
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Endocrinol. , 21 April 2020
Sec. Neuroendocrine Science
Volume 11 - 2020 | https://doi.org/10.3389/fendo.2020.00195
This article is part of the Research Topic Neuroendocrine-Immunological Interactions in Health and Disease View all 26 articles
 Samantha Alvarez-Herrera1†‡
Samantha Alvarez-Herrera1†‡ Raúl Escamilla2†‡
Raúl Escamilla2†‡ Oscar Medina-Contreras3†
Oscar Medina-Contreras3† Ricardo Saracco2†
Ricardo Saracco2† Yvonne Flores2†
Yvonne Flores2† Gabriela Hurtado-Alvarado4†
Gabriela Hurtado-Alvarado4† José Luis Maldonado-García1†
José Luis Maldonado-García1† Enrique Becerril-Villanueva1†
Enrique Becerril-Villanueva1† Gilberto Pérez-Sánchez1†
Gilberto Pérez-Sánchez1† Lenin Pavón1*†
Lenin Pavón1*†Atypical antipsychotics (AAP) or second-generation antipsychotics are the clinical option for schizophrenia treatment during acute psychoses, but they are also indicated for maintenance during lifetime, even though they are being used for other psychiatric conditions in clinical practice such as affective disorders and autism spectrum disorder, among others. These drugs are differentiated from typical antipsychotics based on their clinical profile and are a better choice because they cause fewer side effects regarding extrapyramidal symptoms (EPS). Even though they provide clear therapeutic benefits, AAP induce peripheral effects that trigger phenotypic, functional, and systemic changes outside the Central Nervous System (CNS). Metabolic disease is frequently associated with AAP and significantly impacts the patient's quality of life. However, other peripheral changes of clinical relevance are present during AAP treatment, such as alterations in the immune and endocrine systems as well as the intestinal microbiome. These less studied alterations also have a significant impact in the patient's health status. This manuscript aims to revise the peripheral immunological, endocrine, and intestinal microbiome changes induced by AAP consumption recommended in the clinical guidelines for schizophrenia and other psychiatric disorders.
Antipsychotics have been widely used in clinical psychiatry and neuroscience research for over 68 years since chlorpromazine demonstrated sedative effects in psychotic patients (1). Antipsychotics drugs are classified as typical or atypical according to the clinical effects that they cause (2). Atypical (AAP) or second-generation antipsychotics (SGA) are effective against positive and negative symptoms and improve some domains of cognition of schizophrenia. AAP are the first clinical option to treat various psychiatric conditions because they produce significantly fewer EPS and pose a lower risk of pseudo-parkinsonism and catalepsy in comparison to typical antipsychotics (3, 4).
Even though AAP were initially prescribed for psychotic disorders like schizophrenia, the Food and Drug Administration (FDA) has approved the use of these drugs for the treatment of other psychiatric conditions, including bipolar disorder, major depressive disorder with psychotic features, acute agitation, Tourette syndrome, borderline personality disorder, dementia, and substance-induced psychotic disorder (5) as well as diagnosed psychiatric conditions in children (6).
The pharmacological and adverse effects related to AAP consumption are due to the affinity of these drugs to a broad range of neurotransmitter receptors located in the CNS, peripheral organs, tissues, and cells. Each AAP has its own unique affinity pattern that generates psychiatric and peripheral effects acting at dopamine (DA) D1, D2, D3, D4, adrenergic α-1 and α-2, serotoninergic 5-HT2A and 5-HT2C, histaminergic, and muscarinic receptors (7). Despite their enormous efficacy on psychiatric symptoms and their low rate of EPS, AAP are not without adverse side effects. It is well-known that AAP produce peripheral effects related with metabolic alterations (8) like weight gain, type 2 diabetes, dyslipidemia, and subsequent cardiovascular complications (9, 10).
However, AAP consumption induces other peripheral changes that are clinically relevant but commonly dismissed, such as alterations in the immune and endocrine function as well as the intestinal microbiome. These sets of changes play a significant role in triggering inflammatory and metabolic chronic changes that affect the adequate recovery of patients and their quality of life.
A wide variety of hormones show alterations in their circulatory levels in human and animal models during AAP consumption. Among the affected hormones are those related to glucose metabolism, orexigenic and anorexigenic molecules, and hormones secreted by the hypothalamus or pituitary (11–14).
In patients and experimental models, AAP consumption modifies leukocyte phenotype, and cell count. The evidence demonstrates that macrophages (MQs), dendritic cells (DCs), T and B lymphocytes, neutrophils, and other leukocytes modify their function as well as cytokine production and release, apoptosis, phagocytosis, and Th1-Th2 differentiation (15–17). Additionally, other reports show AAP can change the peripheral levels of pro-inflammatory, anti-inflammatory, and growth factors molecules like C-reactive protein (CRP), interleukin (IL)-1β, IL-6, IL-12, IL-10, tumor necrosis factor (TNF)-α, interferon (IFN)-γ, and other molecules, affecting the systemic condition of the organism (18, 19).
Changes in hormonal and inflammatory levels in patients that consume AAP impact intestinal microbiota. It is important to note that the growing evidence suggests the intestinal microbiome could be involved in the treatment response. Moreover, gut microorganisms might be necessary to the occurrence of adverse effects such as weight gain (20).
In this review, we summarize the clinical and experimental studies that demonstrated the immunological, endocrine, and intestinal microbiome changes induced by the consumption of each AAP approved by the FDA for the treatment of schizophrenia and other psychiatric disorders.
Antipsychotic therapy prevails as a standard and fundamental component for major psychiatric disorders like acute episode of psychoses and the maintenance phase of schizophrenia and schizoaffective disorders (21). These drugs do not act exclusively on the CNS, and the more evident problem related with AAP treatment is the higher risk of developing hyperphagia, hyperglycemia, dyslipidemia, weight gain, diabetes mellitus, and insulin resistance (22), which further develops metabolic and cardiac complications with subsequent reduction in life expectancy, poor patient compliance, and sudden death (4).
Daily clinical practice not only identifies metabolic problems but also three clinically relevant issues that are the result of the chronic consumption of AAP by patients (23), often disregarded by healthcare professionals. The first are the endocrine and immune effects that AAP cause in patients and that will be listed in subsequent sections. The second is that the effects of the cotreatment with AAP and psychiatric medication, despite being more common than which is acknowledged (approximately 66% of psychiatrists use AAPs in combination), (24). The most common causes of co-treatment are: Patients seemed resistant to treatment instead of a monotherapy assay with clozapine or had been diagnosed with two or more psychiatric diagnoses, the clinician overlapped one antipsychotic while another was titrated (switching medication because of lack of response or better security profile), and finally, an effective dose of an AP was not achieved because of intolerance or side effects. Finally, the third issue is the joint effect of AAP consumption together with other drugs as benzodiazepines (adjunctive therapy for acute agitation, comorbid anxiety, or distress), antidepressants (as the adjunctive therapy in schizophrenia for persistent negative symptoms, comorbid major depressive disorder, and suicide risk) (25), APs also are used as adjunctive therapy for treatment-resistant major depressive disorder and major depressive disorder with psychotic features, and mood stabilizers as adjunctive therapy in bipolar disorder, schizoaffective disorder, and ultra-resistant schizophrenia (26).
This evidences the wide therapeutic use of AAP, which have become first-choice drugs to treat schizophrenia and other psychoses due to the lower risk of developing EPS. However, these drugs are highly promiscuous in their interaction with several neurotransmitter receptors as 5-HT and D, expressed in different peripheral cell types, such as leukocytes and gland cells (27–30), which constitutively express these receptors AAP can bind to.
To understand the diversity of effects these drugs systemically induce, we must consider that AAP do not behave equivalently, as shown in Figure 1. They are structurally heterogeneous, and the therapeutic effects, albeit generally equivalent, have particularities firstly explained by their heterogeneous physicochemical interactions with several receptors (see Table 1). The interactions between these drugs and their receptor firstly induces the conformational changes within the receptor structure that result in the activation of the associated heterotrimeric G protein (GPCR) and its consequent activation (53).
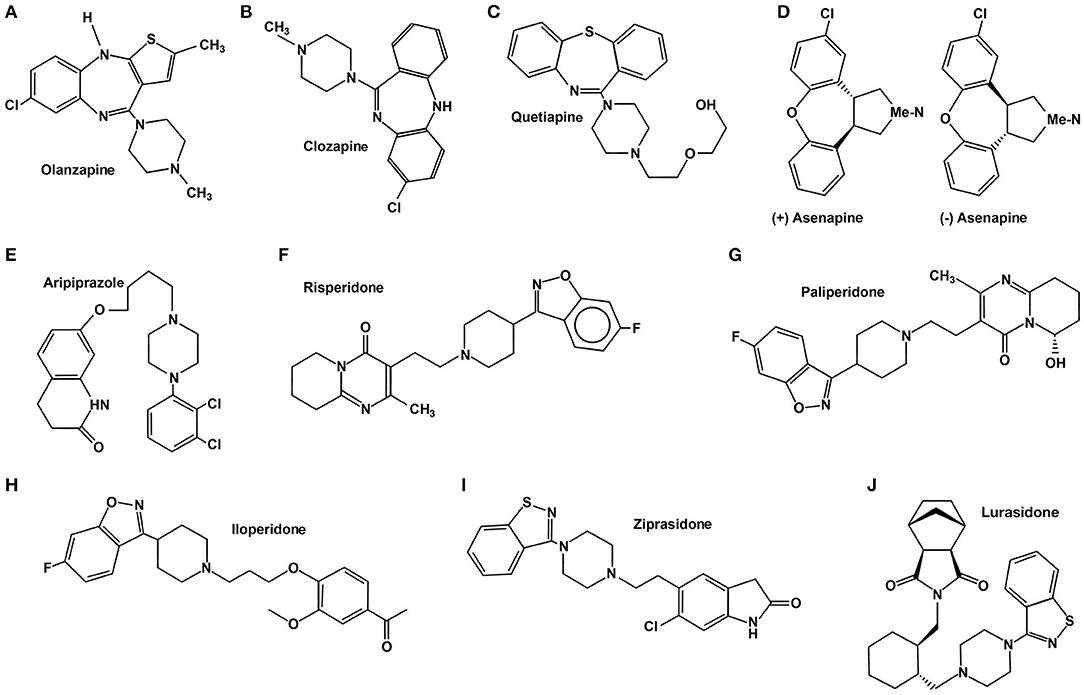
Figure 1. Chemical structure of atypical antipsychotics.
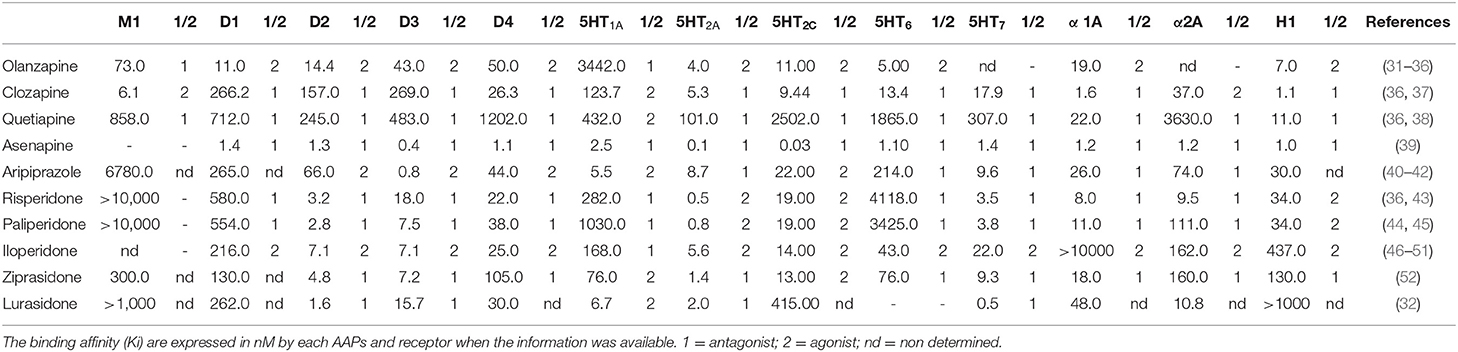
Table 1. Characteristics of APPs interaction with different neurotransmitter receptors.
In addition, over the past decade new mechanisms associated with GPCR function have been discovered, such as the ability of β-arrestins to act as multifunctional proteins and activate multiple mediators like ERK, proto-oncogene tyrosine-protein kinase SRC, nuclear factor-κB, and phosphoinositide 3-kinase (54). The capacity of a ligand to preferentially activate either G protein-dependent signaling or G protein-independent signaling is called “biased agonism” or “functional selectivity.” This innovative new concept reflects the heterogeneity and complexity of the different receptor conformation states it can be transitioning when specifically interacting with stimulants (55). In addition, recent data have demonstrated how receptor functional selectivity is a dynamic and adaptable process, which can also be modified by physiopathological conditions (56).
In addition, it must be considered that, in cases such as cotreatment, the combined effect of two or more AAP or polypharmacy can induce a phenomenon called Convergence of signaling pathways occurring in cells. It can change the overall outcome of signals initiated with different relative strengths of signal, while initial events at the cell membrane may also pose differential consequences for the whole cell (57). These phenomena are reflected in the fluctuations of soluble mediators such as cytokines and hormones in AAP consumers.
In this section we will exhibit the specific immunoendocrine peripheral effects of the 10 most prescribed APs drugs approved by the FDA: olanzapine, clozapine, quetiapine, asenapine, aripiprazole, risperidone, paliperidone, iloperidone, ziprasidone, and lurasidone.
Clozapine was the first AAP developed in 1958 and it was approved by the FDA in 1989 after 31 years of investigations and clinical trials for treatment-resistant schizophrenia (58). Currently, it is considered to be one of the most effective antipsychotics for the treatment of schizophrenia, psychosis, and depression. Nevertheless, it is not the first-line drug of choice due to its range of adverse effects, making compliance an issue for many patients (59).
Clozapine is often discontinued (18) since it has some other potentially dangerous and life-threatening side effects, such as myocarditis, seizures, agranulocytosis, or granulocytopenia, and gastrointestinal hypomotility. It is a 5-HT2A and D4 receptor antagonist. It also shows affinity to D1, D2, D3, D5, α-adrenergic, histaminergic H1, and cholinergic receptors (37) (see Table 1). This fact hinders the understanding of its molecular mechanisms of action and the identification of drug response predictors (60). In addition, this drug is an antagonist of other receptors, such as H1, 5-HT2C, and M3, leading to weight gain and metabolic side effects (61) that include both glycemic dysregulation and insulin resistance (62) (see Table 1).
Clozapine may have several interactions with other drugs as it is metabolized by the hepatic cytochrome P450 (CYP) system. Clozapine is transformed into norclozapine by CYP3A4 and 1A2 and clozapine N-oxide by CYP3A4. Nevertheless, CYP2C19 is also significant at clozapine therapeutic concentration (24%) while the influence of CYP2C9 (12%) and 2D6 (6%) is more modest. Then, blood-level monitoring of clozapine may be needed when inhibitors (such as antifungals, oral contraceptives, fluvoxamine, ciprofloxacin, caffeine, and disulfiram) or inducers (such as rifampicin, omeprazole, phenytoin, phenobarbital, and tobacco smoke) of CYP1A2 and both inhibitors (such as cimetidine, erythromycin, and clarithromycin) and inducers (as carbamazepine and rifampicin) of CYP13A4 are used. It is important to note that tobacco smoking may affect clozapine metabolism through CYP1A2 induction (59).
Endocrine alterations induced in animal models by clozapine administration have been observed at doses of 1–10 mg/kg (63), 7.5 mg/kg (64), 10 mg/kg (65), and 2–20 mg/kg (66). Additionally, clozapine significantly increased leptin levels (67) due to its affinity to M3 receptors, which have been linked to decreased insulin released by β-cells (63, 68), regulated glucose homeostasis, and body weight (69, 70). Indeed, studies have shown that only olanzapine and clozapine have a substantial affinity to M3 receptors. In this sense, the overall increase in leptin levels and its association with BMI suggest that leptin acts as a negative feedback signal in the event of fat increase (63).
A potential metabolic impairment by clozapine via the hypothalamic insulin signaling pathway has been reported in vitro. Using mHypoE-46 and rHypoE-19 neuron cell lines, clozapine impaired insulin-induced phosphorylation of AKT (63). Although clozapine is known to inhibit 5-HT2AR signaling through G protein-dependent mechanisms, it differs from classic GPCR antagonists in that it also induces 5-HT2AR internalization and activates AKT signaling through a 5-HT2AR-mediated event (71). An animal model, where this drug (2.5, 5, 10 mg/kg) was applied intravenously to Wistar rats, showed an acute increase in corticosterone and glucagon levels, which explains the establishment of hyperglycemia (72).
The vasoactive intestinal peptide (VIP) of parasympathetic origin may contribute to clozapine (muscarinic M1-receptor)-induced sialorrhea, an adverse effect created by its synergistic interaction with the antipsychotic in some patients with schizophrenia (73). Therapeutic doses of clozapine may induce reproductive dysfunction through mechanisms involving ovarian mitochondrial dysfunction and oxidative stress (74), an effect explained by the impairment of the mitochondrial respiratory chain. This phenomenon is supported by a study with thirty adult female albino rats that received clozapine (20 mg/kg/day) for 28 days. It was observed that reduced complex I activity (25%) resulted in a 35% decrease in ATP and mitochondrial respiration, thus severely impairing energy production and leading to apoptosis (75).
Resistin is a biomarker of systemic inflammation and likely plays a role as a marker of cardiovascular comorbidity. A study showed that Clozapine (40 μM) inhibited resistin mRNA expression in mouse brown adipocytes (76). However, in 121 schizophrenia patients treated with clozapine (403 mg/day), high serum levels of resistin were associated with smokers in comparison with non-smokers (77).
The interaction between clozapine and its pharmacological target in leukocytes induced inflammatory alterations in cell lines, primary cultures, animal models, and humans. The principal immune alteration associated with clozapine is agranulocytosis (neutrophils <500 cells/mm3), the most severe form of leukopenia affecting approximately 3.9% of users, and others as neutropenia (neutrophil count <1,500 cells/mm3) (78). The risk of neutropenia/agranulocytosis is 0.38% approximately with monitoring and 2.5% without it (79). This was mainly observed in female patients and was directly associated with the time of clozapine use (37, 80). Clozapine by itself was not directly toxic to neutrophils or their progenitors at therapeutic concentrations (79). However, the bioactivation/oxidation of clozapine in neutrophils produced reactive and unstable clozapine metabolites, which induced toxic oxidative stress, leading to neutrophil apoptosis. Metabolites may be cytotoxic to bone marrow stroma, potentially leading to accelerated neutrophil or myelocyte precursor apoptosis (81, 82). An associated genetic susceptibility was detected in 31 patients who developed clozapine-induced agranulocytosis and 38 patients who developed neutropenia in a group of 310 clozapine users. The most significant association was found with mutation NQO2 G1541A, making it one of the candidate markers for the prediction of these adverse effects (83).
The current pharmacovigilance processes, carried out worldwide, have allowed for the identification of three uncommon cases of clozapine-induced drug reaction with eosinophilia and systemic symptoms (84). These three reported cases took place in adults older than 57 years, all of them consuming different drugs previously. Two patients were diagnosed with acute exacerbation of a chronic paranoid schizophrenia and the third presented schizoaffective disorder. In the three cases, 15–22 days into the treatment (200–400 mg/day), the blood levels of clozapine were within the toxic range, while eosinophilia, leukocytosis, and liver abnormalities were detected along with a significant increase in CRP without infection (85, 86).
The phenomenon observed in these patients was secondary to the inflammatory process leading to an increase in circulatory levels of clozapine. It is known that cytokines can inhibit the metabolism of clozapine through cytochrome 4501A2 inhibition (87).
Clozapine is also associated with changes in lymphocyte phenotype and differentiation as well as changes in cytokine secretion. Some evidence showed that clozapine primarily inhibited the expression of 5-HT2A/2C on the membrane of primary T cell cultures and Jurkat and CEM cell lines (29, 72). Additionally, it is known that clozapine in vitro (1.5–7.5 μg/mL) inhibits Th1 differentiation by preventing the expression of transcription factor T-bet but not that of STAT-4 in T cells; clozapine also inhibited Th1 differentiation by blocking the AKT activation pathway (88). Moreover, clozapine (20 μM) promoted the in vitro differentiation of Treg cells and the expression of Foxp3 in splenocytes and lymph node cells from C57BL6/J mice in a model of experimental autoimmune encephalomyelitis (EAE) (15).
Regarding the effects of clozapine in MQs it has been described that clozapine increased IL-10 production and decreased IL-12 secretion in MQs after 5 days of incubation and when it is stimulated with lipopolysaccharide (LPS) for 24 h (89). Similarly, clozapine (10–100 μM) reduced nitric oxide (NO) and IL-12p40 production by LPS-stimulated bone marrow-derived macrophages (BMDM) from female C57BL/6 mice (90).
In studies carried out in animal models, similar effects to those described above in cell cultures have been observed in MQs. In a perinatal phencyclidine rat model, the administration of clozapine increased IL-6 and TNF-α with sex-specific changes (91), which can fit in the theory of “cytokine signature” observed in blood leukocytes from healthy volunteers incubated with clozapine (1 μM) (92). It has also been reported that, in Wistar rats, clozapine (45 mg/kg/day) induced myocarditis related with lymphocytic infiltrates, which induced the release of reactive oxygen species (ROS), cytokines, and TNF-α (93). Additionally, a perinatal model of 90 day-old Wistar rats prenatally treated with LPS reported that daily clozapine (10 mg/kg) significantly reduced IL-1β, TNF-α, and IL-2 levels (60, 94).
Finally, consideration should be given to the changes in the profile of circulating cytokines induced by clozapine consumption. For instance, nine patients with diagnosed schizophrenia or schizoaffective disorder, who were treated with clozapine 100–400 mg/day, showed increased risk of developing fever after the first intake, and IL-6 might play a specific role in the interaction effect between treatment duration and fever development (94, 95). Clozapine has also been shown to increase soluble IL-2 receptor (sIL-2R) and IL-6 levels (96, 97). Similarly, the adipokine resistin was associated with several acute and chronic inflammatory states and promoted the expression of TNF-α and IL-6 by human mononuclear cells (97) (see Table 2).

Table 2. Immunoendocrine peripheral effects induced by atypical antipsychotics.
Risperidone was the second AAP approved by the FDA and is among the most prescribed worldwide (98). Its use was authorized for the treatment of schizophrenia in 1993; it was approved to treat acute manic or mixed episodes of bipolar I disorder as monotherapy or adjunctive drug in 2003 and autism-related irritability in 2006 (99). There are also many varied non-FDA approved uses for risperidone, such as Tourette syndrome (100), major depressive disorder (MDD) (101), anorexia nervosa (102), dementia (103), borderline personality disorder (104), Parkinson's disease psychosis (105), posttraumatic stress disorder (106), and some other psychiatric conditions (107). This drug has liver metabolism; it mainly undergoes 9-hydroxylation that produces active 9-hydroxy-risperidone (OH-RIS) metabolite by CYP2D6 and CYP3A4 to a lesser extent (107). The available formulations of risperidone in the market are oral solution formulation, oral disintegrating tablets, and long-acting injectable (LAI) formulation (108).
The presumed action mechanism of risperidone is associated with the combination of 5-HT2A agonist and D2 antagonist effects with a strong binding affinity for the first one (see Table 1) (109). This drug is also active as an antagonist for other receptors with a lower affinity, such as dopamine D3 and D4, serotonin 5-HT6, 5-HT7, and α-1 and α-2 adrenergic. Risperidone acts as agonist on 5-HT2C serotoninergic receptors and H1 histaminergic receptors (43).
Risperidone consumption has demonstrated to generate hormone alterations in human and animal models. Reports on this AP refer to hormones related with glucose metabolism, adipokines, appetite, and those linked to the adrenal, and gonadal axes, among others. One of the main effects associated with risperidone consumption is elevated PRL levels in patients (110), having a significant incidence of HPRL compared with other APs (111–113). In human and adult studies, the drug caused significant PRL elevation after 44 days of treatment (1–6 mg/day) in 27 of 37 schizophrenic patients, while in a 1 year follow-up (1–6 mg/day) 6 out of 20 patients reported HPRL with decreased PRL, without reaching baseline levels (114). These results were similar to those obtained by Perez-Islas et al., whose report showed that 90% of the men and 87% of the women in the study had PRL levels above the reference range 3 months into the treatment (unspecified dose). These levels were still elevated in 70% of the subjects at 1 year of follow-up with a tendency to decrease (115).
During an acute follow-up (4 weeks), the patients with an increase of more than 20% in PRL levels had a better chance of responding to risperidone (116). However, the chronic consumption gave more information on PRL concentration and its effects; the reports proved that risperidone is associated with chronic HPRL (117). Female patients presented a significant incidence of HPRL as compared with their male counterparts (118). Elevated PRL by risperidone consumption could be associated to higher concentrations of osteocalcin in both genders (119), breast symptoms, discomfort, menstrual changes, and erectile dysfunction (120). In fact, patients who consumed risperidone and showed menstrual disorders had a significant increase in serum PRL levels, showing a correlation between the incidence of elevated PRL and menstrual disorders (121). Other organic and rare alterations, in adult and/or children patients, as granulomatous mastitis (122), amenorrhea (123), galactorrhea (124), acute pancreatitis (125), and pituitary adenoma (126) have also been associated with risperidone consumption.
The reports on PRL levels in children and adolescent patients have shown that these populations present elevated HPRL, which has been reported in 44.9% of autism spectrum disorder (ASD) patients as unrelated to the duration of risperidone treatment (0.25–5 mg/kg, 1.03–158.03 months) (127). Still, there is evidence exposing the possible relation between plasma metabolite levels of risperidone and elevated PRL concentrations (128). Patients from the same population presented an increase in serum PRL during 3 months of follow-up (0.5–4 mg/day); gender, pubertal status, risperidone dosage, psychiatry diagnosis, and personal/family history of autoimmune diseases also affected PRL elevation during treatment (129). Similarly, a meta-analysis reported that pediatric patients treated with risperidone (4–6 mg/day) were found to experience the most significant increase in PRL, followed by patients treated with 1–3 mg/day of risperidone compared with other APs at a different dose (130). In fact, the occurrence of HPRL in this population has been associated to the presence of the C allele of the rs6318 single nucleotide polymorphism (SNP) of the HTR2C gene (0.25–6 mg/day, 0.1–143 months) (131). In addition, HPRL during treatment with risperidone/paliperidone in schizophrenic patients showed an association with rs40184 and rs3863145 variants in SLC6A3 gene of blood leukocyte DNA (132). All the studies that showed elevated PRL levels in the pediatric population are in accordance with a meta-analysis that presented a relation between risperidone treatment and high PRL (130). The possible mechanism by which risperidone causes HPRL is associated with the transcriptional upregulation of neuropeptide Y (NPY) secreted by the arcuate hypothalamic nucleus due to the high affinity of risperidone to 5-HT2A receptors. NPY inhibits tyrosine hydroxylase expression in the paraventricular nucleus and thus reduces DA synthesis, which in turn would diminish the inhibition of PRL expression induced by DA. The reduction in DA would cause the overexpression of PRL in the pituitary and ultimately induce HPRL (133).
Studies have shown that, in addition to PRL increase, other hormone profiles such as estradiol, testosterone, leptin, adiponectin, and insulin could be altered during risperidone treatment. The acute consumption of this drug (2–4 mg/Kg) decreased testosterone and estradiol levels in female patients after a 6-week treatment (134), and the same decrease in estradiol levels was reported in male patients with schizophrenia during 1 year of treatment (2–6 mg/day) (135), although other studies showed that risperidone consumption did not alter the testosterone or estradiol levels in male or female patients (136, 137). Although the mechanism is not clear, risperidone could affect estradiol and testosterone levels by a direct effect on the hypothalamic-hypophysis-gonadal axis, decreasing hormone production (135).
The concentration of leptin has also been proven to increase by 60% in psychotic patients after 4 weeks of risperidone consumption (4–8 mg/day) (138). Other studies showed that the leptin levels of schizophrenic patients with risperidone consumption were higher than those of healthy controls (139, 140). Similarly, there was an increase in leptin among ASD patients during at least 12 months of treatment (0.25–1 mg/day) (141). In treatment-naïve children and adolescents, leptin increased after 3 and 6 months of treatment (unspecified dose) when compared with baseline (142). However, a 5-month treatment (6.1 ± 1.8 mg/day) yielded no changes in leptin concentrations when compared with baseline (143); this data is supported by a meta-analysis that found no significant changes in leptin after risperidone treatment (63). The serum leptin elevation is attributed to weight gain rather than the direct effect of risperidone on leptin metabolism. This hormone is secreted by adipocytes and it is proportional to the mass of stored fat, so the elevation in blood of patients with antipsychotic-induced weight gain could be the result of the increased weight itself (144).
Adiponectin is a molecule with conflicting results on the effect of risperidone consumption. Schizophrenia patients treated with risperidone (unspecified dose, 50.1 ± 82.4 months) reduced plasma levels of adiponectin when compared with healthy subjects (145), and medication-free children showed a decrease in adiponectin levels after 16 weeks of treatment (3–91 months) (146). However, these data do not match with the results of two meta-analysis that reported no association between risperidone treatment and low adiponectin (147, 148).
According with most reports, risperidone consumption increases insulin levels in blood. The treatment with LAI-risperidone (38 ± 2 mg/15 days) for 18 ± 1.6 months showed higher insulin concentrations in patients compared with the control group (140). Children studies show that the treatment for at least 12 months (0.25–1 mg/day) increased insulin levels in ASD patients (141). Similar results were obtained in medication-free children (3–91 months) after 16 weeks of treatment (146).
The changes in hormone levels related with risperidone treatment are also evident in animal models. In a pigtail macaque model, PRL was higher at low (0.025 mg/Kg) and high doses (0.05 mg/Kg) of risperidone during 4-month consumption, with a gradual decline until reaching placebo levels in the post-drug phase (149). In an animal model, male Wistar rats showed reduced α-MSH, agouti-related protein (AgRP), and cocaine- and amphetamine-regulated transcript (CART) concentrations and increased leptin levels vs. the vehicle group after 4 weeks of treatment (2 mg/kg/day) (150). In addition, female Sprague-Dawley rats with depot risperidone exhibited higher glucagon levels (20 mg/day), while daily risperidone (40 mg) increased leptin and ghrelin levels at 4 and 6 weeks (151). In rat ovarian theca cells, risperidone inhibited mitochondrial bioenergetics and steroidogenesis by reducing ATP content (0.1–100 μM, 24 h) and the production of progesterone and androstenedione (1–37 μM, 24 h) (151). Finally, mouse models have reported plasma insulin increased over 8-fold in FVB/N mice 3 h after consumption of risperidone (152).
The adverse effects, involving immune alterations, caused by risperidone have been widely studied. In the immune system, this drug alters leukocyte numbers and levels of humoral inflammatory molecules, and it directly effects the phenotype and function of leukocytes (16, 153, 154).
Risperidone does not require regular clinical monitoring of white blood cell (WBC) count; however, anecdotal evidence has shown that it could modify and reduce the leukocyte count. Different risperidone doses (2–4 mg/day) and the combined treatment of risperidone/paliperidone (2 mg/day/100 mg) caused leukopenia with neutropenia (155) or lymphopenia (153, 156–158) as well as fever (159). Other leukocyte alterations have been described. A case report showed the development of thrombocytopenia in a male paranoid schizophrenia patient with risperidone treatment (4 mg/day) (160). Risperidone has demonstrated the association between its consumption (3 mg/day) and the development of acute eosinophilic pneumonia (AEP) in a male patient under a 6-month treatment (161). All count alterations and developed diseases improved after discontinuing the drug. Nevertheless, there are reported cases of neutropenia induced by risperidone but the incidence rate of cytopenia alterations seems to be very low (162).
The reports that analyze the effect of risperidone consumption on soluble molecules with immune function are very diverse. Even though CRP levels did not show changes in schizophrenic patients during risperidone treatment vs. healthy volunteers (163), there is evidence showing the relationship between elevated CRP and the effects on risperidone metabolism. A case report demonstrated that two females who had consumed risperidone during acute inflammation indicated by elevated CRP exhibited an increase in dose-related serum concentrations of risperidone up to the therapeutic concentration (164, 165). Furthermore, high CRP vs. common CRP values increased risperidone and OH-RIS serum levels by 58.4 and 20%, respectively, in patients with risperidone consumption (166). Contrastingly, other reports showed no correlation between CRP levels and affected risperidone levels in serum concentration (95).
The studies of changes in BDNF levels during treatment are controversial; only relapse schizophrenic males patients showed elevated BDNF after 4 weeks of risperidone consumption (3–6 mg/day), a result that suggests gender should be considered when choosing the pharmacological treatment (167–169). However, other studies reported no alteration whatsoever after risperidone consumption (170, 171).
Cytokines, chemokines, and immunoglobulins (Ig) are inflammatory molecules that have been measured during risperidone treatment and results show changes in blood levels in some of them. In an animal model, risperidone decreased and normalized the plasma levels of IL-6 and TNF-α in n-3 fatty-acid deficient rats when compared with elevated levels of n-3 fatty-acid adequate rats after 40 days of treatment (3 mg/kg/day) (172).
In patients with schizophrenia, the measurement of these molecules in blood has proven the immunomodulatory effect of risperidone at different times of consumption. A 3-month treatment with risperidone (1–6 mg/day) showed significant decreases in serum levels of IL-8, macrophage inflammatory protein (MIP)-1β, fractalkine, TNF-α, IL-7, IL 13, IL-17a, IL-23, and IL-21 (173). The same effect in TNF-α was observed at 40 days of treatment (unspecified doses) with increased IL-10 serum levels in patients with risperidone or clozapine consumption (174). Elevated levels of Interleukin-1 receptor antagonist (IL-1RA) and IL-10 have also been reported after 6 weeks of treatment (unspecified doses) (175) as well as significant decreases in IL-6, IL-10, TNF-α, and IL-4 after 10 weeks of risperidone consumption (4 ± 1.8 mg/day) (176). During 6 months of risperidone treatment (2–6 mg/day), TNF-α levels increased compared with baseline while IL-1β and IL-6 decreased at 1 month and then gradually increased at the end of the follow-up (154). In ASD patients, eotaxin and monocyte chemoattractant protein-1 (MCP-1) levels significantly decreased after 8 weeks of treatment (0.5–1.5 mg/day) (177). Regarding Igs, a report showed that a 4-week treatment with risperidone increased Igγ (IgG) chain levels significantly when compared with baseline (178). Another report showed that from 17 schizophrenic children with high blood titers of platelet-associated antibodies (PAA) only two became PAA-negative following 3 years of treatment. Most of the reports above show evidence that risperidone could reduce the production of the pro-inflammatory molecules caused by psychiatric conditions and support an anti-inflammatory response.
When talking about phenotype and function alterations in immune cells, it seems that risperidone causes significant changes such as a shift in cytokine secretion, cell differentiation, adhesion and phagocytic functions, receptor expression on leukocytes, and gene expression. Firstly, this drug reduces ATP-induced platelet aggregation when platelet-rich plasma of healthy donors is incubated in vitro with risperidone (65 ng/mL) for 30 min (179). An in vitro assay showed that activated peripheral blood mononuclear cell (PBMC) from healthy adults incubated with risperidone (10−7 M, 3–5 days) reduced IFN-γ production and inhibited AKT phosphorylation and T-bet expression, causing reduced Th1 differentiation. During chronic treatment (10−7 M, 28 days), risperidone reduced IFN-γ released by CD4+ T cell subpopulation (180). Similarly, this drug affected cytokine and chemokine production of activated mature monocyte-derived dendritic cells (DCs) of healthy adults, increasing IL-10, IL-6, IL-8, and TNF-α levels and decreasing interferon γ-inducible protein−10 (IP-10) and IL-12 levels (10−7-10−5 M, 3 days). These changes in mature DCs produced a reduction in IFN-γ secretion by activated T cells, causing Th1 suppression and leading to neutrophil death (15, 17).
The increase in IL-10 levels and/or the decrease in IFN-γ production by activated PBMC with risperidone treatment were reproducible in other reports (16). Risperidone also inhibited the adhesion, phagocytosis, and ROS production by activated U937 cells (10−5-10−4 M), decreased IL-6, IL-8, and IL-12, and increased IL-10 production in healthy, stimulated human MQs in vitro (10−6-10−5 M). This effect could support the inhibition of Th1 differentiation (181, 182), although some evidence proposes that this drug suppresses inflammatory (M1 MQs, Th1 lymphocytes) and anti-inflammatory (Th2 lymphocytes, Treg) responses (90).
In the blood of schizophrenic patients, the treatment with risperidone (2–6 mg/day) for 4 weeks showed a decrease in the number of Th17 cells (183). In vitro, differentiated human adipocytes incubated with risperidone (100 ng/ml, 11 days) induced transcription factor NF-κB target genes of IL-1β and IL-8 molecules (184). In schizophrenic patients, risperidone (8 weeks, 2–6 mg/day) modifies the expression of toll-like receptors (TLR), while monocytes CD14+, CD3+CD4+Foxp3+ T, and CD3+CD4+CD25+ T cells increased TLR2 expression, and CD14+ monocytes decreased TLR4 expression (185). Effects on gene expression have been reported; in blood cells of first-episode psychosis patients, risperidone (unspecified doses, 20 days) was associated to the up-regulation of 11 immune system genes, including cytokines and cytokine receptors (SPP1, IL1R1, IL1R2), pattern recognition molecules (TLR1, TLR2, TLR6, dectin-1/CLEC7a), molecules involved in apoptosis (FAS), and BDKRB1, IGF1R, and CR1 (186).
In animal models, a 3-week treatment with risperidone (1.25 mg/Kg/day) in diabetic Wistar rats showed that this drug altered the vascular function by the significant up-regulation of vascular cell adhesion molecule-1 (VCAM-1), intercellular adhesion molecule-1 (ICAM-1), E-selectin, and MCP-1 and TNF-α in aortic tissue homogenate (187). In an experimental autoimmune encephalitis (EAE) model with C57BL/6 mice, risperidone (3 mg/kg/day) reduced the severity of the disease in a dose-dependent manner and down-regulated IL-17a, IL-2, and IL-4 secretion by splenocytes at peak disease (day 15). During chronic EAE phase, risperidone significantly increased the number of splenocytes, Tregs, and CD4+ T cells and increased IFN-γ levels, showing that T cells responded differently to risperidone during the acute and chronic phases of EAE. In addition, activated BMDM of treated mice decreased IL-12 levels but increased IL-10 concentration. These cells modified T cell activation reducing IFN-γ and IL-17 production and enhancing IL-10 levels (188). In RAW 264.7, a macrophage mice line, risperidone activated these cells (20–40 μM for 24, 48, and 72 h) and increased nitric oxide (NO) levels (30–40 μM) as well as apoptosis events by modulating levels of caspases 8 and 3 (20–40 μM at 72 h). The drug also reduced Bcl-2/BAX gene expression ratio (24 h) and, contrary to the above data, increased IL-1, IL-6, TNF-α, and IFN-γ and decreased IL-10 production in a dose-dependent manner. Results of the RAW 264.7 line could show that the continued activation of MQs likely contributes to the development of endocrine disturbances caused by risperidone (91) (see Table 2).
Olanzapine (2-methyl-4-(4-methyl-1-piperazinyl)-10H-thieno[2,3-b][1,5]benzodiazepine) belongs to the thienobenzodiazepine class and is structurally similar to clozapine (see Figure 1) (189). It was authorized for the treatment of schizophrenia in 1996 and bipolar I acute manic or mixed episodes in 2000 (43, 109, 190). There are also many varied non-FDA-approved uses for quetiapine, such as dementia-related behavioral problems, bipolar depression, psychotic depression, SSRI-resistant major depression, personality disorders, post-traumatic stress disorder, and Tourette syndrome in children and adolescents (191). Olanzapine is metabolized in the liver by direct glucuronidation and cytochrome P450 (CYP) oxidation and generates two metabolites, 10-N-glucuronide and 4'-N-desmethylolanzapine, which lack pharmacological activity (192).
The therapeutic effect of this drug is associated to the antagonism of D2, D3, 5HT2a, and 5-HT2c receptors, although it exhibits an antagonist effect on other receptors such as 5-HT1C, 5-HT6, 5-HT7, α-1A, α-2A, H1, M1, M3, and 5-HT1A (see Table 1) (31–35).
This drug has been associated with the decrease in cell counts. There are few reports that evidence leukopenia is induced by olanzapine consumption (2.5–10 mg/day) during the first 35 days of treatment (193, 194). This phenomenon is associated with the covalent bonding between neutrophils and a reactive nitrenium ion, the oxidized form of olanzapine. It has been proposed that this reactive metabolite is responsible for the effect in neutrophils (195). Other studies have reported that this drug is associated with a decrease in eosinophils. Three reported cases showed that males with schizophrenia developed eosinophilia during olanzapine treatment (10–20 mg); in all cases, the problem was solved suspending the medication (193, 194, 196). This drug also modifies receptor expression; according to a report, 30 first-episode psychotic patients treated with olanzapine (15–25 mg/day) for 30 days showed a decrease in D1, D2, 5-HT2A, and transforming growth factor (TGF)-β mRNA expression in PBMC as well as an increase in IL-6, IL-1β, and TGF-β blood levels (197).
The increase in PRL caused by the consumption of most of the AAPs is also observed in the consumption of olanzapine. However, the evidence of this effect is contradictory since there is also proof of the decrease during the consumption of this drug. It has been reported that according to the period of consumption, the effects on PRL blood levels change (111, 198, 199): The consumption during short periods (<2 weeks) or the intake of a single dose does not alter PRL levels (111, 200).
There are two case reports of women aged 29 and 49 years with bipolar affective disorder and delusional disorder who exhibited HPRL associated with olanzapine consumption during 24 weeks (5–20 mg/day) (201). Most reports evidence the increase of this hormone: A study with 72 patients (33 women and 39 men) who were administered olanzapine (10–20 mg/day) for 3 weeks or more showed an increase in PRL levels only in female patients (202). Similar effects were detected in 49 schizophrenia patients (24 women and 25 men) treated with 15–30 mg/day of olanzapine for 4 weeks (203) and those patients (27) with chronic consumption (10–15 mg/day, 8 years) (204). In fact, a study with healthy volunteers showed that participants with no psychopathology who received one dose of olanzapine (10 mg) exhibited an increase in PRL levels (58). On the other hand, other studies showed the decrease of PRL after olanzapine consumption. In a study with 22 participants, PRL levels were reduced only in women with schizophrenia or schizoaffective disorder at 6 and 12 months of treatment (5–20 mg/day), while levels in men showed no difference (198). The decrease in PRL levels was also reported in 37 first-episode psychosis patients who consumed olanzapine (unspecified dose) for 1 year (115). The PRL elevation is associated with the interaction between olanzapine and D2 receptors on lactotroph cells. This phenomenon hampers the interaction between DA and its receptor, so DA cannot inhibit PRL production. The alterations depend on gender, genetic predisposition, dose, and time of consumption (115, 190, 201, 205).
Interestingly, the chronic administration of olanzapine is associated with the development of hyperinsulinemia and insulin resistance; the decrease in insulin sensitivity was reported in 29 healthy individuals after 10 days of olanzapine treatment (10 mg/day) (206). The increase in fasting insulin was reported in 25 schizophrenic patients with olanzapine consumption (5–20 mg/day) for 13 weeks (207). It has been postulated that alterations in insulin synthesis may be due to the stimulation of M3 receptors in β-pancreatic cells (70, 208).
It has also been described that olanzapine is associated with an increase in leptin blood levels (63), although there is little evidence that shows no changes in leptin blood levels during treatment. The increase in this hormone was shown in 18 schizophrenic male patients after 9 months of olanzapine consumption (5–20 mg/day) (209). Another study reported that 23 schizophrenic patients showed the same effect after 8 weeks of olanzapine treatment, and the increase in leptin concentration was correlated with elevated IL-1 receptor antagonist (IL-1ra) serum levels (210). In fact, the report of Tsuneyama and cols. described that the increase in leptin levels was observed only in schizophrenic female participants treated with olanzapine for 1 year (12 male and 19 female) (211). However, 12 schizophrenic patients exhibited no changes in appetite or leptin concentrations after a 5-month treatment (mean: 25 mg/day) (143). Although the mechanism by which this drug affects leptin secretion is unclear, the effect could be secondary to the interaction between the drug and H1 receptors on the hypothalamus and nucleus accumbens. Additionally, the genetic predisposition is crucial for the development of alterations associated with leptin function (212).
The data of reports on the olanzapine effect in ghrelin levels exhibit contradictions in the conclusions; some studies in schizophrenic patients show that this drug reduces ghrelin concentration after a 6-week treatment (213) and chronic consumption (8.3 ± 7.5 years, 10–20 mg/day) (214). In contrast, no changes in ghrelin levels or appetite were shown in 13 patients with schizophrenia treated with olanzapine for 5 months (143). However, an animal model with Wistar rats showed that the acute consumption of this drug (1 mg/kg) increased the concentration of postprandial ghrelin compared to controls (215). The mechanism of ghrelin alterations is not clear but these changes are associated with leptin resistance. It has been proposed that olanzapine exhibits a direct effect on hypothalamic neurocircuits that regulate ghrelin synthesis, causing an altered leptin/ghrelin ratio (212). There is only one report that measured cortisol levels during olanzapine treatment: Hahn and cols. reported that healthy individuals who received a single dose of olanzapine (10 mg) exhibited a decrease in cortisol serum levels compared with baseline (58).
Other immune alterations associated with olanzapine consumption are the modulation of cytokine secretion and production, depending on the consumption time of the drug (216). There are in vitro studies in PBMC of healthy individuals and THP-1 line (10−4 M for 72 h) that demonstrate a reduction in mRNA expression of IL-1β, IL-6, IL-10, and TNF-α, IL-6, TNF-α, and IL-10. Similarly, stimulation in THP-1 cells resulted in a significant decrease in the expression and secretion of IL-1β and TNF-α (217). Other studies in schizophrenic patients with prolonged consumption reported changes in cytokine levels. After a 24-month olanzapine treatment (unspecified dose), 95 schizophrenic patients with metabolic syndrome showed lower concentrations of BDNF (P < 0.012) and higher values of TNF-α as compared to 121 patients only diagnosed with schizophrenia (218). Also, out of 28 patients with chronic olanzapine consumption (unspecified dose) 14 were insulin-resistant and had a higher concentration of TNF-α, IL-6, IL-1β, and IL-8 with a positive correlation between these values and insulin resistance (210). Similarly, female Sprague-Dawley rats and female BALB/c mice, after 8 weeks of treatment (10 mg/kg/day), exhibited a significant increase in TNF-α, IL-6, IL-1β, and IL-8 levels, in addition to insulin resistance (219). Some evidence suggests that the effect of olanzapine under cytokine secretion is gender-dependent; female Sprague-Dawley rats given olanzapine (low dose 2 mg/day; high dose 4 mg/day) for 3 weeks showed increased IL-8 levels, while males showed TNF-α concentration during low dose consumption, proving a gender-dependent difference. Also, compared with those of the control group, IL-6 levels were reduced in males after both doses of olanzapine while IL-1β concentration was reduced in females after a low dose (208).
Olanzapine can also modulate TLR expression in leukocytes. A study that evaluated 24 schizophrenic patients after 8 weeks of treatment (10–25 mg/day) exhibited that this drug increased TLR2 expression and decreased TLR4 and TLR5 in CD14+ monocytes. Treg and Tact cells reduced TLR2 and increased TLR5 expression (186). In 23 patients diagnosed with schizophrenia and treated with olanzapine for 8 weeks, IL-1RA was overexpressed, which correlated with the increase in leptin (210) (see Table 2).
Quetiapine is an AAP derived from benzothiazepine (220) (see Figure 1) and used for the treatment of psychotic symptoms in a wide range of disorders. Its use was authorized for the treatment of schizophrenia in 1997; it was authorized to treat unipolar and bipolar disorders in 2003 and bipolar depression in 2006. There are also many varied non-FDA-approved uses for quetiapine, such as anxiety, delirium, obsessive compulsive disorder, and the combined treatment of major depressive disorder (MDD) with antidepressants (221). Quetiapine metabolism, which comprises several steps as sulfoxidation, N- and O- dealkylation, and 7-hydroxilation by the CYP3A of the cytochrome P-450 system, produces N-desalkylquetiapine (norquetiapine), an active metabolite of quetiapine (222). Quetiapine is considered a multifunctional drug since it acts on three systems: dopaminergic, serotonergic, and noradrenergic (223). It shows high affinity for serotonin (5-HT) and DA type-2 receptors, slightly higher for the serotonergic than the dopaminergic. Contrastingly, lower affinity has been reported for type-1 receptors of both systems: D1 and 5-HT1A. Moreover, it is known that quetiapine also has affinity for histaminergic (H1) and adrenergic systems (α-1 and α-2) (see Table 1) (38).
It is well-known that most SGAs produce HPRL; however, quetiapine is considered among the safest medications due to its lower incidence of HPRL. Such properties have been associated to its lower affinity to Sackett et al. (149) and fast dissociation rate from DA receptors (224). In schizophrenia patients with sexual dysfunction, the treatment usually begins with PRL-sparing antipsychotics, switching to quetiapine in a second phase (225). In fact, quetiapine has been reported to revert HPRL in 175 patients with schizophrenia after a 2-week treatment (300–700 mg/day) (226).
On the other hand, several studies have shown the adverse endocrine effects produced by the administration of APs. In patients (12) with schizophrenia, quetiapine consumption induced significant insulin resistance. Nine months after administration, it led to a reduction in insulin sensitivity, as a result of a deficient secretion of insulin by the β-pancreatic cells (227). After a 10-month treatment with quetiapine, 16 youths (9–18 years) showed decreased levels of insulin associated with an impairment in β-pancreatic cell function (228). In mice, quetiapine administration (10 mg/kg) induced an increase in plasma levels of glucose but not in insulin, suggesting an insulin-blocker role of quetiapine in the insulin-secretory compensation mechanism (152), a finding supported by in vitro studies (229). McNamara et al., demonstrated that stearoyl-CoA desaturase-1 (scd-1), an enzyme involved in triglyceride biosynthesis and whose up-regulation showed a positive correlation with quetiapine consumption, could be involved in both sensitivity and insulin resistance (230). Moreover, the higher activity of scd-1 has been suggested as a risk factor for diabetes in humans (231), which reinforces the link between scd-1 and the adverse effects of quetiapine consumption. On the other hand, studies in rats have suggested that quetiapine-induced hyperglycemia was produced by increased levels of glucagon and suppressed glucagon-like peptide-1 (GLP-1) more than insulin resistance (66). Disturbances in glucagon and GLP-1 caused serious alterations in glucose metabolism because of stimulated hepatic glucose production (232).
In healthy volunteers aged 18–21 years, a dose of 150 mg/day of quetiapine was tested and the results showed an increase in PRL and growth hormone (GH) after 60 and 210 min of administration, respectively; in contrast, cortisol showed a decrease at 240 min and no changes were observed in ACTH (233). The alterations observed in GH levels by quetiapine consumption might be attributed to the high affinity and antagonism between the drug and H1 receptors. It should be noted that PRL in healthy volunteers showed a different behavior than that observed in patients, but, importantly, the sampling time used in healthy volunteers was very short. However, data in healthy volunteers are controversial, since other reports have shown no effects on PRL, but on ACTH due to the consumption of quetiapine in short sampling periods (234). Disturbances in ACTH and cortisol could be due to alterations in functioning of hypothalamus-pituitary-adrenal (HPA) axis in psychiatric patients, but the exact mechanism remains unclear. Moreover, quetiapine has revealed affectations in the levels of thyroxine (T4) and thyroid-stimulating hormone (TSH) (cases reports) with doses of 300–350 mg/day which induced a decrease in T4 and free T4, whereas TSH was increased (235).
Although the precise mechanism by which quetiapine induces adverse endocrine effects is not fully clear yet, some studies have focused their efforts on shedding light on this issue; nevertheless, more works are required to clarify this point.
Quetiapine consumption also affects the immune system. In patients with schizophrenia, quetiapine (600–1,200 mg/day, case reports) is associated with neutropenia, leukopenia (236–239), agranulocytosis, and thrombocytopenia (240). The mechanism by which quetiapine causes these adverse effects is still unclear, but some authors have proposed that this drug acts directly as a cytotoxic agent on immune cells, thus producing cell death; additionally, some products of quetiapine oxidation could induce apoptosis by oxidative stress (241). Other authors have suggested a bone marrow depression by quetiapine consumption, which could be produced by an inhibitory effect on leukopoiesis. It has even been proposed that quetiapine may act as a hapten, inducing antibody formation, complement activation, and cell death (237).
Studies in vitro have demonstrated the capability of quetiapine to alter the levels of some cytokines (242). Himmerich et al. demonstrated that this drug reduced IL-2 levels in whole blood cells, whereas it increased the levels of TNF-α and IL-17 (243). In PBMC cultures (LPS-stimulated) from patients with schizophrenia, quetiapine raised the levels of anti-inflammatory cytokines (IL-4 and IL-10) and lowered the pro-inflammatory ones (IFN-γ) (181). The anti-inflammatory properties of quetiapine may be explained by its capacity to suppress the NF-κB pathway activation. Quetiapine not only inhibited the expression NF-κB but also reverted its translocation from the cytosol to the nucleus, thus affecting its activation as well. These properties could explain quetiapine effects on cytokine expression (244) and have led experts to consider it a therapeutic alternative in some neuroinflammatory diseases.
Neurotrophins (NTs) are a group of neural growth factors that regulate survival, maintenance, cell differentiation, and synaptic plasticity in the CNS. But, their activity is not limited to the CNS: Cells of the immune system also express both NTs and their receptors (245), which in turn strongly contributes to the connection between neuronal dysregulation and inflammation (246). BDNF has been considered a potential biomarker of psychiatric disorders (247, 248). In patients with first episode psychosis, serum BDNF levels were increased after a 12-week treatment with quetiapine (200 or 400 mg/day). This rise in BDNF showed a positive correlation with the clinical improvement of patients, suggesting an indirect neurotrophic role of quetiapine through BDNF (249) (see Table 2).
Ziprasidone is a psychotropic agent commonly used in the treatment of schizophrenia (250) and bipolar disorder (251, 252) since its approval by the FDA in 2001. It is a benzisothiazolyl-3-yl-piperazine-type AAP (see Figure 1) with potent pharmacological antagonism to 5-HT2A and D2 receptors. However, it also acts on H1, M1, α1 and α2 receptors with less affinity (253, 254). The high affinity of ziprasidone to 5-HT2A as compared to D2 is an important characteristic of this drug. However, the pharmacological antagonism of ziprasidone toward D2 makes a lot of sense considering its antipsychotic effects, whereas the role of 5-HT2A receptors is still unclear (see Table 1). Still, it has been proposed that the antagonism against 5-HT2A stimulates the activity of DA in mesocortical pathways (52). Ziprasidone is metabolized almost fully, excreting only 5 % of the original drug intact. Aldehyde oxidase and cytochrome CYPA34 are the two main pathways by which ziprasidone is metabolized (255).
Ziprasidone slightly disturbs PRL levels and causes low extrapyramidal effects (256). There is a case study that reported elevated PRL levels after 9 days of ziprasidone administration (80 mg/day) (257, 258). Moreover, other studies have shown that ziprasidone suppresses the activity of the HPA-axis (n = 11, healthy volunteers; 40 mg/day), reducing the levels of nocturnal cortisol excretion, likely due to its antagonism toward H1 and α1 adrenergic receptors (259). Studies on the adverse effects produced by ziprasidone are scarce and more research is needed.
On the other hand, the immune alterations caused by ziprasidone consumption are few. There is no sufficient evidence supporting ziprasidone causes neutropenia, but there is a case report of agranulocytosis (120 mg/day); however, this effect was attributed to a combined activity of ziprasidone and mirtazapine (260). In vitro studies have shown that ziprasidone and its metabolites have cytotoxic, cytostatic, and genotoxic effects on peripheral blood lymphocyte cultures, causing a reduction in mitotic, proliferation, and nuclear division indexes (261).
In RAW macrophage cell line cultures, ziprasidone can induce inflammatory response. RAW cells exposed to ziprasidone (75 ng/L) showed increased levels of NO and ROS; moreover, they showed significantly higher levels of IL-1, IL-6, TNF-α, and IFN-γ but reduced levels of IL-10 (262). Several case reports have shown that ziprasidone induced allergic responses, such as Kounis syndrome (20 mg; IM) (263), pedal edema (80 mg/day) (264), urticaria, and angioedema (120 mg/day) (265). Little is known about the adverse effects of ziprasidone, but some studies have demonstrated minor effects in the endocrine system. On the other hand, special attention should be paid to the allergic response observed after ziprasidone administration, which can be explained by the high levels of IgE and the complement proteins C3 and C4 observed in patients (264). However, it is still unclear how ziprasidone induces this response (see Table 2).
Aripiprazole acts as a stabilizer of the dopamine-serotonin system. Its use was authorized for the treatment of schizophrenia in 2002 (266); in 2006 it was approved to treat bipolar disorder (mania or mixed episodes) (267), and major depressive disorder (as adjunctive drug) (268). In 2009 it was finally approved for the treatment of autism-related irritability (269). There are also non-FDA-approved uses for this drug such as Tourette syndrome and substance abuse disorders (270–273). Aripiprazole is metabolized in the liver by cytochrome P450, CYP2D6, and CYP3A4 by dehydrogenation, hydroxylation, and N-dealkylation. Its active metabolite, dehydro-aripiprazole, represents around 40% of the parent drug levels in plasma (274, 275). Despite the use of SGAs, this drug has several advantages for the treatment of multiple mood disorders, even if its consumption affects patients' metabolism (276–278).
Aripiprazole is a quinolinone derivate (see Figure 1); its pharmacological activity is based on its activity as a partial agonist of D2 and 5-HT1A receptors and as an antagonist of 5HT2A. Furthermore, aripiprazole exhibits a moderate affinity to α1 adrenergic and histaminergic H1 receptors. When compared to other typical and atypical APs, aripiprazole has a higher affinity to both states of D2 receptors (see Table 1) (40–42).
There are few reports of hormonal alterations caused by the consumption of aripiprazole, possibly because this drug develops fewer hormonal effects than other AAPs. There are multicentric studies that evaluate the tolerability, efficacy, and safety of aripiprazole in schizophrenia and other mood disorders for up to 52 weeks of treatment (15 mg/day) (279–282). The administration of aripiprazole (15 mg/day) is recommended for the control of HPRL associated to chronic consumption of other AAPs such as risperidone, amisulpride, olanzapine (270, 275, 283), and benzamide, and it helps to maintain improvement in the positive and negative symptoms of patients (134, 284, 285). In fact, aripiprazole is prescribed as a substitute for treatments with AAPs when the patients show no signs of clinical response or when they exhibit severe symptoms of sexual dysfunction associated with HPRL (134, 282, 286, 287). Although there are few cases of patients with an increase in PRL during treatment (288–290), aripiprazole is considered a safe drug.
There is minimal evidence on its metabolic activity, yet aripiprazole is known to play a partially protective role (291–295). Concerning research of aripiprazole-induced effects in animal models (Wistar rats) and cell lines (rHypoE-19), beneficial changes over metabolic parameters such as risk dyslipidemia and body weight have been found (72, 296).
Regarding the immune effects caused by aripiprazole consumption, there is evidence that shows this drug produces significant changes, such as cell count and changes in cytokine secretion, response to ROS, and gene expression. Although reports on the adverse effects of aripiprazole are scant compared to other AAPs, there is minimal evidence of its effect on the decrease in white blood cell count (297). A 10-year old with attention deficit hyperactivity disorder (ADHD) treated with aripiprazole (5 mg/day) showed a lower absolute neutrophil count (ANC). Additionally, a 50-year old Caucasian woman with schizophrenia developed neutropenia after aripiprazole consumption (15 mg/day) for 5 days (298), and a 21-year old Asian man with a conduct disorder showed a drop in WBC and neutropenia during aripiprazole treatment (297, 298). In all cases, the discontinuation of aripiprazole resulted in the normalization of WBC count and ANC, suggesting that the long-term bone marrow suppression by this drug plays a role in repeated antipsychotic consumption.
Some studies have shown that this drug affect cytokines secretion toward an anti-inflammatory profile: A meta-analysis involving 505 patients treated with aripiprazole showed a relationship between cytokine levels (TNF-α and IFN-γ) and their possible role as state and trait markers (86, 299). Another report described that aripiprazole consumption (5–30 mg/day, 3 months) reduced TNF-α, IL-8, IL-21, IL-13, IL-17, and fractalkine (CXCL1) levels in 31 first-episode psychotic patients; the effect in these molecules exhibited a positive correlation with clinical improvement (174). Another study also demonstrated a decrease in IL−1β, IL-6, TNF-α, IL-12, IL-23, IL-4, and IFN-γ under aripiprazole treatment with a dose from 10 mg/day (week 1) to a maximum of 30 mg/day (weeks 2, 3, and 4) (300).
In vitro studies confirm those data, since PBMC from healthy subjects and THP-1 cells incubated with aripiprazole (10–5 μM) exhibited a decrease in the expression of pro-inflammatory cytokines IL-1β, IL-6, and TNF-α and reduced the levels of IL-2, IL-9, IP-10 (CXCL10), and MIP-1β (CCL4) in the supernatant (217). The anti-inflammatory effect shown by this drug could be associated to the decrease in gene expression of cyclooxygenase (COX)-2 and inducible nitric oxide synthase (iNOS), causing lower levels of NO, prostaglandin 2 (PGE 2), and TNF-α (301). Furthermore, It is known that RAW264.7 cells treated with aripiprazole (20 μM) inhibited the interaction of the second messengers TAK1, MKK4, and MKK7 on AP-1, and, and Syk, which play a key role in the NF-κB signaling pathway (301). Aripiprazole also acts as an antioxidant improving the response to ROS. Studies in murine (2 mg/kg) and in vitro (5 μM) models showed that this drug increased the activity of glutathione peroxidase (GSH-Px) and superoxide dismutase (SOD) enzymes, promoting a decrease in the concentration of NO in supernatants and TNF-α, IL-1α, IL-2, and IL-10 in mice serum levels. This antioxidant activity is related to the input of intracellular [Ca2+], which allows for ROS regulation and the decrease in inflammation cytokines (302, 303).
Some evidence suggests that this drug also modifies the gene expression of relevant genes; an in vitro study using primary human adipose-derived stem cells (ADSCs) demonstrated that aripiprazole (100 ng/mL) increased the expression of key genes involved in cell cycle (ANAPC2, CD14), apoptosis (BCL2), nuclear and transporter receptors (PPARα, PPARγ, ABCA1, LEPR, INSR), transcript factors (CEBPA, SREBF1, NF-KB1), signal transduction (IRS1, SIRT1), adipogenic markers, lipid metabolism, adipokines (ADFP, FABPN, LPL, ACSL1, ADIPOQ, LEP) and cytokines and chemokines (TNF-α, IL-1β, IL-8, MCP-1). These results support the role of AAPs in the recruitment of MQs to adipose tissue by increasing MCP-1 and the risk of metabolic syndrome associated with drug treatment (185). However, this drug showed no significant immunotoxic effects in ICR mice and C6 glioma and RAW264.7 cells (50 mg/kg) when no alterations in organs or cell lines were found (304).
In summary, there is little evidence on the hormonal and immune effects of aripiprazole, as well as its partially protective role (291–295). These effects allow aripiprazole to suitably treat schizophrenia and bipolar disorder (304) (see Table 2).
Paliperidone, or 9-hydroxy-risperidone (see Figure 1), is the most significant active metabolite of risperidone. The FDA approved this drug for the treatment of schizophrenia in 2006 (305). Paliperidone is a monotherapy drug for short-term and maintenance treatment of schizophrenia as well as monotherapy or adjunct drug for the short-term treatment of schizoaffective disorder (306–308). It has also been used in the treatment of bipolar disorder (309), borderline personality disorder (310), Huntington's disease (311), ASD, and ADHD (312); however, it has not been approved to treat any of these last clinical conditions. Paliperidone is a racemic mixture of (+)-paliperidone and (–)-paliperidone enantiomers that undergo minimal hepatic metabolism (44). The available pharmaceutical formulations of this drug are oral immediate-release formulation, oral extended-release (ER) formulation, and intramuscular depot formulation (305).
The therapeutic activity of paliperidone is comparable with that of risperidone itself; its action mechanism is unknown, but it likely acts through a combination of 5-HT2A agonism and D2 receptor antagonism (see Table 1) (44). This drug is also active as an antagonist for other receptors such as D3, D4 dopaminergic receptors, 5-HT1A, 5-HT1B, and 5-HT1D serotoninergic receptors, and α-1 and α-2 adrenergic receptors, although it also acts as agonist to 5-HT2C and 5- H1 histaminergic receptors (45).
The immunoendocrine alterations cited in this section are related to paliperidone; effects on 9-hydroxy-risperidone by risperidone consumption and its subsequent metabolism are not mentioned. The most representative endocrine alteration reported after paliperidone consumption is the increase in PRL or HPRL (207, 308, 313, 314). This alteration can produce prolactin-related adverse effects (PRL-RAEs) or be asymptomatic (315).
HPRL induced by risperidone/paliperidone treatment in schizophrenic patients was presented in association with rs40184 and rs3863145 variants in the SLC6A3 gene of blood leukocyte DNA (132). According to several reports, paliperidone produced high HPRL incidence when compared vs. other SGAs in adults (dosage 7.03 ± 3.63 mg/day) (111) and pediatric patients (130). Paliperidone consumption showed an association between PRL, sex, and age (113, 130), although Druyts and cols. reported no differences between females and males (314). This drug increased PRL levels, yet some reports have shown that the switch from risperidone or paliperidone ER to paliperidone palmitate treatment (PP, an intramuscular depot formulation) reduced PRL concentration (316, 317) as well as sexual dysfunction (316), a common PRL-RAE. Similarly, patients with sexual dysfunction presented higher PRL as compared with no sexual dysfunction patients (318). Adolescent patients with PRL-RAE showed higher PRL levels when compared against patients without PRL-RAE (1.5–12 mg/day) (319). According to the literature above, different formulations of paliperidone could cause this alteration in pediatric and adult patients. The precise mechanism by which paliperidone increases PRL levels is unclear; however, it corresponds to D2 receptor blockade (320).
There are a few reports that show changes in other hormonal profiles in patients during paliperidone consumption. Although other AAPs mentioned in this review induce dysregulation in glucose metabolism, paliperidone does not modify serum levels of insulin. The acute and chronic treatment with paliperidone did not alter serum insulin levels and β-cell function with the homeostatic model assessment (HOMA-B) (207, 313, 321). However, a case report showed increased insulin secretion, causing hypoglycemia in a schizophrenic female patient (322).
The reports on immune alterations induced by paliperidone consumption are a few yet diverse. Several cases of schizophrenic patients showed that paliperidone treatment decreased leukocyte counts. Monotherapy with paliperidone produced leukopenia and neutropenia (323); still, the combined use of paliperidone depot/risperidone (100–2 mg/day) resulted in leukopenia and lymphopenia but risperidone alone did not (159). Agranulocytosis was reported in a patient when switching from risperidone to paliperidone treatment (6 mg/day) (324). The treatment with paliperidone ER/valproic acid (12–1,000 mg/day) caused leukopenia and neutropenia in a patient with schizoaffective disorder (325). In all cases, the cytopenic alterations were normalized after discontinuing the consumption of paliperidone. Some proposed mechanisms of AP-induced blood dyscrasia, such as paliperidone, include direct bone marrow suppression, antibody formation against hematologic precursors, and peripheral WBC destruction (326).
Paliperidone increases BDNF concentration during acute treatment. The serum levels of BDNF in first-episode schizophrenia patients increased after a 12-week paliperidone treatment negatively correlated with a reduction rate of the positive and negative symptoms scale (PANSS) score (unspecified dose) (327). However, the paliperidone ER treatment during 8 weeks did not increase BDNF serum concentration (unspecified dose) (328).
In blood, peripheral cells of patients with EPS (acute dystonia and drug-induced parkinsonism) showed a constructed network enriched in different biological processes related to pathways of NF-κB, an important transcription factor for immune response, (12.85 ± 2.85 mg/day) (329) in patients with paliperidone or risperidone treatment. In vitro, U-937 human cell line decreased cell survival with 25 and 50 μM/mL of paliperidone (330) (see Table 2).
The FDA approved asenapine for the treatment of schizophrenia (331) and bipolar disorders (332) in 2019. This drug is a new AAP with unique features that was introduced in Japan in 2016, and it is the only AP used sublingually; its chemical structure of (±)-Asenapine can be described as a tetracyclic framework wherein N-methylpyrrolidine ring fuses at third and fourth positions with chlorophenyl phenyl ether in a trans geometry (333) (see Figure 1). This drug is metabolized rapidly in a process mediated by glucuronidation and demethylation pathways that induce two non-active metabolites, asenapine N-glucuronide and asenapine N-desmethyl carbamoyl glucuronide (334). Asenapine has subnanomolar and nanomolar affinities for diverse and numerous subtypes of aminergic G protein coupled receptors (GPCRs) associated to 5-HT, norepinephrine (NE), DA, and histamine (H) (335, 336). Still, the antagonist activity at 5-HT1A, 5-HT1B, 5-HT2A, 5-HT2C, 5-HT5A, 5-HT6, and 5-HT7 may contribute to the antimanic and antidepressant effects of asenapine (see Table 1) (39).
Endocrine deleterious side effects induced by asenapine consumption were reported in PRL and insulin blood levels. Asenapine displays more potent antagonist activity toward 5-HT2A receptor than D2 receptor (337, 338), that is why it has a low propensity to cause PRL elevation (331, 339, 340); Therefore, this drug is one of the AP treatments of choice for breast cancer patients (341). Nevertheless, research groups reported that 2.3% of patients with bipolar disorder who received asenapine monotherapy had PRL levels ≥ 4 times the upper limit of the normal range, compared with those who received a placebo (332, 342), In contrast, 9% of patients with schizophrenia who received asenapine (5 and 10 mg twice daily) had PRL levels over 2-fold the upper limit of the normal range compared with those who received a placebo (343).
Insulin altered levels are associated with glucose metabolism disturbances and the evidence shows that asenapine modifies the blood levels of these hormones. In 302 patients (aged 10–17 years) with bipolar I disorder in manic or mixed episodes who were treated with asenapine (2, 5, or 10 mg twice, daily) for 3 weeks, the mean change from baseline in fasting insulin was significant when compared to controls. In all cases, the patients treated with asenapine increased their body weight (344). Contrastingly, no changes in insulin resistance were detected in adult female Sprague Dawley rats treated with asenapine (0.01, 0.05, 0.1, 0.5, 1.0 mg/kg) (67).
As described above, asenapine can interact with 5-HT, NE, DA, and H receptors expressed in leukocytes (28, 29, 345, 346). Then, the administration or consumption of this drug could induce changes in the inflammatory response in patients, although the evidence of this effect is very scarce. There is only one report of a case of pityriasis rosea secondary to asenapine consumption. A biopsy of the lesions evidenced superficial and deep perivascular and interstitial dermatitis with eosinophils and dermal perivascular lymphocytic infiltrate, as well as minimal parakeratosis and spongiosis (347). Although there was no molecular explanation of this phenomenon, we may speculate that this patient had an alteration in neurotransmitter receptors (density or functional alteration) expressed by leukocytes, becoming more susceptible to this aberrant inflammatory response secondary to asenapine consumption (see Table 2).
Little is known about the effects of AAPs on the microbiota; however, a small body of evidence suggests they cause severe adverse effects. Olanzapine and risperidone induced an increase in Firmicutes and a decrease in Bacteroidetes, as well as metabolic alterations as a result of a shift toward a potentially obesogenic bacterial profile associated with short-chain fatty acids and inflammation in adults (348), children (349, 350), and rodents (351). These changes were also gender-dependent (349, 352, 353), with females showing a higher pro-inflammatory cytokine (IL-8 and IL-1β) response in circulation and macrophage infiltration; still, microbiota dysbiosis was equally present in males and females.
AAPs have a potent antibiotic effect, inducing a profound dysbiosis in the gut microbiota, either chronically or after short-term administration (205). Antibiotic co-administration resulted in further changes in microbiota composition. Interestingly, these antibiotic-dependent changes in microbiota diversity reduced the side effects, including macrophage infiltration. Furthermore, experiments in germ-free mice showed no alteration in their metabolic profile (352–355), indicating a clear role of the microbiota in the metabolic dysfunction associated with AAPs (352). Finally, fecal transplants from risperidone-treated mice induced excess weight gain in control mice (354). These alterations have been associated to a decrease in Bifidobacterium, Escherichia coli, and Lactobacillus and an increase in Clostridium coccoides (353).
Risperidone in vitro altered the colon microbiota just 24 h after administration, inducing specific metabolites (350). Probiotic treatment has shown a protective effect, restoring the Bacteroidetes:Firmicutes ratio, without reducing the AAPs effect (356).
The bidirectional communication between the SNC with other peripheral systems occurs by the release of soluble molecules that interact with their receptors. Any cell in the organism that bears a functional receptor for a molecule will respond when they interact. The complex structure that confers pharmacological non-specificity to AAPs allows for the interaction with the receptors they have an affinity for, not only in the CNS but also in all body cells. This result leads to the therapeutic effect of AAPs in various psychiatric conditions and their possible ability to modify the endocrine and immune systems as well as the gut microbiota. The therapeutic effect of AAPs is exhibited by the antagonism in CNS receptors that are involved in the pathophysiology of the disease. In schizophrenia, for example, the positive and negative symptoms decrease due to the AAP-receptor interaction in the mesocortical and mesolimbic pathways, although HPRL is caused by the antagonism of receptors in the tuberoinfundibular pathway. In addition, the antagonism of neurotransmitter receptors on leukocytes and glandular cells have immune and endocrine effects. The effect of each AAP is unique and depends on specificity and affinity characteristics.
AAPs are drugs prescribed for various psychiatric conditions due to their high efficiency and low rate of extrapyramidal effects. However, these drugs have systemic effects that are not only metabolic but also related to changes in endocrine and immune responses. Having greater knowledge of these immune, endocrine, and microbiota effects, allows clinicians to have a broader point of view and more significant criteria to prescribe these drugs to patients, considering that the adverse effects can modify the systemic response and generate undesirable effects, with a direct impact on the patients' quality of life. It is necessary to start a new generation of drugs that support the resolution of psychiatric symptoms with higher specificity to prevent acute adverse effects and the patients' systemic deterioration by chronic consumption.
SA-H, RE, and LP: conceptualization. All authors: writing-original draft preparation and writing-review and editing. LP: supervision.
The study was supported by Instituto Nacional de Psiquiatría Ramón de la Fuente Muñiz Projects SC-15-24-14; NC150048SECITI; SECITI 0048/2014; NC16044.0; NC092318.0 and FOSISS: SALUD-2017-1-289800. SA-H is a doctoral student from Programa de Doctorado en Ciencias Biomédicas, Universidad Nacional Autónoma de México (UNAM) and she was supported by CONACYT-fellowship number 594780.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We acknowledge and appreciate the support of Raul Cardoso and José L. Calderon, from the Department of Biomedical Illustration of INPRFM. Thanks to León Luka for breaking paradigms and taking us beyond our limits.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2020.00195/full#supplementary-material
1. Cunningham Owens D, Johnstone EC. The development of antipsychotic drugs. Brain Neurosci Adv. (2018) 2:2398212818817498. doi: 10.1177/2398212818817498
2. Meltzer HY. Update on typical and atypical antipsychotic drugs. Annu Rev Med. (2013) 64:393–406. doi: 10.1146/annurev-med-050911-161504
3. Zhang J-P, Gallego JA, Robinson DG, Malhotra AK, Kane JM, Correll CU. Efficacy and safety of individual second-generation vs. first-generation antipsychotics in first-episode psychosis: a systematic review and meta-analysis. Int J Neuropsychopharmacol. (2013) 16:1205–18. doi: 10.1017/S1461145712001277
4. Singh R, Bansal Y, Medhi B, Kuhad A. Antipsychotics-induced metabolic alterations: recounting the mechanistic insights, therapeutic targets and pharmacological alternatives. Eur J Pharmacol. (2019) 844:231–40. doi: 10.1016/j.ejphar.2018.12.003
5. Chokhawala K, Stevens L. Antipsychotic Medications. Treasure Island, FL: StatPearls Publishing (2019).
6. Lee ES, Vidal C, Findling RL. A focused review on the treatment of pediatric patients with atypical antipsychotics. J Child Adolesc Psychopharmacol. (2018) 28:582–605. doi: 10.1089/cap.2018.0037
7. Xu H, Zhuang X. Atypical antipsychotics-induced metabolic syndrome and nonalcoholic fatty liver disease: a critical review. Neuropsychiatr Dis Treat. (2019) 15:2087–99. doi: 10.2147/NDT.S208061
8. Murray R, Correll CU, Reynolds GP, Taylor D. Atypical antipsychotics: recent research findings and applications to clinical practice: proceedings of a symposium presented at the 29th Annual European college of neuropsychopharmacology congress, 19 September 2016, Vienna, Austria. Ther Adv Psychopharmacol. (2017) 7:1–14. doi: 10.1177/2045125317693200
9. Wei Xin Chong J, Hsien-Jie Tan E, Chong CE, Ng Y, Wijesinghe R. Atypical antipsychotics: a review on the prevalence, monitoring, and management of their metabolic and cardiovascular side effects. Ment Heal Clin. (2016) 6:178–84. doi: 10.9740/mhc.2016.07.178
10. De Hert M, Detraux J, van Winkel R, Yu W, Correll CU. Metabolic and cardiovascular adverse effects associated with antipsychotic drugs. Nat Rev Endocrinol. (2012) 8:114–26. doi: 10.1038/nrendo.2011.156
11. Kowalchuk C, Castellani LN, Chintoh A, Remington G, Giacca A, Hahn MK. Antipsychotics and glucose metabolism: how brain and body collide. Am J Physiol Metab. (2019) 316:E1–15. doi: 10.1152/ajpendo.00164.2018
12. Guest PC. Insulin resistance in schizophrenia. Adv Exp Med Biol. (2019) 1134:1–16. doi: 10.1007/978-3-030-12668-1_1
13. Misiak B, Bartoli F, Stramecki F, Samochowiec J, Lis Michałand Kasznia J, Jarosz K, et al. Appetite regulating hormones in first-episode psychosis: a systematic review and meta-analysis. Neurosci Biobehav Rev. (2019) 102:362–70. doi: 10.1016/J.NEUBIOREV.2019.05.018
14. Karanikas E, Ntouros E, Oikonomou D, Floros G, Griveas I, Garyfallos G. Evidence for hypothalamus-pituitary-adrenal axis and immune alterations at prodrome of psychosis in males. Psychiatry Investig. (2017) 14:703–7. doi: 10.4306/pi.2017.14.5.703
15. Chen ML, Tsai TC, Lin YY, Tsai YM, Wang LK, Lee MC, Tsai FM. Antipsychotic drugs suppress the AKT/NF-κB pathway and regulate the differentiation of T-cell subsets. Immunol Lett. (2011) 140:81–91. doi: 10.1016/j.imlet.2011.06.011
16. Chen ML, Tsai TC, Wang LK, Lin YY, Tsai YM, Lee MC, et al. Risperidone modulates the cytokine and chemokine release of dendritic cells and induces TNF-α-directed cell apoptosis in neutrophils. Int Immunopharmacol. (2012) 12:197–204. doi: 10.1016/j.intimp.2011.11.011
17. Chen ML, Tsai TC, Wang LK, Lin YY, Tsai YM, Lee MC, et al. Clozapine inhibits Th1 cell differentiation and causes the suppression of IFN-γ production in peripheral blood mononuclear cells. Immunopharmacol Immunotoxicol. (2012) 34:686–94. doi: 10.3109/08923973.2011.651535
18. Petrikis P, Voulgari PV, Tzallas AT, Boumba VA, Archimandriti DT, Zambetas D, et al. Changes in the cytokine profile in first-episode, drug-naïve patients with psychosis after short-term antipsychotic treatment. Psychiatry Res. (2017) 256:378–83. doi: 10.1016/J.PSYCHRES.2017.07.002
19. Capuzzi E, Bartoli F, Crocamo C, Clerici M, Carrà G. Acute variations of cytokine levels after antipsychotic treatment in drug-naïve subjects with a first-episode psychosis: a meta-analysis. Neurosci Biobehav Rev. (2017) 77:122–8. doi: 10.1016/j.neubiorev.2017.03.003
20. Cussotto S, Clarke G, Dinan TG, Cryan JF. Psychotropics and the microbiome: a chamber of secrets. Psychopharmacology. (2019) 236:1411–32. doi: 10.1007/s00213-019-5185-8
21. Jeon S, Kim Y-K. Unresolved issues for utilization of atypical antipsychotics in schizophrenia: antipsychotic polypharmacy and metabolic syndrome. Int J Mol Sci. (2017) 18:2174. doi: 10.3390/ijms18102174
22. Reynolds GP, Kirk SL. Metabolic side effects of antipsychotic drug treatment – pharmacological mechanisms. Pharmacol Ther. (2010) 125:169–79. doi: 10.1016/j.pharmthera.2009.10.010
23. Correll CU, Rummel-Kluge C, Corves C, Kane JM, Leucht S. Antipsychotic combinations vs monotherapy in schizophrenia: a meta-analysis of randomized controlled trials. Schizophr Bull. (2009) 35:443–57. doi: 10.1093/schbul/sbn018
24. Fleischhacker WW, Uchida H. Critical review of antipsychotic polypharmacy in the treatment of schizophrenia. Int J Neuropsychopharmacol. (2014) 17:1083–93. doi: 10.1017/S1461145712000399
25. Liu Y, Zhou X, Qin B, Del Giovane C, Zhang Y, Xie P. Efficacy, quality of life, and acceptability outcomes of atypical antipsychotic augmentation treatment for treatment-resistant depression: protocol for a systematic review and network meta-analysis. Syst Rev. (2014) 3:133. doi: 10.1186/2046-4053-3-133
26. Baandrup L. Polypharmacy in schizophrenia. Basic Clin Pharmacol Toxicol. (2020) 126:183–92. doi: 10.1111/bcpt.13384
27. Kapur S, Zipursky RB, Remington G. Clinical and theoretical implications of 5-HT2 and D2 receptor occupancy of clozapine, risperidone, and olanzapine in schizophrenia. Am J Psychiatry. (1999) 156:286–93. doi: 10.1176/ajp.156.2.286
28. Arreola R, Alvarez-Herrera S, Pérez-Sánchez G, Becerril-Villanueva E, Cruz-Fuentes C, Flores-Gutierrez EO, et al. Immunomodulatory effects mediated by dopamine. J Immunol Res. (2016) 2016:3160486. doi: 10.1155/2016/3160486
29. Arreola R, Becerril-Villanueva E, Cruz-Fuentes C, Velasco-Velázquez MA, Garcés-Alvarez ME, Hurtado-Alvarado G, et al. Immunomodulatory effects mediated by serotonin. J Immunol Res. (2015) 2015:354957. doi: 10.1155/2015/354957
30. Zhang Y, Zheng R, Meng X, Wang L, Liu L, Gao Y. Pancreatic endocrine effects of dopamine receptors in human islet cells. Pancreas. (2015) 44:925–9. doi: 10.1097/MPA.0000000000000357
31. Chue P, Chue J. A review of olanzapine pamoate. Expert Opin Pharmacother. (2012) 13:1661–70. doi: 10.1517/14656566.2012.686169
32. Ishibashi T, Horisawa T, Tokuda K, Ishiyama T, Ogasa M, Tagashira R, et al. Pharmacological profile of lurasidone, a novel antipsychotic agent with potent 5-hydroxytryptamine 7 (5-HT 7) and 5-HT 1A receptor activity. J Pharmacol Exp Ther. (2010) 334:171–81. doi: 10.1124/jpet.110.167346
33. Zajdel P, Kos T, Marciniec K, Satała G, Canale V, Kaminski K, et al. Novel multi-target azinesulfonamides of cyclic amine derivatives as potential antipsychotics with pro-social and pro-cognitive effects. Eur J Med Chem. (2018) 145:790–804. doi: 10.1016/J.EJMECH.2018.01.002
34. Fernández J, Alonso JM, Andrés JI, Cid JM, Díaz A, Iturrino L, et al. Discovery of new tetracyclic tetrahydrofuran derivatives as potential broad-spectrum psychotropic agents. J Med Chem. (2005) 48:1709–12. doi: 10.1021/jm049632c
35. Ablordeppey SY, Altundas R, Bricker B, Zhu XY, Suresh Kumar EVK, Jackson T, et al. Identification of a butyrophenone analog as a potential atypical antipsychotic agent: 4-[4-(4-Chlorophenyl)-1,4-diazepan-1-yl]-1-(4-fluorophenyl)butan-1-one. Bioorg Med Chem. (2008) 16:7291–301. doi: 10.1016/j.bmc.2008.06.030
36. Cutler AJ, Kalali AH, Mattingly GW, Kunovac J, Meng X. Long-term safety and tolerability of iloperidone: results from a 25-week, open-label extension trial. CNS Spectr. (2013) 18:43–54. doi: 10.1017/S1092852912000764
37. Munshi T, Mazhar M, Hassan T. Clozapine reinitiation following a “red result” secondary to chemotherapy. Neuropsychiatr Dis Treat. (2013) 9:1267–71. doi: 10.2147/NDT.S49028
38. Keating GM, Robinson DM. Quetiapine: a review of its use in the treatment of bipolar depression. Drugs. (2007) 67:1077–95. doi: 10.2165/00003495-200767070-00008
39. Reynolds GP. Receptor mechanisms of antipsychotic drug action in bipolar disorder – focus on asenapine. Ther Adv Psychopharmacol. (2011) 1:197–204. doi: 10.1177/2045125311430112
40. Keck PE, McElroy SL. Aripiprazole: a partial dopamine D2 receptor agonist antipsychotic. Expert Opin Investig Drugs. (2003) 12:655–62. doi: 10.1517/eoid.12.4.655.23750
41. Burris KD, Molski TF, Xu C, Ryan E, Tottori K, Kikuchi T, et al. Aripiprazole, a novel antipsychotic, is a high-affinity partial agonist at human dopamine D2 receptors. J Pharmacol Exp Ther. (2002) 302:381–9. doi: 10.1124/jpet.102.033175
42. Goodnick PJ, Jerry JM. Aripiprazole: profile on efficacy and safety. Expert Opin Pharmacother. (2002) 3:1773–1781. doi: 10.1517/14656566.3.12.1773
43. Mauri MC, Paletta S, Maffini M, Colasanti A, Dragogna F, Di Pace C, et al. Clinical pharmacology of atypical antipsychotics: an update. EXCLI J. (2014) 13:1163–91.
44. Chue P, Chue J. A review of paliperidone palmitate. Expert Rev Neurother. (2012) 12:1383–97. doi: 10.1586/ern.12.137
45. Wang SM, Han C, Lee SJ, Patkar AA, Pae CU, Fleischhacker WW. Paliperidone: a review of clinical trial data and clinical implications. Clin Drug Investig. (2012) 32:497–512. doi: 10.2165/11634440
46. Kalkman HO, Subramanian N, Hoyer D. Extended radioligand binding profile of iloperidone: a broad spectrum dopamine/serotonin/norepinephrine receptor antagonist for the management of psychotic disorders. Neuropsychopharmacology. (2001) 25:904–14. doi: 10.1016/S0893-133X(01)00285-8
47. Kalkman HO, Feuerbach D, Lötscher E, Schoeffter P. Functional characterization of the novel antipsychotic iloperidone at human D2, D3, α2C, 5-HT6, and 5-HT1A receptors. Life Sci. (2003) 73:1151–59. doi: 10.1016/S0024-3205(03)00419-3
48. Richelson E, Souder T. Binding of antipsychotic drugs to human brain receptors focus on newer generation compounds. Life Sci. (2000) 68:29–39. doi: 10.1016/s0024-3205(00)00911-5
49. Roth BL, Lopez E, Patel S, Kroeze WK. The multiplicity of serotonin receptors: uselessly diverse molecules or an embarrassment of riches? Neuroscientist. (2000) 6:252–62. doi: 10.1177/107385840000600408
50. Kongsamut S, Roehr JE, Cai J, Hartman HB, Weissensee P, Kerman LL, et al. Iloperidone binding to human and rat dopamine and 5-HT receptors. Eur J Pharmacol. (1996) 317:417–23. doi: 10.1016/S0014-2999(96)00840-0
51. Caccia S, Pasina L, Nobili A. New atypical antipsychotics for schizophrenia: iloperidone. Drug Des Devel Ther. (2010) 4:33–48. doi: 10.2147/dddt.s6443
52. Conley RR, Kelly DL. Current status of antipsychotic treatment. Curr Drug Targets CNS Neurol Disord. (2002) 1:123–8. doi: 10.2174/1568007024606221
53. Aringhieri S, Carli M, Kolachalam S, Verdesca V, Cini E, Rossi M, et al. Molecular targets of atypical antipsychotics: from mechanism of action to clinical differences. Pharmacol Ther. (2018) 192:20–41. doi: 10.1016/j.pharmthera.2018.06.012
54. Rajagopal S, Rajagopal K, Lefkowitz RJ. Teaching old receptors new tricks: biasing seven-transmembrane receptors. Nat Rev Drug Discov. (2010) 9:373–86. doi: 10.1038/nrd3024
55. Kenakin T. New concepts in pharmacological efficacy at 7TM receptors: IUPHAR review 2. Br J Pharmacol. (2013) 168:554–75. doi: 10.1111/j.1476-5381.2012.02223.x
56. Kaya AI, Onaran HO, Özcan G, Ambrosio C, Costa T, Balli S, et al. Cell contact-dependent functional selectivity of β2-adrenergic receptor ligands in stimulating cAMP accumulation and extracellular signal-regulated kinase phosphorylation. J Biol Chem. (2012) 287:6362–74. doi: 10.1074/jbc.M111.301820
57. Kenakin T. Signaling bias in drug discovery. Expert Opin Drug Discov. (2017) 12:321–33. doi: 10.1080/17460441.2017.1297417
58. Hahn MK, Wolever TMS, Arenovich T, Teo C, Giacca A, Powell V, et al. Acute effects of single-dose olanzapine on metabolic, endocrine, and inflammatory markers in healthy controls. J Clin Psychopharmacol. (2013) 33:740–46. doi: 10.1097/JCP.0b013e31829e8333
59. De Berardis D, Rapini G, Olivieri L, Di Nicola D, Tomasetti C, Valchera A, et al. Safety of antipsychotics for the treatment of schizophrenia: a focus on the adverse effects of clozapine. Ther Adv drug Saf. (2018) 9:237–56. doi: 10.1177/2042098618756261
60. Nikolić-Kokić A, Tatalović N, Nestorov J, Mijović M, Mijusković A, Miler M, et al. Clozapine, ziprasidone, and sertindole-induced morphological changes in the rat heart and their relationship to antioxidant enzymes function. J Toxicol Environ Health A. (2018) 81:844–53. doi: 10.1080/15287394.2018.1495587
61. Yuen JWY, Wu C, Wang CK, Kim DD, Procyshyn RM, Honer WG, et al. A comparison of the effects of clozapine and its metabolite norclozapine on metabolic dysregulation in rodent models. Neuropharmacology. (2019) 23:107717. doi: 10.1016/j.neuropharm.2019.107717
62. Nakazawa T, Kikuchi M, Ishikawa M, Yamamori H, Nagayasu K, Matsumoto T, et al. Differential gene expression profiles in neurons generated from lymphoblastoid B-cell line-derived iPS cells from monozygotic twin cases with treatment-resistant schizophrenia and discordant responses to clozapine. Schizophr Res. (2017) 181:75–82. doi: 10.1016/j.schres.2016.10.012
63. Potvin S, Zhornitsky S, Stip E. Antipsychotic-induced changes in blood levels of leptin in schizophrenia: a meta-analysis. Can J Psychiatry. (2015) 60:S26–34. Available at: http://www.ncbi.nlm.nih.gov/pubmed/25886677
64. Houseknecht KL, Robertson AS, Zavadoski W, Gibbs EM, Johnson DE, Rollema H. Acute effects of atypical antipsychotics on whole-body insulin resistance in rats: implications for adverse metabolic effects. Neuropsychopharmacology. (2007) 32:289–97. doi: 10.1038/sj.npp.1301209
65. Tulipano G, Rizzetti C, Bianchi I, Fanzani A, Spano P, Cocchi D. Clozapine-induced alteration of glucose homeostasis in the rat: the contribution of hypothalamic-pituitary-adrenal axis activation. Neuroendocrinology. (2007) 85:61–70. doi: 10.1159/000100981
66. Smith GC, Vickers MH, Cognard E, Shepherd PR. Clozapine and quetiapine acutely reduce glucagon-like peptide-1 production and increase glucagon release in obese rats: implications for glucose metabolism and food choice behaviour. Schizophr Res. (2009) 115:30–40. doi: 10.1016/j.schres.2009.07.011
67. Boyda HN, Procyshyn RM, Pang CCY, Hawkes E, Wong D, Jin CH, et al. Metabolic side-effects of the novel second-generation antipsychotic drugs asenapine and iloperidone: a comparison with olanzapine. PLoS ONE. (2013) 8:e53459. doi: 10.1371/journal.pone.0053459
68. Tschoner A, Engl J, Rettenbacher M, Edlinger M, Kaser S, Tatarczyk T, et al. Effects of six second generation antipsychotics on body weight and metabolism - risk assessment and results from a prospective study. Pharmacopsychiatry. (2009) 42:29–34. doi: 10.1055/s-0028-1100425
69. Bymaster FP, Felder CC, Tzavara E, Nomikos GG, Calligaro DO, Mckinzie DL. Muscarinic mechanisms of antipsychotic atypicality. Prog Neuropsychopharmacol Biol Psychiatry. (2003) 27:1125–43. doi: 10.1016/j.pnpbp.2003.09.008
70. Weston-Green K, Huang X-F, Deng C. Second generation antipsychotic-induced type 2 diabetes: a role for the muscarinic M3 receptor. CNS Drugs. (2013) 27:1069–80. doi: 10.1007/s40263-013-0115-5
71. Schmid CL, Streicher JM, Meltzer HY, Bohn LM. Clozapine acts as an agonist at serotonin 2A receptors to counter MK-801-induced behaviors through a βarrestin2-independent activation of Akt. Neuropsychopharmacology. (2014) 39:1902–13. doi: 10.1038/npp.2014.38
72. Kowalchuk C, Kanagasundaram P, Belsham DD, Hahn MK. Antipsychotics differentially regulate insulin, energy sensing, and inflammation pathways in hypothalamic rat neurons. Psychoneuroendocrinology. (2019) 104:42–8. doi: 10.1016/j.psyneuen.2019.01.029
73. Heppner KM, Perez-Tilve D. GLP-1 based therapeutics: simultaneously combating T2DM and obesity. Front Neurosci. (2015) 9:92. doi: 10.3389/fnins.2015.00092
74. Ekström J, Godoy T, Loy F, Riva A. Parasympathetic vasoactive intestinal peptide (VIP): a likely contributor to clozapine-induced sialorrhoea. Oral Dis. (2014) 20:90–6. doi: 10.1111/odi.12139
75. Khalaf HA, Elmorsy E, Mahmoud E-HM, Aggour AM, Amer SA. The role of oxidative stress in ovarian toxicity induced by haloperidol and clozapine-a histological and biochemical study in albino rats. Cell Tissue Res. (2019) 378:371–83. doi: 10.1007/s00441-019-03067-x
76. Margulska A, Kozłowska E, Wysokinski A. Effect of clozapine dose and concentration on fasting concentration of appetite regulating peptides. Psychiatry Res. (2018) 260:473–7. doi: 10.1016/j.psychres.2017.12.018
77. Oh J-E, Cho YM, Kwak S-N, Kim J-H, Lee KW, Jung H, et al. Inhibition of mouse brown adipocyte differentiation by second-generation antipsychotics. Exp Mol Med. (2012) 44:545–53. doi: 10.3858/emm.2012.44.9.062
78. Basta-Kaim A, Szcz?eny E, Leśkiewicz M, Głombik K, Slusarczyk J, Budziszewska B, et al. Maternal immune activation leads to age-related behavioral and immunological changes in male rat offspring - The effect of antipsychotic drugs. Pharmacol Rep. (2012) 64:1400–10. doi: 10.1016/S1734-1140(12)70937-4
79. Tunsirimas N, Pariwatcharakul P, Choovanichvong S, Ratta-apha W. Clozapine-induced agranulocytosis and leukopenia: incidence, associated factors, and rate of hematologic adverse-effects monitoring in psychiatric out-patient services in Thailand. Asian J Psychiatr. (2019) 41:13–6. doi: 10.1016/j.ajp.2019.03.002
80. Capllonch A, de Pablo S, de la Torre A, Morales I. Increase in white cell and neutrophil counts during the first eighteen weeks of treatment with clozapine in patients admitted to a long-term psychiatric care inpatient unit. Rev Psiquiatr Salud Ment. (2018) 11:94–100. doi: 10.1016/j.rpsm.2016.03.005
81. Pessina A, Turlizzi E, Bonomi A, Guizzardi F, Cavicchini L, Croera C, et al. In vitro toxicity of clozapine, olanzapine, and quetiapine on granulocyte-macrophage progenitors (GM-CFU). Pharmacopsychiatry. (2006) 39:20–2. doi: 10.1055/s-2006-931475
82. Goto A, Yoshimi A, Nagai T, Ukigai M, Mouri A, Ozaki N, et al. Human neutrophils show decreased survival upon long-term exposure to clozapine. Hum Psychopharmacol. (2017) 32:1–5. doi: 10.1002/hup.2629
83. Gardiner E, Carroll A, Tooney PA, Cairns MJ. Antipsychotic drug-associated gene-miRNA interaction in T-lymphocytes. Int J Neuropsychopharmacol. (2014) 17:929–43. doi: 10.1017/S1461145713001752
84. Van Der Weide K, Loovers H, Pondman K, Bogers J, Van Der Straaten T, Langemeijer E, et al. Genetic risk factors for clozapine-induced neutropenia and agranulocytosis in a Dutch psychiatric population. Pharmacogenomics J. (2017) 17:471–8. doi: 10.1038/tpj.2016.32
85. Petrikis P, Voulgari PV, Tzallas AT, Boumba VA, Archimandriti DT, Zambetas D, et al. Changes in the cytokine profile in first-episode, drug-naïve patients with psychosis after short-term antipsychotic treatment. Psychiatry Res. (2017) 256:378–83. doi: 10.1016/j.psychres.2017.07.002
86. Capuzzi E, Bartoli F, Crocamo C, Clerici M, Carrà G. Acute variations of cytokine levels after antipsychotic treatment in drug-naïve subjects with a first-episode psychosis: a meta-analysis. Neurosci Biobehav Rev. (2017) 77:122–8. doi: 10.1016/J.NEUBIOREV.2017.03.003
87. Sanader B, Grohmann R, Grötsch P, Schumann T, Toto S, Fernando P, et al. Clozapine-induced DRESS syndrome: a case series from the AMSP multicenter drug safety surveillance project. Pharmacopsychiatry. (2019) 52:156–9. doi: 10.1055/a-0586-8983
88. Yin J, Albert RH, Tretiakova AP, Jameson BA. 5-HT1B receptors play a prominent role in the proliferation of T-lymphocytes. J Neuroimmunol. (2006) 181:68–81. doi: 10.1016/j.jneuroim.2006.08.004
89. Zareie P, Connor B, La Flamme AC. Amelioration of experimental autoimmune encephalomyelitis by clozapine is not associated with defective CD4 T cell responses. J Neuroinflammation. (2017) 14:1–10. doi: 10.1186/s12974-017-0842-5
90. Chen ML, Wu S, Tsai TC, Wang LK, Tsai FM. Regulation of macrophage immune responses by antipsychotic drugs. Immunopharmacol Immunotoxicol. (2013) 35:573–80. doi: 10.3109/08923973.2013.828744
91. O'Sullivan D, Green L, Stone S, Zareie P, Kharkrang M, Fong D, et al. Treatment with the antipsychotic agent, risperidone, reduces disease severity in experimental autoimmune encephalomyelitis. PLoS ONE. (2014) 9: e104430. doi: 10.1371/journal.pone.0104430
92. Nikolić T, Petronijević M, Sopta J, Velimirović M, Stojković T, Jevtić DoŽudić G, et al. Haloperidol affects bones while clozapine alters metabolic parameters - sex specific effects in rats perinatally treated with phencyclidine. BMC Pharmacol Toxicol. (2017) 18:65. doi: 10.1186/s40360-017-0171-4
93. Szuster-Ciesielska A, Słotwinska M, Stachura A, Marmurowska-Michałowska H, Kandefer-Szerszen M. Neuroleptics modulate cytokine and reactive oxygen species production in blood leukocytes of healthy volunteers. Arch Immunol Ther Exp. (2004) 52:59–67.
94. Haack MJ, Bak MLFJ, Beurskens R, Maes M, Stolk LML, Delespaul PAEG. Toxic rise of clozapine plasma concentrations in relation to inflammation. Eur Neuropsychopharmacol. (2003) 13:381–5. doi: 10.1016/S0924-977X(03)00042-7
95. Hefner G, Shams MEE, Unterecker S, Falter T, Hiemke C. Inflammation and psychotropic drugs: the relationship between C-reactive protein and antipsychotic drug levels. Psychopharmacology. (2016) 233:1695–705. doi: 10.1007/s00213-015-3976-0
96. Hung YP, Wang CSM, Yen CN, Chang HC, Chen PS, Lee IH, et al. Role of cytokine changes in clozapine-induced fever: a cohort prospective study. Psychiatry Clin Neurosci. (2017) 71:395–402. doi: 10.1111/pcn.12508
97. Miller BJ, Buckley P, Seabolt W, Mellor A, Kirkpatrick B. Meta-analysis of cytokine alterations in schizophrenia: clinical status and antipsychotic effects. Biol Psychiatry. (2011) 70:663–71. doi: 10.1016/j.biopsych.2011.04.013
98. Ayano G. Second generation antipsychotics: pharmacodynamics, therapeutic effects indications and associated metabolic side effects: review of articles. J Schizophr Res. (2016) 3:1027.
99. Chopko TC, Lindsley CW. Classics in chemical neuroscience: risperidone. ACS Chem Neurosci. (2018) 9:1520–9. doi: 10.1021/acschemneuro.8b00159
100. Roth J. The colorful spectrum of tourette syndrome and its medical, surgical and behavioral therapies. Park Relat Disord. (2018) 46:S75–9. doi: 10.1016/j.parkreldis.2017.08.004
101. Spielmans GI, Berman MI, Linardatos E, Rosenlicht NZ, Perry A, Tsai AC. Adjunctive atypical antipsychotic treatment for major depressive disorder: a meta-analysis of depression, quality of life, and safety outcomes. PLoS Med. (2013) 10:e1001403. doi: 10.1371/journal.pmed.1001403
102. Couturier J, Isserlin L, Spettigue W, Norris M. Psychotropic medication for children and adolescents with eating disorders. Child Adolesc Psychiatr Clin N Am. (2019) 28:583–92. doi: 10.1016/j.chc.2019.05.005
103. Kassm SA, Naja W, Hoertel N, Limosin F. Prise en charge pharmacologique des idées délirantes associées à un syndrome démentiel. Gériatrie Psychol Neuropsychiatr du Vieil. (2019) 17:317–26. doi: 10.1684/PNV.2019.0813
104. Belli H, Ural C, Akbudak M. Borderline personality disorder: bipolarity, mood stabilizers and atypical antipsychotics in treatment. J Clin Med Res. (2012) 4:301–8. doi: 10.4021/jocmr1042w
105. Yuan M, Sperry L, Malhado-Chang N, Duffy A, Wheelock V, Farias S, et al. Atypical antipsychotic therapy in Parkinson's disease psychosis: a retrospective study. Brain Behav. (2017) 7:1–6. doi: 10.1002/brb3.639
106. Courtois C, Sonis J, Brown L, Seattle W, Cook J, Fairbank J, et al. Summary of the clinical practice guideline for the treatment of posttraumatic stress disorder (PTSD) in adults. Am Psychol. (2019) 74:596–607. doi: 10.1037/amp0000473
108. Madaan V, Bestha DP, Kolli V, Jauhari S, Burket RC. Clinical utility of the risperidone formulations in the management of schizophrenia. Neuropsychiatr Dis Treat. (2011) 7:611–20. doi: 10.2147/NDT.S14385
109. Kusumi I, Boku S, Takahashi Y. Psychopharmacology of atypical antipsychotic drugs: from the receptor binding profile to neuroprotection and neurogenesis. Psychiatry Clin Neurosci. (2015) 69:243–58. doi: 10.1111/pcn.12242
110. Grahovac T, RuŽić K, Medved P, Pavešić-Radonja A, Dadić-Hero E. Hyperprolactinaemia - a risperidone side-effect. Psychiatr Danub. (2010) 22:120–22.
111. Park YM, Lee SH, Lee BH, Lee KY, Lee KS, Kang SG, et al. Prolactin and macroprolactin levels in psychiatric patients receiving atypical antipsychotics: a preliminary study. Psychiatry Res. (2016) 239:184–9. doi: 10.1016/j.psychres.2016.03.015
112. Lally J, Ajnakina O, Stubbs B, Williams HR, Colizzi M, Carra E, et al. Hyperprolactinaemia in first episode psychosis - A longitudinal assessment. Schizophr Res. (2017) 189:117–25. doi: 10.1016/j.schres.2017.07.037
113. Bonete Llácer JM, Martínez Hortelano A, Richart Albelda B. Hyperprolactinemia in psychotic patients treated in monotherapy with long-acting injectable antipsychotics. Int J Psychiatry Clin Pract. (2019) 23:189–93. doi: 10.1080/13651501.2019.1576905
114. Cešková E, Prikryl R, Kašpárek T, Ondrušová M. Prolactin levels in risperidone treatment of first-episode schizophrenia. Int J Psychiatry Clin Pract. (2004) 8:31–6. doi: 10.1080/1365150031004786
115. Pérez-Iglesias R, Mata I, Martínez-García O, Garcia-Unzueta MT, Amado JA, Valdizán EM, et al. Long-term effect of haloperidol, olanzapine, and risperidone on plasma prolactin levels in patients with first-episode psychosis. J Clin Psychopharmacol. (2012) 32:804–8. doi: 10.1097/JCP.0b013e318272688b
116. Charan A, Shewade DG, Rajkumar RP, Chandrasekaran A. Relation between serum prolactin levels and antipsychotic response to risperidone in patients with schizophrenia. Psychiatry Res. (2016) 240:209–13. doi: 10.1016/j.psychres.2016.04.001
117. Liu J, Sun J, Shen X, Guo W, Zhi S, Song G, et al. Randomized controlled trial comparing changes in serum prolactin and weight among female patients with first-episode schizophrenia over 12 months of treatment with risperidone or quetiapine. Shanghai Arch Psychiatry. (2014) 26:88–94. doi: 10.3969/j.issn.1002-0829.2014.02
118. Suzuki Y, Fukui N, Watanabe J, Ono S, Sugai T, Tsuneyama N, et al. Gender differences in the relationship between the risperidone metabolism and the plasma prolactin levels in psychiatric patients. Prog Neuro-Psychopharmacol Biol Psychiatry. (2010) 34:1266–8. doi: 10.1016/j.pnpbp.2010.07.003
119. Kinon BJ, Liu-Seifert H, Stauffer VL, Jacob J. Bone loss associated with hyperprolactinemia in patients with schizophrenia: are there gender differences? Clin Schizophr Relat Psychoses. (2013) 7:115–23. doi: 10.3371/CSRP.KISE.020113
120. An F-R, Yang R, Wang Z-M, Ungvari GS, Ng CH, Chiu HFK, et al. Hyperprolactinemia, prolactin-related side effects and quality of life in Chinese psychiatric patients. Compr Psychiatry. (2016) 71:71–6. doi: 10.1016/j.comppsych.2016.08.009
121. Takechi K, Yoshioka Y, Kawazoe H, Tanaka M, Takatori S, Kobayashi M, et al. Psychiatric patients with antipsychotic drug-induced hyperprolactinemia and menstruation disorders. Biol Pharm Bull. (2017) 40:1775–8. doi: 10.1248/bpb.b17-00053
122. Holla SN, Vittalrao AMB, Kamath A, Kamalkishore MK, Ommurugan B. Risperidone induced granulomatous mastitis secondary to hyperprolactinemia in a non-pregnant woman-a rare case report in a bipolar disorder. J Clin Diagnostic Res. (2017) 11:FD01–3. doi: 10.7860/JCDR/2017/20733.9278
123. Shagufta S, Farooq F, Khan AM, Dar K, Mohit A. Risperidone-induced amenorrhea in floridly psychotic female. Cureus. (2017) 9:e1683. doi: 10.7759/cureus.1683
124. Sakaguchi S, Aizawa K. Galactorrhea induced by risperidone. Intern Med. (2020) 58:3609–10. doi: 10.2169/internalmedicine.3224-19
125. Kawabe K, Ueno SI. A case of acute pancreatitis associated with risperidone treatment. Clin Psychopharmacol Neurosci. (2014) 12:67–8. doi: 10.9758/cpn.2014.12.1.67
126. Rad F, Buica AM, Anghel GC, Stancu M, Dobrescu I. Hormonal imbalance and pituitary adenoma during antipsychotic treatment in an adolescent with bipolar affective disorder. Riv Psichiatr. (2019) 54:37–9. doi: 10.1708/3104.30939
127. Hongkaew Y, Ngamsamut N, Puangpetch A, Vanwong N, Srisawasdi P, Chamnanphon M, et al. Hyperprolactinemia in thai children and adolescents with autism spectrum disorder treated with risperidone. Neuropsychiatr Dis Treat. (2015) 11:191–6. doi: 10.2147/NDT.S76276
128. Roke Y, Buitelaar JK, Boot AM, Tenback D, Van Harten PN. Risk of hyperprolactinemia and sexual side effects in males 10-20 years old diagnosed with autism spectrum disorders or disruptive behavior disorder and treated with risperidone. J Child Adolesc Psychopharmacol. (2012) 22:432–9. doi: 10.1089/cap.2011.0109
129. Margari L, Matera E, Petruzzelli MG, Simone M, Lamanna AL, Pastore A, et al. Prolactin variations during risperidone therapy in a sample of drug-naive children and adolescents. Int Clin Psychopharmacol. (2015) 30:103–8. doi: 10.1097/YIC.0000000000000063
130. Balijepalli C, Druyts E, Zoratti MJ, Wu P, Kanji S, Rabheru K, et al. Change in prolactin levels in pediatric patients given antipsychotics for schizophrenia and schizophrenia spectrum disorders: a network meta-analysis. Schizophr Res Treat. (2018) 2018:543034. doi: 10.1155/2018/1543034
131. Dos Santos Júnior A, Henriques TB, De Mello MP, Neto APF, Paes LA, Torre OH Della, et al. Hyperprolactinemia in children and adolescents with use of risperidone: clinical and molecular genetics aspects. J Child Adolesc Psychopharmacol. (2015) 25:738–48. doi: 10.1089/cap.2015.0094
132. Osmanova DZ, Freidin MB, Fedorenko OY, Pozhidaev IV, Boiko AS, Vyalova NM, et al. A pharmacogenetic study of patients with schizophrenia from West Siberia gets insight into dopaminergic mechanisms of antipsychotic-induced hyperprolactinemia. BMC Med Genet. (2019) 20(Suppl. 1):47. doi: 10.1186/s12881-019-0773-3
133. Sun WW, Li LY, Huang XF, Shi YC, Yang HQ, Song ZY, et al. The central mechanism of risperidone-induced hyperprolactinemia. Prog Neuro-Psychopharmacology Biol Psychiatry. (2017) 76:134–9. doi: 10.1016/j.pnpbp.2017.03.009
134. Jiang XJ, Wu FX, Zhang JP, Shi L, Hu JQ, Zhu HZ, et al. Effects of risperidone and aripiprazole on serum levels of prolactin, testosterone and estradiol in female patients with schizophrenia. Drug Res. (2018) 68:410–14. doi: 10.1055/s-0044-102093
135. Piriu G, Torac E, Gaman LE, Iosif L, Tivig IC, Delia C, et al. Clozapine and risperidone influence on cortisol and estradiol levels in male patients with schizophrenia. J Med Life. (2015) 8:548–51.
136. Konarzewska B, Galinska-Skok B, Waszkiewicz N, Łazarczyk-Kirejczyk J, Małus A, Simonienko K, et al. Association between serum testosterone levels, body mass index (BMI) and insulin in male patients with schizophrenia treated with atypical antipsychotics - Olanzapine or risperidone. Neuroendocrinol Lett. (2014) 35:50–7.
137. Bishop JR, Rubin LH, Reilly JL, Pavuluri MN, Sweeney JA. Risperidone-associated prolactin elevation and markers of bone turnover during acute treatment. Ther Adv Psychopharmacol. (2012) 2:95–102. doi: 10.1177/2045125312442080
138. Yanik T, Kursungoz C, Sutcigil L, Ak M. Weight Gain in risperidone therapy: investigation of peripheral hypothalamic neurohormone levels in psychotic patients. J Clin Psychopharmacol. (2013) 33:608–13. doi: 10.1097/JCP.0b013e318297980e
139. Tsai MC, Chang CM, Liu CY, Chang PY, Huang TL. Association of serum levels of leptin, ghrelin, and adiponectin in schizophrenic patients and healthy controls. Int J Psychiatry Clin Pract. (2011) 15:106–11. doi: 10.3109/13651501.2010.550400
140. Doknic M, Maric NP, Britvic D, Pekic S, Damjanovic A, Miljic D, et al. Bone remodeling, bone mass and weight gain in patients with stabilized schizophrenia in real-life conditions treated with long-acting injectable risperidone. Neuroendocrinology. (2011) 94:246–54. doi: 10.1159/000329391
141. Srisawasdi P, Vanwong N, Hongkaew Y, Puangpetch A, Vanavanan S, Intachak B, et al. Impact of risperidone on leptin and insulin in children and adolescents with autistic spectrum disorders. Clin Biochem. (2017) 50:678–85. doi: 10.1016/j.clinbiochem.2017.02.003
142. Baeza I, Vigo L, de la Serna E, Calvo-Escalona R, Merchán-Naranjo J, Rodríguez-Latorre P, et al. The effects of antipsychotics on weight gain, weight-related hormones and homocysteine in children and adolescents: a 1-year follow-up study. Eur Child Adolesc Psychiatry. (2017) 26:35–46. doi: 10.1007/s00787-016-0866-x
143. Smith RC, Rachakonda S, Dwivedi S, Davis JM. Olanzapine and risperidone effects on appetite and ghrelin in chronic schizophrenic patients. Psychiatry Res. (2012) 199:159–63. doi: 10.1016/J.PSYCHRES.2012.03.011
144. Endomba FT, Tankeu AT, Nkeck JR, Tochie JN. Leptin and psychiatric illnesses: does leptin play a role in antipsychotic-induced weight gain? Lipids Health Dis. (2020) 19:22. doi: 10.1186/s12944-020-01203-z
145. Sugai T, Suzuki Y, Fukui N, Ono S, Watanabe J, Tsuneyama N, et al. Dysregulation of adipocytokines related to second-generation antipsychotics in normal fasting glucose patients with schizophrenia. J Clin Psychopharmacol. (2012) 32:390–93. doi: 10.1097/JCP.0b013e3182524393
146. Scahill L, Jeon S, Boorin SJ, McDougle CJ, Aman MG, Dziura J, et al. Weight gain and metabolic consequences of risperidone in young children with autism spectrum disorder. J Am Acad Child Adolesc Psychiatry. (2016) 55:415–23. doi: 10.1016/j.jaac.2016.02.016
147. Bartoli F, Lax A, Crocamo C, Clerici M, Carrà G. Plasma adiponectin levels in schizophrenia and role of second-generation antipsychotics: a meta-analysis. Psychoneuroendocrinology. (2015) 56:179–89. doi: 10.1016/j.psyneuen.2015.03.012
148. Bartoli F, Crocamo C, Clerici M, Carrà G. Second-generation antipsychotics and adiponectin levels in schizophrenia: a comparative meta-analysis. Eur Neuropsychopharmacol. (2015) 25:1767–74. doi: 10.1016/j.euroneuro.2015.06.011
149. Sackett G, Unis A, Crouthamel B. Some effects of risperidone and quetiapine on growth parameters and hormone levels in young pigtail macaques. J Child Adolesc Psychopharmacol. (2010) 20:489–93. doi: 10.1089/cap.2010.0018
150. Kursungoz C, Ak M, Yanik T. Effects of risperidone treatment on the expression of hypothalamic neuropeptide in appetite regulation in Wistar rats. Brain Res. (2015) 1596:146–55. doi: 10.1016/j.brainres.2014.10.070
151. Horska K, Ruda-Kucerova J, Karpisek M, Suchy P, Opatrilova R, Kotolova H. Depot risperidone-induced adverse metabolic alterations in female rats. J Psychopharmacol. (2017) 31:487–99. doi: 10.1177/0269881117691466
152. Savoy YE, Ashton MA, Miller MW, Nedza FM, Spracklin DK, Hawthorn MH, et al. Differential effects of various typical and atypical antipsychotics on plasma glucose and insulin levels in the mouse: evidence for the involvement of sympathetic regulation. Schizophr Bull. (2010) 36:410–8. doi: 10.1093/schbul/sbn104
153. Manfredi G, Solfanelli A, Dimitri G, Cuomo I, Sani G, Kotzalidis GD, et al. Risperidone-induced leukopenia: a case report and brief review of literature. Gen Hosp Psychiatry. (2013) 35:102.e3–102.e6. doi: 10.1016/j.genhosppsych.2012.03.009
154. Noto C, Ota VK, Gouvea ES, Rizzo LB, Spindola LMN, Honda PHS, et al. Effects of risperidone on cytokine profile in drug-naïve first-episode psychosis. Int J Neuropsychopharmacol. (2014) 18:1–8. doi: 10.1093/ijnp/pyu042
155. Elmorsy E, Al-Ghafari A, Aggour AM, Mosad SM, Khan R, Amer S. Effect of antipsychotics on mitochondrial bioenergetics of rat ovarian theca cells. Toxicol Lett. (2017) 272:94–100. doi: 10.1016/j.toxlet.2017.03.018
156. Tseng CC. Neutropenia during risperidone treatment. J Neuropsychiatry Clin Neurosci. (2011) 23:E19. doi: 10.1176/jnp.23.4.jnpe19
157. Morrison M, Schultz A, Sanchez DL, Catalano MC, Catalano G. Leukopenia associated with risperidone treatment. Curr Drug Saf. (2017) 12:126–30. doi: 10.2174/1574886312666170531072837
158. Sy-Cherng Woon L, Tee CK, Gan LLY, Deang KT, Chan LF. Olanzapine-induced and risperidone-induced leukopenia: a case of synergistic adverse reaction? J Psychiatr Pract. (2018) 24:121–4. doi: 10.1097/PRA.0000000000000292
159. Raj V, Druitt T, Purushothaman S, Dunsdon J. Risperidone/paliperidone induced neutropenia and lymphopenia. Aust N Z J Psychiatry. (2013) 47:291–2. doi: 10.1177/0004867412460594
160. Kailasam VK, Chima V, Nnamdi U, Sharma K, Shah K. Risperidone-induced reversible neutropenia. Neuropsychiatr Dis Treat. (2017) 13:1975–77. doi: 10.2147/NDT.S141472
161. Bhattacharjee D, Yerrapragada D, Chogtu B, Thomson SR. Risperidone induced isolated thrombocytopenia: a rare adverse event. Psychopharmacol Bull. (2018) 48:47–9.
162. Rizos E, Tsigkaropoulou E, Lambrou P, Kanakaki M, Chaniotou A, Alevyzakis E, et al. Risperidone-induced acute eosinophilic pneumonia. In Vivo. (2013) 27:651–4.
163. Rettenbacher MA, Hofer A, Kemmler G, Fleischhacker WW. Neutropenia induced by second generation antipsychotics: a prospective investigation. Pharmacopsychiatry. (2010) 43:41–4. doi: 10.1055/s-0030-1249071
164. Diaz FJ, Pérez-Iglesias R, Mata I, Martínez-Garcia O, Vázquez-Barquero JL, de Leon J, et al. Possible effects of some antipsychotic drugs on C-reactive protein in a drug-naïve psychotic sample. Schizophr Res. (2010) 121:207–12. doi: 10.1016/j.schres.2010.06.002
165. Lin CC, Chang CM, Liu CY, Huang TL. Increased high-sensitivity C-reactive protein levels in Taiwanese schizophrenic patients. Asia-Pacific Psychiatry. (2013) 5:E58–63. doi: 10.1111/appy.12078
166. Hefner G, Falter T, Bruns K, Hiemke C. Elevated risperidone serum concentrations during acute inflammation, two cases. Int J Psychiatry Med. (2015) 50:335–44. doi: 10.1177/0091217415610313
167. Ajami A, Hosseini SH, Taghipour M, Khalilian A. Changes in serum levels of brain derived neurotrophic factor and nerve growth factor-beta in schizophrenic patients before and after treatment. Scand J Immunol. (2014) 80:36–42. doi: 10.1111/sji.12158
168. Kudlek Mikulic S, Mihaljevic-Peles A, Sagud M, Bajs Janovic M, Ganoci L, Grubisin J, et al. Brain-derived neurotrophic factor serum and plasma levels in the treatment of acute schizophrenia with olanzapine or risperidone: 6-week prospective study. Nord J Psychiatry. (2017) 71:513–20. doi: 10.1080/08039488.2017.1340518
169. Yoshimura R, Ueda N, Hori H, Ikenouchi-Sugita A, Umene-Nakano W, Nakamura J. Different patterns of longitudinal changes in plasma levels of catecholamine metabolites and brain-derived neurotrophic factor after administration of atypical antipsychotics in first episode untreated schizophrenic patients. World J Biol Psychiatry. (2010) 11:256–61. doi: 10.3109/15622970802309617
170. Scherf-Clavel M, Weidner A, Deckert J, Menke A, Unterecker S. Pathological concentration of c-reactive protein is correlated to increased concentrations of quetiapine, but not of risperidone, olanzapine and aripiprazole in a naturalistic setting. Pharmacopsychiatry. (2019) 53:30–5. doi: 10.1055/a-0869-8053
171. Rizos EN, Papadopoulou A, Laskos E, Michalopoulou PG, Kastania A, Vasilopoulos D, et al. Reduced serum BDNF levels in patients with chronic schizophrenic disorder in relapse, who were treated with typical or atypical antipsychotics. World J Biol Psychiatry. (2010) 11:251–5. doi: 10.3109/15622970802182733
172. Bosshart H. Supra-therapeutic plasma concentrations of haloperidol induce moderate inhibition of lipopolysaccharide-induced interleukin-8 release in human monocytes. Ann Transl Med. (2016) 4:396. doi: 10.21037/atm.2016.10.56
173. McNamara RK, Jandacek R, Rider T, Tso P. Chronic risperidone normalizes elevated pro-inflammatory cytokine and C-reactive protein production in omega-3 fatty acid deficient rats. Eur J Pharmacol. (2011) 652:152–6. doi: 10.1016/j.ejphar.2010.11.010
174. Juncal-Ruiz M, Riesco-Dávila L, Ortiz-García de la Foz V, Martínez-Garcia O, Ramírez-Bonilla M, Ocejo-Viñals JG, et al. Comparison of the anti-inflammatory effect of aripiprazole and risperidone in 75 drug-naïve first episode psychosis individuals: a 3 months randomized study. Schizophr Res. (2018) 202:226–33. doi: 10.1016/j.schres.2018.06.039
175. Ajami A, Abedian F, Hamzeh Hosseini S, Akbarian E, Alizadeh-Navaei R, Taghipour M. Serum TNF-α, IL-10 and IL-2 in schizophrenic patients before and after treatment with risperidone and clozapine. Iran J Immunol. (2014) 11:200–09.
176. De Witte L, Tomasik J, Schwarz E, Guest PC, Rahmoune H, Kahn RS, et al. Cytokine alterations in first-episode schizophrenia patients before and after antipsychotic treatment. Schizophr Res. (2014) 154:23–9. doi: 10.1016/j.schres.2014.02.005
177. Lo LH, Shiea J, Huang TL. Rapid detection of alteration of serum IgG in patients with schizophrenia after risperidone treatment by matrix-assisted laser desorption ionization/time-of-flight mass spectrometry. Rapid Commun Mass Spectrom. (2016) 30:2645–9. doi: 10.1002/rcm.7753
178. Song X, Fan X, Li X, Zhang W, Gao J, Zhao J, et al. Changes in pro-inflammatory cytokines and body weight during 6-month risperidone treatment in drug naïve, first-episode schizophrenia. Psychopharmacology. (2014) 231:319–25. doi: 10.1007/s00213-013-3382-4
179. Choi JE, Widjaja F, Careaga M, Bent S, Ashwood P, Hendren RL. Change in plasma cytokine levels during risperidone treatment in children with autism. J Child Adolesc Psychopharmacol. (2014) 24:586–9. doi: 10.1089/cap.2013.0108
180. Chen ML, Wu S, Tsai TC, Wang LK, Tsai FM. Regulation of neutrophil phagocytosis of Escherichia coli by antipsychotic drugs. Int Immunopharmacol. (2014) 23:550–7. doi: 10.1016/j.intimp.2014.09.030
181. Al-Amin MM, Uddin MMN, Reza HM. Effects of antipsychotics on the inflammatory response system of patients with schizophrenia in peripheral blood mononuclear cell cultures. Clin Psychopharmacol Neurosci. (2013) 11:144–51. doi: 10.9758/cpn.2013.11.3.144
182. Krause D, Weidinger E, Dippel C, Riedel M, Schwarz MJ, Müller N, et al. Impact of different antipsychotics on cytokines and tryptophan metabolites in stimulated cultures from patients with schizophrenia. Psychiatr Danub. (2013) 25:389–97.
183. Noto MN, Maes M, Nunes SOV, Ota VK, Rossaneis AC, Verri WA, et al. Activation of the immune-inflammatory response system and the compensatory immune-regulatory system in antipsychotic naive first episode psychosis. Eur Neuropsychopharmacol. (2019) 29:416–31. doi: 10.1016/j.euroneuro.2018.12.008
184. Ding M, Song X, Zhao J, Gao J, Li X, Yang G, et al. Activation of Th17 cells in drug naïve, first episode schizophrenia. Prog Neuro-Psychopharmacology Biol Psychiatry. (2014) 51:78–82. doi: 10.1016/j.pnpbp.2014.01.001
185. Sárvári AK, Veréb Z, Uray IP, Fésüs L, Balajthy Z. Atypical antipsychotics induce both proinflammatory and adipogenic gene expression in human adipocytes in vitro. Biochem Biophys Res Commun. (2014) 450:1383–9. doi: 10.1016/j.bbrc.2014.07.005
186. Kéri S, Szabó C, Kelemen O. Antipsychotics influence Toll-like receptor (TLR) expression and its relationship with cognitive functions in schizophrenia. Brain Behav Immun. (2017) 62:256–64. doi: 10.1016/j.bbi.2016.12.011
187. Mantere O, Trontti K, García-González J, Balcells I, Saarnio S, Mäntylä T, et al. Immunomodulatory effects of antipsychotic treatment on gene expression in first-episode psychosis. J Psychiatr Res. (2019) 109:18–26. doi: 10.1016/j.jpsychires.2018.11.008
188. Aboul-Fotouh S, Elgayar N. Atypical antipsychotics such as risperidone, but not paliperidone, worsen vascular endothelial function via upregulation of adhesion molecules VCAM-1, ICAM-1, and E-selectin in diabetic rats. Can J Physiol Pharmacol. (2013) 91:1119–26. doi: 10.1139/cjpp-2013-0185
189. Da Cruz Jung IE, Machado AK, Da Cruz IBM, Barbisan F, Azzolin VF, Duarte T, et al. Haloperidol and risperidone at high concentrations activate an in vitro inflammatory response of RAW 264.7 macrophage cells by induction of apoptosis and modification of cytokine levels. Psychopharmacology. (2016) 233:1715–23. doi: 10.1007/s00213-015-4079-7
190. Bymaster FP, Calligaro DO, Falcone JF, Marsh RD, Moore NA, Tye NC, et al. Radioreceptor binding profile of the atypical antipsychotic olanzapine. Neuropsychopharmacology. (1996) 14:87–96. doi: 10.1016/0893-133X(94)00129-N
191. Eisenberg Center at Oregon Health & Science University EC at OH& S. Off -Label Use of Atypical Antipsychotic Drugs: A Summary for Clinicians and Policymakers. Agency for Healthcare Research and Quality (US) (2007). Available online at: http://www.ncbi.nlm.nih.gov/pubmed/21938813 (accessed February 20, 2020)
192. Thomas K, Saadabadi A. Olanzapine. (2019). Available at: https://www.ncbi.nlm.nih.gov/books/NBK532903/ (accessed February 24, 2020)
193. Evison M, Holme J, Alaloul M, Doran H, Bishop P, Booton R, et al. Olanzapine-induced eosinophilic pleuritis. Respir Med Case Rep. (2015) 14:24–6. doi: 10.1016/J.RMCR.2014.11.007
194. Huang J, Yu Y, Lin W, Zhang D, Deng Z, Ding Q. Olanzapine-induced peripheral eosinophilia and eosinophilic pleural effusion: a case report. Medicine. (2018) 97:e9996. doi: 10.1097/MD.0000000000009996
195. Ng W, Kennar R, Uetrecht J. Effect of clozapine and olanzapine on neutrophil kinetics: implications for drug-induced agranulocytosis. Chem Res Toxicol. (2014) 27:1104–08. doi: 10.1021/tx500183x
196. Raz A, Bergman R, Eilam O, Yungerman T, Hayek T. A case report of olanzapine-induced hypersensitivity syndrome. Am J Med Sci. (2001) 321:156–8. doi: 10.1097/00000441-200102000-00008
198. Matei VP, Purnichi T, Mihailescu A, Grigoras R. Prolactin level in patients with first episode schizophrenia treated for one year with atypical antipsychotics. Acta Endocrinol. (2018) 14:483–90. doi: 10.4183/aeb.2018.483
199. Kishimoto T, Hagi K, Nitta M, Kane JM, Correll CU. Long-term effectiveness of oral second-generation antipsychotics in patients with schizophrenia and related disorders: a systematic review and meta-analysis of direct head-to-head comparisons. World Psychiatry. (2019) 18:208–24. doi: 10.1002/wps.20632
200. Yasui-Furukori N, Furukori H, Sugawara N, Tsuchimine S, Fujii A, Inoue Y, et al. Prolactin fluctuation over the course of a day during treatments with three atypical antipsychotics in schizophrenic patients. Hum Psychopharmacol Clin Exp. (2010) 25:236–42. doi: 10.1002/hup.1110
201. Barata PC, Santos MJ, Melo JC, Maia T. Olanzapine-induced hyperprolactinemia: two case reports. Front Pharmacol. (2019) 10:846. doi: 10.3389/fphar.2019.00846
202. Suzuki Y, Sugai T, Fukui N, Watanabe J, Ono S, Tsuneyama N, et al. Differences in plasma prolactin levels in patients with schizophrenia treated on monotherapy with five second-generation antipsychotics. Schizophr Res. (2013) 145:116–9. doi: 10.1016/J.SCHRES.2012.12.027
203. Wu XL, Wang JH, Hu SH, Tao J. Serum prolactin levels and the acute-phase efficacy in drug-naïve schizophrenia treated with ziprasidone and olanzapine (translated version). East Asian Arch Psychiatry. (2012) 22:7–11.
204. Feng S, Melkersson K. Metabolic parameters and long-term antipsychotic treatment: a comparison between patients treated with clozapine or olanzapine. Neuro Endocrinol Lett. (2012) 33:493–8.
205. Davey KJ, O'Mahony SM, Schellekens H, O'Sullivan O, Bienenstock J, Cotter PD, et al. Gender-dependent consequences of chronic olanzapine in the rat: effects on body weight, inflammatory, metabolic and microbiota parameters. Psychopharmacology. (2012) 221:155–69. doi: 10.1007/s00213-011-2555-2
206. Sacher J, Mossaheb N, Spindelegger C, Klein N, Geiss-Granadia T, Sauermann R, et al. Effects of olanzapine and ziprasidone on glucose tolerance in healthy volunteers. Neuropsychopharmacology. (2008) 33:1633–41. doi: 10.1038/sj.npp.1301541
207. Huang M, Yu L, Pan F, Lu S, Hu S, Hu J, et al. A randomized, 13-week study assessing the efficacy and metabolic effects of paliperidone palmitate injection and olanzapine in first-episode schizophrenia patients. Prog Neuro-Psychopharmacology Biol Psychiatry. (2018) 81:122–30. doi: 10.1016/j.pnpbp.2017.10.021
208. Li H, Peng S, Li S, Liu S, Lv Y, Yang N, et al. Chronic olanzapine administration causes metabolic syndrome through inflammatory cytokines in rodent models of insulin resistance. Sci Rep. (2019) 9:1582. doi: 10.1038/s41598-018-36930-y
209. Raposo NRB, Ferreira AS, Gattaz WF. Body mass index increase, serum leptin, adiponectin, neuropeptide y and lipid levels during treatment with olanzapine and haloperidol. Pharmacopsychiatry. (2011) 44:169–72. doi: 10.1055/s-0031-1280793
210. Lin Y, Peng Y, He S, Xu J, Shi Y, Su Y, et al. Serum IL-1ra, a novel biomarker predicting olanzapine-induced hypercholesterolemia and hyperleptinemia in schizophrenia. Prog Neuro-Psychopharmacology Biol Psychiatry. (2018) 84:71–8. doi: 10.1016/J.PNPBP.2018.01.020
211. Tsuneyama N, Suzuki Y, Sawamura K, Sugai T, Fukui N, Watanabe J, et al. Effect of serum leptin on weight gain induced by olanzapine in female patients with schizophrenia. PLoS ONE. (2016) 11:e0149518. doi: 10.1371/journal.pone.0149518
212. Panariello F, Polsinelli G, Borlido C, Monda M, De Luca V. The role of leptin in antipsychotic-induced weight gain: genetic and non-genetic factors. J Obes. (2012) 2012:572848. doi: 10.1155/2012/572848
213. Basoglu C, Oner O, Ates AM, Algul A, Semiz UB, Ebrinc S, et al. Association between symptom improvement and change of body mass index, lipid profile, and leptin, ghrelin, and cholecystokinin levels during 6-week olanzapine treatment in patients with first-episode psychosis. J Clin Psychopharmacol. (2010) 30:636–8. doi: 10.1097/JCP.0b013e3181f0580e
214. Chen VC-H, Wang T-N, Lu M-L, Chou J-Y, Ju P-C, Wu J-Y, et al. Weight gain and ghrelin level after olanzapine monotherapy. Prog Neuro-Psychopharmacology Biol Psychiatry. (2011) 35:632–5. doi: 10.1016/j.pnpbp.2011.01.010
215. Goetz RL, Miller BJ. Meta-analysis of ghrelin alterations in schizophrenia: effects of olanzapine. Schizophr Res. (2019) 206:21–6. doi: 10.1016/J.SCHRES.2018.11.036
216. Zabotina T, Nasyrova RF, Sosin DN, Sosina Ershov, Grunina N, Krupitsky. The effect of antipsychotic drug on monoamine receptors in peripheral blood mononuclear cells: Affinity linked mechanism. Biomeditsinskaya Khimiya. (2018) 64:201–07. doi: 10.18097/PBMC20186402201
217. Stapel B, Sieve I, Falk CS, Bleich S, Hilfiker-Kleiner D, Kahl KG. Second generation atypical antipsychotics olanzapine and aripiprazole reduce expression and secretion of inflammatory cytokines in human immune cells. J Psychiatr Res. (2018) 105:95–102. doi: 10.1016/j.jpsychires.2018.08.017
218. Zhang C, Fang X, Yao P, Mao Y, Cai J, Zhang Y, et al. Metabolic adverse effects of olanzapine on cognitive dysfunction: a possible relationship between BDNF and TNF-alpha. Psychoneuroendocrinology. (2017) 81:138–43. doi: 10.1016/J.PSYNEUEN.2017.04.014
219. Hatziagelaki E, Tsiavou A, Gerasimou C, Vavougios G, Spathis A, Laskos E, et al. Effects of olanzapine on cytokine profile and brain-derived neurotrophic factor in drug-naive subjects with first-episode psychosis. Exp Ther Med. (2019) 17:3071–6. doi: 10.3892/etm.2019.7285
220. DeVane CL, Nemeroff CB. Clinical pharmacokinetics of quetiapine. Clin Pharmacokinet. (2001) 40:509–22. doi: 10.2165/00003088-200140070-00003
221. Riedel M, Müller N, Strassnig M, Spellmann I, Severus E, Möller H-J. Quetiapine in the treatment of schizophrenia and related disorders. Neuropsychiatr Dis Treat. (2007) 3:219–35. doi: 10.2147/nedt.2007.3.2.219
222. Kim D-W, Weon K-Y, Hong E-P, Chung EK, Lee K-T. Comparative physicochemical and pharmacokinetic properties of quetiapine and its active metabolite norquetiapine. Chem Pharm Bull. (2016) 64:1546–54. doi: 10.1248/cpb.c16-00223
223. López-Muñoz F, Álamo C. Active metabolites as antidepressant drugs: the role of norquetiapine in the mechanism of action of quetiapine in the treatment of mood disorders. Front Psychiatry. (2013) 4:102. doi: 10.3389/fpsyt.2013.00102
224. Carboni L, Negri M, Michielin F, Bertani S, Fratte SD, Oliosi B, et al. Slow dissociation of partial agonists from the D2 receptor is linked to reduced prolactin release. Int J Neuropsychopharmacol. (2012) 15:645–56. doi: 10.1017/S1461145711000824
225. Nunes LVA, Moreira HC, Razzouk D, Nunes SOV, Mari JDJ. Strategies for the treatment of antipsychotic-induced sexual dysfunction and/or hyperprolactinemia among patients of the schizophrenia spectrum: a review. J Sex Marital Ther. (2012) 38:281–301. doi: 10.1080/0092623X.2011.606883
226. Bushe C, Sniadecki J, Bradley AJ, Poole Hoffmann V. Comparison of metabolic and prolactin variables from a six-month randomised trial of olanzapine and quetiapine in schizophrenia. J Psychopharmacol. (2010) 24:1001–9. doi: 10.1177/0269881108101783
227. Oriot P, Feys J-L, Mertens de Wilmars S, Misson A, Ayache L, Fagnart O, et al. Insulin sensitivity, adjusted beta-cell function and adiponectinaemia among lean drug-naive schizophrenic patients treated with atypical antipsychotic drugs: a nine-month prospective study. Diabetes Metab. (2008) 34:490–6. doi: 10.1016/j.diabet.2008.03.003
228. Ngai YF, Sabatini P, Nguyen D, Davidson J, Chanoine J-P, Devlin AM, et al. Quetiapine treatment in youth is associated with decreased insulin secretion. J Clin Psychopharmacol. (2014) 34:359–64. doi: 10.1097/JCP.0000000000000118
229. Melkersson K, Jansson E. The atypical antipsychotics quetiapine, risperidone and ziprasidone do not increase insulin release in vitro. Neuro Endocrinol Lett. (2005) 26:205–8.
230. McNamara RK, Jandacek R, Rider T, Tso P, Cole-Strauss A, Lipton JW. Atypical antipsychotic medications increase postprandial triglyceride and glucose levels in male rats: relationship with stearoyl-CoA desaturase activity. Schizophr Res. (2011) 129:66–73. doi: 10.1016/j.schres.2011.03.016
231. Ran H, Zhu Y, Deng R, Zhang Q, Liu X, Feng M, et al. Stearoyl-CoA desaturase-1 promotes colorectal cancer metastasis in response to glucose by suppressing PTEN. J Exp Clin Cancer Res. (2018) 37:54. doi: 10.1186/s13046-018-0711-9
232. Smith GC, Chaussade C, Vickers M, Jensen J, Shepherd PR. Atypical antipsychotic drugs induce derangements in glucose homeostasis by acutely increasing glucagon secretion and hepatic glucose output in the rat. Diabetologia. (2008) 51:2309–17. doi: 10.1007/s00125-008-1152-3
233. De Borja Gonçalves Guerra A, Castel S, Benedito-Silva AA, Calil HM. Neuroendocrine effects of quetiapine in healthy volunteers. Int J Neuropsychopharmacol. (2005) 8:49–57. doi: 10.1017/S1461145704004705
234. Cohrs S, Röher C, Jordan W, Meier A, Huether G, Wuttke W, et al. The atypical antipsychotics olanzapine and quetiapine, but not haloperidol, reduce ACTH and cortisol secretion in healthy subjects. Psychopharmacology. (2006) 185:11–8. doi: 10.1007/s00213-005-0279-x
235. Kontaxakis VP, Karaiskos D, Havaki-Kontaxaki BJ, Ferentinos P, Papadimitriou GN. Can quetiapine-induced hypothyroidism be reversible without quetiapine discontinuation? Clin Neuropharmacol. (2009) 32:295–6. doi: 10.1097/WNF.0b013e3181a8cbcc
236. Crépeau-Gendron G, L'Heureux S. Quetiapine XR-induced neutropenia: is a clozapine trial still possible for treatment-resistant schizophrenia? A case report. Early Interv Psychiatry. (2015) 9:151–5. doi: 10.1111/eip.12134
237. Croarkin P, Rayner T. Acute neutropenia in a patient treated with quetiapine. Psychosomatics. (2001) 42:368. doi: 10.1176/appi.psy.42.4.368
238. Cowan C, Oakley C. Leukopenia and neutropenia induced by quetiapine. Prog Neuropsychopharmacol Biol Psychiatry. (2007) 31:292–4. doi: 10.1016/j.pnpbp.2006.07.003
239. Almaghrebi AH. Safety of a clozapine trial following quetiapine-induced leukopenia: a case report. Curr Drug Saf. (2019) 14:80–3. doi: 10.2174/1574886313666180807094654
240. Shankar BR. Quetiapine-induced leucopenia and thrombocytopenia. Psychosomatics. (2007) 48:530–31. doi: 10.1176/appi.psy.48.6.530
241. Arslan FC, Aykut DS, Ince C, Tiryaki A. Klinik Psikofarmakoloji Bülteni-bulletin of clinical psychopharmacology neutropenia and thrombocytopenia induced by quetiapine monotherapy: a case report and review of literature. Bull Clin Psychopharmacol. (2016) 26:319–23. doi: 10.5455/bcp.20151219072235
242. Tourjman V, Kouassi É, Koué M-È, Rocchetti M, Fortin-Fournier S, Fusar-Poli P, et al. Antipsychotics' effects on blood levels of cytokines in schizophrenia: a meta-analysis. Schizophr Res. (2013) 151:43–7. doi: 10.1016/j.schres.2013.10.011
243. Himmerich H, Schönherr J, Fulda S, Sheldrick AJ, Bauer K, Sack U. Impact of antipsychotics on cytokine production in-vitro. J Psychiatr Res. (2011) 45:1358–65. doi: 10.1016/j.jpsychires.2011.04.009
244. Zhu S, Shi R, Li V, Wang J, Zhang R, Tempier A, et al. Quetiapine attenuates glial activation and proinflammatory cytokines in APP/PS1 transgenic mice via inhibition of nuclear factor-κB pathway. Int J Neuropsychopharmacol. (2014) 18:pyu022. doi: 10.1093/ijnp/pyu022
245. Hillis J, O'Dwyer M, Gorman AM. Neurotrophins and B-cell malignancies. Cell Mol Life Sci. (2016) 73:41–56. doi: 10.1007/s00018-015-2046-4
246. Manti S, Brown P, Perez MK, Piedimonte G. The role of neurotrophins in inflammation and allergy. In: G. Litwack editor. Vitamins and Hormones. Cambridge MA: Elsevier. p. 313–41. doi: 10.1016/bs.vh.2016.10.010
247. Polyakova M, Stuke K, Schuemberg K, Mueller K, Schoenknecht P, Schroeter ML. BDNF as a biomarker for successful treatment of mood disorders: a systematic & quantitative meta-analysis. J Affect Disord. (2015) 174:432–40. doi: 10.1016/j.jad.2014.11.044
248. Fernandes BS, Gama CS, Maria Ceresér K, Yatham LN, Fries GR, Colpo G, et al. Brain-derived neurotrophic factor as a state-marker of mood episodes in bipolar disorders: a systematic review and meta-regression analysis. J Psychiatr Res. (2011) 45:995–1004. doi: 10.1016/j.jpsychires.2011.03.002
249. Murphy BP, Pang TY, Hannan AJ, Proffitt T-M, McConchie M, Kerr M, et al. Vascular endothelial growth factor and brain-derived neurotrophic factor in quetiapine treated first-episode psychosis. Schizophr Res Treat. (2014) 2014:1–10. doi: 10.1155/2014/719395
250. Kul D, Gumustas M, Uslu B, Ozkan SA. Electroanalytical characteristics of antipsychotic drug ziprasidone and its determination in pharmaceuticals and serum samples on solid electrodes. Talanta. (2010) 82:286–95. doi: 10.1016/j.talanta.2010.04.036
251. Gitlin M, Frye MA. Maintenance therapies in bipolar disorders. Bipolar Disord. (2012) 14(Suppl. 2):51–65. doi: 10.1111/j.1399-5618.2012.00992.x
252. Sacchetti E, Galluzzo A, Valsecchi P. Oral ziprasidone in the treatment of patients with bipolar disorders: a critical review. Expert Rev Clin Pharmacol. (2011) 4:163–79. doi: 10.1586/ecp.10.139
253. Warrington L, Lombardo I, Loebel A, Ice K. Ziprasidone for the treatment of acute manic or mixed episodes associated with bipolar disorder. CNS Drugs. (2007) 21:835–49. doi: 10.2165/00023210-200721100-00004
254. Schmidt AW, Lebel LA, Howard HR, Zorn SH. Ziprasidone: a novel antipsychotic agent with a unique human receptor binding profile. Eur J Pharmacol. (2001) 425:197–201. doi: 10.1016/s0014-2999(01)01188-8
255. Beedham C, Miceli JJ, Obach RS. Ziprasidone metabolism, aldehyde oxidase, and clinical implications. J Clin Psychopharmacol. (2003) 23:229–32. doi: 10.1097/01.jcp.0000084028.22282.f2
256. Ben Amor L. Antipsychotics in pediatric and adolescent patients: a review of comparative safety data. J Affect Disord. (2012) 138:S22–30. doi: 10.1016/j.jad.2012.02.030
257. Raza S, Haq F. Ziprasidone-induced galactorrhea in an adolescent female: a case report. Prim Care Companion J Clin Psychiatry. (2010) 12:PCC.09l00855. doi: 10.4088/PCC.09l00855gry
258. Kopecek M, Bares M, Mohr P. Ziprasidone-induced galactorrhea: a case report. Neuro Endocrinol Lett. (2005) 26:69–70. doi: 10.4088/PCC.09l00855gry
259. Meier A, Neumann AC, Jordan W, Huether G, Rodenbeck A, Rüther E, et al. Ziprasidone decreases cortisol excretion in healthy subjects. Br J Clin Pharmacol. (2005) 60:330–6. doi: 10.1111/j.1365-2125.2005.02431.x
260. Montgomery J. Ziprasidone-related agranulocytosis following olanzapine-induced neutropenia. Gen Hosp Psychiatry. (2006) 28:83–5. doi: 10.1016/j.genhosppsych.2005.08.005
261. Kefelioglu H, Atli Sekeroglu Z, Coşguner G, Kontaş Yedier S, Sekeroglu V. Ziprasidone induces cytotoxicity and genotoxicity in human peripheral lymphocytes. Drug Chem Toxicol. (2017) 40:425–31. doi: 10.1080/01480545.2016.1252920
262. Duarte T, Barbisan F, do Prado-Lima PAS, Azzolin VF, da Cruz Jung IE, Duarte MMMF, et al. Ziprasidone, a second-generation antipsychotic drug, triggers a macrophage inflammatory response in vitro. Cytokine. (2018) 106:101–7. doi: 10.1016/j.cyto.2017.10.017
263. Hamera L, Khishfe BF. Kounis syndrome and ziprasidone. Am J Emerg Med. (2017) 35:493–4. doi: 10.1016/j.ajem.2016.11.061
264. Ku HL, Su TP, Chou YH. Ziprasidone-associated pedal edema in the treatment of schizophrenia. Prog Neuro-Psychopharmacology Biol Psychiatry. (2006) 30:963–4. doi: 10.1016/j.pnpbp.2006.01.020
265. Akkaya C, Sarandol A, Aydogan K, Kirli S. Urticaria and angio-oedema due to ziprasidone. J Psychopharmacol. (2007) 21:550–2. doi: 10.1177/0269881106075273
266. Swainston Harrison T, Perry CM. Aripiprazole. Drugs. (2004) 64:1715–36. doi: 10.2165/00003495-200464150-00010
267. Li DJ, Tseng PT, Stubbs B, Chu CS, Chang HY, Vieta E, et al. Efficacy, safety and tolerability of aripiprazole in bipolar disorder: an updated systematic review and meta-analysis of randomized controlled trials. Prog Neuro-Psychopharmacology Biol Psychiatry. (2017) 79:289–301. doi: 10.1016/j.pnpbp.2017.06.023
268. Jing Y, Guo Z, Kalsekar I, Forbes RA, Hebden T, Thase ME. Dosing patterns of aripiprazole and quetiapine for adjunctive treatment of major depressive disorder (2006–2010). Int Clin Psychopharmacol. (2013) 28:87–90. doi: 10.1097/YIC.0b013e32835ce232
269. Blankenship K, Erickson CA, Stigler KA, Posey DJ, McDougle CJ. Aripiprazole for irritability associated with autistic disorder in children and adolescents aged 6–17 years. Ped Health. (2010) 4:375–81. doi: 10.2217/phe.10.45
270. Stip E, Tourjman V. Aripiprazole in schizophrenia and schizoaffective disorder: a review. Clin Ther. (2010) 32(Suppl. 1):S3–20. doi: 10.1016/j.clinthera.2010.01.021
271. Hirsch LE, Pringsheim T. Aripiprazole for autism spectrum disorders (ASD). Cochrane Database Syst Rev. (2016) 26:CD009043. doi: 10.1002/14651858.CD009043.pub3
272. Brunetti M, Di Tizio L, Dezi S, Pozzi G, Grandinetti P, Martinotti G. Aripiprazole, alcohol and substance abuse: a review. Eur Rev Med Pharmacol Sci. (2012) 16:1346–54.
273. Marcus RN, McQuade RD, Carson WH, Hennicken D, Fava M, Simon JS, et al. The efficacy and safety of aripiprazole as adjunctive therapy in major depressive disorder: a second multicenter, randomized, double-blind, placebo-controlled study. J Clin Psychopharmacol. (2008) 28:156–65. doi: 10.1097/JCP.0b013e31816774f9
274. McGavin JK, Goa KL. Aripiprazole. CNS Drugs. (2002) 16:779–86. doi: 10.2165/00023210-200216110-00008
275. Belmonte C, Ochoa D, Román M, Saiz-Rodríguez M, Wojnicz A, Gómez-Sánchez CI, et al. Influence of CYP2D6, CYP3A4, CYP3A5 and ABCB1 Polymorphisms on pharmacokinetics and safety of aripiprazole in healthy volunteers. Basic Clin Pharmacol Toxicol. (2018) 122:596–605. doi: 10.1111/bcpt.12960
276. Perez Rodriguez A, Tajima-Pozo K, Lewczuk A, Montañes-Rada F. Atypical antipsychotics and metabolic syndrome. Cardiovasc Endocrinol. (2015) 17:460–6. doi: 10.1097/XCE.0000000000000063
277. Pramyothin P, Khaodhiar L. Metabolic syndrome with the atypical antipsychotics. Curr Opin Endocrinol Diabetes Obes. (2010) 17:460–6. doi: 10.1097/MED.0b013e32833de61c
278. Newcomer JW. Second-generation (atypical) antipsychotics and metabolic effects: a comprehensive literature review. CNS Drugs. (2005) 19(Suppl 1):1–93. doi: 10.2165/00023210-200519010-00001
279. Keck PE, Marcus R, Tourkodimitris S, Ali M, Liebeskind A, Saha A, et al. A placebo-controlled, double-blind study of the efficacy and safety of aripiprazole in patients with acute bipolar mania. Am J Psychiatry. (2003) 160:1651–8. doi: 10.1176/appi.ajp.160.9.1651
280. Casey DE, Carson WH, Saha AR, Liebeskind A, Ali MW, Jody D, et al. Aripiprazole Study Group. Switching patients to aripiprazole from other antipsychotic agents: a multicenter randomized study. Psychopharmacology. (2003) 166:391–9. doi: 10.1007/s00213-002-1344-3
281. Kane JM, Meltzer HY, Carson WH, McQuade RD, Marcus RN, Sanchez R. Aripiprazole for treatment-resistant schizophrenia: results of a multicenter, randomized, double-blind, comparison study versus perphenazine. J Clin Psychiatry. (2007) 68:213–23. doi: 10.4088/JCP.v68n0206
282. Fleischhacker WW, Baker RA, Eramo A, Sanchez R, Tsai LF, Peters-Strickland T, et al. Effects of aripiprazole once-monthly on domains of personal and social performance: results from 2 multicenter, randomized, double-blind studies. Schizophr Res. (2014) 59:415–20. doi: 10.1016/j.schres.2014.09.019
283. Zhao J, Song X, Ai X, Gu X, Huang G, Li X, et al. Adjunctive aripiprazole treatment for risperidone-induced hyperprolactinemia: an 8-week randomized, open-label, comparative clinical trial. PLoS ONE. (2015) 10:e0139717. doi: 10.1371/journal.pone.0139717
284. Chen CK, Huang YS, Ree SC, Hsiao CC. Differential add-on effects of aripiprazole in resolving hyperprolactinemia induced by risperidone in comparison to benzamide antipsychotics. Prog Neuro-Psychopharmacology Biol Psychiatry. (2010) 134:1495–9. doi: 10.1016/j.pnpbp.2010.08.012
285. Kelly DL, Powell MM, Wehring HJ, Sayer MA, Kearns AM, Hackman AL, et al. Adjunct aripiprazole reduces prolactin and prolactin-related adverse effects in premenopausal women with psychosis: results from the DAAMSEL clinical trial. J Clin Psychopharmacol. (2018) 38:317–26. doi: 10.1097/JCP.0000000000000898
286. Berardis D, Fornaro M, Serroni N, Marini S, Piersanti M, Cavuto M, et al. Treatment of antipsychotic-induced hyperprolactinemia: an update on the role of the dopaminergic receptors D2 partial agonist aripiprazole. Recent Pat Endocr Metab Immune Drug Discov. (2014) 8:30–7. doi: 10.2174/1872214807666131229125700
287. Fujioi J, Iwamoto K, Banno M, Kikuchi T, Aleksic B, Ozaki N. Effect of adjunctive aripiprazole on sexual dysfunction in schizophrenia: a preliminary open-label study. Pharmacopsychiatry. (2017) 50:74–8. doi: 10.1055/s-0042-116323
288. Joseph SP. Aripiprazole-induced hyperprolactinemia in a young female with delusional disorder. Indian J Psychol Med. (2016) 38:260–2. doi: 10.4103/0253-7176.183082
289. Saraf G, Behere RV, Venkatasubramanian G, Rao NP, Varambally S, Gangadhar BN. Hyperprolactinemia with aripiprazole: understanding the paradox. Am J Ther. (2014) 21:e80–1. doi: 10.1097/MJT.0b013e3182456de7
290. Luo T, sheng Liu Q, jian Yang Y, Wei B. Aripiprazole for the treatment of duloxetine-induced hyperprolactinemia: a case report. J Affect Disord. (2019) 250:330–2. doi: 10.1016/j.jad.2019.03.006
291. Stroup TS, McEvoy JP, Ring KD, Hamer RH, LaVange LM, Swartz MS, et al. A randomized trial examining the effectiveness of switching from olanzapine, quetiapine, or risperidone to aripiprazole to reduce metabolic risk: comparison of antipsychotics for metabolic problems (CAMP). Am J Psychiatry. (2011) 168:947–56. doi: 10.1176/appi.ajp.2011.10111609
292. Stroup TS, Byerly MJ, Nasrallah HA, Ray N, Khan AY, Lamberti JS, et al. Effects of switching from olanzapine, quetiapine, and risperidone to aripiprazole on 10-year coronary heart disease risk and metabolic syndrome status: results from a randomized controlled trial. Schizophr Res. (2013) 146:190–5. doi: 10.1016/j.schres.2013.01.013
293. Bak M, Fransen A, Janssen J, Van Os J, Drukker M. Almost all antipsychotics result in weight gain: a meta-analysis. PLoS ONE. (2014) 9:e94112. doi: 10.1371/journal.pone.0094112
294. Parabiaghi A, Tettamanti M, D'Avanzo B, Barbato A, GiSAS study group. Metabolic syndrome and drug discontinuation in schizophrenia: a randomized trial comparing aripiprazole olanzapine and haloperidol. Acta Psychiatr Scand. (2016) 133:63–75. doi: 10.1111/acps.12468
295. Kasteng F, Eriksson J, Sennfält K, Lindgren P. Metabolic effects and cost-effectiveness of aripiprazole versus olanzapine in schizophrenia and bipolar disorder. Acta Psychiatr Scand. (2011) 124:214–25. doi: 10.1111/j.1600-0447.2011.01716.x
296. Horska K, Ruda-Kucerova J, Drazanova E, Karpisek M, Demlova R, Kasparek T, et al. Aripiprazole-induced adverse metabolic alterations in polyI: C neurodevelopmental model of schizophrenia in rats. Neuropharmacology. (2017) 123:148–58. doi: 10.1016/j.neuropharm.2017.06.003
297. Felin T, Naveed S, Chaudhary AM. Aripiprazole-induced neutropenia: case report and literature review. J Psychosoc Nurs Ment Health Serv. (2018) 56:21–4. doi: 10.3928/02793695-20180419-02
298. Lim MH, Park J-I, Park TW. A case with neutropenia related with the use of various atypical antipsychotics. Psychiatry Investig. (2013) 10:428. doi: 10.4306/pi.2013.10.4.428
299. Upthegrove R, Manzanares-Teson N, Barnes NM. Cytokine function in medication-naive first episode psychosis: a systematic review and meta-analysis. Schizophr Res. (2014) 155:101–8. doi: 10.1016/j.schres.2014.03.005
300. Sobis J, Rykaczewska-Czerwinska M, wietochowska E, Gorczyca P. Therapeutic effect of aripiprazole in chronic schizophrenia is accompanied by anti-inflammatory activity. Pharmacol Rep. (2015) 67:353–9. doi: 10.1016/j.pharep.2014.09.007
301. Yoo S, Kim MY, Cho JY. Syk and Src-targeted anti-inflammatory activity of aripiprazole, an atypical antipsychotic. Biochem Pharmacol. (2018) 148:1–12. doi: 10.1016/j.bcp.2017.12.006
302. Kato T, Mizoguchi Y, Monji A, Horikawa H, Suzuki SO, Seki Y, et al. Inhibitory effects of aripiprazole on interferon-γ-induced microglial activation via intracellular Ca2+ regulation in vitro. J Neurochem. (2008) 106:815–25. doi: 10.1111/j.1471-4159.2008.05435.x
303. Zargar S, Al-Majed ARA, Wani TA. Potentiating and synergistic effect of grapefruit juice on the antioxidant and anti-inflammatory activity of aripiprazole against hydrogen peroxide induced oxidative stress in mice. BMC Complement Altern Med. (2018) 18:106. doi: 10.1186/s12906-018-2169-x
304. Baek KS, Ahn S, Lee J, Kim JH, Kim HG, Kim E, et al. Immunotoxicological effects of a ripiprazole: in vivo and in vitro studies. Korean J Physiol Pharmacol. (2015) 19:365–72. doi: 10.4196/kjpp.2015.19.4.365
305. Citrome L. Paliperidone: Quo vadis? Int J Clin Pract. (2007) 61:653–62. doi: 10.1111/j.1742-1241.2007.01321.x
306. Greenberg WM, Citrome L. Paliperidone palmitate for schizoaffective disorder: a review of the clinical evidence. Neurol Ther. (2015) 4:81–91. doi: 10.1007/s40120-015-0030-4
307. INVEGA SUSTENNA ® Paliperidone Palmitate Product Information. Available online at: https://www.janssen.com/australia/sites/www_janssen_com_australia/files/prod_files/live/invega_sustenna_pi.pdf (accessed October 2, 2019)
308. Nussbaum AM, Stroup TS. Paliperidone for treatment of schizophrenia. Schizophr Bull. (2008) 34:419–22. doi: 10.1093/schbul/sbn015
309. Marino J, English C, Caballero J, Harrington C. The role of paliperidone extended release for the treatment of bipolar disorder. Neuropsychiatr Dis Treat. (2012) 2012:181–9. doi: 10.2147/NDT.S20675
310. Palomares N, Montes A, Díaz-Marsá M, Carrasco JL. Effectiveness of long-acting paliperidone palmitate in borderline personality disorder. Int Clin Psychopharmacol. (2015) 30:338–41. doi: 10.1097/YIC.0000000000000095
311. van Oosterom N, Theodoros T. Paliperidone long-acting injections in huntington's disease for motor and behavioural disturbances. Clin Drug Investig. (2019) 39:407–10. doi: 10.1007/s40261-019-00759-8
312. Naguy A, Adel T, Almazeedi I. Paliperidone use in child psychiatry: evidence or diffidence? Pharmacology. (2019) 104:67–70. doi: 10.1159/000500629
313. Hu S, Yao M, Peterson BS, Xu D, Hu J, Tang J, et al. A randomized, 12-week study of the effects of extended-release paliperidone (paliperidone ER) and olanzapine on metabolic profile, weight, insulin resistance, and β-cell function in schizophrenic patients. Psychopharmacology. (2013) 230:3–13. doi: 10.1007/s00213-013-3073-1
314. Druyts E, Zoratti MJ, Toor K, Wu P, Kanji S, Rabheru K, et al. Prolactin-related adverse events and change in prolactin levels in pediatric patients given antipsychotics for schizophrenia and schizophrenia spectrum disorders: a systematic review. BMC Pediatr. (2016) 16:1–14. doi: 10.1186/s12887-016-0710-y
315. Skopek M, Manoj P. Hyperprolactinaemia during treatment with paliperidone. Australas Psychiatry. (2010) 18:261–3. doi: 10.3109/10398561003686763
316. Montalvo I, Ortega L, López X, Solé M, Monseny R, Franch J, et al. Changes in prolactin levels and sexual function in young psychotic patients after switching from long-acting injectable risperidone to paliperidone palmitate. Int Clin Psychopharmacol. (2013) 28:46–9. doi: 10.1097/YIC.0b013e32835ba832
317. Nakamura M, Nagamine T, Sato G, Besho K. Prolactin levels after switching to paliperidone palmitate in patients with schizophrenia. Innov Clin Neurosci. (2016) 13:28–30.
318. Potkin SG, Loze JY, Forray C, Baker RA, Sapin C, Peters-Strickland T, et al. Reduced sexual dysfunction with aripiprazole once-monthly versus paliperidone palmitate: results from QUALIFY. Int Clin Psychopharmacol. (2017) 32:147–54. doi: 10.1097/YIC.0000000000000168
319. Gopal S, Lane R, Nuamah I, Copenhaver M, Singh J, Hough D, et al. Evaluation of potentially prolactin-related adverse events and sexual maturation in adolescents with schizophrenia treated with paliperidone extended-release (ER) for 2 years: a post hoc analysis of an open-label multicenter study. CNS Drugs. (2017) 31:797–808. doi: 10.1007/s40263-017-0437-9
320. Ngamsamut N, Hongkaew Y, Vanwong N, Srisawasdi P, Puangpetch A, Chamkrachangpada B, et al. 9-Hydroxyrisperidone-induced hyperprolactinaemia in thai children and adolescents with autism spectrum disorder. Basic Clin Pharmacol Toxicol. (2016) 119:267–72. doi: 10.1111/bcpt.12570
321. Sliwa JK, Fu DJ, Bossie CA, Turkoz I, Alphs L. Body mass index and metabolic parameters in patients with schizophrenia during long-term treatment with paliperidone palmitate. BMC Psychiatry. (2014) 2014:1–11. doi: 10.1186/1471-244X-14-52
322. Omi T, Riku K, Fukumoto M, Kanai K, Omura Y, Takada H, et al. Paliperidone induced hypoglycemia by increasing insulin secretion. Case Rep Psychiatry. (2016) 2016:1–3. doi: 10.1155/2016/1805414
323. Kim JN, Lee BC, Choi IG, Jon DI, Jung MH. Paliperidone-induced leukopenia and neutropenia: a case report. Prog Neuro-Psychopharmacology Biol Psychiatry. (2011) 35:284–5. doi: 10.1016/j.pnpbp.2010.09.018
324. Wakuda T, Suzuki A, Hasegawa M, Ichikawa D, Yamasue H. Acute agranulocytosis when switching from risperidone to paliperidone. Aust N Z J Psychiatry. (2019) 53:586–7. doi: 10.1177/0004867418821441
325. Matsuura H, Kimoto S, Harada I, Naemura S, Yamamuro K, Kishimoto T. Lithium carbonate as a treatment for paliperidone extended-release-induced leukopenia and neutropenia in a patient with schizoaffective disorder; a case report. BMC Psychiatry. (2016) 16:1–3. doi: 10.1186/s12888-016-0874-x
326. Flanagan RJ, Dunk L. Haematological toxicity of drugs used in psychiatry. Hum Psychopharmacol. (2008) 23(Suppl 1):27–41. doi: 10.1002/hup.917
327. Wu RQ, Lin CG, Zhang W, Lin XD, Chen XS, Chen C, et al. Effects of risperidone and paliperidone on brain-derived neurotrophic factor and N400 in first-episode schizophrenia. Chin Med J. (2018) 131:2297–301. doi: 10.4103/0366-6999.241802
328. Chung YC, Cui Y, Sumiyoshi T, Kim MG, Lee KH. Associations of fatty acids with cognition, psychopathology, and brain-derived neurotrophic factor levels in patients with first-episode schizophrenia and related disorders treated with paliperidone extended release. J Psychopharmacol. (2017) 31:1556–63. doi: 10.1177/0269881117731169
329. Mas S, Gassó P, Parellada E, Bernardo M, Lafuente A. Network analysis of gene expression in peripheral blood identifies mTOR and NF-κB pathways involved in antipsychotic-induced extrapyramidal symptoms. Pharmacogenomics J. (2015) 15:452–60. doi: 10.1038/tpj.2014.84
330. Schmidt AJ, Krieg JC, Clement HW, Hemmeter UM, Schulz E, Vedder H, et al. Effects of quetiapine, risperidone, 9-hydroxyrisperidone and ziprasidone on the survival of human neuronal and immune cells in vitro. J Psychopharmacol. (2010) 24:349–54. doi: 10.1177/0269881108096506
331. Leucht S, Cipriani A, Spineli L, Mavridis D, Örey D, Richter F, et al. Comparative efficacy and tolerability of 15 antipsychotic drugs in schizophrenia: a multiple-treatments meta-analysis. Lancet. (2013) 382:951–62. doi: 10.1016/S0140-6736(13)60733-3
332. Vieta E, Montes JM. A review of asenapine in the treatment of bipolar disorder. Clin Drug Investig. (2018) 38:87–99. doi: 10.1007/s40261-017-0592-2
333. Anugu RR, Mainkar PS, Sridhar B, Chandrasekhar S. The Ireland-claisen rearrangement strategy towards the synthesis of the schizophrenia drug, (+)-asenapine. Org Biomol Chem. (2016) 14:1332–7. doi: 10.1039/c5ob02047f
334. van de Wetering-Krebbers SFM, Jacobs PL, Kemperman GJ, Spaans E, Peeters PAM, Delbressine LPC, et al. Metabolism and excretion of asenapine in healthy male subjects. Drug Metab Dispos. (2011) 39:580–90. doi: 10.1124/dmd.110.036715
335. Hounsou C, Baehr C, Gasparik V, Alili D, Belhocine A, Rodriguez T, et al. From the promiscuous asenapine to potent fluorescent ligands acting at a series of aminergic G-protein-coupled receptors. J Med Chem. (2018) 61:174–88. doi: 10.1021/acs.jmedchem.7b01220
336. Delcourte S, Abrial E, Etiévant A, Rovera R, Arnt J, Didriksen M, et al. Asenapine modulates mood-related behaviors and 5-HT1A/7 receptors-mediated neurotransmission. CNS Neurosci Ther. (2017) 23:518–25. doi: 10.1111/cns.12698
337. Meltzer HY, Massey BW. The role of serotonin receptors in the action of atypical antipsychotic drugs. Curr Opin Pharmacol. (2011) 11:59–67. doi: 10.1016/j.coph.2011.02.007
338. Bobo WV. Asenapine, iloperidone and lurasidone: critical appraisal of the most recently approved pharmacotherapies for schizophrenia in adults. Expert Rev Clin Pharmacol. (2013) 6:61–91. doi: 10.1586/ecp.12.70
339. Samalin L, Charpeaud T, Llorca PM. Asenapine in bipolar I disorder: evidence and place in patient management. Ther Adv Chronic Dis. (2013) 4:5–14. doi: 10.1177/2040622312468933
340. Peuskens J, Pani L, Detraux J, De Hert M. The effects of novel and newly approved antipsychotics on serum prolactin levels: a comprehensive review. CNS Drugs. (2014) 28:421–53. doi: 10.1007/s40263-014-0157-3
341. Rahman T, Clevenger CV, Kaklamani V, Lauriello J, Campbell A, Malwitz K, et al. Antipsychotic treatment in breast cancer patients. Am J Psychiatry. (2014) 171:616–21. doi: 10.1176/appi.ajp.2013.13050650
342. Citrome L. Asenapine review, part II: clinical efficacy, safety and tolerability. Expert Opin Drug Saf. (2014) 13:803–30. doi: 10.1517/14740338.2014.908183
343. Citrome L. Role of sublingual asenapine in treatment of schizophrenia. Neuropsychiatr Dis Treat. (2011) 7:325–39. doi: 10.2147/NDT.S16077
344. Findling RL, Landbloom RL, Szegedi A, Koppenhaver J, Braat S, Zhu Q, et al. Asenapine for the acute treatment of pediatric manic or mixed episode of bipolar i disorder. J Am Acad Child Adolesc Psychiatry. (2015) 54:1032–41. doi: 10.1016/j.jaac.2015.09.007
345. Mommert S, Ratz L, Stark H, Gutzmer R, Werfel T. The histamine H4 receptor modulates the differentiation process of human monocyte-derived M1 macrophages and the release of CCL4/MIP-1β from fully differentiated M1 macrophages. Inflamm Res. (2018) 67:503–13. doi: 10.1007/s00011-018-1140-0
346. Anstead MI, Hunt TA, Carlson SL, Burki NK. Variability of peripheral blood lymphocyte beta-2-adrenergic receptor density in humans. Am J Respir Crit Care Med. (1998) 157:990–2. doi: 10.1164/ajrccm.157.3.9704071
347. Makdisi J, Amin B, Friedman A. Pityriasis rosea-like drug reaction to asenapine. J Drugs Dermatol. (2013) 12:1050–51.
348. Klemettilä JP, Kampman O, Seppälä N, Viikki M, Hämäläinen M, Moilanen E, et al. Resistin as an inflammatorymarker in patients with schizophrenia treated with clozapine. Nord J Psychiatry. (2017) 71:89–95. doi: 10.1080/08039488.2016.1230649
349. Skonieczna-Zydecka K, Łoniewski I, Misera A, Stachowska E, Maciejewska D, Marlicz W, et al. Second-generation antipsychotics and metabolism alterations: a systematic review of the role of the gut microbiome. Psychopharmacology. (2019) 236:1491–512. doi: 10.1007/s00213-018-5102-6
350. Kao AC-C, Spitzer S, Anthony DC, Lennox B, Burnet PWJ. Prebiotic attenuation of olanzapine-induced weight gain in rats: analysis of central and peripheral biomarkers and gut microbiota. Transl Psychiatry. (2018) 8:66. doi: 10.1038/s41398-018-0116-8
351. Yuan X, Zhang P, Wang Y, Liu Y, Li X, Kumar BU, et al. Changes in metabolism and microbiota after 24-week risperidone treatment in drug naïve, normal weight patients with first episode schizophrenia. Schizophr Res. (2018) 201:299–306. doi: 10.1016/j.schres.2018.05.017
352. Bahra SM, Weidemann BJ, Castro AN, Walsh JW, deLeon O, Burnett CML, et al. Risperidone-induced weight gain is mediated through shifts in the gut microbiome and suppression of energy expenditure. EBioMedicine. (2015) 2:1725–34. doi: 10.1016/j.ebiom.2015.10.018
353. Bahr SM, Tyler BC, Wooldridge N, Butcher BD, Burns TL, Teesch LM, et al. Use of the second-generation antipsychotic, risperidone, and secondary weight gain are associated with an altered gut microbiota in children. Transl Psychiatry. (2015) 5:e652. doi: 10.1038/tp.2015.135
354. Davey KJ, Cotter PD, O'Sullivan O, Crispie F, Dinan TG, Cryan JF, et al. Antipsychotics and the gut microbiome: olanzapine-induced metabolic dysfunction is attenuated by antibiotic administration in the rat. Transl Psychiatry. (2013) 3:e309. doi: 10.1038/tp.2013.83
355. Cussotto S, Strain CR, Fouhy F, Strain RG, Peterson VL, Clarke G, et al. Differential effects of psychotropic drugs on microbiome composition and gastrointestinal function. Psychopharmacology. (2019) 236:1671–85. doi: 10.1007/s00213-018-5006-5
Keywords: atypical antipsychotics (AAP), peripheral effects, inflammatory response, endocrine response, microbiome
Citation: Alvarez-Herrera S, Escamilla R, Medina-Contreras O, Saracco R, Flores Y, Hurtado-Alvarado G, Maldonado-García JL, Becerril-Villanueva E, Pérez-Sánchez G and Pavón L (2020) Immunoendocrine Peripheral Effects Induced by Atypical Antipsychotics. Front. Endocrinol. 11:195. doi: 10.3389/fendo.2020.00195
Received: 14 October 2019; Accepted: 18 March 2020;
Published: 21 April 2020.
Edited by:
Ana Rosa Pérez, National Council for Scientific and Technical Research (CONICET), ArgentinaReviewed by:
Antonio Torsello, University of Milano-Bicocca, ItalyCopyright © 2020 Alvarez-Herrera, Escamilla, Medina-Contreras, Saracco, Flores, Hurtado-Alvarado, Maldonado-García, Becerril-Villanueva, Pérez-Sánchez and Pavón. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lenin Pavón, bGt1cmlha2lAaW1wLmVkdS5teA==
†ORCID: Samantha Alvarez-Herrera orcid.org/0000-0001-9747-885X
Raúl Escamilla orcid.org/0000-0002-6772-9731
Oscar Medina-Contreras orcid.org/0000-0002-4432-7780
Ricardo Saracco orcid.org/0000-0002-0004-1318
Yvonne Flores orcid.org/0000-0002-4508-8734
Gabriela Hurtado-Alvarado orcid.org/0000-0002-6137-663X
José Luis Maldonado-García orcid.org/0000-0003-2694-1290
Gilberto Pérez-Sánchez orcid.org/0000-0003-3878-0631
Enrique Becerril-Villanueva orcid.org/0000-0001-7210-9775
Lenin Pavón orcid.org/0000-0002-6067-6868
‡These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.