 Xueli Feng
Xueli Feng Zhengxue Zhao
Zhengxue Zhao
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 27 March 2025
Sec. Models in Ecology and Evolution
Volume 13 - 2025 | https://doi.org/10.3389/fevo.2025.1549532
Determining the suitable areas for pests and the dynamic shift of these areas under climate change is crucial for their management and control. The black planthopper (Ricania speculum) is an economically important pest in China, but knowledge about the suitable areas for its habitation is limited. Therefore, in this study, a Maxent model was constructed using 111 occurrence records and related environmental variables to predict the suitable areas for it to inhabit now and in the future (2041–2060, 2061–2080, and 2081–2100) in China, following by exploration of their dynamic shift. The model results showed that the currently suitable areas are predominantly in central and southern provinces such as Henan, Anhui, Hubei, and Guizhou, while the areas that are highly suitable are the most abundant among the three levels of suitability. Furthermore, the number of suitable areas for R. speculum in the three future periods was predicted to increase compared with that in the current period; the results also revealed that the suitable areas for R. speculum will consistently expand northward. The results of this study deepen our understanding of the impacts of climate change on the distribution of R. speculum and should also be beneficial for farmers and government departments to develop appropriate and cost-effective pest management strategies to reduce economic damage to China’s agricultural systems.
Although the survival and development of human society is dependent on agriculture, thousands of insect pests seriously threaten it (Ali et al., 2023). These pests reduce yields and cause huge economic losses (Oliveira et al., 2013; 2014; Tonnang et al., 2022). As such, a major task in global agricultural production is to reduce the harm inflicted by insect pests.
Climate change is one of the gravest concerns globally. This term refers to the phenomenon of changes in environmental parameters such as temperature, humidity, and precipitation over a long period of time (Skendžić et al., 2021a). There is considerable evidence that climate change has significantly altered the distribution patterns of insect pests (Santana et al., 2019; Wang et al., 2020; Zhang et al., 2023; Chen et al., 2024). This has in turn raised numerous challenges for the management and control of such pests. For example, in newly invaded regions, there is a need to establish special monitoring and prevention measures. Unfortunately, given that ongoing climate change increasingly seems inevitable, new and growing challenges may be on the horizon in the fight against insect pests. In this context, the current priority is to use scientific methods to predict in advance the trends in the distribution of insect pests under future climate change conditions. This would facilitate the establishment of more cost-effective monitoring and prevention strategies.
Ricania speculum (Walker, 1851), commonly known as the black planthopper, belongs to the family Ricaniidae (Hemiptera: Fulgoroidea). This pest feeds on more than 60 plant species (Gao et al., 2022), including some of the most economically important crops, such as apples, coffee trees, oil palm, and tea trees (Li et al., 2011; Xu et al., 2013; Rossi and Lucchi, 2015). The plant damage caused by R. speculum is mainly due to its direct feeding. This pest often gathers on young branches, pierces the host tissue, and absorbs sap, which weakens the growth of the plant, induces leaf curvature and chlorosis, and in severe cases causes the branches to die (Yang, 1989; Yu, 2007; Mifsud et al., 2010; Wilson et al., 2016). China is one of the main regions of R. speculum infestation. In the 20th century, work began to document the damage caused by this pest to various plants in China (Xu and Zhong, 1988; Yang, 1989; Zhong, 1989), and subsequent studies reported pest outbreaks in regions with environmental conditions that are favorable to it (Li et al., 2015). Not only do such favorable conditions aggravate the damage to crops, but the fact that these conditions may change also complicates pest management efforts. There is thus an urgent need to understand the distribution of R. speculum. Determining its current distribution and predicting its future ones should aid in the development of scientific and effective pest management strategies, thereby reducing harm to agricultural production.
At present, the monitoring and management of R. speculum rely on traditional field surveys combined with chemical, biological, and cultural control methods. Field surveys typically involve visual inspections, trapping, and manual specimen collection to assess population distribution and density. Chemical control, such as insecticide applications, is commonly used to reduce pest populations in affected areas (Yu, 2007; Zhang and Xie, 2008; Xu et al., 2013). In some regions, biological control measures, including the introduction of natural predators, have been implemented to suppress R. speculum populations (Zhang and Xie, 2008; Xu et al., 2013). Additionally, cultural practices, such as habitat modification, are employed to limit its establishment and spread (Zhang and Xie, 2008). Although field surveys method provides information on the distribution of R. speculum populations, it has three major limitations. First, manual monitoring and investigations require substantial resources and usually rely on field sampling and manual records, which is laborious and prone to omissions or errors. Second, conventional methods can often only provide distribution data on a limited spatial scale, with it being difficult to cover large regions such as the whole of China. Finally, the changing distribution of R. speculum in the context of climate change cannot be obtained. Given these limitations of conventional methods, there is an urgent need to find a feasible alternative.
Species distribution models predict suitable areas for particular species based on the association between occurrence records and environmental predictors and have been broadly applied to insect pests (Kumar et al., 2016; de Paula et al., 2020; Xu et al., 2020; Hwang et al., 2022). These models can predict the suitable areas for insect pests at present and in the future at a macroscale and reduce monitoring costs and workload. Data on such areas constitute a key reference for the formulation of pest management strategies. To analyze the data, ecologists have developed a large number of species distribution models ranging from simple regression models to complex machine learning algorithms. Among these models, the maximum entropy (Maxent) model, a machine learning algorithm, is the most widely used. Its popularity is related to many factors, especially outstanding predictive accuracy and ease of use (Merow et al., 2013). Given this background, a Maxent model was here applied to the insect pest R. speculum. The objectives of this study are as follows: 1) to predict the suitable areas for R. speculum in China at present and in the future and 2) to investigate the dynamics of suitable areas under climate change.
We compiled 183 occurrence records of R. speculum in China from the literature, specimen collection records from Anshun and Guizhou Universities, and the Global Biodiversity Information Facility (GBIF; https://www.gbif.org/). For the GBIF data, we removed the occurrence records with common biological collection errors and high coordinate uncertainty (>20 km) based on the CoordinateCleaner Package in R 4.2.1 (Zizka et al., 2019). The occurrence records from the literature were georeferenced via Google Earth if information on the latitude and longitude was missing. Furthermore, to improve the performance of the species distribution model, we needed to resolve the potential issue of uneven sampling, such as due to field accessibility and collector preference, which often results in sampling bias in species occurrence records within a given area (Boria et al., 2014). Thus, we used the spThin package in R 4.2.1 to perform spatial thinning at a distance of 20 km (Aiello-Lammens et al., 2015). We finally obtained 111 occurrence records (Figure 1; Supplementary Table S1), which were mainly distributed in the central and southern provinces of China, such as Hubei, Guizhou, and Yunnan.

Figure 1. Occurrence records of R. speculum in China.
The current environmental variables included 19 bioclimatic variables (Bio1–Bio19) and 2 topographic variables (altitude and terrain roughness). Taking into account the collinearity between environmental variables is also a key step in building species distribution models. Therefore, we calculated the pairwise Pearson correlation coefficients (r) among the values of all pairs of the environmental variables obtained from the occurrence records using the sjPlot packages in R 4.2.1 (Lüdecke, 2024). We selected only one of a pair of variables if r between them was ≥0.8. Then, the variance inflation factor (VIF) of the selected environmental variables was also computed via usdm packages in R 4.2.1 (Naimi et al., 2014), and the environmental variables with the highest VIF values were subsequently removed in sequence until the remaining environmental variables had a VIF value of <5. Typically, VIF values of <5 among environmental variables indicate negligible collinearity. Finally, we selected six environmental variables: mean diurnal range (Bio2), mean temperature of the wettest quarter (Bio8), precipitation of the driest month (Bio14), precipitation seasonality (Bio15), precipitation of the warmest quarter (Bio18), and terrain roughness.
To predict suitable areas for habitation by the pest in the future, we accessed the future bioclimatic variables for the periods 2041–2060, 2061–2080, and 2081–2100 in the Coupled Model Intercomparison Project Phase 6. We used only future climate data provided by Shared Socioeconomic Pathways 245 (SSP245), a moderate-emissions scenario, to avoid extreme predictions (Hwang et al., 2022). Future bioclimatic variables in SSP245 are the means of the three Global Climate Models: CanESM5, IPSL-CM6A-LR, and MIROC6. These models have been widely used in previous studies (Zhao et al., 2024a; 2024b). Future data on terrain roughness are expressed as current terrain roughness because this should not change significantly over a period of several decades.
Bioclimatic variables and altitude were obtained from the WorldClim data website (https://www.worldclim.org), and terrain roughness was derived from the ENVIREM dataset created by Title and Bemmels (2018). The spatial resolution of these environmental variables is 2.5 arc-min.
As the parameter feature class (FC) and regularization multiplier (RM) in Maxent models are highly correlated with model complexity, previous research suggests they should be optimized (Phillips and Dudík, 2008; Muscarella et al., 2014; Radosavljevic et al., 2014). FC is an expanded set of transformations of the original environmental variables. Linear (L), quadratic (Q), product (P), threshold (T), and hinge (H) FC are provided in the Maxent model and can determine the kinds of constraints. Specifically, L ensures that the mean of the environmental variable is close to its observed value (Phillips et al., 2006). Q constrains the variance of the environmental variable close to its observed value (Phillips et al., 2006). P constrains the covariance of two variables to be close to its observed value (Phillips et al., 2006). T constrains the proportion of estimated distributions, with values for the variable above the threshold close to the observed proportion (Elith et al., 2011). H constrains the mean of the variable above the threshold under the estimated distribution close to its mean above the threshold in the observed proportion (Elith et al., 2011). The RM value seeks to balance the model fit and complexity (Velasco and González-Salazar, 2019). In short, the FC and RM values can be combined to achieve optimal model complexity.
We tuned the Maxent model by setting the different FC combinations (“L,” “H,” “LQ,” “LQH,” “LQHP,” “LQHPT”) and RM values ranging from 0.5 to 4, with increments of 0.5 (Zhao et al., 2024c). In this process, we used the checkerboard2 approach for partitioning the occurrence data and selected the best model using the lowest AICc score (Muscarella et al., 2014). Finally, a feature class combination of LQ and a regularization multiplier value of 0.5 were obtained to run the Maxent model. Meanwhile, we applied settings of 5-fold cross-validation, loglog output, and 10,000 background points throughout the study region (Zhao et al., 2024c). The workflow for fitting the Maxent model is also shown in Figure 2.
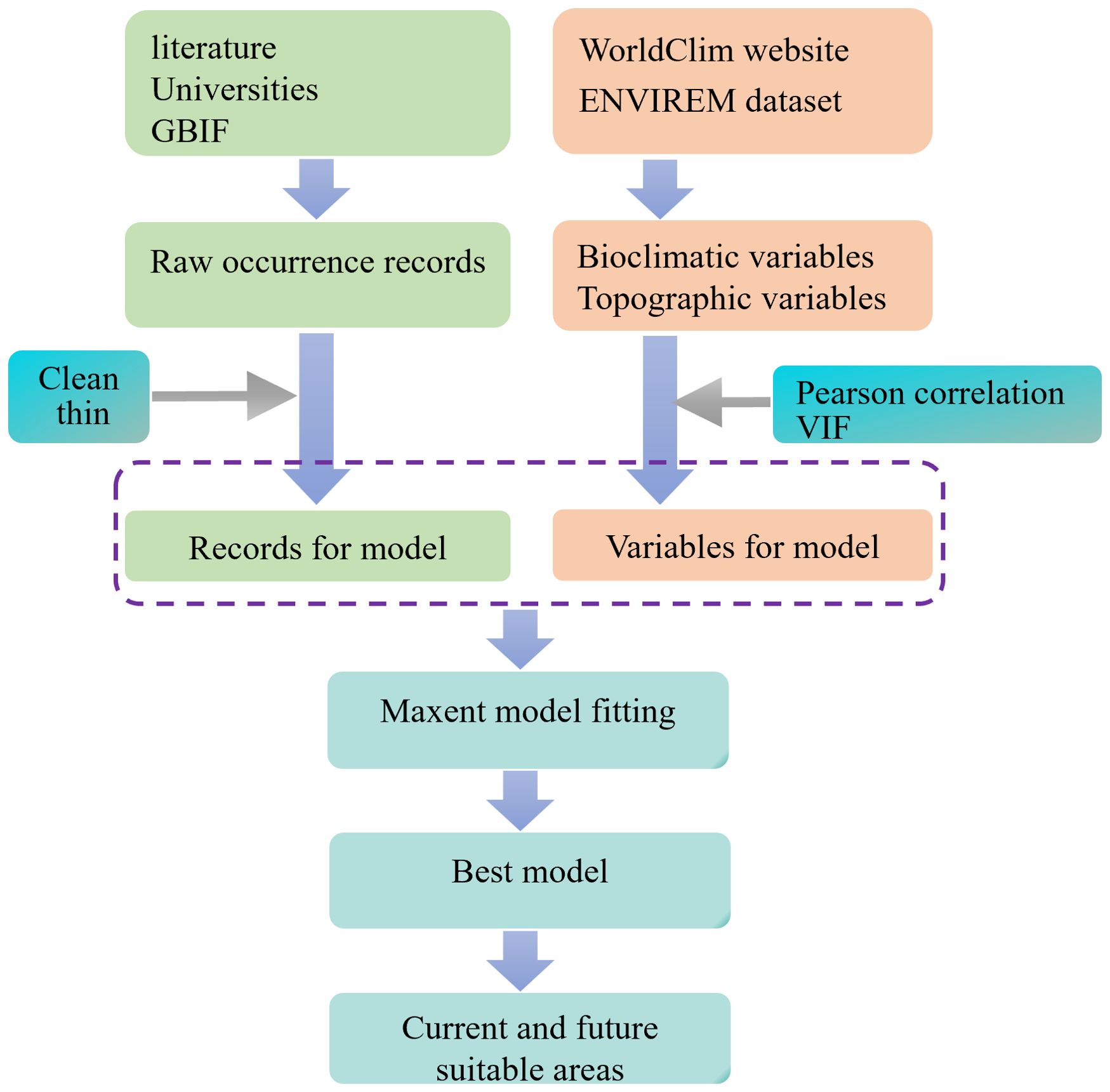
Figure 2. The workflow for fitting the Maxent model.
The relative contributions of the environmental variables to the Maxent model were estimated using the percent contribution and permutation importance metrics. To assess the performance of the Maxent model fitting, we employed the area under the receiver operating characteristic curve (AUC), with a value of >0.75 being considered useful (Elith, 2002). The above processes were performed in the raster (Hijmans, 2023), ENMeval (Kass et al., 2021), and dismo packages (Hijmans et al., 2023) in R 4.2.1. The code to build the Maxent model is in Supplementary Table S2.
For the Maxent model constructed in this study, we follow the eleven obligatory subsections of the ODMAP protocol (Zurell et al., 2020), which five data-related subsections (focal taxon/taxa, location, biodiversity data overview, predictor type, spatial and temporal scale), two conceptual subsections (hypotheses, underlying assumptions) and four technical subsections (SDM algorithms, model workflow, and the software and data used).
The Maxent model produces a continuous raster with values from 0 to 1 representing habitat suitability. Based on this, we defined suitable/unsuitable areas for R. speculum in China using the maximum training sensitivity plus specificity Cloglog threshold (0.228). This threshold was chosen because it is one of the best threshold selection methods (Liu et al., 2005). Furthermore, suitable areas were classified into three levels in terms of their suitability: low (0.228–0.4), moderate (0.4–0.6), and high (0.6–1) (Wei et al., 2020; Wang et al., 2021). This classification provides a clear gradient of habitat suitability, making it essential for targeted management efforts.
To summarize changes in distribution under climate change, we used the SDMtoolbox in ArcGIS 10.7 software (Brown, 2014) to obtain the distribution change types and centroid change. Distribution change types consist of contraction, expansion, and unchanged areas in the species distribution (in km2), while the centroid change reduces species distribution to a single central point and creates a vector describing the magnitude and direction of change over time (Brown, 2014)
The mean AUC value of five replicate runs of the Maxent model was 0.849 (Figure 3), suggesting that it can be used to predict suitable areas for R. speculum in China. The percent contribution metric showed that the mean diurnal range (Bio2) is the most important environmental variable limiting the distribution of R. speculum, followed by the mean temperature of the wettest quarter (Bio8) (Table 1). The remaining four variables are of relatively low importance. The permutation importance metric also demonstrated that these same two variables are the most important for the distribution of this species (Table 1). Response curves show the relationship between the presence probability of R. speculum and environmental variables (Figure 4). The presence probability increases with increases in the variables mean diurnal range (Bio2), precipitation in the driest month (Bio14), precipitation in the warmest quarter (Bio18), and terrain roughness, followed by decreases. In contrast, the presence probability continues to increase as the mean temperature of the wettest quarter (Bio8) increases. For precipitation seasonality (Bio15), the presence probability initially decreases and then increases.
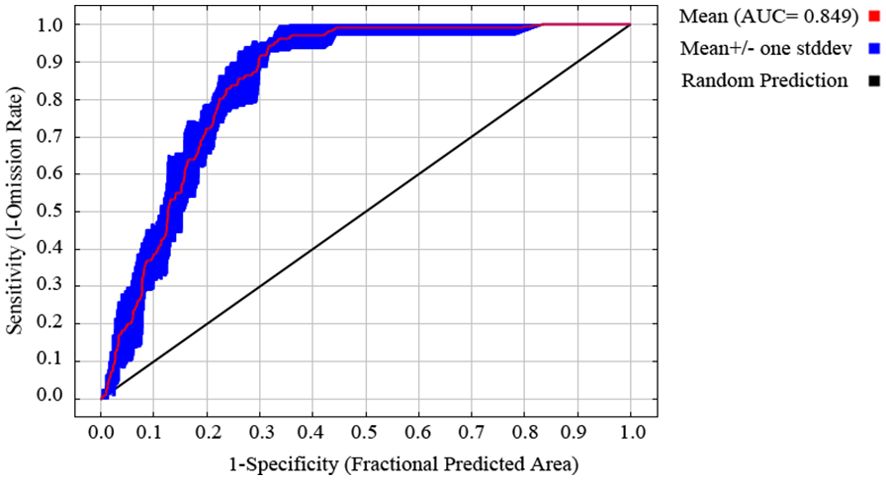
Figure 3. Receiver operating characteristic curve and AUC of Maxent.

Table 1. Importance of the environmental variables.
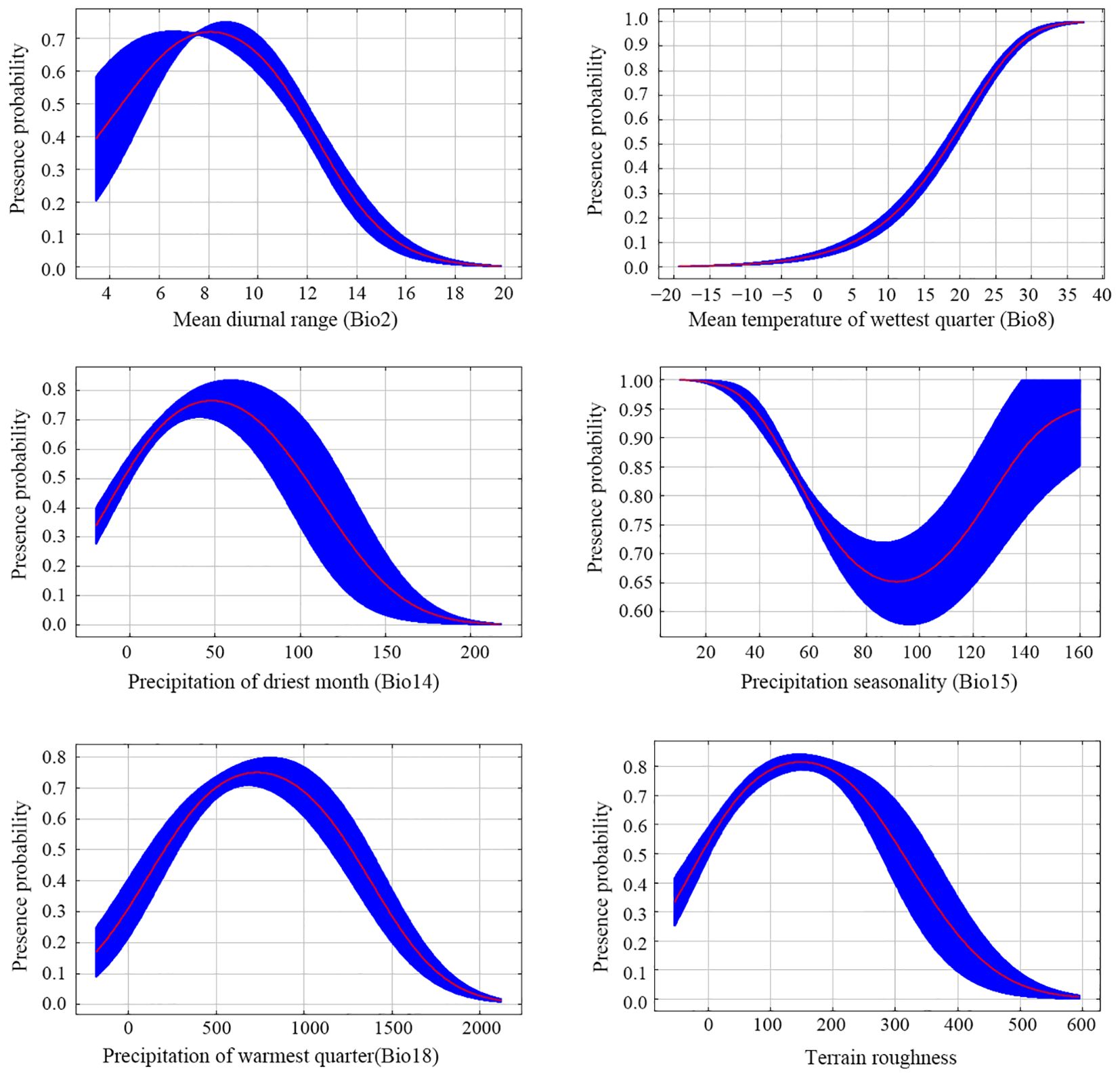
Figure 4. Response curves of the relationship between the presence probability of R. speculum and six environmental variables.
The total suitable area for R. speculum in China was predicted to be 3.02×106 km2 (Table 2) under present-day environmental conditions, being distributed mainly in the central and southern provinces of China (Figure 5), such as Henan, Anhui, Hubei, and Guizhou. In northern China, suitable areas were also detected in southern Liaoning and northern Xinjiang (Figure 5). However, such areas were not identified in Qinghai and Inner Mongolia (Figure 5). The distribution patterns of the three suitable areas present a nested structure. Specifically, the highly suitable area is located in the innermost region and occupies the largest geographical range (Figure 5), reaching 1.69 × 106 km2 (Table 2). The moderately suitable area (6.98 × 105 km2) is nested outside the highly suitable habitat, while the area with low suitability (6.34×105 km2) is nested outside the moderately suitable habitat (Figure 5).
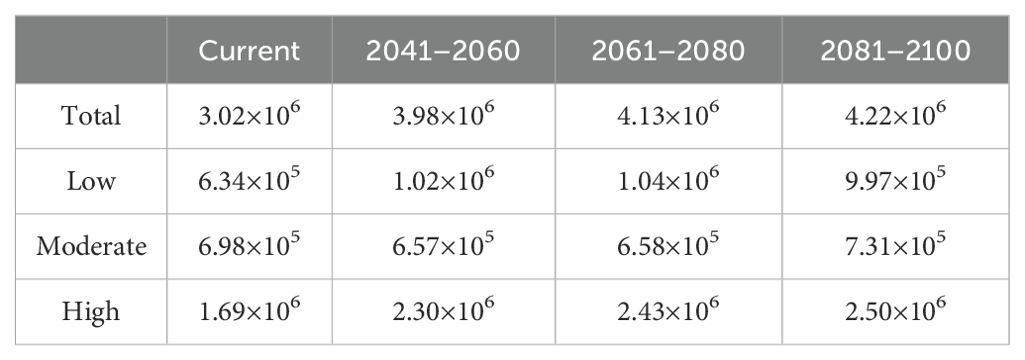
Table 2. Suitable areas (km2) for R. speculum under current and future environmental conditions.
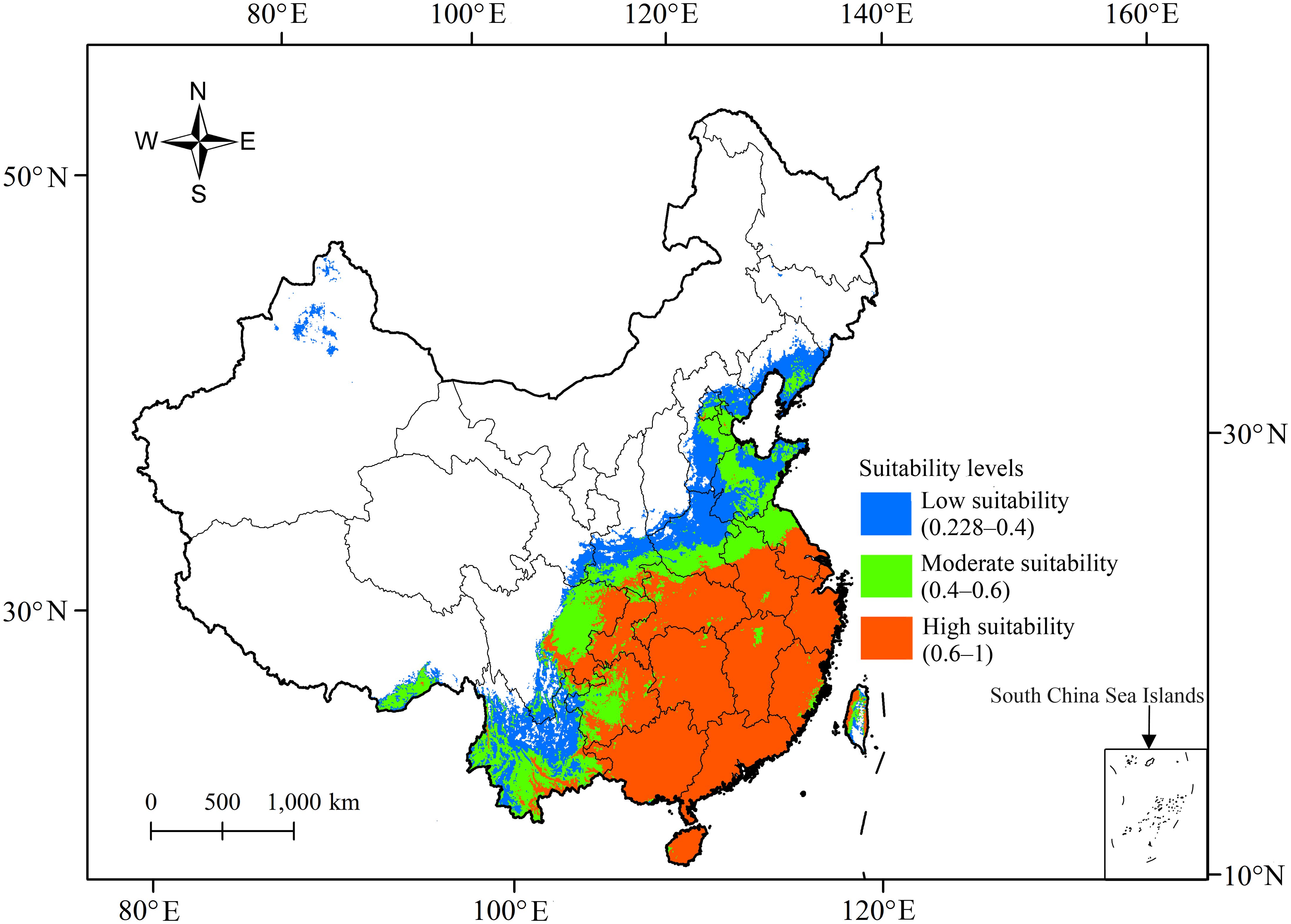
Figure 5. Areas suitable for R. speculum under the current environmental conditions in China.
The suitable area for R. speculum in China under future environmental conditions is larger than that under the conditions in the present day (Figure 6; Table 2). The suitable area was predicted to reach 3.98 × 106 km2 in the period 2041–2060 (Table 2). In this period, the areas with low and high suitability showed increases compared with their sizes in the present day (Figures 5, 6), while the area with moderate suitability was predicted to decrease (Table 2). The areas predicted to be suitable in the period 2061–2080 continued to increase in size, being larger than those of 2041–2060 and the present day (Table 2). Meanwhile, in the period 2081–2100, the suitable areas reach their greatest size, predicted to be 4.22×106 km2 (Table 2). It was further found that the areas with moderate and high suitability in this period were also the largest among all of the periods (Table 2).

Figure 6. Areas suitable for R. speculum under future environmental conditions in China.
In terms of the types of change in the suitable areas for R. speculum in China, only “expansion” and “unchanged” were identified under the three future periods (Figure 7). In 2041–2060, the expansive suitability area was predicted to be 9.56×105 km2 (Table 3), mainly concentrated in northern China, especially in Heilongjiang, Liaoning, Jilin, and Xinjiang (Figure 7). The unchanged suitability area was significantly larger than the expansive suitability area (Figure 7), covering 3.02×106 km2 (Table 3). In 2061–2080 and 2081–2100, the expansive suitability area is predicted to continue to increase northward (Figure 7), while the unchanged suitability area is predicted to be consistent with that in the period 2041–2060 (Table 3). Centroid change more clearly shows the trend of changes in the areas suitable for R. speculum in China under future environmental conditions (Figure 8). Overall, the suitable areas move northward, at distances of 442.25 km for 2041–2060, 55.29 km for 2061–2080, and 31.31 km for 2081–2100 (Figure 8).
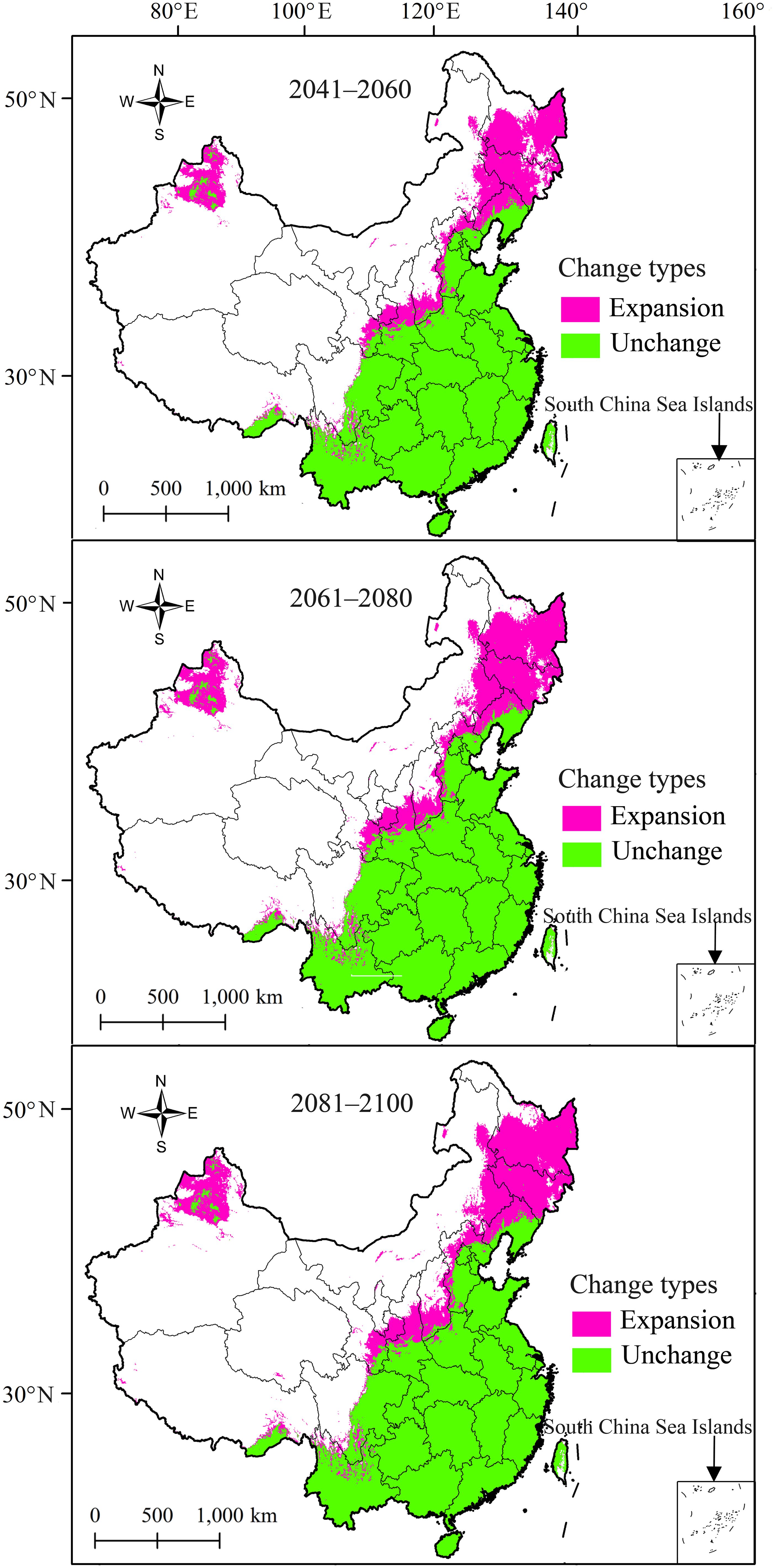
Figure 7. Change in area suitable for R. speculum in China compared with that under current environmental conditions.
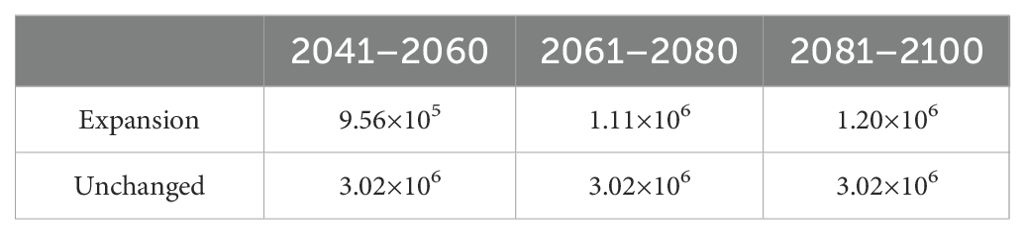
Table 3. Change in the suitable area (km2) for R. speculum under current and future environmental conditions.
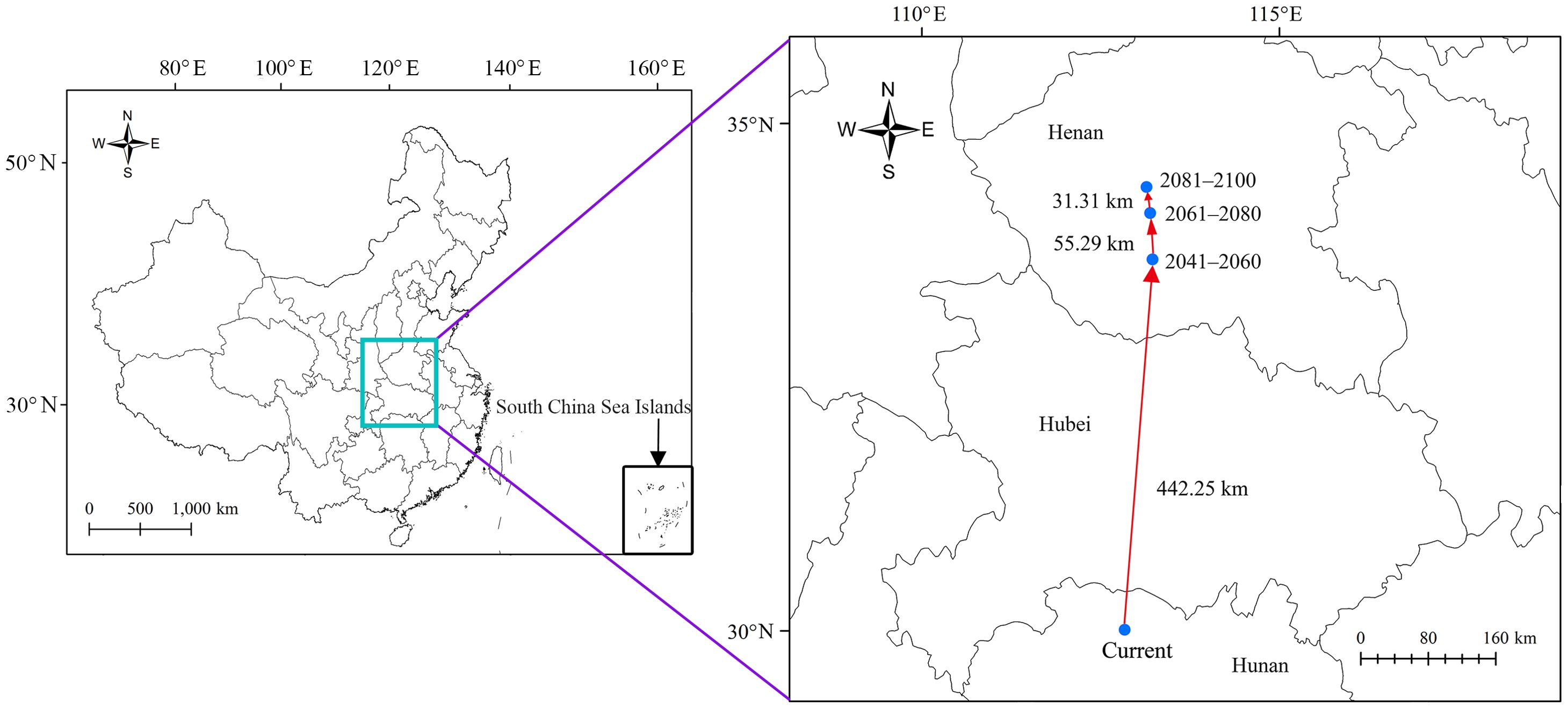
Figure 8. Centroid changes in R. speculum under future environmental conditions in China.
In this study, a Maxent model was constructed to predict the suitable areas for the agricultural pest R. speculum in China at present and in the future. The model results revealed the relative importance of various environmental variables and further indicated that the temperature-related variables (Bio2 and Bio8) had a much greater impact on this species’ distribution than the precipitation- and topography-related ones. This can be explained by the physiological characteristics of insects, in that their body temperature depends on the ambient temperature (Skendžić et al., 2021a), and thus temperature may be the most important environmental variable affecting insect distribution (Kocmánková et al., 2010).
Because of the dominant role of temperature-related variables in the distribution of R. speculum, only small or no suitable areas for this species were detected in China’s colder regions, such as Heilongjiang, Jilin, Xizang, and Inner Mongolia (Figure 5). Our study also predicted that several provinces without occurrence records are suitable habitats for R. speculum, such as Xizang, Xinjiang, Hainan, and Taiwan. At the same time, although Yunnan and Guizhou have the largest numbers of occurrence records and large numbers of suitable areas, the number of highly suitable areas is not large. In contrast, almost the entire land area of some provinces with fewer distribution records (e.g., Hunan, Jiangxi, and Guangdong) was predicted to be highly suitable for this species. These findings suggest that there is a lack of uniformity between different provinces in terms of the surveys performed to date, resulting in major underestimation of the actual distribution of R. speculum in some provinces. In other words, the crops in these provinces are very likely to be attacked by pests, but this has yet to be recorded. Field collection work can be carried out in regions currently lacking distribution records but predicted to contain suitable habitats, to test the validity of the model prediction.
It is well established that insects are sensitive to temperature (Lehmann et al., 2020). Species that fail to adapt and evolve to rising temperatures generally struggle to maintain their populations, whereas other species can thrive and reproduce quickly (Skendžić et al., 2021a). Rising temperatures have thus significantly altered the geographical distributions of insect pests (Bebber et al., 2013; Netherer and Schopf, 2010; Jactel et al., 2019; Skendžić et al., 2021b). From an agricultural perspective, particularly with the goal of improving crop yields, there is an expectation that future climate warming might lead to the shrinkage or even disappearance of pest populations. However, contrary to this expectation, numerous studies have shown that many insect pests are projected to expand their distribution ranges under future climate scenarios (Xu et al., 2020; Zhang et al., 2022; Chen et al., 2024; Wei et al., 2024). This expansion highlights the fact that global warming is creating increasingly favorable conditions for insect pests, enabling their spread and colonization into regions previously unsuitable for them, as observed in Heilongjiang and Jilin regions (Figure 7). The expansion of pest distributions inevitably leads to a range of adverse effects. These include reduced crop yields, along with the costs of establishing the infrastructure, resources, and expertise to address emerging pest populations, which are ultimately borne by farmers and the government. In light of these challenges, it is crucial to develop the tools to predict pest distributions under future climate scenarios, as this can help to prevent pest invasions and mitigate potential damage before it occurs.
The suitable areas for R. speculum and their patterns of change as predicted in this study are helpful for farmers and government departments to formulate pest management strategies. Specifically, such information tells them which regions should be prioritized and what appropriate management strategies should be implemented depending on the particular trends. For instance, highly suitable areas face the greatest risk of species occurrence and spread, so pest management resources (e.g., pesticide application and population monitoring) should be concentrated in these areas to enable more effective control. In addition, in suitable areas that are predicted to expand under climate change, rigorous preventive measures, including stringent quarantine protocols, should be formulated to limit the invasion of R. speculum from other regions. To achieve these goals, local governments should provide financial support to facilitate timely and effective action.
Species distribution models are widely recognized as powerful tools for predicting the distribution of insect pests, and offer insights that are crucial for pest management strategies. However, they are not without limitations, particularly when the key drivers of pest distribution are omitted. For example, the survival and distribution of R. speculum are influenced by variables related to the surrounding biota, including competitive interactions and the presence of associated plants. Extensive research has demonstrated that integrating biotic variables into species distribution models can significantly enhance their accuracy (Giannini et al., 2013; Kass et al., 2019; Stephenson et al., 2022). However, a major limitation of our modeling process is the lack of availability of rasterized data for the relevant biotic variables associated with R. speculum. Consequently, these biotic variables could not be included in the current modeling framework, potentially reducing the accuracy of the predictions. In future work, it will be essential to address this limitation to refine the model and improve its utility for understanding the distribution of R. speculum.
The current study fitted a Maxent model to predict the suitable areas for the insect pest R. speculum in China under both current and future environmental conditions. It also investigated the dynamics of these areas over time. The results revealed that the suitable areas for this species under present-day environmental conditions were primarily concentrated in the central and southern provinces of the country, with a nested distribution pattern observed among areas of low, moderate, and high suitability. Under future environmental conditions, the areas predicted to be suitable were larger than those under current environmental conditions, suggesting that the population will increase in size and demonstrate a northward shift. The data on areas predicted to be suitable for R. speculum in this study provide valuable information for the management of this pest and efforts to control it in response to changing environmental conditions.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author/s.
The manuscript presents research on animals that do not require ethical approval for their study.
XF: Conceptualization, Writing – original draft, Writing – review & editing. ZZ: Data curation, Formal analysis, Writing – review & editing. YW: Data curation, Formal analysis, Writing – review & editing. YZ: Data curation, Funding acquisition, Project administration, Supervision, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the Scientific Research Platform of Education Department of Guizhou Province (No. Qianjiaoji(2022)052), Guizhou Provincial Science and Technology Projects (No. QKHJC(2024)youth290), Scientific Research Project of High Education Department of Guizhou Province (No. Qianjiaoji (2024) 214), Doctoral Fund Project of Anshun University (No. 202215), and Anshun Science and Technology Planning Project (No. Anshikeping (2020)2).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1549532/full#supplementary-material
Aiello-Lammens M. E., Boria R. A., Radosavljevic A., Vilela B., Anderson R. P. (2015). spThin: an r package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 38, 541–545.
Ali M. A., Abdellah I. M., Eletmany M. R. (2023). Towards sustainable management of insect pests: Protecting food security through ecological intensification. Int. J. Chem. Biochem. Sci. 24, 386–394.
Bebber D. P., Ramotowski M. A., Gurr S. J. (2013). Crop pests and pathogens move polewards in a warming world. Nat. Clim. Change 3, 985–988. doi: 10.1038/nclimate1990
Boria R. A., Olson L. E., Goodman S. M., Anderson R. P. (2014). Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 275, 73–77. doi: 10.1016/j.ecolmodel.2013.12.012
Brown J. L. (2014). SDMtoolbox: a python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 5, 694–700. doi: 10.1111/mee3.2014.5.issue-7
Chen J., Jiang K., Li Y., Wang S., Bu W. (2024). Climate change effects on the diversity and distribution of soybean true bugs pests. Pest Manage. Sci. 80, 5157–5167. doi: 10.1002/ps.v80.10
de Paula A. S., Barreto C., Neven L. (2020). Potential distribution of Nysius simulans (Hemiptera: Lygaeidae) in soybean crops in South America under current and future climate. J. Econ. Entomol. 113, 1702–1710. doi: 10.1093/jee/toaa089
Elith J. (2002). “Quantitative Methods for Modeling Species Habitat: Comparative Performance and an Application to Australian Plants,” in Quantitative Methods for Conservation Biology. Eds. Ferson S., Burgman M. (New York: Springer).
Elith J., Phillips S. J., Hastie T., Dudík M., Chee Y. E., Yates C. J. (2011). A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 17, 43–57. doi: 10.1111/j.1472-4642.2010.00725.x
Gao T., Brożek J., Dai W. (2022). Fine-structural morphology of the mouthparts of the polyphagous invasive planthopper, Ricania speculum (Walker)(Hemiptera: Fulgoromorpha: Ricaniidae). Insects 13, 843. doi: 10.3390/insects13090843
Giannini T. C., Chapman D. S., Saraiva A. M., Alves-dos-Santos I., Biesmeijer J. C. (2013). Improving species distribution models using biotic interactions: a case study of parasites, pollinators and plants. Ecography 36, 649–656. doi: 10.1111/j.1600-0587.2012.07191.x
Hijmans R. (2023). raster: Geographic Data Analysis and Modeling (R package version 3.6-23). Vienna, Austria: R Foundation for Statistical Computing. Available online at: https://CRAN.R-project.org/package=raster.
Hijmans R. J., Phillips S., Leathwick J., Elith J. (2023). dismo: Species Distribution Modeling (R package version 1.3-14). Vienna, Austria: R Foundation for Statistical Computing. Available online at: https://CRAN.R-project.org/package=dismo.
Hwang J. H., Kim S. H., Yoon S., Jung S., Kim D. H., Lee W. H. (2022). Evaluation of spatial distribution of three major Leptocorisa (Hemiptera: Alydidae) pests using MaxEnt model. Insects 13, 750. doi: 10.3390/insects13080750
Jactel H., Koricheva J., Castagneyrol B. (2019). Responses of forest insect pests to climate change: not so simple. Curr. Opin. Insect Sci. 35, 103–108. doi: 10.1016/j.cois.2019.07.010
Kass J. M., Anderson R. P., Espinosa-Lucas A., Juárez-Jaimes V., Martínez-Salas E., Botello F., et al. (2019). Biotic predictors with phenological information improve range estimates for migrating monarch butterflies in Mexico. Ecography 43, 341–352. doi: 10.1111/ecog.2020.v43.i3
Kass J. M., Muscarella R., Galante P. J., Bohl C. L., Buitrago-Pinilla G. E., Boria R. A., et al. (2021). ENMeval 2.0: Redesigned for customizable and reproducible modeling of species’ niches and distributions. Methods Ecol. Evol. 12, 1602–1608. doi: 10.1111/2041-210X.13628
Kocmánková E., Trnka M., Juroch J., Dubrovský M., Semerádová D., Možný M., et al. (2010). Impact of climate change on the occurrence and activity of harmful organisms. Plant Prot. Sci. 45, S48–S52. doi: 10.17221/2835-PPS
Kumar S., Yee W. L., Neven L. G. (2016). Mapping global potential risk of establishment of Rhagoletis pomonella (Diptera: Tephritidae) using MaxEnt and CLIMEX niche models. J. Econ. Entomol. 109, 2043–2053. doi: 10.1093/jee/tow166
Lehmann P., Ammunét T., Barton M., Battisti A., Eigenbrode S. D., Jepsen J. U., et al. (2020). Complex responses of global insect pests to climate warming. Front. Ecol. Environ. 18, 141–150. doi: 10.1002/fee.v18.3
Li J. L., Shao Y. H., Tang J. C., Li J. X., Tang H., Li D. (2011). The spatial distribution and the sampling technique of adult Ricania speculum (Walker). Guangdong Agric. Sci. 19, 81–83.
Li Z. G., Wang L. C., Ye J. W., Xu H. L., Dai J. Q., Han S. C. (2015). Tropism of Ricania speculum (Walker) adults to different colors and host plants in mangrove forest. J. South. Agric. 46, 1624–1627. doi: 10.3969/jissn.2095-1191.2015.09.1624
Liu C., Berry P. M., Dawson T. P., Pearson R. G. (2005). Selecting thresholds of occurrence in the prediction of species distributions. Ecography 28, 385–393. doi: 10.1111/j.0906-7590.2005.03957.x
Lüdecke D. (2024). sjPlot: Data Visualization for Statistics in Social Science (R package version 2.8.17). Vienna, Austria: R Foundation for Statistical Computing. Available online at: https://CRAN.R-project.org/package=sjPlot.
Merow C., Smith M. J., Silander J. A. (2013). A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings matter. Ecography 36, 1058–1069. doi: 10.1111/j.1600-0587.2013.07872.x
Mifsud D., Cocquempot C., Mühlethaler R., Wilson M., Streito J. C. (2010). “Other Hemiptera Sternorrhyncha (Aleyrodidae, Phylloxeroidea, and Psylloidea) and Hemiptera Auchenorrhyncha,” in Alien terrestrial arthropods of Europe. Ed. Roques A. (Bulgaria: BioRisk).
Muscarella R., Galante P. J., Soley-Guardia M., Boria R. A., Kass J. M., Uriarte M., et al. (2014). ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 5, 1198–1205. doi: 10.1111/mee3.2014.5.issue-11
Naimi B., Hamm N., Groen T. A., Skidmore A. K., Toxopeus A. G. (2014). Where is positional uncertainty a problem for species distribution modelling. Ecography 37, 191–203. doi: 10.1111/j.1600-0587.2013.00205.x
Netherer S., Schopf A. (2010). Potential effects of climate change on insect herbivores in European forests—General aspects and the pine processionary moth as specific example. For. Ecol. Manage. 259, 831–838.
Oliveira C. M., Auad A. M., Mendes S. M., Frizzas M. R. (2013). Economic impact of exotic insect pests in Brazilian agriculture. J. Appl. Entomol. 137, 1–15. doi: 10.1111/jen.2012.137.issue-1-2
Oliveira C. M., Auad A. M., Mendes S. M., Frizzas M. R. (2014). Crop losses and the economic impact of insect pests on Brazilian agriculture. Crop Protect. 56, 50–54. doi: 10.1016/j.cropro.2013.10.022
Phillips S. J., Anderson R. P., Schapire R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecol. Model. 190, 231–259. doi: 10.1016/j.ecolmodel.2005.03.026
Phillips A. J., Dudík M. (2008). Modeling of species distributions with Maxent new extensions. Ecography 31, 161–175. doi: 10.1111/j.0906-7590.2008.5203.x
Radosavljevic A., Anderson R. P., Araújo M. (2014). Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 41, 629–643. doi: 10.1111/jbi.2014.41.issue-4
Rossi E., Lucchi A. (2015). The Asian planthopper Ricania speculum (Walker) (Homoptera: Ricaniidae) on several crops in Italy: a potential threat to the EPPO region? EPPO Bull. 45, 119–122. doi: 10.1111/epp.2015.45.issue-1
Santana P. A., Kumar L., Da Silva R. S., Pereira J. L., Picanço M. C. (2019). Assessing the impact of climate change on the worldwide distribution of Dalbulus maidis (DeLong) using MaxEnt. Pest Manage. Sci. 75, 2706–2715. doi: 10.1002/ps.v75.10
Skendžić S., Zovko M., Pajač Živković I., Lešić V., Lemić D. (2021b). Effect of climate change on introduced and native agricultural invasive insect pests in Europe. Insects 12, 985. doi: 10.3390/insects12110985
Skendžić S., Zovko M., Živković I. P., Lešić V., Lemić D. (2021a). The impact of climate change on agricultural insect pests. Insect pests 12, 440. doi: 10.3390/insects12050440
Stephenson F., Gladstone-Gallagher R. V., Bulmer R. H., Thrush S. F., Hewitt J. E. (2022). Inclusion of biotic variables improves predictions of environmental niche models. Divers. Distrib. 28, 1373–1390. doi: 10.1111/ddi.13546
Title P. O., Bemmels J. B. (2018). ENVIREM: an expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche modeling. Ecography 41, 291–307. doi: 10.1111/ecog.2017.v41.i2
Tonnang H. E., Sokame B. M., Abdel-Rahman E. M., Dubois T. (2022). Measuring and modelling crop yield losses due to invasive insect pests under climate change. Curr. Opin. Insect Sci. 50, 100873. doi: 10.1016/j.cois.2022.100873
Velasco J. A., González-Salazar C. (2019). Akaike information criterion should not be a “test” of geographical prediction accuracy in ecological niche modelling. Ecol. Inform. 51, 25–32. doi: 10.1016/j.ecoinf.2019.02.005
Wang F., Wang D., Guo G., Zhang M., Lang J., Wei J. (2021). Potential distributions of the invasive barnacle scale Ceroplastes cirripediformis (Hemiptera: Coccidae) under climate change and implications for its management. J. Econ. Entomol. 114, 82–89. doi: 10.1093/jee/toaa245
Wang R. L., Yang H., Wang M. T., Zhang Z., Huang T. T., Wen G., et al. (2020). Predictions of potential geographical distribution of Diaphorina citri (Kuwayama) in China under climate change scenarios. Sci. Rep. 10, 9202. doi: 10.1038/s41598-020-66274-5
Wei J., Niu M., Zhang H., Cai B., Ji W. (2024). Global potential distribution of invasive species Pseudococcus viburni (Hemiptera: Pseudococcidae) under climate change. Insects 15, 195. doi: 10.3390/insects15030195
Wei J., Peng L., He Z., Lu Y., Wang F. (2020). Potential distribution of two invasive pineapple pests under climate change. Pest Manage. Sci. 76, 1652–1663. doi: 10.1002/ps.v76.5
Wilson S. W., Rossi E., Lucchi A. (2016). Descriptions of the adult genitalia and immatures of the Asian planthopper Ricania speculum (Hemiptera: Fulgoroidea: Ricaniidae) recently introduced to Italy. Ann. Entomol. Soc Am. 109, 899–905. doi: 10.1093/aesa/saw056
Xu D. P., Li X. Y., Jin Y. W., Zhuo Z. H., Yang H. J., Hu J. M., et al. (2020). Influence of climatic factors on the potential distribution of pest Heortia vitessoides Moore in China. Glob. Ecol. Conserv. 23, e01107. doi: 10.1016/j.gecco.2020.e01107
Xu H., Liu Z., Bao Q., Zeng L., Jiang S. (2013). Damage and control of Ricania speculum in mangrove in Futian, Shenzhen. Guangdong Sci. Tech. 29, 26–30.
Xu G. J., Zhong S. T. (1988). A preliminary report on the kinds of lanternflies in citrus orchard in Yichang Hubei. J. Huazhong Agric. Univ. 7, 196–198.
Yang C. T. (1989). Ricaniidae of Taiwan (Homoptera: Fulgoroidea) (Taiwan: Taiwan Museum Special Publication Series).
Yu A. (2007). Biological characteristics of Ricania speculum and its control. Jiangxi For. Sci. Technol. 3, 34–35. doi: 10.16259/j.cnki.36-1342/s.2007.03.014
Zhang Y., Hughes A. C., Zhao Z., Li Z., Qin Y. (2022). Including climate change to predict the global suitable area of an invasive pest: Bactrocera correcta (Diptera: Tephritidae). Jiangxi For. Sci. Technol. 34, e02021. doi: 10.1016/j.gecco.2022.e02021
Zhang X. D., Xie S. G. (2008). Damage and control strategies of R. speculum on pomelo in Chenzhou. South China Fruits 37, 26–27. doi: 10.13938/j.issn.1007-1431.2008.03.012
Zhang Y. B., Zhao Z. X., Wang Y. J., Liu T. L. (2023). Suitable habitats for Cicadella viridis and Evacanthus interruptus (Hemiptera: Cicadellidae) with global climate change. J. Entomol. Sci. 58, 215–229. doi: 10.18474/JES22-36
Zhao Z. X., Feng X. L., Wang Y. J., Zhou Z. X., Zhang Y. B. (2024a). Potential suitability areas of Sitobion miscanthi in China based on the MaxEnt model: implications for management. Crop Prot 183, 106755. doi: 10.1016/j.cropro.2024.106755
Zhao Z. X., Yang L., Chen X. S. (2024b). Current and future potential distribution of two bamboo pests in China: Anaka burmensis and Cicadella viridis (Hemiptera, Cicadellidae). Zookeys 1203, 197–210. doi: 10.3897/zookeys.1203.118978
Zhao Z. X., Yang L., Long J. K., Chang Z. M., Chen X. S. (2024c). Predicting suitable areas for Metcalfa pruinosa (Hemiptera: Flatidae) under climate change and implications for management. J. Insect Sci. 24, 7. doi: 10.1093/jisesa/ieae053
Zhong S. T. (1989). Biological observation and control of Ricania speculum in citrus orchard. Chin. Citrus 18, 32. doi: 10.13938/j.issn.1007-1431.1989.04.018
Zizka A., Silvestro D., Andermann T., Azevedo J., Duarte Ritter C., Edler D., et al. (2019). CoordinateCleaner: Standardized cleaning of occurrence records from biological collection databases. Methods Ecol. Evol. 10, 744–751.
Keywords: climate change, MAXENT model, suitable area, pest management, Ricania speculum
Citation: Feng X, Zhao Z, Wang Y and Zhang Y (2025) Potentially suitable areas for Ricania speculum (Hemiptera: Ricaniidae) in China under climate change. Front. Ecol. Evol. 13:1549532. doi: 10.3389/fevo.2025.1549532
Received: 21 December 2024; Accepted: 06 March 2025;
Published: 27 March 2025.
Edited by:
Sergio Noce, Foundation Euro-Mediterranean Center on Climate Change (CMCC), ItalyReviewed by:
Maurici Ruiz, University of the Balearic Islands, SpainCopyright © 2025 Feng, Zhao, Wang and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yubo Zhang, enliNzgxMjEwQHNpbmEuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.