 Yu-Qian Shi
Yu-Qian Shi Xi-Ting Zhu
Xi-Ting Zhu Su-Na Zhang
Su-Na Zhang Yi-Fu Ma
Yi-Fu Ma Yan-Hua Han2
Yan-Hua Han2 Yue-Hui Zhang
Yue-Hui Zhang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Endocrinol. , 01 August 2023
Sec. Reproduction
Volume 14 - 2023 | https://doi.org/10.3389/fendo.2023.1172481
This article is part of the Research Topic Aging and the Oocyte View all 6 articles
Normal levels of reactive oxygen species (ROS) play an important role in regulating follicular growth, angiogenesis and sex hormone synthesis in ovarian tissue. When the balance between ROS and antioxidants is disrupted, however, it can cause serious consequences of oxidative stress (OS), and the quantity and quality of oocytes will decline. Therefore, this review discusses the interrelationship between OS and premature ovarian insufficiency (POI), the potential mechanisms and the methods by which antioxidants can improve POI through controlling the level of OS. We found that OS can mediate changes in genetic materials, signal pathways, transcription factors and ovarian microenvironment, resulting in abnormal apoptosis of ovarian granulosa cells (GCs) and abnormal meiosis as well as decreased mitochondrial Deoxyribonucleic Acid(mtDNA) and other changes, thus accelerating the process of ovarian aging. However, antioxidants, mesenchymal stem cells (MSCs), biological enzymes and other antioxidants can delay the disease process of POI by reducing the ROS level in vivo.
POI is an important medical issue concerning women of reproductive age. It is a clinical syndrome defined by loss of ovarian activity before the age of 40 years. Over the years, several terms have been used to describe this condition, such as ‘primary ovarian insufficiency’, ‘premature menopause’ and ‘premature ovarian failure’. Although the latest guideline from the European Society of Human Reproduction and Embryology (ESHRE) adopted ‘premature ovarian insufficiency’ as the standard nomenclature (1). The diagnostic criteria are not fully standardized, yet the differences varied little. The latest diagnostic criteria proposed in the latest guidelines are primary or secondary amenorrhea or spaniomenorrhea of>4 months with onset before 40 year of age, and elevated follicle-stimulating hormone (FSH)>25IU/L on 2 assays at>4 weeks’ interval. Estradiol level is low, and anti-Müllerian hormone (AMH) levels have usually collapsed (2). The prevalence of POI varies among ethnic groups across the world, and studies show a prevalence that ranges from 1% (3) up to 5.5% (4). If not treated quickly, women living with POI face the short- and long-term consequences of prolonged hypoestrogenism and can develop symptoms such as lack of libido, vaginal dryness, mood disorders, rise in cardiovascular risk and cognitive impairment (5). Decreased mineral density promotes osteopenia and osteoporosis, as well as bone fractures. The health provider should be aware of POI signs and symptoms in order to promote prompt diagnosis (6). The most accepted ovarian aging theory emphasizes that the ability of ovarian GCs to counteract the ROS pathogenic effect is decreased (7), which ultimately leads to the reduction of the quantity and quality of follicles (8, 9). Reactive oxygen species (ROS) are two electron reduction products of oxygen, including superoxide anion, hydrogen peroxide, hydroxyl radical, lipid peroxides, protein peroxides and peroxides formed in nucleic acids (10). They are maintained in a dynamic balance by a series of reduction-oxidation (redox) reactions in biological systems and act as signaling molecules to drive cellular regulatory pathways (11, 12). The production of ROS can be induced by various factors such as heavy metals, tobacco, smoke, drugs, xenobiotics, pollutants and radiation (13). Excessive oxidative stress derived from ROS accumulation deregulates the antioxidative defense system, which is closely associated with various diseases (14, 15). ROS are produced in almost every subcellular organelle in the cell, including the plasma membrane, cytoplasm, mitochondria, core, endoplasmic reticulum, Golgi, and others (16). The main sources of cellular ROS are inflammation and mitochondrial dysfunction, as ROS are produced by immune cells in response to infection, injury and aerobic metabolism (17). In ovarian tissue, normal levels of ROS play an important regulatory role in follicular growth, intrathecal angiogenesis and sex hormone synthesis. Nevertheless, when oocytes are exposed to high concentrations of oxygen molecules or chemical derivatives of oxygen, they are susceptible to cellular damage, which may be manifested by impaired follicular oocyte development, POI and decreased reproductive function (18). The development of POI is associated with excessive accumulation of ROS in the ovary, which accelerates the aging of ovarian cells and reduces ovarian function (19). Studies have shown that perfluorononanoic acid (PFNA) can induce OS in oocytes, destroy spindle assembly, mitochondrial function and DNA structure, and induce apoptosis and hinder oocyte maturation in vitro (20). In addition, high levels of OS-mediated ROS were also found to block GCs development and induce apoptosis in the hydrogen peroxide(H2O2)–treated POI mouse model, ultimately leading to follicular atresia (21, 22). As a product of OS, the pathological effect of ROS has been confirmed in many female reproductive diseases such as endometriosis, polycystic ovary syndrome, spontaneous abortion, hydatidiform mole and pre eclampsia (23). Therefore, it is of great significance to study the role of ROS, a product of OS, in the pathogenesis of POI for exploring new OS-targeting drugs.
Recently, the second-generation sequencing has been used in large POI families to determine that the pathogenic factors mainly focus on DNA damage repair, homologous recombination, and meiosis (24). In this part, therefore, we discuss the role of OS-mediated DNA damage and alterations in genetic material including apoptosis, mtDNA dysfunction, meiotic abnormalities, and telomere shortening in the pathogenesis of POI. Environmental factors seem to be the major determinants in ovarian reserve or premature menopause during the prenatal period or adult life (25). Perfluoroalkyl and polyfluoroalkyl substances (PFASs) are persistent synthetic chemicals that are widely used in industrial applications and often detectable in humans (26). Perfluorohexanesulfonate (PFHxS), perfluorononanoate (PFNA), perfluorooctanoate (PFOA), and perfluorooctanesulfonate (PFOS) are used as stain, water, and grease repellents in a wide range of consumer products. Human exposure to PFAS mainly occurs orally via the intake of contaminated food, water, and dust. A population-based study (27)showed that high exposure to PFOA, PFOS and PFHxS was linked to an increased risk of POI in humans. Jiao et al. (20)found that PFNA can induce OS, destroy spindle assembly, disrupt mitochondrial function as well as DNA damage, and cause oocyte apoptosis to interfere with oocyte maturation in vitro. Animal experiments have shown that exposure of mice to PFOS (0.1 mg/kg) inhibited ovarian hormone production and impaired follicular development and ovulation by selectively reducing acetylation of ovarian steroidogenic acutely regulated promoter histones, suggesting that high exposure to PFOS and PFOA may reduce ovarian follicular reserve (28). Similarly, mycotoxins, which are secondary metabolites of various fungi, have had a significant impact on food contamination in many countries, especially in developing countries. Fusarium species produce many trichothecene mycotoxins; one of them is Nivalenol (NIV). Previous studies have shown that NIV exposure in female rats disrupts the reproductive system and reduces fertility. The earlier studies (29)have demonstrated that NIV exposure in female rats disrupts the reproductive system and reduces fertility.Wang et al. found that type B Nivalenol exposure was also capable of mediating OS, leading to meiotic cell cycle arrest and further inducing failure of polar body extrusion in POI mouse oocytes, inducing autophagy and early cell apoptosis (30).
ROS are a group of short-lived, highly reactive, oxygenated products of OS. Their response pathway to OS-induced DNA damage has been widely demonstrated, with DNA damage and chromosome missegregation that can further lead to apoptosis (31). ROS can induce DNA damage and affect DNA damage response(DDR) by mediating genotoxin-induced damage, influencing double-strand breaks (DSBs) sensing, impacting signal transduction within DDR, interfering with cell cycle progression, and inducing p53 transcriptional response and apoptosis (32). Numerous studies have shown that ROS induces DNA breaks as well as decreased base oxidation and DNA repair in GCs, reducing the transfer of nutrients and survival factors to the oocyte, leading to apoptosis and initiating follicular atresia, which may be a key factor in the progression of POI (32–34). Apoptosis in GCs is the cellular mechanism of follicular atresia in mammals, the main process of ovarian follicle and oocyte loss, and the underlying cause of ovarian aging (35). Chemotherapeutic drugs with reproductive toxicity such as cyclophosphamide (CTX) or its main metabolite acrolein can cause excessive production of ROS in vivo, damage DNA structure and function in GCs, and trigger ovarian DNA repair responses and induce apoptosis in oocytes and thus leading to irreversible POI (33, 36). Sha et al. (33) and Jiao et al. (20) both demonstrated that oxidants such as H2O2 and PFNA can mediate OS leading to oxidative DNA damage and cause early apoptosis in GCs cells. ROS are capable of directly inducing DNA damage by oxidizing nucleoside bases (e.g., forming 8-oxoguanine), which can lead to G-T or G-A translocations if not repaired (37). In addition, ROS are able to oxidize deoxyribonucleoside triphosphates (dNTPs) to affect polymerase activity and reduce the rate of replication in vitro (38, 39). Deltamethrin, a class of synthetic pyrethroids, is known as one of the most effective insecticides, because it kills mites and ticks by generating ROS and RNS that directly cause OS damage. Deltamethrin was found to mediate OS to attack DNA and produce genotoxicity, neurotoxicity and reproductive toxicity, including micronucleus induction, chromosomal and nuclear abnormalities and DNA breakage (40). For this reason, it is capable of causing ultrastructural changes such as substrate stripping, border folds, and irregular nuclear membranes in young oocytes (stages I-III) of ticks, interfering with oocyte development and producing reproductive toxicity (41). In addition, Deltamethrin (DM) as an insecticide is able to mediate OS to produce high levels of ROS levels, cause DSBs in DNA of oocytes and induce oocyte apoptosis by regulating Bcl-2-associated X protein (Bax) and B-cell lymphoma-2 (Bcl-2) protein expression, then reduce ovarian reserve function (42).
Mitochondria are a major source of ROS and they not only regulate the level of ROS produced but are also very sensitive to OS damage. ROS accumulation contributes to mitochondrial DNA damage, which further leads to mitochondrial dysfunction (43). Excessive accumulation of ROS causes damage to oocytes and GCs, which in turn leads to ovarian senescence and reduced ovarian reserve. When mitochondrial function is impaired, a mitochondrial stress response mechanism, also known as mitochondrial phagocytosis, is initiated to prevent further damage and restore mitochondrial homeostasis (9, 44). The maternal inheritance of POI and the significant dependence of folliculogenesis on mitochondrial number and energetics suggest that extensive mitochondrial defects may be involved (45). Miao C et al. (46) found that granulosa cell insufficiency in POI was associated with elevated ROS levels and reduced mitochondrial membrane potential (MMP) and adenosine triphosphate(ATP), suggesting considerable mitochondrial dysfunction. Moreover, Bakhshalizadeh S et al. found that mitochondrial ribosomal protein L50 (MRPL50) error variants destabilize mitochondrial ribosomes, leading to oxidative phosphorylation deficiency and syndrome POI (47), highlighting the importance of mitochondrial support in ovarian development and function. Mitochondria are extremely abundant in the oocyte, not only providing energy but also acting as signal transduction hubs to regulate physiological activities such as intracellular signaling, calcium homeostasis, apoptosis and autophagy, which are essential for oocyte quality (48, 49). Mitochondria produce ATP using redox reactions in the respiratory chain complex located in the cell membrane. However, a small amount of ROS is produced in mitochondria due to electron leakage in the electron transport chain (ETC). Physiologically produced ROS act as signal transducers for successful oocyte maturation and ovulation, facilitating the restoration of diploidy in the follicular microenvironment to terminate meiosis in oocytes (50). When mitochondrial dysfunction leads to excessive accumulation of ROS, nevertheless, the mitochondrial genomic DNA responsible for encoding the core components of the ETC lacks histone protection, antioxidant defense and effective DNA repair systems that are more vulnerable to damage (51). Wang et al. (52) proposed mechanisms of mitochondrial-driven ovarian aging including accumulation of mtDNA mutations, mitochondrial dysfunction, impaired fusion and fission, modified membrane potential, altered metabolism and defective ETC function, and reduced mtDNA contents. Bonomi, M et al (45). found significant mtDNA depletion by Taqman copy number assay in women with POI and poor responders to ovarian stimulation compared to 43 women of similar age with intact ovarian reserve or 53 very old women with previous physiological menopause (P < 0.001). Ding Y et al. (53) used PCR-Sanger sequencing to find that POI patients present with mutations or high variability in mitochondrial transfer Ribonucleic acid (mt-tRNA) genes, and these mutations are able to lead to impaired mitochondrial protein synthesis, significantly reduced ATP levels, and elevated ROS levels, strongly suggesting that these mt-tRNA mutations may contribute to mitochondrial dysfunction and play an active role in the progression and pathogenesis of POI. Using a zebrafish Oxr1a mutant, Xu H et al. (54)explored the role of Oxr1a in oocyte maturation and POI. oxr1a deletion leads to imbalance in the redox state of oocytes and mitochondrial dysfunction, entailing the development of a POF-like phenotype. PFNA exposure increases the level of reactive oxygen species, inducing abnormal mitochondrial distribution and increased mitochondrial membrane potential resulting in mitochondrial dysfunction in mouse oocytes (20). DM is also known to mediate OS that causes abnormal mitochondrial distribution, decreases mitochondrial membrane potential, and reduces oocyte quality (42).
OS can affect meiotic progression and also interrupt the normal assembly of meiotic structures, ultimately inducing failure of oocyte maturation and accelerating disease progression in POI (55, 56). H2O2 is a well known signaling agent and is the most direct and effective agent causing OS in cells. It is the major biological ROS, a multifunctional pleiotropic physiological signaler for many essential metabolic functions, and is often used to construct oocyte oxidative stress models and models of early-onset ovarian insufficiency. Single-cell RNA sequencing data showed that H2O2-treated oocytes caused OS mainly through two pathways: “meiosis” and “oxidative phosphorylation”. Furthermore, oxidative stress was observed in oocytes treated with H2O2 to interfere with the mitochondrial distribution pattern and membrane potential abnormalities in oocytes. H2O2 induces elevated endogenous pro-apoptosis-related molecules (Bax, Bak) and reduced anti-apoptotic molecules (Bcl-2, Bcl-xL) in ovarian granulosa cells and regulates apoptosis in ovarian sarcomeres via the ROS-JNK-p53 pathway (57, 58). The H2O2-driven Fenton reaction is able to disrupt spindle morphology and chromosome movement in oocytes during meiosis (59). It has been shown that high levels of ROS in oocytes cause spindle defects, polar body abnormalities and chromosomal mislocalization by acting on microtubules and causing them correctly attached to the centromere (60). Jiao et al. confirmed that PFNA exposure can mediate OS to produce reproductive toxicity in animals by inducing p-extracellular regulated kinase (ERK)1/2 mislocalization which leads to spindle abnormalities, defective aggregation of microtubule organizing center (MTOC) and chromosomal misalignment in MI mouse oocytes (20). Moreover, ROS cause DNA DSBs, block the meiotic progression of oocytes from reaching metaphase II (MII), inhibit follicle maturation and induce adverse pregnancy outcomes (56, 61). Sirtuins molecules are able to extend lifespan, regulate metabolism and repair DNA damage, and possess single ADP-ribosyltransferase or deacylase activity (62). Sirtuins molecules are able to extend lifespan, regulate metabolism and repair DNA damage, and possess single adenosine diphosphate (ADP)-ribosyltransferase (ART) or deacylase activity (62). Zhang et al. (63) found that exposure to the chemotherapeutic drug cisplatin caused high levels of ROS in mouse with POI, significantly disrupting cytoskeletal assembly which includes spindle formation and actin polymerization, thereby destroying meiotic organelle dynamics and arrangement, leading to oocyte meiotic failure and accelerating the decline of ovarian function, which was reversed by tea polyphenol supplementation to regulate ROS levels. Another study also supported the conclusion that high levels of ROS and apoptosis can disrupt the cytoskeleton and mitochondrial integrity, inhibit the meiotic process in oocytes, disrupt the structure of the cumulus cells, and reduce its fertilization potential, which can also be reversed by melatonin (MT) through inhibition of ROS levels (64).
Telomeres are complexes of specialized repetitive DNA sequences and shelter proteins located at the ends of eukaryotic chromosomes. Its basic function is to protect linear chromosome ends from being mistaken for a broken end and being incorrectly repaired by the cell itself or undergoing processes such as DNA end joining or DNA recombination (65). As of 2009, Blackburn, Greider and Szostak were awarded the Nobel Prize for the discovery of the protective effect of telomeres and telomerase on chromosome ends. In women, the effects of reproductive aging on oocyte quality are mainly explained by telomere shortening (66). Telomeres are progressively shortened with each replication due to the inability of cell division to replicate DNA to chromosome ends, resulting in the loss of about 20-50 base pairs of per cell division. In addition, OS also accelerates telomere shortening and induces DNA damage response, impaired cell proliferation, oocyte senescence or apoptosis (17). A review that included 44 studies suggested that POI, diminished ovarian reserve and infertility are associated with shorter telomere length, hence the term telomere shortening is also called “a marker of aging” (67). High levels of ROS often lead to chromosomal instability or abnormalities, spindle defects, reduced mitochondrial function and telomere shortening in older oocytes (over 35 years of age) (68). Butts et al. observed shorter granulocyte TL and lower telomeric DNA in patients with occult ovarian insufficiency (OOI) (69), as well as in POI (70). It was shown that ROS induce local telomeric single-strand breaks (SSBs) by affecting transcription or shelterin binding of telomeric repeat-containing RNA (TERRA) at telomeres and cause collapse of replication fork and telomere loss in somatic cells (71, 72). When telomeres reach extremely short lengths, they can no longer perform their chromosomal end protection function. Since the six-membered protein complex (called shelterin) lacks sufficient binding sites at the very short telomeres, it remodels the telomeres into a “cap” structure. The loss of the telomere “cap” triggers misidentification of DNA damage response proteins at chromosome breaks, exposing and degrading chromosome ends. Robinson L.G et al. found that congenital dyskeratosis is a telomerase mutation-based disorder that may affect oocyte quality and developmental capacity through telomerase dysfunction or mutation or result in telomere shortening, ultimately leading to reduced ovarian reserve (73).
Signaling pathways, as enzymatic response pathways, can transmit molecular signals from extracellular to cellular in order to function, and the pathogenesis of POI involves the involvement of related signaling pathways (see Figure 1). High levels of ROS can mediate signaling pathways such as Phosphatidylinositol 3 kinase/protein kinase B (PI3K/Akt), mitogen-activated protein kinase (MAPK), and Kelch-like ECH-associated protein 1 (Keap1)-nuclear factor erythroid 2-related factor 2 (Nrf2)-antioxidant response elements (ARE) involved in ovarian oxidative damage, GCs apoptosis, follicular atresia, and other biological processes associated with POI (74). In contrast, the transforming growth factor (TGF)-β/Smads signaling pathway can regulate cellular autophagy to reverse the manifestation of ovarian aging caused by OS.
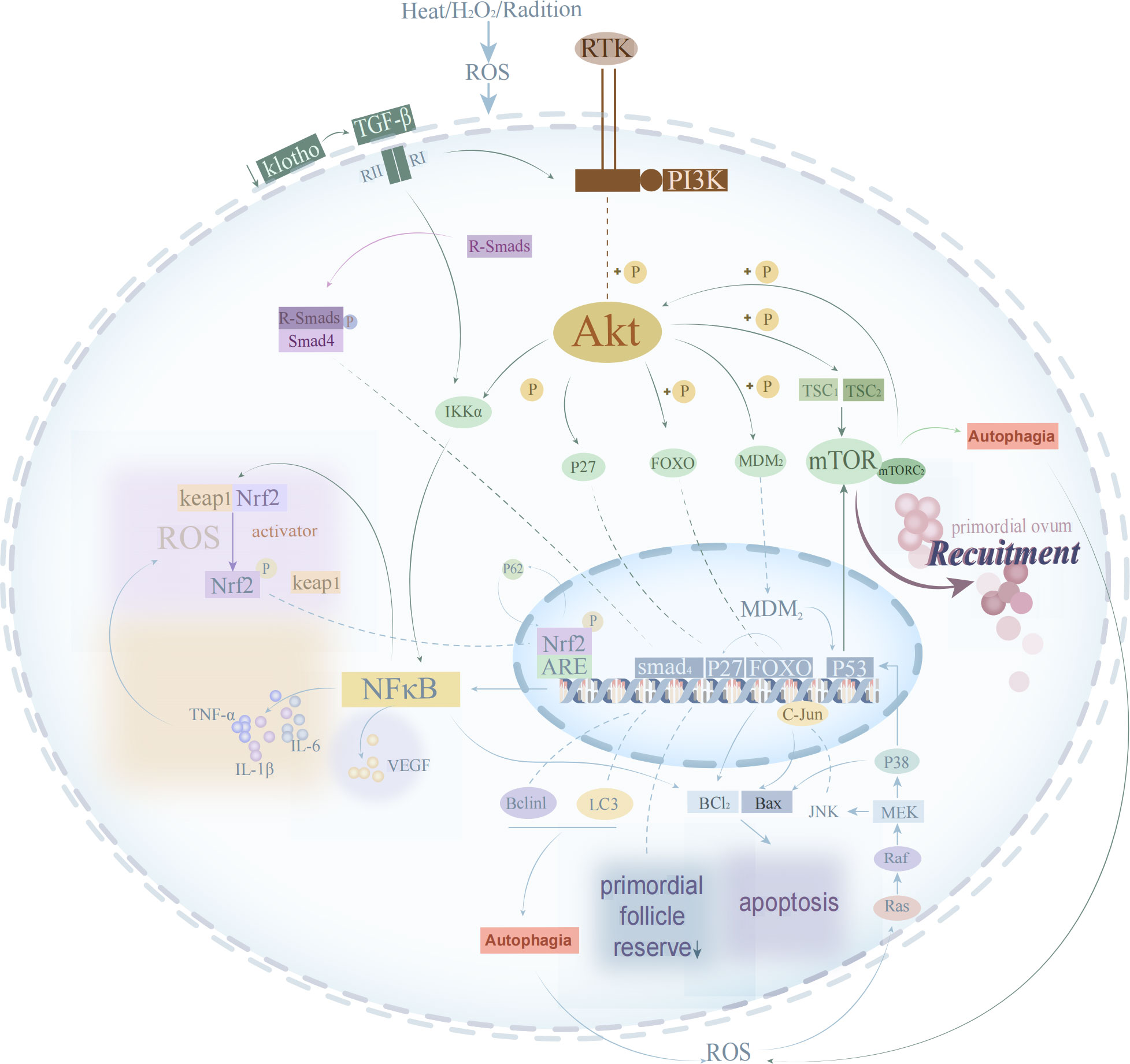
Figure 1 The process of oxidative stress signal transduction in oocyte.
The PI3K/Akt signaling pathway promotes oocyte proliferation and differentiation and inhibits apoptosis (74). If abnormally blocked, the development of primordial follicles is inhibited which leads to the development of POI (75). The PI3K-Akt-mammalian target of rapamycin (mTOR) signaling pathway is competent to regulate the HPG axis (76), and the activated Akt pathway further causes a cascade of signaling pathway responses. Phosphorylated Akt can phosphorylate a series of downstream target proteins such as forkhead box type O transcription factor-3a (FOXO3a) (anti-proliferation and apoptosis), P27 (maintenance of primordial follicular reserve), Bcl-xL Bcl-2 associated death promoter (BAD) (a pro-regulatory member of the Bcl-2 family, involved in cellular mitochondrial regulatory pathways) and mTOR (controls protein biosynthesis and regulates cell growth) (74). PI3K/Akt inhibits primordial follicle overactivation by acting on the phosphorylation site of FoxO protein to avoid excessive depletion of primordial follicles and prolong ovarian reserve and fertility (77). Loss of Klotho causes attenuation of FOXO3 through PI3K/Akt signaling and disrupts the balance of oxidative and antioxidant systems, ultimately leading to POI (18). Quercetin (antioxidant) can restore ovarian function and inhibit OS by activating PI3K-Akt-FoxO3a signaling pathway and increasing superoxide dismutase (SOD) and glutathione peroxidase (GPX) expression in POI rat (78). In the case of follicular depletion, anti-Mullerian hormone (AMH) produced by GCs of early developing ovarian follicles is also involved in protecting the primordial follicles (PMFs) pool by inhibiting FOXO3/FOXO3A phosphorylation (leading to PMF activation) to induce ovarian autophagy (70). The PI3K/AKT/mTOR signaling pathway can regulate cell proliferation and differentiation, inhibit apoptosis by activating ribosomal kinases and promote cell cycle progression (79). Cheng Y et al. (80) found that knockdown of the TSC1/TSC2 gene was able to activate mTOR of phosphatase and tensin homology deleted on chromosome 10 (PTEN)/tuberous sclerosis complex 1 (TSC1)/tuberous sclerosis complex 2 (TSC2) in germ cells leading to primordial follicle recruitment, POI and infertility (81). Zhao Di (82) discover that the Chinese herbal medicine Erxian Decoction could promote GCs proliferation and inhibit apoptosis and exert a protective effect on cisplatin-injured ovarian GCs. On the one hand, it could promote the cell cycle transition from G0 to S phase of GCs by down-regulating the cell cycle blocking protein P27kip1; on the other hand, it could down-regulate the expression of FOXO3a and the pro-apoptotic protein Bim.
MAPK signaling pathway regulates apoptosis through complex mechanisms such as enhanced c-Myc expression, P53 phosphorylation, involvement in Fas/Fas L-mediated apoptosis, activation of c-Jun and c-fos, and induction of Bax translocation. Scholars have cloned and identified six MAPK subfamilies in mammalian cells: they are c-Jun N-terminal kinase(JNK) 1/2/3, ERK1/2, p38MAPK (p38α/β/γ/δ), ERK7/8, ERK3/4, and ERK5/BMK1 (large MAP kinase 1) (83). When different subfamilies are activated by upstream kinases, they regulate physiological processes such as inflammation, stress, cell growth, cell development, differentiation and death by phosphorylating multiple substrates such as transcription factors (84). Xu et al. (85) found that zinc pretreatment significantly reduced ROS levels in mouse embryonic osteoblasts (MC3T3-E1) via the MAPK/ERK signaling pathway and prevented OS-induced apoptosis in mouse.The JNK has been shown to induce apoptosis or growth inhibition and protect the organism from OS. Upon activation of the upstream kinases MKK4 and MKK7 (i.e. MAP2K) of the JNK pathway by different upstream MAP3Ks, respectively (83), JNKs translocate from the cytoplasm to the nucleus (86) and activate the transcription factor c-Jun translocated to the nucleus for regulating the expression of the pro- or anti-apoptotic genes Bax and Bcl-2 (87, 88). Sun et al. (89) knocked down isocitrate dehydrogenase-1 (IDH1) in a CUS mouse model and detected that elevated ROS levels in its human ovarian GCs, which induced autophagy activation and triggered cell cycle arrest in the S and G2/M phases of human ovarian GCs, which was reversed by the ROS scavenger N-acetylcysteine (NAC). Additionally, activation of ERK pathway was found to be involved in autophagy-related cell proliferation inhibition and cellular senescence, as well as JNK and p38 MAPK signaling pathways were found to be involved in regulating cell cycle arrest-related cell proliferation inhibition and senescence in IDH1 knockdown human ovarian GCs.
TGF-β)/Smads signaling is an important transduction pathway that regulates follicular development, and abnormalities in either process of the transduction pathway may lead to impaired signaling, resulting in impaired follicular recruitment, inhibition of follicular growth and development, accelerated follicular atresia, and POI (74). Extensive studies have shown that the transforming growth factor (TGF)-β/smad signaling pathway can regulate GCs differentiation and apoptosis, interfere with follicular development and ovulation, and is a potential mechanism for autophagy disorders in POI. Autophagy is a cytolytic metabolic pathway that helps regulate the various stages of oocyte death and follicular atresia development, and the expression or activation of autophagy-related proteins is most pronounced in high stress conditions (e.g., starvation, heat, and hypoxia) to inhibit apoptotic signals in ovarian oocytes and/or granulosa cells and promote oocyte and follicle survival, thereby ensuring fertility (90). Autophagy plays a key role in the anti-aging process through lysosomal degradation and recycling of proteins and organelles, and also plays an important part in ovarian homeostasis by reducing the oxidative load of oocytes (91). Klotho is a transmembrane enzyme that controls insulin sensitivity in organisms and inhibits OS and inflammation, whose loss leads to premature aging (62). It was indicated that reduced Klotho expression in the POI mouse model activated the TGF-β/smad signaling pathway, which in turn downregulated autophagy-related factors such as Lc3 and Beclin1 in GCs.Then, autophagy was attenuated, ROS clearance was reduced, and ROS levels increased, which subsequently led to further POI (92).
The Keap1, Nrf2 and ARE signaling pathway is a classical pathway to resist endogenous and exogenous OS and a key link to inhibit the inflammatory response against endogenous and exogenous OS, which is closely associated with inflammatory diseases such as aging (93). Under physiological conditions, Keap1 targets Nrf2 for ubiquitin-dependent proteasomal degradation. During OS, Keap1 is inactivated and ubiquitination of Nrf2 is halted, leading to accumulation and activation of newly synthesized Nrf2. Thus, Nrf2 translocation to the nucleus induces the transcription of cytoprotective genes that regulate the redox state of cells to maintain cellular homeostasis (94). Nrf2 is an essential sensor and regulator of chemical homeostasis in ovarian cells that protects them from aggressive effects by managing metabolic detoxification and exerting endogenous antioxidant effects (95). In parallel with the activation of oxidative damage, Nrf2 and its cytoplasmic binding protein cooperate with Keap1 to bind to the antioxidant response element (ARE) in the target gene promoter and promote the transcription of the antioxidant gene heme oxygenase-1 (HO-1), thereby regulating the redox state of the cell to maintain cellular homeostasis (96). Chen Y et al. also verified this conclusion that ovarian tissues of POI model rats suffered from oxidative damage and the expression of Keap-1 protein was increased in ovarian tissues, while the expression of Nrf2/HO-1 protein was decreased (97). Several studies have demonstrated that Keap1-Nrf2-ARE pathway-related drugs treat age-related eye diseases through anti-OS (98). In addition, Nrf2 deficiency upregulates nuclear factor-kappa B (NF-κB) activity, leading to increased inflammatory cytokine release (99), while Nrf2 up-regulation leads to an attenuated inflammatory response in rodent models of liver and kidney injury (100). However, NF-κB activity can regulate Nrf2-mediated antioxidant gene expression. In this context, Yu et al. (93) have demonstrated that overexpression of the typical NF-κB subunit p65 increased nuclear Keap1 levels, leading to reduced Nrf2/ARE signaling. Chen (101) found that TCM activated the Keap1-Nrf2-ARE signaling pathway and downstream antioxidant enzymes SOD, heme oxygenase-1 and quinone oxidoreductase (NQO1) activities to improve ovarian function, reduce OS and inflammatory status in POI-immune model mouse. The Nrf2/HO-1 pathway also removes ROS by activating Nrf2 downstream factors, HO-1 and sequential enzymatic reactions, and the anti-senescence protein klotho inhibits ovarian and endothelial cell senescence by activating the Nrf2 pathway (102). Shang Z et al. found that excess ROS were removed by activation of HO-1, a factor downstream of Nrf2, to produce a continuous enzymatic response. The expression of Nrf2 and HO-1 decreased after D-galactose (D-gal) treatment, but increased after red ginseng extract (RGE) treatment (103).
OS damages biomolecules and forms a series of cascade reactions that cause endogenous damage in the organism. The core of OS is the anti-apoptotic proteins (104), FOXO (105), P53 (106), Methylation of N6 adenosine/N6-methyladenosine (M6A) (107), and other regulatory factors that activate or inhibit signaling pathways and alter the structural and functional properties of host molecules, and transcription factor dysfunction can accelerate the onset of POI. Currently, the clinical mechanism of action for the treatment of POI includes promoting the stable expression of anti-apoptotic proteins, FOXO, P53, and exosomes, thereby reducing the damage to the ovaries caused by OS.
Free radicals, such as ROS, have been shown to play an important role in apoptosis. Due to the uniqueness of the ovarian physiological environment and follicular development process, apoptosis of ovarian GCs and oocytes synergistically causes follicular atresia and accelerates the process of ovarian aging. The Bcl2 family is an important regulator of the endogenous pathway mediating apoptosis and can be divided into two groups: one is the regulators represented by Bcl-2 that inhibit apoptosis and suppress cell death caused by various cytotoxic factors; the other is the regulators represented by Bax that promote apoptosis and whose overexpression can antagonize the protective effect of Bcl-2 and cause cell death (108). The balance of Bax/Bcl-2 relationship can regulate apoptosis and maintain the stability of the cellular environment (109). In turn, Bcl-2-associated athanogene 3(BAG-3), as an anti-apoptotic molecular chaperone protein, also attenuates ovarian damage caused by OS. Zhu Rong et al. found that up-regulation of Bcl-2 and down-regulation of Bax inhibited follicular atresia and GCs apoptosis in POI (110). Thus, apoptosis and OS form a “vicious cycle” during POI, and therefore the stable expression of apoptosis-related proteins is important for ovarian aging (111).
The Bcl-2 family has a role in regulating ovarian GCs apoptosis at all stages of follicular growth and development (112).Bcl-2 is localized at sites of reactive oxygen species production such as mitochondria, ER and nuclear membranes and is a key regulator of the signals leading to activation of cysteine aspartases. Edlich F found that Bcl-2 protein prevents the release of apoptosis-associated proteins from mitochondria and resists ovarian apoptosis through aquaporin-1 protein (113). When Bcl-2 expression increases, mitochondrial membrane potential enhances and membrane permeability stabilizes, hindering the release of cytochrome C and preventing its activation of the downstream caspase family of apoptotic factors, which strengthens GCs against apoptosis and prevents the effects of OS-induced apoptosis on ovarian function (104, 114, 115). Bcl-2, despite not affecting oxygen radical production, prevents oxidative damage to GCs components, including lipid membranes (116, 117). However, it has still been suggested that Bcl-2 overexpression, although protecting primary fibroblasts from OS-mediated reduction in cell proliferation, does not prevent premature senescence (118).
BAX is a pro-apoptotic protein that is a key executioner of cell death by inducing permeabilization of the outer mitochondrial membrane, leading to mitochondrial regulation of cell death (119). If BAX expression increases, it causes a decrease in mitochondrial membrane potential and an increase in membrane permeability, which in turn releases cytochrome C into the cytoplasm, activates the caspase family, accelerates apoptosis of GCs, contributes to follicular atresia, and accelerates disease progression in POI (120, 121). Nevertheless, when OS-mediated ROS levels are abnormally elevated in oocytes and GCs, it causes the opening of the mitochondrial permeability transition pore, upregulates the expression of the pro-regulatory protein Bax or activates the Caspase family (122), accelerating the onset of POI.
BAG-3 is a key element of cellular proteostasis, a molecular chaperone protein that significantly promotes Bcl-2 anti-apoptosis and is involved in biological processes such as stress response, proliferation, migration and apoptosis (123). BAG3 interacts with the proteasome and regulates its activity to maintain cell survival and prolong cell life by inhibiting apoptosis for anti-aging purposes (124, 125). It was found that endothelial-specific BAG3 knockdown in mouse models significantly enhanced OS-related endothelial damage and vascular remodeling, affecting ovarian vascular function and thus accelerating ovarian aging (126). Furthermore, stress stimulation induced the expression of BAG-3, which promoted cell survival. One mechanism by which BAG3 is endowed with anti-apoptotic effects is that it binds to anti-apoptotic proteins such as κB kinase and γ inhibitors and prevents their transport to the proteasome by heat shock proteins 70 (HSP70), therefore exerting anti-apoptotic effects (127), and can attenuate the stress response. Thus, it is clear that the expression of BAG-3 can achieve its anti-apoptotic effect by inhibiting OS and attenuate the damage to ovaries by OS.
FOXO is an important class of transcription factors and are important determinants of aging, whichare involved in several processes such as cell cycle arrest, metabolism and apoptosis, and DNA damage repair (128, 129). FOXO1 and FOXO3 are highly expressed in GCs of ovarian follicles, and FOXO3a functions as an inhibitor of follicular activation at the earliest stages of follicular growth (130), and its sustained expression leads to retarded oocyte and follicular development and delays oocyte aging. In contrast, OS can activate the PI3K/AKT pathway and lead to inactivation of FOXO3a phosphorylation, resulting in the development of POI (131). Furthermore, Foxo1-mediated changes in signaling can induce low-grade chronic inflammation in vivo (132). Dabravolski S.A et al. (133) found that OS and chronic inflammation are capable of interacting and promoting each other to synergistically impair ovarian function. Beyond that, the abnormal increase in FOXO transcriptional activity can also alter the Bax/Bcl-2 ratio by regulating the expression of BH3-only protein Bim, “a direct activator of the pro-apoptotic protein Bax”, which ultimately leads to cysteine activation, DNA fragmentation and mitochondrial dysfunction, hence accelerates OS, forming a vicious cycle that speeds up the aging of oocytes (134). Cui et al. found that overexpression of FOXO aggravates OS, but deacetylation of FOXO1 by Silent Information Regulator 3 (SIRT3) can correct excessive OS (135), thus suggesting that stable expression of FOXO can maintain normal ovarian function.
P53 is a tumor suppressor gene that mediates cellular signaling pathways to regulate normal cellular life activities and intracellular redox homeostasis (136, 137). P53 is strictly regulated by the E3 ubiquitin ligase murine double minute 2(MDM2) through an autoregulatory feedback loop, and downregulation of MDM2 activity can increase the level and stability of P53, which further inhibits oocyte proliferation in order to prevent oocyte apoptosis or senescence (138). Haraguchi H et al. found that abnormal increases of P53 in ovarian GCs were able to reduce oocyte quality and ovarian function (139) and induce the appearance of premature aging phenotypes (140). Wang further discovered that OS can upregulate MDM2 and reduce P53 stability, while accelerating cellular senescence, namely, the MDM2 and P53 can form a feedback regulatory loop (141). When MDM2 is absent, however, it leads to significant P53 nuclear accumulation in oocytes, which improves the fertility potential of a mouse model resembling the human POI phenotype (142). Stable expression of P53 not only regulates oocyte quality, but also regulates oocyte autophagy. Autophagy, as a catabolic pathway, can serve as a source of energy to maintain cell cycle arrest and sustain DNA repair activities (143). Moreover, P53 is also capable of mediating cellular autophagy to remove obsolete intracellular mitochondria and alleviate OS and its induced senescence in oocytes.
Internal modifications of messenger RNA (mRNA) are used to maintain the stability of mRNA and the most common internal modifications include M6A, Methylation of N1 adenosine/N1-methyladenosine (M1A), and 5-methylcytosine (M5C) (144). The M6A dynamic reversible modifications of RNA require methyltransferases such as methyltransferase-like 3 (METTL3) (145) and demethylases such as fat mass- and obesity-associated (FTO) (146) protein to jointly complete the regulation of mRNA function. Li Wanjing et al. (147) identified that ovarian reserve function could be improved by regulating ovarian RNA-M6A methylation modification, and another study confirmed that the M6A content in the RNA of POI patients and mice was significantly higher than that of the control group (148). Downregulation of M6A demethylases including FTO protein and ALKB homolog 5 (ALKBH5) leads to increased M6A in aged ovarian GCs, while knockdown of GCs in FTO shows a more rapid ovarian premature aging phenotype (149), thus increased M6A impairs ovarian function and accelerates the process of ovarian aging. Nrf2 is an endogenous antioxidant pathway, and it has been found that FTO can increase Nrf2 expression by mediating M6A demethylation of Nrf2 mRNA, thereby inhibiting OS (150), which shows that a decrease in M6A concentration is beneficial in alleviating OS. Additionally, M6A modifications can regulate cellular ROS levels through different mechanisms, and imbalanced OS and antioxidant imbalance can also increase M6A levels (107). It is thus hypothesized that OS causes an increase in M6A in mRNA, impairing ovarian function and accelerating the process of POI, but further confirmation is needed.
The sirtuins are an evolutionarily conserved family of nicotinamide adenine dinucleotide (NAD)-dependent histone deacetylases that consist of seven members including SIRT1-7, which are involved in many biological processes such as cell survival, senescence, proliferation, and apoptosis by regulating inflammation, OS, and mitochondrial function (151, 152). Zhang et al. found a positive correlation between sirtuins and treated with chemotherapy mouse models (153). Another study showed that SIRT1 deficiency significantly increased ROS and inflammation levels, and SIRT3 can promote mitochondrial oxidative metabolism in response to nutritional stress and membrane depolarization to attenuate OS (154), thus it can be assumed that sirtuins can mediate OS involved in ovarian aging. Sirtuins-mediated OS and inflammation also lead to the expression of genes like Nrf2, NF-kB, interleukin (IL)-1, FOXO and P53, which also damage oocytes (155). SIRT2 and SIRT3 are able to positively regulate FOXO3 and increase ovarian reserve; conversely, defective sirtuins expression reduces follicle number and accelerates the depletion of PMFs (156). Yang, M., et al. found that increased expression of SIRT1, SIRT2, and SIRT3 reduced P53 acetylation levels, inhibited P53 hyperactivation, and diminished follicular apoptosis and atresia. Meanwhile, the upregulation of sirtuins inhibited P53-dependent cell cycle arrest and apoptosis (157), reducing abnormal follicular atresia to maintain normal ovarian function (158, 159).
The ovarian microenvironment refers to the environment of cell survival in which various intercellular and humoral components of the ovary participate together to constitute. It contains rich microenvironmental signals such as gap junctions, paracrine, autocrine, endocrine and exocrine signaling factors that facilitate intercellular communication. Oxidative damage to biomolecules caused by OS can lead to the production and release of cytokines through initiating endogenous damage related molecular patterns in the organism. Cytokines further recognize receptors and activate downstream signaling pathways, such as nuclear factor kB (NF-kB), janus kinase (JAK), signal transducer and activator of transcription (STAT) and MAPK, thus providing increased release of cytokines and inflammatory factors, recruiting and activating more inflammatory cells and causing a chronic inflammatory response in the body. ROS are oxygen-containing reactive chemicals that are involved in cell signaling and promote cell survival, proliferation, and differentiation at a physiological level (160). Once ROS levels exceed cellular antioxidant levels, they react with DNA, proteins, lipids and carbohydrates; leading to DNA strand breaks as well as protein and lipid oxidation. Not only do they impair ovarian function and accelerate the onset of premature ovarian failure (161–163).
Existing studies have implicated that OS is of great importance in the development and perpetuation of inflammation, increasing the levels of inflammatory factors such as tumor necrosis factor-α (TNF-α), IL-1β, IL-6, and TGF-β in vivo (164). Han et al. (165) detected that elevated levels of IL-4, IL-1β, and IL-6 in follicular fluid samples and GCs from POI patients, and further found that IL-4 could activate the PI3K/AKT signaling pathway, promote apoptosis, and inhibit GCs proliferation in in vitro experiments. Besides, inflammatory factors can also activate NF-κB signaling pathway and affect the expression of follicle-stimulating hormone receptor(FSHR) through glucose-regulated protein 78-mediated ER stress to promote GCs proliferation. Luo et al. (158) noticed that decreased nuclear p65 expression in ovarian GCs and reduced number of sinus follicles in a mouse model of NF-κB inactivation, showing that OS can increase the concentration of inflammatory factors and further decline the expression of NF-κB to participate in the POI process. However, over-activation of NF-κB can induce the expression of cytokines such as IL- 1β, adhesion molecules, immune receptors, and inflammation-related enzymes, which aggravate the damage to the ovary by OS and thus form a vicious circle, thereby suggesting that stable expression of inflammatory factors is beneficial for the maintenance of ovarian function. Mantawy EM et al. (166) discovered that salicin was able to ameliorate radiation-induced POI by decreasing the expression of radiation-induced ovarian damage markers including NF-κB, TNF-α, inducible nitric oxide synthase (iNOS) and cyclooxygenase (COX)-2 and down-regulating the TGF-β/MAPK signaling pathway to impede POI-related inflammatory and apoptotic signaling pathways. We found in another study that in a fluorosis POI model, inflammatory factors IL-6 and IL-8 were significantly increased, ovarian ultrastructure was obviously damaged and showed premature failure, and there was a positive correlation with the dose of fluoride dosing, which indicated that the concentration of pro-inflammatory factors was positively correlated with the degree of POI (167). While some potential drugs for the treatment of POI can reduce the levels of inflammatory factors and modulate the ovarian microenvironment thus exerting a therapeutic effect on POI For example, resveratrol can lead to a significant decrease in TNF-α and IL-6 concentrations in the mice’s ovary (159), lacrimalin down-regulates peripheral blood CC chemokine receptor 9 (CCR9), C-X-C-motif chemokine receptor 3 (CXCR3) and CD4 T lymphocyte counts and IL-12 secretion (168).
Lipids maintain the structural integrity of cells. Lipids containing polyunsaturated fatty acids are susceptible to oxidation triggered by free radicals, causing chain reactions to form lipid peroxidation products such as malonaldehyde and 4-hydroxynonenoic acid, which alter the fluidity and permeability of cell membranes and cause cellular dysfunction in aging and most age-related diseases. Disruption of redox homeostasis is a key phenotype of POI (169–172) and is closely associated with lipid peroxidation. Among the lipid oxidation metabolites, arachidonic acid metabolism is dramatically altered. Arachidonic acid may induce apoptosis by attenuating oocyte proliferation, migration and viability in response to mitochondrial membrane depolarization and activation of cystathione-3, -8 and -9 (173–175). 4-Hydroxynonenoic acid disrupts signal transduction and protein activity, as well as induces chronic inflammation and triggers apoptosis in the ovary under conditions of OS (176), contributing to decreased reserve function and ovarian aging.
The accumulation of oxidatively modified proteins is a signature of aging of organisms in vivo and cellular replicative senescence in vitro (177). Protein carbonylation is one of the most deleterious irreversible oxidative protein modifications and is the initiating factor for mitochondrial dysfunction and ER stress (178), which is also considered to be a major marker of OS-related diseases (179), with elevated concentrations in senescent cells (180). One research identified by mass spectrometry that almost half of the carbonylated proteins are located in mitochondria, reflecting the preferential accumulation of damaged proteins in mitochondria during cellular senescence (181), and impaired mitochondrial function is capable of inducing OS responses. In a research (182), PFOA was found to considerably increase the production of ROS, protein carbonylation and DNA damage in oocytes, impairing the maturation and viability of porcine oocytes and affecting their fertility. Protein damage leading to carbonyl formation originates from the direct oxidation of amino acid side chains, but can also be obtained through the formation of protein adducts of lipid peroxidation products and dicarbonyl glycation compounds (183). Under OS conditions, consequently, protein peroxidation and lipid peroxidation interact and promote each other, speeding up the occurrence of POI.
Several factors that promote and inhibit angiogenesis around the follicle work together to regulate ovarian angiogenesis, and vascular endothelial growth factor (VEGF) plays a key role in ovarian angiogenesis, especially during follicular development and ovulation (184). Selection and ovulation of the dominant follicle depends on a well-functioning vascular supply and permeability within the antral follicle, maintaining the normal function of the ovary accordingly. Instead, damage to the endothelium and inadequate capillary network formation in the membrane cell layer can lead to follicular atresia or degeneration (185, 186). Insufficient blood supply leads to a hypoxic state in the follicular fluid and inhibition of angiogenesis which can cause disruption of follicular development, ovulation and ovarian endocrine function (187). There is also some research showing that ovarian tissue VEGF protein expression is clearly lower in rats with POI, demonstrating that follicle development is closely linked to angiogenesis (188). Mitochondrial ROS is instrumental in promoting angiogenic conversion in resting endothelial cells (189), and OS impairs lysosomal clearance of damaged DNA, enhances stimulator of interferon genes (STING) signaling, and up-regulates VEGF expression in senescent cells, which is beneficial for angiogenesis (190). Hence, good regulation of OS promotes angiogenesis and tissue repair (191), stimulates ovarian endothelial cell motility and proliferation in vitro or vivo, boosts follicle development, and reduces apoptosis (192). Nonetheless, vascular endothelial dysfunction occurs when the increase of immune cells in the vessel wall leads to the release of inflammatory cytokines and induces the production of ROS, which eventually leads to the generation of excessive OS (193), entailing a disruption of vascular homeostasis. OS-mediated epithelial cell injury may also contribute to a decrease in VEGF levels (194), and it has been evidenced that young women with POI have evident vascular endothelial dysfunction (195), so maintaining OS at physiological level is beneficial for the stability of the ovarian microenvironment and slows down the process of ovarian aging.
Exosomes are small membrane vesicles (30-150 nm) containing complex RNAs and proteins, which are crucial for intercellular communication and information transfer and are able to improve POI by inhibiting apoptosis and promoting angiogenesis (196, 197). Some of these healthy cells, including MSCs, neural progenitor cells and astrocytes, have antioxidant damage and their derived exosomes show the same efficacy as parental cells (198, 199). It was shown that miR-144-5 p carried by bone marrow mesenchymal stem cell-derived exosomes inhibited the expression of apoptosis-related proteins by suppressing the PTEN, thereby up-regulating the PI3K/ATK pathway to promote ovarian follicle recovery after chemotherapy and improving follicle morphology in POI mouse. This conclusion was verified by Yang, Meiling et al. (200), Exosomes inhibited cisplatin-induced GCs apoptosis and increased cell viability. In contrast, OS can notably affect the number of exosomes by regulating the degradation of multivesicular bodies. High levels of OS promote the degradation of intracellular multivesicular bodies by activating autophagy, lessening the amount of exosomes, thereby influencing exosome function, reducing the antioxidant effect of exosomes, and exacerbating the damage of OS on oocytes (201). Qu et al. determined that human umbilical cord MSC-derived exosomes carrying miR-126-3p promoted angiogenesis in POI rats and attenuated apoptosis of ovarian GCs, highlighting the potential of miR-126-3p-containing exosomes as an effective therapeutic strategy for POI treatment (202). In addition, Sun et al. (203) discovered that miR-644-5p carried by bone mesenchymal stem cell-derived exosomes could inhibit apoptosis of ovarian GCs by targeting cellular p53, suggesting the potential of exosome-carried micro Ribose Nucleic Acid(miRNA) to treat POI and restore ovarian function through their antioxidant effects.
The key to counteracting ovarian functional impairment in POI patients caused by OS is to restore the dynamic balance between ROS and the antioxidant system by reducing the production of ROS on the one hand and increasing the activity of the endogenous antioxidant system or supplementing exogenous antioxidant substances to enhance the ability to scavenge ROS on the other hand (204). The endogenous antioxidant system of the organism consists of two main components, an enzymatic scavenging system consisting of various antioxidant enzymes such as SOD and catalase (CAT), and a non-enzymatic scavenging system comprising antioxidants including GSH and MT (205). Furthermore, herbal extracts such as ginsenosides, curcumin (CUR), resveratrol and stem cell transplantation also perform important roles in the fight against OS, and we will introduce several antioxidants with outstanding efficacy and good research prospects (Table 1).
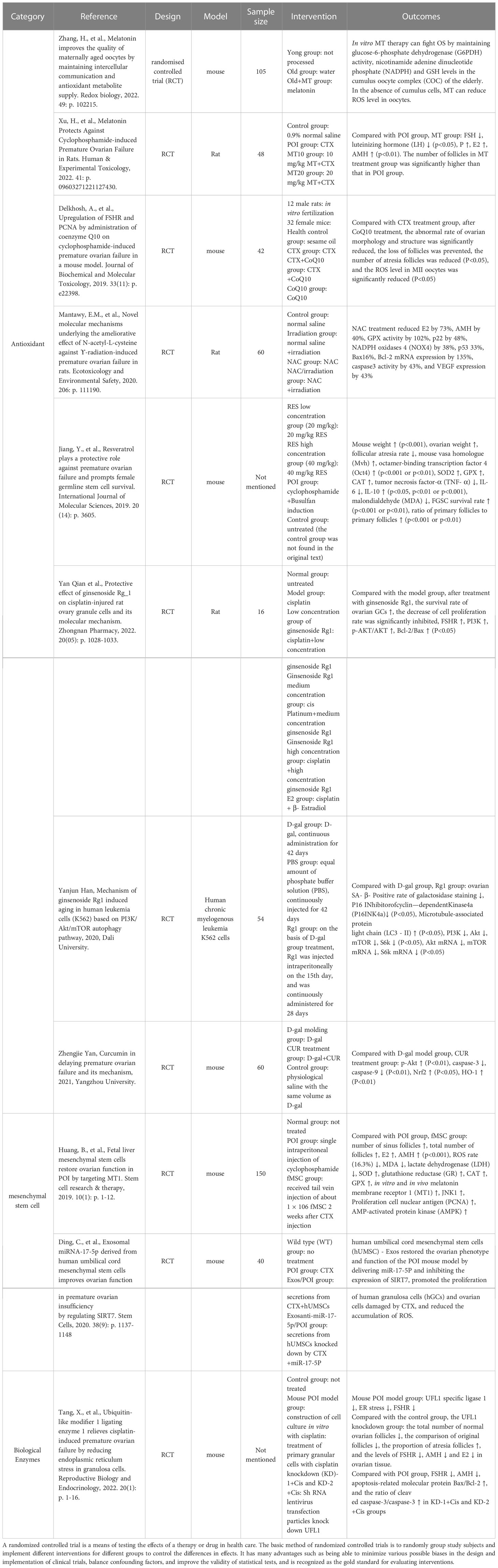
Table 1 The application of antioxidants.
MT, an amine hormone produced by the pineal gland in mammals and humans, is secreted in tissues such as the ovary and placenta (206). In the long-term decline of ovarian aging, melatonin treatment is able to regulate ovarian function through anti-oxidative stress and also protect the follicular pool, oocyte quantity and quality (207). Multiple studies have indicated that MT protects mice’ and hamsters’ GCs and oocytes from ROS damage by reducing OS (208, 209), thereby protecting ovarian function and delaying the development of POI. Osatd-Rahimi N et al. (210) found that melatonin increases the activity of CAT, SOD and thiols, thus it has a protective effect against oxidative stress. Additionally, melatonin plays an effective role in reducing malondialdehyde (MDA) levels, which are a product of lipid peroxidation in cell membranes. It can directly scavenge reactive oxygen radicals, promote the synthesis of the reactive oxygen scavenger GSH in the body, and mitigate OS damage to mitochondrial proteins and DNA (211). Cyclophosphamide (CTX), an alkylating agent with the highest risk of ovarian damage, is commonly used in the treatment of malignancies such as breast cancer (212). According to studies, CTX is catalyzed and degraded in vivo by hepatic microsomal enzymes, resulting in the production of phosphoramidite mustard and acrylic acid, which have cytotoxic effects (213). During the course of treatment, abnormal blood circulation and associated stress damage to the body may result in the release of large amounts of free radicals. It promotes peroxidation and releases large amounts of ROS, which damages the mitochondrial and cellular membranes of ovarian granulosa cells, induces an inflammatory cascade, releases alarm signals (alarms), and initiates cellular responses (214). Whereas granulosa cells in the ovary provide an important environment for oocyte formation and development (215). CTX alters follicular growth, oocyte maturation and apoptosis or atresia in the ovary due to the production of ROS and inflammatory substances. Xu et al. (216) examined that MT improved POI in a cyclophosphamide-induced POI in rat model by maintaining ovarian hormonal status, enhancing ovarian index, promoting follicular development, and inhibiting GCs apoptosis. Animal and human trials showed that short-term use of MT was safe and long-term MT treatment caused only minor adverse effects comparable to placebo (217).
Sphingosine-1-phosphate (S1P) is a biologically active sphingolipid that mediates various biological processes such as apoptosis, immune response and inflammation, and can substantially reduce oocyte apoptosis initiated by various stimuli such as radiation and chemotherapeutic drugs, effectively protecting ovarian tissue to alleviate POI (218). It was found that OS can induce cellular regulation by altering the balance of Cer/S1P, and with elevated Ceramide (Cer)/sphingosine-1-phosphate (S1P) levels, cells tend to move in a direction favorable to life activity. Conversely, cells tend to die (219), so S1P is an important antioxidant in reducing apoptosis, and thus treating POI. There are fewer reports on its safety, and one study detected that S1P treatment interferes with the clinical effects of anticancer drugs during chemotherapy, and its anti-apoptotic effects may inhibit normal atresia of DNA-damaged oocytes during follicular development (206).
Coenzyme Q10 (CoQ10) is a fat-soluble quinone, a natural antioxidant found in the mitochondrial respiratory chain. Exogenous supplementation of CoQ10 has been shown to improve ovarian function in mouse (220). It has a beneficial effect on POI patients by reducing OS, preventing mtDNA mutations caused by peroxidative damage, restoring mitochondrial function, counteracting ovarian physiological aging and improving ovarian reserve function for enhancing ovarian responsiveness (221). Delkhosh A et al. (220) discovered that CoQ10 markedly improved ovarian histological morphology and the number of atretic follicles in CTX-treated mouse ovaries by down-regulating ROS levels. It was also shown that CoQ10 is well tolerated, has few adverse effects, and is highly safe (222).
N-Acetyl-L-cysteine (NAC) is a thiol antioxidant whose main role is to disturb the generation of free radicals and to scavenge those already generated (223). Mantawy EM et al. (224) revealed that NAC inhibits radiotherapy-induced POI by down-regulating nicotinamide adenine dinucleotide phosphate oxidase (NOX) and p22 proteins and enhancing antioxidant defense to eliminate oxidative damage, and suppresses p53-dependent apoptotic mechanisms by reducing multiple pathways including MAPK signaling pathway, and also reduces follicular atresia and increases ovarian reserve by inhibiting radiation-triggered primordial follicular loss, thereby restraining radiotherapy-induced POI in rats. At the same time, it preserves ovarian function and structure, and plays a positive role in POI patients. Clinical studies have proven that NAC has a good safety and tolerability profile (225).
Vitamin C and E are classic antioxidant molecules. Vitamin C converts oxidized glutathione to reduced glutathione. The α-tocopherol in vitamin E can directly scavenge excess ROS from the body, modulate antioxidant responses, and enhance the expression of antioxidant enzymes like SOD (226), thus exerting an antioxidant stress effect and favorably affecting POI patients. Resveratrol (RES), a naturally occurring dietary antioxidant flavonoid, is the most recognized antioxidant and the strongest activator of the longevity gene SIRT1. It can effectively improve ovarian function and female germline stem cell (FGSC) regeneration in POI models by alleviating OS, inflammation, and mechanisms involving the Hh signaling pathway (159), while FGSCs can initiate meiosis in the ovary to produce oocytes and expand follicular reserve to preserve fertility. RES also apparently rescues the activity of antioxidant enzymes including GPX, SOD, and CAT in POI ovaries (159) and counteracts OS, which plays an essential role in the treatment of POI. The ginseng extract ginsenoside Rg1 has a protective effect on ovarian function and can delay ovarian aging through its antioxidant, anti-aging, anti-inflammatory and estrogenic activities (227). Rg1, a ginseng extract, not only can inhibit GCs apoptosis and promote the proliferation and growth of GCs by up-regulating the expression of FSHR and activating the FSHR-PI3K-AKT pathway to protect ovarian function in rat model (228) and thus attenuate POI. D-gal was able to successfully induce POF models by impairing follicular development, decreasing the apoptosis rate of granulosa cells, lowering E2 levels, and increasing the level of FSH in vivo. Studies have shown that D-gal can induce oxidative stress directly in the body, and that increased levels of galactose in women’s blood lead to a decrease in the number of ovarian follicles and damage to oocytes and ovarian granulosa cells, thus leading to POI (229, 230). Rg1 can also can postpone D-galactose (D-gal)-induced POI through the PI3K-Akt-mTOR autophagy signaling pathway mouse model of ovarian senescence, effectively facilitating the secretion of estrogen and luteinizing hormone, and increasing the expression of SOD and CAT, thereby alleviating ovarian oxidative damage in mice model (231, 232) and delaying the development of POI. Aside from this, Tang’s rat experiments demonstrated that ginsenoside Rb1 has a protective effect on ovarian function by the mechanism that the estrogenic activity of ginsenoside can regulate ovarian function, reverse disorders in sex hormone levels, reduce apoptosis of ovarian granulosa cells, and delay ovarian aging (233). We have discovered that CUR (the active ingredient of turmeric rhizome) possesses a positive effect on POI patients by potently inhibiting D-gal-induced 0S, apoptosis and ovarian damage through various mechanisms including Nrf2/HO-1 and PI3K/Akt signaling pathways (234) (193). Compared with resveratrol and vitamin C, the antioxidant capacity of CUR is superior (234).
Relevant animal and clinical studies have confirmed that both the above natural compounds and herbal extracts have a good safety profile in animal models and human models (235–239).
Numerous studies have recently reported promising therapeutic effects of MSC for POI treatment (240) (241–243). MSCs can secrete a variety of factors, including cytokines and exosomes, that lead to POI recovery through several mechanisms, involving reduction of apoptosis and inflammation and induction of angiogenesis (244). Stem cell transplantation is a treatment that has the ability to repair damaged ovarian tissues, restore ovarian elasticity, and reduce ovarian tissue fibrosis. In recent years, MSCs for the treatment of POI have become a hot research topic in this field. The mechanisms include MSC homing, promotion of cell proliferation, inhibition of apoptosis, differentiation, anti-inflammatory and immunomodulation, secretion of various cytokines, regulation of genes, and regulation of cell autophagy (245). The main MSCs used for the treatment of POI are bone marrow mesenchymal stem cells, adipose mesenchymal stem cells (ADMSCs), placental mesenchymal stem cells (PMSCs), amniotic mesenchymal stem cells (AMSCs), umbilical cord mesenchymal stem cells (UCMSCs), amniotic fluid mesenchymal stem cells (AFMSCs), and menstrual stem cells (MenSCs). We have discovered that MSCs can play an anti-OS role by secreting IL-6, VEGF and other related cytokines and exosomes, activating NQO1/MAPK and other related pathways, increasing the production of antioxidant-acting enzymes, and prohibiting ROS production, which in turn improves mice’ ovarian function (246). For example, fetal mesenchymal stem cells (fMSCs) prominently reduce oxidative damage, increase oxidative protection, improve anti-apoptotic effects, and inhibit apoptotic gene expression in vivo and vitro (247), protecting ovarian function in POI patients. Exosomes have been recognized to have a vital role in the treatment of POI by MSC, which are carriers of intercellular signaling and drug therapy, and miRNA is a class of single-stranded non-coding RNA in exosomes. Several studies have found that human mesenchymal stem cells derived from bone marrow (BMSC)is at work in the fight against oxidative stress. Proteomic analysis revealed that various antioxidant mediators secreted by BMSCs, such as cyclophilin A, cyclophilin B, thioxapurine, DJ-1, heat shock protein 27, and peroxycin-1, exhibited significant antioxidant stress effects. Proteomic analysis revealed that various antioxidant mediators secreted by BMSCs, such as cyclophilin A, cyclophilin B, thioxapurine, DJ-1, heat shock protein 27, and peroxycin-1, exhibited significant antioxidant stress effects (248). Insulin-like growth factor 1 (IGF1) released from BMSCs also has potent antioxidant capacity (249). On top of that, BMSC transplantation affects SOD activity and malondialdehyde content by decreasing cyclin-dependent kinase inhibitor 2A(P16) expression and increasing proliferating cell nuclear antigen, so as to improve the morphology and function of the ovary (250). Exosomes and miRNAs have the capacity to regulate the expression of downstream target genes to inhibit apoptosis of various cells in the ovary, so as to improve ovarian function (251) and slow down the progression of POI. For instance, human umbilical cord MSC-derived exosome microRNA-17-5P (miR-17-5p) can reduce ROS accumulation by inhibiting SIRT7 expression to combat OS and thus improve POI (252).
Although MSC has obvious therapeutic effects in animal models of POI, its clinical application still suffers from the issues of insufficient cell source, immunogenicity, passaging culture, ethics, safety and efficacy, which need to be evaluated more comprehensively through more in-depth clinical studies (245, 253).
Endogenous antioxidant systems exist within the body and cells, including antioxidant enzymes such as SOD, GPX, glutathione sulfhydryltransferase (GST), and CAT, in addition to GSH as a scavenger of ROS (205, 223). Superoxide anions in ROS form a spontaneous conversion to H2O2 catalyzed by SOD, and then CAT and GPX further convert H2O2 to O2 and H2O, which exerts antioxidant stress effects in POI patients (254). In addition, phase II detoxification enzymes have defensive effects against oxidative damage caused by exogenous chemical exposure, mainly including HO-1, HQO1 and γ-glutamylcysteine ligase (γ-GCL), which can moderate POI to some extent by counteracting OS. HO-1 is a classical phase II detoxifying enzyme involved in preventing the generation of more active hydroxyl groups from H2O2, and also catalyzes the production of biliverdin from heme, of which the latter can form bilirubin, and biliverdin and bilirubin are powerful free radical scavengers in the body. It has been proved in mouse that γ-GCLase catalyzes the formation of GSH and is the most critical rate-limiting enzyme in GSH biosynthesis (254). Furthermore, OS results in ER stress, and Ufm1-specific ligase 1 (UFL1) alleviates cisplatin-induced apoptosis in ovarian GCs by attenuating ER stress, thereby relieving POI to some extent (254).The key to POI is the reduction in the quantity and quality of oocytes, which results in symptoms such as hot flashes, excessive sweating, vaginal dryness, and even causes serious consequences such as infertility.
The key to POI is the reduction in the quantity and quality of oocytes, which results in symptoms such as hot flashes, excessive sweating, vaginal dryness. Accordingly, it even causes serious consequences such as infertility. The role of apoptosis, abnormal meiosis, decreased mtDNA numbers, and ovarian microenvironment in POI is well known, but the mechanism of action is unclear. This review proposes that OS can mediate changes in genetic material, signaling pathways, transcription factors and ovarian microenvironment to accelerate oocyte and GCs senescence. Antioxidants are capable of counteracting the adverse effects of OS in the ovary and exhibit good therapeutic effects in the prevention and treatment of POI. We focus on exploring the mechanism of action of antioxidants, mesenchymal stem cells, and bioenzymes against OS in POI. Therefore, studying the relationship between OS and POI will not only help us understand the mechanism of action, but will also provide new insights into the protection of the female reproductive system and infertility.
The data of this article can be obtained with the permission of the corresponding author.
Y-QS put forward the article idea and wrote the article. X-TZ wrote articles. S-NZ wrote articles. Y-FM translated and polished the manuscript. Y-HH revised the article. YJ provided technical support for the article. Y-HZ revised the article and provided financial support. All authors contributed to the article and approved the submitted version.
The National Natural Science Foundation of China (grant no. 82074259), the Project of Cultivation Project of Outstanding Youth Fund of Heilongjiang University of Traditional Chinese Medicine (grant no. 2018jc02), “Outstanding Young Academic Leaders” Scientific Research Project of Heilongjiang University of Chinese Medicine; Special project on the popularization of traditional Chinese medicine classics in Heilongjiang Province (ZYW2022-100).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. POI, Webber L, Davies M, Anderson R, Bartlett J, Braat D, et al. ESHRE Guideline: management of women with premature ovarian insufficiency. Hum Reprod (2016) 31(5):926–37. doi: 10.1093/humrep/dew027
2. Christin-Maitre S, Givony M, Albarel F, Bachelot A, Bidet M, Blanc JV, et al. Position statement on the diagnosis and management of premature/primary ovarian insufficiency (except Turner Syndrome). In: Annales d'Endocrinologie. France: Elsevier (2021).
3. Coulam CB, Adamson SC, Annegers JF. Incidence of premature ovarian failure. Obstetrics gynecology (1986) 67(4):604–6.
4. Mishra GD, Pandeya N, Dobson AJ, Chung HF, Anderson D, Kuh D, et al. Early menarche, nulliparity and the risk for premature and early natural menopause. Hum Reprod (2017) 32(3):679–86. doi: 10.1093/humrep/dew350
5. Panay N, Anderson RA, Nappi RE, Vincent AJ, Vujovic S, Webber L, et al. Premature ovarian insufficiency: an international menopause society white paper. Climacteric (2020) 23(5):426–46. doi: 10.1080/13697137.2020.1804547
6. Lambrinoudaki I, Paschou SA, Lumsden MA, Faubion S, Makrakis E, Kalantaridou S, et al. Premature ovarian insufficiency: a toolkit for the primary care physician. Maturitas (2021) 147:53–63. doi: 10.1016/j.maturitas.2020.11.004
7. Taún JJ. Aetiology of age-associated aneuploidy: a mechanism based on the ‘free radical theory of ageing’. MHR: Basic Sci Reprod Med (1995) 1(4):163–5. doi: 10.1093/humrep/10.6.1563
8. Shi L, Zhang J, Lai Z, Tian Y, Fang L, Wu M, et al. Long-term moderate oxidative stress decreased ovarian reproductive function by reducing follicle quality and progesterone production. PloS One (2016) 11(9):e0162194. doi: 10.1371/journal.pone.0162194
9. Seli E, Wang T, Horvath TL. Mitochondrial unfolded protein response: a stress response with implications for fertility and reproductive aging. Fertility sterility (2019) 111(2):197–204. doi: 10.1016/j.fertnstert.2018.11.048
10. Lushchak VI. Free radicals, reactive oxygen species, oxidative stress and its classification. Chemico-biological Interact (2014) 224:164–75. doi: 10.1016/j.cbi.2014.10.016
11. Zorov DB, Juhaszova M, Sollott SJ. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol Rev (2014) 94(3):909–50. doi: 10.1152/physrev.00026.2013
12. Zhang J, Wang X, Vikash V, Ye Q, Wu D, Liu Y, et al. ROS and ROS-mediated cellular signaling. Oxid Med Cell Longevity (2016) 2016. doi: 10.1155/2016/4350965
13. Sahoo BM, Banik BK, Borah P, Jain A. Reactive oxygen species (ROS): key components in cancer therapies. Anti-Cancer Agents Medicinal Chem (Formerly Curr Medicinal Chemistry-Anti-Cancer Agents) (2022) 22(2):215–22. doi: 10.2174/1871520621666210608095512
14. Scialò F, Fernández-Ayala DJ, Sanz A. Role of mitochondrial reverse electron transport in ROS signaling: potential roles in health and disease. Front Physiol (2017) 8:428. doi: 10.3389/fphys.2017.00428
15. El-Kenawi A, Ruffell B. Inflammation, ROS, and mutagenesis. Cancer Cell (2017) 32(6):727–9. doi: 10.1016/j.ccell.2017.11.015
16. Sun Y, Lu Y, Saredy J, Wang X, Drummer C Iv, Shao Y, et al. ROS systems are a new integrated network for sensing homeostasis and alarming stresses in organelle metabolic processes. Redox Biol (2020) 37:101696. doi: 10.1016/j.redox.2020.101696
17. Lin J, Epel E. Stress and telomere shortening: Insights from cellular mechanisms. Ageing Res Rev (2022) 73:101507. doi: 10.1016/j.arr.2021.101507
18. Xie T, Ye W, Liu J, Zhou L, Song Y. The emerging key role of Klotho in the hypothalamus–pituitary–ovarian axis. Reprod Sci (2021) 28(2):322–31. doi: 10.1007/s43032-020-00277-5
19. Fukai T, Ushio-Fukai M. Superoxide dismutases: role in redox signaling, vascular function, and diseases. Antioxidants Redox Signaling (2011) 15(6):1583–606. doi: 10.1089/ars.2011.3999
20. Jiao X, Liu N, Xu Y, Qiao H. Perfluorononanoic acid impedes mouse oocyte maturation by inducing mitochondrial dysfunction and oxidative stress. Reprod Toxicol (2021) 104:58–67. doi: 10.1016/j.reprotox.2021.07.002
21. Cao Y, Shen M, Jiang Y, Sun SC, Liu H. Melatonin reduces oxidative damage in mouse granulosa cells via restraining JNK-dependent autophagy. Reproduction (2018) 155(3):307–19. doi: 10.1530/REP-18-0002
22. Schieber M, Chandel NS. ROS function in redox signaling and oxidative stress. Curr Biol (2014) 24(10):R453–62. doi: 10.1016/j.cub.2014.03.034
23. Murphy AA, Palinski W, Rankin S, Morales AJ, Parthasarathy S. Evidence for oxidatively modified lipid-protein complexes in endometrium and endometriosis. Fertility sterility (1998) 69(6):1092–4. doi: 10.1016/S0015-0282(98)00087-9
24. Jiao X, Ke H, Qin Y, Chen ZJ. Molecular genetics of premature ovarian insufficiency. Trends Endocrinol Metab (2018) 29(11):795–807. doi: 10.1016/j.tem.2018.07.002
25. Peng H, Zeng L, Zhu L, Luo S, Xu L, Zeng , et al. Zuogui Pills inhibit mitochondria-dependent apoptosis of follicles in a rat model of premature ovarian failure. J ethnopharmacology (2019) 238:111855. doi: 10.1016/j.jep.2019.111855
26. Yang L, Zhao Y, Xu H, Ma Y, Wang L, Ma J, et al. Network pharmacology-based prediction and verification of the potential mechanisms of He’s Yangchao formula against diminished ovarian reserve. Evidence-Based Complementary Altern Med (2022) 2022. doi: 10.1155/2022/8361808
27. Zhang S, Tan R, Pan R, Xiong J, Tian Y, Wu J, et al. Association of perfluoroalkyl and polyfluoroalkyl substances with premature ovarian insufficiency in Chinese women. J Clin Endocrinol Metab (2018) 103(7):2543–51. doi: 10.1210/jc.2017-02783
28. Feng X, Wang X, Cao X, Xia Y, Zhou R, Chen L. Chronic exposure of female mice to an environmental level of perfluorooctane sulfonate suppresses estrogen synthesis through reduced histone H3K14 acetylation of the StAR promoter leading to deficits in follicular development and ovulation. Toxicological Sci (2015) 148(2):368–79. doi: 10.1093/toxsci/kfv197
29. Bianco G, Fontanella B, Severino L, Quaroni A, Autore G, Marzocco S. Nivalenol and deoxynivalenol affect rat intestinal epithelial cells: a concentration related study. PloS One (2012) 7(12):e52051. doi: 10.1371/journal.pone.0052051
30. Wang Y, Xing CH, Zhang HL, Pan ZN, Sun SC. Exposure to nivalenol declines mouse oocyte quality via inducing oxidative stress-related apoptosis and DNA damage. Biol Reprod (2021) 105(6):1474–83. doi: 10.1093/biolre/ioab171
31. Aguiar PH, Furtado C, Repolês BM, Ribeiro GA, Mendes IC, Peloso EF, et al. Oxidative stress and DNA lesions: the role of 8-oxoguanine lesions in Trypanosoma cruzi cell viability. PloS Negl Trop Dis (2013) 7(6):e2279. doi: 10.1371/journal.pntd.0002279
32. Srinivas US, Tan BWQ, Vellayappan BA, Jeyasekharan AD. ROS and the DNA damage response in cancer. Redox Biol (2019) 25:101084. doi: 10.1016/j.redox.2018.101084
33. Sha C, Chen L, Lin L, Li T, Wei H, Yang M, et al. TRDMT1 participates in the DNA damage repair of granulosa cells in premature ovarian failure. Aging (Albany NY) (2021) 13(11):15193. doi: 10.18632/aging.203080
34. Xiong Y, Yin Q, Jin E, Chen H, He S. Selenium attenuates chronic heat stress-induced apoptosis via the inhibition of endoplasmic reticulum stress in mouse granulosa cells. Molecules (2020) 25(3):557. doi: 10.3390/molecules25030557
35. Wang X, Wang L, Xiang W. Mechanisms of ovarian aging in women: a review. J Ovarian Res (2023) 16(1):1–22. doi: 10.1186/s13048-023-01151-z
36. Ganesan S, Keating AF. Phosphoramide mustard exposure induces DNA adduct formation and the DNA damage repair response in rat ovarian granulosa cells. Toxicol Appl Pharmacol (2015) 282(3):252–8. doi: 10.1016/j.taap.2014.11.017
37. Salehi F, Behboudi H, Kavoosi G, Ardestani SK. Oxidative DNA damage induced by ROS-modulating agents with the ability to target DNA: A comparison of the biological characteristics of citrus pectin and apple pectin. Sci Rep (2018) 8(1):1–16. doi: 10.1038/s41598-018-32308-2
38. Meng Y, Chen CW, Yung MMH, Sun W, Sun J, Li Z, et al. DUOXA1-mediated ROS production promotes cisplatin resistance by activating ATR-Chk1 pathway in ovarian cancer. Cancer Lett (2018) 428:104–16. doi: 10.1016/j.canlet.2018.04.029
39. Graindorge D, Martineau S, Machon C, Arnoux P, Guitton J, Francesconi S, et al. Singlet oxygen-mediated oxidation during UVA radiation alters the dynamic of genomic DNA replication. PloS One (2015) 10(10):e0140645. doi: 10.1371/journal.pone.0140645
40. Lu Q, Sun Y, Ares I, Anadón A, Martínez M, Martínez-Larrañaga MR, et al. Deltamethrin toxicity: A review of oxidative stress and metabolism. Environ Res (2019) 170:260–81. doi: 10.1016/j.envres.2018.12.045
41. Sreelekha K, Chandrasekhar L, Kartha HS, Ravindran R, Juliet S, Ajithkumar KG, et al. Ultrastructural analysis of oocytes of Rhipicephalus (Boophilus) annulatus during postengorgement period as a tool to evaluate the cytotoxic effects of amitraz and deltamethrin on the germinative cells. Veterinary Parasitol (2017) 247:113–20. doi: 10.1016/j.vetpar.2017.10.006
42. Jia Z, Zhang JW, Zhou D, Xu DQ, Feng XZ. Deltamethrin exposure induces oxidative stress and affects meiotic maturation in mouse oocyte. Chemosphere (2019) 223:704–13. doi: 10.1016/j.chemosphere.2019.02.092
43. Sreerangaraja Urs DB, Wu WH, Komrskova K, Postlerova P, Lin YF, Tzeng CR, et al. Mitochondrial function in modulating human granulosa cell steroidogenesis and female fertility. Int J Mol Sci (2020) 21(10):3592. doi: 10.3390/ijms21103592
44. Lemasters JJ. Selective mitochondrial autophagy, or mitophagy, as a targeted defense against oxidative stress, mitochondrial dysfunction, and aging. Rejuvenation Res (2005) 8(1):3–5. doi: 10.1089/rej.2005.8.3
45. Bonomi M, Somigliana E, Cacciatore C, Busnelli M, Rossetti R, Bonetti S, et al. Blood cell mitochondrial DNA content and premature ovarian aging. PLoS One (2012) 7(8):e42423. doi: 10.1371/journal.pone.0042423
46. Miao C, Zhao Y, Chen Y, Wang R, Ren N, Chen B, et al. Investigation of He’s Yang Chao recipe against oxidative stress-related mitophagy and pyroptosis to improve ovarian function. Front Endocrinol (2023) 14:1077315. doi: 10.3389/fendo.2023.1077315
47. Bakhshalizadeh S, Hock DH, Siddall NA, Kline BL, Sreenivasan R, Bell KM, et al. Deficiency of the mitochondrial ribosomal subunit, MRPL50, causes autosomal recessive syndromic premature ovarian insufficiency. Hum Genet (2023) 142(7):879–907. doi: 10.1007/s00439-023-02563-z
48. Wang L, Tang J, Wang L, Tan F, Song H, Zhou J, et al. Oxidative stress in oocyte aging and female reproduction. J Cell Physiol (2021) 236(12):7966–83. doi: 10.1002/jcp.30468
49. Xing C, Wang Y, Liu JC, Pan ZN, Zhang HL, Sun SC, et al. Melatonin reverses mitochondria dysfunction and oxidative stress-induced apoptosis of Sudan I-exposed mouse oocytes. Ecotoxicology Environ Saf (2021) 225:112783. doi: 10.1016/j.ecoenv.2021.112783
50. Shkolnik K, Tadmor A, Ben-Dor S, Nevo N, Galiani D, Dekel N. Reactive oxygen species are indispensable in ovulation. Proc Natl Acad Sci (2011) 108(4):1462–7. doi: 10.1073/pnas.1017213108
51. Hou X, Zhu S, Zhang H, Li C, Qiu D, Ge J, et al. Mitofusin1 in oocyte is essential for female fertility. Redox Biol (2019) 21:101110. doi: 10.1016/j.redox.2019.101110
52. Kudryavtseva AV, Krasnov GS, Dmitriev AA, Alekseev BY, Kardymon OL, Sadritdinova AF, et al. Mitochondrial dysfunction and oxidative stress in aging and cancer. Oncotarget (2016) 7(29):44879. doi: 10.18632/oncotarget.9821
53. Ding Y, Xia BH, Zhuo GC, Zhang CJ, Leng JH. Premature ovarian insufficiency may be associated with the mutations in mitochondrial tRNA genes. Endocrine J (2019) 66(1):81–8. doi: 10.1507/endocrj.EJ18-0308
54. Xu H, Mao X, Nie Z, Li Y. Oxr1a prevents the premature ovarian failure by regulating oxidative stress and mitochondrial function in zebrafish. Free Radical Biol Med (2023) 203:102–13. doi: 10.1016/j.freeradbiomed.2023.04.002
55. Zhang X, Wu XQ, Lu S, Guo YL, Ma X. Deficit of mitochondria-derived ATP during oxidative stress impairs mouse MII oocyte spindles. Cell Res (2006) 16(10):841–50. doi: 10.1038/sj.cr.7310095
56. Jiao X, Ding Z, Meng F, Zhang X, Wang Y, Chen F, et al. The toxic effects of Fluorene-9-bisphenol on porcine oocyte in vitro maturation. Environ Toxicol (2020) 35(2):152–8. doi: 10.1002/tox.22851
57. Roca-Agujetas V, de Dios C, Lestón L, Marí M, Morales A, Colell A, et al. Recent insights into the mitochondrial role in autophagy and its regulation by oxidative stress. Oxid Med Cell Longevity (2019) 2019. doi: 10.1155/2019/3809308
58. Li L. Oxidative stress-induced autophagy in porcine ovarian granulosa cells and its effect on apoptosis. China: Nanjing Agricultural University (2016).
59. Shaeib F, Banerjee J, Maitra D, Diamond MP, Abu-Soud HM. Impact of hydrogen peroxide-driven Fenton reaction on mouse oocyte quality. Free Radical Biol Med (2013) 58:154–9. doi: 10.1016/j.freeradbiomed.2012.12.007
60. Han L, Wang H, Li L, Li X, Ge J, Reiter RJ. Melatonin protects against maternal obesity-associated oxidative stress and meiotic defects in oocytes via the SIRT 3-SOD 2-dependent pathway. J Pineal Res (2017) 63(3):e12431. doi: 10.1111/jpi.12431
61. Jensen TK, Andersen LB, Kyhl HB, Nielsen F, Christesen HT, Grandjean P, et al. Association between perfluorinated compound exposure and miscarriage in Danish pregnant women. PloS One (2015) 10(4):e0123496. doi: 10.1371/journal.pone.0123496
63. Luo S, Ruan X, Mueck AO. The effect of Kuntai capsule on ovarian function in cisplatin-induced premature ovarian insufficiency rats. Front Endocrinol (2023) 13:1097165. doi: 10.3389/fendo.2022.1097165
64. Miao Y, Zhou C, Bai Q, Cui Z, ShiYang X, Lu Y, et al. The protective role of melatonin in porcine oocyte meiotic failure caused by the exposure to benzo (a) pyrene. Hum Reprod (2018) 33(1):116–27. doi: 10.1093/humrep/dex331
65. Kaeberlein M, Rabinovitch PS, Martin GM. Healthy aging: the ultimate preventative medicine. Science (2015) 350(6265):1191–3. doi: 10.1126/science.aad3267
66. Kalmbach KH, Antunes DM, Kohlrausch F, Keefe DL. Telomeres and female reproductive aging. in Seminars in reproductive medicine. United States: Thieme Medical Publishers (2015).
67. Vasilopoulos E, Fragkiadaki P, Kalliora C, Fragou D, Docea AO, Vakonaki E, et al. The association of female and male infertility with telomere length. Int J Mol Med (2019) 44(2):375–89. doi: 10.3892/ijmm.2019.4225
68. Kordowitzki P. Oxidative stress induces telomere dysfunction and shortening in human oocytes of advanced age donors. Cells (2021) 10(8):1866. doi: 10.3390/cells10081866
69. Butts S, Riethman H, Ratcliffe S, Shaunik A, Coutifaris C, Barnhart K. Correlation of telomere length and telomerase activity with occult ovarian insufficiency. J Clin Endocrinol Metab (2009) 94(12):4835–43. doi: 10.1210/jc.2008-2269
70. Xu X, Chen X, Zhang X, Liu Y, Wang Z, Wang P, et al. Impaired telomere length and telomerase activity in peripheral blood leukocytes and granulosa cells in patients with biochemical primary ovarian insufficiency. Hum Reprod (2017) 32(1):201–7. doi: 10.1097/01.ogx.0000513118.72411.50
71. Barnes RP, Fouquerel E, Opresko PL. The impact of oxidative DNA damage and stress on telomere homeostasis. Mech Ageing Dev (2019) 177:37–45. doi: 10.1016/j.mad.2018.03.013
72. Jiang HL, Cao LQ, Chen HY. Protective effects ROS up-regulation on premature ovarian failure by suppressing ROS-TERT signal pathway. Eur Rev Med Pharmacol Sci (2018) 22(19):6198–204. doi: 10.26355/eurrev_201810_16025
73. Robinson LG Jr, Pimentel R, Wang F, Kramer YG, Gonullu DC, Agarwal S, et al. Impaired reproductive function and fertility preservation in a woman with a dyskeratosis congenita. J Assisted Reprod Genet (2020) 37(5):1221–5. doi: 10.1007/s10815-020-01758-x
74. Wang Y, Teng X, Liu J. Research progress on the effect of traditional Chinese medicine on signal pathway related to premature ovarian insufficiency. Evidence-Based Complementary Altern Med (eCAM) (2022) 2022. doi: 10.1155/2022/7012978
75. John GB, Shidler MJ, Besmer P, Castrillon DH. Kit signaling via PI3K promotes ovarian follicle maturation but is dispensable for primordial follicle activation. Dev Biol (2009) 331(2):292–9. doi: 10.1016/j.ydbio.2009.05.546
76. Wang F, Xiao KZ, Lin CQ, Wang ZC. Regulatory effects of PI3K signaling on HPG axis in mammalians. Chin J Cell Biol (2013) 35:1068–72. doi: 10.1016/j.apsb.2019.06.005
77. Pelosi E, Omari S, Michel M, Ding J, Amano T, Forabosco A, et al. Constitutively active Foxo3 in oocytes preserves ovarian reserve in mice. Nat Commun (2013) 4(1):1–7. doi: 10.1038/ncomms2861
78. Zheng S, Ma M, Chen Y, Li M. Effects of quercetin on ovarian function and regulation of the ovarian PI3K/Akt/FoxO3a signalling pathway and oxidative stress in a rat model of cyclophosphamide-induced premature ovarian failure. Basic Clin Pharmacol Toxicol (2022) 130(2):240–53. doi: 10.1111/bcpt.13696
79. Liu J, Wu DC, Qu LH, Liao HQ, Li MX. The role of mTOR in ovarian neoplasms, polycystic ovary syndrome, and ovarian aging. Clin Anat (2018) 31(6):891–8. doi: 10.1002/ca.23211
80. Cheng Y, Kim J, Li XX, Hsueh AJ. Promotion of ovarian follicle growth following mTOR activation: synergistic effects of AKT stimulators. PloS One (2015) 10(2):e0117769. doi: 10.1371/journal.pone.0117769
81. Adhikari D, Liu K. mTOR signaling in the control of activation of primordial follicles. United States: Taylor & Francis (2010) p. 1673–4.
82. Zhao D. Exploring the protective mechanism of Erxian Tang against cisplatin-injured rat ovarian granulosa cells based on FOXO3a transcription factor. China: Peking University (2017).
83. Krishna M, Narang H. The complexity of mitogen-activated protein kinases (MAPKs) made simple. Cell Mol Life Sci (2008) 65(22):3525–44. doi: 10.1007/s00018-008-8170-7
84. Bogoyevitch MA, Kobe B. Uses for JNK: the many and varied substrates of the c-Jun N-terminal kinases. Microbiol Mol Biol Rev (2006) 70(4):1061–95. doi: 10.1128/MMBR.00025-06
85. Xu S, Yang Y, Han S, Wu Z. ZIP1 and zinc inhibits fluoride-induced apoptosis in MC3T3-E1 cells. Biol Trace element Res (2014) 159(1):399–409. doi: 10.1007/s12011-014-9935-5
86. Mizukami Y, Yoshioka K, Morimoto S, Yoshida K. A novel mechanism of JNK1 activation: nuclear translocation and activation of JNK1 during ischemia and reperfusion. J Biol Chem (1997) 272(26):16657–62. doi: 10.1074/jbc.272.26.16657
87. Zhang P, Miller BS, Rosenzweig SA, Bhat NR. Activation of C-jun N-terminal kinase/stress-activated protein kinase in primary glial cultures. J Neurosci Res (1996) 46(1):114–21. doi: 10.1002/(SICI)1097-4547(19961001)46:1<114::AID-JNR14>3.0.CO;2-5
88. Kops GJ, Dansen TB, Polderman PE, Saarloos I, Wirtz KW, Coffer PJ, et al. Forkhead transcription factor FOXO3a protects quiescent cells from oxidative stress. Nature (2002) 419(6904):316–21. doi: 10.1038/nature01036
89. Sun J, Guo Y, Fan Y, Wang Q, Zhang Q, Lai D. Decreased expression of IDH1 by chronic unpredictable stress suppresses proliferation and accelerates senescence of granulosa cells through ROS activated MAPK signaling pathways. Free Radical Biol Med (2021) 169:122–36. doi: 10.1016/j.freeradbiomed.2021.04.016
90. Stringer JM, Alesi LR, Winship AL, Hutt KJ. Beyond apoptosis: evidence of other regulated cell death pathways in the ovary throughout development and life. Hum Reprod Update (2023) 29(4):434–56. doi: 10.1093/humupd/dmad005
91. Winship AL, Stringer JM, Liew SH, Hutt KJ. The importance of DNA repair for maintaining oocyte quality in response to anti-cancer treatments, environmental toxins and maternal ageing. Hum Reprod Update (2018) 24(2):119–34. doi: 10.1093/humupd/dmy002
92. Jin J, Jin L, Lim SW, Yang CW. Klotho deficiency aggravates tacrolimus-induced renal injury via the phosphatidylinositol 3-kinase-Akt-forkhead box protein O pathway. Am J Nephrol (2016) 43(5):357–65. doi: 10.1159/000446447
93. Yu M, Li H, Liu Q, Liu F, Tang L, Li C, et al. Nuclear factor p65 interacts with Keap1 to repress the Nrf2-ARE pathway. Cell signalling (2011) 23(5):883–92. doi: 10.1016/j.cellsig.2011.01.014
94. Lu MC, Ji JA, Jiang ZY, You QD. The Keap1–Nrf2–ARE pathway as a potential preventive and therapeutic target: an update. Medicinal Res Rev (2016) 36(5):924–63. doi: 10.1002/med.21396
95. Zhang Q, Liu J, Duan H, Li R, Peng W, Wu C. Activation of Nrf2/HO-1 signaling: An important molecular mechanism of herbal medicine in the treatment of atherosclerosis via the protection of vascular endothelial cells from oxidative stress. J advanced Res (2021) 34:43–63. doi: 10.1016/j.jare.2021.06.023
96. Younis NS, Abduldaium MS, Mohamed ME. Protective effect of geraniol on oxidative, inflammatory and apoptotic alterations in isoproterenol-induced cardiotoxicity: role of the Keap1/Nrf2/HO-1 and PI3K/Akt/mTOR pathways. Antioxidants (2020) 9(10):977. doi: 10.3390/antiox9100977
97. Chen Y, Zhao R, Li X, Luan YP, Xing LW, Zhang XJ, et al. Preventive electroacupuncture alleviates oxidative stress and inflammation via Keap1/Nrf2/HO-1 pathway in rats with cyclophosphamide-induced premature ovarian insufficiency. BioMed Res Int (2022) 2022. doi: 10.1155/2022/6718592
98. Cai ZY, Fu MD, Liu K, Duan XC. Therapeutic effect of Keap1-Nrf2-ARE pathway-related drugs on age-related eye diseases through anti-oxidative stress. Int J Ophthalmol (2021) 14(8):1260. doi: 10.18240/ijo.2021.08.19
99. Pan H, Wang H, Wang X, Zhu L, Mao L. The absence of nrf2 enhances nf-b-dependent inflammation following scratch injury in mouse primary cultured astrocytes. Mediators Inflammation (2012) 2012. doi: 10.1155/2012/217580
100. Abd El-Twab SM, Hussein OE, Hozayen WG, Bin-Jumah M, Mahmoud AM. Chicoric acid prevents methotrexate-induced kidney injury by suppressing NF-κB/NLRP3 inflammasome activation and up-regulating Nrf2/ARE/HO-1 signaling. Inflammation Res (2019) 68(6):511–23. doi: 10.1007/s00011-019-01241-z
101. Chen S. Clinical study on improving early-onset ovarian insufficiency with kidney tonifying and blood activating soup and the mechanism of regulating Keap1/Nrf2/ARE signaling pathway in model mice. China: Nanjing University of Traditional Chinese Medicine (2021).
102. Romero A, San Hipólito-Luengo Á, Villalobos LA, Vallejo S, Valencia I, Michalska P, et al. The angiotensin-(1-7)/Mas receptor axis protects from endothelial cell senescence via klotho and Nrf2 activation. Aging Cell (2019) 18(3):e12913. doi: 10.1111/acel.12913
103. Shang Z, Fan M, Zhang J, Wang Z, Jiang S, Li W. Red ginseng improves D-galactose-induced premature ovarian failure in mice based on network pharmacology. Int J Mol Sci (2023) 24(9):8210. doi: 10.3390/ijms24098210
104. Takahashi A, Masuda A, Sun M, Centonze VE, Herman B. Oxidative stress-induced apoptosis is associated with alterations in mitochondrial caspase activity and Bcl-2-dependent alterations in mitochondrial pH (pHm). Brain Res Bull (2004) 62(6):497–504. doi: 10.1016/j.brainresbull.2003.07.009
105. Dharshini LC, Vishnupriya S, Sakthivel KM, Rasmi RR. Oxidative stress responsive transcription factors in cellular signalling transduction mechanisms. Cell Signalling (2020) 72:109670. doi: 10.1016/j.cellsig.2020.109670
106. Yang H, Xie Y, Yang D, Ren D. Oxidative stress-induced apoptosis in granulosa cells involves JNK, p53 and Puma. Oncotarget (2017) 8(15):25310. doi: 10.18632/oncotarget.15813
107. Yang B, Chen Q. Cross-talk between oxidative stress and m6A RNA methylation in cancer Vol. 2021. United States: Oxidative Medicine and Cellular Longevity (2021).
108. Yu B, Sun ZJ. Apoptosis-related genes bcl-2, bax, bad and breast cancer. Chin J Basic Clin Gen Surg (2007) 06:739–42. doi: 10.4149/endo_2013_04_189
109. Zhu R, Guo XB, Yang MY, Chen XZ, Hu YD, Yang Z, et al. Effects of tonifying kidney and invigorating blood soup on Bax/Bcl-2 and cleaved Caspase-3 protein expression in nude mice with bone metastasis model of breast cancer. Chin J Exp Formulary (2023) 29(08):133–41. doi: 10.13422/j.cnki.syfjx.20222426
110. Xu H, Xia Y, Qin J, Xu J, Li C, Wang Y. Effects of low intensity pulsed ultrasound on expression of B-cell lymphoma-2 and BCL2-Associated X in premature ovarian failure mice induced by 4-vinylcyclohexene diepoxide. Reprod Biol Endocrinol (2021) 19(1):1–11. doi: 10.1186/s12958-021-00799-w
111. Cai L, Zong DK, Tong GQ, Li L. Apoptotic mechanism of premature ovarian failure and rescue effect of Traditional Chinese Medicine: a review. J Traditional Chin Med (2021) 41(3):491. doi: 10.19852/j.cnki.jtcm.2021.03.017
112. Zhang JB, Lang QL, Zhu JL. Role of the BCL-2 gene family in regulating apoptosis. Chin J Traditional Chin Med (2005) 3(003):16–8.
113. Edlich F. BCL-2 proteins and apoptosis: Recent insights and unknowns. Biochem Biophys Res Commun (2018) 500(1):26–34. doi: 10.1016/j.bbrc.2017.06.190
114. Willis S, Day CL, Hinds MG, Huang DC. The Bcl-2-regulated apoptotic pathway. J Cell Sci (2003) 116(20):4053–6. doi: 10.1242/jcs.00754
115. Wu J, Zheng GT, Zhang L, Tang ZQ, Wang L, Su SH, et al. Expression of apoptosis-regulating proteins Bcl-2, Bax, and p53 in ovarian granulosa cells with polycystic ovary syndrome and their significance. New Med (2019) 50(07):536–9.
116. Korsmeyer SJ, Shutter JR, Veis DJ, Merry DE, Oltvai ZN. Bcl-2/Bax: a rheostat that regulates an anti-oxidant pathway and cell death. In: Seminars in cancer biology (1993).
117. Cekanova M, Fernando RI, Siriwardhana N, Sukhthankar M, Parra C, Woraratphoka J, et al. BCL-2 family protein, BAD is down-regulated in breast cancer and inhibits cell invasion. Exp Cell Res (2015) 331(1):1–10. doi: 10.1016/j.yexcr.2014.11.016
118. López-Diazguerrero NE, López-Araiza H, Conde-Perezprina JC, Bucio L, Cárdenas-Aguayo MC, Ventura JL, et al. Bcl-2 protects against oxidative stress while inducing premature senescence. Free Radical Biol Med (2006) 40(7):1161–9. doi: 10.1016/j.freeradbiomed.2005.11.002
119. Spitz AZ, Gavathiotis E. Physiological and pharmacological modulation of BAX. Trends Pharmacol Sci (2021) 43(3):206–20. doi: 10.1016/j.tips.2021.11.001
120. Cai X, Fu H, Wang Y, Liu Q, Wang X. Depletion of GPSM1 enhances ovarian granulosa cell apoptosis via cAMP-PKA-CREB pathway in vitro. J Ovarian Res (2020) 13(1):1–10. doi: 10.1186/s13048-020-00740-6
121. Liang M, Zhou J, Sun X, He C, Zhang K, Hu K. Effects of bisphenol A on apoptosis and ovarian development in mouse ovarian antral follicular granulosa cells. J South Med Univ (2021) 41(01):93–9. doi: 10.12122/j.issn.1673-4254.2021.01.13
122. Pe A Blanco A, García Sáez AJ. Bax, Bak and beyond—mitochondrial performance in apoptosis. FEBS J (2018) 285(3):416–31. doi: 10.1111/febs.14186
123. Doong H, Vrailas A, Kohn EC. What's in the ‘BAG’?–a functional domain analysis of the BAG-family proteins. Cancer Lett (2002) 188(1-2):25–32. doi: 10.1016/S0304-3835(02)00456-1
124. Baeken MW, Behl C. On the origin of BAG (3) and its consequences for an expansion of BAG3's role in protein homeostasis. J Cell Biochem (2022) 123(1):102–14. doi: 10.1002/jcb.29925
125. Chen Y, Yang LN, Cheng L, Tu S, Guo SJ, Le HY, et al. Bcl2-associated athanogene 3 interactome analysis reveals a new role in modulating proteasome activity. Mol Cell Proteomics (2013) 12(10):2804–19. doi: 10.1074/mcp.M112.025882
126. Zhang N, Zhang Y, Miao W, Shi C, Chen Z, Wu B, et al. An unexpected role for BAG3 in regulating PARP1 ubiquitination in oxidative stress-related endothelial damage. Redox Biol (2022) 50:102238. doi: 10.1016/j.redox.2022.102238
127. De Marco M, Falco A, Iaccarino R, Raffone A, Mollo A, Guida M, et al. An emerging role for BAG3 in gynaecological malignancies. Br J Cancer (2021) 125(6):789–97. doi: 10.1038/s41416-021-01446-2
128. Link W. Introduction to FOXO biology. FOXO Transcription Factors (2019) p:1–9. doi: 10.1007/978-1-4939-8900-3_1
129. Martins R, Lithgow GJ, Link W. Long live FOXO: unraveling the role of FOXO proteins in aging and longevity. Aging Cell (2016) 15(2):196–207. doi: 10.1111/acel.12427
130. Castrillon DH, Miao L, Kollipara R, Horner JW, DePinho RA. Suppression of ovarian follicle activation in mice by the transcription factor Foxo3a. Science (2003) 301(5630):215–8. doi: 10.1126/science.1086336
131. Xin M, He J, Jia Z, Wu Y, Zhang Y, Yang W, et al. Study of warming kidney and nourishing blood formula to improve oocyte quality and embryonic development potential in senior female rats. J Capital Med Univ (2018) 6(39):815–20.
132. Chung HY, Lee EK, Choi YJ, Kim JM, Kim DH, Zou Y, et al. Molecular inflammation as an underlying mechanism of the aging process and age-related diseases. J Dental Res (2011) 90(7):830–40. doi: 10.1177/0022034510387794
133. Dabravolski SA, Nikiforov NG, Eid AH, Nedosugova LV, Starodubova AV, Popkova TV, et al. Mitochondrial dysfunction and chronic inflammation in polycystic ovary syndrome. Int J Mol Sci (2021) 22(8):3923. doi: 10.3390/ijms22083923
134. Shukla S, Rizvi F, Raisuddin S, Kakkar P. FoxO proteins’ nuclear retention and BH3-only protein Bim induction evoke mitochondrial dysfunction-mediated apoptosis in berberine-treated HepG2 cells. Free Radical Biol Med (2014) 76:185–99. doi: 10.1016/j.freeradbiomed.2014.07.039
135. Cui Z, Zhao X, Amevor FK, Du X, Wang Y, Li D, et al. Therapeutic application of quercetin in aging-related diseases: SIRT1 as a potential mechanism. Front Immunol (2022) 13:943321. doi: 10.3389/fimmu.2022.943321
136. Liu M, Wang RB, Xing JH, Tang YX. Atractylenolide inhibits apoptosis and oxidative stress of HTR-8/SVneo cells by activating MAPK/ERK signalling in preeclampsia. Phytomedicine (2021) 93:153773. doi: 10.1016/j.phymed.2021.153773
137. Liu X, Fan L, Lu C, Yin S, Hu H. Functional role of p53 in the regulation of chemical-induced oxidative stress. Oxid Med Cell Longevity (2020) 2020. doi: 10.1155/2020/6039769
138. Lessel D, Wu D, Trujillo C, Ramezani T, Lessel I, Alwasiyah MK, et al. Dysfunction of the MDM2/p53 axis is linked to premature aging. J Clin Invest (2017) 127(10):3598–608. doi: 10.1172/JCI92171
139. Haraguchi H, Hirota Y, Saito-Fujita T, Tanaka T, Shimizu-Hirota R, Harada M, et al. Mdm2-p53-SF1 pathway in ovarian granulosa cells directs ovulation and fertilization by conditioning oocyte quality. FASEB J (2019) 33(2):2610–20. doi: 10.1096/fj.201801401R
140. Hotchkiss RS, Strasser A, McDunn JE, Swanson PE. Cell death. New Engl J Med (2009) 361(16):1570–83. doi: 10.1056/NEJMra0901217
141. Wang Z, Li B. Mdm2 links genotoxic stress and metabolism to p53. Protein Cell (2010) 1(12):1063–72. doi: 10.1007/s13238-010-0140-9
142. Zhang CX, Zhang Q, Xie YY, He XY, Xiang C, Hou XS, et al. Mouse double minute 2 actively suppresses p53 activity in oocytes during mouse folliculogenesis. Am J Pathol (2017) 187(2):339–51. doi: 10.1016/j.ajpath.2016.09.023
143. Galati S, Boni C, Gerra MC, Lazzaretti M, Buschini A. Autophagy: a player in response to oxidative stress and DNA damage. Oxid Med Cell Longevity (2019) 2019. doi: 10.1155/2019/5692958
144. Li Y, Xiao J, Bai J, Tian Y, Qu Y, Chen X, et al. Molecular characterization and clinical relevance of m6A regulators across 33 cancer types. Mol Cancer (2019) 18(1):1–6. doi: 10.1186/s12943-019-1066-3
145. Xia H, Zhong C, Wu X, Chen J, Tao B, Xia X, et al. Mettl3 mutation disrupts gamete maturation and reduces fertility in zebrafish. Genetics (2018) 208(2):729–43. doi: 10.1534/genetics.117.300574
146. Huff S, Tiwari SK, Gonzalez GM, Wang Y, Rana TM. m6A-RNA demethylase FTO inhibitors impair self-renewal in glioblastoma stem cells. ACS Chem Biol (2021) 16(2):324–33. doi: 10.1021/acschembio.0c00841
147. Li W, Deng D, Wang J, Zhang J, Xv J. Effect of nourishing essence and seeding jade soup on RNA-m~6A modification in a model of ovarian reserve hypofunction in rats. Chin J Traditional Chin Med Inf (2023) 30(05):109–13. doi: 10.19879/j.cnki.1005-5304.202206605
148. Ding C, Zou Q, Ding J, Ling M, Wang W, Li H, et al. Increased N6-methyladenosine causes infertility is associated with FTO expression. J Cell Physiol (2018) 233(9):7055–66. doi: 10.1002/jcp.26507
149. Jiang ZX, Wang YN, Li ZY, Dai ZH, He Y, Chu K, et al. The m6A mRNA demethylase FTO in granulosa cells retards FOS-dependent ovarian aging. Cell Death Dis (2021) 12(8):1–10. doi: 10.1038/s41419-021-04016-9
150. Hou L, Li S, Li S, Wang R, Zhao M, Liu X, et al. FTO inhibits oxidative stress by mediating m6A demethylation of Nrf2 to alleviate cerebral ischemia/reperfusion injury. J Physiol Biochem (2023) 79(1):133–46. doi: 10.1007/s13105-022-00929-x
151. Zhang GZ, Deng YJ, Xie QQ, Ren EH, Ma ZJ, He XG, et al. Sirtuins and intervertebral disc degeneration: roles in inflammation, oxidative stress, and mitochondrial function. Clinica chimica Acta (2020) 508:33–42. doi: 10.1016/j.cca.2020.04.016
152. Carafa V, Rotili D, Forgione M, Cuomo F, Serretiello E, Hailu GS, et al. Sirtuin functions and modulation: from chemistry to the clinic. Clin Epigenet (2016) 8(1):1–21. doi: 10.1186/s13148-016-0224-3
153. Zhang J, Fang L, Lu Z, Xiong J, Wu M, Shi L, et al. Are sirtuins markers of ovarian aging? Gene (2016) 575(2):680–6. doi: 10.1016/j.gene.2015.09.043
154. Yang W, Nagasawa K, Münch C, Xu Y, Satterstrom K, Jeong S, et al. Mitochondrial sirtuin network reveals dynamic SIRT3-dependent deacetylation in response to membrane depolarization. Cell (2016) 167(4):985–1000.e21. doi: 10.1016/j.cell.2016.10.016
155. Singh V, Ubaid S. Role of silent information regulator 1 (SIRT1) in regulating oxidative stress and inflammation. Inflammation (2020) 43(5):1589–98. doi: 10.1007/s10753-020-01242-9
156. Tatone C, Di Emidio G, Barbonetti A, Carta G, Luciano AM, Falone S, et al. Sirtuins in gamete biology and reproductive physiology: emerging roles and therapeutic potential in female and male infertility. Hum Reprod Update (2018) 24(3):267–89. doi: 10.1093/humupd/dmy003
157. Yang M, Peng Y, Liu W, Zhou M, Meng Q, Yuan C. Sirtuin 2 expression suppresses oxidative stress and senescence of nucleus pulposus cells through inhibition of the p53/p21 pathway. Biochem Biophys Res Commun (2019) 513(3):616–22. doi: 10.1016/j.bbrc.2019.03.200
158. Luo X, Xu J, Zhao R, Qin J, Wang X, Yan Y, et al. The role of inactivated NF-κB in premature ovarian failure. Am J Pathol (2022) 192(3):468–83. doi: 10.1016/j.ajpath.2021.12.005
159. Jiang Y, Zhang Z, Cha L, Li L, Zhu D, Fang Z, et al. Resveratrol plays a protective role against premature ovarian failure and prompts female germline stem cell survival. Int J Mol Sci (2019) 20(14):3605. doi: 10.3390/ijms20143605
160. Yang L, Zhao Q, Li Y, Zheng Z, Kong X, Shu C, et al. The role of oxidative stress and natural antioxidants in ovarian aging. Front Pharmacol (2021) 11:617843. doi: 10.3389/fphar.2020.617843
161. Unver E, Tosun M, Olmez H, Kuzucu M, Cimen FK, Suleyman Z. The effect of taxifolin on cisplatin-induced pulmonary damage in rats: A biochemical and histopathological evaluation. Mediators Inflammation (2019) 2019. doi: 10.1155/2019/3740867
162. Tan BL, Norhaizan ME. Effect of high-fat diets on oxidative stress, cellular inflammatory response and cognitive function. Nutrients (2019) 11(11):2579. doi: 10.3390/nu11112579
163. Xia S, Sun T & Wu J. Free radicals, inflammation and aging. Pract Geriatrics (2014) 28(02):100–3.
164. Lugrin J, Rosenblatt-Velin N, Parapanov R, Liaudet L. The role of oxidative stress during inflammatory processes. Biol Chem (2014) 395(2):203–30. doi: 10.1515/hsz-2013-0241
165. Han Y, Yao R, Yang Z, Li S, Meng W, Zhang Y, et al. Interleukin-4 activates the PI3K/AKT signaling to promote apoptosis and inhibit the proliferation of granulosa cells. Exp Cell Res (2022) 412(1):113002. doi: 10.1016/j.yexcr.2021.113002
166. Mantawy EM, Said RS, Abdel-Aziz AK. Mechanistic approach of the inhibitory effect of chrysin on inflammatory and apoptotic events implicated in radiation-induced premature ovarian failure: emphasis on TGF-β/MAPKs signaling pathway. Biomedicine Pharmacotherapy (2019) 109:293–303. doi: 10.1016/j.biopha.2018.10.092
167. Wang F, Li Y, Wang N, Liu Z, Zhou Y, Liu Y, et al. Effects of oxidative stress-inflammatory factors on premature ovarian failure in fluorosis. Modern Prev Med (2017) 44(12):2296–9.
168. Lin J, Nie X, Xiong Y, Gong Z, Chen J, Chen C, et al. Fisetin regulates gut microbiota to decrease CCR9+/CXCR3+/CD4+ T-lymphocyte count and IL-12 secretion to alleviate premature ovarian failure in mice. Am J Trans Res (2020) 12(1):203.
169. Gaschler MM, Stockwell BR. Lipid peroxidation in cell death. Biochem Biophys Res Commun (2017) 482(3):419–25. doi: 10.1016/j.bbrc.2016.10.086
170. Niki E. Lipid peroxidation products as oxidative stress biomarkers. Biofactors (2008) 34(2):171–80. doi: 10.1002/biof.5520340208
171. Negre-Salvayre A, Auge N, Ayala V, Basaga H, Boada J, Brenke R, et al. Pathological aspects of lipid peroxidation. Free Radical Res (2010) 44(10):1125–71. doi: 10.3109/10715762.2010.498478
172. Li Y, Zhao T, Li J, Xia M, Li Y, Wang X, et al. Oxidative stress and 4-hydroxy-2-nonenal (4-HNE): implications in the pathogenesis and treatment of aging-related diseases. J Immunol Res (2022) 2022. doi: 10.1155/2022/2233906
173. Cantonero C, Sánchez-Collado J, Lopez JJ, Salido GM, Rosado JA, Redondo PC. Arachidonic acid attenuates cell proliferation, migration and viability by a mechanism independent on calcium entry. Int J Mol Sci (2020) 21(9):3315. doi: 10.3390/ijms21093315
174. Liang C, Zhang X, Qi C, Hu H, Zhang Q, Zhu X, et al. UHPLC-MS-MS analysis of oxylipins metabolomics components of follicular fluid in infertile individuals with diminished ovarian reserve. Reprod Biol Endocrinol (2021) 19(1):1–14. doi: 10.1186/s12958-021-00825-x
175. Wolf LA, Laster SM. Characterization of arachidonic acid-induced apoptosis. Cell Biochem biophysics (1999) 30(3):353–68. doi: 10.1007/BF02738119
176. Breitzig M, Bhimineni C, Lockey R, Kolliputi N. 4-Hydroxy-2-nonenal: a critical target in oxidative stress? Am J Physiology-Cell Physiol (2016) 311(4):C537–43. doi: 10.1152/ajpcell.00101.2016
177. Hamon MP, Ahmed EK, Baraibar MA, Friguet B. Proteome oxidative modifications and impairment of specific metabolic pathways during cellular senescence and aging. Proteomics (2020) 20(5-6):1800421. doi: 10.1002/pmic.201800421
178. Hauck AK, Bernlohr DA. Oxidative stress and lipotoxicity. J Lipid Res (2016) 57(11):1976–86. doi: 10.1194/jlr.R066597
179. Akagawa M. Protein carbonylation: molecular mechanisms, biological implications, and analytical approaches. Free Radical Res (2021) 55(4):307–20. doi: 10.1080/10715762.2020.1851027
180. Bollineni RC, Hoffmann R, Fedorova M. Proteome-wide profiling of carbonylated proteins and carbonylation sites in HeLa cells under mild oxidative stress conditions. Free Radical Biol Med (2014) 68:186–95. doi: 10.1016/j.freeradbiomed.2013.11.030
181. Ahmed EK, Rogowska-Wrzesinska A, Roepstorff P, Bulteau AL, Friguet B. Protein modification and replicative senescence of WI-38 human embryonic fibroblasts. Aging Cell (2010) 9(2):252–72. doi: 10.1111/j.1474-9726.2010.00555.x
182. Mario T, Yvonne D, Veronica S, Alejandro D, Juan RM, Diana F, et al. Effects of perfluorooctanoic acid in oxidative stress generation, DNA damage in cumulus cells, and its impact on in vitro maturation of porcine oocytes. Environ Toxicol (2022) 37(6):1394–403. doi: 10.1002/tox.23492
183. Baraibar MA, Ladouce R, Friguet B. Proteomic quantification and identification of carbonylated proteins upon oxidative stress and during cellular aging. J Proteomics (2013) 92:63–70. doi: 10.1016/j.jprot.2013.05.008
184. Du X-Y. Progress in the study of the effect of vascular neogenesis-related factors on follicular growth and development. Chin J Family Plann (2014) 22(7):502–4. doi: 10.1155/2022/7012978
185. Duffy DM, Ko C, Jo M, Brannstrom M, Curry TE. Ovulation: parallels with inflammatory processes. Endocrine Rev (2019) 40(2):369–416. doi: 10.1210/er.2018-00075
186. Akiyama I, Yoshino O, Osuga Y, Shi J, Harada M, Koga K, et al. Bone morphogenetic protein 7 increased vascular endothelial growth factor (VEGF)-a expression in human granulosa cells and VEGF receptor expression in endothelial cells. Reprod Sci (2014) 21(4):477–82. doi: 10.1177/1933719113503411
187. Guo X, Yi H, Li TC, Wang Y, Wang H, Chen X. Role of vascular endothelial growth factor (VEGF) in human embryo implantation: clinical implications. Biomolecules (2021) 11(2):253. doi: 10.3390/biom11020253
188. Zhou M, Yv X, Mao H. Effects of Erxian Tang on serum inflammatory factors, related growth factors and sex hormone levels in rats with premature ovarian failure model. Chin J Traditional Chin Med (2021) 39(09):240–3. doi: 10.13193/j.issn.1673-7717.2021.09.060
189. Fukai T, Ushio-Fukai M. Cross-talk between NADPH oxidase and mitochondria: role in ROS signaling and angiogenesis. Cells (2020) 9(8):1849. doi: 10.3390/cells9081849
190. Chen Q, Tang L, Zhang Y, Wan C, Yu X, Dong Y, et al. STING up-regulates VEGF expression in oxidative stress-induced senescence of retinal pigment epithelium via NF-κB/HIF-1α pathway. Life Sci (2022) 293:120089. doi: 10.1016/j.lfs.2021.120089
191. Huang Y, Nan G. Oxidative stress-induced angiogenesis. J Clin Neurosci (2019) 63:13–6. doi: 10.1016/j.jocn.2019.02.019
192. Zhou F, Song Y, Liu X, Zhang C, Li F, Hu R, et al. Si-Wu-Tang facilitates ovarian function through improving ovarian microenvironment and angiogenesis in a mouse model of premature ovarian failure. J Ethnopharmacology (2021) 280:114431. doi: 10.1016/j.jep.2021.114431
193. Weng Y, Wu S, Luo J, Lu C, Hu Y, Liang Y, et al. Modulatory effects of GDF15 on oxidative stress-induced vascular endothelial injury and angiogenesis. J Guangxi Med Univ (2022) 39(05):693–8. doi: 10.16190/j.cnki.45-1211/r.2022.05.002
194. Kanazawa H, Yoshikawa J. Elevated oxidative stress and reciprocal reduction of vascular endothelial growth factor levels with severity of COPD. Chest (2005) 128(5):3191–7. doi: 10.1378/chest.128.5.3191
195. Kalantaridou SN, Naka KK, Papanikolaou E, Kazakos N, Kravariti M, Calis KA, et al. Impaired endothelial function in young women with premature ovarian failure: normalization with hormone therapy. J Clin Endocrinol Metab (2004) 89(8):3907–13. doi: 10.1210/jc.2004-0015
196. Kalluri R, Le Bleu VS. The biology, function, and biomedical applications of exosomes. Science (2020) 367(6478):eaau6977. doi: 10.1126/science.aau6977
197. Shen K, Jia Y, Wang X, Zhang J, Liu K, Wang J, et al. Exosomes from adipose-derived stem cells alleviate the inflammation and oxidative stress via regulating Nrf2/HO-1 axis in macrophages. Free Radical Biol Med (2021) 165:54–66. doi: 10.1016/j.freeradbiomed.2021.01.023
198. Bodart-Santos V, de Carvalho LRP, de Godoy MA, Batista AF, Saraiva LM, Lima LG, et al. Extracellular vesicles derived from human Wharton’s jelly mesenchymal stem cells protect hippocampal neurons from oxidative stress and synapse damage induced by amyloid-β oligomers. Stem Cell Res Ther (2019) 10(1):1–13. doi: 10.1186/s13287-019-1432-5
199. Yao J, Zheng J, Cai J, Zeng K, Zhou C, Zhang J, et al. Extracellular vesicles derived from human umbilical cord mesenchymal stem cells alleviate rat hepatic ischemia-reperfusion injury by suppressing oxidative stress and neutrophil inflammatory response. FASEB J (2019) 33(2):1695–710. doi: 10.1096/fj.201800131RR
200. Yang M, Lin L, Sha C, Li T, Zhao D, Wei H, et al. Bone marrow mesenchymal stem cell-derived exosomal miR-144-5p improves rat ovarian function after chemotherapy-induced ovarian failure by targeting PTEN. Lab Invest (2020) 100(3):342–52. doi: 10.1038/s41374-019-0321-y
201. Zhang W, Liu R, Chen Y, Wang M, Du J. Crosstalk between oxidative stress and exosomes. Oxid Med Cell Longevity (2022) 2022. doi: 10.1155/2022/3553617
202. Qu Q, Liu L, Cui Y, Liu H, Yi J, Bing W, et al. miR-126-3p containing exosomes derived from human umbilical cord mesenchymal stem cells promote angiogenesis and attenuate ovarian granulosa cell apoptosis in a preclinical rat model of premature ovarian failure. Stem Cell Res Ther (2022) 13(1):1–15. doi: 10.1186/s13287-022-03056-y
203. Sun B, Ma Y, Wang F, Hu L, Sun Y. miR-644-5p carried by bone mesenchymal stem cell-derived exosomes targets regulation of p53 to inhibit ovarian granulosa cell apoptosis. Stem Cell Res Ther (2019) 10(1):1–9. doi: 10.1186/s13287-019-1442-3
204. Qin Y. Research progress of antioxidants to improve ovarian function. Chin J Reprod Health (2021) 32(01):91–4.
205. Jia R. Study on the toxic effect and mechanism of action of iodoacetic acid on antioxidant enzymes. China: Yantai University (2020).
206. Jang H, Hong K, Choi Y. Melatonin and fertoprotective adjuvants: prevention against premature ovarian failure during chemotherapy. Int J Mol Sci (2017) 18(6):1221. doi: 10.3390/ijms18061221
207. Song C, Peng W, Yin S, Zhao J, Fu B, Zhang J, et al. Melatonin improves age-induced fertility decline and attenuates ovarian mitochondrial oxidative stress in mice. Sci Rep (2016) 6(1):35165. doi: 10.1038/srep35165
208. Zhang H, Li C, Wen D, Li R, Lu S, Xu R, et al. Melatonin improves the quality of maternally aged oocytes by maintaining intercellular communication and antioxidant metabolite supply. Redox Biol (2022) 49:102215. doi: 10.1016/j.redox.2021.102215
209. Xie QE, Wang MY, Cao ZP, Du X, Ji DM, Liang D, et al. Melatonin protects against excessive autophagy-induced mitochondrial and ovarian reserve function deficiency though ERK signaling pathway in Chinese hamster ovary (CHO) cells. Mitochondrion (2021) 61:44–53. doi: 10.1016/j.mito.2021.09.009
210. Osatd-Rahimi N, Saburi E, Karimi S, Boustan A, Ebrahimzadeh-Bideskan A. The therapeutic effect of melatonin on female offspring ovarian reserve and quality in BALB/c mice after exposing their mother to methamphetamine during pregnancy and lactation. Iranian J Basic Med Sci (2023) 26(2):208. doi: 10.22038/IJBMS.2022.66660.14636
211. Cruz MH, Leal CL, Cruz JF, Tan DX, Reiter RJ. Essential actions of melatonin in protecting the ovary from oxidative damage. Theriogenology (2014) 82(7):925–32. doi: 10.1016/j.theriogenology.2014.07.011
212. Lambertini M, Partridge AH. Cyclophosphamide-free adjuvant chemotherapy for the potential prevention of premature ovarian insufficiency and infertility in young women with breast cancer. United States: Oxford University Press (2021) p. 1274–6.
213. Spears N, Lopes F, Stefansdottir A, Rossi V, De Felici M, Anderson RA, et al. Ovarian damage from chemotherapy and current approaches to its protection. Hum Reprod Update (2019) 25(6):673–93. doi: 10.1093/humupd/dmz027
214. Lin L, Gao W, Chen Y, Li T, Sha C, Chen L, et al. Reactive oxygen species-induced SIAH1 promotes granulosa cells' senescence in premature ovarian failure. J Cell Mol Med (2022) 26(8):2417–27. doi: 10.1111/jcmm.17264
215. Zhang Y, Yan Z, Qin Q, Nisenblat V, Chang HM, Yu Y, et al. Transcriptome landscape of human folliculogenesis reveals oocyte and granulosa cell interactions. Mol Cell (2018) 72(6):1021–1034.e4. doi: 10.1016/j.molcel.2018.10.029
216. Xu H, Bao X, Kong H, Yang J, Li Y, Sun Z, et al. Melatonin protects against cyclophosphamide-induced premature ovarian failure in rats. Hum Exp Toxicol (2022) 41:09603271221127430. doi: 10.1177/09603271221127430
217. Andersen LP, Gögenur I, Rosenberg J, Reiter RJ. The safety of melatonin in humans. Clin Drug Invest (2016) 36(3):169–75. doi: 10.1007/s40261-015-0368-5
218. Jia Q, Zhang N, Kaiyun. Research progress on the protective effect of sphingosine 1-phosphate on the reproductive system. World abstract latest Med Inf (2019) 19(08):110–2. doi: 10.19613/j.cnki.1671-3141.2019.08.052
219. Shi Y. The role of histone acetylation regulation by sphingosine 1-phosphate in oxidative stress-induced insulin resistance. China: Tianjin Medical University (2011).
220. Delkhosh A, Delashoub M, Tehrani AA, Bahrami AM, Niazi V, Shoorei H, et al. Upregulation of FSHR and PCNA by administration of coenzyme Q10 on cyclophosphamide-induced premature ovarian failure in a mouse model. J Biochem Mol Toxicol (2019) 33(11):e22398. doi: 10.1002/jbt.22398
221. Mantawy EM, Said RS, Kassem DH, Abdel-Aziz AK, Badr AM. Novel molecular mechanisms underlying the ameliorative effect of N-acetyl-L-cysteine against ?-radiation-induced premature ovarian failure in rats. Ecotoxicology Environ Saf (2020) 206:111190. doi: 10.1016/j.ecoenv.2020.111190
222. He H, Qin D. Research progress of coenzyme Q10 in the treatment of osteoporosis. Chin Prescription Drugs (2022) 20(02):176–8.
223. Qin Y. Progress in the study of antioxidants to improve ovarian function. Chin J Reprod Health (2021) 32(1):91–4.
224. Zhao M, Li J, liang Q, Pan C, Cui W, et al. The preventive and curative effects of coenzyme Q10 in patients with reduced ovarian reserve function. Chin Med Herald (2020) 39(01):73–8.
225. Papi A, Di Stefano AF, Radicioni M. Pharmacokinetics and safety of single and multiple doses of oral N-acetylcysteine in healthy chinese and caucasian volunteers: An open-label, phase I clinical study. Adv Ther (2021) 38(1):468–78. doi: 10.1007/s12325-020-01542-4
226. Perumpail BJ, Li AA, John N, Sallam S, Shah ND, Kwong W, et al. The role of vitamin E in the treatment of NAFLD. Diseases (2018) 6(4):86. doi: 10.3390/diseases6040086
227. Xueling L, Kun MA, Wenhua T, Zhongkun XU, Gang L, Chunyan HU, et al. Natural products for treatment of premature ovarian failure: a narrative review. J Traditional Chin Med (2023) 43(3):606. doi: 10.19852/j.cnki.jtcm.20230227.002
228. Yan Q, Shi D, Yang J, He Y, Zhao P. Protective effect of ginsenoside Rg_1 on cisplatin-injured rat ovarian granulosa cells and its molecular mechanism. Zhongnan Pharmacol (2022) 20(05):1028–33.
229. Banerjee S, Chakraborty P, Saha P, Bandyopadhyay SA, Banerjee S, Kabir SN. Ovotoxic effects of galactose involve attenuation of follicle-stimulating hormone bioactivity and up-regulation of granulosa cell p53 expression. PloS One (2012) 7(2):e30709. doi: 10.1371/journal.pone.0030709
230. Guerrero NV, Singh RH, Manatunga A, Berry GT, Steiner RD, Elsas LJ 2nd, et al. Risk factors for premature ovarian failure in females with galactosemia. J Pediatr (2000) 137(6):833–41. doi: 10.1067/mpd.2000.109148
231. Liu X, Zhao Z, Zhou Y, Wang C, Han Y, Zhou W, et al. Role of PI3K/Akt/mTOR autophagic pathway in ginsenoside Rg_1 in delaying D-gal-induced premature ovarian failure in a mouse model of premature ovarian failure. Chin J Traditional Chin Med (2020) 45(24):6036–42. doi: 10.19540/j.cnki.cjcmm.20200901.405
232. Yan Z. Study on the effect of curcumin in delaying premature ovarian failure and its mechanism. China: Yangzhou University (2021).
233. Tang Z. Effects of ginsenoside Rb_1 on sex hormones in rats with premature ovarian failure caused by chemotherapy. Contemp Med (2013) 19(05):30+115.
234. Hoppe C, Freuding M, Büntzel J, Münstedt K, Hübner J. Clinical efficacy and safety of oral and intravenous vitamin C use in patients with malignant diseases. J Cancer Res Clin Oncol (2021) 147(10):3025–42. doi: 10.1007/s00432-021-03759-4
235. Samare-Najaf M, Zal F, Safari S, Koohpeyma F, Jamali N. Stereological and histopathological evaluation of doxorubicin-induced toxicity in female rats’ ovary and uterus and palliative effects of quercetin and vitamin E. Hum Exp Toxicol (2020) 39(12):1710–24. doi: 10.1177/0960327120937329
236. Zeng L, Yang T, Yang K, Yu G, Li J, Xiang W, et al. Efficacy and safety of curcumin and curcuma longa extract in the treatment of arthritis: a systematic review and meta-analysis of randomized controlled trial. Front Immunol (2022) 13. doi: 10.3389/fimmu.2022.891822
237. Movahed A, Raj P, Nabipour I, Mahmoodi M, Ostovar A, Kalantarhormozi M, et al. Efficacy and safety of resveratrol in type 1 diabetes patients: a two-month preliminary exploratory trial. Nutrients (2020) 12(1):161. doi: 10.3390/nu12010161
238. Han Y. Exploring the mechanism of ginsenoside Rg1-induced senescence in human leukemia cells (K562) based on PI3K/Akt/mTOR autophagy pathway. China: Dali University (2020).
239. Li Z, Zheng J, Tian Y, Huang X. Recent advances and mechanisms of mesenchymal stem cell therapy for premature ovarian failure. Chin Tissue Eng Res (2022) 26(01):141–7.
240. Park HS, Chugh RM, Seok J, Cetin E, Mohammed H, Siblini H, et al. Comparison of the therapeutic effects between stem cells and exosomes in primary ovarian insufficiency. Stem Cell Res Ther (2023) 14(1):165. doi: 10.21203/rs.3.rs-2151593/v1
241. Mohamed SA, Shalaby SM, Abdelaziz M, Brakta S, Hill WD, Ismail N, et al. Human mesenchymal stem cells partially reverse infertility in chemotherapy-induced ovarian failure. Reprod Sci (2018) 25:51–63. doi: 10.1177/1933719117699705
242. Seok J, Park H, Choi JH, Lim JY, Kim KG, Kim GJ. Placenta-derived mesenchymal stem cells restore the ovary function in an ovariectomized rat model via an antioxidant effect. Antioxidants (2020) 9(7):591. doi: 10.3390/antiox9070591
243. Huang Y, Zhu M, Liu Z, Hu R, Li F, Song Y, et al. Bone marrow mesenchymal stem cells in premature ovarian failure: Mechanisms and prospects. Front Immunol (2022) 13:997808. doi: 10.3389/fimmu.2022.997808
244. Jalalie L, Rezaie MJ, Jalili A, Rezaee MA, Vahabzadeh Z, Rahmani MR, et al. Distribution of the CM-Dil-labeled human umbilical cord vein mesenchymal stem cells migrated to the cyclophosphamide-injured ovaries in C57BL/6 mice. Iranian Biomed J (2019) 23(3):200. doi: 10.29252/ibj.23.3.200
245. El-Tantawy WH, Haleem ENAA. Therapeutic effects of stem cell on hyperglycemia, hyperlipidemia, and oxidative stress in alloxan-treated rats. Mol Cell Biochem (2014) 391(1):193–200. doi: 10.1007/s11010-014-2002-x
246. Huang B, Qian C, Ding C, Meng Q, Zou Q, Li H. Fetal liver mesenchymal stem cells restore ovarian function in premature ovarian insufficiency by targeting MT1. Stem Cell Res Ther (2019) 10(1):1–12. doi: 10.1186/s13287-019-1490-8
247. Li R, Jia H, Zhang C, Zhao R, Dong J, Cheng Y, et al. Role of exosomes and miRNAs in the mechanism and diagnosis of early-onset ovarian insufficiency. Chin Tissue Eng Res (2023) 27(15):2453–60.
248. Pires AO, Mendes-Pinheiro B, Teixeira FG, Anjo SI, Ribeiro-Samy S, Gomes ED, et al. Unveiling the differences of secretome of human bone marrow mesenchymal stem cells, adipose tissue-derived stem cells, and human umbilical cord perivascular cells: a proteomic analysis. Stem Cells Dev (2016) 25(14):1073–83. doi: 10.1089/scd.2016.0048
249. Gabr H, Rateb MA, El Sissy MH, Ahmed Seddiek H, Ali Abdelhameed Gouda S. The effect of bone marrow-derived mesenchymal stem cells on chemotherapy induced ovarian failure in albino rats. Microscopy Res technique (2016) 79(10):938–47. doi: 10.1002/jemt.22725
250. Wang Z, Yang T, Liu S, Chen Y. Effects of bone marrow mesenchymal stem cells on ovarian and testicular function in aging Sprague-Dawley rats induced by D-galactose. Cell Cycle (2020) 19(18):2340–50. doi: 10.1080/15384101.2020.1806434
251. Ding C, Zhu L, Shen H, Lu J, Zou Q, Huang C, et al. Exosomal miRNA-17-5p derived from human umbilical cord mesenchymal stem cells improves ovarian function in premature ovarian insufficiency by regulating SIRT7. Stem Cells (2020) 38(9):1137–48. doi: 10.1002/stem.3204
252. Fu YX, Ji J, Shan F, Li J, Hu R. Human mesenchymal stem cell treatment of premature ovarian failure: new challenges and opportunities. Stem Cell Res Ther (2021) 12(1):1–13. doi: 10.1186/s13287-021-02212-0
253. Zheng X. Study on the mechanism of antioxidant enzymes and phase II detoxification enzyme system in oxidative damage of mouse lungs caused by diesel exhaust particulate matter. China: Chinese Center for Disease Control and Prevention (2019).
Keywords: oxidative stress, aging and oocyte, premature ovarian insufficiency, antioxidant, mitochondrial dysfunction
Citation: Shi Y-Q, Zhu X-T, Zhang S-N, Ma Y-F, Han Y-H, Jiang Y and Zhang Y-H (2023) Premature ovarian insufficiency: a review on the role of oxidative stress and the application of antioxidants. Front. Endocrinol. 14:1172481. doi: 10.3389/fendo.2023.1172481
Received: 28 February 2023; Accepted: 03 July 2023;
Published: 01 August 2023.
Edited by:
Marc Yeste, University of Girona, SpainReviewed by:
Svetlana Vujovic, University of Belgrade, SerbiaCopyright © 2023 Shi, Zhu, Zhang, Ma, Han, Jiang and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yue-Hui Zhang, Y2hpemlzaHVpLTA0QDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.