 Yujie Si1†
Yujie Si1† Hongxiang Li
Hongxiang Li Cunqi Liu
Cunqi Liu- 1College of Life Sciences, Hebei University, Baoding, China
- 2Hebei Key Laboratory of Wetland Ecology and Conservation, Hengshui University, Hengshui, China
- 3National Key Lab of Regional and Urban Ecological Protection, Chinese Research Academy of Environmental Sciences, Beijing, China
Distance-decay relationships (DDRs) are a useful method for describing the spatial distribution of biological communities. However, the patterns of macroinvertebrate DDRs and the mechanisms of community assembly in continuous lotic-lentic (river-lake) systems remain poorly understood. We compared the spatial and temporal patterns and biogeographic patterns of macroinvertebrate communities in different water types (river vs. lake) in the temperate zone of China. Data were collected from 152 sampling sites in the Daqing River Basin over two years, and multivariate statistical analyses were conducted. These results provide the first description of biogeographical patterns of macroinvertebrate communities in the Daqing River in temperate China. Macroinvertebrates from both the river and lake exhibited contrasting community compositions, likely due to habitat differences. All macroinvertebrate communities showed a significant distance-decay pattern in both waterbody types, with stronger DDRs in the lake. In the lake, spatial and environmental variables contributed to the spatial distribution of macroinvertebrate communities, while in the river, spatial variables were slightly more important than environmental variables. Spatial variables influenced macroinvertebrate community distribution directly and impacted environmental variables indirectly. Importantly, these communities were strongly driven by stochastic processes. Additionally, macroinvertebrate taxa formed ecologically and taxonomically distinct groups, potentially structured by both deterministic and stochastic processes. The normalized stochasticity ratio (NST) quantified the community-building process based on the neutral model, indicating that deterministic processes may dominate in rivers, while stochastic processes may dominate in lakes. The findings highlight the contrasting roles of deterministic and stochastic processes in shaping macroinvertebrate communities in lotic and lentic systems, providing new insights into the mechanisms of community assembly in continuous river-lake ecosystems.
1 Introduction
Studying species spatial distribution patterns and the underlying mechanisms is fundamental for community ecology, macroecology, and biogeography (Sutherland et al., 2013). Turnover, nestedness, species replacement, and species richness differences can inform the determination of community spatial distribution patterns (Soininen et al., 2011; Carvalho et al., 2012). The spatial distribution patterns of macroinvertebrate communities in rivers vary significantly across different seasons (Yang et al., 2023). Additionally, macroinvertebrate communities exhibit distinct spatial distribution patterns in lakes, displaying varying patterns across different habitat types such as macrophyte or phytoplankton-dominated waters and open water (Dong et al., 2024). However, further understanding of the variation in the spatial distribution of macroinvertebrates in different waterbody types in a continuum river-lake system is needed.
Distance decay relationships (DDRs) are commonly used to examine spatial distribution patterns in community ecology (Soininen et al., 2011). They demonstrate how community similarity decreases with increasing geographical or environmental distance. This decay can represent community composition across a gradient of spatial scales (Anderson et al., 2011). Recent studies of spatial distribution in macroinvertebrate communities have mainly documented the α diversity dynamics in rivers (Li et al., 2020; Buffagni, 2021) or lakes (Zhang et al., 2019; Yang et al., 2020). Only one study has considered the spatial distribution differences between macroinvertebrate communities in rivers and lakes (Zhao et al., 2024). Therefore, studies on the DDR patterns of macroinvertebrate communities in river-lake systems are limited, and it is still unknown whether macroinvertebrate communities exhibit comparable DDR patterns in river-lake systems to those observed in soil bacteria, plankton, and fungal communities (Martiny et al., 2011; Jin et al., 2022; Zhao et al., 2019).
Clarifying the key driving factors of the spatial structure of communities is vital in community ecology (Cottenie, 2005). At large spatial scales, community similarity tends to decrease with distance due to the reduction in environmental similarity, dispersal limitations, and niche width differences (Nekola and White, 1999). In this scenario, the beta diversity of highly dispersing taxa demonstrates a greater environmental influence and a weaker spatial structure (limited by dispersal), whereas the beta diversity of poorly dispersing taxa exhibits a stronger spatial structure and less environmental influence (Li et al., 2022). Previous studies have indicated that both environmental and spatial factors contribute to community dissimilarity. However, the relative importance of these factors may be contingent upon spatial extent, seasonal dynamics, or geographical connectivity (Heino et al., 2012; Chaparro et al., 2018; Jamoneau et al., 2018). Compared with lentic habitats in lakes, lotic habitats in rivers have a faster water velocity, a greater degree of spatial heterogeneity and oxygen concentration, and larger - sized substrate particles (Thorp et al., 2001). Macroinvertebrates exhibit diverse dispersal modes (e.g., passive and/or active movement along river corridors or flying in the air) and a wide range of dispersal abilities; hence, they are optimal study organisms for understanding the mechanisms regulating beta diversity (Heino and Peckarsky, 2014). Despite increasing research on the relationship between macroinvertebrates and abiotic factors, there are still gaps in understanding the impact of their synergistic interactions on macroinvertebrate distribution (Heino et al., 2021; Lemm et al., 2021). Furthermore, few studies have been conducted on the influence of abiotic factors on macroinvertebrates in different waterbody types (Zhao et al., 2024).
Community assembly processes are an important research topic in aquatic ecology. The relative importance of stochastic and deterministic processes in aquatic community assembly has received considerable attention (Qu et al., 2018; Göthe et al., 2017). Aquatic community can be divided into two distinct categories: species sorting (deterministic) and neutral (stochastic) (Ning et al., 2020). The species sorting by niche differentiation hypothesis is that heterogeneity in environmental factors can shape local communities (Leibold et al., 2004). In contrast, neutral processes, such as stochastic dispersal and ecological drift, can influence community assembly. Neutral theory posits that all species are functionally equivalent and that drift is the primary driver of community composition (Hubbell, 2001). Östman et al. (2010) proposed that the relative importance of species sorting and neutral processes is contingent upon the environmental heterogeneity among sites, with greater environmental variation facilitating species sorting. Species sorting and dispersal are thought to be fundamental and competing processes that determine community structure (Leibold et al., 2004; Astorga et al., 2012). Species sorting includes environmental filtration and species interaction, whereas dispersal is a key regional process that refers to the movement of individuals between local communities in a landscape. Small-scale dispersal occurs uniformly, while large-scale dispersal is limited by distance (Cottenie, 2005).
Here, we focused on macroinvertebrate communities in river and lake systems to examine the joint effects of environmental filtering and spatial structuring. We hypothesized that: (1) in most cases, macroinvertebrate communities have significant DDR patterns in lakes and rivers, but the slopes in the lake are steeper than that those in the river; (2) macroinvertebrate communities are primarily affected by spatial and environmental factors in both the river and lake; and (3) the role of stochastic processes is more prominent in the lake, but deterministic processes are more prominent in the river.
2 Materials and methods
2.1 Study area and sampling sites
The Daqing River system (38°10′–40°102′ N, 113°39′′–117°34′′ E) is located in the central part of the Haihe Basin and crosses 64 counties and districts in the Shanxi, Hebei, Beijing, and Tianjin provinces. Its upper reaches are divided into two branches, the south and the north, which merge into Baiyangdian Lake (BYDL). The BYDL (38°3′–40°4′ N, 113°39′′–116°12′′ E), located in the northern part of the North China Plain, is a major part of the Daqing River water system (Zhao et al., 2011) (Figure 1). The BYDL is a flood-relieving, stagnant, and comprehensively utilized lake connecting the Daqing River’s mountain areas and the plains. The Daqing River’s lower reaches enter the sea through the mainstream of Hai River in the north and Duliujian River in the south. In this study, we focused on the spatial distribution of macroinvertebrate communities in river-lake systems. The sample points were selected to cover the upstream, BYDL, and downstream regions, thereby reflecting the observed differences in the overall characteristics of the macroinvertebrate communities within the basin. A total of 152 samples were collected, 83 in 2019 and 69 in 2020, of which 69 were overlapping sample sites and 14 were not collected due to unforeseen circumstances.
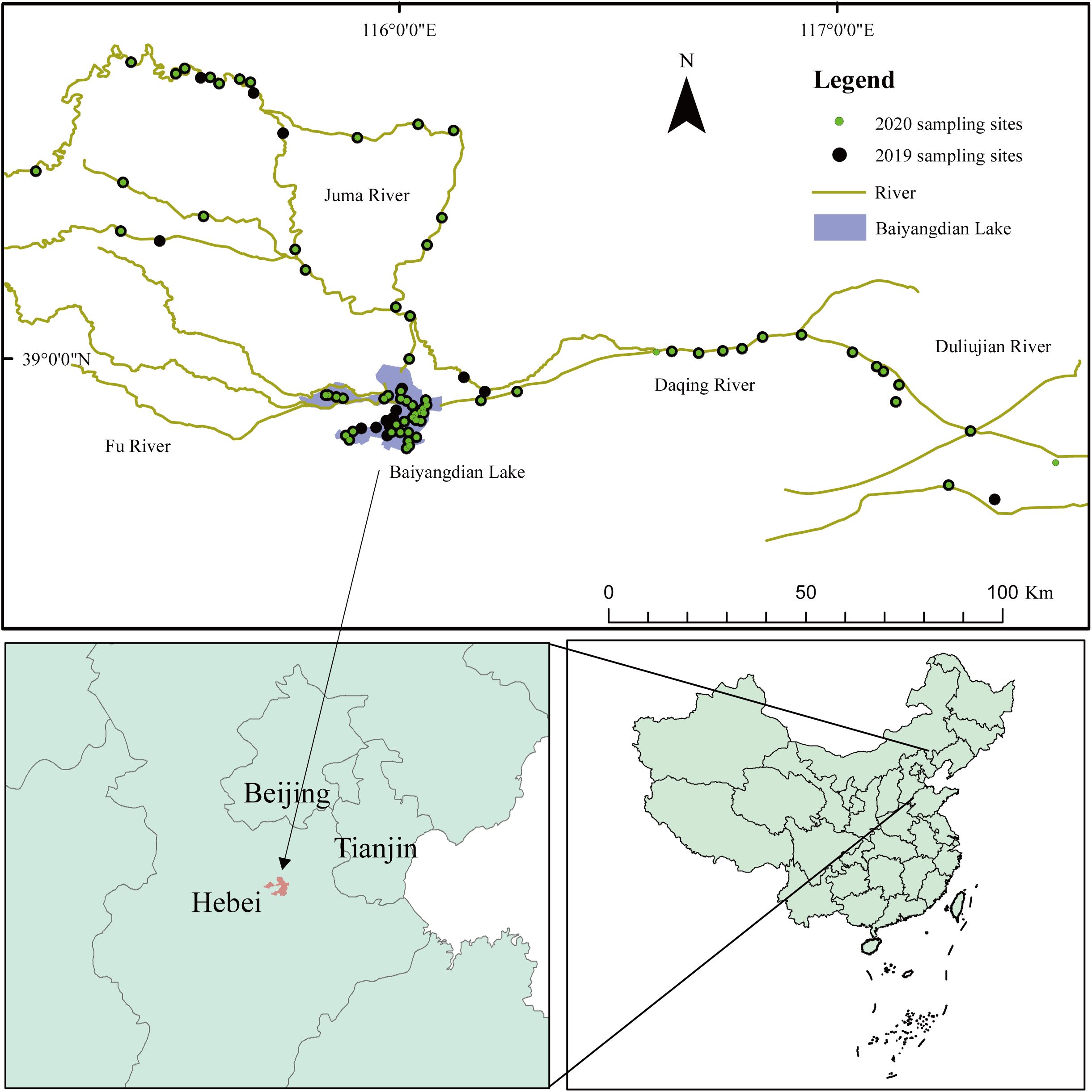
Figure 1. The 152 studied sties in Daqinghe River system in subtropical and tropical China. The surface water and macroinvertebrate samples were collected from September in 2019 and 2020.
2.2 Benthic macroinvertebrates
A combination of type D trawls and modified Peterson mud harvesters were used to collect macroinvertebrate samples, and three replicate samples were collected at each sampling site (Zhang et al., 2021). The three replicate samples were mixed into one sample. After being sieved through a 60-mesh screen (250 μm) and removing any impurities, the samples were preserved with 75% alcohol and fixed with formaldehyde. A small amount of soda or borax was added to neutralize the formaldehyde to avoid corrosion of the calcareous shells, and insects and crustaceans were fixed with 75% alcohol in the sample bottles. The macroinvertebrates were identified to the lowest possible taxonomic level using appropriate identification guides under a microscope (Morse et al., 1994; Costas et al., 2018).
2.3 Environmental variables
Physicochemical parameters, including water temperature (WT; ℃), dissolved oxygen (DO; mg/L), electrical conductivity (EC;μs/m), pH, and turbidity were measured using a YSI multiparameter probe (Yellow Spring, Ohio, USA). Water samples were collected from each sampling point using 500 mL plastic bottles and transported to the laboratory for water quality analysis. The ammonium nitrogen (NH4-N), total phosphorus (TP), and total nitrogen (TN) contents were determined by UV spectrophotometry. The chemical oxygen demand (COD) was determined by potassium dichromate colorimetry. The measurements were conducted according to the relevant standards for water chemistry measurement in China (Huang et al., 1999). The environmental factors for the 152 samples under investigation are presented in Supplementary Table S1. The water samples were collected before the macroinvertebrate samples to avoid any disturbance caused by the collection of the latter.
2.4 Spatial factors
Spatial factors were employed as proxies for dispersal processes (Borcard et al., 2004). To create spatial factors, a Moran feature vector map (MEM) based on the Euclidean distances between sites was created according to geographic coordinates (x and y) (Blanchet et al., 2008). The principal coordinates of neighbor matrices (PCNM) is a method for the decomposition of spatial relationships among sampling sites, based on the diagonalization of a spatial weighted matrix (Bellier et al., 2007). For PCNM, the Euclidean distance matrix between the sampling points was calculated, and then the scale of the distance matrix was reduced. During this process, a Euclidean distance threshold needs to be set. Distances smaller than the threshold were retained, while distances larger than this threshold were set to four times the threshold. Quantitative ecologists have improved the traditional PCNM method, which can model any type of spatial structure. Thus, any similarity matrix can replace the Euclidean distance matrix to analyze the spatial structure, which is called distance-based Moran’s eigenvector maps (dbMEM). MEMs (i.e., spatial eigenvalues) can be used to model the spatial structure of biological assemblages at multiple scales, and they can represent dispersal processes, biogeographic effects, and environmental autocorrelation (Dray et al., 2012). In this study, the MEM vectors that were associated with positive eigenvalues and a significant Moran’s I were retained as they represented a positive spatial autocorrelation. The MEM was constructed using the “PCNM” package in R (Legendre et al., 2015).
2.5 Data analysis
All of the analyses were performed in R (version 4.3.2) unless otherwise indicated (R Core Team, 2022). The α-diversity index for each sample (i.e., the Shannon-Winner index) and β-diversity index (i.e., the “Bray-Curtis” distance) between the pairwise sampling sites were calculated using the “vegan” package. A principle coordinates analysis (PCoA) based on the “Bray-Curtis” distance was performed using the “vegan” package (version 2.6-4) to show the profile of the macroinvertebrate communities. Non-parametric statistical methods based on Adonis (Anderson, 2017) were employed to analyze the differences in macroinvertebrate community composition within rivers and lakes using the “vegan” package (Oksanen et al., 2019).
To address whether the DDR patterns of macroinvertebrate communities are variable with scale, the relationship between macroinvertebrate community similarity and geographic distance at the continuum river-lake system was investigated in 2019 and 2020. The study area was divided into two waterbody types (river and lake) and the distance decay relationship was calculated for each sampling point within the two spaces. The relationship between macroinvertebrate community similarity and geographic distance was calculated by linear least squares regression using the “vegan” package (Martiny et al., 2011).
To test the overall correlation between the two matrices of macroinvertebrate community and environmental factors and identify the potential driving factors of the formation of ecological patterns at the macro level, a mantel test was applied. However, a mantel test is highly dependent on the randomness and independence of the data. The importance of the effect of spatial and environmental variables on macroinvertebrate community variation was assessed separately based on redundancy analyses (RDA) and variance partitioning analysis. Variance Partitioning Analysis can determine the proportion of changes in community structure due to a given environmental factor. The contribution of each environmental factor and the interaction among the different environmental factors to the change in biomes can be calculated using sequencing analysis. However, the traditional varpart function in the “vegan” package can only handle four groups of explanatory variables. To improve normality and mean square deviation before the statistical analyses, the macroinvertebrate data were Hellinger transformed, and the environmental parameters were log (x+1) transformed except for the pH. For the RDA, spatial factors were calculated from the geographic distances using a PCoA of dbMEMs (Peres-Neto et al., 2006). The geographical factor (dbMEM) was determined using the “pcnm” function in the “vegan” package. To reduce multicollinearity between the spatial and environmental factors, variables with variance inflation factors less than 10 were retained in the analysis. The Euclidean ambient distance for each sample point in the two spaces was calculated to determine whether the environmental variables of the two water types were different.
To analyze the relative importance of the effect of the geographical distance and environmental factors on macroinvertebrate communities, a multiple regression on matrices (MRM) approach was employed. A MRM can consider multiple candidate models at the same time, evaluate each model, and select the model combination that can best explain the dynamic change in the vegetation community. However, the process of model selection is subjective. In this study, MRM was used for the different waterbody types (i.e., river and lake). The macroinvertebrate community dissimilarity matrix was calculated based on the abundance‐weighted UniFrac distance. A modified MRM code based on the R package “ecodist” (Goslee, 2007) was used to elucidate potential relationships between macroinvertebrate β-diversity and factors in the river and lake. The R2 values of the MRM model represent the total explanatory power of all the factors involved in the model, while the partial regression coefficient (i.e., b) represents the relative contribution of each factor. To remove covariant factors, non-significant factors were removed (Harrell, 2001). The MRM model was then performed again following the methods detailed in (Martiny et al., 2011).
To assess the importance of the effect of different processes on macroinvertebrate communities, the direct and indirect effects of spatial processes, physicochemical processes, and nutrients on macroinvertebrate communities were modeled using the “plspm” package (Liu et al., 2019). The corresponding influencing processes were represented by factors at different scales. The initial model included all possible pathways between the macroinvertebrate community composition, spatial geographic factors (MEM), physicochemical factors (WT, EC, DO, Turbidity, pH, and COD), and nutrients (TN, TP, and NH4-N). A PCoA was used to summarize the variation in the distribution of the macroinvertebrate communities. The variables with loadings < 0.6 were removed and the goodness of fit index and R2 were used to estimate the performance of the model. The “lavaan” package was used to perform partial least squares path model (PLS-PM) analysis (Mamet et al., 2019).
To ascertain the potential influence of stochastic processes on community assembly, the NCM (neutral community model) was used to predict the relationship between macroinvertebrate detection frequency and their relative abundance across the wider metacommunity (Sloan et al., 2006). The model presented here is an adaptation of the neutral theory adjusted to macroinvertebrate populations (Hubbell, 2001). In this model, Nm is an estimate of dispersal between communities, and it can be used to determine the correlation between occurrence frequency and regional relative abundance, with N representing the metacommunity size and m indicating the immigration rate. The parameter R2 represents the overall fit to the neutral model (Sloan et al., 2006). All the fitting statistics were calculated with 1000 bootstrap replicates, and 95% confidence intervals were generated. The new mathematical framework of quantitative description of ecological stochastic processes that was proposed by Ning et al. (2019) was used, and the NST (normalized stochasticity ratio)() was employed to measure the importance of deterministic (homogenous and heterogenous) and stochastic (dispersal limitation and drifts) assembly processes. The assembly was considered to be dominated by deterministic processes (with a ratio of < 50%) or stochastic processes with a ratio of > 50% (Ning et al., 2019).
3 Results
3.1 Composition, structure and diversity of the macroinvertebrate communities in river and lake equations
In both years, the river and lake macroinvertebrate communities comprised 72 taxa distributed across three phyla, six classes, and 15 orders (Supplementary Table S1). Arthropoda was mainly distributed in the river, while Mollusca was widely distributed in the lake (Supplementary Figure S1). Specifically, the relative abundance of Annelida and Arthropoda was lower in the lake than in the river, whereas Mollusca was higher (Supplementary Figure S1). The macroinvertebrate α-diversity significantly differed between the river and lake in 2019. Specifically, compared with the lake, the observed Shannon and Simpson indexes were significantly lower in the river (Supplementary Figure S1). Neither the Simpson Index nor the Shannon Index of the lake and river were significantly different in 2020. No significant difference in richness was observed inter-annually (Supplementary Figure S2).
The PCoA results demonstrated a degree of dissimilarity between the samples, with the river and lake showing significant spatial variability, which was further supported by the non-parametric statistical approaches (R2 = 0.09, P = 0.001, 2019; R2 = 0.09, P = 0.001, 2020; Supplementary Figure S3). The PCoA analysis showed clear separation of communities by sampling location, with all the samples essentially grouped into two geographic clusters (i.e., river and lake; Supplementary Figure S3).
3.2 Spatial distribution of the macroinvertebrate communities in river and lake
The community similarity of the lake was significantly higher than that of the river in 2019 and 2020 (Figure 2). Significant DDRs were identified in both the river and lake macroinvertebrate communities, with the similarity of the macroinvertebrate communities significantly decreasing with increasing geographic distance based on Bray-Curtis similarity (P < 0.01, Figure 2). Moreover, between the two waterbody types, there were significant differences in the slopes of the DDRs. In the lake, the DDR slope was steeper, while the slope was flatter and the correlation was weaker in the river (Figure 2). In addition, the DDR patterns of the macroinvertebrate community differed in the river between 2019 and 2020 (Figure 2). Furthermore, significant DDRs were observed both upstream and downstream in the river (Supplementary Figure S4).
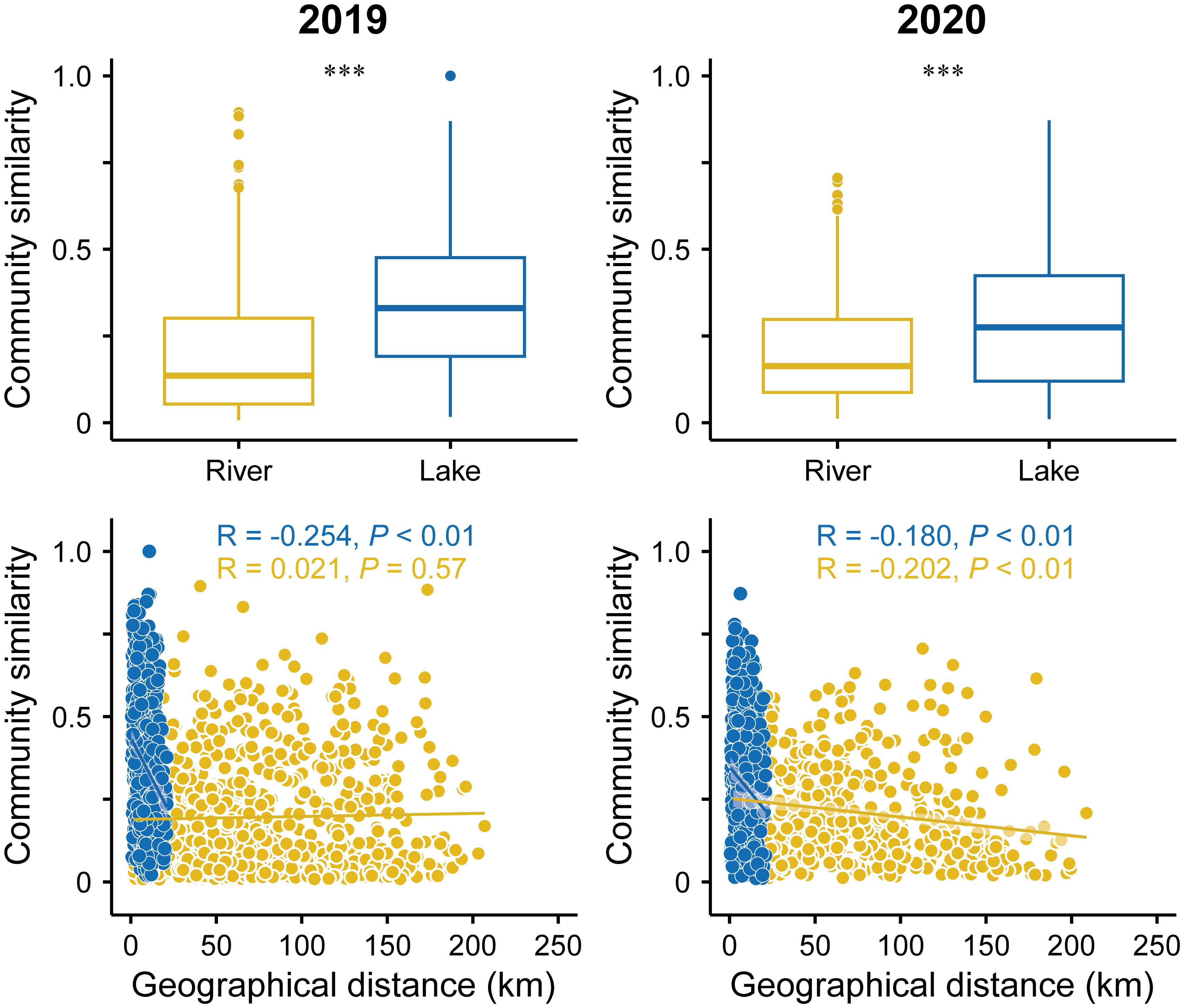
Figure 2. Distance decay relationships based on the Bray-Curtis similarity of macroinvertebrates community and geographical distance at two scales (within rivers, and within lake). Significant differences (***P < 0.01; Wilcoxon rank-sum test) between the two scales are indicated by asterisks. Boxes and error bars indicate the 25th/75th and 5th/95th percentiles, respectively; middle line of the boxes is the median. The shaded area around the lines covers 95 % confidence.
Environmental distance decay was also significantly different in the different waterbody types in 2019 and 2020. In 2019, there was a significant environmental distance decay pattern for the macroinvertebrate communities in the lake, but the pattern was weaker in the river (Supplementary Figure S5).
3.3 Spatial and environmental variables driving the differences in the macroinvertebrate communities
The Mantel test was employed to identify which environmental variables exhibited a significant correlation with the macroinvertebrate community. In 2019, pH showed a relatively weak relationship with the macroinvertebrate communities in the river (r <0.2, P < 0.01). Longitude (r > 0.2, P < 0.01) and TP (r > 0.2, P < 0.05) showed significant correlations with the macroinvertebrate composition in the lake. In 2020, WT had significant correlations with the macroinvertebrate community composition (r > 0.2, P < 0.05) in both the river and lake. Additionally, pH had strong and significant correlations with the macroinvertebrate community composition in the lake (r > 0.2, P < 0.05; Figure 3).
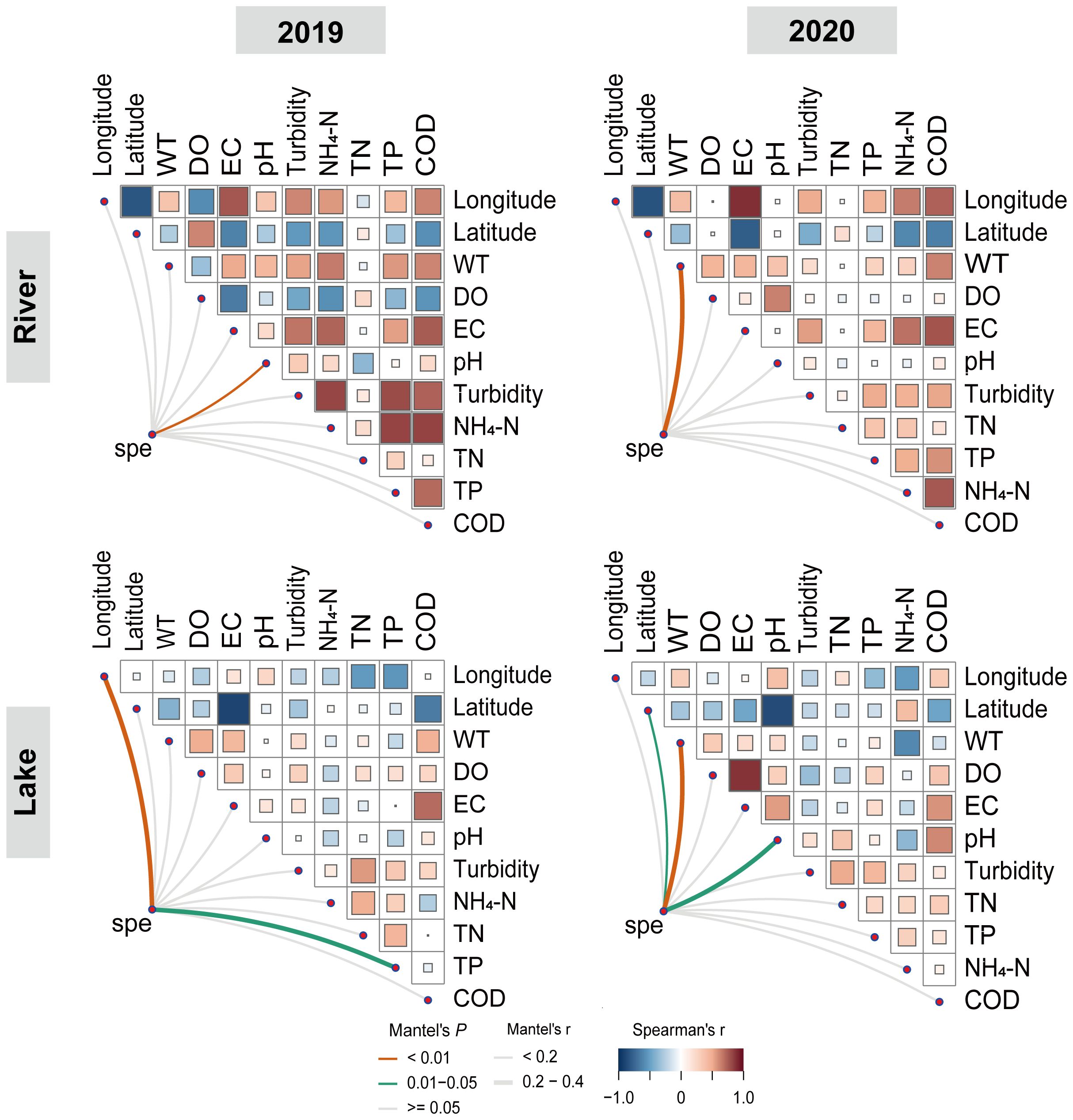
Figure 3. The paired comparison of geographic and environmental factors is shown in the bottom, and the color gradient represents Spearman’s correlation coefficient. Through partial Mantel tests, the composition of the macroinvertebrates communities is related to each geographic orenvironmental factor. The line width represents partial the Mantel’s r statistics of the corresponding correlation, and the line color represents the significance based on 999 permutations.
According to the VPA, the macroinvertebrate communities were influenced by both spatial and environmental factors. Macroinvertebrate communities in rivers were slightly more affected by spatial factors than environmental factors. Whereas, in 2020, macroinvertebrate communities in the lake were more affected by environmental factors than spatial factors, and, in 2019, they were more affected by spatial factors than environmental factors (Figure 4).
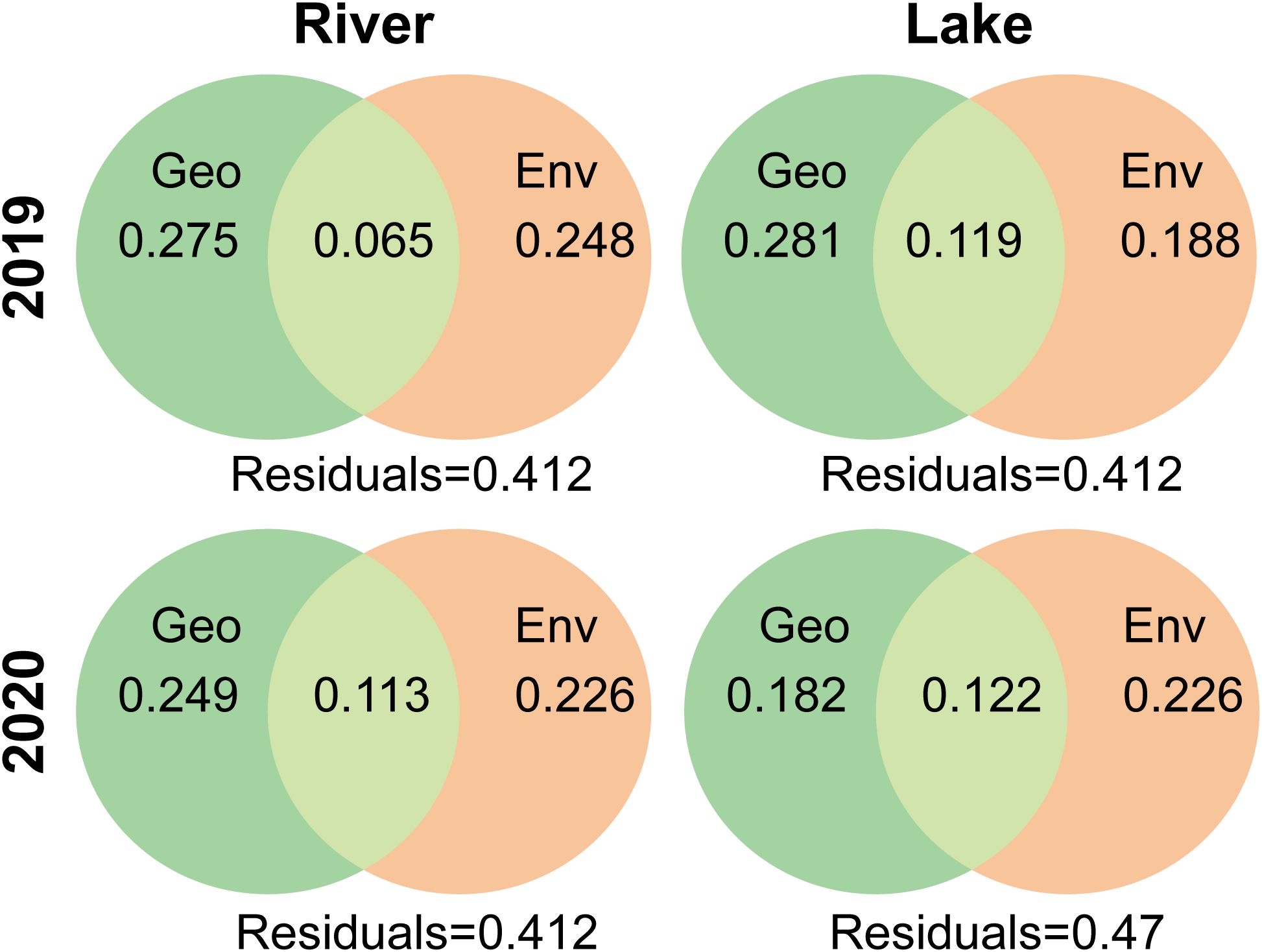
Figure 4. Venn diagrams illustrating the results of variation partitioning for macroinvertebrate communities in 2019, macroinvertebrate communities in 2020 by Env (environmental variables) and Geo (geographical factors) in the two spatial levels: river and lake. Residuals (Res) are shown in the lower corner. All fractions are based on adjusted R2 values shown as percentages of total variation.
In the MRM analyses, pH was the most important abiotic factor in explaining community variation in the river (b = -0.254, P < 0.05) in 2019. In the lake, the dominant factor driving changes in macroinvertebrate communities was geographical distance (b = -0.163, P < 0.05; Supplementary Table S1). In 2020, geographical distance and WT were the most important environmental factors in explaining community variation in the river (b = -0.314, P < 0.05; b = -0.522, P < 0.05). In the lake, the most important driver of changes in macroinvertebrate communities was WT (b = -0.488, P < 0.001; Supplementary Table S3). The varying importance of geographic distance and environmental factors reflected the different underlying variability across spatial-temporal scales in macroinvertebrate communities (Supplementary Table S3).
We used PLS-PM analysis to explore the potential direct and indirect impacts of geographical factors, physicochemical factors, and nutrients on macroinvertebrate community composition (Supplementary Figure S6). The variation in the PCoA was explained by geographical factors, physicochemical factors, and nutrient variables. Spatial variables directly influenced macroinvertebrate community distribution, and they indirectly exerted influence via environmental factors. In 2019, the geographical variables, nutrients, and physicochemicals all had positive effects on the PCoA in the river, while they had negative effects in the lake. In 2020, the physicochemicals in the river directly affected the PCoA, while nutrients directly affected the PCoA in the lake.
3.4 Community assembly mechanisms of macroinvertebrate communities
The NCM accurately predicted the relationship between the occurrence frequency of the macroinvertebrates and variations in their relative abundance. The NCM explained 36.4%; 2019, river). 81%; 2019, lake). 69.6%;2020, river), and 83.1%;2020, lake) of community variance (Figure 5). The NST results demonstrated that stochastic processes played a large role in shaping the macroinvertebrate community composition in the lake, but deterministic process dominated in the river (Figure 6).
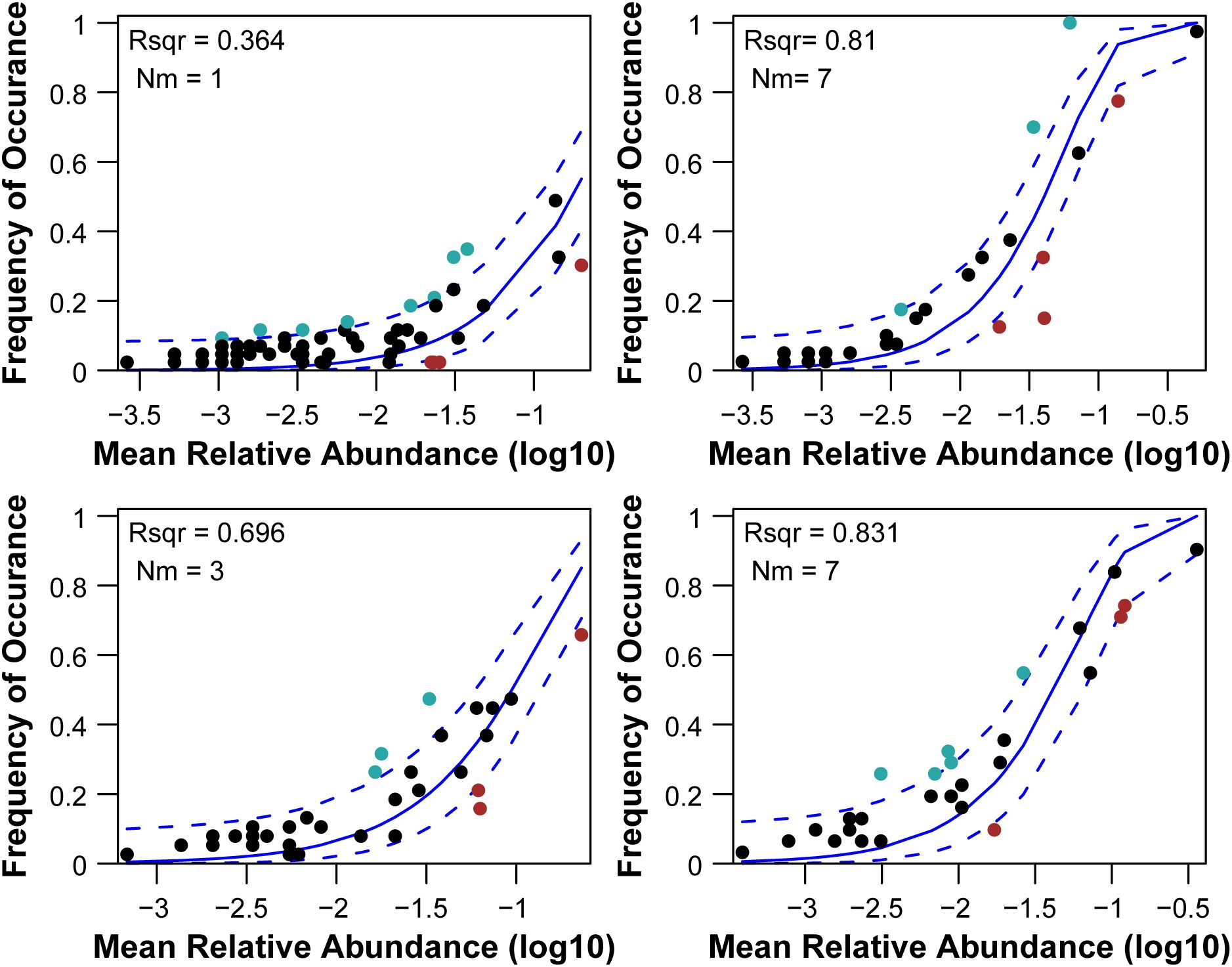
Figure 5. Fit of the neutral community model (NCM), displaying the predicted occurrence frequencies against the relative abundance of aquatic macroinvertebrates in river and lake. The blue solid lines depict the optimal fit to Sloan’s neutral model, and the dashed lines delineate the 95 % confidence intervals around the model prediction. R2: goodness of fit to the neutral model.

Figure 6. Normalized stochasticity ratio (NST). The value of 0.5 is regarded as the boundary between a stochastic (> 0.5) and deterministic (< 0.5) assembly.
4 Discussion
4.1 Water type distance decay patterns in macroinvertebrate communities
The distance decay of similarity concept is inherent in many previous studies (Preston, 1962) and has been reported for various taxa and geographical settings (Whittaker, 1960; Soininen et al., 2007). In this study the waterbody type dependence of DDR patterns was investigated in a river-lake system. The results supported the hypothesis that the river zone decays at a slower rate when each spatial region of the Daqing River system was examined separately. The decay rates were more gradual in the river zone than in the lake. Furthermore, the results supported the other hypotheses to some extent, as the similarity of the macroinvertebrates in the lake decreased with distance. However, the similarity of the riverine macroinvertebrates did not differ significantly with increasing distance. The disparities among the different spatial ecoregions highlight the diverse factors that affect organisms at varying spatial levels (Maloney and Munguia, 2011). Some studies have shown that significant DDRs occur in macroinvertebrate communities (Astorga et al., 2012; Maloney and Munguia, 2011; Soininen et al., 2007). The relationship between macroinvertebrate community similarity and geographical distance may be stronger in lakes (smaller spatial scales). Our results are also consistent with the importance of geographic distance being limited to relatively small spatial scales and taxa with poor dispersal abilities (Astorga et al., 2012).
Our most noteworthy discovery was that the slope of the DDR became less steep as the spatial scale for the macroinvertebrate communities increased, which supports the hypothesis of Maloney and Munguia (2011). This suggested that macroinvertebrate community similarity decreased rapidly with the increase in geographic distance (particularly in the lake). This may be due to several ecological mechanisms. Lakes tend to have more homogeneous environmental conditions and lower habitat complexity than rivers, which allows for stronger effects of dispersal limitation and environmental filtering on community structure. In addition, the smaller the spatial scale of lakes, the greater the influence of local environmental factors, and the faster community similarity declines with distance. In addition, connectivity between lake habitats may be more homogeneous, leading to more consistent dispersal processes across the lake. These factors combine to influence distance decay patterns in lake macroinvertebrate communities. We also found that the slope of the DDRs was significantly more gradual in the river than in the lake. As macroinvertebrates are dispersed throughout the entire basin by water currents, the selective pressure exerted by the environment can significantly alter the community composition. Usually, the greater the disparity in the environmental conditions between localities, the more pronounced the variation in the macroinvertebrate communities (Heino et al., 2015b; Kärnä et al., 2015). This finding is consistent with other studies in phytoplankton, diatoms, and zooplankton (Bellier et al., 2014; Mazaris et al., 2010; Marquardt et al., 2018).
There may be several reasons for the waterbody type’s dependence on macroinvertebrates DDRs. First, the environmental heterogeneity was significantly larger at the river scale than at the lake scale (Supplementary Figure S7). Second, the initial community similarity of adjacent sampling stations in the lake was high, which usually results in strong distance decay (Soininen et al., 2011). Third, the dispersal rate strongly regulates the compositional response of macroinvertebrate communities (Li et al., 2022; Zhao et al., 2024). The downstream areas were more impacted by human activities and exhibited greater homogenization compared with the headwaters. This led to a decrease in both taxonomic and functional diversity within the macroinvertebrate communities. Hence, human activities played a significant role in community shift, indirectly contributing to biogenic homogenization through alterations in physical and chemical properties, and subsequently influencing ecological processes (Ma et al., 2023).
4.2 Spatial and environmental variables shaped the macroinvertebrate communities
Variance partitioning revealed substantial combined effects of the spatial and environmental variables. The Daqing River Basin spanned approximately 300 km. Therefore, the effects of spatial and environmental variables on the macroinvertebrate assemblages in the river were not surprising. The findings emphasize the significant impact of spatial variables on the variations in macroinvertebrate communities throughout the river (Figure 4). In the river, macroinvertebrate communities with a greater geographical distance between them exhibited increasingly larger differences, which confirms the influence of spatial variables (Figure 2). However, being surrounded or divided by farmlands, ditches, or paved roads related to agricultural activities may cause severe dispersal limitations for macroinvertebrates in the river (Guan et al., 2024).
A high degree of environmental heterogeneity was found within the lake and in the river, and both spatial and environmental factors influenced the community composition. The results of the VPA and MRM demonstrated that spatial factors exerted a slightly more pronounced influence on the structure of the macroinvertebrate communities than environmental factors. This is consistent with the findings observed in the Hun-Tai watershed (Zhou et al., 2020). The results of the VPA analysis in this study indicated that spatial factors played a significant role in shaping the macroinvertebrate communities in the river in both 2019 and 2020. In contrast, the lake region exhibited disparate outcomes between the two years, with environmental factors serving as the primary driver of community differences in 2020 (Spasojevic and Suding, 2012). This may be attributed to the high environmental heterogeneity observed in the lake in 2020. Geographical factors, physicochemical factors, and nutrients affected the macroinvertebrate community spatial distribution as found in other aquatic communities (Dong et al., 2024; Zhao et al., 2024).
The geographical factors (PCNMs) indirectly affected the distribution of the macroinvertebrate communities by altering the environmental factors. The macroinvertebrate communities in the river were not nutrition-restricted, while the lake had lower nutrient levels than the river (Supplementary Figure S7). This may be due to the dilution effect of the lake on nutrients. Nutrient restriction affects primary productivity and, therefore, macroinvertebrate communities. To illustrate the effects of the spatial and environmental factors on macroinvertebrate communities’ spatial distribution, field observation should be conducted more frequently over a longer duration.
4.3 Assembly process of macroinvertebrate communities
The NCM model exhibited a good fit for the majority of species, thereby substantiating the significant role of stochastic processes (i.e., dispersal and ecological drift) in the assembly of macroinvertebrate communities. The distance decay pattern of benthic macroinvertebrate communities in different water body types further confirmed the significance of stochastic processes (Figure 2; Supplementary Figure S4). According to Hubbell’s Neutral Theory (Hubbell, 2001), due to dispersal limitation, community similarity is expected to decline with distance (Chase and Myers, 2011). Several studies have supported these findings. For instance, (Li et al., 2022) demonstrated that the benthic community structure in South Chinese bays was primarily driven by stochastic processes (R2 = 0.90; NCM).
In this study, the R2 value of the NCM parameter was slightly higher in the lake than in the river, and based on the Nm values, the macroinvertebrate dispersal between the sampling sites may have been higher in the lake than in the river (assuming a similar metacommunity size, Figure 5). This suggests that the effect of stochastic processes on the macroinvertebrates was stronger in the lake (Swenson and Enquist, 2009). Widespread dispersal is more likely to occur in highly connected water bodies (Tolonen et al., 2017), such as lakes (Cai et al., 2019) and main rivers (Brown and Swan, 2010). These results may be attributed to the higher habitat homogeneity in the lake compared with the river. High diffusion rates can reduce the effects of environmental selection and ecological drift to some extent. The wide distribution of the macroinvertebrates had a relatively small spatial impact on the community, suggesting that dispersal limitations had a weak effect on the spatial turnover of the macroinvertebrate communities in the lake.
Nevertheless, the NCM did not explain all the variation in the macroinvertebrate community, indicating that there might be other community assembly mechanisms, resulting in a non-neutral distribution. These other mechanisms include environmental selection and species interactions (Heino et al., 2015a). Stochastic processes establish the relationship between species occurrence and mean relative abundance (conforming to the neutral distribution), while environmental selection may modify this relationship. Species that are more adapted to local conditions have a higher frequency of occurrence irrespective of their relative abundance. This results in a non-neutral distribution of species occurrence. Another potential explanation is the varying migration or dispersal capabilities among species (Li et al., 2022). A higher occurrence frequency might indicate a higher dispersal ability than expected, and a lower occurrence frequency might indicate a lower dispersal ability than anticipated.
The relative importance of stochasticity in macroinvertebrate community building was further quantified by the NST based on a neutral model. The NST allows for more precise categorization of the relative contributions of stochastic and deterministic processes in benthic community construction and provides a more comprehensive perspective for understanding community construction mechanisms. The results suggested that deterministic processes may have a high influence in river macroinvertebrate community construction (Zhao et al., 2024), whereas construction may be more random in lake macroinvertebrate communities (Guan et al., 2024).
In summary, both ecological niche-related processes (environmental conditions and species interactions) and neutral processes (dispersal) were important for benthic community formation in this study, and both should be considered when exploring community assembly processes.
4.4 Future research
In this study, the diversity trends reflective of the river-lake system in temperate China. Additionally, the analysis was based on a two-year data set and does not include the full range of conditions that can occur in rivers and lakes, including wet periods, dry periods, and stable periods. Therefore, long-term data should be collected.
This study compared different regions and different years. Future research could use remote sensing techniques, such as satellite or unmanned aerial vehicle imagery. This information could provide large-scale, consistent, and repeatable measurements of land cover and water attributes, allowing for the validation of these research findings and determining the extent to which the observed patterns are generalizable. Remote sensing could be used to incorporate high-resolution hydrological conditions, such as wet and dry conditions, and it has reproducible frameworks (Valerio et al., 2024; Yue et al., 2023), which could enhance the understanding of the impacts of seasonal flow patterns on community composition. It could also be helpful for studying the dispersion of macroinvertebrates (Dong et al., 2021), which is strongly influenced by habitat landscape connectivity.
5 Conclusion
Our multi-scale data provided empirical support for macroinvertebrate communities being affected by the waterbody type in a river-lake system. This study demonstrated that the DDR patterns in macroinvertebrate communities varied in river-lake systems. The slopes of the DDRs were found to be steeper within the lake, and flatter in the river. The findings provide compelling evidence that both spatial and environmental variables contributed to the macroinvertebrate DDRs and affected the distribution of the macroinvertebrate communities in the river-lake system. In the river, spatial variables had a larger effect. Spatial variables directly influenced macroinvertebrate community distribution and indirectly influenced environmental variables. Stochastic processes were dominant and maintained community aggregation in the lake. Deterministic processes were dominant that and maintained macroinvertebrate community in the river. In order to preserve these communities and maintain their ecological functions, relevant conservation strategies should be considered. Firstly, protecting and restoring the natural connectivity of water bodies between lakes and rivers is essential to promote dispersal and gene flow between populations. Second, reducing anthropogenic disturbances, such as pollution and habitat destruction, can help maintain the environmental heterogeneity of diverse macroinvertebrate communities. Thirdly, the implementation of monitoring programs can facilitate the collection of data to assess the effectiveness of conservation efforts and adjust management strategies over time. By integrating these we can conserve biodiversity and ecological integrity in river-lake systems.
In summary, this study contributed to our comprehension of the spatial distribution of macroinvertebrate communities and revealed the underlying mechanisms by which macroinvertebrate communities assemble across a range of spatial extents.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
YS: Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. JW: Formal analysis, Investigation, Writing – review & editing. HXL: Formal analysis, Writing – review & editing. HL: Data curation, Methodology, Writing – review & editing. PH: Conceptualization, Methodology, Writing – review & editing. LJ: Funding acquisition, Investigation, Writing – original draft, Writing – review & editing. CL: Funding acquisition, Methodology, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by the National Natural Science Foundation of China (32401332), the Biodiversity Survey and Assessment Project of the Ministry of Ecology and Environment, China (2019HJ2096001006), the Hebei Province Wetland Ecology and Protection key laboratory open fund, China (hklk202003).
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1538755/full#supplementary-material
References
Anderson M. J. (2017). “Permutational multivariate analysis of variance (PERMANOVA)” in Wiley StatsRef: Statistics Reference Online., 1–15. doi: 10.1002/9781118445112.stat07841
Anderson M. J., Crist T. O., Chase J. M., Vellend M., Inouye B. D., Freestone A. L., et al. (2011). Navigating the multiple meanings of beta diversity: a road map for the practicing ecologist. Ecol. Letters. 14, 19–28. doi: 10.1111/j.1461-0248.2010.01552.x
Astorga A., Oksanen J., Luoto M., Soininen J., Virtanen R., Muotka T. (2012). Distance decay of similarity in freshwater communities: do macro- and microorganisms follow the same rules? Global Ecol. Biogeography 21, 365–375. doi: 10.1111/j.1466-8238.2011.00681.x
Bellier E., Grøtan V., Engen S., Schartau A. K., Herfindal I., Finstad A. G. (2014). Distance decay of similarity, effects of environmental noise and ecological heterogeneity among species in the spatio-temporal dynamics of a dispersal-limited community. Ecography 37, 172–182. doi: 10.1111/j.1600-0587.2013.00175.x
Bellier E., Monestiez P., Durbec J.-P., Candau Jean-Noël (2007). Identifying spatial relationships at multiple scales: principal coordinates of neighbor matrices (PCNM) and geostatistical approaches. Ecography 30, 385–399. doi: 10.1111/j.2007.0906-7590.04911.x
Blanchet F. G., Legendre P., Borcard D. (2008). Modeling directional spatial processes in ecological data. Ecol. Model. 215, 325–336. doi: 10.1016/j.ecolmodel.2008.04.001
Borcard D., Legendre P., Avois-Jacquet C., Tuomisto H. (2004). Dissecting the spatial structure of ecological data at multiple scales. Ecology 85, 1826–1832. doi: 10.1890/03-3111
Brown B. L., Swan C. M. (2010). Dendritic network structure constrains metacommunity properties in riverine ecosystems. J. Anim. Ecol. 79, 571–580. doi: 10.1111/j.1365-2656.2010.01668.x
Buffagni A. (2021). The lentic and lotic characteristics of habitats determine the distribution of benthic macroinvertebrates in Mediterranean rivers. Freshw. Biol. 66, 13–34. doi: 10.1111/fwb.13596
Cai Y., Zhang Y., Hu Z., Deng J., Qin B., Yin H., et al. (2019). Metacommunity ecology meets bioassessment: Assessing spatio-temporal variation in multiple facets of macroinvertebrate diversity in human-influenced large lakes. Ecol. Indic. 103, 713–721. doi: 10.1016/j.ecolind.2019.03.016
Carvalho J. C., Cardoso P., Gomes P. (2012). Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Global Ecol. Biogeography 21, 760–771. doi: 10.1111/j.1466-8238.2011.00694.x
Chaparro G., Horváth Z., O’Farrell I., Ptacnik R., Hein T. (2018). Plankton metacommunities in floodplain wetlands under contrasting hydrological conditions. Freshw. Biol. 63, 380–391. doi: 10.1111/fwb.13076
Chase J. M., Myers J. A. (2011). Disentangling the importance of ecological niches from stochastic processes across scales. Philos. Trans. R. Soc. B-Biological 366, 2351–2363. doi: 10.1098/rstb.2011.0063
Costas N., Pardo I., Méndez-Fernández L., Martínez-Madrid M., Rodríguez P. (2018). Sensitivity of macroinvertebrate indicator taxa to metal gradients in mining areas in Northern Spain. Ecol. Indic. 93, 207–218. doi: 10.1016/j.ecolind.2018.04.059
Cottenie K. (2005). Integrating environmental and spatial processes in ecological community dynamics. Ecol. Lett. 8, 1175–1182. doi: 10.1111/j.1461-0248.2005.00820.x
Dong R., Peng K., Zhang Q., Heino J., Cai Y., Gong Z. (2024). Spatial and temporal variation in lake macroinvertebrate communities is decreased by eutrophication. Environ. Res. 243, 117872. doi: 10.1016/j.envres.2023.117872
Dong R., Wang Y., Lu C., Lei G., Wen L. (2021). The seasonality of macroinvertebrate β diversity along the gradient of hydrological connectivity in a dynamic river-floodplain system. Ecol. Indic. 121, 107112. doi: 10.1016/j.ecolind.2020.107112
Dray S., Pélissier R., Couteron P., Fortin M.-J., Legendre P., Peres-Neto P. R., et al. (2012). Community ecology in the age of multivariate multiscale spatial analysis. Ecol. Monogr. 82, 257–275. doi: 10.1890/11-1183.1
Goslee G. C. (2007). The ecodist package for dissimilarity based analysis of ecological data. J. Stat. Software 22, 1–15. doi: 10.18637/jss.v022.i07
Göthe E., Baattrup-Pedersen A., Wiberg-Larsen P., Graeber D., Kristensen E. A., Friberg N. (2017). Environmental and spatial controls of taxonomic versus trait composition of stream biota. Freshw. Biol. 62, 397–413. doi: 10.1111/fwb.12875
Guan Q., He F., Li Z., Cai Y., Kang Y., Zhang Z., et al. (2024). Contrasting diversity patterns and drivers of aquatic macroinvertebrates in floodplain and non-floodplain wetlands. Sci. Total Environ. 945, 174045. doi: 10.1016/j.scitotenv.2024.174045
Harrell F. E. Jr. (2001). Regression modeling strategies, with applications to linear models, survival analysis and logistic regression. (Springer). Available online at: https://link.springer.com/book/10.1007/9781475734621?page=1.
Heino J., Alahuhta J., Bini L. M., Cai Y., Heiskanen A., Hellsten S., et al. (2021). Lakes in the era of global change: moving beyond single-lake thinking in maintaining biodiversity and ecosystem services. Biol. Rev. 96, 89–106. doi: 10.1111/brv.12647
Heino J., Grönroos M., Soininen J., Virtanen R., Muotka T. (2012). Context dependency and metacommunity structuring in boreal headwater streams. Oikos 121, 537–544. doi: 10.1111/j.1600-0706.2011.19715.x
Heino J., Melo A. S., Bini L. M. (2015b). Reconceptualising the beta diversity-environmental heterogeneity relationship in running water systems. Freshw. Biol. 60, 223–235. doi: 10.1111/fwb.12502
Heino J., Melo A. S., Siqueira T., Soininen J., Valanko S., Bini L. M. (2015a). Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshw. Biol. 60, 845–869. doi: 10.1111/fwb.12533
Heino J., Peckarsky B. L. (2014). Integrating behavioral, population and large-scale approaches for understanding stream insect communities. Curr. Opin. Insect Sci. 2, 7–13. doi: 10.1016/j.cois.2014.06.002
Huang X., Chen W., Cai Q. (1999). “Standard methods for observation and analysis in chinese ecosystem research network e survey,” in Observation and Analysis of Lake Ecology (Standards Press of China, Beijing).
Hubbell S. P. (2001). A unified neutral theory of biodiversity and biogeography (Princeton: Princeton University Press).
Jamoneau A., Passy S. I., Soininen J., Leboucher T., Tison-Rosebery J. (2018). Beta diversity of diatom species and ecological guilds: Response to environmental and spatial mechanisms along the stream watercourse. Freshw. Biol. 63, 62–73. doi: 10.1111/fwb.12980
Jin L., Chen H.-H., Xue Y., Soininen J., Yang J. (2022). The scale-dependence of spatial distribution of reservoir plankton communities in subtropical and tropical China. Sci. Total Environ. 845, 157179–157179. doi: 10.1016/j.scitotenv.2022.157179
Kärnä O.-M., Grönroos M., Antikainen H., Hjort J., Ilmonen J., Paasivirta L., et al. (2015). Inferring the effects of potential dispersal routes on the metacommunity structure of stream insects: as the crow flies, as the fish swims or as the fox runs? J. Anim. Ecol. 84, 1342–1353. doi: 10.1111/1365-2656.12397
Legendre P., Fortin M. J., Borcard D. (2015). Should the Mantel test be used in spatial analysis. Methods in Ecol. Evol. 6, 1239–1247. doi: 10.1111/2041-210X.12425
Leibold M. A., Holyoak M., Mouquet N., Amarasekare P., Chase J. M., Hoopes M. F., et al. (2004). The metacommunity concept: a framework for multi-scale community ecology. Ecol. Lett. 7, 601–613. doi: 10.1111/j.1461-0248.2004.00608.x
Lemm J. U., Venohr M., Globevnik L., Stefanidis K., Panagopoulos Y., Gils J., et al. (2021). Multiple stressors determine river ecological status at the European scale: Towards an integrated understanding of river status deterioration. Global Change Biol. 27, 1962–1975. doi: 10.1111/gcb.15504
Li Z., Xing Y., Liu Z., Chen X., Jiang X., Xie Z., et al. (2020). Seasonal changes in metacommunity assembly mechanisms of benthic macroinvertebrates in a subtropical river basin. Sci. Total Environ. 729, 139046. doi: 10.1016/j.scitotenv.2020.139046
Li T., Zhang M. H., Li B., Cai G. Q., Li S., Nie X. (2022). Co-occurrence patterns and community assembly mechanisms of benthic foraminiferal communities in South Chinese bays. Ecol. Indic. 144, 109489. doi: 10.1016/j.ecolind.2022.109489
Liu L. M., Chen H. H., Liu M., Yang J. R., Xiao P., Wilkinson D. M., et al. (2019). Response of the eukaryotic plankton community to the cyanobacterial biomass cycle over 6 years in two subtropical reservoirs. ISME J. 13, 2196–2208. doi: 10.1038/s41396-019-0417-9
Ma Y., Yu Z., Jia S., Wu N., Yin K., Wang Y., et al. (2023). Multiple anthropogenic stressors influence the taxonomic and functional homogenization of macroinvertebrate communities on the mainstream of an urban-agricultural river in China. J. Environ. Manage. 341, 118017–118017. doi: 10.1016/j.jenvman.2023.118017
Maloney K. O., Munguia P. (2011). Distance decay of similarity in temperate aquatic communities: effects of environmental transition zones, distance measure, and life histories. Ecography 34, 287–295. doi: 10.1111/j.1600-0587.2010.06518.x
Mamet S. D., Redlick E., Brabant M., Lamb E. G., Helgason B. L., Stanley K. G., et al. (2019). Structural equation modeling of a winnowed soil microbiome identifies how invasive plants re-structure microbial networks. ISME J. 13, 1988–1996. doi: 10.1038/s41396-019-0407-y
Marquardt G. C., Blanco S., Bicudo C. (2018). Distance decay as a descriptor of the diatom compositional variation in tropical reservoirs. Mar. Freshw. Res. 69, 105–105. doi: 10.1071/mf17003
Martiny J. B., Eisen J. A., Penn K., Allison S. D., Horner-Devine M.C. (2011). Drivers of bacterial β-diversity depend on spatial scale. P. Natl. Acad. Sci. U.S.A. 108, 7850–7854. doi: 10.1073/pnas.1016308108
Mazaris A. D., Moustaka-Gouni M., Michaloudi E., Bobori D. C. (2010). Biogeographical patterns of freshwater micro- and macroorganisms: a comparison between phytoplankton, zooplankton and fish in the eastern Mediterranean. J. Biogeography 37, 1341–1351. doi: 10.1111/j.1365-2699.2010.02294.x
Morse J. C., Yang L., Tian L. (1994). Aquatic insects of China useful for monitoring water quality. (Nanjing: Hohai University Press).
Nekola J. C., White P. S. (1999). The distance decay of similarity in biogeography and ecology. J. Biogeography 26, 867–878. doi: 10.1046/j.1365-2699.1999.00305.x
Ning D., Deng Y., Tiedje J. M., Zhou J. (2019). A general framework for quantitatively assessing ecological stochasticity. Proc. Natl. Acad. Sci. United States America 116, 16892–16898. doi: 10.1073/pnas.1904623116
Ning D., Yuan M., Wu L., Zhang Y., Guo X., Zhou X., et al. (2020). A quantitative framework reveals ecological drivers of grassland microbial community assembly in response to warming. Nat. Commun. 11, 4717. doi: 10.1038/s41467-020-18560-z
Oksanen J., Blanchet F. G., Friendly M., Kindt R., Legendre P., McGlinn D., et al. (2019). Vegan: community ecology package. R package version 2.5-6. Available online at: https://CRAN.R-project.org/package=vegan.
Östman Ö., Drakare S., Kritzberg E. S., Langenheder S., Logue J. B., Lindström E. S. (2010). Regional invariance among microbial communities. Ecol. Lett. 13, 118–127. doi: 10.1111/j.1461-0248.2009.01413.x
Peres-Neto P. R., Legendre P., Dray S., Borcard D. (2006). Variation partitioning of species data matrices: estimation and comparison of fractions. Ecology 87, 2614–2625. doi: 10.1890/0012-9658(2006)87[2614:vposdm]2.0.co;2
Preston F. W. (1962). The canonical distribution of commonness and rarity: part II. Ecology 43, 410–432. doi: 10.2307/1933371
Qu Y., Wu N., Guse B., Fohrer N. (2018). Riverine phytoplankton shifting along a lentic-lotic continuum under hydrological, physiochemical conditions and species dispersal. Sci. Total Environ. 619, 1628–1636. doi: 10.1016/j.scitotenv.2017.10.139
R Core Team (2022). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/ (Accessed May 13, 2024).
Sloan W. T., Lunn M., Woodcock S., Head I. M., Nee S., Curtis T. P. (2006). Quantifying the roles of immigration and chance in shaping prokaryote community structure. Environ. Microbiol. 8, 732–740. doi: 10.1111/j.1462-2920.2005.00956.x
Soininen J., Korhonen J. J., Karhu J., Vetterli A. (2011). Disentangling the spatial patterns in community composition of prokaryotic and eukaryotic lake plankton. Limnology Oceanography 56, 508–520. doi: 10.4319/lo.2011.56.2.0508
Soininen J., McDonald R., Hillebrand H. (2007). The distance decay of similarity in ecological communities. Ecography 30, 3–12. doi: 10.1111/j.0906-7590.2007.04817.x
Spasojevic M. J., Suding K. N. (2012). Inferring community assembly mechanisms from functional diversity patterns: the importance of multiple assembly processes. J. Ecol. 100, 652–661. doi: 10.1111/j.1365-2745.2011.01945x
Sutherland W. J., Freckleton R. P., Godfray H. C. J., Beissinger S. R., Benton T., Cameron D. D., et al. (2013). Identification of 100 fundamental ecological questions. J. Ecol. 101, 58–67. doi: 10.1111/1365-2745.12025
Swenson N. G., Enquist B. J. (2009). Opposing assembly mechanisms in a Neotropical dry forest: implications for phylogenetic and functional community ecology. Ecology 90, 2161–2170. doi: 10.1890/08-1025.1
Thorp J. H., Covich A. P., Thorp J. H., Covich A. P. (2001). Ecology and classification of north American freshwater invertebrates. 2nd ed. (San Diego (California): Academic Press).
Tolonen K. T., Vilmi A., Karjalainen S.-M., Hellsten S., Sutela T., Heino J. (2017). Ignoring spatial effects results in inadequate models for variation in littoral macroinvertebrate diversity. Oikos 126, 852–862. doi: 10.1111/oik.03587
Valerio F., Godinho S., Ferraz G., Pita R., Gameiro J., Silva B., et al. (2024). Multi-temporal remote sensing of inland surface waters: A fusion of sentinel-1&2 data applied to small seasonal ponds in semiarid environments. Int. J. Appl. Earth Observation Geoinformation 135, 104283. doi: 10.1016/j.jag.2024.104283
Whittaker R. H. (1960). Vegetation of the siskiyou mountains, oregon and California. Ecol. Monogr. 30, 279–338. doi: 10.2307/1943563
Yang Z., He S., Feng T., Lin Y., Chen M., Li Q., et al. (2023). Spatial variation in the community structure and response of benthic macroinvertebrates to multiple environmental factors in mountain rivers. J. Environ. Manage. 341, 118027. doi: 10.1016/j.jenvman.2023.118027
Yang Y., Yi Y., Zhou Y., Wang X., Zhang S., Yang Z. (2020). Spatio-temporal variations of benthic macroinvertebrates and the driving environmental variables in a shallow lake. Ecol. Indic. 110, 105948. doi: 10.1016/j.ecolind.2019.105948
Yue L., Li B., Zhu S., Yuan Q., Shen H. (2023). A fully automatic and high-accuracy surface water mapping framework on Google Earth Engine using Landsat time-series. Int. J. Digital Earth. 16, 210–233. doi: 10.1080/17538947.2023.2166606
Zhang Y., Cheng L., Li K., Zhang L., Cai Y., Wang X., et al. (2019). Nutrient enrichment homogenizes taxonomic and functional diversity of benthic macroinvertebrate assemblages in shallow lakes. Limnol. Oceangr. 64, 1047–1058. doi: 10.1002/lno.11096
Zhang Q., Yang T., Wan X., H. Wang Y., Wang W. (2021). Community characteristics of benthic macroinvertebrates and identification of environmental driving factors in rivers in semi-arid areas - A case study of Wei River Basin, China. Ecol. Indic. 121, 107153. doi: 10.1016/j.ecolind.2020.107153
Zhao J., Gao Q., Zhou J., Wang M., Liang Y., Sun B., et al. (2019). The scale dependence of fungal community distribution in paddy soil driven by stochastic and deterministic processes. Fungal Ecol. 42, 100856–100856. doi: 10.1016/j.funeco.2019.07.010
Zhao X., Ma Y., Xie H., Du C., Zhan A., Xu J., et al. (2024). Spatial distribution of benthic taxonomic and functional diversity in the Yellow River Basin: From ecological processes to associated determinant factors. Environ. Int. 188, 108745–108745. doi: 10.1016/j.envint.2024.108745
Zhao Y., Xia X., Yang Z., Xia. N. (2011). Temporal and spatial variations of nutrients in baiyangdian lake, north China. J. Environ. Inf. 17, 102–108. doi: 10.3808/jei.201100192
Zhou S., Wu N., Zhang M., Peng W., He F., Guo K., et al. (2020). Local environmental, geo-climatic and spatial factors interact to drive community distributions and diversity patterns of stream benthic algae, macroinvertebrates and fishes in a large basin, Northeast China. Ecol. Indic. 117, 106673. doi: 10.1016/j.ecolind.2020.106673
Keywords: macroinvertebrate, Daqing River, river-lake system, spatial distribution, distance-decay relationship, community assembly
Citation: Si Y, Wang J, Li H, Li H, He P, Jin L and Liu C (2025) Waterbody type dependence of the spatial distribution of macroinvertebrate communities in temperate China. Front. Ecol. Evol. 13:1538755. doi: 10.3389/fevo.2025.1538755
Received: 03 December 2024; Accepted: 07 March 2025;
Published: 27 March 2025.
Edited by:
Matteo Marcantonio, Université Catholique de Louvain, BelgiumReviewed by:
Francesco Valerio, Centro de Investigacao em Biodiversidade e Recursos Geneticos (CIBIO-InBIO), PortugalTan Lu, Chinese Academy of Sciences (CAS), China
Copyright © 2025 Si, Wang, Li, Li, He, Jin and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cunqi Liu, bGl1Y3VucWlAc2luYS5jb20=; Lei Jin, aGJ1amlubGVpQGhidS5lZHUuY24=
†These authors have contributed equally to this work