated protein kinase and nuclear factor-kappa B signaling pathways (67). Mice treated with 4 mg/kg curcumin nanocrystals recover the negative effects of cyclophosphamide in sperm activity, sperm chromatin condensation, and seminiferous tubule architecture, meanwhile have a reduction in germ cells apoptosis induced by the treatment with 200 mg/kg cyclophosphamide (IP) (68). Mice with colon cancer treated with 5 mg/kg cisplatin (IP) have a reduction in sperm count which is recovered after 5 weeks of treatment with 10 mg/kg ellagic acid (oral) (69). Rats receiving 250 mg/kg vitamin E with 500 mg/kg L-carnitine control the oxidative stress induced by the treatment with 20 mg/kg methotrexate (IP) via reducing malondialdehyde and increasing superoxide dismutase (70). In rabbits, the treatment with 4 mg/kg resveratrol (intravenous) for 8 weeks ameliorates the DNA fragmentation and abnormal DNA integrity in epididymal sperms induced by 5 mg/kg paclitaxel (71). In addition, 20 µM resveratrol induces the proliferation of spermatogonia stem cells in vitro. In mice, 100 mg/kg/day resveratrol for 2 weeks promotes the proliferation of germ cells thereby reversing the loss of spermatogenic cells in the testis and sperm cells in the epididymis induced by 30 mg/kg busulfan treatment (gavage) (72).
Cancer incidence is increasing worldwide and men are more affected than women. Current cancer treatments involve surgery, chemotherapy and radiation, which have side-effects such as infertility. Moreover, male infertility is responsible for 50% of couples with fertility problems. Adult cancer patients are subjected to invasive and expensive techniques to recover fertility after cancer treatment such as cryopreservation of sperm cells and in vitro fertilization (Figure 1). However, prepuberal patients without sperm cells are unable to access to these techniques. In this age group, cryopreserving testicular tissue to culture germ and sperm cells and do intracytoplasmic injection seems to be another invasive, expensive technique that is unsuccessful in humans. In this way, looking for alternative treatments for cancer and male infertility is a need to alleviate patient suffering post-cancer survival. Natural products rich in bioactive compounds are increasing interest in this scenario as alternative, novel, non-invasive agents to prevent and treat cancer preserving male fertility. Curcumin, ellagic acid, and resveratrol seem to be potential compounds that recover testicular function by increasing proliferation, reducing oxidative stress, inflammation, and apoptosis of germ cells. However, further research regarding bioavailability, solubility and metabolism of these natural compounds must be taken in consideration to improve the current therapy approaches.
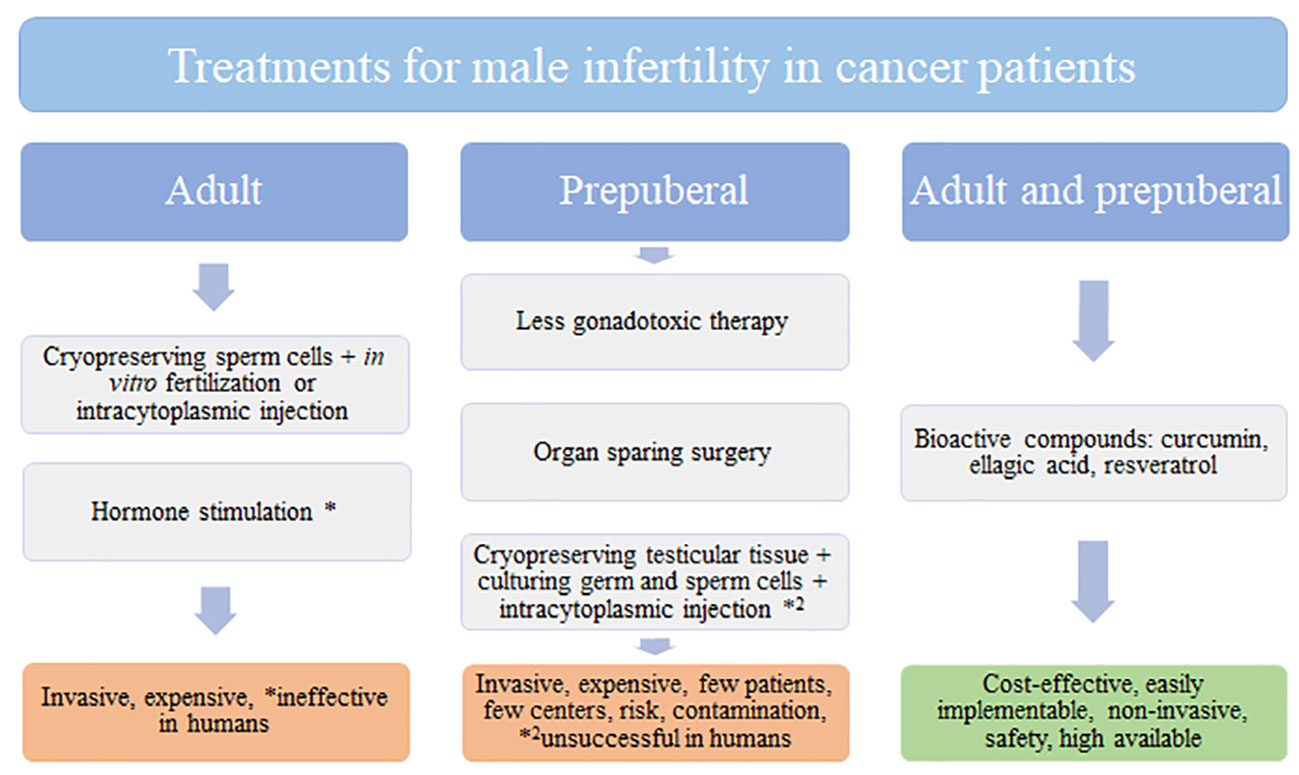
Figure 1 Treatments for male infertility in cancer patients.
LS: Conceptualization, Funding acquisition, Writing – original draft, Writing – review & editing. MM-C: Funding acquisition, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by ANID FONDECYT iniciación 11230112 (LS), Concurso de Investigación con Colaboración Internacional de la Universidad Finis Terrae (LS), ANID FONDECYT regular 1220097 (MM-C).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Agarwal A, Baskaran S, Parekh N, Cho CL, Henkel R, Vij S, et al. Male infertility. Lancet (2021) 397(10271):319–33. doi: 10.1016/S0140-6736(20)32667-2
2. Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin (2023) 73(1):17–48. doi: 10.3322/caac.21763
3. Yumura Y, Takeshima T, Komeya M, Karibe J, Kuroda S, Saito T. Long-term fertility function sequelae in young male cancer survivors. World J Mens Health (2023) 41(2):255–71. doi: 10.5534/wjmh.220102
4. Claessens JJM, Penson A, Bronkhorst EM, Kremer LCM, van Dulmen-den Broeder E, van der Heiden-van der Loo M, et al. Desire for children among male survivors of childhood cancer: A DCCSS LATER study. Cancer. (2023) 129(9):1432–42. doi: 10.1002/cncr.34685
5. Eugeni E, Arato I, Del Sordo R, Sidoni A, Garolla A, Ferlin A, et al. Fertility preservation and restoration options for pre-pubertal male cancer patients: current approaches. Front Endocrinol (Lausanne). (2022) 13:877537. doi: 10.3389/fendo.2022.877537
6. Zafar MI, Mills KE, Baird CD, Jiang H, Li H. Effectiveness of nutritional therapies in male factor infertility treatment: A systematic review and network meta-analysis. Drugs. (2023) 83(6):531–46. doi: 10.1007/s40265-023-01853-0
7. Li KP, Yang XS, Wu T. The effect of antioxidants on sperm quality parameters and pregnancy rates for idiopathic male infertility: A network meta-analysis of randomized controlled trials. Front Endocrinol (Lausanne). (2022) 13:810242. doi: 10.3389/fendo.2022.810242
8. Krausz C, Riera-Escamilla A. Genetics of male infertility. Nat Rev Urol. (2018) 15(6):369–84. doi: 10.1038/s41585-018-0003-3
9. Fainberg J, Kashanian JA. Recent advances in understanding and managing male infertility. F1000Res. (2019) 8:2019. doi: 10.12688/f1000research.17076.1
10. Leisegang K, Sengupta P, Agarwal A, Henkel R. Obesity and male infertility: Mechanisms and management. Andrologia. (2021) 53(1):e13617. doi: 10.1111/and.13617
11. Giulioni C, Maurizi V, Castellani D, Scarcella S, Skrami E, Balercia G, et al. The environmental and occupational influence of pesticides on male fertility: A systematic review of human studies. Andrology. (2022) 10(7):1250–71. doi: 10.1111/andr.13228
12. Yang F, Li L, Chen JP, Liu XQ, Zhong CL, Yang Y, et al. Couple′s infertility in relation to male smoking in a Chinese rural area. Asian J Androl. (2017) 19(3):311. doi: 10.4103/1008-682X.168685
13. Evans-Hoeker EA, Eisenberg E, Diamond MP, Legro RS, Alvero R, Coutifaris C, et al. Major depression, antidepressant use, and male and female fertility. Fertil Steril. (2018) 109(5):879–87. doi: 10.1016/j.fertnstert.2018.01.029
14. Chinyere Nsonwu-Anyanwu A, Raymond Ekong E, Jeremiah Offor S, Francis Awusha O, Chukwuma Orji O, Idiongo Umoh E, et al. Heavy metals, biomarkers of oxidative stress and changes in sperm function: A case-control study. Int J Reprod Biomed (2019) 17(3):163–74. doi: 10.18502/ijrm.v17i3.4515
15. Sukhn C, Awwad J, Ghantous A, Zaatari G. Associations of semen quality with non-essential heavy metals in blood and seminal fluid: data from the Environment and Male Infertility (EMI) study in Lebanon. J Assist Reprod Genet (2018) 35(9):1691–701. doi: 10.1007/s10815-018-1236-z
16. Zhang C, Chen J, Ma S, Sun Z, Wang Z. Microplastics may be a significant cause of male infertility. Am J Mens Health (2022) 16(3):15579883221096548. doi: 10.1177/15579883221096549
17. Virant-Klun I, Imamovic-Kumalic S, Pinter B. From oxidative stress to male infertility: review of the associations of endocrine-disrupting chemicals (Bisphenols, phthalates, and parabens) with human semen quality. Antioxidants (Basel). (2022) 11(8):1691–1701. doi: 10.3390/antiox11081617
18. Dashora A, Wagner T, Berney DM. Testicular tumors: new developments in germ cell and sex cord stromal tumors. Surg Pathol Clin (2022) 15(4):729–43. doi: 10.1016/j.path.2022.07.007
19. Vakalopoulos I, Dimou P, Anagnostou I, Zeginiadou T. Impact of cancer and cancer treatment on male fertility. Hormones. (2015) 14(4):579–589. doi: 10.14310/horm.2002.1620
20. Foran D, Chen R, Jayasena CN, Minhas S, Tharakan T. The use of hormone stimulation in male infertility. Curr Opin Pharmacol (2023) 68:102333. doi: 10.1016/j.coph.2022.102333
21. Hansen ML, Thulstrup AM, Bonde JP, Olsen J, Håkonsen LB, Ramlau-Hansen CH. Does last week’s alcohol intake affect semen quality or reproductive hormones? A cross-sectional study among healthy young Danish men. Reprod Toxicol (2012) 34(3):457–62. doi: 10.1016/j.reprotox.2012.06.004
22. Anifandis G, Bounartzi T, Messini CI, Dafopoulos K, Sotiriou S, Messinis IE. The impact of cigarette smoking and alcohol consumption on sperm parameters and sperm DNA fragmentation (SDF) measured by Halosperm(®). Arch Gynecol Obstet. (2014) 290(4):777–82. doi: 10.1007/s00404-014-3281-x
23. Cutillas-Tolín A, Mínguez-Alarcón L, Mendiola J, López-Espín JJ, Jørgensen N, Navarrete-Muñoz EM, et al. Mediterranean and western dietary patterns are related to markers of testicular function among healthy men. Hum Reprod (2015) 30(12):2945–55. doi: 10.1093/humrep/dev236
24. Afeiche MC, Williams PL, Gaskins AJ, Mendiola J, Jørgensen N, Swan SH, et al. Meat intake and reproductive parameters among young men. Epidemiology. (2014) 25(3):323–30. doi: 10.1097/EDE.0000000000000092
25. Eslamian G, Amirjannati N, Rashidkhani B, Sadeghi MR, Baghestani AR, Hekmatdoost A. Dietary fatty acid intakes and asthenozoospermia: a case-control study. Fertil Steril. (2015) 103(1):190–8. doi: 10.1016/j.fertnstert.2014.10.010
26. Afeiche MC, Gaskins AJ, Williams PL, Toth TL, Wright DL, Tanrikut C, et al. Processed meat intake is unfavorably and fish intake favorably associated with semen quality indicators among men attending a fertility clinic. J Nutr (2014) 144(7):1091–8. doi: 10.3945/jn.113.190173
27. Jensen TK, Heitmann BL, Blomberg Jensen M, Halldorsson TI, Andersson AM, Skakkebæk NE, et al. High dietary intake of saturated fat is associated with reduced semen quality among 701 young Danish men from the general population. Am J Clin Nutr (2013) 97(2):411–8. doi: 10.3945/ajcn.112.042432
28. Salas-Huetos A, Bulló M, Salas-Salvadó J. Dietary patterns, foods and nutrients in male fertility parameters and fecundability: a systematic review of observational studies. Hum Reprod Update. (2017) 23(4):371–89. doi: 10.1093/humupd/dmx006
29. Ferramosca A, Zara V. Diet and male fertility: the impact of nutrients and antioxidants on sperm energetic metabolism. Int J Mol Sci (2022) 23(5):2542–2558. doi: 10.3390/ijms23052542
30. Saez Lancellotti TE, Boarelli PV, Romero AA, Funes AK, Cid-Barria M, Cabrillana ME, et al. Semen quality and sperm function loss by hypercholesterolemic diet was recovered by addition of olive oil to diet in rabbit. PloS One (2013) 8(1):e52386. doi: 10.1371/journal.pone.0052386
31. Simón L, Funes AK, Monclús MA, Colombo R, Cabrillana ME, Saez Lancellotti TE, et al. Manchette-acrosome disorders and testicular efficiency decline observed in hypercholesterolemic rabbits are recovered with olive oil enriched diet. PloS One (2018) 13(8):e0202748. doi: 10.1371/journal.pone.0202748
32. Funes AK, Avena MV, Ibañez J, Simón L, Ituarte L, Colombo R, et al. Extra-virgin olive oil ameliorates high-fat diet-induced seminal and testicular disorders by modulating the cholesterol pathway. Andrology. (2023) 11(6):1203–17. doi: 10.1111/andr.13398
33. Melnikovova I, Fait T, Kolarova M, Fernandez EC, Milella L. Effect of lepidium meyenii walp. on semen parameters and serum hormone levels in healthy adult men: A double-blind, randomized, placebo-controlled pilot study. Evidence-Based Complementary Altern Med (2015) 2015:1–6. doi: 10.1155/2015/324369
34. Wu W, hong LJ, Ping YC, Chua Z. [A comparative study of the effects of Acanthopanacis senticosi injection, theophylline and caffeine on human sperm mobility in vitro]. Zhonghua Nan Ke Xue. (2009) 15(3):278–81.
35. Khani B, Bidgoli SR, Moattar F, Hassani H. Effect of sesame on sperm quality of infertile men. J Res Med Sci (2013) 18(3):184–7.
36. Uno UUU, Ndifon TB, Esua IS, Ekaluo UB. Ameliorating potential of sesame (Sesamum indicum L.) on caffeine induced sperm toxicity in male albino rats. Asian J Biol Sci (2019) 12:204–9. doi: 10.3923/ajbs.2019.204.209
37. Gupta A, Mahdi AA, Shukla KK, Ahmad MK, Bansal N, Sankhwar P, et al. Efficacy of Withania somnifera on seminal plasma metabolites of infertile males: A proton NMR study at 800MHz. J Ethnopharmacol (2013) 149(1):208–14. doi: 10.1016/j.jep.2013.06.024
38. Bai X, Tang Y, Li Q, Chen Y, Liu D, Liu G, et al. Network pharmacology integrated molecular docking reveals the bioactive components and potential targets of Morinda officinalis–Lycium barbarum coupled-herbs against oligoasthenozoospermia. Sci Rep (2021) 11(1):2220. doi: 10.1038/s41598-020-80780-6
39. Opuwari C, Monsees T. Green tea consumption increases sperm concentration and viability in male rats and is safe for reproductive, liver and kidney health. Sci Rep (2020) 10(1):15269. doi: 10.1038/s41598-020-72319-6
40. Osama E, Galal AAA, Abdalla H, El-Sheikh SMA. Chlorella vulgaris ameliorates testicular toxicity induced by deltamethrin in male rats via modulating oxidative stress. Andrologia. (2019) 51(3):e13214. doi: 10.1111/and.13214
41. Eissa MM, Ahmed MM, Abd Eldaim MA, Orabi SH, Elbaz HT, Mohamed MA, et al. Methanolic extract of Chlorella vulgaris protects against sodium nitrite-induced reproductive toxicity in male rats. Andrologia (2020) 52(11):e13811. doi: 10.1111/and.13811
42. Farag MR, Abd EL-Aziz RM, Ali HA, Ahmed SA. Evaluating the ameliorative efficacy of Spirulina platensis on spermatogenesis and steroidogenesis in cadmium-intoxicated rats. Environ Sci pollut Res (2016) 23(3):2454–66. doi: 10.1007/s11356-015-5314-9
43. Abd El-Hakim YM, Mohamed WA, El-Metwally AE. Spirulina platensis attenuates furan reprotoxicity by regulating oxidative stress, inflammation, and apoptosis in testis of rats. Ecotoxicol Environ Saf. (2018) 161:25–33. doi: 10.1016/j.ecoenv.2018.05.073
44. El-Desoky GE, Bashandy SA, Alhazza IM, Al-Othman ZA, Aboul-Soud MAM, Yusuf K. Improvement of mercuric chloride-induced testis injuries and sperm quality deteriorations by spirulina platensis in rats. PloS One (2013) 8(3):e59177. doi: 10.1371/journal.pone.0059177
45. Ibrahim IA, Shalaby AA, Abd Elaziz RT, Bahr HI. Chlorella vulgaris or Spirulina platensis mitigate lead acetate-induced testicular oxidative stress and apoptosis with regard to androgen receptor expression in rats. Environ Sci pollut Res (2021) 28(29):39126–38. doi: 10.1007/s11356-021-13411-w
46. Güner Ö, Güner A, Yavaşoğlu A, Karabay Yavaşoğlu NÜ, Kavlak O. Ameliorative effect of edible Halopteris scoparia against cadmium-induced reproductive toxicity in male mice: A biochemical and histopathologic study. Andrologia. (2020) 52(6):e13591. doi: 10.1111/and.13591
47. Kong ZL, Sudirman S, Hsu YC, Su CY, Kuo HP. Fucoxanthin-rich brown algae extract improves male reproductive function on streptozotocin-nicotinamide-induced diabetic rat model. Int J Mol Sci (2019) 20(18):4485. doi: 10.3390/ijms20184485
48. Mitra S, Patra T, Saha D, Ghosh P, Mustafi SM, Varghese AC, et al. Sub-chronic cadmium and lead compound exposure induces reproductive toxicity and development of testicular germ cell neoplasia in situ in murine model: Attenuative effects of resveratrol. J Biochem Mol Toxicol (2022) 36(7):e23058. doi: 10.1002/jbt.23058
49. Najafi A, Daghigh Kia H, Hamishehkar H, Moghaddam G, Alijani S. Effect of resveratrol-loaded nanostructured lipid carriers supplementation in cryopreservation medium on post-thawed sperm quality and fertility of roosters. Anim Reprod Sci (2019) 201:32–40. doi: 10.1016/j.anireprosci.2018.12.006
50. Shabani Nashtaei M, Nekoonam S, Naji M, Bakhshalizadeh S, Amidi F. Cryoprotective effect of resveratrol on DNA damage and crucial human sperm messenger RNAs, possibly through 5’ AMP-activated protein kinase activation. Cell Tissue Bank. (2018) 19(1):87–95. doi: 10.1007/s10561-017-9642-5
51. Momeni H, Eskandari N. Curcumin protects the testis against cadmium-induced histopathological damages and oxidative stress in mice. Hum Exp Toxicol (2020) 39(5):653–61. doi: 10.1177/0960327119895564
52. Desai KR, Rajput DK, Patel PB, Highland HN. Ameliorative effects of curcumin on artesunate-induced subchronic toxicity in testis of swiss albino male mice. Dose-Response. (2015) 13(2):155932581559239. doi: 10.1177/1559325815592393
53. Yang SH, He JB, Yu LH, Li L, Long M, Liu MD, et al. Protective role of curcumin in cadmium-induced testicular injury in mice by attenuating oxidative stress via Nrf2/ARE pathway. Environ Sci pollut Res (2019) 26(33):34575–83. doi: 10.1007/s11356-019-06587-9
54. Gómez-Elías MD, Rainero Cáceres TS, Giaccagli MM, Guazzone VA, Dalton GN, De Siervi A, et al. Association between high-fat diet feeding and male fertility in high reproductive performance mice. Sci Rep (2019) 9(1):18546. doi: 10.1038/s41598-019-54799-3
55. Zhang W, Tian Z, Qi X, Chen P, Yang Q, Guan Q, et al. Switching from high-fat diet to normal diet ameliorate BTB integrity and improve fertility potential in obese male mice. Sci Rep (2023) 13(1):14152. doi: 10.1038/s41598-023-41291-2
56. Abarikwu SO, Onuah CL, Singh SK. Plants in the management of male infertility. Andrologia (2020) 52(3):e13509. doi: 10.1111/and.13509
57. Martins RVL, Silva AMS, Duarte AP, Socorro S, Correia S, Maia CJ. Natural products as protective agents for male fertility. BioChem. (2021) 1(3):122–47. doi: 10.3390/biochem1030011
58. Haque A, Brazeau D, Amin AR. Perspectives on natural compounds in chemoprevention and treatment of cancer: an update with new promising compounds. Eur J Cancer. (2021) 149:165–83. doi: 10.1016/j.ejca.2021.03.009
59. Subramaniam S, Selvaduray KR, Radhakrishnan AK. Bioactive compounds: natural defense against cancer? Biomolecules. (2019) 9(12):758. doi: 10.3390/biom9120758
60. Simón L, Arazo-Rusindo M, Quest AFG, Mariotti-Celis MS. Phlorotannins: novel orally administrated bioactive compounds that induce mitochondrial dysfunction and oxidative stress in cancer. Antioxidants. (2023) 12(9):1734. doi: 10.3390/antiox12091734
61. Sanhueza S, Simón L, Cifuentes M, Quest AFG. The adipocyte–macrophage relationship in cancer: A potential target for antioxidant therapy. Antioxidants. (2023) 12(1):126. doi: 10.3390/antiox12010126
62. Wong KF, Yuan Y, Luk JM. Tripterygium wilfordii bioactive compounds as anticancer and anti-inflammatory agents. Clin Exp Pharmacol Physiol (2012) 39(3):311–20. doi: 10.1111/j.1440-1681.2011.05586.x
63. Bordbar H, Esmaeilpour T, Dehghani F, Panjehshahin MR. Stereological study of the effect of ginger’s alcoholic extract on the testis in busulfan-induced infertility in rats. Iran J Reprod Med (2013) 11(6):467–72.
64. Mohammadi F, Nikzad H, Taghizadeh M, Taherian A, Azami-Tameh A, Hosseini SM, et al. Protective effect of Zingiber officinale extract on rat testis after cyclophosphamide treatment. Andrologia. (2014) 46(6):680–6. doi: 10.1111/and.12135
65. Wang PT, Sudirman S, Hsieh MC, Hu JY, Kong ZL. Oral supplementation of fucoxanthin-rich brown algae extract ameliorates cisplatin-induced testicular damage in hamsters. Biomedicine Pharmacotherapy. (2020) 125:109992. doi: 10.1016/j.biopha.2020.109992
66. Mercantepe T, Unal D, Tümkaya L, Yazici Z. Protective effects of amifostine, curcumin and caffeic acid phenethyl ester against cisplatin-induced testis tissue damage in rats. Exp Ther Med (2018) 15(4):3404–3412. doi: 10.3892/etm.2018.5819
67. Ilbey YO, Ozbek E, Cekmen M, Simsek A, Otunctemur A, Somay A. Protective effect of curcumin in cisplatin-induced oxidative injury in rat testis: mitogen-activated protein kinase and nuclear factor-kappa B signaling pathways. Hum Reproduction. (2009) 24(7):1717–25. doi: 10.1093/humrep/dep058
68. Poojary KK, Nayak G, Vasani A, Kumari S, Dcunha R, Kunhiraman JP, et al. Curcumin nanocrystals attenuate cyclophosphamide-induced testicular toxicity in mice. Toxicol Appl Pharmacol (2021) 433:115772. doi: 10.1016/j.taap.2021.115772
69. Goyal Y, Koul A, Ranawat P. Ellagic acid ameliorates cisplatin toxicity in chemically induced colon carcinogenesis. Mol Cell Biochem (2019) 453(1–2):205–15. doi: 10.1007/s11010-018-3446-1
70. Yüncü M, Bükücü N, Bayat N, Sencar L, Tarakçioğlu M. The effect of vitamin E and L-carnitine against methotrexate-induced injury in rat testis. Turk J Med Sci (2015) 45:517–25. doi: 10.3906/sag-1409-39
71. Ili P, Sari F, Bucak MN, Öztürk C, Güngör Ş, Ataman MB. DNA damaging effect of paclitaxel in the epididymal sperms as a chemotherapeutic agent and possible remedies to prevent this effect: A study on reproductive potential of male cancer patients of reproductive age. Theriogenology. (2019) 132:201–11. doi: 10.1016/j.theriogenology.2019.04.015
Keywords: fertility, testicular, chemotherapy, radiation, tumor, resveratrol, curcumin
Citation: Simón L and Mariotti-Celis MS (2024) Bioactive compounds as potential alternative treatments to prevent cancer therapy-induced male infertility. Front. Endocrinol. 14:1293780. doi: 10.3389/fendo.2023.1293780
Received: 13 September 2023; Accepted: 29 December 2023;
Published: 18 January 2024.
Edited by:
C Yan Cheng, St. Francis College, United StatesReviewed by:
Wei Zhang, Marine Bioproducts Cooperative Research Centre, AustraliaCopyright © 2024 Simón and Mariotti-Celis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Layla Simón, bHNpbW9uQHVmdC5jbA==; María Salomé Mariotti-Celis, bW1hcmlvdHRpQHVmdC5jbA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.