
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Endocrinol., 31 May 2023
Sec. Systems Endocrinology
Volume 14 - 2023 | https://doi.org/10.3389/fendo.2023.1159714
This article is part of the Research TopicGenetic variants and metabolic diseases, volume IIView all 13 articles
 Qiaoli Zeng1,2,3†
Qiaoli Zeng1,2,3† Bing Tan1,2,4†Fengqiong Han5†Xiujuan Huang6
Bing Tan1,2,4†Fengqiong Han5†Xiujuan Huang6 Jinzhi Huang7*Yue Wei8*
Jinzhi Huang7*Yue Wei8* Runmin Guo1,2,3,9*
Runmin Guo1,2,3,9*Background: The solute carrier family 30 A8 zinc transporter (SLC30A8) plays a crucial role in insulin secretion. This study aimed to investigate the impact of SLC30A8 gene polymorphisms on gestational diabetes mellitus (GDM).
Methods: The research objective was to select 500 patients with GDM and 502 control subjects. Rs13266634 and rs2466293 were genotyped using the SNPscan™ genotyping assay. Statistical tests, such as the chi-square test, t-test, logistic regression, ANOVA, and meta-analysis, were conducted to determine the differences in genotypes, alleles, and their associations with GDM risk.
Results: Statistically significant differences were observed in age, pregestational BMI, SBP, DBP, and parity between individuals with GDM and healthy subjects (P < 0.05). After adjusting for these factors, rs2466293 remained significantly associated with an increased risk of GDM in overall subjects (GG+AG vs. AA: OR = 1.310; 95% CI: 1.005-1.707; P = 0.046, GG vs. AA: OR = 1.523; 95% CI: 1.010-2.298; P = 0.045 and G vs. A: OR = 1.249; 95% CI: 1.029-1.516; P = 0.024). Rs13266634 was still found to be significantly associated with a decreased risk of GDM in individuals aged ≥ 30 years (TT vs. CT+CC: OR = 0.615; 95% CI: 0.392-0.966; P = 0.035, TT vs. CC: OR = 0.503; 95% CI: 0.294-0.861; P = 0.012 and T vs. C: OR =0.723; 95% CI: 0.557-0.937; P = 0.014). Additionally, the haplotype CG was found to be associated with a higher risk of GDM (P < 0.05). Furthermore, pregnant women with the CC or CT genotype of rs13266634 exhibited significantly higher mean blood glucose levels than those with the TT genotype (P < 0.05). Our findings were further validated by the results of a meta-analysis.
Conclusion: The SLC30A8 rs2466293 polymorphism was found to be associated with an increased risk of GDM, while rs13266634 was associated with a decreased risk of GDM in individuals aged ≥ 30 years. These findings provide a theoretical basis for GDM testing.
Gestational diabetes mellitus (GDM) is a global concern, and its incidence has increased by over 30% in numerous countries during the past few years (1, 2). GDM is characterized by β-cell dysfunction, insulin resistance, and abnormal glucose utilization (3, 4), but its pathogenesis is not yet clear. Increasing evidence indicates that environmental and genetic factors are implicated in the development of GDM. Single nucleotide polymorphisms (SNPs) are a common type of genetic variation, and polymorphisms in different genes may be associated with GDM (5).
The solute carrier family 30 A8 zinc transporter (SLC30A8) gene encodes ZnT8, which is primarily expressed in pancreatic β-cells and is in charge of delivering zinc from the cytoplasm into insulin vesicles (6). SLC30A8 is involved in the secretion of insulin (7). The zinc stabilizes the insulin hexamer in secretory insulin vesicles, making it resistant to degradation (8). Insulin packaged into secretory vesicles can be released immediately upon glucose stimulation (7). The rs13266634 polymorphism is a missense C to T variant in exon 9 of the SLC30A8 gene, and the amino acid changes from arginine (R) to tryptophan (W) at position 325 (8). Thus, rs13266634 has been thought to be related to diabetes risk, as it affects the expression of SLC30A8, and negative regulation of ZnT8 is considered to disrupt the stability of insulin molecules (9). The polymorphism rs2466293 is in the 3′-UTR of the SLC30A8 gene, and rs2466293 may impact SLC30A8 post-transcriptional regulation by binding to miRNA (10). MiRNAs are closely related to gene level regulation; hence, rs2466293 in the seed sites of miRNA targets can create or disrupt miRNA-binding sites that further influence disease susceptibility (11). In this context, this study researched the influence of rs13266634 and rs2466293 polymorphisms on GDM risk.
From 1 August 1 2021 to 31 January 31 2022, a total of 1,002 unrelated Chinese Han pregnant women (500 GDM cases and 502 controls) were recruited for our study at the obstetric clinic of Shunde Maternal and Child Health Hospital, Guangdong Medical University. All individuals underwent a routine 75-gram oral glucose tolerance test (OGTT) during 24-28 weeks of gestation. A control group consisting of pregnant women at 24 to 28 weeks of gestation was selected over the same period. The inclusion criteria were as follows: voluntarily provided written informed consent, not previously diagnosed with diabetes, Han nationality, aged ≥ 18 years, no pregnancy complications, and not taking hypoglycemic medicines. Participants who did not meet the above criteria were excluded.
Information including age, height, pregestational weight, parity (primipara or multipara), blood pressure, race, pregnancy condition, and other clinical information were obtained at 24-28 gestational weeks. Pregestational body mass index (pre-BMI, Kg/m2) was calculated as pregestational weight (Kg) divided by height squared (m2). The Chinese standards for obesity were as follows: underweight (< 18.5 Kg/m2), normal (18.5-24 Kg/m2), overweight (24-28 Kg/m2), and obese (≥ 28 kg/m2).
A total of 2 mL of EDTA-treated blood was immediately stored in the freezer. Genomic DNA was extracted and purified from blood cells by a QIAamp DNA Blood Kit (Qiagen, Germany). Genotypes of candidate SNPs were determined using the SNPscan™ genotyping assay (Genesky Technologies Inc., Shanghai, China). Pre-experiments were conducted before formal experiments. In order to check the genotyping data accuracy, 6% of the samples were randomly selected for duplicate analysis using Sanger sequencing.
Continuous variables following normal distribution were reported as means ± SD, and the independent sample t-test was used to determine the differences between the relevant parameters of the two groups. In cases where the assumption of normality was violated, non-parametric tests were employed. Qualitative data were analyzed using the chi-square (χ2) test. The Hardy-Weinberg equilibrium (HWE) test, assessed through the goodness-of-fit χ2, was used to ensure that the control group was representative of the population. The risk of GDM was evaluated using six genetic models, namely, codominant homozygous, codominant heterozygous, dominant, recessive, overdominant, and allele models, through the χ2 test and logistic regression analysis. Crude and adjusted odds ratios (ORs) and their corresponding 95% confidence intervals (CIs) were presented, with adjustments made for covariates such as age, pre-BMI, etc. Stratified analysis was performed to further examine the potential influence of age and pre-BMI on the results. The frequency distribution of haplotypes was calculated using Haploview 4.2 software. The association between SNPs and blood glucose levels was investigated using one-way ANOVA. For multiple comparisons, the least significant difference (LSD) method was used. Statistical analyses were performed using SPSS 20.0 (SPSS Inc., Chicago, IL, USA), and a P-value < 0.05 was considered statistically significant.
The study included 500 GDM cases and 502 non-diabetic controls for the evaluation of the SLC30A8 genotype. Table 1 presents the clinical baseline information and stratified features. The mean age, pre-BMI, systolic blood pressure (SBP), diastolic blood pressure (DBP), fasting plasma glucose (FPG), 1 h-PG, and 2 h-PG were significantly higher in the GDM group than in the control group (P < 0.05). Furthermore, there was a significant difference in parity between the GDM and control groups (P < 0.05).
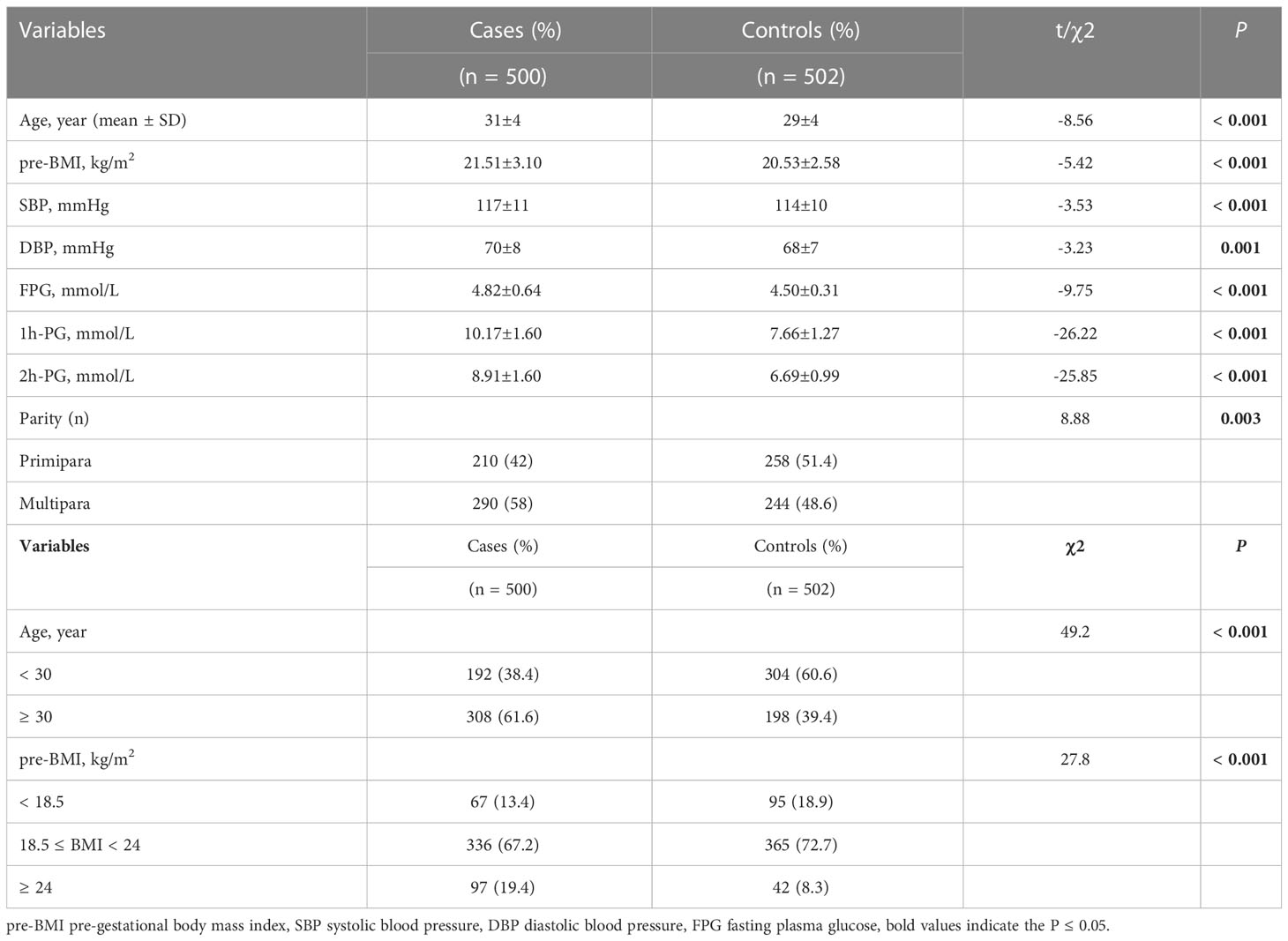
Table 1 Basic and stratified characteristic of participants of the study.
Table 2 presents the minor allele frequency (MAF) and the results of the HWE analysis for two SNPs in the control group. The results were in conformity with HWE (P > 0.05). Table 3 shows the ORs with corresponding 95% CIs and associated P values estimated for the relationship between genotypes and GDM in the six models (codominant homozygous, codominant heterozygous, dominant, recessive, overdominant, and allele models) for each polymorphism. SLC30A8 rs2466293 was found to be significantly associated with an increased risk of GDM in the dominant model (GG+AG vs. AA: OR = 1.288; 95% CI: 1.003-1.655; P = 0.047), codominant homozygous model (GG vs. AA: OR = 1.499; 95% CI: 1.014-2.217; P = 0.043), and allele model (G vs. A: OR = 1.237; 95% CI: 1.029-1.487; P = 0.023). Further evaluation was performed using a logistic regression method to adjust for age, pre-BMI, SBP, DBP, and parity. The results indicated a strong association between SLC30A8 rs2466293 and an increased risk of GDM in the dominant model (GG+AG vs. AA: OR = 1.310; 95% CI: 1.005-1.707; P = 0.046), codominant homozygous model (GG vs. AA: OR = 1.523; 95% CI: 1.010-2.298; P = 0.045), and allele model (G vs. A: OR = 1.249; 95% CI: 1.029-1.516; P = 0.024). However, no significant association was found in rs13266634.
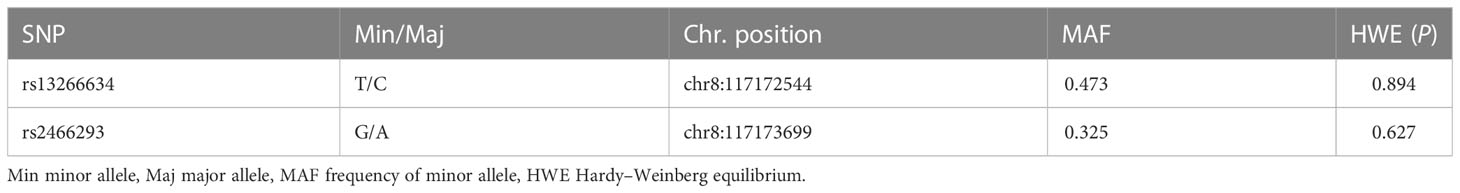
Table 2 SNPs information and HWE test in the controls.
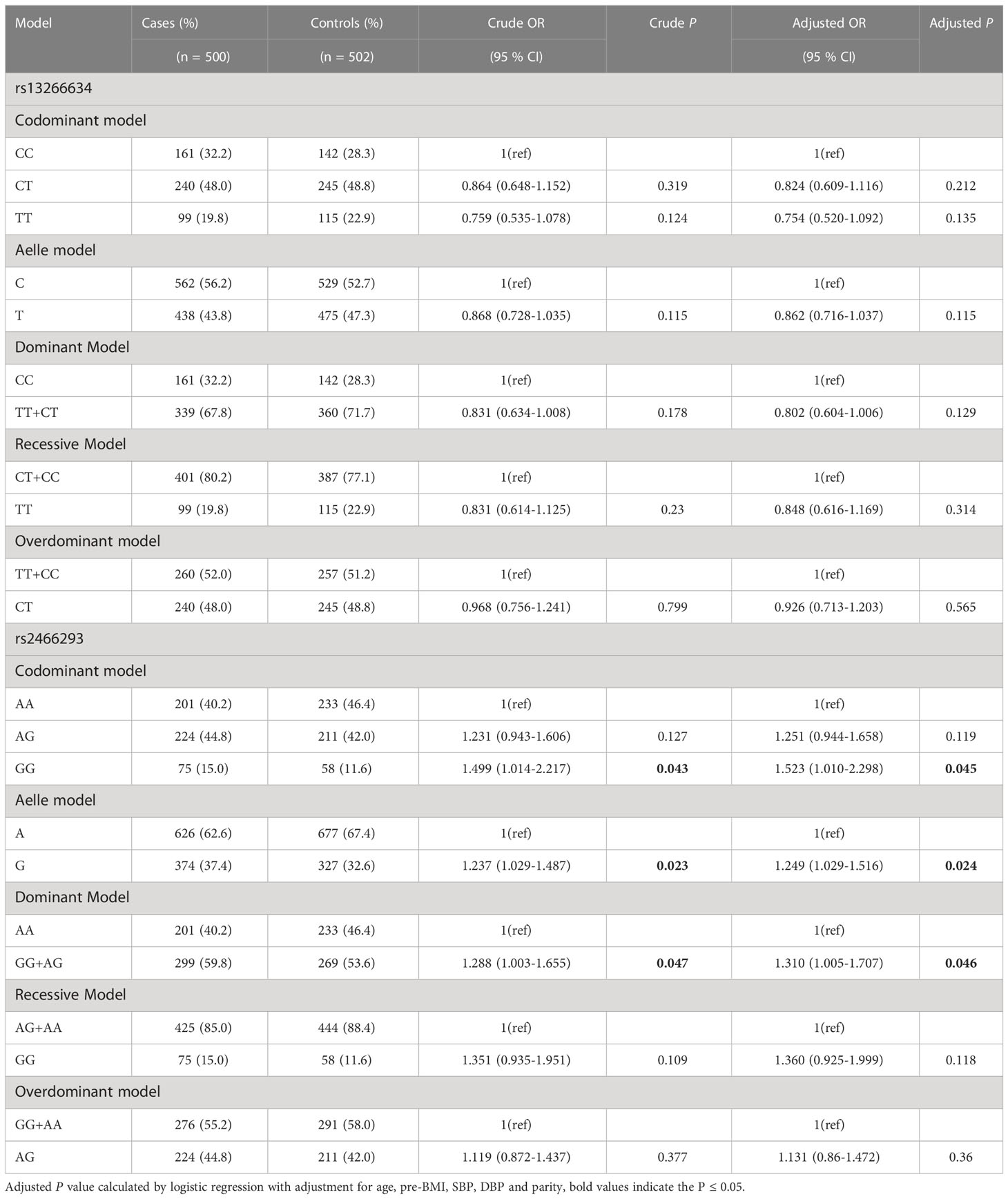
Table 3 The associations between SNPs in SLA30C8 gene and GDM risk in overall subjects.
Subsequently, the associations between two SNPs and susceptibility to GDM in six models were tested using stratified analysis for age or pre-BMI. Notably, protective roles were detected in subjects aged ≥ 30 years for rs13266634 under the dominant model (TT+CT vs. CC: OR = 0.648; 95% CI: 0.431-0.975; P = 0.037), codominant homozygous (TT vs. CC: OR = 0.517; 95% CI: 0.307-0.872; P = 0.013) and allele model (T vs. C: OR = 0.728; 95% CI: 0.565-0.938; P = 0.014). After adjustments, rs13266634 was significantly associated with lower GDM risk under the recessive model (TT vs. CT+CC: OR = 0.615; 95% CI: 0.392-0.966; P = 0.035), codominant homozygous model (TT vs. CC: OR = 0.503; 95% CI: 0.294-0.861; P = 0.012) and allele model (T vs. C: OR =0.723; 95% CI: 0.557-0.937; P = 0.014) (Table 4). Moreover, these associations were more evident in subjects aged ≥ 30 years for rs2466293 under the dominant model (GG+AG vs. AA: OR = 1.445; 95% CI: 1.007-2.073; P = 0.045) and allele model (G vs. A: OR = 1.337; 95% CI: 1.024-1.747; P = 0.033). After these abovementioned factors were adjusted, rs2466293 was significantly related to higher GDM odds under the dominant model (GG+AG vs. AA: OR = 1.579; 95% CI: 1.086-2.295; P = 0.017), codominant heterozygous (AG vs. AA: OR = 1.519; 95% CI: 1.020-2.263; P = 0.040), and allele model (G vs. A: OR = 1.399; 95% CI: 1.064-1.839; P = 0.016) (Table 4). However, no significant associations were found in subjects aged < 30 years (Supplementary Table 1). Nevertheless, the results indicated no significant relationship between rs13266634 or rs2466293 and GDM susceptibility in subjects in the pre-BMI stratified analysis.
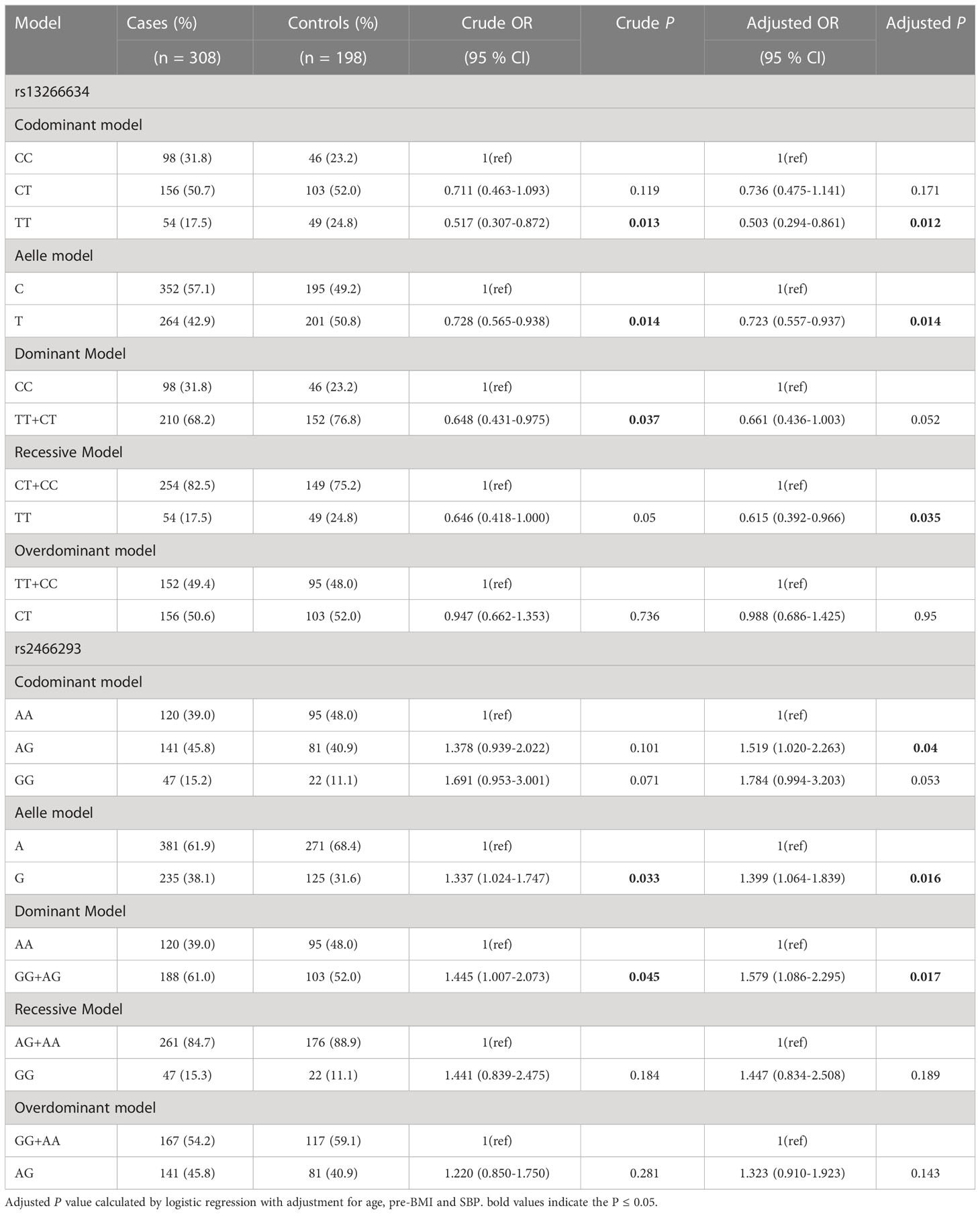
Table 4 The associations between SNPs in SLA30C8 gene and GDM risk in subjects aged ≥ 30 years.
The study found that two SNPs, rs13266634 and rs2466293, were in strong linkage disequilibrium (D′ > 0.99) with each other (Figure 1). The CG haplotype consisting of these SNPs was significantly associated with higher GDM risk (OR = 1.231; 95% CI: 1.024-1.48; P = 0.026). In addition, the age-stratified analysis revealed that haplotype CG was associated with higher GDM risk in subjects aged ≥ 30 years (OR = 1.328; 95% CI: 1.016-1.734; P = 0.037), while haplotype TA was associated with lower GDM risk in subjects aged ≥ 30 years (OR = 0.722; 95% CI: 0.560-0.931; P = 0.011). However, no significant associations were found with age < 30 years (Table 5).
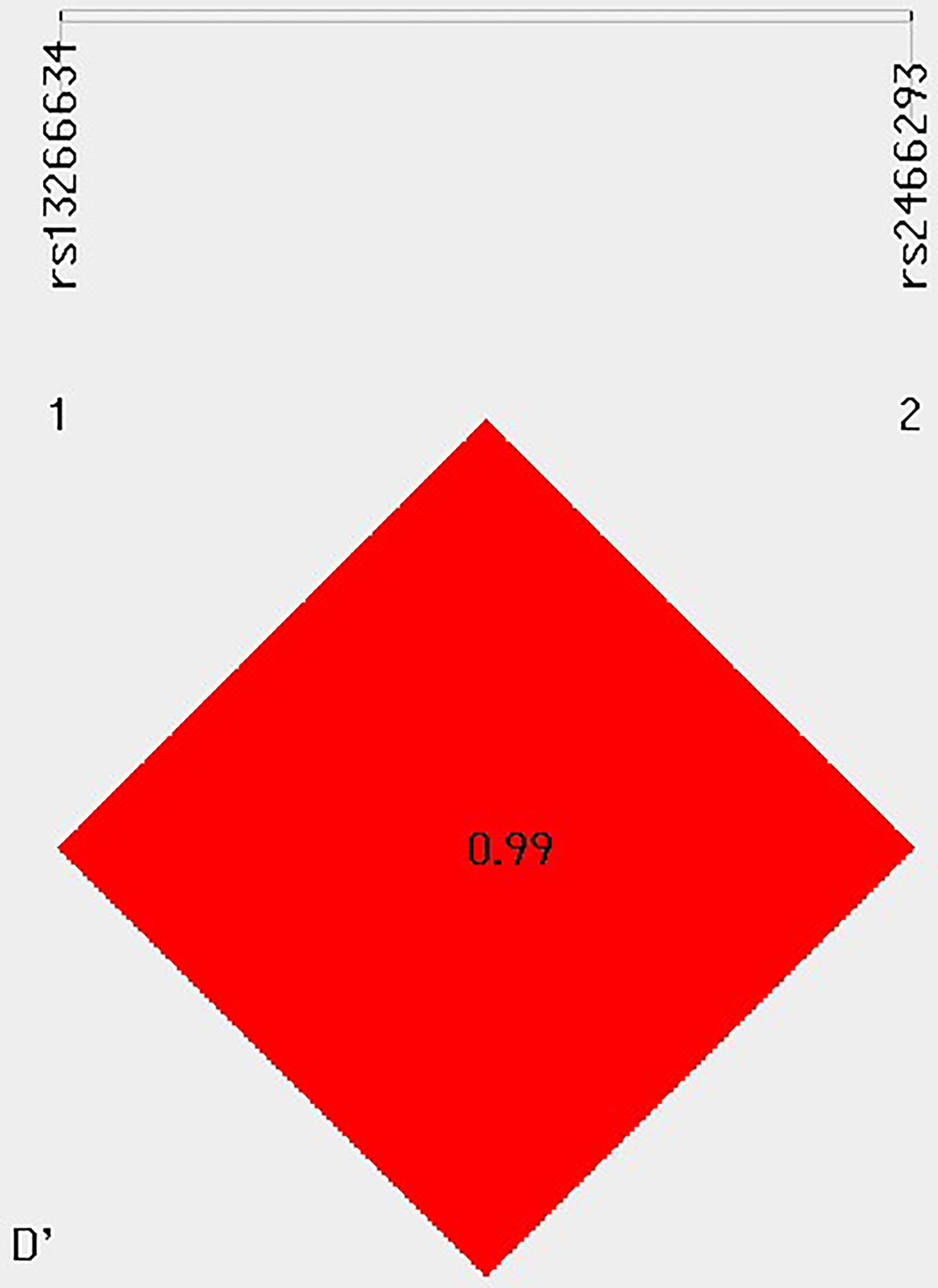
Figure 1 Linkage disequilibrium (LD) between multiple loci of the SLC30A8 gene (rs13266634 C/T and rs2466293 A/G).
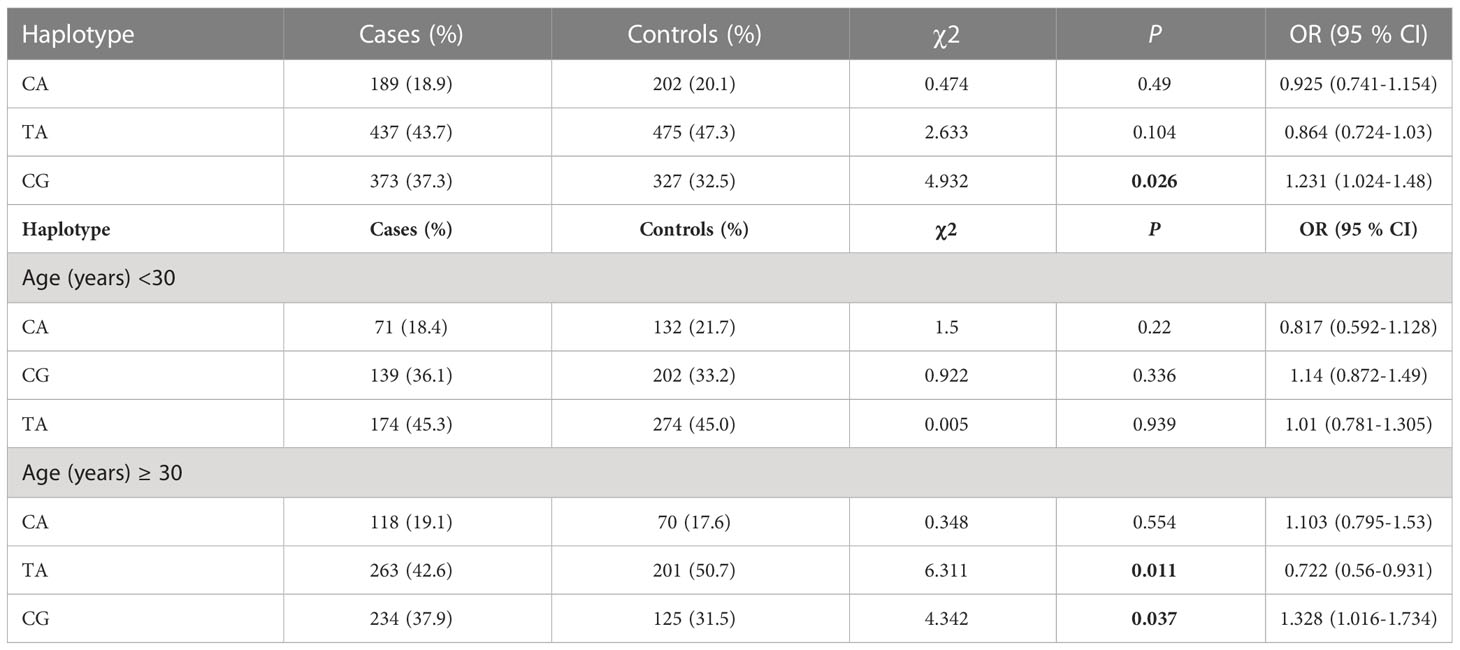
Table 5 Haplotype analysis of the rs132666342 and rs2466293 SNPs of the SLA30C8 gene for the GDM and controls.
The fasting glucose and 1-h PG levels of pregnant women with different genotypes were analyzed by age stratification (Table 6). The results showed that the glucose indexes of the rs13266634 CC genotype were higher than those of the TT genotype in subjects aged ≥ 30 years (all P < 0.05), and the 1-h PG level of the CC genotype was significantly higher than the CT genotype.
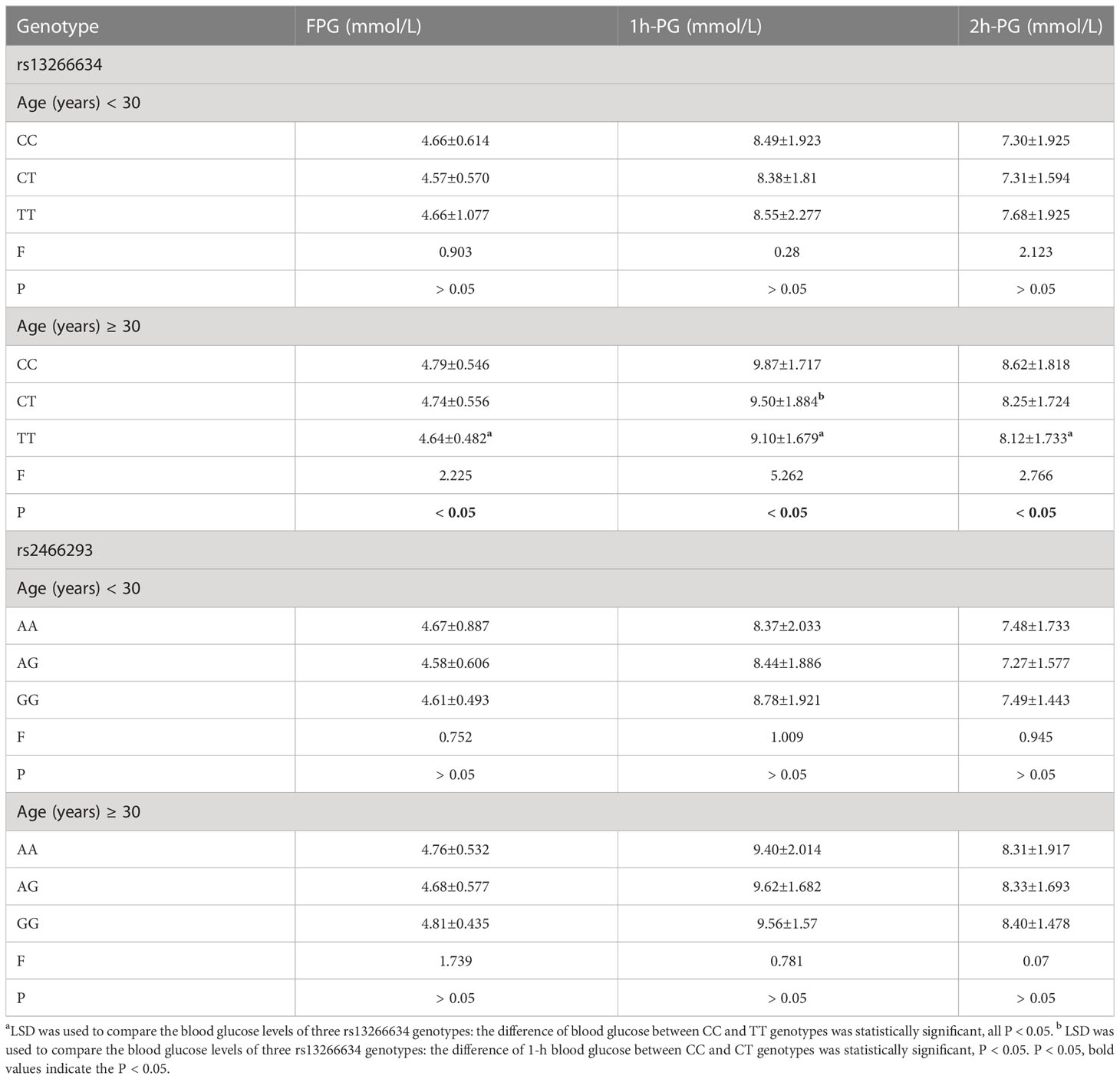
Table 6 Association between SNPs polymorphisms genotype and blood glucose levels.
Relevant references were searched for based on the PubMed and Google Scholar databases to evaluate the relationship between SLC30A8 rs13266634 or rs2466293 and GDM. Eight eligible studies were included in the rs13266634 and GDM analysis, and two studies were related to SLC30A8 rs2466293 and GDM. In total, the fixed-effects model was used for analysis. Rs13266634 was shown to be significantly associated with a decreased risk of GDM in the following models: dominant model (TT+CT vs. CC: OR = 0.751; 95% CI: 0.674-0.838; P < 0.001), recessive model (TT vs. CT+CC: OR = 0.736; 95% CI: 0.629-0.861; P < 0.001), overdominant model (CT vs. TT+CC: OR = 0.878; 95% CI: 0.789-0.977; P < 0.001), codominant homozygous model (TT vs. CC: OR = 0.643; 95% CI: 0.542-0.763; P < 0.001), codominant heterozygous model (CT vs. CC: OR = 0.789; 95% CI: 0.703-0.885; P < 0.001), and allele model (T vs. C: OR = 0.795; 95% CI: 0.734-0.860; P < 0.001) (Figure 2). In addition, SLC30A8 rs2466293 was associated with increased GDM risk in the dominant model (GG+AG vs. AA: OR = 1.184; 95% CI: 1.013-1.383; P = 0.034), recessive model (GG vs. AG + AA : OR = 1.408; 95% CI: 1.135-1.747; P = 0.002), codominant homozygous model (GG vs. AA : OR = 1.474; 95% CI: 1.167-1.861; P = 0.001), and allele model (G vs. A: OR = 1.195; 95% CI: 1.069-1.336; P = 0.002), and no significant association was found in other genetic models (Figure 3). There was no obvious evidence of publication bias in the genetic models, and these results are consistent with Egger’s tests (all P > 0.05).

Figure 2 Meta-analysis with a fixed effects model for the association between SLC30A8 rs13266634 and GDM susceptibility. (A) dominant model, TT+CT vs. CC (B) recessive model, TT vs. CT+CC (C) overdominant model, CT vs. TT +CC (D) codominant homozygous model,TT vs.CC (E) codominant heterozygous model, CT vs.CC (F) allele model, T vs. (C) OR: odds ratio, CI: confidence interval, I-squared: measure to quantify the degree of heterogeneity in meta-analyses.

Figure 3 Meta-analysis with a fixed effects model for the association between SLC30A8 rs2466293 and GDM susceptibility. (A) dominant model,GG+AG vs. AA (B) recessive model,GG vs. AG + AA (C) overdominant model, AG vs. GG +AA (D) codominant homozygous model, GG vs. AA (E) codominant heterozygous model, AG vs.AA (F) allele model, G vs. (A) OR: odds ratio, CI: confidence interval, I-squared: measure to quantify the degree of heterogeneity in meta-analyses.
The role of genetic factors in the GDM process has been verified by previous findings (12). Rs13266634 is a non-synonymous SNP in SLC30A8, and a protective role for the rs13266634 T allele, which reduces GDM risk, has been proposed in a Swedish population (8). In contrast, six studies from Brazil, the United States, Denmark, the Republic of Korea, and China failed to replicate the results (13–18). Therefore, further verification is necessary. Moreover, rs2466293 is a polymorphism in miRNA-binding sites (miR-binding SNP). Recent findings indicated that rs2466293 impacted the development of GDM (19), but more extensive research is needed for verification. This research paper conducted a case-control study to estimate the association of SLC30A8 rs13266634 or rs2466293 with GDM among six different genetic models in a Chinese population.
In the research process, we explored the relationship between SLC30A8 gene multiformity and GDM risk. In the overall analysis, the findings indicated that SLC30A8 rs13266634 showed no association with GDM risk, but SLC30A8 rs2466293 was shown to be significantly related to increased GDM risk under the dominant (GG+AG), codominant homozygous (GG), and allele(G) genetic models that were unadjusted and adjusted for age, pre-BMI, SBP, DBP, and parity. In this study, women with GDM were older than healthy controls. It has been pointed out that the prevalence of GDM increases with age, and the incidence is higher among women over 30 years of age (20). Therefore, further studies used a cutoff point of 30 years of age and analyzed the association between polymorphic variants and GDM after stratification by age. Interestingly, after adjusting for age, pre-pregnancy BMI, and SBP, our findings indicated that the SNP rs13266634 in SLC30A8 was found to have a protective effect against GDM risk in subjects aged ≥30 years under the recessive and homozygous dominant genetic models, while SLC30A8 rs2466293 was significantly associated with increased GDM risk in patients aged ≥30 years under the dominant and heterozygous dominant genetic models. These results are in accordance with some scholarly studies (8, 13, 19). Furthermore, the CG haplotype, comprised of SNPs rs13266634 and rs2466293, was significantly associated with an increased risk of GDM in the overall analysis. In a further analysis stratified by age, the CG haplotype was also associated with an increased risk of GDM in individuals aged ≥ 30 years, while the TA haplotype was associated with a reduced risk of GDM in the same age group. These results suggest that the T allele of rs13266634 in SLC30A8 can be considered a protective factor for GDM, while the G allele of rs2466293 may be a risk factor for GDM.
Wang et al. found that the C allele of rs2466293 increased susceptibility to GDM in the Chinese population (19), which was consistent with our research findings. In addition, our study found that the TT homozygous genotype of rs13266634 and the T allele decreased the risk of developing GDM in subjects aged ≥ 30 years. Similarly, previous research has demonstrated that the T allele of rs13266634 protects against the risk of GDM in the Swedish population (8), which was consistent with our findings. Moreover, in populations of Filipinos, Swedes, Koreans, and Chinese individuals, there was evidence of an association between the C allele of SLC30A8 rs13266634 and a higher risk of GDM (8, 13, 14, 18). However, other studies have not found any association between rs13266634 and the risk of GDM in populations of Danes and Europeans (15–17). Inconsistencies in these results may be related to differences in ethnicity, environment, or limited study sample sizes. Therefore, a comprehensive meta-analysis was carried out with a larger number of different populations (ethnicities) to identify the relationship of SLC30A8 SNPs with GDM risk. Rs13266634 was demonstrated to have a protective effect in every genetic model (P < 0.05) in eight eligible studies (including our study), and significant findings of rs13266634 could also be observed in both the Caucasian and Asian subgroups. SLC30A8 rs2466293 was found to be significantly related to higher GDM risk in the relevant models (codominant homozygous and allele models) (P < 0.05) based on two Chinese population studies.
GDM and T2DM are considered to have similar pathogenesis. In a study of diabetic mice, SLC30A8 gene expression levels were inhibited in the pancreas of animals with this pathology, indicating that it is related to diabetes (9). Studies have shown that the SLC30A8 rs13266634 C allele is associated with glucose regulation in GWASs (21, 22). In addition, studies based on fluorescence and radiation have proposed a hypothesis that the rs13266664-T allele reduces SLC30A8 activity, which changes insulin synthesis and reduces GDM susceptibility based on this mechanism (23–25). In addition, genetic variation in the 3’UTR, a miRNA target gene, can affect the interaction between miRNA and target mRNA. We queried the rs2466293 polymorphism located using the “MirSNP” database (http://bioinfo.life.hust.edu.cn/miRNASNP/). According to the results, it can be inferred that rs2466293 creates eight and destroys three putative miRNA target sites, which may impact the expression of SLC30A8 and lead to a higher risk of GDM. However, functional research is necessary to further confirm its mechanism.
According to the abovementioned research, this study obtained a conclusion that the age and pre-BMI of the GDM group were significantly higher than those of the control group, and logistic regression analysis indicated that the increase in age and pre-BMI were important risk factors for GDM. SBP, DBP, and parity in the GDM group were significantly higher than those in the other group. It can be inferred that patients with GDM were prone to pregnancy-induced hypertension syndrome. Moreover, a previous study found that the SLC30A8 rs13266634 C allele was correlated with higher fasting glucose levels among women with gestational high BMI (26). Our study also showed that the SLC30A8 rs13266634 C allele had an influence on higher fasting glucose, 1-h, and 2-h glucose levels among pregnant women over the age of 30 years, which was similar to the results of previous studies. The SLC30A8 rs13266634 C allele may affect the normal secretion of insulin. Wang et al. found a significant relationship between the C allele of rs2466293 with higher plasma glucose (19), but no differences were found in our study. Therefore, further relevant research is necessary.
There are still several limitations in this study. First, due to the modest sample size of the GDM and control groups, future studies need to validate our observations in a larger cohort. Second, the data used in this study were insufficient, such as the lack of fasting insulin data, to accurately measure and evaluate pancreatic islet β-cell function. Finally, the study subjects were limited to Chinese individuals, and additional research is necessary to confirm our findings in diverse populations.
In conclusion, in subjects aged ≥ 30 years, SLC30A8 rs13266634 exhibited a protective relationship against GDM susceptibility, while the results indicated associations of rs2466293 with the risk of GDM. The haplotype CG was also associated with a higher risk of GDM, and the haplotype TA was associated with a lower risk of GDM in subjects aged ≥ 30 years. In general, our findings provide more clues for studying the precise mechanism of the development of GDM.
The original contributions presented in the study are publicly available. This data can be found here: PRJEB61053, ERZ16808993.
The study was agreed by the Ethics Committee of Shunde Maternal and Child Health Hospital of Guangdong Medical University. The patients/participants provided their written informed consent to participate in this study.
QZ, BT and FH contributed equally to this study. QZ, FH and BT collected clinical data and samples. QZ, XH and JH performed data analyses. QZ, YW and RG wrote the manuscript. JH and YW supervised the whole research. All authors contributed to the article and approved the submitted version.
Support from the National Natural Science Foundation of China (81873649); Doctoral scientific research Initiate funding project of Shunde Women and Children’s Hospital of Guangdong Medical University (Maternity and Child Healthcare Hospital of Shunde Foshan) (2020BSQD007); Medical Research Project of Foshan Health Bureau (20210289); Guangdong Medical University Research Foundation (GDMUM2020008); Youth Talent Project of Shunde Women and Children’s Hospital of Guangdong Medical University (Maternity and Child Healthcare Hospital of Shunde Foshan) (2023QNRC023).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2023.1159714/full#supplementary-material
1. Ferrara A. Increasing prevalence of gestational diabetes mellitus: a public health perspective [published correction appears in diabetes care. Diabetes Care (2007) 30 Suppl 2:S141–6. doi: 10.2337/dc07-s206
2. Zhu Y, Zhang C. Prevalence of gestational diabetes and risk of progression to type 2 diabetes: a global perspective. Curr Diabetes Rep (2016) 16(1):7. doi: 10.1007/s11892-015-0699-x
3. Chen L, Magliano DJ, Zimmet PZ. The worldwide epidemiology of type 2 diabetes mellitus–present and future perspectives. Nat Rev Endocrinol (2011) 8(4):228–36. doi: 10.1038/nrendo.2011.183
4. Martin AO, Simpson JL, Ober C, Freinkel N. Frequency of diabetes mellitus in mothers of probands with gestational diabetes: possible maternal influence on the predisposition to gestational diabetes. Am J Obstet Gynecol (1985) 151(4):471–5. doi: 10.1016/0002-9378(85)90272-8
5. Huopio H, Cederberg H, Vangipurapu J, Hakkarainen H, Pääkkönen M, Kuulasmaa T, et al. Association of risk variants for type 2 diabetes and hyperglycemia with gestational diabetes. Eur J Endocrinol (2013) 169(3):291–7. doi: 10.1530/EJE-13-0286
6. Chimienti F, Devergnas S, Pattou F, Schuit F, Garcia-Cuenca R, Vanderwalle B, et al. In vivo expression and functional characterization of the zinc transporter ZnT8 in glucose-induced insulin secretion. J Cell Sci (2006) 119(Pt 20):4199–206. doi: 10.1242/jcs.03164
7. Maruthur NM, Clark JM, Fu M, Linda Kao WH, Shuldiner AR. Effect of zinc supplementation on insulin secretion: interaction between zinc and SLC30A8 genotype in old order Amish. Diabetologia (2015) 58(2):295–303. doi: 10.1007/s00125-014-3419-1
8. Dereke J, Palmqvist S, Nilsson C, Landin-Olsson M, Hillman M. The prevalence and predictive value of the SLC30A8 R325W polymorphism and zinc transporter 8 autoantibodies in the development of GDM and postpartum type 1 diabetes. Endocrine (2016) 53(3):740–6. doi: 10.1007/s12020-016-0932-7
9. Seman NA, Mohamud WN, Östenson CG, Brismar K, Gu HF. Increased DNA methylation of the SLC30A8 gene promoter is associated with type 2 diabetes in a Malay population. Clin Epigenet (2015) 7(1):30. doi: 10.1186/s13148-015-0049-5
10. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell (2004) 116(2):281–97. doi: 10.1016/s0092-8674(04)00045-5
11. Ryan BM, Robles AI, Harris CC. Genetic variation in microRNA networks: the implications for cancer research [published correction appears in nat rev cancer. Nat Rev Cancer (2010) 10(6):389–402. doi: 10.1038/nrc2867
12. Tsai PJ, Roberson E, Dye T. Gestational diabetes and macrosomia by race/ethnicity in Hawaii. BMC Res Notes (2013) 6:395. doi: 10.1186/1756-0500-6-395
13. He M, Ban B, Bian D, Li P, Zhang M, Man D, et al. Association of rs13266634 polymorphism in zinc transporter solute carrier family 30-member 8 gene with gesta-tional diabetes mellitus. Chin J Diabetes Mellit (2013) 5(1):38–43.
14. Benny P, Ahn HJ, Burlingame J, Lee MJ, Miller C, Chen J, et al. Genetic risk factors associated with gestational diabetes in a multi-ethnic population. PloS One (2021) 16(12):e0261137. doi: 10.1371/journal.pone.0261137
15. Teleginski A, Welter M, Frigeri HR, Réa RR, Souza EM, Alberton D, et al. Leptin (rs7799039) and solute carrier family 30 zinc transporter (rs13266634) polymorphisms in Euro-Brazilian pregnant women with gestational diabetes. Genet Mol Res (2017) 16(1). doi: 10.4238/gmr16019515
16. Stuebe AM, Wise A, Nguyen T, Herring A, North KE, Siega-Riz AM. Maternal genotype and gestational diabetes. Am J Perinatol (2014) 31(1):69–76. doi: 10.1055/s-0033-1334451
17. Lauenborg J, Grarup N, Damm P, Borch-Johnsen K, Jørgensen T, Pederson O, et al. Common type 2 diabetes risk gene variants associate with gestational diabetes. J Clin Endocrinol Metab (2009) 94(1):145–50. doi: 10.1210/jc.2008-1336
18. Cho YM, Kim TH, Lim S, Choi SH, Shin HD, Lee HK, et al. Type 2 diabetes-associated genetic variants discovered in the recent genome-wide association studies are related to gestational diabetes mellitus in the Korean population. Diabetologia (2009) 52(2):253–61. doi: 10.1007/s00125-008-1196-4
19. Wang X, Li W, Ma L, Ping F, Liu J, Wu X, et al. Investigation of miRNA-binding site variants and risk of gestational diabetes mellitus in Chinese pregnant women. Acta Diabetol (2017) 54(3):309–16. doi: 10.1007/s00592-017-0969-y
20. Murgia C, Berria R, Minerba L, Sulis S, Murenu M, Portoghese E, et al. Risk assessment does not explain high prevalence of gestational diabetes mellitus in a large group of sardinian women. Reprod Biol Endocrinol (2008) 6:26. doi: 10.1186/1477-7827-6-26
21. Sladek R, Rocheleau G, Rung J, Dina C, Shen L, Serre D, et al. A genome-wide association study identifies novel risk loci for type 2 diabetes. Nature (2007) 445(7130):881–5. doi: 10.1038/nature05616
22. Scott LJ, Mohlke KL, Bonnycastle LL, Willer CJ, Li Y, Duren WL, et al. A genome-wide association study of type 2 diabetes in finns detects multiple susceptibility variants. Science (2007) 316(5829):1341–5. doi: 10.1126/science.1142382
23. Carvalho S, Molina-López J, Parsons D, Corpe C, Maret W, Hogstrand C. Differential cytolocation and functional assays of the two major human SLC30A8 (ZnT8) isoforms. J Trace Elem Med Biol (2017) 44:116–24. doi: 10.1016/j.jtemb.2017.06.001
24. Kim I, Kang ES, Yim YS, Ko SJ, Jeong SH, Rim JH, et al. A low-risk ZnT-8 allele (W325) for post-transplantation diabetes mellitus is protective against cyclosporin a-induced impairment of insulin secretion. Pharmacogenom J (2011) 11(3):191–8. doi: 10.1038/tpj.2010.22
25. Nicolson TJ, Bellomo EA, Wijesekara N, Loder MK, Baldwin JM, Gyulkhandanyan AV, et al. Insulin storage and glucose homeostasis in mice null for the granule zinc transporter ZnT8 and studies of the type 2 diabetes-associated variants. Diabetes (2009) 58(9):2070–83. doi: 10.2337/db09-0551
Keywords: gestational diabetes mellitus, solute carrier family 30 A8 zinc transporter, SNP, rs13266634, rs2466293, case-control study
Citation: Zeng Q, Tan B, Han F, Huang X, Huang J, Wei Y and Guo R (2023) Association of solute carrier family 30 A8 zinc transporter gene variations with gestational diabetes mellitus risk in a Chinese population. Front. Endocrinol. 14:1159714. doi: 10.3389/fendo.2023.1159714
Received: 06 February 2023; Accepted: 12 May 2023;
Published: 31 May 2023.
Edited by:
Tarunveer Singh Ahluwalia, Steno Diabetes Center Copenhagen (SDCC), DenmarkReviewed by:
Gopi Sundaramoorthy, Madras Diabetes Research Foundation, IndiaCopyright © 2023 Zeng, Tan, Han, Huang, Huang, Wei and Guo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jinzhi Huang, aHVhbmdqemdkQDE2My5jb20=; Yue Wei, d2VpeXVlMTM4QDE2My5jb20=; Runmin Guo, cnVubWluLmd1b0BnZG11LmVkdS5jbg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.