 Zishan Ahmad Wani
Zishan Ahmad Wani Javid Ahmad Dar
Javid Ahmad Dar Aamir Nazir Lone1
Aamir Nazir Lone1 Shreekar Pant
Shreekar Pant- 1Terrestrial Ecology and Modelling (TEaM) Lab, Department of Environmental Science and Engineering, SRM University-AP, Amaravati, Andhra Pradesh, India
- 2Centre for Geospatial Technology, SRM University-AP, Amaravati, Andhra Pradesh, India
- 3Centre for Biodiversity Studies, Baba Ghulam Shah Badshah University, Rajouri, Jammu and Kashmir, India
- 4Department of Biology, College of Science, King Khalid University, Abha, Saudi Arabia
Prioritizing native and endemic species for conservation is fundamental to achieve broader objectives of safeguarding biodiversity, as these species are vulnerable to extinction risks. Forecasting the climatic niche of these species through species distribution models can be crucial for their habitat conservation and sustainable management in future. In this study, an ensemble modelling approach was used to predict the distribution of Bergenia stracheyi, a native alpine plant species of Himalayan region. The results revealed that the distribution of B. stracheyi is primarily influenced by Annual Mean Temperature (Bio1) and Annual Precipitation (Bio12). Ensemble model predictions revealed that under the current climatic conditions, the suitable habitats for B. stracheyi are distributed across higher elevations of Jammu and Kashmir and future ensemble model predictions indicate that, across all future climatic scenarios, the majority of the currently suitable habitats will remain suitable for the species. The model predicts a significant expansion in suitable habitats for B. stracheyi, particularly under more severe climate change scenarios (RCP8.5). However, some areas currently identified as suitable, including parts of the Pir Panjal range and Mirpur (Pakistan), are projected to become unsuitable for the species in the future. These shifts in plant distribution may have far-reaching consequences for ecosystem functioning and stability and the services provided to human communities. Additionally, these shifts may lead to mismatches between the plant phenological events and pollinators potentially causing more ecological disruptions. Thus, the predicted range shifts in the distribution of B. stracheyi highlight the importance of local conservation measures to mitigate the impacts of climate change.
1 Introduction
Global biodiversity is declining at an unprecedented rate and with nearly a million plant and animal species at risk of extinction—a threat projected to worsen over the next few decades (Turnhout and Purvis, 2020; Palombo, 2021). This decline in biodiversity results from both anthropogenic and natural causes. Among various drivers, climate change is perceived as one of the most severe threats to biodiversity (Ripple et al., 2017; Arneth et al., 2020; Roman-Palacios and Wiens, 2020; Verrall and Pickering, 2020; Wani et al., 2024a). In the modern anthropocene era, climate shifts pose a serious threat to the overall functioning and stability of ecosystems (Wan et al., 2016) and have long-term effects, leading to ecological niche shifts in various plant species (Lenoir et al., 2020). Changes in regional climate such as alteration in precipitation patterns and humidity, and reduced snow cover significantly affect biodiversity of that region as climate variables primarily govern the geographic range of species distributions (Weiskopf et al., 2020; Pepin et al., 2022; Shakoor et al., 2023; Wang et al., 2024). While several species suffer from climate change, but some are able to adapt to climate change by moving to more favorable locations (Vitasse et al., 2021; Manes et al., 2021a) or by utilizing adaptive genetic systems or phenotypic plasticity to withstand environmental changes (Anderson and Song, 2020; Pazzaglia et al., 2021). The distributional ranges of species are altered due to their cumulative adaptive responses to climate change (Taheri et al., 2021). The steadily rising concentration of greenhouse gases threatens the survival of various plant species, and disrupts the stability and functionality of ecosystems worldwide (Kumar et al., 2024; Chaudhry and Sidhu, 2022; Shrestha et al., 2022; Zandalinas et al., 2022). However, the Himalayan biodiversity hotspot is particularly vulnerable to climate change with three-fold faster warming than the global average (Shrestha et al., 2012). Climate is expected to affect the growth, phenology, and distribution of Himalayan plants, posing significant challenges to their survival in the future (Sarkar et al., 2024; Rana et al., 2024; Mishra et al., 2024). Further, climate change is closely linked to shifts in species distribution (Permesan, 2006; Sekar et al., 2024), thus highlighting the urgent need to understand the dynamics of species distribution and ecological niches (Javeed et al., 2024). Furthermore, under these critical scenarios prioritizing native and endemic species for conservation is essential to achieving broader biodiversity conservation goals, as these species are vulnerable to extinction (Manes et al., 2021b). Thus, forecasting the climatic niche of native and endemic species using species distribution models (SDMs) is crucial for habitat conservation and the sustainable management of species in the future (Profirio et al., 2014).
Species distribution modelling (SDM) is a useful technique for predicting range shifts and identifying habitats suitable for species conservation in response to climate change (Zellmer et al., 2019). SDM has significant potential to support adaptive conservation initiatives and ultimately contribute to biodiversity conservation (Guisan et al., 2013). SDM is an emerging field in ecology for predicting species distributions and has gained increasing importance in the context of growing awareness of environmental change and its ecological consequences (Miller, 2010). It allows for the assessment of a wide range of biodiversity phenomena, including potential geographic distributions, future distributions under climate change scenarios, species invasions, crop damage by pest organisms, biodiversity conservation priorities, and species range shifts (Javeed et al., 2024). Various algorithms are used in SDM typically integrating the species occurrence data and environmental variables to identify suitable habitats (Kaky et al., 2020; Frans et al., 2022). Endemic species have a limited geographic range, making them more vulnerable to extinction due to natural and anthropogenic drivers (Kraus et al., 2023; Wani et al., 2023). Given this susceptibility of endemic plants, information on population status, distribution range and threats has significant conservation implications (Abro et al., 2024). It is therefore imperative to understand the distribution of endemic species and SDMs are increasing recognized as a valuable tool to predict suitable habitats under current and future climatic scenarios (Qazi et al., 2022). In the present study, an ensemble of multiple algorithms was used to predict the distribution of Bergenia stracheyi, a native alpine plant species of the Himalayan region. The present study aims to: a) predict the current distribution of B. stracheyi using an ensemble modelling approach; b) predict the distribution of B. stracheyi under changing climatic scenarios; c) assess the range shifts in the distribution of B. stracheyi under anticipated climate change; d) identify the most important bioclimatic variables determining the distribution of B. stracheyi.
2 Materials and methods
2.1 Study area
Jammu and Kashmir, formerly a state of India (bifurcated into two Union Territories, Jammu-Kashmir and Ladakh) lies between the coordinates 32°17′ to 37°20′ N and73°25′ to 80°30′ E with an elevation range of 225-8,253 m asl (Figure 1). Climatically, the study area is divided into sub-tropical (Jammu), temperate (Kashmir) and cold desert (Ladakh) in south, middle and the east, respectively. Owing to its varied topography, altitude, and climate, the region supports a rich biodiversity and provides a complex habitat for numerous rare, endemic, and threatened plant species. However, several plant species have faced threats over the years due to habitat loss, fragmentation, deforestation, invasive species introduction, overgrazing, overexploitation, land-use changes, a large influx of tourists, road construction, and political unrest (Pant and Pant, 2011; Tali et al., 2019; Mir et al., 2020; Wani et al., 2022a). A total of 429 species of phanerogams, representing 256 genera in 87 families, have been documented in different threat assessment studies in the State (Hamid et al., 2020). This indicates that a significant portion of state’s biodiversity is threatened by the anthropogenic disturbances and climate change (Pant and Pant, 2011; Khuroo et al., 2020).
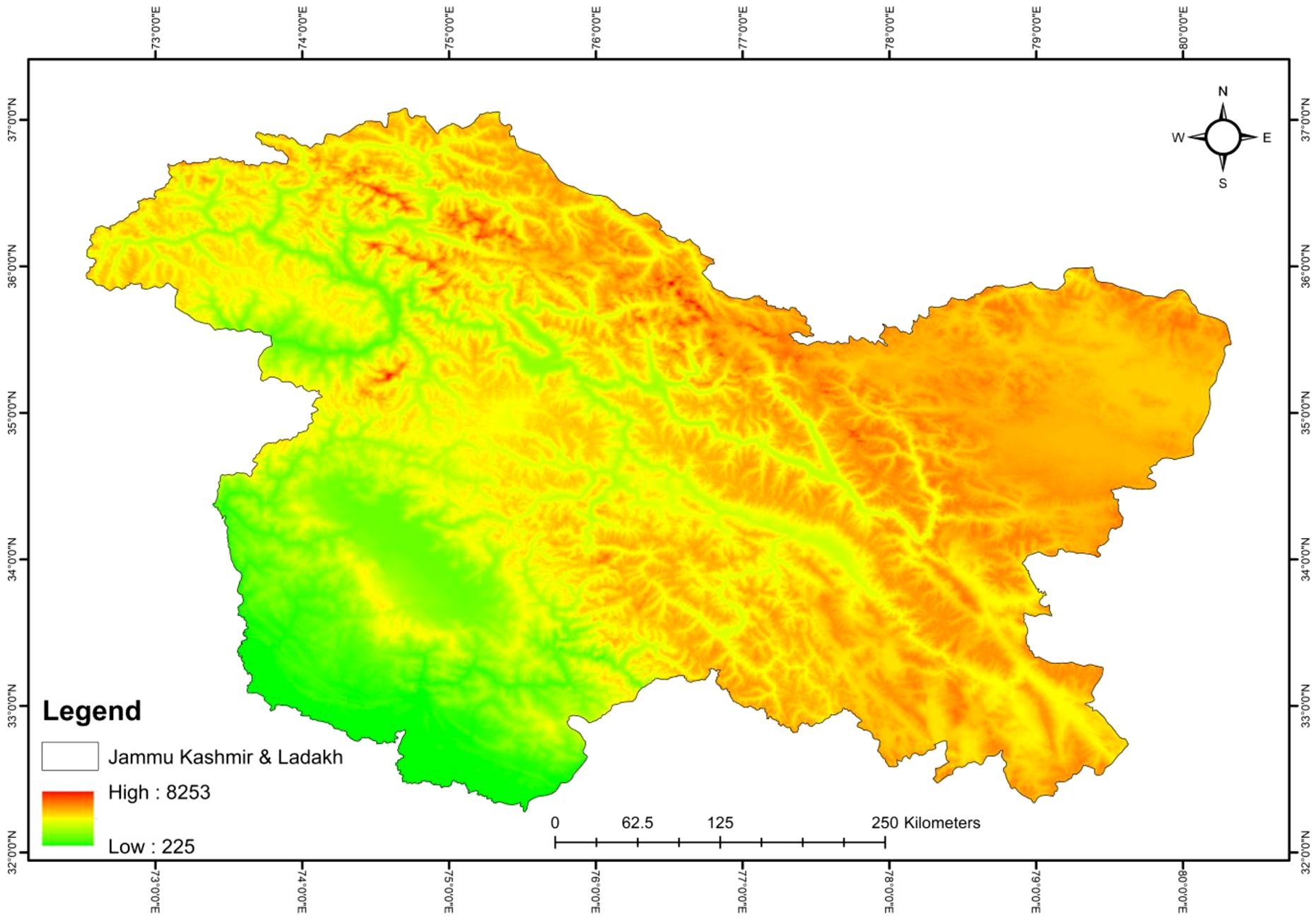
Figure 1. Location and elevation map of the study area.
2.2 Target plant species
Bergenia stracheyi (Hook.f. & Thomson) Engl., a member of the family Saxifragaceae is a herbaceous plant native to the Himalayas, from Afghanistan to Uttarakhand. It is particularly common in the western Himalaya, occurring at elevations between 3300 and 4800 m asl (Tiwari et al., 2017; Flowers of India assessed on 26th November, 2024). B. stracheyi thrives in nutrient rich soil, preferring shady habitats, often growing within rocks crevices (Figure 2). Morphologically, it is distinguished by its massive rootstock reserve and broad petiolar sheath (Chauhan et al., 2016). The rhizomes of B. stracheyi are solid, dark brown, and longitudinally grooved. These rhizomes are traditionally used to treat ailments such as renal calculi, and burns; while their astringent and laxative properties make them beneficial for digestive disorders (Siddiq et al., 2012; Tiwari et al., 2017; Ali et al., 2014). Owing to the presence of biologically active compounds like bergenin (Siddiq et al., 2012), B. stracheyi holds significant medicinal value, particularly in Ayurveda and Unani healthcare systems (Karki et al., 2021). Additionally, plant extracts have been reported to exhibit anti-oxidant and anti-microbial properties (Karki et al., 2021).

Figure 2. Field photographs showing the Habitat (A) and morphology (B) of Bergenia stracheyi.
2.3 Species occurrence data
A total of thirty-six occurrence records of B. stracheyi were collected through intensive field surveys conducted between 2018 and 2023 and was further supplemented with data from the Global Biodiversity Information Facility (GBIF) (http://www.gbif.org accessed 05 June 2024) using the ‘gbif’ function available in ‘dismo’ package in R statistical software version 4.0.3. A crucial pre-processing step in species distribution modeling (SDM) is spatial rarefaction, particularly when occurrence points are unevenly distributed or in clustered within the study area. Occurrence records are often collected from easily accessible locations, such as areas near roads, human settlements, or well-studied sites, leading to the spatial bias (Boria et al., 2014; Ahmad et al., 2019). If left unaddressed, spatial bias can lead to over-predictions in certain regions, resulting in flawed model outputs (Boria et al., 2014; Wani et al., 2022b). To mitigate this, occurrence records were spatially rarefied, removing autocorrelated points and reducing multiple occurrences to a single point within 1×1 km grid. After spatial rarefaction, a final geo-referenced dataset with 31 occurrence points was compiled for modelling the distribution of B. stracheyi.
2.4 Environmental data
Nineteen bioclimatic variables were obtained from the WorldClim database, ver. 1.4 (http://www.worldclim.org) at a 1 km spatial resolution to model the current distribution of B. stracheyi (Table 1). Future climatic data for two representative concentration pathways (RCP 4.5 and RCP 8.5) for the time periods 2050 and 2070 was sourced from the 5th assessment report (AR5) of the Intergovernmental Panel for Climate Change (IPCC). The Hadley Global Environment Model 2-Earth System (HADGEM2-ES) was selected due to its ability to simulate enhanced climate induced ecosystem and hydrological processes (Chakraborty et al., 2016). HADGEM2-ES is widely recognized for its robust simulation capabilities and has been extensively used in climate research to project future climatic conditions under various greenhouse gas emission scenarios. It has been widely applied in predicting species distributions in the Himalayan region (Gajurel et al., 2014; Chakraborty et al., 2016; He et al., 2019; Li et al., 2020; Singh et al., 2022). The data represents simulations for four representative concentration pathways (RCP2.6, RCP4.5, RCP6.0, and RCP8.5), depicting optimistic and pessimistic approaches for the years 2050 and 2070 (Moss et al., 2010). RCP 4.5 assumes that greenhouse gas emissions will be moderate at first then stabilize as a result of substantial mitigation efforts and by the year 2100, radiative forcing is expected to stabilize at about 4.5 W/m². Contradictory, RCP 8.5 assumes a continued high use of fossil fuels and the absence of major efforts to reduce emissions and projects radiative forcing to reach 8.5 W/m² by 2100 (Farooq et al., 2023; Chanzi et al., 2023).
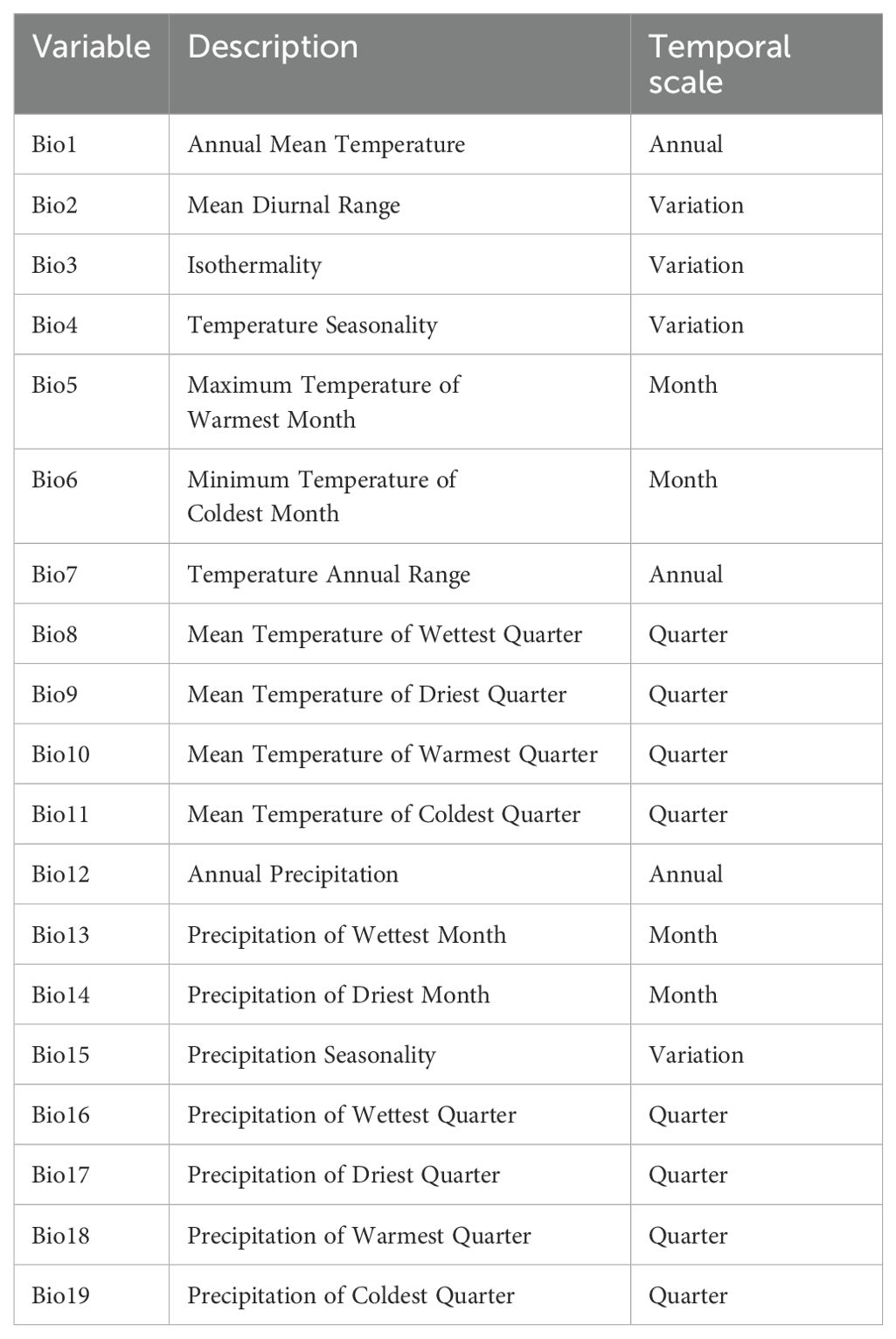
Table 1. List of 19 bioclimatic variables used for habitat suitability modelling of selected plant species downloaded from WorldClim database, ver. 1.4 (http://www.worldclim.org).
In ecological modeling, multi-collinearity among predictor variables can negatively impact model performance, leading to over-fitting, poor generalization, and unreliable results (Dormann et al., 2013a; Amiri et al., 2022). In particular, highly correlated environmental variables can induce redundancy, making it difficult to isolate the individual effects of each variable on the species distribution. To mitigate the impact of multi-collinearity and enhance the robustness of the model, Pearson correlation analysis was used to assess the degree of autocorrelation between bioclimatic variables. For correlation analysis, values for bioclimatic variables for all occurrence points were extracted by using “extract values to points” in ArcGIS 10.8. The exported data was imported into ORIGIN 10.0 software for correlation analysis. Pairs of variables with a correlation coefficient (r) greater than 0.7 were considered highly correlated. Following Peterson et al. (2011), only one variable from each highly correlated pair was retained for modelling, while the others were excluded. This approach ensured that only the most independent variables contributed to the model, minimizing collinearity and improving accuracy. The same set of least correlated variables was used to predict the current and future distribution of B. stracheyi.
2.5 Modelling technique
Current and future distribution modelling and forecasting were performed using the ‘biomod2’ package, designed for ensemble species distribution modeling (Thuiller et al., 2009), within R statistical software (v 4.0.3; R Core Team, 2021). It provides a comprehensive framework for modelling species distributions and predicting their potential geographic ranges under current and future environmental conditions. Widely used in ecology and conservation biology, it is valued for its flexibility, ease of use, and ability to integrate multiple modeling algorithms (Zhang et al., 2024). By allows users to combine results from multiple algorithms to create ensemble models, biomod2 enhances prediction reliability and reduces biases associated with individual models (Gu et al., 2024). In the present study, an ensemble of 10 algorithms viz., Generalized Linear Model (GLM), Generalized Additive Models (GAM), Generalized Boosted Models (GBM), Classification Tree Analysis (CTA), Flexible Discriminant Analysis (FDA), Artificial Neural Networks (ANN), Maximum Entropy (MaxEnt), Multivariate Adaptive Regression Splines (MARS), Random Forest (RF), and Surface Response Envelope (SRE) were used to create species distribution maps.
One thousand pseudo-absences within the study area were generated and to lessen potential sample bias in the generation of pseudo-absences, the procedure was replicated three times following Wu et al. (2024). For model calibration, 80% of the occurrence data was used for training and 20% was used for testing and the process was repeated thrice to reduce uncertainty. Model performance was assessed using cross validation using Area under Curve (AUC), and True Skills Statistics (TSS) as evaluation metrics. The AUC evaluates the performance of a binary classification model by analyzing the Receiver Operating Characteristic (ROC) curve. AUC values range from 0-1, with higher values indicating better model discrimination between presence and absence (Wani et al., 2024b). The TSS assesses the model accuracy by considering both omission and commission errors. Its value ranges from -1 to +1, where higher values indicate better model performance (Freitas et al., 2019). Ensemble model for each climatic scenario and time period combination were constructed using two approaches: committee averaging and weighted mean. The ensemble modelling process incorporated all repetitions and pseudo-absence sets from the algorithm with the highest accuracy score. This resulted in five ensemble models corresponding to current climatic suitabilities and four models corresponding to future climatic suitabilities representing RCP 4.5 and 8.5 for the time periods 2050 and 2070. For the evaluation of relative importance of each climatic variable in governing the current and future distribution of the selected species, permutation procedure was used following Rather et al. (2022).
The RangeSize function in biomod2 package was used to quantify and represent the range change over future climatic scenarios. From the output of the package, information about absolute metrics viz., “Loss, Absent, Stable and Gain” is obtained. Loss is calculated as the number of suitable pixels predicted to be lost under changing climatic scenarios; gain as the number of pixels that are currently unsuitable and predicted to become suitable in future, absent as the number of pixels that are neither suitable nor predicted to be suitable in future and stable as the number of pixels currently suitable and predicted to remain suitable in future. Three additional relative metrics (Percentage Loss, Percentage gain and Range change) were derived from the four absolute metrics following Rather et al. (2022) and Wani et al. (2024b).
3 Results and discussion
With committee averaging TSS and AUC values of 0.56 and 0.87 and weighted mean TSS and AUC values of 0.63 and 0.89, respectively, the final ensemble model performed well in predicting the target species’ distribution. In contrast to the other algorithms used, ANN and SRE showed the lowest accuracy, while GBM, RF, MaxEnt, and GLM performed fairly well. Other algorithms like FDA, MARS, GAM, and CTA showed intermediate performances (Figure 3). Fair performance of GBM, RF, MaxEnt, and GLM in ensemble modeling approaches has also been documented in other ensemble modelling studies (Mohammady et al., 2021; Edalat et al., 2022; Wani et al., 2022b).
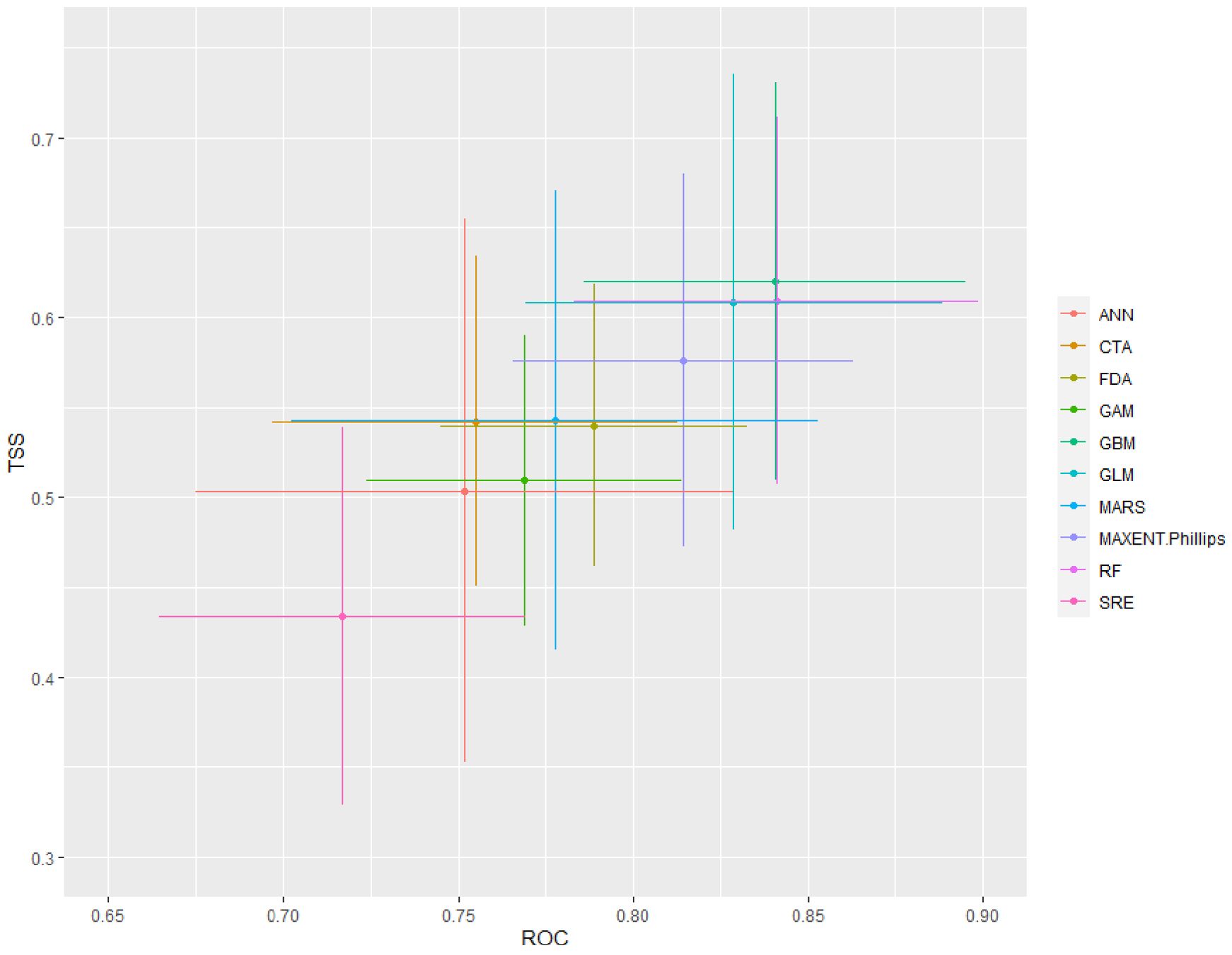
Figure 3. Mean algorithmic model evaluation scores for (B) stracheyi based on two distinct evaluation metrics, ROC (AUC) and TSS.
3.1 Variable importance
Following Pearson’s correlation analysis, five variables were selected for modelling the distribution of B. stracheyi under current climatic conditions (Figure 4). These variables include Bio1 (Annual Mean Temperature), Bio2 (Mean Diurnal Range), Bio4 (Temperature Seasonality), Bio12 (Annual Precipitation) and Bio15 (Precipitation Seasonality). By selecting variables with lower correlation, the final dataset reflects a reduced redundancy in the environmental data, expected to lead to better model performance and more reliable ecological predictions. Reducing multicolinearity among variables is crucial for ensuring that the model predictions are not skewed by the inclusion of highly correlated predictors (Dormann et al., 2013b). Among the selected variables, distribution of B. stracheyi is most strongly influenced by Bio1, Bio12 and Bio2 (Table 2). Bio1 represents the average annual temperature, integrating both daily and seasonal variations. Changes in annual mean temperature can direct or indirect affect the physiology and metabolic activities, thereby impacting species distribution. Bio12, which measures total annual precipitation (in millimeters), plays a crucial role in shaping species distribution by affecting soil moisture levels and soil-plant-atmosphere-continuum (Wani et al., 2024b). Furthermore, the dependence on Bio2 suggests that regions with reduced diurnal fluctuations provide more stable environments, which are likely preferred by B. stracheyi. In contrast, extreme diurnal variations may increase physiological stress, potentially affecting the distribution and survival of species (Venkat and Muneer, 2022).
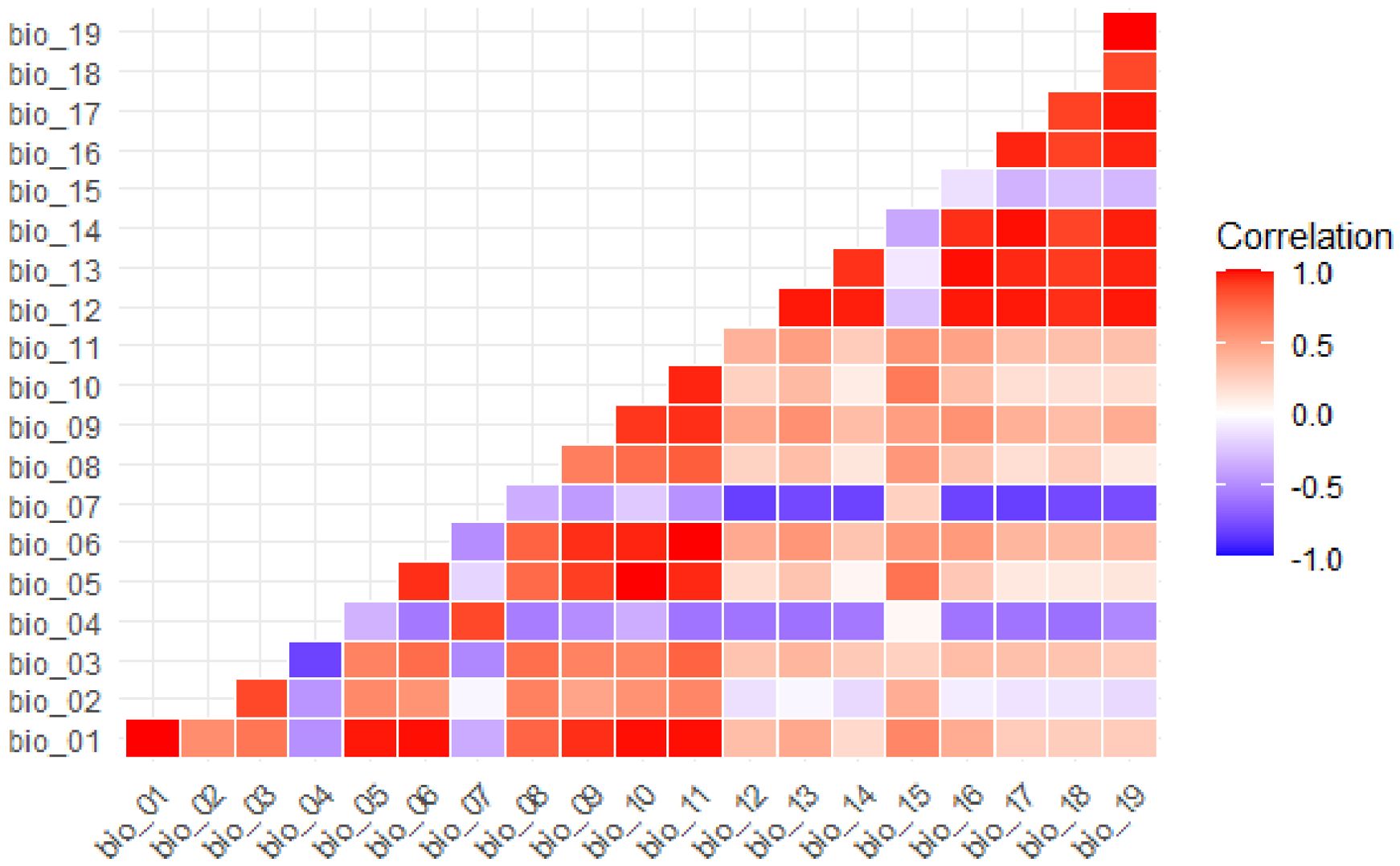
Figure 4. Plot showing the Pearson correlation between the 19 bioclimatic variables.
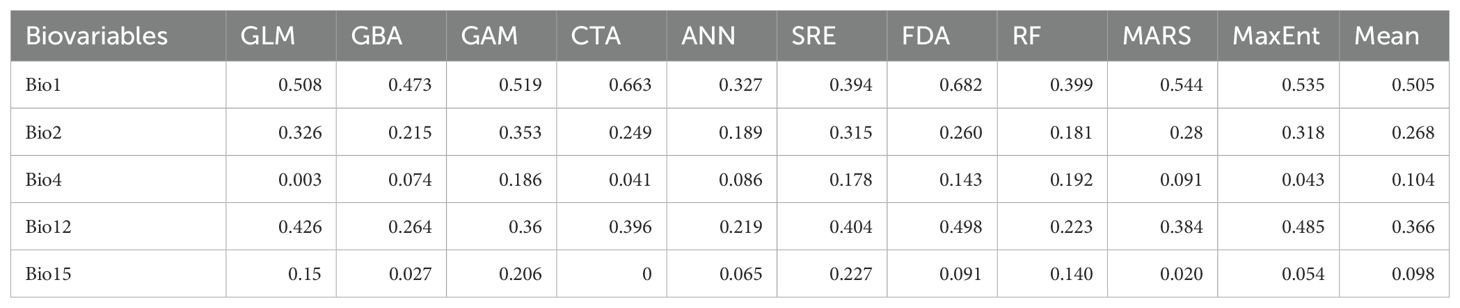
Table 2. Individual and total algorithmic importance scores for the chosen bioclimatic variables.
3.2 Predicted distribution and range change dynamics
Findings of the ensemble model revealed that under the current climatic conditions, ideal habitats for B. stracheyi spread throughout the higher elevations of Jammu and Kashmir most appropriately towards Kargil, Leh, Gilgit-Baltistan, Mirpur, Ghizar, and Rajouri-Poonch region. In Kashmir Valley, Sonamarg, Gulmarg and parts of Shopian district are predicted to be highly suitable for the plant and certain areas of Kupwara, Bandipora, Baramulla, and Ganderbal are predicted to be moderately suitable for B. stracheyi. Lower elevations are predicted to be unsuitable for its growth (Figure 5), supported by the fact that the targeted plant is an alpine species with its distribution reported from the higher elevations of the Himalayan region (Tiwari et al., 2017). Future ensemble model predictions indicated that, across all future climatic scenarios, the majority of the habitats that are currently suitable will continue to be so. However, the species will shift towards the north and southeast in some currently unsuitable habitats, like northern Gilgit-Baltistan, northeastern Leh, and some parts of Budgam and Baramulla (Figure 6).
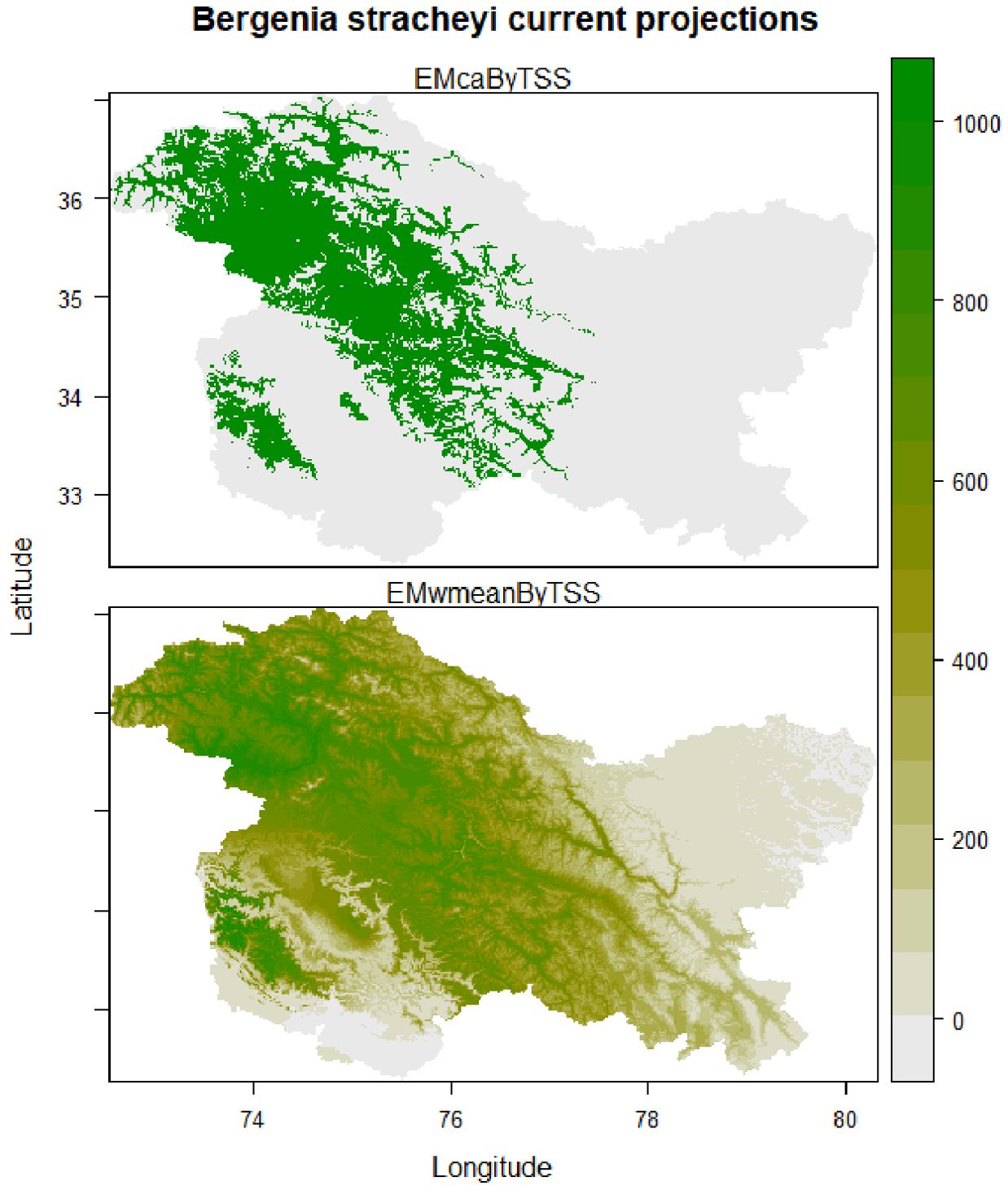
Figure 5. Predicted distribution of (B) stracheyi under present climatic conditions.
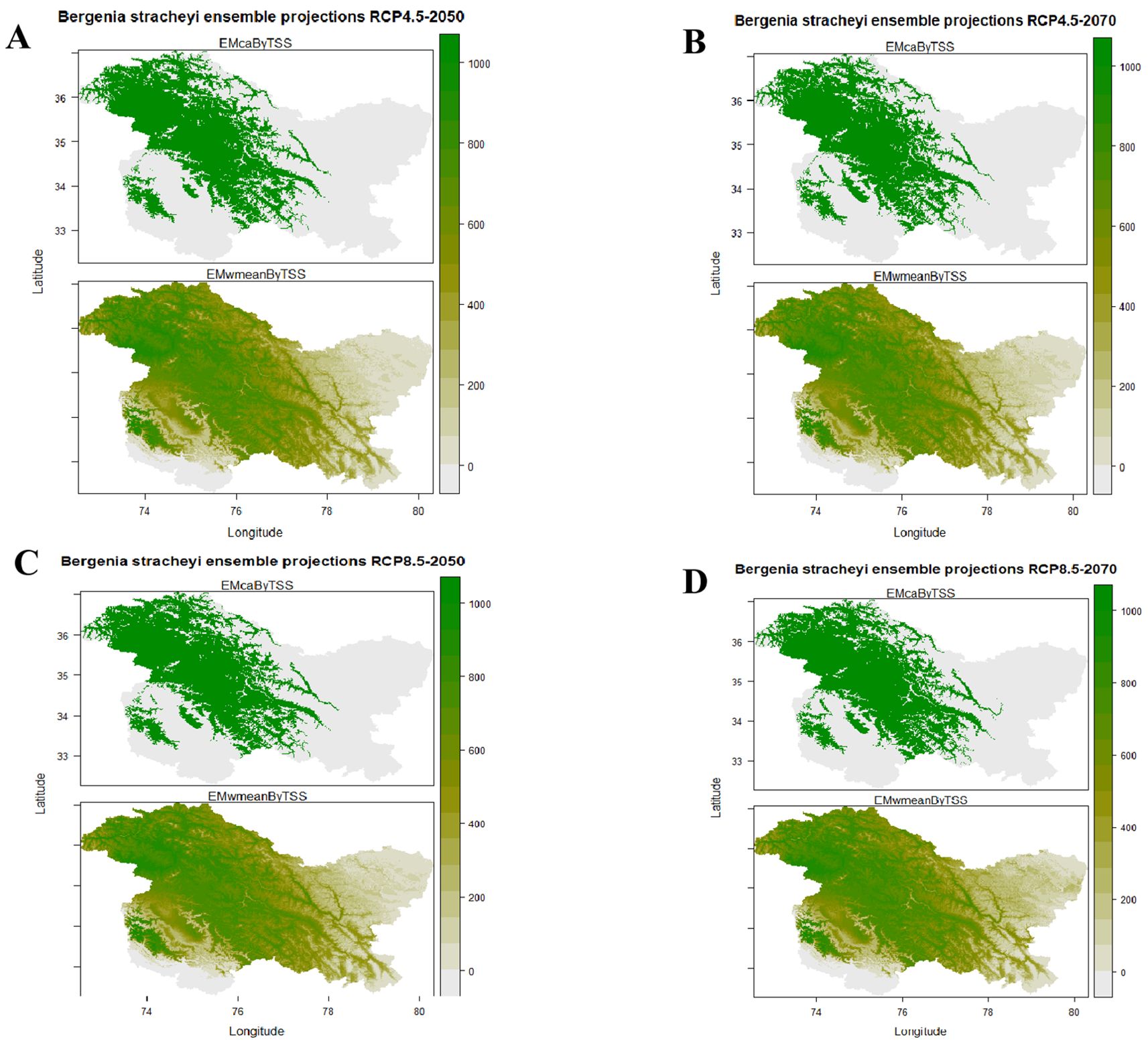
Figure 6. Predicted future distribution of (B) stracheyi under (A) RCP 4.5 (2050); (B) RCP 4.5 (2070); (C) RCP 8.5 (2050) and (D) RCP 8.5 (2070).
Using RCP 4.5 and RCP 8.5 allows comparative assessment of how potential distribution of B. stracheyi may shift under contrasting climate futures. These RCP’s have been extensively validated and widely used in ecological research, ensuring consistency with previous studies, and facilitating comparisons with historical trends. RCPs provide reliable insights into the impacts of different greenhouse gas concentration trajectories on species distribution and habitat suitability (del Río et al., 2021; Shrestha et al., 2021). The model projects a considerable increase in suitable habitats for B. stracheyi, particularly under more severe climate change scenarios (RCP8.5). The range change depicts a considerable increase approximately 42.57% under RCP4.5 for 2050 to as high as 57.31% under RCP8.5 for 2070, as per committee averaging. A similar trend is observed when using weighted mean calculations, with gains ranging from 41.40% to 55.86% across scenarios and timeframes (Table 3). Qiu et al. (2024) have also predicted that the suitable habitat of B. stracheyi is going to expand in future under SSP1-2.6 and SSP2-45.
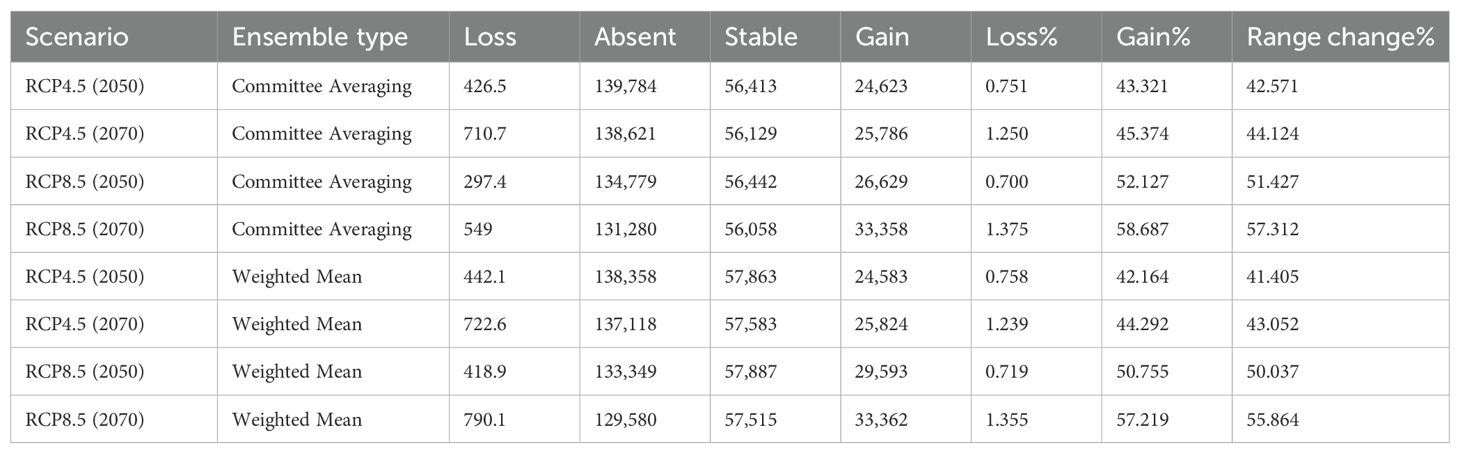
Table 3. Range shift statistics for B. stracheyi under future climate change scenarios in comparison to the current climate conditions (the values are given in km2).
Results from the ensemble modelling indicate that most of the currently suitable habitats for B. stracheyi are projected to remain suitable in future, as represented by the purple patches in Figure 7. This stability is particularly significant for conservation planning, as it suggests that protecting these areas could maintain the species core populations, despite ongoing climatic changes. The persistence of current habitats could be attributed to the adaptability of B. stracheyi across a range of climatic conditions, as well as the relative stability of these regions under future scenarios. These findings suggest that B. stracheyi has the potential to exploit emerging ecological niches that are expected to become more favorable due to shifts in temperature and precipitation patterns driven by climate change. B. stracheyi exhibits lithotriptic property meaning it has the ability to break stones, which serves as an adaptative advantage, allowing it to thrive in environments that are often unsuitable for other plant species (Kumar and Tyagi, 2013). Additionally, its unique morphology characterized by thick and fleshy leaves, enables efficient nutrient and water storage, helping the plant withstand harsh alpine conditions (Pandey et al., 2017).
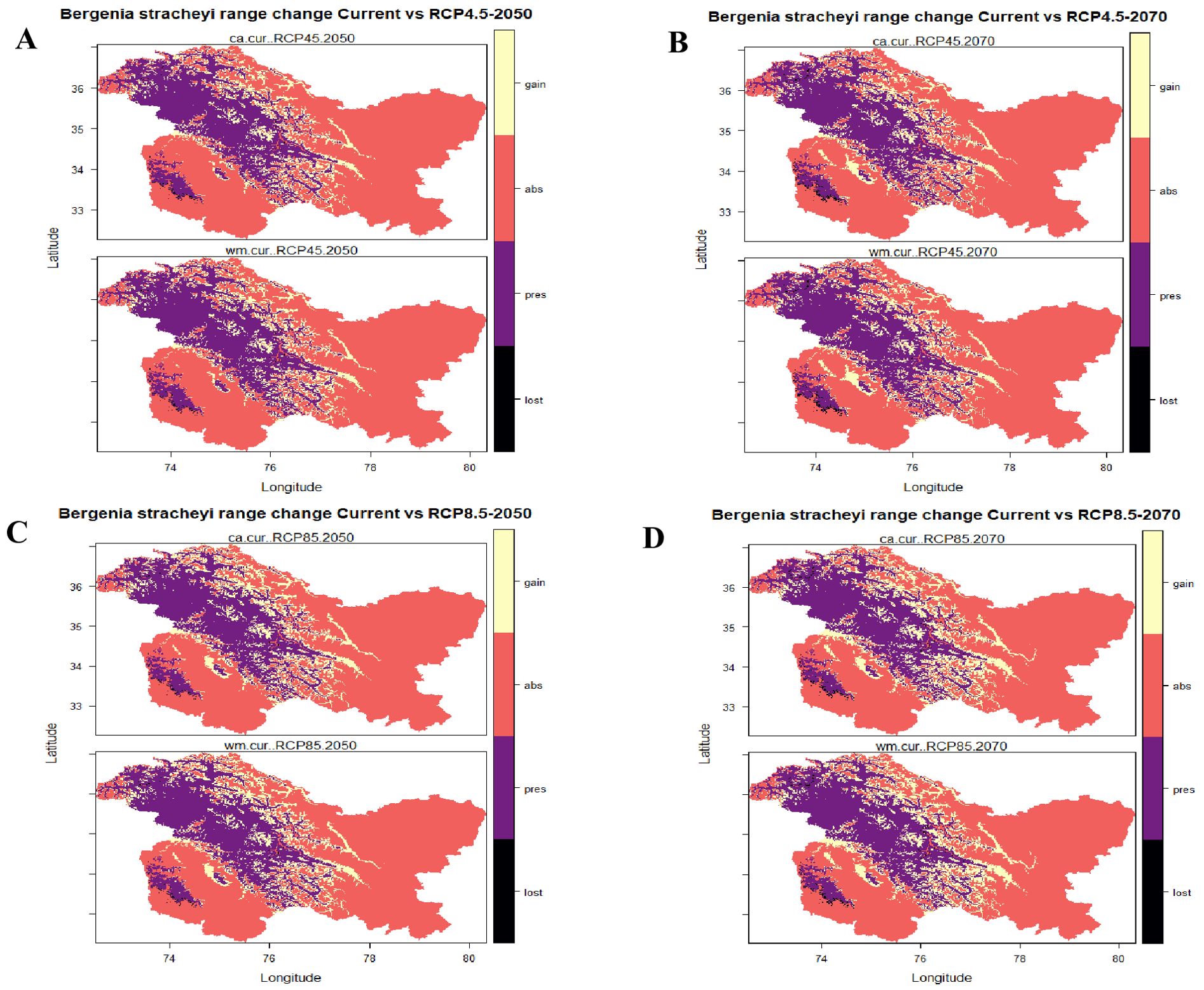
Figure 7. Range change dynamics showing the loss, gain, absent and presence under future climatic scenarios (A) RCP 4.5 (2050); (B) RCP 4.5 (2070); (C) RCP 8.5 (2050); and (D) RCP 8.5 (2070).
Some areas currently deemed unsuitable for B. stracheyi are projected to become suitable in the future, indicated by yellow patches in Figure 6. These regions are primarily located in northern Gilgit-Baltistan, northeastern Leh, and parts of Budgam and Baramulla. The species is predicted to shift its distribution northward and southeastward, aligning with findings from Qiu et al. (2024), who reported that two Himalayan species of Bergenia, B. ciliata and B. stracheyi are expected to expand their ranges in similar directions. The areas predicted to become suitable for B. stracheyi in the future are expected to experience climatic conditions that align with its ecological requirements. This expansion may contribute to an overall increase in population size and genetic diversity, providing an opportunity for adaptive resilience under changing climatic conditions. However, despite the projected habitat gains, some areas currently identified as suitable, including parts of Rajouri-Poonch and Mirpur (Pakistan), are expected to become unsuitable in the future (black patches in Figure 7). These habitat losses are likely driven by climatic shifts that exceed the tolerance range of B. stracheyi.
Climate change is profoundly reshaping plant species distributions worldwide, with significant consequences for ecosystems, biodiversity, and human societies (Kelly and Goulden, 2008; Pecl et al., 2017; Mosoh et al., 2024). Rising temperatures, changing precipitation patterns, and more frequent extreme weather events are forcing many plant species to shift their ranges to higher latitudes or elevations in search of suitable habitats (Muluneh, 2021; Yang et al., 2024; Zhao et al., 2024). With rising temperatures, B. stracheyi is predicted to expand its distribution northward and southeastward. Similar range shifts have been predicted for other plant species in the Himalayan region (Telwala et al., 2013; Manish et al., 2016; He et al., 2019; Manish, 2022; Wani et al., 2022a; Qiu et al., 2024; Satish et al., 2024). Such shifts in plant distribution have far-reaching consequences for ecosystem functioning and stability and the services these species provide to human communities. Additionally, these shifts may disrupt plant-pollinator interactions, leading to phenological mismatches with potentially severe ecological consequences (Karthik et al., 2021; Shivanna, 2022). As climate change disrupts the timing of phenological events, plants may flower before their pollinators become active or vice versa, potentially leading to reproductive failures and population declines. The predicted range shifts in the distribution of B. stracheyi underscore the urgency of implementing local conservation measures to mitigate the impacts of climate change.
4 Limitations of the study
Although this study employs a robust methodology and well performing models, certain limitations exist. The distribution of B. stracheyi was primarily modeled using nineteen bioclimatic variables, which may have led to an overestimation of actual range. Incorporating additional variables such as topographical, soil features, land-use, and biotic interactions could further refine the model and provide a comprehensive understanding of the species distribution (Qiu et al., 2024). Furthermore, the future climatic projections in this study were based on CMIP5 (IPCC AR5) which is now considered obsolete. The latest CMIP6 framework introduces updated climate scenarios known as Shared Socioeconomic Pathways (SSPs), developed by the energy modelling community, offering more refined and policy relevant climate projections.
5 Conclusion
B. stracheyi is an important medicinal plant native to the Himalayas, occurring at elevations between 3300 and 4800 m asl. In the present study, an ensemble modelling approach was used to predict the current and future distribution of B. stracheyi under the anticipated climate change scenarios for the time periods 2050 and 2070. The findings of the study revealed that the distribution of B. stracheyi is predominantly determined by temperature and precipitation variables impeding that alterations in temperature and precipitation can have a considerable direct or indirect effect on the plant, affecting its distribution. Results of the ensemble modelling revealed that most of the currently suitable habitats for B. stracheyi are likely to remain suitable in future. Some currently unsuitable areas for the plant are expected to become suitable in future, allowing the species to expand its distribution northward and southeastward. Further, some currently areas are predicted to become unsuitable for the plant in future. Thus, overall B. stracheyi is predicted to show major range change shifts under future climatic scenarios. Findings of the present study endorse and lay a reliable foundation for conservation planning of B. stracheyi. Further, present study recommends that distribution of B. stracheyi should be predicted using the latest future climatic scenarios and all possible biotic and abiotic variables for better model predictions.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
ZW: Data curation, Investigation, Methodology, Software, Writing – original draft, Writing – review & editing. JD: Data curation, Supervision, Validation, Visualization, Writing – review & editing. AL: Writing – review & editing. SP: Conceptualization, Methodology, Supervision, Visualization, Writing – review & editing. SS: Funding acquisition, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors extend their appreciation to the Deanship of Research and Graduate Studies at King Khalid University for funding this work through Large Group Project under grant number RGP2/49/45.
Acknowledgments
Zishan Ahmad Wani acknowledges the Department of Environmental Science and Engineering, SRM University-AP for providing University Post Doctoral Fellowship.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abro T. W., Desta A. B., Debie E., Alemu D. M. (2024). Endemic plant species and threats to their sustainability in Ethiopia: A systematic review. Trees Forests People 100634.
Ahmad R., Khuroo A. A., Charles B., Hamid M., Rashid I., Aravind N. A. (2019). Global distribution modelling, invasion risk assessment and niche dynamics of Leucanthemum vulgare (Ox-eye Daisy) under climate change. Sci. Rep. 9 (1), 1–15. doi:10.1038/s41598-019-47859-1
Ali I., Bibi S., Hussain H., Bano F., Ali S., Khan S. W., et al. (2014).Biological activities of Suaeda heterophylla and Bergenia stracheyi. Asian Pac. J. Trop. Dis. 4, S885–S889. doi: 10.1016/S2222-1808(14)60752-0
Amiri M., Tarkesh M., Shafiezadeh M. (2022). Modelling the biological invasion of Prosopis juliflora using geostatistical-based bioclimatic variables under climate change in arid zones of southwestern Iran. J. Arid Land 14, 203–224.
Anderson J. T., Song B. H. (2020). Plant adaptation to climate change—Where are we? J. Systematics Evol. 58, 533–545.
Arneth A., Shin Y. J., Leadley P., Rondinini C., Bukvareva E., Kolb M., et al. (2020). Post-2020 biodiversity targets need to embrace climate change. Proc. Natl. Acad. Sci. 117, 30882–30891.
Boria R. A., Olson L. E., Goodman S. M., Anderson R. P. (2014). Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 275, 73–77.
Chakraborty A., Joshi P. K., Sachdeva K. (2016). Predicting distribution of major forest tree species to potential impacts of climate change in the central Himalayan region. Ecol. Eng. 97, 593–609.
Chanzi G., Bushesha M., Munishi S., Karia A. (2023). Application of mann kendal sen’s slope estimator in trend analysis of historical and future precipitation and temperature in the kilombero river basin. Huria: J. Open Univ. Tanzania 30, 127–150.
Chaudhry S., Sidhu G. P. S. (2022). Climate change regulated abiotic stress mechanisms in plants: a comprehensive review. Plant Cell Rep. 41 (1), 1–31. doi: 10.1007/s00299-021-02759-5
Chauhan R., Ruby K., Dwivedi J. (2016). Antioxidant, lipid peroxidation and astringency study of hydroethanolic root extracts of Bergenia ligulata, Bergenia ciliata and B. stracheyi. Eur. J. Medicinal Plants 15, 1–10. doi: 10.9734/EJMP/2016/25370
del Río S., Canas R., Cano E., Cano-Ortiz A., Musarella C., Pinto-Gomes C., et al. (2021). Modelling the impacts of climate change on habitat suitability and vulnerability in deciduous forests in Spain. Ecol. Indic. 131, 108202. doi: 10.1016/j.ecolind.2021.108202
Dormann C. F., Elith J., Bacher S., Buchmann C., Carl G., Carre G., et al. (2013a). Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27–46. doi: 10.1111/j.1600-0587.2012.07348.x
Dormann C. F., Elith J., Bacher S., Buchmann C., Carl G., Carré G., et al. (2013b). Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27–46. doi: 10.1111/j.1600-0587.2012.07348.x
Edalat M., Dastres E., Jahangiri E., Moayedi G., Zamani A., Pourghasemi H. R., et al. (2022). Spatial mapping Zataria multiflora using different machine-learning algorithms. Catena 212, 106007. doi: 10.1016/j.catena.2021.106007
Farooq A., Farooq N., Akbar H., Hassan Z. U., Gheewala S. H. (2023). A critical review of climate change impact at a global scale on cereal crop production. Agronomy 13, 162. doi: 10.3390/agronomy13010162
Frans V. F., Augé A. A., Fyfe J., Zhang Y., McNally N., Edelhoff H., et al. (2022). Integrated SDM database: Enhancing the relevance and utility of species distribution models in conservation management. Methods Ecol. Evol. 13, 243–261. doi: 10.1111/2041-210X.13736
Freitas G. H., Costa L. M., Silva P. H., Chaves A. V., Ribeiro L. C., Rodrigues M. (2019). Spatial ecology and conservation of the microendemic ovenbird CipoCinclodes (Cinclodes espinhacensis) from the Brazilian highlands. J. Field Ornithology 90, 128–142. doi: 10.1111/jofo.2019.90.issue-2
Gajurel J. P., Werth S., Shrestha K. K., Scheidegger C. (2014). Species distribution modeling of Taxus wallichiana (Himalayan yew) in Nepal Himalaya. Asian J. Conserv. Biol. 3, 127–134.
Guisan A., Tingley R., Baumgartner J. B., Naujokaitis-Lewis I., Sutcliffe P. R., Tulloch A. I., et al. (2013). Predicting species distributions for conservation decisions. Ecol. Lett. 16 (12), 1424–1435. doi: 10.1111/ele.12189
Gu R., Wei S., Li J., Zheng S., Li Z., Liu G., et al. (2024). Predicting the impacts of climate change on the geographic distribution of moso bamboo in China based on biomod2 model. Eur. J. For. Res. 143, 1499–1512.
Hamid M., Khuroo A. A., Ahmad R., Rasheed S., Malik A. H., Dar G. H. (2020). Threatened flora of Jammu and Kashmir state. Biodiversity Himalaya: Jammu Kashmir State 957–995.
He X., Burgess K. S., Yang X. F., Ahrends A., Gao L. M., Li D. Z. (2019). Upward elevation and northwest range shifts for alpine Meconopsis species in the Himalaya–Hengduan Mountains region. Ecol. Evol. 9, 4055–4064.
Javeed B., Ridwan Q., Huang D., Wani Z. A., Siddiqui S., Yassin H. M., et al. (2024). Ecological niche modelling: a global assessment based on bibliometric analysis. Front. Environ. Sci. 12, 1376213.
Kaky E., Nolan V., Alatawi A., Gilbert F. (2020). A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: A case study with Egyptian medicinal plants. Ecol. Inf. 60, 101150.
Karki S., Chowdhury S., Nath S., Dora K., Murmu P. (2021). Phytochemistry and ethnomedicinal use of Bergenia species-a miraculous herb. Indian J. Anim. Health 60, 143–152.
Karthik S., Reddy M. S., Yashaswini G. (2021). “Climate change and its potential impacts on insect-plant interactions,” in The nature, causes, effects and mitigation of climate change on the environment, 10.
Kelly A. E., Goulden M. L. (2008). Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. 105, 11823–11826. doi: 10.1073/pnas.0802891105
Khuroo A. A., Mehraj G., Muzafar I., Rashid I., Dar G. H. (2020). “Biodiversity conservation in Jammu and Kashmir state: current status and future challenges,” in Biodiversity of the Himalaya: Jammu and Kashmir State, 1049–1076.
Kraus D., Enns A., Hebb A., Murphy S., Drake D. A. R., Bennett B. (2023). Prioritizing nationally endemic species for conservation. Conserv. Sci. Pract. 5, e12845. doi: 10.1111/csp2.12845
Kumar A., Shabnam S., Oraon P. R., Malik M. S. (2024). “Forest health in a changing scenario of climate change,” in Sustainable forest resources management. (Apple Academic Press), 109–129.
Kumar V., Tyagi D. (2013). Phytochemical screening and free-radical scavenging activity of B. stracheyi. J. Pharmacognosy Phytochem. 2, 175–180.
Lenoir J., Bertrand R., Comte L., Bourgeaud L., Hattab T., Murienne J., et al. (2020). Species better track climate warming in the oceans than on land. Nat. Ecol. Evol. 4, 1044–1059. doi: 10.1038/s41559-020-1198-2
Li P., Zhu W., Xie Z., Qiao K. (2020). Integration of multiple climate models to predict range shifts and identify management priorities of the endangered Taxus wallichiana in the Himalaya–Hengduan Mountain region. J. Forestry Res. 31, 2255–2272. doi: 10.1007/s11676-019-01009-5
Manes S., Costello M. J., Beckett H., Debnath A., Devenish-Nelson E., Grey K. A., et al. (2021a). Endemism increases species’ climate change risk in areas of global biodiversity importance. Biol. Conserv. 257, 109070. doi: 10.1016/j.biocon.2021.109070
Manes S., Costello M. J., Beckett H., Debnath A., Devenish-Nelson E., Grey K. A., et al. (2021b). Endemism increases species’ climate change risk in areas of global biodiversity importance. Biol. Conserv. 257, 109070. doi: 10.1016/j.biocon.2021.109070
Manish K. (2022). Medicinal plants in peril due to climate change in the Himalaya. Ecol. Inf. 68, 101546. doi: 10.1016/j.ecoinf.2021.101546
Manish K., Telwala Y., Nautiyal D. C., Pandit M. K. (2016). Modelling the impacts of future climate change on plant communities in the Himalaya: a case study from Eastern Himalaya, India. Modeling Earth Syst. Environ. 2, 1–12. doi: 10.1007/s40808-016-0163-1
Miller J. (2010). Species distribution modeling. Geogr. Compass 4, 490–509. doi: 10.1111/j.1749-8198.2010.00351.x
Mir A. H., Tyub S., Kamili A. N. (2020). Ecology, distribution mapping and conservation implications of four critically endangered endemic plants of Kashmir Himalaya. Saudi J. Biol. Sci. 27 (9), 2380–2389. doi: 10.1016/j.sjbs.2020.05.006
Mishra A., Jugran H. P., Sekar K. C., Talukdar G. (2024). “Plant phenological shifts in the Indian himalayan region,” in Warming mountains: implications for livelihood and sustainability (Springer Nature Switzerland, Cham), 85–104.
Mohammady M., Pourghasemi H. R., Yousefi S., Dastres E., Edalat M., Pouyan S., et al. (2021). Modeling and prediction of habitat suitability for Ferula gummosa medicinal plant in a mountainous area. Natural Resour. Res. 30, 4861–4884. doi: 10.1007/s11053-021-09940-3
Mosoh D. A., Prakash O., Khandel A. K., Vendrame W. A. (2024). Preserving earth’s flora in the 21st century: climate, biodiversity, and global change factors since the mid-1940s. Front. Conserv. Sci. 5, 1383370.
Moss R. H., Edmonds J. A., Hibbard K. A., Manning M. R., Rose S. K., Van Vuuren D. P., et al. (2010). The next generation of scenarios for climate change research and assessment. Nature 463, 747–756.
Muluneh M. G. (2021). Impact of climate change on biodiversity and food security: a global perspective—a review article. Agric. Food Secur. 10, 1–25. doi: 10.1186/s40066-021-00318-5
Palombo M. R. (2021). Thinking about the biodiversity loss in this changing world. Geosciences 11, 370. doi: 10.3390/geosciences11090370
Pandey R., Kumar B., Meena B., Srivastava M., Mishra T., Tiwari V., et al. (2017). Major bioactive phenolics in Bergenia species from the Indian Himalayan region: Method development, validation and quantitative estimation using UHPLC-QqQLIT-MS/MS. PloS One 12, e0180950.
Pant S., Pant V. S. (2011). Status and conservation management strategies for threatened plants of Jammu and Kashmir. J. Phytology 3.
Pazzaglia J., Reusch T. B., Terlizzi A., Marín-Guirao L., Procaccini G. (2021). Phenotypic plasticity under rapid global changes: The intrinsic force for future seagrasses survival. Evolutionary Appl. 14, 1181–1201. doi: 10.1111/eva.13212
Pecl G. T., Araújo M. B., Bell J. D., Blanchard J., Bonebrake T. C., Chen I. C., et al. (2017). Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 355, eaai9214.
Pepin N. C., Arnone E., Gobiet A., Haslinger K., Kotlarski S., Notarnicola C., et al. (2022). Climate changes and their elevational patterns in the mountains of the world. Rev. Geophysics 60, e2020RG000730.
Permesan C. (2006). Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 37, 637–669.doi: 10.1146/annurev.ecolsys.37.091305.110100
Peterson A. T., Soberón J., Pearson R. G., Anderson R. P., Martínez-Meyer E., Nakamura M., et al. (2011). “Ecological niches and geographic distributions (MPB-49),” in Ecological niches and geographic distributions (MPB-49) (Princeton University Press).
Profirio L. L., Harris R. M. B., Lefroy E. C., Hugh S., Gould S. F., Lee G., et al. (2014). Improving the use of species distribution models in conservation planning and management under climate change. PloS One 9, e113749.
Qazi A. W., Saqib Z., Zaman-ul-Haq M. (2022). Trends in species distribution modelling in context of rare and endemic plants: a systematic review. Ecol. Processes 11, 1–11.
Qiu L., Fu Q. L., Jacquemyn H., Burgess K. S., Cheng J. J., Mo Z. Q., et al. (2024). Contrasting range changes of Bergenia (Saxifragaceae) species under future climate change in the Himalaya and Hengduan Mountains Region. Theoret. Appl. Climatol. 155 (3), 1927–1939. doi: 10.1007/s00704-023-04746-0
R Core Team. (2021). R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
Rana N., Manish K., Pandit M. K. (2024). Effect of climate change on the flowering phenology of Rhododendron arboreum Sm. in the Western Himalaya. J. Asia-Pacific Biodiversity.
Rather Z. A., Ahmad R., Khuroo A. A. (2022). Ensemble modelling enables identification of suitab le sites for habitat restoration of threatened biodiversity under climate change: A case study of Himalayan Trillium. Ecol. Eng. 176, 106534.
Ripple W. J., Wolf C., Ne,wsome T. M., Hoffmann M., Wirsing A. J., McCauley. D. J. (2017). Extinction risk is most acute for the world’s largest and smallest vertebrates. Proc. Natl. Acad. Sci. 114, 10678–10683.
Roman-Palacios C., Wiens J. J. (2020). Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. United States America 117, 4211–4217.
Sarkar D., Jagannivsan H., Debnath A., Talukdar G. (2024). A systematic review on the potential impact of future climate change on India’s biodiversity using species distribution model (SDM) studies: trends, and data gaps. Biodiversity Conserv. 33, 3399–3415.
Satish K. V., Srivastava P. K., Behera M. D., Khan M. L., Gwal S., Srivastava S. K. (2024). Ensemble of machine learning and global circulation models coupled with geospatial databases for niche mapping of Bell Rhododendron under climate change. Geocarto Int. 39, 2421233.
Sekar K. C., Thapliyal N., Bhojak P., Bisht K., Pandey A., Mehta P., et al. (2024). Early signals of climate change impacts on alpine plant diversity in Indian Himalaya. Biodiversity Conserv. 1–27.
Shakoor A., Albasher G., Farooq T. H. (2023). Climate change on the brink: Time for urgent action. Ecol. Inf. 78, 102286.
Shivanna K. R. (2022). Climate change and its impact on biodiversity and human welfare. Proc. Indian Natl. Sci. Acad. 88, 160–171.
Shrestha B., Tsiftsis S., Chapagain D. J., Khadka C., Bhattarai P., Kayastha Shrestha N., et al. (2021). Suitability of habitats in Nepal for Dactylorhiza hatagirea now and under predicted future changes in climate. Plants 10, 467.
Shrestha U. B., Gautam S., Bawa K. S. (2012). Widespread climate change in the Himalayas and associated changes in local ecosystems. PloS One 7 (5), e36741. doi: 10.1371/journal.pone.0036741
Shrestha U. B., Lamsal P., Ghimire S. K., Shrestha B. B., Dhakal S., Shrestha S., et al. (2022). Climate change-induced distributional change of medicinal and aromatic plants in the Nepal Himalaya. Ecol. Evol. 12 (8), e9204. doi: 10.1002/ece3.9204
Siddiq F., Fatima I., Malik A., Afza N., Iqbal L., Lateef M., et al. (2012). Biologically active bergenin derivatives from B. stracheyi. Chem. biodiversity 9, 91–98. doi: 10.1002/cbdv.201100003
Singh L., Kanwar N., Bhatt I. D., Nandi S. K., Bisht A. K. (2022). Predicting the potential distribution of Dactylorhiza hatagirea (D. Don) Soo-an important medicinal orchid in the West Himalaya, under multiple climate change scenarios. PloS One 17, e0269673.
Taheri S., Naimi B., Rahbek C., Araújo M. B. (2021). Improvements in reports of species redistribution under climate change are required. Sci. Adv. 7, eabe1110. doi: 10.1126/sciadv.abe1110
Tali B. A., Khuroo A. A., Nawchoo I. A., Ganie A. H. (2019). Prioritizing conservation of medicinal flora in the Himalayan biodiversity hotspot: an integrated ecological and socioeconomic approach. Environ. Conserv. 46 (2), 147–154. doi: 10.1017/S0376892918000425
Telwala Y., Brook B. W., Manish K., Pandit M. K. (2013). Climate-induced elevational range shifts and increase in plant species richness in a Himalayan biodiversity epicentre. PloS One 8, e57103. doi: 10.1371/journal.pone.0057103
Thuiller W., Lafourcade B., Engler R., Araujo M. B. (2009). BIOMOD–a platform for ensemble forecasting of species distributions. Ecography 32 (3), 369–373. doi: 10.1111/j.1600-0587.2008.05742.x
Tiwari V., Meena B., Nair K. N., Upreti D. K., Tamta S., Rana T. S. (2017). Assessment of genetic diversity and population structure of B. stracheyi (Saxifragaceae) in the Western Himalaya (India). Biochem. Systematics Ecol. 70, 205–210. doi: 10.1016/j.bse.2016.12.001
Turnhout E., Purvis A. (2020). Biodiversity and species extinction: categorization, calculation, and communication. Griffith Law Rev. 29, 669–685. doi: 10.1080/10383441.2020.1925204
Venkat A., Muneer S. (2022). Role of circadian rhythms in major plant metabolic and signaling pathways. Front. Plant Sci. 13, 836244. doi: 10.3389/fpls.2022.836244
Verrall B., Pickering C. M. (2020). Alpine vegetation in the context of climate change: A global review of past research and future directions. Sci. Total Environ. 748, 141344. doi: 10.1016/j.scitotenv.2020.141344
Vitasse Y., Ursenbacher S., Klein G., Bohnenstengel T., Chittaro Y., Delestrade A., et al. (2021). Phenological and elevational shifts of plants, animals and fungi under climate change in the European Alps. Biol. Rev. 96, 1816–1835. doi: 10.1111/brv.12727
Wan J. Z., Wang C. J., Yu. F. H. (2016). Risk hotspots for terrestrial plant invaders under climate change at the global scale. Environ. Earth Sci. 75. doi: 10.1007/s12665-016-5826-8
Wang H., Bin-Bin W., Cui P., Yao-Ming M., Wang Y., Jian-Sheng H., et al. (2024). Disaster effects of climate change in High-Mountain Asia: State of art and scientific challenges. Adv. Climate Change Res. doi: 10.1016/j.accre.2024.06.003
Wani Z. A., Bhat J. A., Negi V. S., Satish K. V., Siddiqui S., Pant S. (2022a). Conservation Priority Index of species, communities, and habitats for biodiversity conservation and their management planning: A case study in Gulmarg Wildlife Sanctuary, Kashmir Himalaya. Front. Forests Global Change 5, 995427. doi: 10.3389/ffgc.2022.995427
Wani Z. A., Khan S., Satish K. V., Haq S. M., Pant S., Siddiqui S. (2024b). Ensemble modelling reveals shrinkage of suitab le habitat for Himalayan Boxwood (Buxus wallichiana Bail.) under climate change-implications for conservation. Phytocoenologia 52. doi: 10.1127/phyto/2024/0427
Wani I. A., Khan S., Verma S., Al-Misned F. A., Shafik H. M., El-Serehy H. A. (2022b). Predicting habitat suitability and niche dynamics of Dactylorhiza hatagirea and Rheum webbianum in the Himalaya under projected climate change. Sci. Rep. 12, 13205. doi: 10.1038/s41598-022-16837-5
Wani Z. A., Pant S., Bhat J. A., Shukla G. (2024a). Distribution and survival of medicinal and aromatic plants is threatened by the anticipated climate change. Trees Forests People 16, 100549. doi: 10.1016/j.tfp.2024.100549
Wani Z. A., Satish K. V., Islam T., Dhyani S., Pant S. (2023). Habitat suitability modelling of Buxus wallichiana Bail.: an endemic tree species of Himalaya. Vegetos 36, 583–590. doi: 10.1007/s42535-022-00428-w
Weiskopf S. R., Rubenstein M. A., Crozier L. G., Gaichas S., Griffis R., Halofsky J. E., et al. (2020). Climate change effects on biodiversity, ecosystems, ecosystem services, and natural resource management in the United States. Sci. Total Environ. 733, 137782. doi: 10.1016/j.scitotenv.2020.137782
Wu C., Li S., Zhou Y., Hu X., Feng J. (2024). High lability of global niche and range in the Giant African Snail (Lissachatina fulica): small niche expansions resulting in large range shifts. Ecol. Indic. 151, 110328. doi: 10.1016/j.ecolind.2023.110328
Yang L., Zhu X., Song W., Shi X., Huang X. (2024). Predicting the potential distribution of 12 threatened medicinal plants on the Qinghai-Tibet Plateau, with a maximum entropy model. Ecol. Evol. 14, e11042. doi: 10.1002/ece3.11042
Zandalinas S. I., Balfagón D., Gómez-Cadenas A., Mittler R. (2022). Plant responses to climate change: metabolic changes under combined abiotic stresses. J. Exp. Bot. 73 (11), 3339–3354. doi: 10.1093/jxb/erac073
Zellmer A. J., Claisse J. T., Williams C. M., Schwab S., Pondella D. J. (2019). Predicting optimal sites for ecosystem restoration using stacked-species distribution modeling. Front. Mar. Sci. 6, 3. doi: 10.3389/fmars.2019.00003
Zhang X., Wu W., Liang Y. (2024). Analysis of the potential distribution of shoot blight of larch in China based on the optimized maxEnt and biomod2 ensemble models. Forests 15, 1313. doi: 10.3390/f15081313
Keywords: native and endemic, medicinal plants, climate change, ensemble modelling, range change, Himalayan region
Citation: Wani ZA, Dar JA, Lone AN, Pant S and Siddiqui S (2025) Habitat suitability modelling and range change dynamics of Bergenia stracheyi under projected climate change scenarios. Front. Ecol. Evol. 13:1561640. doi: 10.3389/fevo.2025.1561640
Received: 16 January 2025; Accepted: 31 March 2025;
Published: 22 April 2025.
Edited by:
Fernanda Michalski, Universidade Federal do Amapá, BrazilReviewed by:
Rupesh Dhyani, University of Giessen, GermanyShiv Paul, Himalayan Forest Research Institute (HFRI), India
Copyright © 2025 Wani, Dar, Lone, Pant and Siddiqui. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Javid Ahmad Dar, amF2aWRhaG1hZC5kQHNybWFwLmVkdS5pbg==