 Juan Ignacio Bertucci1
Juan Ignacio Bertucci1 Ayelén Melisa Blanco1,2
Ayelén Melisa Blanco1,2 Lakshminarasimhan Sundarrajan1Jithine Jayakumar Rajeswari1
Lakshminarasimhan Sundarrajan1Jithine Jayakumar Rajeswari1 Cristina Velasco1,2
Cristina Velasco1,2 Suraj Unniappan1*
Suraj Unniappan1*- 1Laboratory of Integrative Neuroendocrinology, Department of Veterinary Biomedical Sciences, University of Saskatchewan, Saskatoon, SK, Canada
- 2Laboratorio de Fisioloxìa Animal, Departamento de Bioloxía Funcional e Ciencias da Saúde, Facultade de Bioloxía and Centro de Investigación Mariña, Universidade de Vigo, Vigo, Spain
Endocrine factors regulate food intake and growth, two interlinked physiological processes critical for the proper development of organisms. Somatic growth is mainly regulated by growth hormone (GH) and insulin-like growth factors I and II (IGF-I and IGF-II) that act on target tissues, including muscle, and bones. Peptidyl hormones produced from the brain and peripheral tissues regulate feeding to meet metabolic demands. The GH-IGF system and hormones regulating appetite are regulated by both internal (indicating the metabolic status of the organism) and external (environmental) signals. Among the external signals, the most notable are diet availability and diet composition. Macronutrients and micronutrients act on several hormone-producing tissues to regulate the synthesis and secretion of appetite-regulating hormones and hormones of the GH-IGF system, eventually modulating growth and food intake. A comprehensive understanding of how nutrients regulate hormones is essential to design diet formulations that better modulate endogenous factors for the benefit of aquaculture to increase yield. This review will discuss the current knowledge on nutritional regulation of hormones modulating growth and food intake in fish.
Introduction
Physiological processes in fish, as well as in other vertebrates, are subject to complex regulatory mechanisms that act in response to both internal and external signals (1–4). Signals provided by the environment, along with internal cues are sensed and centrally integrated, providing information about the metabolic status. This enables fish to determine whether conditions are ripe to feed, grow, reproduce and save energy (4, 5). Among the external signals that can influence feeding behavior and growth in fish, one of the most important is food (6, 7). Both food availability and food composition exert a critical control of these processes, primarily by acting on the hormones in charge of their endocrine control. The main aim of this review is to summarize the recent advances on the role of feeding and fasting, as well as of dietary macro- and micronutrients, on the regulation of appetite- and growth-regulating hormones in fish. A better understanding of the effects of feeding status and diet composition on the expression and release of those hormones could be beneficial to determine the effects of a specific diet or feeding regime on fish health, growth, and development, which could be crucial in aquaculture. This review will also aim to identify gaps in knowledge and directions for future research regarding this important topic.
Nutrients and Their Importance in Fish
Nutrients are organic compounds involved in biochemical reactions that produce energy and are constituents of cellular biomass (8). They are divided into two broad groups: macronutrients and micronutrients. Macronutrients are classified into carbohydrates (CHO), proteins and lipids, and are needed in relatively large amounts since they are the primary source to generate the energy organisms require to survive, grow, and reproduce. These nutrients can be stored within the body for later use, or be utilized, leading to the somatic growth of the animal (9). The micronutrients comprise of vitamins and minerals and are needed in smaller quantity, although they have several critical roles in cellular processes. The nutritional requirements regarding the composition and proportion of different nutrients present in the diet vary among species and within species, is determined by various extrinsic and intrinsic factors such as environmental conditions, stage of life cycle, sex, and reproductive state (10). The importance and main roles of each nutrient in fish metabolism are summarized in Table 1.
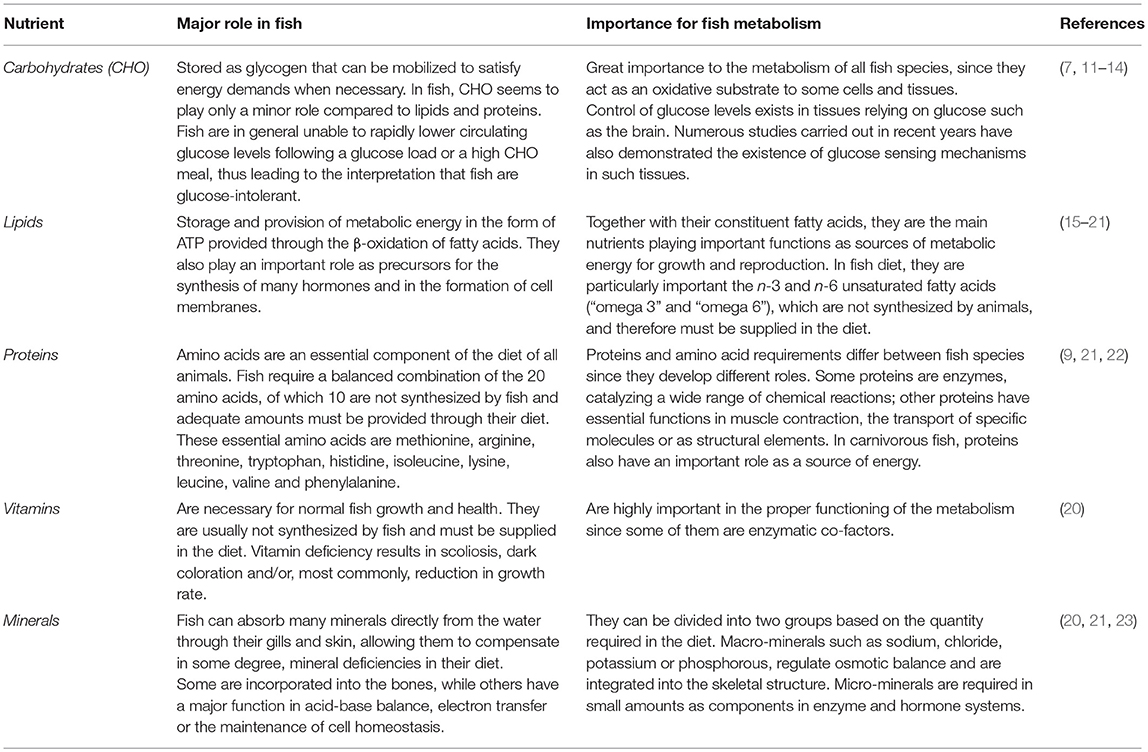
Table 1. The role of principal macronutrients in fish metabolism.
Organisms coordinate their growth and development with nutrient level fluctuations in their environment, and therefore they must be able to sense their internal and external nutrient levels (24). In mammals, sensing mechanisms regulate specific processes such as food intake, hormone secretion, and energy expenditure to maintain energy homeostasis (25–27). The sensing of nutrients may occur directly or indirectly. In the first case, the nutrient molecule binds to its sensor. In the second case, nutrient abundance is detected through a related molecule (8). These detection processes occur in both central and peripheral tissues in fish. At the central level, the brain integrates metabolic information related to nutrient availability, satiety/hunger signals and hormones related to adiposity. As a result of such signal integration, a response is generated in peripheral tissues aiming to modulate the metabolism (28, 29). At the periphery, metabolic regulation by the sensing systems occurs directly or indirectly through the action of endocrine effectors (28).
Hormones Regulating Food Intake
The regulation of food intake involves the integration of exogenous and endogenous factors to supply the energy necessary to support biological processes. Such regulation is achieved by the endocrine system, which secretes hormones and regulates the activity of the cells by transferring information between the organs. The major organs that secrete hormones involved in regulating appetite are the brain (hypothalamus) and gastrointestinal tissues. The following section will provide a brief description of the main hormones with a critical role in feeding regulation.
Brain Hormones/Neuropeptides
Neuropeptide Y (NPY)
Neuropeptide Y, a 36 kDa amino acid protein, belongs to the NPY family of peptides, which also includes the pancreatic polypeptide (PNP) and peptide tyrosine (PYY). All these peptides share a common three-dimensional structure composed of polyproline coil and an amphipathic helix. The structure of the neuropeptide Y family is tightened by hydrophobic interactions between prolines and the helix (30). NPY is mainly secreted by the neurosecretory cells in the hypothalamus and is abundantly expressed in the brain, pituitary, spleen, gastrointestinal tract, kidney, testis and smooth muscles (31, 32). In teleosts, NPY has been shown to play an essential role in stimulating feeding [see Volkoff (33) for review].
Agouti Related Protein (AgRP)
AgRP is a 128 amino acid neuropeptide released by the NPY/AgRP neurons, and is an endogenous antagonist of melanocortin receptors 3 and 4 (MC3R and MC4R) (34). It is mainly expressed in the brain, but it is also found in several peripheral tissues, including ovary, muscle and ventral skin (34). AgRP receptor is highly localized at the site of the paraventricular nucleus, the dorsal motor nucleus of the vagus nerve and also in the raphe nucleus, all areas that are highly involved in energy homeostasis. AgRP acts as an orexigenic factor in fish, by antagonizing the activity of MC4R (33).
Proopiomelanocortin (POMC)
POMC is a 267 amino acid peptide secreted by the hypothalamic neurons located in the arcuate nucleus, as well as the corticotropic cells of the anterior pituitary, the melanotropic cells of the pars intermedia and skin melanocytes (35). In vertebrates, precursors of POMC has three domains, namely N-terminal pro-γ-melanocyte stimulating hormone (MSH), adrenocorticotropic hormone (ACTH) and C-terminal β-lipotropin, which are cleaved by the action of prohormone convertases. The most important of these derivatives are α-MSH, which plays a vital role in suppressing feeding by acting as an agonist at the anorectic MC4R (33), and ACTH, which regulates the secretion of glucocorticoids from the adrenal glands (36, 37).
Cocaine- and Amphetamine-Regulated Transcript (CART)
CART was isolated from rat striatum upon injection of cocaine and amphetamine, two psychomotor stimulants (38). In goldfish, two forms of CART precursor exist, namely, CARTI that encodes a 117 amino acid pro-CART, and CART-II which encodes a 120 amino acid pro-CART (39). Both CART precursors have been reported to be abundantly expressed in brain, pituitary and also in other peripheral tissues such as eye, interrenal tissues, and gonads in goldfish (39). CART exerts multiple physiological functions in fish, including the inhibition of appetite (39), regulation of the stress response (40) and energy balance (41).
Orexins
Orexins/hypocretins consist of two orexins, orexin-A, and orexin-B, both cleaved from the same precursor, prepro-orexin (42, 43). In fish, both the prepro-orexin RNA and the peptides A and B have been shown to be abundant in the hypothalamus (44), as well as in the gastrointestinal tract (33). Two heptahelical G-protein coupled receptors are known to mediate orexin functions. Orexins have been reported to have a significant role in increasing feeding behavior and locomotor activity (33), and they were also implicated in the regulation of sleep, energy homeostasis and circadian cycle (45).
Melanin-Concentrating Hormone (MCH)
Melanin-concentrating hormone (MCH), a 17 amino acid cyclic peptide, was initially isolated from the brain of chum salmon (46). Two genes, MCH1 and MCH2, have been identified in zebrafish and pufferfish (47). The MCH receptor was identified as the GPCR SLC-1, later termed as MCH-R1. MCH-R1 couples to different G-proteins and plays an essential role in activating different signaling pathways. The MCH-R1 is preferentially expressed in the brain, particularly in the hypothalamus, areas of the cortex, arcuate and ventromedial nuclei, and olfactory lobes. MCH mainly acts on the melanophores regulating the color change and also lightens the skin in fish. Besides this, MCH seems to have a role in the regulation of feeding, although such a function is still unclear in fish [see Volkoff (33) for review].
Nesfatin-1
Nesfatin-1 is an 82 amino acid peptide originally isolated from the rat hypothalamus and is encoded in the nucleobindin-2 (NUCB2) gene (48). It is proposed that NUCB2 is cleaved by the prohormone convertases into three different peptides, namely nesfatin-1 (82 amino acids), nesfatin-2 (85–163 amino acids) and nesfatin-3 (166–396 amino acids), respectively. Of all these peptides, nesfatin-1 has been shown to have biological activity. The 30-amino acid mid-segment of nesfatin-1 is considered as the bioactive core and has been shown to affect appetite, the hypothalamus-pituitary-ovarian axis, and to modulate intracellular Ca2+ signaling in mammals (49). Among non-mammals, nesfatin-1 has been studied in various fish species, including goldfish (50), Ya-fish (51), and rainbow trout (52). In goldfish, nesfatin-1-like immunoreactivity has been found in the hypothalamus, particularly in the nucleus lateralis tuberis (NLT) (50). Exogenous administration of nesfatin-1 has been shown to cause anorectic actions in goldfish (50).
Hormones Primarily Arising From Peripheral Tissues
Ghrelin
Ghrelin has been identified in numerous fish species. It consists of 28 amino acids in mammals and has 12 to 25 amino acids in fish depending on the species. The gene encoding the protein was identified within the chromosome 3 and consists of four exons. Ghrelin exerts its physiological functions by binding to the growth hormone secretagogue receptor-1a (GHS-R1a/ghrelin receptor) (53). This peptide is mainly released in the stomach (or its equivalent in stomachless species), although other tissues have been shown to synthesize the hormone, particularly the hypothalamus (54). The central role described for ghrelin is its potent orexigenic role (33), but there are other well-known physiological roles for this hormone, including the regulation of GH release from the pituitary, a role in energy balance regulation, and cardiovascular effects, among others (55–57).
Cholecystokinin (CCK)
CCK is a peptide characterized by a C-terminal tetrapeptide sequence. The structure of pro-CCK consists of three sulfated tyrosine residues, which play a crucial role in the activation of CCK receptors (58). CCK binds to two receptor subtypes, CCK-A receptor (CCK1) and CCK-B receptor (CCK2), which are primarily localized at the site of the gastrointestinal tract and the brain (59). In fish, CCK and its cleavage sites suggest that the precursor protein (pro-CCK) is processed into octapeptides and are fully sulfated. CCK plays an essential role in the regulation of feeding, influences digestion and activates satiety signals (60). In goldfish, acute administration of CCK resulted in the suppression of food intake (61), likely by acting on NPY and orexin-A (62).
Peptide YY
Peptide YY, a member of the Y family of peptides, is a 36 amino acid gut-brain hormone that is known to have anorectic actions in goldfish (63). PYY is released from the endocrine cells of ileum and colon (64), and binds to the NPY receptor 1 which is abundantly expressed in the brain and gut of fish. Similar to NPY, PYY plays an essential role in signaling between the enteric nervous system and central nervous system in fish (31, 65).
Glucagon-Like-Peptide-1 (GLP-1)
GLP-1, another anorexigenic intestinal peptide, belongs to the family of glucagon-like peptides encoded in the preproglucagon gene. In fish, the pancreas synthesizes glucagon and GLP-1, while the intestine secretes oxyntomodulin (66), and all of these peptides are processed from the proglucagon in the nervous system and intestine. GLP mRNAs have been identified in several teleosts, and its receptor (GLP-1R) has been successfully cloned in zebrafish and goldfish (67). Apart from reducing food intake, GLP-1 has been involved in gastric emptying and plays an important role in regulating liver glycogenolysis and gluconeogenesis (66).
Leptin
Leptin is a 16 kDa protein encoded in chromosome 7. While mammalian leptin is mainly produced by the adipose tissue, the liver appears to be the main leptin production site in fish (68). The structure of leptin resembles that of growth hormone, belonging to the family of tetrahelical cytokines. In fish, this hormone has been shown to affect adipogenesis (57), and to increase lipolysis while reducing lipogenesis in liver (69). Besides this, leptin decreases food intake in several fish species (33), likely by stimulating the anorexigenic neuropeptides POMC and CART (70).
Hormonal Regulators of Growth
Regulation of growth in fish, as well as in most of the vertebrates, is coordinated by the GH-IGF system (71). A summary of each component of the GH-IGF system as well as their principal function related to growth is presented in this section.
Growth Hormone
Growth hormone (GH) is an endocrine regulator of many physiological processes in vertebrates. In fish, GH is involved in almost all physiological processes including osmotic balance, lipid, protein and CHO metabolism, reproduction and growth. Moreover, studies have indicated that GH also affects behavioral aspects, such as appetite (72) and foraging (73) in rainbow trout and transgenic Atlantic salmon, respectively. In fish, GH is released from the adenohypophysis in response to hypothalamic signals, and exerts its effects on target tissues (74).
Growth Hormone Receptors
In fish cells, GH binds to its receptors GHR-I and GHR-II (growth hormone receptor I and II, respectively) to exert its biological actions (75). GH receptors (GHRs) are widely distributed in tissues, but the primary expression is in the liver (or hepatopancreas). In that tissue, the important response to GH binding its receptors is the release of the insulin-like growth factor I (IGF-I). In other tissues, GHRs also mediate the growth-promoting effects of GH, although the liver is still the place in which GHRs have a significant role in the somatic growth regulation.
Insulin-Like Growth Factors
As GHRs, IGF-I and IGF-II are expressed in several tissues, but the main expression is in fish liver. Both factors play a key role in the promotion of cellular proliferation and differentiation in vertebrates (76–78). These and other biological functions of IGFs are mediated by binding to specific transmembrane receptors, present in fish as well as in mammals (79). Apart from growth, IGF-I has also been associated with fish metabolism, development, reproduction and osmoregulation in seawater (74). The IGF-II mRNA has been detected in the liver as well as in the brain, heart, kidney, gills, gastrointestinal tract, pancreatic islets, skeletal muscle and gonads of fish (74). The widespread gene expression of IGF-II detected by RT-qPCR in both juvenile and adult fish contrasts the findings in mammals, in which its expression seems to be relevant only during the early stages of development (80). A role in metabolism regulation by IGF-II was demonstrated in muscle cells from trout (81), indicating that this factor could act as a metabolic hormone in fish. There is evidence that GH regulates igf-II gene in all tissues in fish (82–84), while GH most likely regulates only the expression of the igf-I gene in vertebrates. This situation makes fish an excellent model to study species-specific differences in the growth system.
Nutritional Regulation of Hormones Modulating Food Intake in Fish
Feeding
The nutritional status is an important modulator of appetite-regulating hormones in fish. In this respect, several central and peripheral appetite regulators are affected by a single meal, showing periprandial fluctuations in their expression and/or secretion levels. Brain hormones showing such changes in fish include NPY (85, 86), orexin (87–90), CART (87, 88, 91, 92), and nesfatin-1 (93). In general terms, appetite-stimulating or orexigenic factors, including NPY (85, 86), and orexin (89, 90), have been shown to display periprandial changes characterized by higher expression levels before or at mealtime, and lower levels post feeding, suggesting that they are hunger signals. By contrast, appetite inhibiting or anorexigenic signals, such as CART (91, 92), were found to be mainly upregulated after a meal, thus acting as a postprandial satiety signal.
As for peripheral appetite-regulating hormones, the most studied in terms of periprandial fluctuations have been ghrelin, CCK, PYY, and leptin. Circulating levels of ghrelin (in its acylated form) were found to rise pre-prandially in goldfish (94), supporting the role of acyl-ghrelin as a meal initiator in this teleost. Consistently, Unniappan et al. (95) described a significant decrease in total ghrelin in circulation after a meal in goldfish, although this postprandial decrease in plasma ghrelin levels seems to be species-specific as it was detected neither in rainbow trout (96) nor in Mozambique tilapia (97). Ghrelin has also been shown to display periprandial fluctuations in terms of gene expression, although different profiles have been described depending on the species [Atlantic cod, (90); gibel carp, (98); goldfish, (94, 95); Mozambique tilapia, (97, 99); Schizothorax prenanti, (100); zebrafish, (101)]. The periprandial changes of CCK have been only studied in terms of its mRNA expression. Although species-specific changes were observed (88, 89), periprandial variations of cck expression are mainly characterized by an increase in mRNA levels after a meal in the intestine and/or the hypothalamus [dourado, (87); Schizottorax prenanti, (102)]. A similar periprandial profile was described for pyy in the hypothalamus of the Mexican blind cavefish (89) and Schizottorax prenanti (103), in the brain of goldfish (63), and in the brain and gut of Siberian sturgeon (104). Finally, leptin seems only to vary postprandially, although such variations are species- and tissue-specific. Thus, postprandial increases in leptin mRNA expression has been observed in the brain and liver of the mandarin fish (105), in the liver of goldfish (106) and in the visceral adipose tissue of the Atlantic salmon (107), but not in the brain of goldfish (106), the brain and intestine of pacu (88), or the liver of Schizothorax prenanti (102).
Food Deprivation
Food deprivation has been shown to regulate the secretion and expression of appetite-regulating hormones. Table 2 includes a summary of the available studies within the literature and shows the main effect of fasting periods of different duration on the circulating levels and the mRNA expression of the main appetite regulators in several fish species. In general terms, as expected, fasting has been found to upregulate the levels of orexigens and decrease the levels of anorexigens, but several exceptions have been observed depending on the duration of the fasting period, the tissue analyzed and the species (see Table 2 for results and references). In general an upregulation of orexigens and the GH-IGF system should result in an increase in growth. However, without a complete profile of the redundant endocrine milieu, such conclusions are not valid. In addition, a major limitation of many of the studies is that only mRNA expression was determined. Without understanding more about the peptide synthesis in its major tissue sources, and its levels in circulation, it is difficult to reach conclusions on the effects of nutrient status on these hormones.

Table 2. Effects of fasting periods of different duration on the circulating levels and the mRNA expression of the main appetite-regulating hormones in several fish species.
Diet Composition
Another nutritional aspect influencing the appetite-regulating hormones is the composition of diets. This is of great importance as there is significant interest in fisheries and aquaculture in modulating fish growth and reproduction by altering diet and/or endocrine milieu. Therefore, it is important to understand the dietary regulation of hormones, as they have remarkable effects on both reproduction and growth. Although the literature available on this is not very large, several studies have described that altering the macronutrient (i.e., carbohydrates, proteins, and fat) composition of the diet has significant effects on the secretion and/or expression of appetite-regulating hormones in fish.
Carbohydrates (CHO)
Few studies have been performed in fish describing the effects of CHO on appetite regulators. In 2002, Narnaware and Peter described that feeding goldfish CHO-enriched diets significantly reduces NPY mRNA levels in brain areas (175). Additionally, this macronutrient has been shown to upregulate preproghrelin mRNA expression in the pituitary (176). An in vitro assay performed to test the effects of glucose on the expression of appetite-regulating hormones revealed that exposure of goldfish intestinal fragments to different concentrations of this monosaccharide leads to a downregulation of nucb2/nesfatin-1 mRNAs and an upregulation of preproghrelin mRNA expression (177).
Proteins
High-protein diets also modulate important appetite-regulating hormonal systems. A study performed in sea bream revealed that fish fed on high protein diets has higher mRNA expression levels of preproghrelin and cck than those fish fed on diets containing a lower amount of protein (178). In goldfish, feeding diets enriched in this type of macronutrient results in a significant increase in nucb2/nesfatin-1 expression in the pituitary, and a significant decrease in nucb2/nesfatin-1 mRNAs in the gut and in preproghrelin mRNAs in the liver (176). Accordingly, expression of both nucb2/nesfatin-1 and preproghrelin was reduced by direct exposure of goldfish intestinal and hepatopancreas fragments to L-tryptophan (177). Very recently, Volkoff et al. demonstrated that the replacement of dietary fish protein with soy protein does not produce major changes in the expression of cart, orexin, cck, and leptin in the pacu (88). Similarly, varying the diet lysine to arginine ratio has been described to not significantly modify the expression of npy and cck in juvenile cobia (86). Leucine reduced leptin secretion from the adipocytes of food-restricted rainbow trout (57).
Lipids
Intake of fat-enriched diets has been described to reduce the gene expression of npy in the goldfish telencephalon-preoptic area (175), and to increase the mRNA levels of nucb2/nesfatin-1 in the hypothalamus and liver, and of preproghrelin in the pituitary of goldfish (176). Treatment of goldfish intestine with different fatty acids in vitro revealed that fatty acids, in general, downregulate NUCB2/nesfatin-1 in the intestine, but only the longer and highly unsaturated fatty acids inhibit preproghrelin (177). Jönsson and coworkers showed that rainbow trout fed a normal-protein/high-lipid diet tends to have higher plasma ghrelin levels than those fed a high-protein/low-lipid diet (96). Apart from the above-mentioned studies, which were all carried out in adult fish, few studies have been performed at larvae or post-larvae state to study the effects of the replacement of the dietary fat source on the expression of metabolic hormones. Bertucci and coworkers demonstrated that the replacement of dietary fish oil with sunflower oil leads to a decrease in nucb2/nesfatin-1 mRNA expression in pejerrey larvae (179). Additionally, it was described that Senegalese sole post-larvae fed with diets containing soybean oil have higher cart1 and cck mRNA levels in the brain, but lower peripheral cck levels than larvae fed cod liver oil (180).
Nutritional Regulation of the GH-IGF System and Its Influence on Fish Growth
Nutritional Status
The main environmental factor that regulates the GH-IGF system is the nutritional status (181). Table 3 summarizes the effects of fasting on the expression of components of the GH-IGF system in different fish species. During fasting, growth ceases, and energy is mobilized from tissues to support metabolism. This is mainly mediated by changes in the GH-IGF system: plasma levels of GH generally increase while plasma levels of IGFs decrease (182). These changes are explained by a phenomenon known as liver GH resistance, which is characterized by the fact that hepatocytes become resistant to GH, resulting in decreased IGF production despite elevated GH (197). These changes in GH-IGF system during fasting could be adaptive jn response to the high GH plasmatic levels together with low levels of IGF-I (and also insulin) induces lipolysis making fatty acids available to peripheral tissues (197). Using this mechanism, fish are capable to usually maintain its internal functions during fasting, avoiding somatic growth.
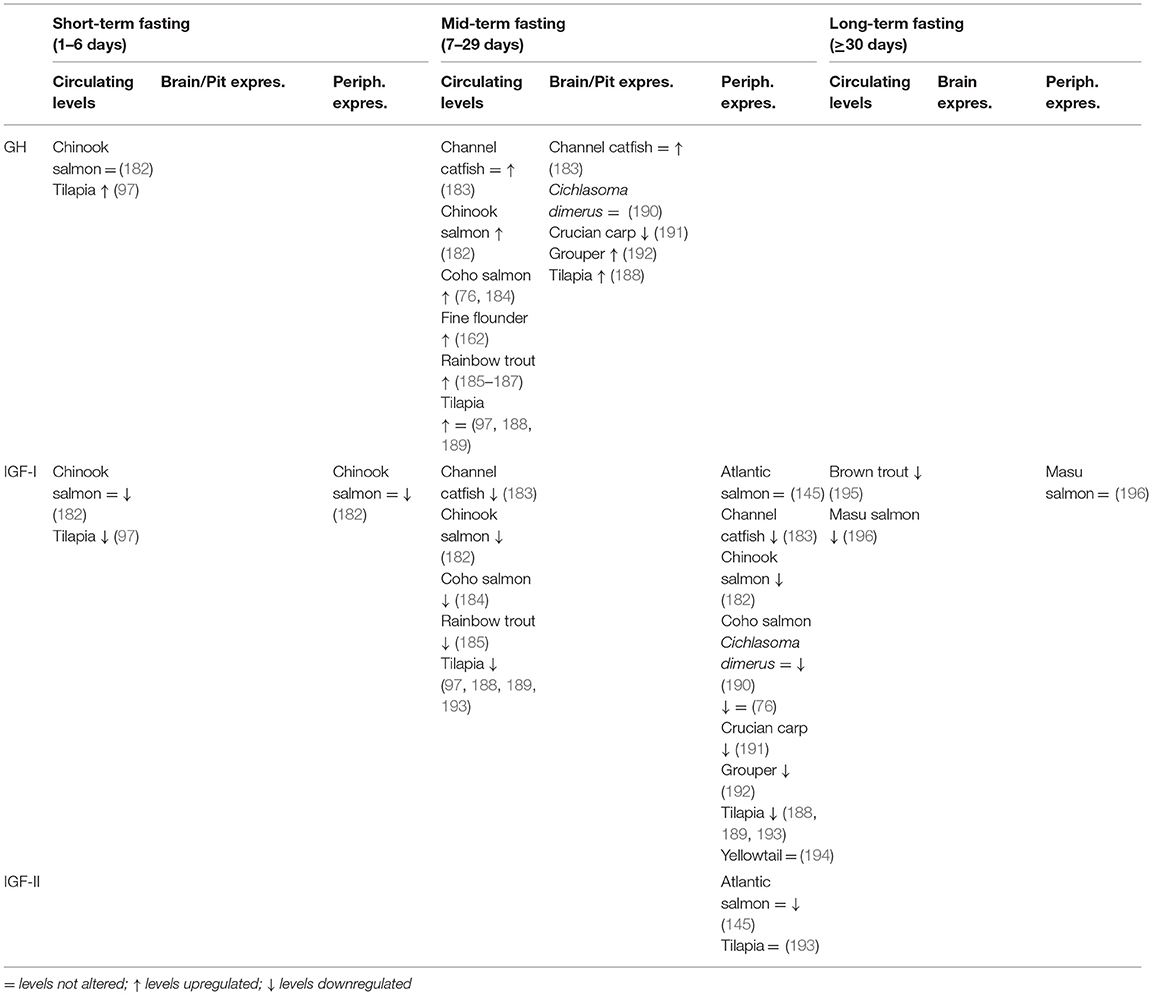
Table 3. Effects of fasting periods of different duration on the circulating levels and the mRNA expression of the GH-IGF system endocrine components in several fish species.
Studies on the effects of feed quantity are available regarding the GH-IGF system. In general, increased feed ration results in inverse changes in GH-IGF system hormones, compared to fasting (182, 198–200). This state in which plasmatic GH levels are low and IGFs are high is correlated with an increase in the somatic growth rate of fish (200). However, if feed ration size is considerable, plasmatic GH levels are high, and GHRs and IGFs levels remain low. This probably diminishes feed utilization for growth (199).
Diet Composition
CHO
Diet composition is another important factor regulating the GH-IGF system and somatic growth (197). Several works have been carried out in the past years aiming to determine the effect of dietary carbohydrates, especially glucose, on GH in fish. Rodgers et al. (201) demonstrated that when tilapia pituitaries were incubated in the presence of varying glucose concentrations, the quantity of GH released is inversely related to glucose concentration in the culture media. However, Riley et al. (202) found no changes in pituitary GH mRNA or plasma GH in response to intraperitoneal (IP) glucose injection. In the liver, IP glucose treatment significantly elevates the levels of GHSR mRNAs. Although the IGF-I mRNA expression was not altered by the IP glucose injection, the IGF-I plasma levels were significantly reduced in tilapia (202). In an in vitro assay carried out by our group (203), it was found that incubation of goldfish hepatopancreas with different glucose concentrations significantly increases the expression of ghr-I, ghr-II, igf-I, and igf-II mRNAs at 4 h. The increase in GHR and GH mRNAs caused by glucose could be related its insulinotropic effect, as such outcomes were demonstrated in mammals (204). IGFs were postulated as regulators of glucose uptake in fish (205), and in mammals (206), likely through the modulation of GLUT-1 glucose transporter (207). GH has been shown to have a hyperglycemic effect in several species of fish, and it is glycogenolytic, glycolytic and gluconeogenetic in some tissues, including the liver, brain, and gill (208). Therefore, IGFs seem to play a role in maintaining the balance between GH and insulin to promote normal carbohydrate metabolism.
Proteins
Dietary protein seems to regulate the hepatic IGF-I expression and secretion, as it was demonstrated in mirror carp (209) and Nile tilapia juveniles (210). In both cases, authors found a significant correlation between dietary protein levels and hepatic IGF-I mRNA expression. Moreover, Qiang et al. (210) found that the increase in the dietary protein content not only increases the IGF-I mRNA expression but also increases the plasmatic IGF-I levels. These authors also reported a significant correlation between the fish-specific growth rate (SGR) and the IGF-I plasmatic levels and/or IGF-I hepatic mRNA expression. In gibel carp (211), it was found that an increase in dietary protein levels leads to an increase in SGR and hepatic IGF-I mRNA levels, although a decrease in both parameters was observed with extremely high protein levels in the diet. Moreover, Tu et al. (211) reported that GH mRNA expression in pituitary shows the opposite trend compared to hepatic IGF-I mRNA. Pérez-Sánchez et al. (199) found that gilthead seabream fed on low protein diets had significantly higher GH, but lower hepatic GH binding and lower IGF-I levels than fish fed on higher protein diets. This situation resembles the one discussed previously in which fish under fasting shows diminished IGFs production and elevated GH. Therefore, low and very high dietary protein levels may influence the GH-IGF axis through the same mechanism as fasting, possibly by direct control of pituitary hormone secretion by circulating nutrients.
Lipids
Dietary lipids have also been shown to modulate fish growth. In several fish species, an increase in the dietary lipid content or a decrease in the protein/lipid ratio was shown to have a negative effect on growth, as reported for the Senegalese sole (212), the turbot (213), and the flounder (214). Although a few studies show the effect of dietary lipids on fish growth performance, there is little knowledge on their effect on the regulation of the GH-IGF system. In Senegalese sole, it was demonstrated that an increase in dietary lipid content increases the hepatic IGF-I mRNA expression, and this was inversely correlated with the somatic growth (215). In pejerrey, an increase from 10 to 21% in the dietary lipid amount generates a decrease in the hepatopancreatic GHR-II mRNA expression, while no changes in somatic growth were found (216). In largemouth bass, the effects of different carbohydrates/lipids (CHO/L) ratios on the GH-IGF system, as well as on the somatic growth, were studied. Authors found that CHO/L ratios from 0.32 to 2.36 significantly upregulate GH mRNA expression and downregulate IGF-I mRNA expression. Higher ratios did not exert any further effects on them. A positive correlation between hepatic IGF-I mRNA levels and specific growth rates with varying dietary CHO/LIP ratios was found (217). All these findings indicate that dietary lipid level can differentially regulate the growth endocrine axis, at least at the transcription level. That can be directly associated with the role of these hormones in regulating lipid homeostasis, and particularly with the direct lipolytic effects of GH and the promotion of tissue growth by IGF-I. Moreover, results presented here also indicate that hepatic IGF-I mRNA as well as its plasma abundance could be a reliable index to assess growth and nutritional fitness.
Several studies have been focused on the effect of fatty acids present in dietary lipids and in the replacement of dietary fish oil by vegetable oil on fish growth. In most of the cases, an increase in the dietary amount of fatty acids from vegetable sources causes an increase in GH mRNA expression and/or circulating levels, while diminishes the IGF-I plasmatic and GHR-I mRNA levels (179, 203, 218). The hepatic mRNA expression of GHR-II and IGF-II seems to be constitutive and not affected by dietary fatty acids in sea bream (218), while in pejerrey the replacement of dietary fish oil by sunflower oil increases their mRNA expression (179). As a general conclusion, large replacements of high-unsaturated fatty acids by low-unsaturated fatty acids in fish diet lead to the GH-IGF system pattern to resemble one observed during fasting.
Micronutrients
The effect of micronutrients on the expression of GH-IGF system components was not extensively studied. To the best of our knowledge there is only one work reporting an effect of vitamins on igf-I expression (219). In this, the authors found that 4% of dietary vitamins generate an increase in the IGF-I mRNA expression in sea bass larvae after 38 days of the experiment, compared with diets containing lower or higher amounts.
Future Perspectives
The role of nutrition as a potent modulator of the endocrine system governing feeding behavior and growth in fish is described in this review. Aquaculture research is a topic of increasing interest due to the demand for sustainable food production. That makes it essential to summarize the knowledge generated in the last past years in order to identify the weak points and to determine the direction in which new studies should be focused. As described here, food availability and feed composition are two of the most influent external signals modulating appetite-regulating and growth modulatory hormones. Future research should, therefore, be focused on tapping into this knowledge, expanding it in depth, and exploring its use to enhance feeding and growth in fish. Such approaches will eventually lead to increased yield in cultured fish.
It is also interesting to note the species-specific differences in the effect of feeding on the expression of appetite-regulating hormones and the GH-IGF system. Such differences could be related to the feeding behavior of specific fish species, as the carnivorous, herbivorous and omnivorous fish tend to fall into three different groups of responses. This fact also reflects the flexibility of metabolic systems, considering that the same components present in all fish species could generate distinct responses from the same food stimuli. Therefore, it is critical to bear in mind that results obtained from one species of fish might not apply to another.
As a final consideration, it would be interesting to broaden our knowledge of the crosstalk between hormones regulating food intake and those regulating growth. For instance, GH and IGF-I not only regulate somatic growth but can at the same time modulate lipid and carbohydrate metabolism, respectively (220). Others, including ghrelin, could stimulate food intake (221), and also GH release from pituitary (222, 223). Likewise, it has been shown that leptin directly regulates the expression of IGFs and GHRs in fish hepatopancreas (224). These are a few examples of the crosslinks mentioned above, that serve to highlight the close interaction that exists between hormones controlling food intake and growth to ensure the proper growth and development of fish. Thus, comprehensive approaches to determine both metabolic and growth regulatory hormone responses to nutritional challenges are more desirable from an aquaculture perspective. Future research must focus to identify gaps in knowledge, including the ones identified above. In addition, the use of alternate feed ingredients, use of endogenous feeding and growth regulatory factors as feed additives, and employing hormones using targeted molecular and cellular approaches should be explored to modulate growth rate and yield in cultured species.
Author Contributions
JB prepared larger sections and complied the manuscript. AB, LS, JR, and CV wrote smaller sections. SU provided the idea, outline, provided feedback, and corrected the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The research on fish metabolism in the laboratory of SU has been supported by Discovery Grants from the Natural Sciences and Engineering Research Council (NSERC) of Canada. Research lab infrastructure was purchased using a Leaders Opportunities Fund from the Canada Foundation for Innovation (CFI), and an Establishment Grant from the Saskatchewan Health Research Foundation (SHRF).
References
1. Douros JD, Baltzegar DA, Mankiewicz J, Taylor J, Yamaguchi Y, Lerner DT, et al. Control of leptin by metabolic state and its regulatory interactions with pituitary growth hormone and hepatic growth hormone receptors and insulin like growth factors in the tilapia (Oreochromis mossambicus). Gen Comp Endocrinol. (2017) 240:227–37. doi: 10.1016/j.ygcen.2016.07.017
2. Mommsen TP. Paradigms of growth in fish. Comp Biochem Physiol B Biochem Mol Biol. (2001) 129:207–19. doi: 10.1016/S1096-4959(01)00312-8
3. Näslund E, Hellström PM. Appetite signaling: From gut peptides and enteric nerves to brain. Physiol Behav. (2007) 92:256–62. doi: 10.1016/j.physbeh.2007.05.017
4. Reindl KM, Sheridan MA. Peripheral regulation of the growth hormone-insulin-like growth factor system in fish and other vertebrates. Comp Biochem Physiol A Mol Integr Physiol. (2012) 163:231–45. doi: 10.1016/j.cbpa.2012.08.003
5. Kageyama H, Takenoya F, Shiba K, Shioda S. Neuronal circuits involving ghrelin in the hypothalamus-mediated regulation of feeding. Neuropeptides. (2010) 44:133–8. doi: 10.1016/j.npep.2009.11.010
6. Crespo CS, Cachero AP, Jiménez LP, Barrios V, Ferreiro EA. Peptides and food intake. Front Endocrinol. (2014) 5:58. doi: 10.3389/fendo.2014.00058
7. Michel M, Volkoff H, Conde-Sieira M, Soengas JL. Nutrient sensing systems in fish: impact on food intake regulation and energy homeostasis. Nutr Sens Mech Fish. (2017) 10:1–21. doi: 10.3389/fnins.2016.00603
8. Efeyan A, Comb WC, Sabatini DM. Nutrient-sensing mechanisms and pathways. Nature. (2015) 517:302–10. doi: 10.1038/nature14190
9. Gatlin DM. Food intake in fish. In: Houlihan D, Boujard T, Jobling M, editors. COST Action 827. Oxford, UK: Blackwell Science (2001). p. 259. doi: 10.1016/S0044-8486(02)00066-2
10. Council NR. Nutrient Requirements of Horses. 6th ed. Washington, DC: The National Academies Press (2007).
11. Mommsen TP. (2002). Carbohydrates in fish nutrition effects on growth, glucose metabolism and hepatic enzymes.
12. Polakof S, Panserat S, Soengas JL, Moon TW. Glucose metabolism in fish: a review. J Comp Physiol B Biochem Syst Environ Physiol. (2012) 182:1015–45. doi: 10.1007/s00360-012-0658-7
13. Moon TW. Glucose intolerance in teleost fish: fact or fiction? Comp Biochem Physiol B Biochem Mol Biol. (2001) 129:243–9. doi: 10.1016/S1096-4959(01)00316-5
14. Soengas JL, Aldegunde M. Energy metabolism of fish brain. Comp Biochem Physiol B Biochem Mol Biol. (2002) 131:271–96. doi: 10.1016/S1096-4959(02)00022-2
15. Sargent J, Tocher D, Bell J. The lipids. Fish Nutr. (2002) 181–257. doi: 10.1016/B978-012319652-1/50005-7
16. Torstensen BE, Lie Ø, Frøyland L. Lipid metabolism and tissue composition in Atlantic salmon (Salmo salar L.)- Effects of capelin oil, palm oil, and oleic scid-enriched sunflower oil as dietary lipid sources. Lipids. (2000) 35:653–64. doi: 10.1007/s11745-000-0570-6
17. Tocher DR, Fraser AJJ, Sargent JRR, Gamble JCC. Fatty acid composition of phospholipids and neutral lipids during embryonic and early larval development in atlantic herring (Clupea harengus, L.). Lipids. (1985) 20:69–74. doi: 10.1007/BF02534210
18. Sheridan MA. Regulation of Lipid-Metabolism in Poikilothermic Vertebrates. Comp Biochem Physiol B Biochemistry Mol Biol. (1994) 107:495–508. doi: 10.1016/0305-0491(94)90176-7
19. Taylor P, Tocher DR. Reviews in fisheries science metabolism and functions of lipids and fatty acids in teleost fish. Rev Fish Sci. (2010) 37–41. doi: 10.1080/713610925
20. Craig S, Helfrich LA. Understanding Fish Nutrition Feeds, and Feeding. Virginia Coop Ext (2002) p. 1–18. Available online at: https://pubs.ext.vt.edu/420/420-256/420-256.html
21. Huntingford F, Jobling M, Kadri S. Aquaculture and Behavior. Chichester, UK: John Wiley & Sons (2011).
22. Langhans W. Appetite regulation. In: Lobley GE, White A, MacRae JC, editors. Protein Metabolism and Nutrition. Wageningen Pers (1999). p. 225–51.
23. Lawrence JM. Preliminary results on the use of potassium permanganate to counteract the effects of rotenone on fish. Progress Fish-Culturist. (1956) 18:15–21.
24. Chantranupong L, Wolfson RL, Sabatini DM. Nutrient-sensing mechanisms across evolution. Cell. (2015) 161:67–83. doi: 10.1016/j.cell.2015.02.041
25. Morgan K, Obici S, Rossetti L. Hypothalamic responses to long-chain fatty acids are nutritionally regulated. J Biol Chem. (2004) 279:31139–48. doi: 10.1074/jbc.M400458200
26. Pocai A, Obici S, Schwartz GJ, Rossetti L. A brain-liver circuit regulates glucose homeostasis. Cell Metab. (2005) 1:53–61. doi: 10.1016/j.cmet.2004.11.001
27. Roh E, Song DK, Kim, M-S. Emerging role of the brain in the homeostatic regulation of energy and glucose metabolism. Exp Mol Med. (2016) 48:e216. doi: 10.1038/emm.2016.4
28. Morton GJ, Meek TH, Schwartz MW. Neurobiology of food intake in health and disease. Nat Rev Neurosci. (2014) 15:367–78. doi: 10.1038/nrn3745
29. Rogers RC, Ritter S, Hermann GE, Rc R, Ritter S, Hindbrain HGE. Hindbrain cytoglucopenia-induced increases in systemic blood glucose levels by 2-deoxyglucose depend on intact astrocytes and adenosine release. Am J Physiol Regul Integr Comp Physiol. (2016) 310:1102–8. doi: 10.1152/ajpregu.00493.2015
30. Lerch M, Mayrhofer M, Zerbe O. Structural similarities of micelle-bound peptide YY (PYY) and neuropeptide Y (NPY) are related to their affinity profiles at the Y receptors. J Mol Biol. (2004) 339:1153–1168. doi: 10.1016/j.jmb.2004.04.032
31. Cerdá-Reverter JM, Larhammar D. Neuropeptide Y family of peptides: structure, anatomical expression, function, and molecular evolution. Biochem Cell Biol. (2000) 78:371–92. doi: 10.1139/o00-004
32. Campos VF, Collares T, Deschamps JC, Seixas FK, Dellagostin OA, Lanes CF, et al. Identification, tissue distribution and evaluation of brain neuropeptide Y gene expression in the Brazilian flounder Paralichthys orbignyanus. J Biosci. (2010) 35:405–13. doi: 10.1007/s12038-010-0046-y
33. Volkoff H. The neuroendocrine regulation of food intake in fish: a review of current knowledge. Front Neurosci. (2016) 10:540. doi: 10.3389/fnins.2016.00540
34. Cerdá-Reverter JM, Peter RE. Endogenous melanocortin antagonist in fish: structure, brain mapping, and regulation by fasting of the goldfish agouti-related protein gene. Endocrinology. (2003) 144:4552–61. doi: 10.1210/en.2003-0453
35. Cerdá-Reverter JM, Canosa LF. Neuroendocrine systems of the fish brain. In: Bernier NJ, Van Der Kraak G, Farrell AP, Brauner CJ, editors. Fish Neuroendocrinology: Fish Physiology. (2009) 28, p. 3–74.
36. Noon LA, Franklin JM, King PJ, Goulding NJ, Hunyady L, Clark AJ. Failed export of the adrenocorticotrophin receptor from the endoplasmic reticulum in non-adrenal cells: evidence in support of a requirement for a specific adrenal accessory factor. J Endocrinol. (2002) 174:17–25. doi: 10.1677/joe.0.1740017
37. Metherell LA, Chapple JP, Cooray S, David A, Becker C, Rüschendorf F, et al. Mutations in MRAP, encoding a new interacting partner of the ACTH receptor, cause familial glucocorticoid deficiency type 2. Nat Genet. (2005) 37:166–70. doi: 10.1038/ng1501
38. Douglass J, McKinzie AA, Couceyro P. PCR differential display identifies a rat brain mRNA that is transcriptionally regulated by cocaine and amphetamine. J Neurosci. (1995) (3 Pt 2):2471–81. doi: 10.1523/JNEUROSCI.15-03-02471.1995
39. Volkoff H, Peter RE. Characterization of two forms of cocaine- and amphetamine-regulated transcript (CART) peptide precursors in goldfish: molecular cloning and distribution, modulation of expression by nutritional status, and interactions with leptin. Endocrinology. (2001) 2:5076–88. doi: 10.1210/endo.142.12.8519
40. Shewale SA, Gaupale TC, Bhargava S. Temperature dependent changes in cocaine-and amphetamine regulated transcript (CART) peptide in the brain of tadpole, Sylvirana temporalis. Gen Comp Endocrinol. (2015) 220:61–9. doi: 10.1016/j.ygcen.2014.06.025
41. Gilon P. Cocaine-and amphetamine-regulated transcript: a novel regulator of energy homeostasis expressed in a subpopulation of pancreatic islet cells. Diabetologia. (2016) 59:1855–9. doi: 10.1007/s00125-016-4052-y
42. Ferguson AV, Samson WK. The orexin/hypocretin system: a critical regulator of neuroendocrine and autonomic function. Front Neuroendocrinol. (2003) 24:141–50. doi: 10.1016/S0091-3022(03)00028-1
43. Rodgers RJ, Ishii Y, Halford JC, Blundell JE. Orexins and appetite regulation. Neuropeptides. (2002) 36:303–25 doi: 10.1016/S0143-4179(02)00085-9
44. Hagan JJ, Leslie RA, Patel S, Evans ML, Wattam TA, Holmes S, et al. Orexin A activates locus coeruleus cell firing and increases arousal in the rat. Proc Natl Acad Sci USA. (1999) 96:10911–10916. doi: 10.1073/pnas.96.19.10911
45. Volkoff H, Peter RE. Interactions between orexin A, NPY and galanin in the control of food intake of the goldfish, Carassius auratus. Regul Pept. (2001) 101:59–72. doi: 10.1016/S0167-0115(01)00261-0
46. Kawauchi H, Kawazoe I, Tsubokawa M, Kishida M, Baker BI. Characterization of melanin-concentrating hormone in chum salmon pituitaries. Nature. (1983) 305:321–3. doi: 10.1038/305321a0
47. Logan DW, Bryson-Richardson RJ, Pagán KE, Taylor MS, Currie PD, Jackson IJ. The structure and evolution of the melanocortin and MCH receptors in fish and mammals. Genomics. (2003) 81:184–91. doi: 10.1016/S0888-7543(02)00037-X
48. Oh, -I. S., Shimizu H, Satoh T, Okada S, Sachika A, Inoue K, et al. Identification of nesfatin-1 as a satiety molecule in the hypothalamus. Nature. (2006) 443:709–12. doi: 10.1038/nature05162
49. Iwasaki Y, Nakabayashi H, Kakei M, Shimizu H, Mori M, Yada T. Nesfatin-1 evokes Ca2+ signaling in isolated vagal afferent neurons via Ca2+ influx through N-type channels. Biochem Biophys Res Commun. (2009) 390:958–62. doi: 10.1016/j.bbrc.2009.10.085
50. Gonzalez R, Kerbel B, Chun A, Unniappan S. Molecular, cellular and physiological evidences for the anorexigenic actions of nesfatin-1 in goldfish. PLoS ONE. (2010) 5:e15201. doi: 10.1371/journal.pone.0015201
51. Lin F, Zhou C, Chen H, Wu H, Xin Z, Liu J, et al. Molecular characterization, tissue distribution and feeding related changes of NUCB2A/nesfatin-1 in Ya-fish (Schizothorax prenanti). Gene (2014) 536:238–46. doi: 10.1016/j.gene.2013.12.031
52. Caldwell LK, Pierce AL, Riley LG3m Duncan CA, Nagler JJ. Plasma nesfatin-1 is not affected by long-term food restriction and does not predict rematuration among iteroparous female rainbow trout (Oncorhynchus mykiss). PLoS ONE. (2014) 9:e85700. doi: 10.1371/journal.pone.0085700
53. Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H, Kangawa K, et al. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature. (1999) 402:656–60. doi: 10.1038/45230
54. Kaiya H, Kojima M, Hosoda H, Moriyama S, Takahashi A, Kawauchi H, et al. Peptide purification, complementary deoxyribonucleic acid (DNA) and genomic DNA cloning, and functional characterization of ghrelin in rainbow trout. Endocrinology. (2003) 144:5215–26. doi: 10.1210/en.2003-1085
55. Kaiya H, Miyazato M, Kangawa K, Peter RE, Unniappan S. Ghrelin: a multifunctional hormone in non-mammalian vertebrates. Comp Biochem Physiol A Mol Integr Physiol. (2008) 149:109–28. doi: 10.1016/j.cbpa.2007.12.004
56. Jönsson E. The role of ghrelin in energy balance regulation in fish. Gen Comp Endocrinol. (2013) 187:79–85. doi: 10.1016/j.ygcen.2013.03.013
57. Salmerón C, Johansson M, Asaad M, Angotzi AR, Rønnestad I, Stefansson SO, et al. Roles of leptin and ghrelin in adipogenesis and lipid metabolism of rainbow trout adipocytes in vitro. Comp Biochem Physiol A Mol Integr Physiol. (2015) 188:40–8. doi: 10.1016/j.cbpa.2015.06.017
58. Beinfeld MC. Biosynthesis and processing of pro CCK: recent progress and future challenges. Life Sci. (2003) 72:747–57. doi: 10.1016/S0024-3205(02)02330-5
59. Kurokawa T, Suzuki T, Hashimoto H. Identification of gastrin and multiple cholecystokinin genes in teleost. Peptides. (2003) 24:227–35. doi: 10.1016/S0196-9781(03)00034-2
60. Aldman G, Holmgren S. Intraduodenal fat and amino acids activate gallbladder motility in the rainbow trout, Oncorhynchus mykiss. Gen Comp Endocrinol. (1995) 100:27–32. doi: 10.1006/gcen.1995.1128
61. Himick BA, Peter RE. CCK/gastrin-like immunoreactivity in brain and gut, and CCK suppression of feeding in goldfish. Am J Physiol. (1994) 267(3 Pt 2):R841–51.
62. Volkoff H, Eykelbosh AJ, Peter RE. Role of leptin in the control of feeding of goldfish Carassius auratus: interactions with cholecystokinin, neuropeptide Y and orexin A, and modulation by fasting. Brain Res. (2003) 972:90–109. doi: 10.1016/S0006-8993(03)02507-1
63. Gonzalez R, Unniappan S. Molecular characterization, appetite regulatory effects and feeding related changes of peptide YY in goldfish. Gen Comp Endocrinol. (2010) 166:273–9. doi: 10.1016/j.ygcen.2009.09.008
64. Batterham RL, Cowley MA, Small CJ, Herzog H, Cohen MA, Dakin CL, et al. Gut hormone PYY(3-36) physiologically inhibits food intake. Nature. (2002) 418:650–4. doi: 10.1038/nature00887
65. Cerdá-Reverter JM, Martínez-Rodríguez G, Zanuy S, Carrillo M, Larhammar D. Molecular evolution of the neuropeptide Y (NPY) family of peptides: cloning of three NPY-related peptides from the sea bass (Dicentrarchus labrax). Regul Pept. (2000) 95:25–34. doi: 10.1016/S0167-0115(00)00132-4
66. Plisetskaya EM, Mommsen TP. Glucagon and glucagon-like peptides in fishes. Int Rev Cytol. (1996) 168:187–257. doi: 10.1016/S0074-7696(08)60885-2
67. Yeung CM, Mojsov S, Mok PY, Chow BK. Isolation and structure-function studies of a glucagon-like peptide 1 receptor from goldfish Carassius auratus: identification of three charged residues in extracellular domains critical for receptor function. Endocrinology. (2002) 143:4646–54. doi: 10.1210/en.2002-220694
68. Londraville RL, Macotela Y, Duff RJ, Easterling MR, Liu Q, Crespi EJ. Comparative endocrinology of leptin: assessing function in a phylogenetic context. Gen Comp Endocrinol. (2014) 203:146–57. doi: 10.1016/j.ygcen.2014.02.002
69. Song YF, Wu K, Tan XY, Zhang LH, Zhuo MQ, Pan YX, et al. Effects of recombinant human leptin administration on hepatic lipid metabolism in yellow catfish Pelteobagrus fulvidraco: in vivo and in vitro studies. Gen Comp Endocrinol. (2015) 212:92–9. doi: 10.1016/j.ygcen.2015.01.022
70. Gong N, Jönsson E, Björnsson BT. Acute anorexigenic action of leptin in rainbow trout is mediated by the hypothalamic Pi3k pathway. J Mol Endocrinol. (2016) 56:227–38. doi: 10.1530/JME-15-0279
71. Musumeci G, Trovato FM, Avola R, Imbesi R, Castrogiovanni P. Serotonin/growth hormone/insulin-like growth factors axis on pre- and post-natal development: a contemporary review. OA Anat. (2013) 1:1–7.
72. Johnsson JI, Björnsson BT. Growth hormone increases growth rate, appetite and dominance in juvenile rainbow trout, Oncorhynchus mykiss. Anim. Behav. (1994) 48:177–86. doi: 10.1006/anbe.1994.1224
73. Abrahams MV, Sutterlin A. The foraging and antipredator behaviour of growth-enhanced transgenic Atlantic salmon. Anim Behav. (1999) 58:933–42. doi: 10.1006/anbe.1999.1229
74. Reinecke M, Björnsson BT, Dickhoff WW, McCormick SD, Navarro I, Power DM, et al. Growth hormone and insulin-like growth factors in fish: where we are and where to go. Gen Comp Endocrinol. (2005) 142:20–4. doi: 10.1016/j.ygcen.2005.01.016
75. Pérez-Sánchez J, Calduch-Giner JA, Mingarro M, de Celis SVR, Gomez-Requeni P, Saera-Vila A, et al. Overview of fish growth hormone family. New insights in genomic organization and heterogeneity of growth hormone receptors. Fish Physiol Biochem. (2002) 27:243–58. doi: 10.1023/B:FISH.0000032729.72746.c8
76. Duan C, Plisetskaya EM. Nutritional regulation of insulin-like growth factor-I mRNA expression in salmon tissues. J Endocrinol. (1993) 139:243–52. doi: 10.1677/joe.0.1390243
77. Duguay SJ, Park LK, Samadpour M, Dickhoff WW. Nucleotide sequence and tissue distribution of three insulin-like growth factor I prohormones in salmon. Mol Endocrinol. (1992) 6:1202–10. doi: 10.1210/mend.6.8.1406698
78. Shamblott MJ, Chen TT. Identification of a second insulin-like growth factor in a fish species. Proc Natl Acad Sci USA. (1992) 89 8913–7. doi: 10.1073/pnas.89.19.8913
79. Perrot V, Moiseeva EB, Gozes Y, Chan SJ, Funkenstein B. Insulin-like growth factor receptors and their ligands in gonads of a hermaphroditic species, the gilthead seabream (Sparus aurata): expression and cellular localization. Biol Reprod. (2000) 63:229–41. doi: 10.1095/biolreprod63.1.229
80. Reinecke M, Collet C. The phylogeny of the insulin-like growth factors. Int Rev Cytol. (1998) 183:1–94. doi: 10.1016/S0074-7696(08)60142-4
81. Codina M, Castillo J, Gutiérrez J, Navarro I. Role of IGF-II in the metabolism and proliferation of rainbow trout muscle cells. Gen Comp Endocrinol. (2004) (Abstract 5th ISFE, O41).
82. Shamblott M, Cheng C, Bolt D, Chen T. Appearance of insulin-like growth factor mRNA in the liver and pyloric ceca of a teleost in response to exogenous growth hormone. Proc Natl Acad Sci USA. (1995) 92:6943–6. doi: 10.1073/pnas.92.15.6943
83. Tse MCL, Vong QP, Cheng CHK, Chan KM. PCR-cloning and gene expression studies in common carp (Cyprinus carpio) insulin-like growth factor-II. Biochim Biophys Acta. (2002) 1575:63–74. doi: 10.1016/S0167-4781(02)00244-0
84. Vong QP, Chan KM, Cheng CHK. Quantification of common carp (Cyprinus carpio) IGF-I and IGF-II mRNA by real-time PCR: differential regulation of expression by GH. J Endocrinol. (2003) 178:513–21. doi: 10.1677/joe.0.1780513
85. Campos VF, Robaldo RB, Deschamps JC, Seixas FK, McBride AJA, Marins LF, et al. Neuropeptide Y gene expression around meal time in the Brazilian flounder Paralichthys orbignyanus. J Biosci. (2012) 37:227–32. doi: 10.1007/s12038-012-9205-7
86. Nguyen MV, Jordal A-EO, Espe M, Buttle L, Lai HV, Rønnestad I. Feed intake and brain neuropeptide Y (NPY) and cholecystokinin (CCK) gene expression in juvenile cobia fed plant-based protein diets with different lysine to arginine ratios. Comp Biochem Physiol A Mol Integr Physiol. (2013) 165:328–37. doi: 10.1016/j.cbpa.2013.04.004
87. Volkoff H, Sabioni RE, Cyrino JEP. Appetite regulating factors in dourado, Salminus brasiliensis: cDNA cloning and effects of fasting and feeding on gene expression. Gen Comp Endocrinol. (2016) 237:34–42. doi: 10.1016/j.ygcen.2016.07.022
88. Volkoff H, Estevan Sabioni R, Coutinho LL, Cyrino JEP. Appetite regulating factors in pacu (Piaractus mesopotamicus): tissue distribution and effects of food quantity and quality on gene expression. Comp Biochem Physiol A Mol Integr Physiol. (2017) 203:241–54. doi: 10.1016/j.cbpa.2016.09.022
89. Wall A, Volkoff H. Effects of fasting and feeding on the brain mRNA expressions of orexin, tyrosine hydroxylase (TH), PYY and CCK in the Mexican blind cavefish (Astyanax fasciatus mexicanus). Gen Comp Endocrinol. (2013) 183:44–52. doi: 10.1016/j.ygcen.2012.12.011
90. Xu M, Volkoff H. Molecular characterization of prepro-orexin in Atlantic cod (Gadus morhua): cloning, localization, developmental profile and role in food intake regulation. Mol Cell Endocrinol. (2007) 271:28–37. doi: 10.1016/j.mce.2007.03.003
91. Yuan D, Wei R, Wang T, Wu Y, Lin F, Chen H, et al. Appetite regulation in Schizothorax prenanti by three CART genes. Gen Comp Endocrinol. (2015) 224:194–204. doi: 10.1016/j.ygcen.2015.08.015
92. Zhang X, Gao Y, Tang N, Qi J, Wu Y, Hao J, et al. One evidence of cocaine- and amphetamine-regulated transcript (CART) has the bidirectional effects on appetite in Siberian sturgeon (Acipenser baerii). Fish Physiol Biochem. (2018) 44:411–22. doi: 10.1007/s10695-017-0444-2
93. Hatef A, Shajan S, Unniappan S. Nutrient status modulates the expression of nesfatin-1 encoding nucleobindin 2A and 2B mRNAs in zebrafish gut, liver and brain. Gen Comp Endocrinol. (2015) 215:51–60. doi: 10.1016/j.ygcen.2014.09.009
94. Blanco AM, Gómez-Boronat M, Redondo I, Valenciano AI, Delgado MJ. Periprandial changes and effects of short- and long-term fasting on ghrelin, GOAT, and ghrelin receptors in goldfish (Carassius auratus). J. Comp. Physiol B. (2016) 186:727–38. doi: 10.1007/s00360-016-0986-0
95. Unniappan S, Canosa LF, Peter RE. Orexigenic actions of ghrelin in goldfish: feeding-induced changes in brain and gut mRNA expression and serum levels, and responses to central and peripheral injections. Neuroendocrinology. (2004) 79:100–8. doi: 10.1159/000076634
96. Jönsson E, Forsman A, Einarsdottir IE, Kaiya H, Ruohonen K, Björnsson BT. Plasma ghrelin levels in rainbow trout in response to fasting, feeding and food composition, and effects of ghrelin on voluntary food intake. Comp Biochem Physiol A Mol Integr Physiol. (2007) 147:1116–24. doi: 10.1016/j.cbpa.2007.03.024
97. Fox BK, Breves JP, Hirano T, Grau EG. Effects of short- and long-term fasting on plasma and stomach ghrelin, and the growth hormone/insulin-like growth factor I axis in the tilapia, Oreochromis mossambicus. Domest Anim Endocrinol. (2009) 37:1–11. doi: 10.1016/j.domaniend.2009.01.001
98. Zhou C, Zheng J, Lei L, Yuan D, Zhu C, Ye H, et al. Evidence that ghrelin may be associated with the food intake of gibel carp (Carassius auratus gibelio). Fish Physiol. Biochem. (2016) 42:1637–46. doi: 10.1007/s10695-016-0246-y
99. Peddu SC, Breves JP, Kaiya H, Gordon Grau E, Riley LG. Pre- and postprandial effects on ghrelin signaling in the brain and on the GH/IGF-I axis in the Mozambique tilapia (Oreochromis mossambicus). Gen Comp Endocrinol. (2009) 161:412–8. doi: 10.1016/j.ygcen.2009.02.008
100. Wei R, Liu T, Zhou C, Zhang X, Yuan D, Wang T, et al. Identification, tissue distribution and regulation of preproghrelin in the brain and gut of Schizothorax prenanti. Regul Pept. (2013) 186:18–25. doi: 10.1016/j.regpep.2013.07.002
101. Hatef A, Yufa R, Unniappan S. Ghrelin O-Acyl Transferase in Zebrafish is an evolutionarily conserved peptide upregulated during calorie restriction. Zebrafish. (2015) 12:327–38. doi: 10.1089/zeb.2014.1062
102. Yuan D, Wang T, Zhou C, Lin F, Chen H, Wu H, et al. Leptin and cholecystokinin in Schizothorax prenanti: molecular cloning, tissue expression, and mRNA expression responses to periprandial changes and fasting. Gen Comp Endocrinol. (2014) 204:13–24. doi: 10.1016/j.ygcen.2014.05.013
103. Yuan D, Zhou C, Wang T, Lin F, Chen H, Wu H, et al. Molecular characterization and tissue expression of peptide YY in Schizothorax prenanti: effects of periprandial changes and fasting on expression in the hypothalamus. Regul Pept. (2014) 190–1:32–8. doi: 10.1016/j.regpep.2014.03.004
104. Chen H, Zhang X, Hao J, Chen D, Liu J, Gao Y, et al. Molecular cloning, expression analysis, and appetite regulatory effect of peptide YY in Siberian sturgeon (Acipenser baerii). Gene (2015) 563:172–9. doi: 10.1016/j.gene.2015.03.028
105. Yuan X, Li A, Liang X-F, Huang W, Song Y, He S, et al. Leptin expression in mandarin fish Siniperca chuatsi (Basilewsky): regulation by postprandial and short-term fasting treatment. Comp Biochem Physiol A Mol Integr Physiol. (2016) 194:8–18. doi: 10.1016/j.cbpa.2016.01.014
106. Tinoco AB, Nisembaum LG, Isorna E, Delgado MJ, de Pedro N. Leptins and leptin receptor expression in the goldfish (Carassius auratus). Regulation by food intake and fasting/overfeeding conditions. Peptides. (2012) 34:329–35. doi: 10.1016/j.peptides.2012.02.001
107. Moen A-GG, Finn RN. Short-term, but not long-term feed restriction causes differential expression of leptins in Atlantic salmon. Gen Comp Endocrinol. (2013) 183:83–8. doi: 10.1016/j.ygcen.2012.09.027
108. Murashita K, Kurokawa T, Ebbesson LOE, Stefansson SO, Rønnestad I. Characterization, tissue distribution, and regulation of agouti-related protein (AgRP), cocaine- and amphetamine-regulated transcript (CART) and neuropeptide Y (NPY) in Atlantic salmon (Salmo salar). Gen Comp Endocrinol. (2009) 162:160–71. doi: 10.1016/j.ygcen.2009.03.015
109. Ji W, Ping H-C, Wei K-J, Zhang G-R, Shi Z-C, Yang R-B, et al. Ghrelin, neuropeptide Y (NPY) and cholecystokinin (CCK) in blunt snout bream (Megalobrama amblycephala): cDNA cloning, tissue distribution and mRNA expression changes responding to fasting and refeeding. Gen Comp Endocrinol. (2015) 223:108–19. doi: 10.1016/j.ygcen.2015.08.009
110. Narnaware YK, Peter RE. Effects of food deprivation and refeeding on neuropeptide Y (NPY) mRNA levels in goldfish. Comp Biochem Physiol B Biochem Mol Biol. (2001) 129:633–7. doi: 10.1016/S1096-4959(01)00359-1
111. Silverstein JT, Breininger J, Baskin DG, Plisetskaya EM. Neuropeptide Y-like gene expression in the salmon brain increases with fasting. Gen Comp Endocrinol. (1998) 110:157–65. doi: 10.1006/gcen.1998.7058
112. Babichuk NA, Volkoff H. Changes in expression of appetite-regulating hormones in the cunner (Tautogolabrus adspersus) during short-term fasting and winter torpor. Physiol Behav. (2013) 120:54–63. doi: 10.1016/j.physbeh.2013.06.022
113. Yan P, Jia J, Yang G, Wang D, Sun C, Li W. Duplication of neuropeptide Y and peptide YY in Nile tilapia Oreochromis niloticus and their roles in food intake regulation. Peptides. (2017) 88:97–105. doi: 10.1016/j.peptides.2016.12.010
114. Pitts PM, Volkoff H. Characterization of appetite-regulating factors in platyfish, Xiphophorus maculatus (Cyprinodontiformes Poeciliidae). Comp Biochem Physiol A Mol Integr Physiol. (2017) 208:80–8. doi: 10.1016/j.cbpa.2017.03.018
115. Wei R, Zhou C, Yuan D, Wang T, Lin F, Chen H, et al. Characterization, tissue distribution and regulation of neuropeptideY in Schizothorax prenanti. J Fish Biol. (2014) 85:278–91. doi: 10.1111/jfb.12413
116. Kamijo M, Kojima K, Maruyama K, Konno N, Motohashi E, Ikegami T, et al. Neuropeptide Y in tiger puffer (Takifugu rubripes): distribution, cloning, characterization, and mRNA expression responses to prandial condition. Zoolog Sci. (2011) 28:882–90. doi: 10.2108/zsj.28.882
117. MacDonald E, Volkoff H. Cloning, distribution and effects of season and nutritional status on the expression of neuropeptide Y (NPY), cocaine and amphetamine regulated transcript (CART) and cholecystokinin (CCK) in winter flounder (Pseudopleuronectes americanus). Horm Behav. (2009) 56:58–65. doi: 10.1016/j.yhbeh.2009.03.002
118. MacDonald E, Volkoff H. Neuropeptide Y (NPY), cocaine- and amphetamine-regulated transcript (CART) and cholecystokinin (CCK) in winter skate (Raja ocellata): cDNA cloning, tissue distribution and mRNA expression responses to fasting. Gen Comp Endocrinol. (2009) 161:252–61. doi: 10.1016/j.ygcen.2009.01.021
119. Hosomi N, Furutani T, Takahashi N, Masumoto T, Fukada H. Yellowtail neuropeptide Y: molecular cloning, tissue distribution, and response to fasting. Fish Sci. (2014) 80:483–92. doi: 10.1007/s12562-014-0711-4
120. Yokobori E, Azuma M, Nishiguchi R, Kang KS, Kamijo M, Uchiyama M, et al. Neuropeptide Y stimulates food intake in the Zebrafish, Danio rerio. J Neuroendocrinol. (2012) 24:766–73. doi: 10.1111/j.1365-2826.2012.02281.x
121. Jørgensen EH, Bernier NJ, Maule AG, Vijayan MM. Effect of long-term fasting and a subsequent meal on mRNA abundances of hypothalamic appetite regulators, central and peripheral leptin expression and plasma leptin levels in rainbow trout. Peptides. (2016) 86:162–70. doi: 10.1016/j.peptides.2015.08.010
122. Wei R, Yuan D, Wang T, Zhou C, Lin F, Chen H, et al. Characterization, tissue distribution and regulation of agouti-related protein (AgRP) in a cyprinid fish (Schizothorax prenanti). Gene. (2013) 527:193–200. doi: 10.1016/j.gene.2013.06.003
123. Agulleiro MJ, Cortés R, Leal E, Ríos D, Sánchez E, Cerdá-Reverter JM. Characterization, tissue distribution and regulation by fasting of the agouti family of peptides in the sea bass (Dicentrarchus labrax). Gen Comp Endocrinol. (2014) 205:251–9. doi: 10.1016/j.ygcen.2014.02.009
124. Wan Y, Zhang Y, Ji P, Li Y, Xu P, Sun X. Molecular characterization of CART, AgRP, and MC4R genes and their expression with fasting and re-feeding in common carp (Cyprinus carpio). Mol Biol Rep. (2012) 39:2215–23. doi: 10.1007/s11033-011-0970-4
125. Song Y, Golling G, Thacker TL, Cone RD. Agouti-related protein (AGRP) is conserved and regulated by metabolic state in the zebrafish, Danio rerio. Endocrine (2003) 22:257–65. doi: 10.1385/ENDO:22:3:257
126. Kang D-Y, Kim H-C. Functional relevance of three proopiomelanocortin (POMC) genes in darkening camouflage, blind-side hypermelanosis, and appetite of Paralichthys olivaceus. Comp Biochem Physiol B Biochem Mol Biol. (2015) 179:44–56. doi: 10.1016/j.cbpb.2014.09.002
127. Cerdá-Reverter JM, Schiöth HB, Peter RE. The central melanocortin system regulates food intake in goldfish. Regul Pept. (2003) 115:101–13. doi: 10.1016/S0167-0115(03)00144-7
128. Leder EH, Silverstein JT. The pro-opiomelanocortin genes in rainbow trout (Oncorhynchus mykiss): duplications, splice variants, and differential expression. J Endocrinol. (2006) 188:355–63. doi: 10.1677/joe.1.06283
129. Liu S, Zhang C, Peng G. Effects of starvation on the expression of feeding related neuropeptides in the larval zebrafish hypothalamus. Yi Chuan Hered. (2016) 38:821–30. doi: 10.16288/j.yczz.16-087
130. Subhedar N, Barsagade VG, Singru PS, Thim L, Clausen JT. Cocaine- and amphetamine-regulated transcript peptide (CART) in the telencephalon of the catfish, Clarias gariepinus: distribution and response to fasting, 2-deoxy-D-glucose, glucose, insulin, and leptin treatments. J Comp Neurol. (2011) 519:1281–300. doi: 10.1002/cne.22569
131. Nishio S-I, Gibert Y, Berekelya L, Bernard L, Brunet F, Guillot E, et al. Fasting induces CART down-regulation in the zebrafish nervous system in a cannabinoid receptor 1-dependent manner. Mol Endocrinol Baltim Md. (2012) 26:1316–26. doi: 10.1210/me.2011-1180
132. Kehoe AS, Volkoff H. Cloning and characterization of neuropeptide Y (NPY) and cocaine and amphetamine regulated transcript (CART) in Atlantic cod (Gadus morhua). Comp Biochem Physiol A Mol Integr Physiol. (2007) 146:451–61. doi: 10.1016/j.cbpa.2006.12.026
133. Kobayashi Y, Peterson BC, Waldbieser GC. Association of cocaine- and amphetamine-regulated transcript (CART) messenger RNA level, food intake, and growth in channel catfish. Comp Biochem Physiol A Mol Integr Physiol. (2008) 151:219–25. doi: 10.1016/j.cbpa.2008.06.029
134. Abbott M, Volkoff H. Thyrotropin Releasing Hormone (TRH) in goldfish (Carassius auratus): role in the regulation of feeding and locomotor behaviors and interactions with the orexin system and cocaine- and amphetamine regulated transcript (CART). Horm Behav. (2011) 59:236–45. doi: 10.1016/j.yhbeh.2010.12.008
135. Volkoff H. Appetite regulating peptides in red-bellied piranha, Pygocentrus nattereri: cloning, tissue distribution and effect of fasting on mRNA expression levels. Peptides. (2014) 56:116–24. doi: 10.1016/j.peptides.2014.03.022
136. Novak CM, Jiang X, Wang C, Teske JA, Kotz CM, Levine JA. Caloric restriction and physical activity in zebrafish (Danio rerio). Neurosci Lett. (2005) 383:99–104. doi: 10.1016/j.neulet.2005.03.048
137. Yokobori E, Kojima K, Azuma M, Kang KS, Maejima S, Uchiyama M, et al. Stimulatory effect of intracerebroventricular administration of orexin A on food intake in the zebrafish, Danio rerio. Peptides (2011) 32:1357–62. doi: 10.1016/j.peptides.2011.05.010
138. Kang D-Y, Kim H-C. Functional characterization of two melanin-concentrating hormone genes in the color camouflage, hypermelanosis, and appetite of starry flounder. Gen Comp Endocrinol. (2013) 189:74–83. doi: 10.1016/j.ygcen.2013.04.025
139. Tuziak SM, Volkoff H. Melanin-concentrating hormone (MCH) and gonadotropin-releasing hormones (GnRH) in Atlantic cod, Gadus morhua: tissue distributions, early ontogeny and effects of fasting. Peptides. (2013) 50:109–18. doi: 10.1016/j.peptides.2013.10.005
140. Takahashi A, Tsuchiya K, Yamanome T, Amano M, Yasuda A, Yamamori K, et al. Possible involvement of melanin-concentrating hormone in food intake in a teleost fish, barfin flounder. Peptides. (2004) 25:1613–22. doi: 10.1016/j.peptides.2004.02.022
141. Mizusawa K, Amiya N, Yamaguchi Y, Takabe S, Amano M, Breves JP, et al. Identification of mRNAs coding for mammalian-type melanin-concentrating hormone and its receptors in the scalloped hammerhead shark Sphyrna lewini. Gen Comp Endocrinol. (2012) 179:78–87. doi: 10.1016/j.ygcen.2012.07.023
142. Wang T, Yuan D, Zhou C, Lin F, Wei R, Chen H, et al. Molecular characterization of melanin-concentrating hormone (MCH) in Schizothorax prenanti: cloning, tissue distribution and role in food intake regulation. Fish Physiol Biochem. (2016) 42:883–93. doi: 10.1007/s10695-015-0182-2
143. Tuziak SM, Volkoff H. A preliminary investigation of the role of melanin-concentrating hormone (MCH) and its receptors in appetite regulation of winter flounder (Pseudopleuronectes americanus). Mol Cell Endocrinol. (2012) 348:281–96. doi: 10.1016/j.mce.2011.09.015
144. Berman JR, Skariah G, Maro GS, Mignot E, Mourrain P. Characterization of two melanin-concentrating hormone genes in zebrafish reveals evolutionary and physiological links with the mammalian MCH system. J Comp Neurol. (2009) 517:695–710. doi: 10.1002/cne.22171
145. Hevrøy EM, Azpeleta C, Shimizu M, Lanzén A, Kaiya H, Espe M, et al. Effects of short-term starvation on ghrelin, GH-IGF system, and IGF-binding proteins in Atlantic salmon. Fish Physiol Biochem. (2011) 37:217–32. doi: 10.1007/s10695-010-9434-3
146. Feng K, Zhang G-R, Wei K-J, Xiong B-X. Molecular cloning, tissue distribution, and ontogenetic expression of ghrelin and regulation of expression by fasting and refeeding in the grass carp (Ctenopharyngodon idellus). J Exp Zool Part Ecol Genet Physiol. (2013) 319:202–12. doi: 10.1002/jez.1784
147. Eom J, Hong M, Cone RD, Song Y. Zebrafish ghrelin is expressed in pancreatic endocrine cells and regulated by metabolic state. Biochem Biophys Res Commun. (2013) 439:115–20. doi: 10.1016/j.bbrc.2013.08.017
148. Amole N, Unniappan S. Fasting induces preproghrelin mRNA expression in the brain and gut of zebrafish, Danio rerio. Gen Comp Endocrinol. (2009) 161:133–7. doi: 10.1016/j.ygcen.2008.11.002
149. Murashita K, Kurokawa T, Nilsen TO, Rønnestad I. Ghrelin, cholecystokinin, and peptide YY in Atlantic salmon (Salmo salar): molecular cloning and tissue expression. Gen Comp Endocrinol. (2009) 160:223–35. doi: 10.1016/j.ygcen.2008.11.024
150. Song Y, Zhao C, Liang X-F, He S, Tian C, Cheng X, et al. Effects of fasting, temperature, and photoperiod on preproghrelin mRNA expression in Chinese perch. Fish Physiol Biochem. (2017) 43:803–12. doi: 10.1007/s10695-016-0335-y
151. Zhou C, Zhang X, Liu T, Wei R, Yuan D, Wang T, et al. Schizothorax davidi ghrelin: cDNA cloning, tissue distribution and indication for its stimulatory character in food intake. Gene. (2014) 534:72–7. doi: 10.1016/j.gene.2013.10.012
152. Terova G, Rimoldi S, Bernardini G, Gornati R, Saroglia M. Sea bass ghrelin: molecular cloning and mRNA quantification during fasting and refeeding. Gen Comp Endocrinol. (2008) 155:341–51. doi: 10.1016/j.ygcen.2007.05.028
153. Koven W, Schulte P. The effect of fasting and refeeding on mRNA expression of PepT1 and gastrointestinal hormones regulating digestion and food intake in zebrafish (Danio rerio). Fish Physiol Biochem. (2012) 38:1565–75. doi: 10.1007/s10695-012-9649-6
154. Picha ME, Strom CN, Riley LG, Walker AA, Won ET, Johnstone WM, et al. Plasma ghrelin and growth hormone regulation in response to metabolic state in hybrid striped bass: effects of feeding, ghrelin and insulin-like growth factor-I on in vivo and in vitro GH secretion. Gen Comp Endocrinol. (2009) 161:365–72. doi: 10.1016/j.ygcen.2009.01.026
155. Riley LG, Fox BK, Breves JP, Kaiya H, Dorough CP, Hirano T, et al. Absence of effects of short-term fasting on plasma ghrelin and brain expression of ghrelin receptors in the tilapia, Oreochromis mossambicus. Zoolog Sci. (2008) 25:821–7. doi: 10.2108/zsj.25.821
156. Matsuda K, Miura T, Kaiya H, Maruyama K, Uchiyama M, Kangawa K, et al. Stimulatory effect of n-octanoylated ghrelin on locomotor activity in the goldfish, Carassius auratus. Peptides. (2006) 27:1335–40. doi: 10.1016/j.peptides.2005.10.011
157. Volkoff H. Cloning, tissue distribution and effects of fasting on mRNA expression levels of leptin and ghrelin in red-bellied piranha (Pygocentrus nattereri). Gen Comp Endocrinol. (2015) 217–218:20–27. doi: 10.1016/j.ygcen.2015.05.004
158. Feng K, Zhang G, Wei K, Xiong B, Liang T, Ping H. Molecular characterization of cholecystokinin in grass carp (Ctenopharyngodon idellus): cloning, localization, developmental profile, and effect of fasting and refeeding on expression in the brain and intestine. Fish Physiol Biochem. (2012) 38:1825–34. doi: 10.1007/s10695-012-9679-0
159. Micale V, Campo S, D'Ascola A, Guerrera MC, Levanti MB, German,à A, et al. Cholecystokinin in white sea bream: molecular cloning, regional expression, and immunohistochemical localization in the gut after feeding and fasting. PLoS ONE. (2012) 7:e52428. doi: 10.1371/journal.pone.0052428
160. Murashita K, Fukada H, Hosokawa H, Masumoto T. Cholecystokinin and peptide Y in yellowtail (Seriola quinqueradiata): molecular cloning, real-time quantitative RT-PCR, and response to feeding and fasting. Gen Comp Endocrinol. (2006) 145:287–97. doi: 10.1016/j.ygcen.2005.09.008
161. Huising MO, Geven EJW, Kruiswijk CP, Nabuurs SB, Stolte EH, Spanings FAT, et al. Increased leptin expression in common Carp (Cyprinus carpio) after food intake but not after fasting or feeding to satiation. Endocrinology. (2006) 147:5786–97. doi: 10.1210/en.2006-0824
162. Fuentes EN, Kling P, Einarsdottir IE, Alvarez M, Valdés JA, Molina A, et al. Plasma leptin and growth hormone levels in the fine flounder (Paralichthys adspersus) increase gradually during fasting and decline rapidly after refeeding. Gen Comp Endocrinol. (2012) 177:120–7. doi: 10.1016/j.ygcen.2012.02.019
163. Fuentes EN, Safian D, Einarsdottir IE, Valdés JA, Elorza AA, Molina A, et al. Nutritional status modulates plasma leptin, AMPK and TOR activation, and mitochondrial biogenesis: implications for cell metabolism and growth in skeletal muscle of the fine flounder. Gen Comp Endocrinol. (2013) 186:172–80. doi: 10.1016/j.ygcen.2013.02.009
164. Kling P, Rønnestad I, Stefansson SO, Murashita K, Kurokawa T, Björnsson BT. A homologous salmonid leptin radioimmunoassay indicates elevated plasma leptin levels during fasting of rainbow trout. Gen Comp Endocrinol. (2009) 162:307–12. doi: 10.1016/j.ygcen.2009.04.003
165. Zhang H, Chen H, Zhang Y, Li S, Lu D, Zhang H, et al. Molecular cloning, characterization and expression profiles of multiple leptin genes and a leptin receptor gene in orange-spotted grouper (Epinephelus coioides). Gen Comp Endocrinol. (2013) 181:295–305. doi: 10.1016/j.ygcen.2012.09.008
166. Shpilman M, Hollander-Cohen L, Ventura T, Gertler A, Levavi-Sivan B. Production, gene structure and characterization of two orthologs of leptin and a leptin receptor in tilapia. Gen Comp Endocrinol. (2014) 207:74–85. doi: 10.1016/j.ygcen.2014.05.006
167. Gambardella C, Gallus L, Amaroli A, Terova G, Masini MA, Ferrando S. Fasting and re-feeding impact on leptin and aquaglyceroporin 9 in the liver of European sea bass (Dicentrarchus labrax). Aquaculture. (2012) 354–5:1–6. doi: 10.1016/j.aquaculture.2012.04.043
168. Johnson RM, Johnson TM, Londraville RL. Evidence for leptin expression in fishes. J Exp Zool. (2000) 286:718–24. doi: 10.1002/(SICI)1097-010X(20000601)286:7<718::AID-JEZ6>3.0.CO;2-I
169. Johansson M, Morgenroth D, Einarsdottir IE, Gong N, Björnsson BT. Energy stores, lipid mobilization and leptin endocrinology of rainbow trout. J Comp Physiol B. (2016) 186:759–73. doi: 10.1007/s00360-016-0988-y
170. Won ET, Baltzegar DA, Picha ME, Borski RJ. Cloning and characterization of leptin in a Perciform fish, the striped bass (Morone saxatilis): control of feeding and regulation by nutritional state. Gen Comp Endocrinol. (2012) 178:98–107. doi: 10.1016/j.ygcen.2012.04.019
171. Chen T, Chen S, Ren C, Hu C, Tang D, Yan A. Two isoforms of leptin in the White-clouds Mountain minnow (Tanichthys albonubes): differential regulation by estrogen despite similar response to fasting. Gen Comp Endocrinol. (2016) 225:174–84. doi: 10.1016/j.ygcen.2015.08.002
172. Gorissen M, Bernier NJ, Nabuurs SB, Flik G, Huising MO. Two divergent leptin paralogues in zebrafish (Danio rerio) that originate early in teleostean evolution. J Endocrinol. (2009) 201:329–39. doi: 10.1677/JOE-09-0034
173. Jørgensen EH, Martinsen M, Strøm V, Hansen KER, Ravuri CS, Gong N, et al. Long-term fasting in the anadromous Arctic charr is associated with downregulation of metabolic enzyme activity and upregulation of leptin A1 and SOCS expression in the liver. J Exp Biol. (2013) 216:3222–30. doi: 10.1242/jeb.088344
174. Morini M, Pasquier J, Dirks R, van den Thillart G, Tomkiewicz J, Rousseau K, et al. Duplicated leptin receptors in two species of eel bring new insights into the evolution of the leptin system in vertebrates. PLoS ONE. (2015) 10:e0126008. doi: 10.1371/journal.pone.0126008
175. Narnaware YK, Peter RE. Influence of diet composition on food intake and neuropeptide Y (NPY) gene expression in goldfish brain. Regul Pept. (2002) 103:75–83. doi: 10.1016/S0167-0115(01)00342-1
176. Blanco AM, Bertucci JI, Delgado MJ, Valenciano AI, Unniappan S. Tissue-specific expression of ghrelinergic and NUCB2/nesfatin-1 systems in goldfish (Carassius auratus) is modulated by macronutrient composition of diets. Comp Biochem Physiol A Mol Integr Physiol. (2016) 195:1–9. doi: 10.1016/j.cbpa.2016.01.016
177. Bertucci JI, Blanco AM, Canosa LF, Unniappan S. Glucose, amino acids and fatty acids directly regulate ghrelin and NUCB2/nesfatin-1 in the intestine and hepatopancreas of goldfish (Carassius auratus) in vitro. Comp Biochem Physiol A Mol Integr Physiol. (2017) 206:24–35. doi: 10.1016/j.cbpa.2017.01.006
178. Babaei S, Sáez A, Caballero-Solares A, Fernández F, Baanante IV, Metón I. Effect of dietary macronutrients on the expression of cholecystokinin, leptin, ghrelin and neuropeptide Y in gilthead sea bream (Sparus aurata). Gen Comp Endocrinol. (2017) 240:121–8. doi: 10.1016/j.ygcen.2016.10.003
179. Bertucci JI, Tovar MO, Unniappan S, Navarro JC, Canosa LF. Effects of dietary sunflower oil on growth parameters, fatty acid profiles and expression of genes regulating growth and metabolism in the pejerrey (Odontesthes bonariensis) fry. Aquac. Nutr. (2017) 24:748–57. doi: 10.1111/anu.12603
180. Bonacic K, Campoverde C, Gómez-Arbonés J, Gisbert E, Estevez A, Morais S. Dietary fatty acid composition affects food intake and gut-brain satiety signaling in Senegalese sole (Solea senegalensis, Kaup 1858) larvae and post-larvae. Gen Comp Endocrinol. (2016) 228:79–94. doi: 10.1016/j.ygcen.2016.02.002
181. Moriyama S, Ayson FG, Kawauchi H. Growth regulation by insulin-like growth factor-I in fish. Biosci Biotechnol Biochem. (2000) 64:1553–62. doi: 10.1271/bbb.64.1553
182. Pierce AL, Shimizu M, Beckman BR, Baker DM, Dickhoff WW. Time course of the GH/IGF axis response to fasting and increased ration in chinook salmon (Oncorhynchus tshawytscha). Gen Comp Endocrinol. (2005) 140:192–202. doi: 10.1016/j.ygcen.2004.10.017
183. Small BC, Peterson BC. Establishment of a time-resolved fluoroimmunoassay for measuring plasma insulin-like growth factor I (IGF-I) in fish: effect of fasting on plasma concentrations and tissue mRNA expression of IGF-I and growth hormone (GH) in channel catfish (Ictalurus punctatus). Domest Anim Endocrinol. (2005) 28:202–15. doi: 10.1016/j.domaniend.2004.09.002
184. Shimizu M, Cooper KA, Dickhoff WW, Beckman BR. Postprandial changes in plasma growth hormone, insulin, insulin-like growth factor (IGF)-I, and IGF-binding proteins in coho salmon fasted for varying periods. Am J Physiol Regul Integr Comp Physiol. (2009) 297:R352–361. doi: 10.1152/ajpregu.90939.2008
185. Cleveland BM, Weber GM, Blemings KP, Silverstein JT. Insulin-like growth factor-I and genetic effects on indexes of protein degradation in response to feed deprivation in rainbow trout (Oncorhynchus mykiss). Am J Physiol Regul Integr Comp Physiol. (2009) 297:R1332–42. doi: 10.1152/ajpregu.00272.2009
186. Farbridge KJ, Leatherland JF. Temporal changes in plasma thyroid hormone, growth hormone and free fatty acid concentrations, and hepatic 5'-monodeiodinase activity, lipid and protein content during chronic fasting and re-feeding in rainbow trout (Oncorhynchus mykiss). Fish Physiol Biochem. (1992) 10:245–57. doi: 10.1007/BF00004518
187. Johnsson JI, Jönsson E, Björnsson BT. Dominance, nutritional state, and growth hormone levels in rainbow trout (Oncorhynchus mykiss). Horm Behav. (1996) 30:13–21. doi: 10.1006/hbeh.1996.0003
188. Fox BK, Riley LG, Hirano T, Grau EG. Effects of fasting on growth hormone, growth hormone receptor, and insulin-like growth factor-I axis in seawater-acclimated tilapia, Oreochromis mossambicus. Gen Comp Endocrinol. (2006) 148:340–7. doi: 10.1016/j.ygcen.2006.04.007
189. Uchida K, Kajimura S, Riley LG, Hirano T, Aida K, Grau EG. Effects of fasting on growth hormone/insulin-like growth factor I axis in the tilapia, Oreochromis mossambicus. Comp Biochem Physiol A Mol Integr Physiol. (2003) 134:429–39. doi: 10.1016/S1095-6433(02)00318-5
190. Delgadin TH, Pérez Sirkin DI, Di Yorio MP, Arranz SE, Vissio PG. GH, IGF-I and GH receptors mRNA expression in response to growth impairment following a food deprivation period in individually housed cichlid fish Cichlasoma dimerus. Fish Physiol Biochem. (2015) 41:51–60. doi: 10.1007/s10695-014-0005-x
191. Zhong H, Zhou Y, Liu S, Tao M, Long Y, Liu Z, et al. Elevated expressions of GH/IGF axis genes in triploid crucian carp. Gen Comp Endocrinol. (2012) 178:291–300. doi: 10.1016/j.ygcen.2012.06.006
192. Pedroso FL, de Jesus-Ayson EGT, Cortado HH, Hyodo S, Ayson FG. Changes in mRNA expression of grouper (Epinephelus coioides) growth hormone and insulin-like growth factor I in response to nutritional status. Gen Comp Endocrinol. (2006) 145:237–46. doi: 10.1016/j.ygcen.2005.09.001
193. Fox BK, Breves JP, Davis LK, Pierce AL, Hirano T, Grau EG. Tissue-specific regulation of the growth hormone/insulin-like growth factor axis during fasting and re-feeding: Importance of muscle expression of IGF-I and IGF-II mRNA in the tilapia. Gen Comp Endocrinol. (2010) 166:573–80. doi: 10.1016/j.ygcen.2009.11.012
194. Fukada H, Murashita K, Furutani T, Masumoto T. Yellowtail insulin-like growth factor 1: molecular cloning and response to various nutritional conditions. Domest Anim Endocrinol. (2012) 42:220–9. doi: 10.1016/j.domaniend.2011.12.005
195. Baños N, Planas JV, Gutiérrez J, Navarro I. Regulation of plasma insulin-like growth factor-I levels in brown trout (Salmo trutta). Comp Biochem Physiol C Pharmacol Toxicol Endocrinol. (1999) 124:33–40. doi: 10.1016/S0742-8413(99)00044-4
196. Kawaguchi K, Kaneko N, Fukuda M, Nakano Y, Kimura S, Hara A, et al. Responses of insulin-like growth factor (IGF)-I and two IGF-binding protein-1 subtypes to fasting and re-feeding, and their relationships with individual growth rates in yearling masu salmon (Oncorhynchus masou). Comp Biochem Physiol A Mol Integr Physiol. (2013) 165:191–8. doi: 10.1016/j.cbpa.2013.02.029
197. Pérez-Sánchez J, Le Bail P-Y. Growth hormone axis as marker of nutritional status and growth performance in fish. Aquaculture. (1999) 177:117–28. doi: 10.1016/S0044-8486(99)00073-3
198. Dyer AR, Barlow CG, Bransden MP, Carter CG, Glencross BD, Richardson N, et al. Correlation of plasma IGF-I concentrations and growth rate in aquacultured finfish: a tool for assessing the potential of new diets. Aquaculture. (2004) 236:583–92. doi: 10.1016/j.aquaculture.2003.12.025
199. Pérez-Sánchez J, Martí-Palanca H, Kaushik SJ. Nutrient requirements and interactions ration size and protein intake affect circulating growth hormone concentration , hepatic growth hormone binding and plasma insulin-like growth factor-I immunoreactivity in a marine teleost, the gilthead sea bream (Sparus aurata). J. Nutr. (1995) 546–52.
200. Picha ME, Turano MJ, Beckman BR, Borski RJ. Endocrine biomarkers of growth and applications to aquaculture: a minireview of growth hormone, insulin-like growth factor (IGF)-I, and IGF-binding proteins as potential growth indicators in fish. N Am J Aquac. (2008) 70:196–211. doi: 10.1577/A07-038.1
201. Rodgers BD, Helms LM, Grau EG. Effects of fasting, medium glucose, and amino acid concentrations on prolactin and growth hormone release, in vitro, from the pituitary of the tilapia Oreochromis mossambicus. Gen Comp Endocrinol. (1992) 86:344–351.
202. Riley LG, Walker AP, Dorough CP, Schwandt SE, Grau EG. Glucose regulates ghrelin, neuropeptide Y, and the GH/IGF-I axis in the tilapia, Oreochromis mossambicus. Comp Biochem Physiol A Mol Integr Physiol. (2009) 154:541–6. doi: 10.1016/j.cbpa.2009.08.018
203. Bertucci JI, Blanco AM, Canosa LF. Direct actions of macronutrient components on goldfish hepatopancreas in vitro to modulate the expression of ghr-I, ghr-II, igf-I and igf-II mRNAs. Gen Comp Endocrinol. (2017) 250:1–8. doi: 10.1016/j.ygcen.2017.05.014
204. Yakar S, Setser J, Zhao H, Stannard B, Haluzik M, Glatt V, et al. Inhibition of growth hormone action improves insulin sensitivity in liver IGF-1 –deficient mice. J Clin Invest. (2004) 113:96. doi: 10.1172/JCI200417763.Introduction
205. Wood AW, Duan C, Bern HA. Insulin-like growth factor signaling in fish. Int Rev Cytol. (2005) 243:215–85. doi: 10.1016/S0074-7696(05)43004-1
206. Ekström K, Pulkkinen MA, Carlsson-Skwirut C, Brorsson AL, Ma Z, Frystyk J, et al. Tissue IGF-I measured by microdialysis reflects body glucose utilization after rhIGF-I injection in type 1 diabetes. J Clin Endocrinol Metab. (2015) 100:4299–306. doi: 10.1210/jc.2015-2070
207. Baumann MU, Schneider H, Malek A, Palta V, Surbek DV, Sager R, et al. Regulation of human trophoblast GLUT1 glucose transporter by Insulin-Like Growth Factor I (IGF-I). PLoS ONE. (2014) 9:e106037. doi: 10.1371/journal.pone.0106037
208. Sangiao-Alvarellos S, Míguez JM, Soengas JL. Actions of growth hormone on carbohydrate metabolism and osmoregulation of rainbow trout (Oncorhynchus mykiss). Gen Comp Endocrinol. (2005) 141:214–25. doi: 10.1016/j.ygcen.2005.01.007
209. Huang JF, Xu QY, Chang YM. Effects of temperature and dietary protein on the growth performance and IGF-I mRNA expression of juvenile mirror carp (Cyprinus carpio). Aquac Nutr. (2016) 22:283–92. doi: 10.1111/anu.12254
210. Qiang J, Yang H, Wang H, Kpundeh MD, Xu P. Growth and IGF-I response of juvenile Nile tilapia (Oreochromis niloticus) to changes in water temperature and dietary protein level. J Therm Biol. (2012) 37:686–95. doi: 10.1016/j.jtherbio.2012.07.009
211. Tu Y, Xie S, Han D, Yang Y, Jin J, Liu H, et al. Growth performance, digestive enzyme, transaminase and GH-IGF-I axis gene responsiveness to different dietary protein levels in broodstock allogenogynetic gibel carp (Carassius auratus gibelio) CAS III. Aquaculture. (2015) 446:290–7. doi: 10.1016/j.aquaculture.2015.05.003
212. Borges P, Oliveira B, Casal S, Dias J, Conceição L, Valente LMP. Dietary lipid level affects growth performance and nutrient utilisation of Senegalese sole (Solea senegalensis) juveniles. Br J Nutr. (2009) 102:1007–14. doi: 10.1017/S0007114509345262
213. Regost C, Arzel J, Cardinal M, Robin J, Laroche M, Kaushik SJ. Dietary lipid level, hepatic lipogenesis and flesh quality. Nutrition. (2001) 193:291–309. doi: 10.1016/S0044-8486(00)00493-2
214. Lee S-M, Cho SH, Kim K-D. Effects of dietary protein and energy levels on growth and body composition of juvenile flounder Paralichthys olivaceus. J World Aquac Soc. (2000) 31:306–315. doi: 10.1111/j.1749-7345.2000.tb00882.x
215. Campos C, Valente LMP, Borges P, Bizuayehu T, Fernandes JMO. Dietary lipid levels have a remarkable impact on the expression of growth-related genes in Senegalese sole (Solea senegalensis Kaup). J Exp Biol. (2010) 213:200–9. doi: 10.1242/jeb.033126
216. Gómez-Requeni P, Kraemer MN, Canosa LF. Regulation of somatic growth and gene expression of the GH-IGF system and PRP-PACAP by dietary lipid level in early juveniles of a teleost fish, the pejerrey (Odontesthes bonariensis). J Comp Physiol B. (2012) 182:517–30. doi: 10.1007/s00360-011-0640-9
217. Chen N, Jin L, Zhou H, Qiu X. Effects of dietary arginine levels and carbohydrate-to-lipid ratios on mRNA expression of growth-related hormones in largemouth bass, Micropterus salmoides. Gen Comp Endocrinol. (2012) 179:121–7. doi: 10.1016/j.ygcen.2012.08.004
218. Benedito-Palos L, Saera-Vila A, Calduch-Giner JA, Kaushik S, Pérez-Sánchez J. Combined replacement of fish meal and oil in practical diets for fast growing juveniles of gilthead sea bream (Sparus aurata L.): networking of systemic and local components of GH/IGF axis. Aquaculture. (2007) 267:199–212. doi: 10.1016/j.aquaculture.2007.01.011
219. Mazurais D, Darias MJ, Gouillou-Coustans MF, Le Gall MM, Huelvan C, Desbruyères E, et al. Dietary vitamin mix levels influence the ossification process in European sea bass (Dicentrarchus labrax) larvae. Am J Physiol Regul Integr Comp Physiol. (2008) 294:R520–7. doi: 10.1152/ajpregu.00659.2007
220. Bergan-Roller HE, Sheridan MA. The growth hormone signaling system: insights into coordinating the anabolic and catabolic actions of growth hormone. Gen Comp Endocrinol. (2017) 258:119–33. doi: 10.1016/j.ygcen.2017.07.028
221. Unniappan S, Lin X, Cervini L, Rivier J, Kaiya H, Kangawa K, et al. Goldfish ghrelin: Molecular characterization of the complementary deoxyribonucleic acid, partial gene structure and evidence for its stimulatory role in food intake. Endocrinology. (2002) 143:4143–6. doi: 10.1210/en.2002-220644
222. Unniappan S, Peter RE. In vitro and in vivo effects of ghrelin on luteinizing hormone and growth hormone release in goldfish. Am J Physiol Regul Integr Comp Physiol. (2004) 286:R1093–101. doi: 10.1152/ajpregu.00669.2003
223. Grey CL, Chang JP. Ghrelin-induced growth hormone release from goldfish pituitary cells is nitric oxide dependent. Gen Comp Endocrinol. (2012) 179:152–8. doi: 10.1016/j.ygcen.2012.08.012
Keywords: food intake, growth, nutrients, fish, aquaculture
Citation: Bertucci JI, Blanco AM, Sundarrajan L, Rajeswari JJ, Velasco C and Unniappan S (2019) Nutrient Regulation of Endocrine Factors Influencing Feeding and Growth in Fish. Front. Endocrinol. 10:83. doi: 10.3389/fendo.2019.00083
Received: 16 September 2018; Accepted: 30 January 2019;
Published: 28 February 2019.
Edited by:
Encarnación Capilla, University of Barcelona, SpainReviewed by:
Stephane Panserat, Institut National de la Recherche Agronomique (INRA), FranceElisabeth Jönsson Bergman, University of Gothenburg, Sweden
Copyright © 2019 Bertucci, Blanco, Sundarrajan, Rajeswari, Velasco and Unniappan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Suraj Unniappan, c3VyYWoudW5uaWFwcGFuQHVzYXNrLmNh