 Nathália Menezes de Almeida1,2
Nathália Menezes de Almeida1,2 Rayane Ferreira Dias2
Rayane Ferreira Dias2 Dálete Cássia Vieira Alves1,2Sergio Lisboa Machado3
Dálete Cássia Vieira Alves1,2Sergio Lisboa Machado3 Júlia dos Santos Silva2
Júlia dos Santos Silva2 Cecilia Ferreira de Mello2
Cecilia Ferreira de Mello2 Jeronimo Alencar2*
Jeronimo Alencar2*- 1Programa de Pós-Graduação em Biologia Animal, Instituto de Biologia, Universidade Federal Rural do Rio de Janeiro, Seropédica, Brazil
- 2Laboratório de Diptera, Instituto Oswaldo Cruz (FIOCRUZ), Rio de Janeiro, Brazil
- 3Laboratório de Diagnóstico Molecular e Hematologia, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil
Introduction: Phytotelmata are plant structures that collect and retain water, serving as natural larval habitats. Certain species of culicids utilize these water-holding structures for larval development, playing a crucial role in water retention and nutriente cycling, which benefits resident fauna in tropical forests, such as the Atlantic Forest. This study aimed to characterize the taxonomic composition, diversity, species richness, and abundance of mosquitoes within three types of phytotelmata in an Atlantic Forest remnant.
Methods: Monthly monitoring of mosquito populations was conducted over one year, from March 2023 to February 2024, considering abiotic factors such as temperature, pH, and rainfall, which can influence mosquito larval growth and abundance. Sampling occurred in selected phytotelm habitats along trails within the study area.
Results: A total of 2,672 culicid specimens were collected, representing 11 genera and 29 species. The most abundant species were Wyeomyia artrostigma, Culex pleuristriatus, and Onirion personatum. Bamboo internodes yielded the highest abundance of specimens (67.4%), followed by bromeliads (30.7%) and buttress roots (1.9%). This distribution pattern may be due to the varying suitability of diferente phytotelmata as larval habitats, with bamboo internodes providing more favorable environmental conditions for larval survival. Among types of phytotelmata, bamboo internodes had the highest species richness (22 species) and the largest total number of specimens, highlighting this habitat’s importance for the development of many mosquito species. In contrast, buttress roots, despite having fewer specimens, showed the greatest diversity and evenness, indicating a balanced species distribution within this habitat. Kruskal-Wallis analysis revealed significant differences in species abundance among the types of phytotelmata evaluated, suggesting that mosquito population composition and abundance are habitat-dependent.
Discussion: These results support the idea that different phytotelmata offer distinct ecological conditions, influencing the occurrence and development of culicid populations. Notable among the species collected were Aedes albopictus and Haemagogus leucocelaenus, known vectors of human pathogens. This study provides valuable insights into the mosquito fauna associated with phytotelmata in an Atlantic Forest remnant, contributing to our understanding of the diversity and ecological dynamics of these environments. Our findings also have public health implications, demonstrating the presence of vector species in a natural habitat, which could inform future research and disease control strategies.
Introduction
The Atlantic Forest is considered one of the world’s biodiversity hotspots, meaning it is a global priority for biodiversity conservation (Myers et al., 2000; Mittermeier et al., 2004). This biome is therefore of great importance for studying mosquito fauna that serves as vectors of pathogens, primarily due to its high diversity of flora, vertebrates, and invertebrates, which offers multiple niches for mosquitoes (Alencar et al., 2015).
Most culicid species are found in tropical forests, which host a megadiversity of species and encompass a variety of unique microhabitats. Some species utilize phytotelmata-water-retaining aquatic ecosystems formed by plant structures for oviposition. These natural microhabitats (phytotelmata) are not only vital as temporary larval water habitats but also as rich and diverse ecosystems. They sustain a wide range of organisms, including mosquito species that rely on these environments for reproduction, while also providing shelter and food resources for other species, such as arthropods, amphibians, and even small vertebrates (Olano et al., 1997; Forattini et al., 1998; Cunha et al., 2002).
Phytotelmata are important not only as temporary larval water habitats but also as rich and diverse ecosystems that support a variety of organisms, including mosquito species that depend on these environments to reproduce (Kitching, 2000; Jocque et al., 2013; Ramos and do Nascimento Moura, 2019). These natural environments that accumulate water are a clear example of how microhabitats can influence biodiversity on a large organismal scale (Maguire, 1971; Kitching, 2000). Considering these factors when studying tropical ecosystems is essential to understanding the complexity of ecological relationships and the impacts of environmental change.
Due to their unique characteristics and relative ease of manipulation in experiments, phytotelmata have been studied for their biodiversity and used as microcosms for testing ecological hypotheses (Kitching, 2001). Among the larval phytotelm habitats, those associated with carnivorous plants, bamboo internodes, and bromeliads (Bromeliaceae) have received the most attention. These structures include bromeliad leaves, flowers, leaf axils, natural depressions in tree trunks, open fruits, and even fallen leaves on the forest floor. Phytotelmata provide a protected environment with the resources needed for larval development, making them essential for the survival and dispersal of mosquito populations in tropical forest areas. Furthermore, the diversity of phytotelmata within a forest can directly influence the distribution and abundance of different culicid species, reflecting the complexity and interdependence of tropical ecosystems (Fish, 1983; Reis et al., 2010).
Larval habitats such as bromeliads, tree holes, and bamboo internodes have long been considered some of the oldest natural breeding environments for mosquitoes. It is important to emphasize the ability of phytotelmata to store rainwater, in association with organic matter, in quantities sufficient to sustain the viability of these insects’ developmental cycles even after rainfall has ceased (Jenkins and Carpenter, 1946). While phytotelmata can potentially occur in a wide range of environments, these aquatic habitats are most prolific in tropical forests due to the higher concentration of plant species diversity and humidity in these regions (Greeney, 2001).
Climate change, physical aspects of water, and environmental degradation can enhance water availability in phytotelmata. Consequently, these factors can alter the habitat, attracting a variety of organisms that serve as food for mosquito immatures and affecting the diversity of mosquito species (Kitching, 2001).
This trophic dynamic contributes to the structuring of the mosquito community, where different species compete or interact in ways that can influence their populations. The increase in mosquito diversity in these environments may have significant implications for public health, as many species not only cause discomfort to humans due to the hematophagous habits of females but also possess the ability to transmit pathogens (Gould and Higgs, 2008).
Understanding the life cycles and habits of mosquitoes that develop in phytotelm habitats is crucial for the development of new tools and methods for monitoring and controlling these populations, thereby contributing to the formulation of effective strategies for entomological and epidemiological surveillance. In summary, research with this focus provides valuable insights into ecological interactions and population dynamics, contributing significant insights to public health.
In light of the above, this study aimed to evaluate the potential of three types of phytotelmata as larval habitats for mosquitoes by comparing their diversity, richness, and abundance. We also emphasized the temporal variability related to abiotic factors such as temperature, pH, and rainfall and their impact on the mosquito populations in a remnant of the Atlantic Forest. Studies comparing the diversity of mosquito fauna in phytotelmata generally report a higher diversity index in bromeliad species. This is attributed to their complex structures, which create more favorable environmental conditions. Thus, we can hypothesize that the faunal diversity associated with bromeliads surpasses that found in bamboo and buttress roots due to the specific characteristics of each type of phytotelmata and their varying capacities to support biological diversity.
Materials and methods
Ethics statement
The permanent license for the collection, capture, and transport of zoological material was granted by the Chico Mendes Institute for Biodiversity Conservation (ICMBio) and the System for Authorization and Information on Biodiversity (SISBIO) under License No. 84318-4.
Study area
The study was conducted at the Guapiaçu Ecological Reserve (REGUA), a conservation unit located in the municipality of Cachoeiras de Macacu, state of Rio de Janeiro, Brazil (Figure 1). REGUA encompasses approximately 7,385 hectares of Dense Ombrophilous Forest, characterized by a rich diversity of flora and fauna. The reserve is situated within the Atlantic Forest biome and is crucial for biodiversity conservation and protection of the region’s water resources. In the lower areas near the reserve’s boundaries, there is a predominance of pioneer species, which are plants adapted to higher luminosity and disturbance conditions. As one moves to higher altitudes within the reserve, the vegetation becomes more complex and approaches ecological climax, characterized by species that make up the more mature and stable phases of forest succession (Veloso et al., 1991).
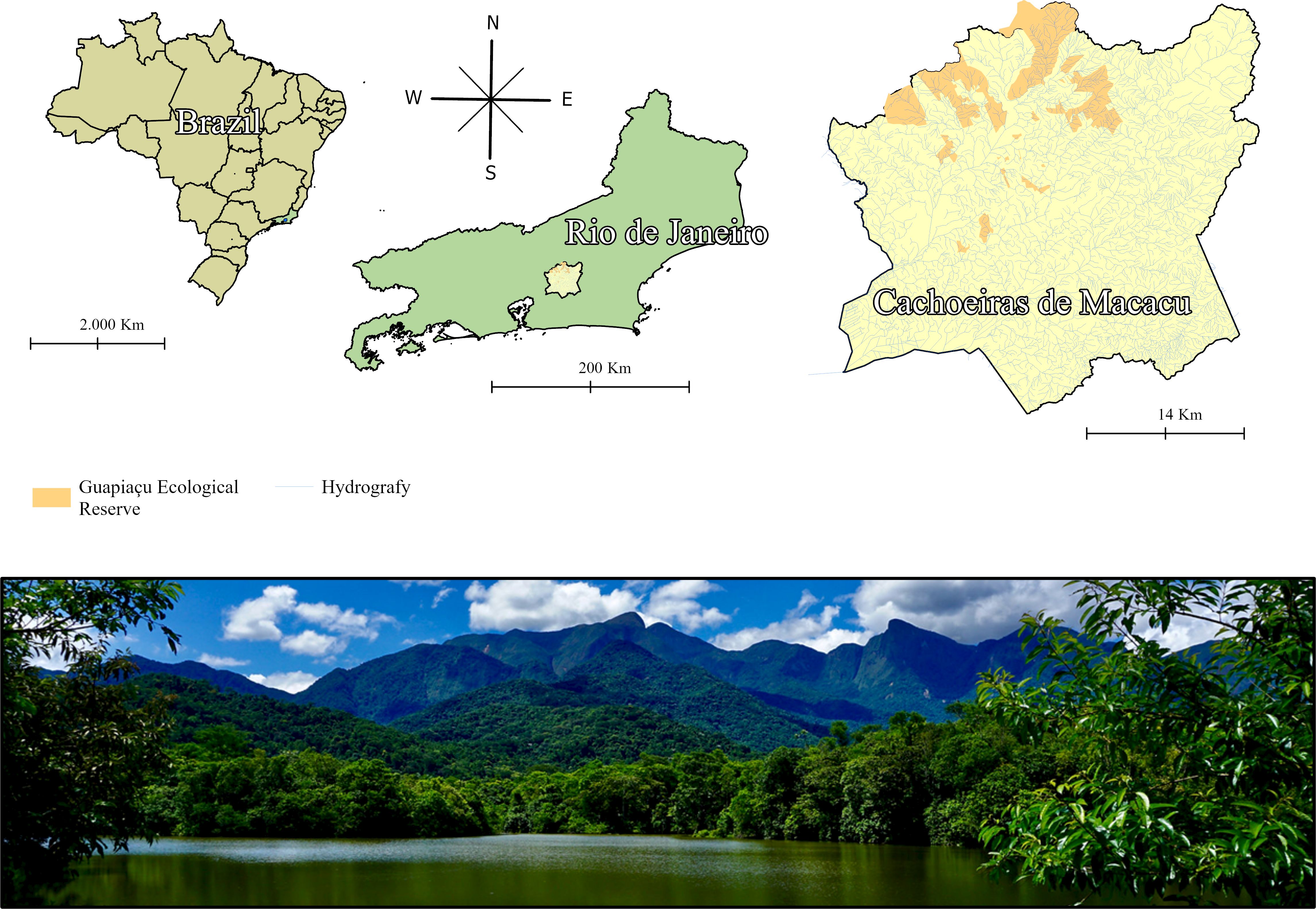
Figure 1. Sampling location at the Guapiaçu Ecological Reserve (REGUA), Cachoeiras de Macacu, Rio de Janeiro, Brazil.
The climate is warm, humid, and lacks a distinctly dry season, classified as type Af according to Köppen (1948). July is the month with the lowest average temperature and rainfall (17.9°C and 59.3 mm), while January and February experience the highest average temperatures (25.3°C) and rainfall (337.8 mm).
The average annual rainfall is 2,095 mm, with December and January as the wettest months, while June and July are the driest. A portion of the REGUA area is included within the Três Picos State Park, and the remainder of the reserve is located in the park’s buffer zone. The vegetation in REGUA is divided into two altitudinal zones: the Submontane Dense Ombrophilous Forest, extending up to 500 m in elevation, and the Montane Dense Ombrophilous Forest, found between 500 and 1,500 m. REGUA constitutes part of the central corridor of the Atlantic Forest, connecting significant conservation units such as the Serra dos Órgãos National Park, the Macaé de Cima Ecological Reserve, and the Paraíso State Ecological Station.
Collections of immatures in phytotelmata
The collections were conducted monthly over two consecutive days, between 9:00 AM and 4:00 PM, from March 2023 to February 2024. Active searches for mosquito immatures were carried out along different trails in the sampling area, inspecting the three most representative types of phytotelmata: bromeliads, bamboo internodes, and buttress roots. Mosquito larvae were sampled in phytotelmata habitats using an active collection method involving the targeted search for larval habitats based on water availability in the selected environments. Ten bamboo plants were selected and artificially perforated using a 12V 1.5Ah lithium battery-powered drill with an 11x150 mm wood bit. Simultaneously, 40 bromeliad plants were chosen and periodically examined—always the same plants—to collect mosquito immatures whenever water was present in the phytotelmata. For buttress root phytotelmata, a single site within the sampling area was identified, where mosquito immatures were collected over 12 months, contingent on the presence of water.
The analysis of three specific types of phytotelmata enables a more controlled and detailed study, ensuring that habitat comparisons are both direct and meaningful. To ensure clarity and objectivity, a subset of phytotelmata was chosen based on prior studies or the characteristics of the local environment, focusing on those deemed most relevant or accessible (Alencar et al., 2016). Sampling was intentionally limited to a smaller number of phytotelmata to control variables better and reduce the influence of unknown or poorly understood factors that might affect the results. It is worth noting that not all phytotelmata are equally suitable for all mosquito species; some may possess characteristics that make them less favorable as larval habitats.
This methodological approach aimed to understand the interaction between these plant species and the occurrence of mosquito immatures in early developmental stages, thereby providing insights into the local ecology and the dynamics of infestation by these insects.
The sampling was conducted using polyethylene pipettes, entomological spoons, and a manual suction pump. The immatures and pupae were pipetted and transferred to 250 ml plastic bags (Whirl-Pak® bags, BioQuip®). Each sample was identified by location, date, type of larval habitat, and water volume. The samples were then transported live, along with water from the phytotelmata to the laboratory. Information on the abiotic variables, temperature, and pH was obtained using an Akso probe, model AK90.
In the laboratory, the immatures were kept in white polyethylene basins filled with water from the original collection site, which was supplemented with dechlorinated water. In the event of evaporation, the pupae were transferred to small containers and maintained until they reached the adult stage.
Species identification
The identification of mosquito species was based on direct observation of morphological characteristics, utilizing both stereomicroscopy and optical microscopy. Identification keys from Lane (1953), Consoli and de Oliveira (1994), and Forattini (2002) were used, supplemented by consultations of species descriptions and diagnostic characteristics. Specimens were subsequently deposited in the Entomological Collection of the Oswaldo Cruz Institute, cataloged as the “Atlantic Forest Collection.”
Data analysis
The assessment and comparison of differences in mosquito community composition across each type of phytotelmata (bamboo internodes, bromeliads, and buttress roots) were conducted using the Shannon-Wiener diversity index (H’) (Shannon, 1948). To determine if there were significant differences between diversity indices, a t-test was calculated at a 5% significance level using the software Past 3.16 (Hammer et al., 2001). Additionally, Evenness (J) was calculated to examine the distribution of individuals among species, which is directly proportional to diversity and inversely proportional to dominance (D). To assess the most abundant mosquito species, the index of species abundance (ISA) was applied, which was then converted to a scale from zero to one using the standardized index of species abundance (SISA), following the definitions by Roberts and Hsi (1979). In this index, a value of 1 represents the most abundant species.
Rarefaction curves, used to observe the trend of species richness with sample size, were generated as a function of the frequency of captured individuals. A rank-abundance curve was applied to characterize the community profile in each studied environment, utilizing the software Past 3.16 (Shannon, 1948). In this method, species are ranked according to their abundances, allowing the assessment of patterns among dominant, intermediate, and rare species, thereby aiding in understanding community structure.
The Kruskal-Wallis analysis, with a 5% significance level, was used to assess significant differences among mosquito populations across the different phytotelmata, using IBM® SPSS® Statistics Version 23. Dunn’s post-hoc test was applied to identify which breeding sites differed from each other. The influence of climatic factors, such as water temperature, pH, and rainfall, was analyzed using Spearman’s correlation coefficient with confidence levels of 95% and 99%, also in IBM® SPSS® Statistics Version 23.
Results
A total of 2,672 culicid specimens were collected, representing 11 genera and 29 species, with seven species showing occurrence rates above 1%. Among the identified specimens, the species with the highest abundance were Wyeomyia arthrostigma (Lutz, 1905) (50.3% and SISA=0.980), Culex pleuristriatus Theobald, 1903 (24.3% and SISA=0.867), and Onirion personatum (Lutz, 1904) (6.0% and SISA=0.553). Of the total, 67.4% of specimens were captured in bamboo internodes, 30.7% in bromeliads, and only 1.9% in buttress roots. In bamboo internodes, the most numerous and abundant species were Wy. arthrostigma (N=1,333; SISA=1.000), followed by On. personatum (N=160; SISA=0.583). In bromeliads, Cx. pleuristriatus (N=638; SISA=0.948) was most abundant, followed by Wy. (Pho.) edwardsi (Lane & Cerqueira, 1942) (N=87; SISA=0.615). In the buttress root habitat, Culex mollis Dyar & Knab, 1906 (N=27; SISA=0.298) was the most abundant (Tables 1, 2).
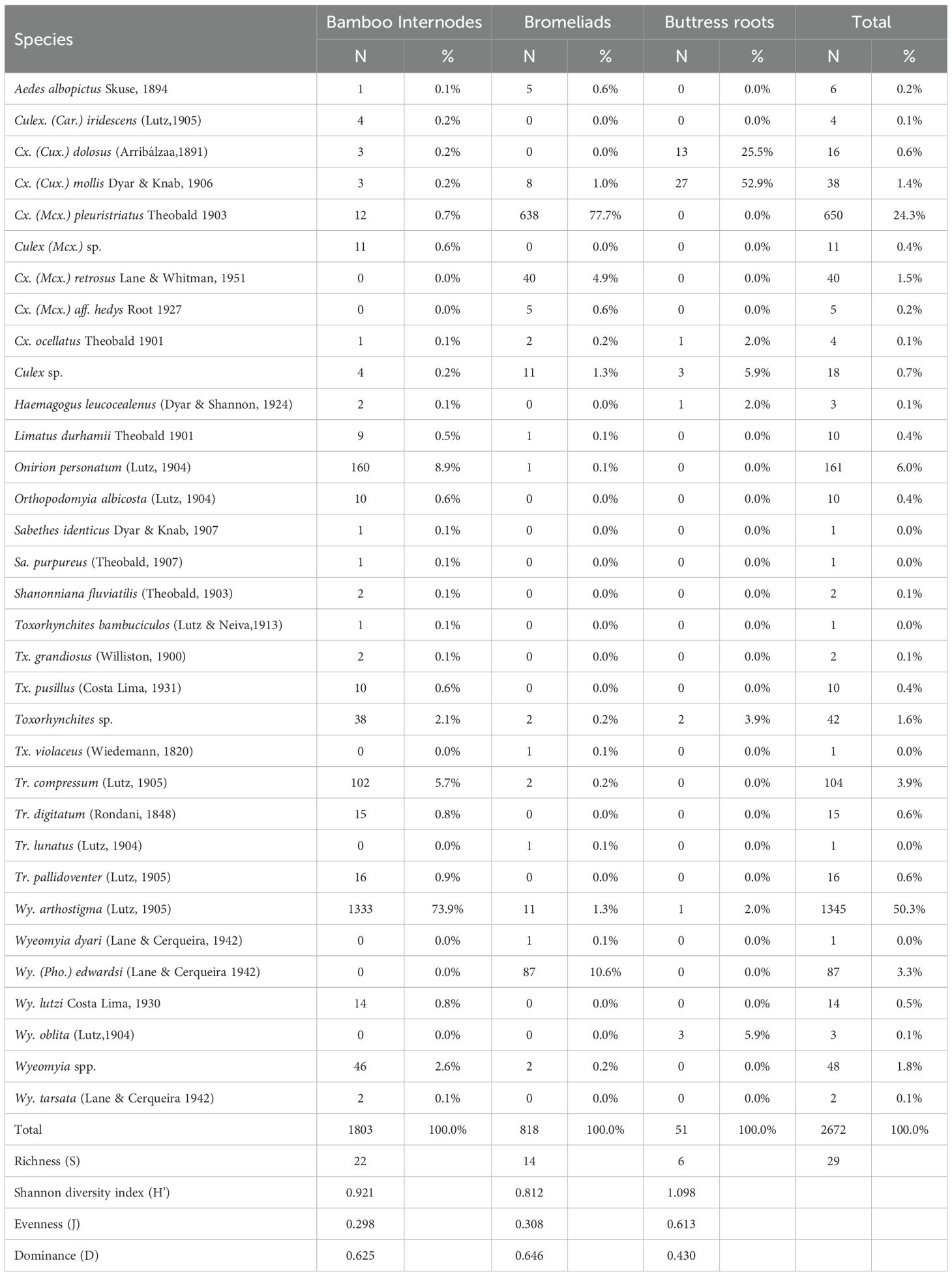
Table 1. Absolute values (N), percentages (%), and ecological indices (richness, dominance, diversity, and Pielou’s evenness) for mosquito immatures by breeding site (bamboo internodes, bromeliads, and buttress roots) at the Guapiaçu Biological Reserve – REGUA, Cachoeiras de Macacu, Rio de Janeiro, Brazil, from March 2023 to February 2024.
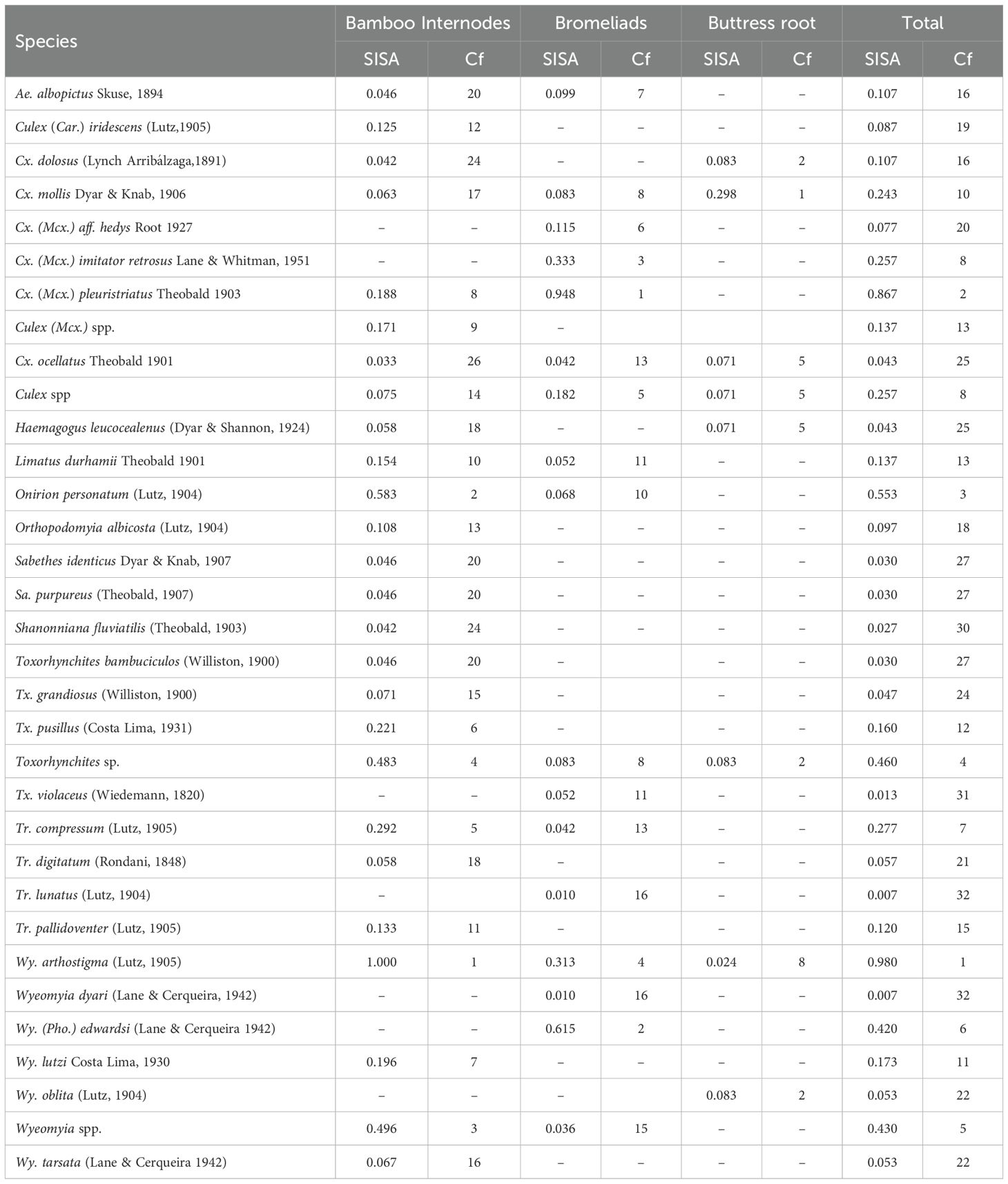
Table 2. Abundance index of species captured at the Guapiaçu Ecological Reserve – REGUA, Cachoeiras de Macacu, Rio de Janeiro, Brazil, indicating the standardized index of species abundance (SISA) and species’ rank (Cf) in different larval habitats.
The bamboo internodes had the highest species richness (S = 22) and the largest number of collected specimens (N = 1,704). However, the greatest diversity and evenness (a more balanced distribution of individuals per species, with less dominance of certain species) were observed in buttress roots (H’ = 1.098 and J = 0.6013). Conversely, bamboo internodes displayed the lowest diversity and evenness, with H’ = 0.298 and J = 0.298, despite their high species richness. This was due to the dominance of a few species, particularly Wy. arthrostigma, which accounted for 73.9% of collected specimens, followed by On. personatum at 3.9. A t-test was conducted to assess differences in diversity across the larval habitats. Results indicated no statistically significant differences in diversity (H’) among bamboo internodes, bromeliads, and buttress roots at the 5% significance level.
Rarefaction curves represent the number of species as a function of the number of collected samples. When the curve approaches stabilization, it indicates that the sampling effort was sufficient to capture most of the species in the area. Bamboo internodes exhibited the highest accumulated species richness, with a taller curve and a tendency toward stabilization, suggesting an almost ideal sampling effort. Bromeliads ranked in an intermediate position, while buttress roots showed lower accumulated richness, with curves exhibiting slight growth toward the end. This suggests additional sampling efforts may reveal more species (Supplementary Figure S1).
Abundance rank curves were created for the three habitats (bamboo internodes, bromeliads, and buttress roots), as shown in Figure 2. In each larval habitat, a different species was dominant: Wy. arthrostigma in bamboo internodes, Cx. pleuristriatus in bromeliads and, to a lesser extent, Cx. mollis in buttress roots. A marked predominance of a few species with high frequency was observed compared to most species with low abundance, particularly in the bamboo and bromeliad habitats, where the steep curves indicate communities with high dominance. In contrast, the abundance distribution in buttress roots was more even, reflecting a higher alpha diversity for this habitat.
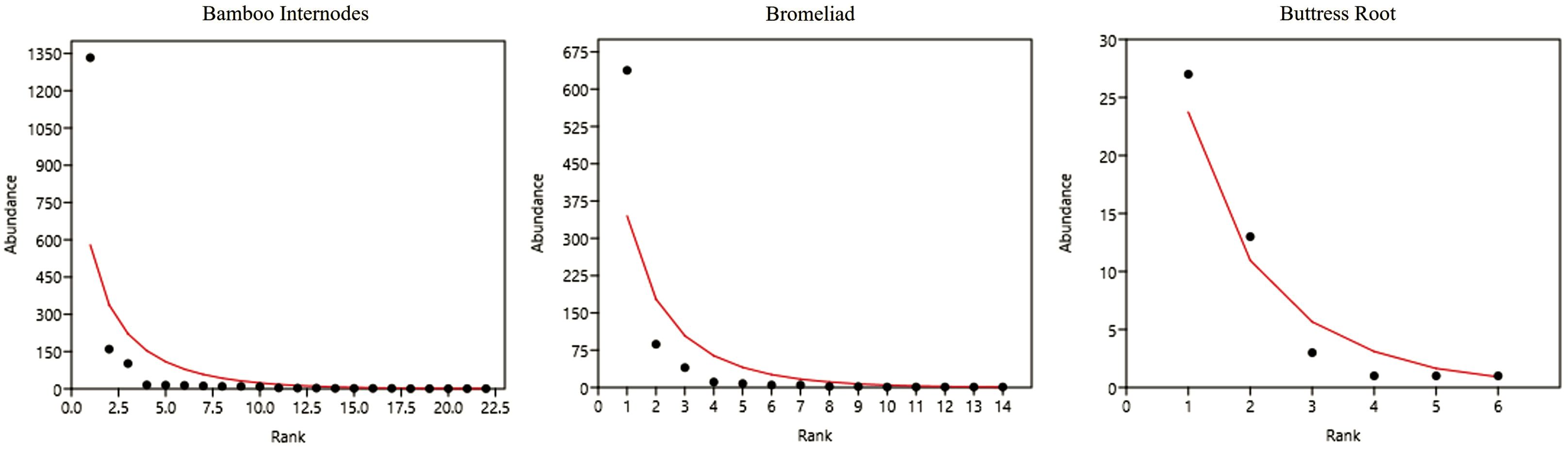
Figure 2. Species abundance rank curves for bamboo internodes, bromeliads, and buttress roots. The Y-axis represents the relative abundance, and the X-axis shows the species rank.
When evaluating mosquito populations in each larval habitat, the Kruskal-Wallis test indicated significant differences between the phytotelmata (p = 0.000). The Dunn post-test was used to identify which habitats showed significant differences from each other (p < 0.05), revealing that populations in bamboo internodes differed from those in buttress roots (p = 0.000).
In bamboo internode habitats, the abundance of Orthopodomyia albicosta (Peryassú, 1908) and Trichoprosopron compressum (Lutz, 1905) decreased with increased water temperature (Spearman’s r = −0.650; r = −0.600). Similarly, an increase in pH was associated with a reduction in On. personatum abundance (Spearman’s r = −0.621). However, rainfall was positively correlated with Or. albicosta (Spearman’s r = 0.622). In bromeliad, a positive correlation was found between water temperature and Wy. arthrostigma (Spearman’s r = 0.621). In buttress roots, no species correlated with the analyzed abiotic factors (Table 3; Supplementary Table S1).
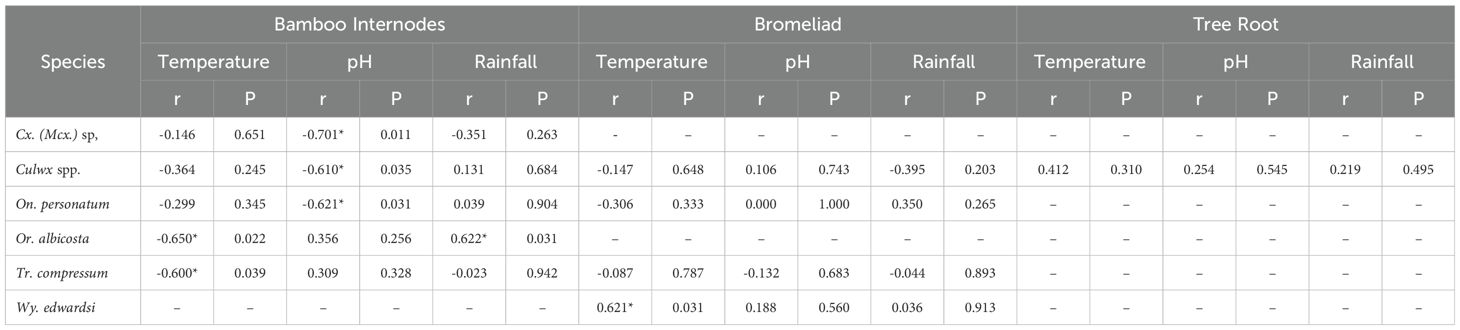
Table 3. Spearman’s correlation coefficient between the abundance of culicids (species with significant correlations) and the abiotic factors—water temperature, pH, and rainfall—from March 2013 to February 2024, Cachoeiras de Macacu, Rio de Janeiro, Brazil. Two-tailed test, with 99% (**) and 95% (*) confidence levels.
Figure 3 shows the population frequency of the seven most abundant species from March 2023 to February 2024. Regarding Culex species, Cx. mollis showed higher abundance in July, August, and November. In contrast, Cx. pleuristriatus had population peaks in April and December, while Culex retrosus Lane & Whitman, 1951 exhibited a population increase during the summer season, specifically in January, February, and March. On. personatum showed multiple peaks throughout the sampling period, with the first peak in June and additional peaks in November and January. Tr. compressum had a single annual peak in May, although it was found in lower numbers in April, June, July, and August. Among Wyeomya species, Wy. (Pho.) edwardsi was predominantly observed in January and February, while Wy. arthrostigma was present in 11 months of the year, except for September (Figure 3).
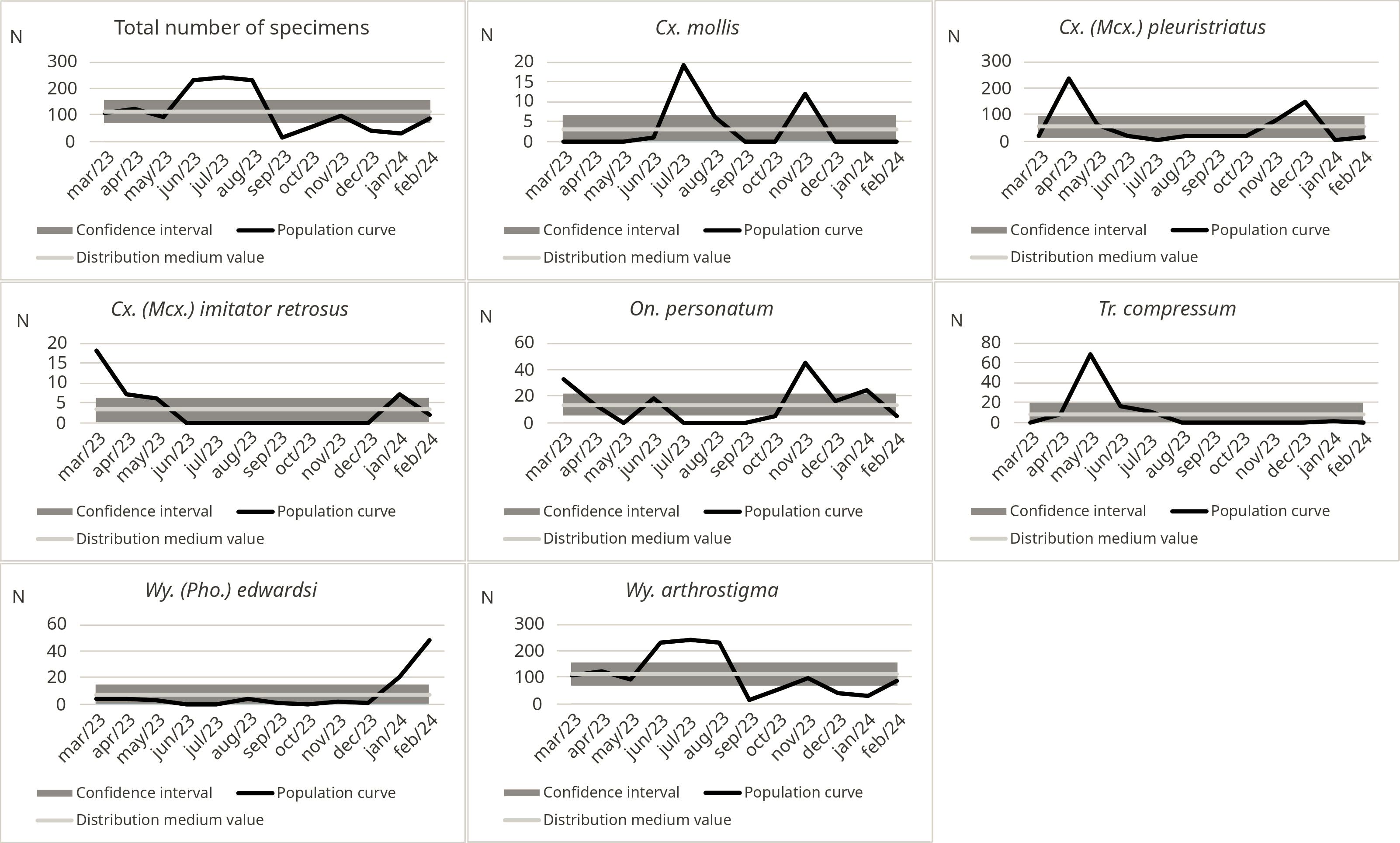
Figure 3. Monthly frequency of the seven most abundant mosquito species and total sum recorded in REGUA, Cachoeiras de Macacu, Brazil, from March 2023 to February 2024.
Considering the total number of specimens, a period of stability was observed between April and May 2023, with around 100 specimens collected. A significant increase in the population curve followed, surpassing the tolerance range in June, July, and August, with more than 200 specimens recorded. Following this peak, the number of specimens gradually decreased, dropping to fewer than 100 in October and reaching the lowest abundance in December 2023, with fewer than 50 specimens. This population fluctuation can be attributed to various factors, including climatic variations, resource availability, and predator interactions. The decrease at the end of the year may be related to decreased temperatures and habitat changes, which influence the activity and life cycle of the collected species. However, a slight increase in the number of specimens was observed in February 2024, suggesting a process of habitat recolonization triggered by the onset of rainfall and the gradual temperature rise (Figure 3).
Discussion
A total of 56 species or genera groups were recorded in a study on mosquito immatures conducted at the Guapiaçu Ecological Reserve, which aimed to evaluate the composition of Culicidae fauna in both natural and artificial larval habitats (Alencar et al., 2016). The most abundant species were Psorophora cingulata (Fabricius, 1805), Culex corniger Theobald, Limatus durhamii Theobald, 1901, Culex usquatus Dyar, 1918, Cx. pleuristriatus, Culex lanei Oliveira Coutinho & Forattini, 1962, and Cx. retrosus. In contrast, the present study recorded only 31 species, with Cx. pleuristriatus as the most abundant, as previously noted by Alencar et al. (2016). The lower species richness observed in our study is likely due to the smaller number of larval habitats evaluated. While we focused on three types of phytotelmata, Alencar et al. (2016) analyzed 13 larval habitats, including both natural and artificial environments.
In the present study, the most abundant species in each phytotelm habitat were Wy. arthrostigma, which was found in the highest numbers in bamboo internodes (Figure 4), corroborating the findings of Nascimento-Pereira et al. (2021); Cx. pleuristriatus, predominant in bromeliads (Figure 5), as reported by Alencar et al. (2016) and Marques and Forattini (2008); and Cx. mollis, in buttress roots (Figure 6), whose immatures are frequently found in tree holes with turbid water and organic material, similar to the phytotelm habitat encountered (Ortega-Morales et al., 2018). The third most frequent species was On. personatum, with 99.38% of specimens (N= 160) found in bamboo internodes and only 0.62% (N= 01) found in bromeliads. In general, the immature forms of this species are commonly found in bamboo internodes and bromeliads (Müller et al., 2014; Stahlhöfer et al., 2022), validating our results.
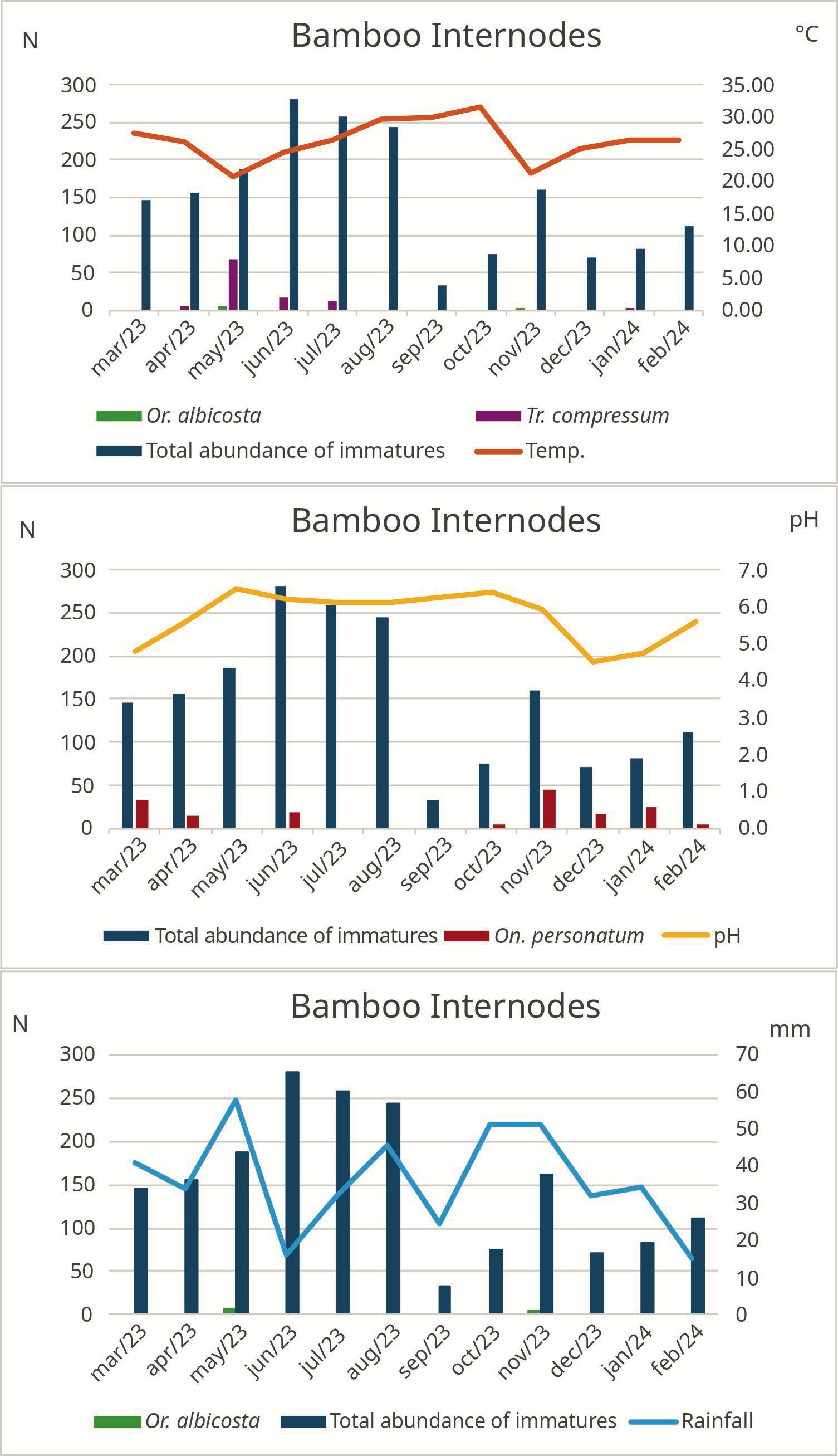
Figure 4. Monthly absolute values of mosquitoes (N), temperature (°C), and pH, considering the total number of specimens and species that showed correlation with abiotic factors, in the bamboo internode phytotelm larval habitat, Cachoeiras de Macacu, Rio de Janeiro, Brazil, from March 2013 to February 2024.
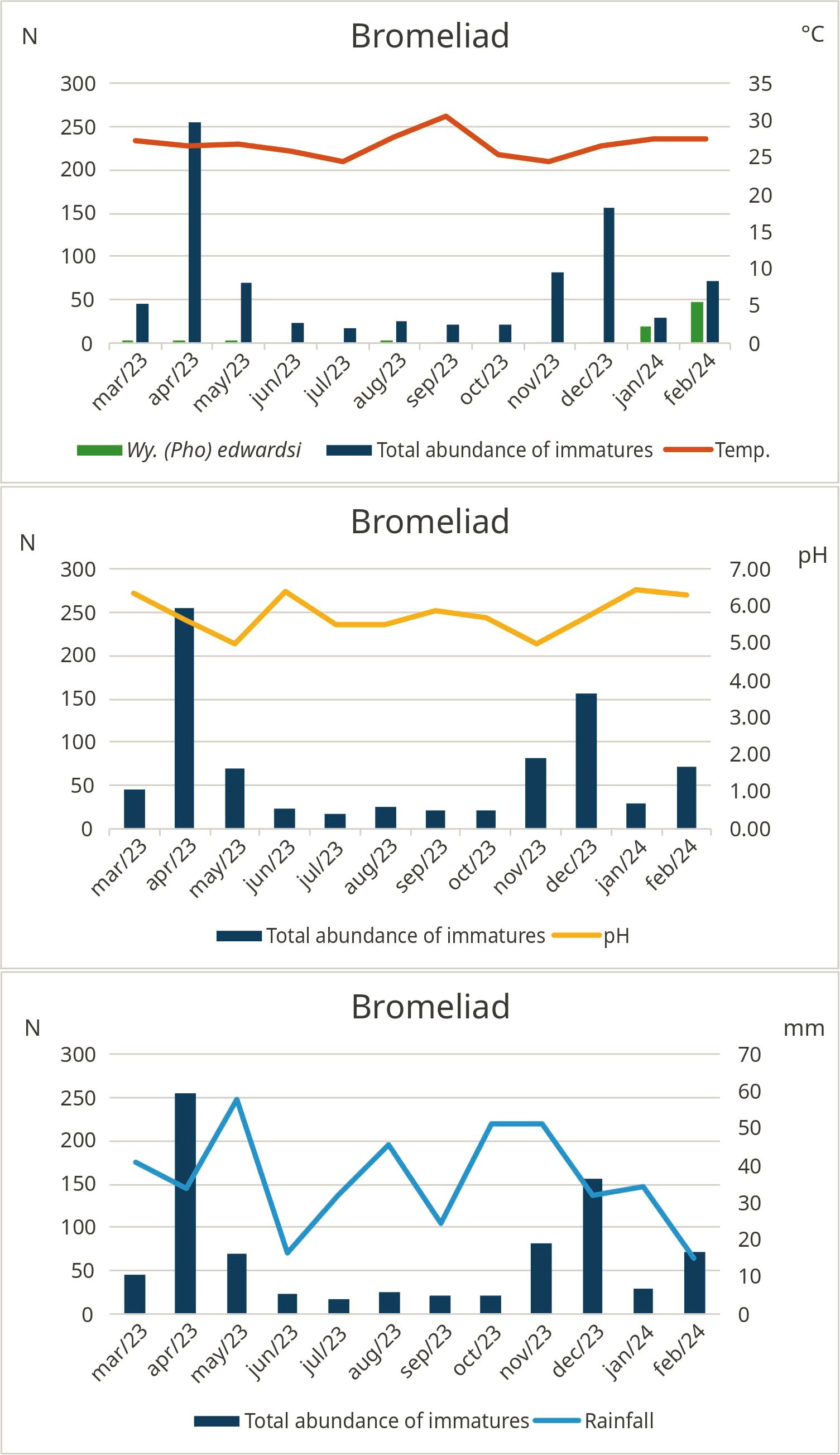
Figure 5. Monthly absolute values of mosquitoes (N), temperature (°C), and pH, considering the total number of specimens and species that showed correlation with abiotic factors, in the bromeliad phytotelm larval habitat, Cachoeiras de Macacu, Rio de Janeiro, Brazil, from March 2013 to February 2024.
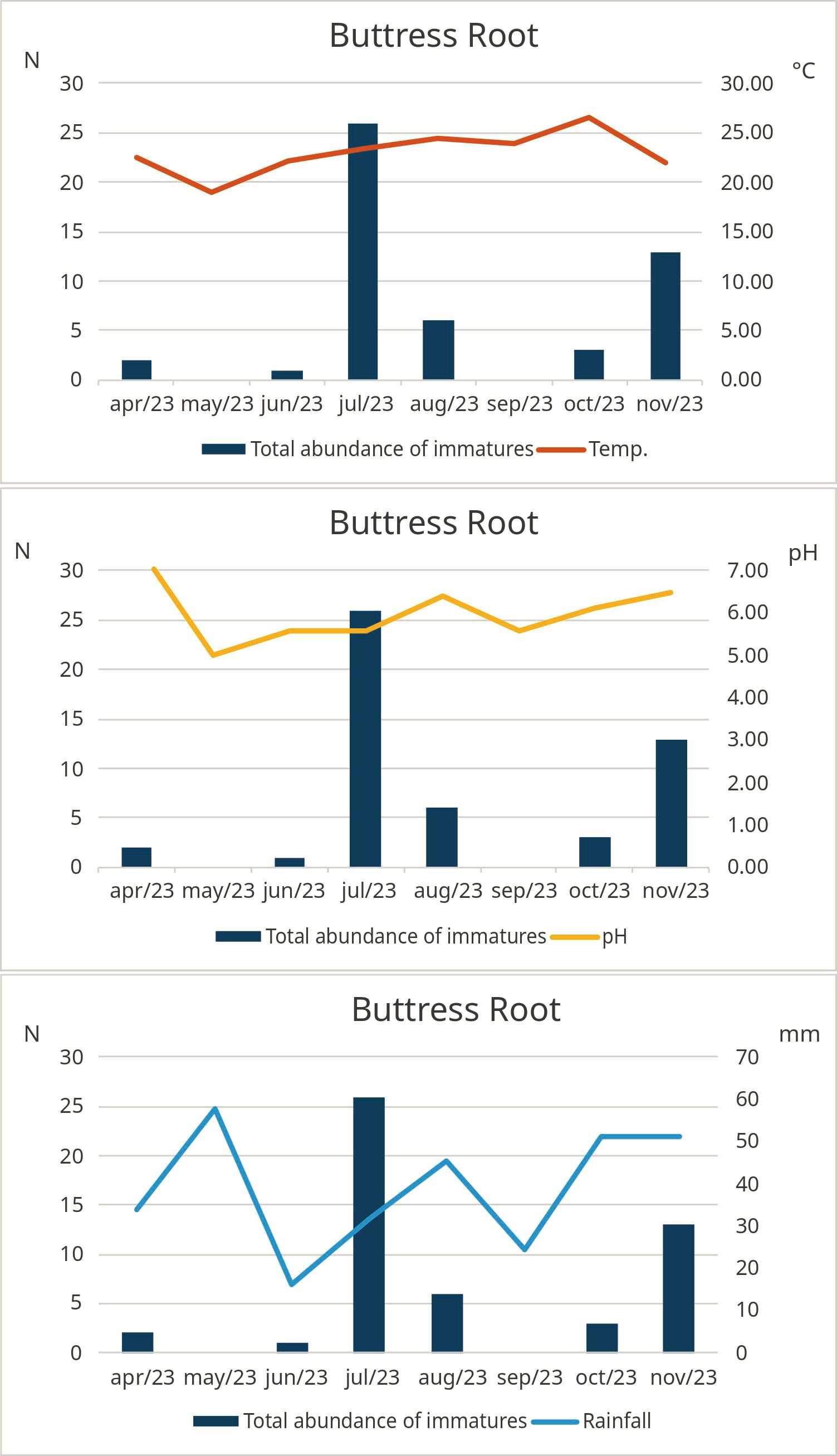
Figure 6. Monthly absolute values of mosquitoes (N), temperature (°C), and pH, considering the total number of specimens in the tree root phytotelm larval habitat, Guapiaçu Ecological Reserve – REGUA, Cachoeiras de Macacu, Rio de Janeiro, Brazil, from March 2013 to February 2024.
The species with the highest population density were found in greater numbers during the rainy months, corroborating the observations of Consoli and de Oliveira (1994), who stated that the rainy season provides favorable environmental and climatic conditions for mosquito proliferation, such as higher average temperatures, elevated relative humidity, and higher water levels in larval habitats. In general, it is expected that a higher volume of water will be present in larval habitats during this period, resulting in greater nutrient transport due to the increased water flow. However, in the present study, only Or. albicosta showed a positive correlation with rainfall. Additionally, Wy. arthrostigma (50.3%) was more abundant during the dry months, which influenced our findings. This effect can be explained by the lower competitiveness during the dry period, as other species were present in smaller numbers.
The diversity of mosquitoes varies across the phytotelm habitats of bamboo, bromeliads, and buttress roots due to differences in the ecological conditions of each environment (Kariyawasam and Wegiriya, 2022). Bromeliads tend to support a higher species diversity because their aquatic cavities offer a more stable environment with lower evaporation and protection from predators, favoring the reproduction of various mosquito species (Cardoso et al., 2015). However, in the present study, buttress roots exhibited the greatest diversity, as they provide moderately stable conditions that promote intermediate biodiversity. In contrast, bamboo internodes showed the lowest diversity, with Wy. arthrostigma strongly predominating, as their cavities have lower water retention capacity and are more susceptible to climatic variations, limiting the presence of other species. These differences emphasize the importance of physical and environmental conditions in determining biodiversity in phytotelm habitats, a relationship well-documented in studies on mosquito ecology (da Silva et al., 2004).
Bamboo internodes had the highest abundance of mosquitoes, followed by bromeliads. These habitats may serve as nutrient reservoirs for mosquito immatures during unfavorable climatic periods, such as the dry season, which could explain the higher number of specimens found (Macdonald and Traub, 1960). da Silva et al. (2007) emphasized that larval habitats like bromeliads, due to their small size, simplicity, and abundance, offer advantages for ecological studies in tropical regions, such as investigations into community structures, colonization, and dispersion.
The higher abundance of mosquitoes in bamboo internodes can be explained by the ability of these habitats to provide stable resources and essential nutrients for larval development, especially under adverse climatic conditions when other aquatic environments may be limited (Bastos et al., 2021). Bromeliads, which are common habitats that are easily colonized, also play an important part by offering favorable conditions for colonization and the maintenance of mosquito populations (Frank, 1983; Torreias Sr da et al., 2010).
Species of Haemagogus and Sabethes are recognized as primary sylvatic vectors of yellow fever (Arnell, 1973; Vasconcelos, 2003). However, the occurrence of these species in a given region can be influenced by ecological variables such as habitat type. In the present study, Haemagogus leucocelaenus (Dyar & Shannon), 1924, was found in low abundance in bamboo internodes and buttress roots. In contrast, Alencar et al. (2015, 2016), conducting studies in the same sampling area, reported a greater diversity of larval habitats but found no Haemagogus species. This discrepancy may indicate an ecological specialization of Hg. leucocelaenus in specific environments, suggesting that this species may prefer or be better adapted to certain habitat conditions. Thus, the diversity of microhabitats and the environmental conditions present at the sampled locations are key determinants for the occurrence and distribution of these species, highlighting the importance of a detailed approach when investigating the biodiversity and ecological interactions of mosquitoes. Therefore, analyzing diverse habitats and considering local environmental conditions are essential to better understanding the dynamics of mosquito populations and their potential interactions with pathogen transmission, such as yellow fever.
Forattini (2002) reported that Sabethes immatures develop in phytotelm larval habitats, such as tree hollows, bamboo internodes, leaf axils, and water stored in cut tree trunks. Furthermore, they exhibit both predatory and cannibalistic behaviors. In the present study, a low representation of species from this genus was observed, which may be attributed to this ecological behavior. Competition between Sabethes immatures and other organisms sharing the same microhabitats may also contribute to the decline in their populations. The specialization of immatures in specific environments can limit their distribution and abundance, making them more vulnerable to environmental changes and resource availability (Medeiros-Sousa et al., 2015). These findings emphasize the importance of more in-depth studies on the ecology of these organisms and the factors influencing their presence in different habitats. Similarly, immatures of Toxorhynchites species, known for being major predators, also showed a low number of specimens in the current study. Leite et al. (2024), who focused solely on bamboo internodes, reported a reduced number of specimens from this genus.
We collected important species responsible for pathogen transmission, such as Ae. albopictus Skuse, 1894 and Hg. leucocelaenus. Although Ae. albopictus is widely distributed in forested areas of the Atlantic Forest (Lourenço-de-Oliveira et al., 2004), it was not found in large abundance, possibly due to its preference for ovipositing in artificial containers, such as tires and bottles (de Albuquerque et al., 2000; Lopes, 2002). Wyeomyia oblita (Lutz, 1904) was exclusively recorded in buttress roots, confirming previous findings (Kumm, 1933), although it was also observed in perforated bamboo (Ceretti-Júnior et al., 2014). On. personatum was the third most abundant species, found in large numbers in bamboo internodes and fewer in bromeliads, in line with literature reports of its presence in both bamboo internodes and bromeliads (Müller et al., 2014; Stahlhöfer et al., 2022).
We observed a positive correlation between water temperature and Wy. arthrostigma in bromeliads, as well as a positive correlation with rainfall in bamboo internodes. However, a negative correlation was identified between water temperature and Or. albicosta and Tr. compressum in bamboo internodes. The abiotic factor temperature alters mosquito population dynamics by influencing the development of immature stages and reproduction (Mordecai et al., 2019). Although water availability in these larval habitats is essential for mosquito development, excess water during the rainier months was likely drained or wasted by the plants, which may explain the lack of a significant correlation in mosquito populations (Müller and Marcondes, 2007). In general, the populations remained stable, even under adverse conditions. Another important factor was the consistency of rainfall throughout the months, with accumulations ranging from 15.3 mm to 57.7 mm, ensuring relatively uniform hydric conditions. Additionally, On. personatum showed a negative correlation with pH, although no specific studies in the literature address the effect of pH on this species. However, the work by Leite et al. (2024) suggests that mosquito immatures can inhabit high pH ranges, indicating some tolerance to variations regarding this abiotic factor.
The biological behavior of mosquitoes involves the interaction between intrinsic species factors and the environmental conditions of the breeding site. Mosquitoes can select a wide range of habitats for reproduction. Examples include Wyeomyia smithii Coquillet, 1901, which exploits the carnivorous plant Sarracenia purpurea (Bradshaw and Holzapfel, 1989), Tripteroides bambusa Lutz, 1919, which predominates in bamboo internodes (Sunahara and Mogi, 1998; Sunahara and Mogi, 2002), and Toxorhynchites species, which tend to lay eggs in tree holes and other types of phytotelmata.
The selection of these larval habitats can be influenced by abiotic factors such as temperature, water quality, resource availability, predator presence, and habitat persistence. These factors, along with rainfall, can affect immature mosquitoes’ development, growth, and survival (Clements, 1992). Alencar et al. (2016) found that Wyeomyia edwardsi was recorded exclusively in bromeliads, supporting the hypothesis that the species’ larval habitat at REGUA shows a lack of variability in oviposition site selection. In contrast, Wy. arthrostigma was the only species of the genus present in all three types of larval habitats, demonstrating its ability to adapt to different environmental conditions.
During the sampling period of this study, we found that the seven most frequently encountered species were not present in all of the months analyzed. The total abundance of species in bromeliads was higher in April and December, while in bamboo internodes, abundance peaked in June, July, and August. Marteis et al. (2017) reported that in the Caatinga biome, the highest abundance in bromeliads occurred in June and August 2014. In contrast, Bastos et al. (2021), studying mosquitoes in a remnant of the Atlantic Forest, found the highest abundance in bamboo internodes during August and September. These results support the hypothesis that seasonal fluctuations in mosquito abundance are influenced by complex interactions between biotic and abiotic factors. Thus, the differences observed in these studies may be linked to factors such as fauna composition and regional climate variations. For buttress roots, the highest abundance was recorded in July.
Alencar et al. (2016), in their study on mosquito ecology at REGUA, found that the species most abundant in this phytotelmata (buttress roots) was Cx. usquatus, suggesting a changing trend, as the most abundant species in the present study was Cx. mollis. Mosquito species are influenced by various environmental and climatic factors, such as habitat structure, temperature, rainfall, humidity, and resource availability (Juliano, 2009). According to Ward (2008), the availability of water and organic matter determines the viability of the culicid life cycle. Cx. mollis was encountered in all types of phytotelmata evaluated, demonstrating physiological and behavioral traits that allow it to exploit diverse habitats.
During the dry period, typical of the winter months (June, July, and August), Cx. pleuristriatus and Cx. retrosus were not detected. It is important to note that mosquitoes are highly adaptable and sensitive to changes in their environment, and species may experience shifts in their distribution and abundance over time, depending on the factors mentioned above (Juliano, 2009).
Conclusion
Our study on mosquitoes utilizing phytotelm habitats for their development revealed distribution and abundance patterns of species, with emphasis on Wy. arthrostigma, Cx. pleuristriatus, and On. personatum. Bamboo internodes were the larval habitats with the highest number of specimens, followed by bromeliads and buttress roots. This distribution pattern reflects the environmental suitability of different phytotelmata for mosquito larval development, with bamboo internodes being the most favorable, likely due to their higher water retention capacity and environmental stability.
The highest species richness was observed in bamboo internodes (N=22), reinforcing the importance of this habitat as a primary development site for most of the species present. In contrast, although housing fewer specimens, buttress roots exhibited the greatest diversity and evenness, suggesting that this environment offers more balanced conditions for the coexistence of different mosquito species. These results highlight the importance of considering the diversity of phytotelmata when studying the ecology of culicid populations, as each type provides specific resources that affect species survival and reproductive success.
Our research emphasizes the ecological complexity of phytotelmata and their relevance to biodiversity, providing valuable insights for monitoring mosquito vectors of pathogens in tropical areas such as the Atlantic Forest. The REGUA is an important protected environmental area, attracting visitors from around the world due to its biodiversity and the ecological significance of its ecosystems. In this context, the presence of species such as Hg. leucocelaenus and Ae. albopictus, recognized pathogen vectors, underscores the importance of continuous monitoring and its implications for public health risks. These species play key roles in pathogen transmission, making the study of their population dynamics, life cycle, and the impact of environmental variations essential.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
NA: Methodology, Writing – original draft, Writing – review & editing. RD: Methodology, Writing – review & editing. DA: Writing – original draft, Writing – review & editing. SM: Methodology, Writing – original draft. JS: Writing – original draft, Writing – review & editing. CM: Methodology, Writing – original draft. JA: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. I declare that all sources of funding received for the research have been submitted. This work was carried out with the support of CNPq, FAPERJ, and CAPES, as well as Conselho Nacional de Desenvolvimento Científico e Tecnológico (Grant number: 303286/2021-0, JA) and Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (E-26/200.956/2002/2022, JA).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1525202/full#supplementary-material
References
Alencar J., De Mello C. F., Guimarães A. É., Gil-Santana H. R., Silva J. D. S., Santos-Mallet J. R., et al. (2015). Culicidae community composition and temporal dynamics in Guapiaçu Ecological Reserve, Cachoeiras de Macacu, Rio de Janeiro, Brazil. PloS One 10, e0122268. doi: 10.1371/journal.pone.0122268
Alencar J., de Mello C. F., Serra-Freire N. M., Guimarães A. É., Gil-Santana H. R., Gleiser R. M. (2016). Biodiversity and temporal distribution of immature culicidae in the atlantic forest, rio de janeiro state, Brazil. PloS One 11, e0159240. doi: 10.1371/journal.pone.0159240
Arnell J. H. (1973). Mosquitoes studies (Diptera: Culicidae). XXXII. A revision of the genus Haemagogus. Contributions of the American Entomological Institute 10, 1–174.
Bastos A. Q., Leite P. J., de Mello C. F., Maia D. A., MaChado S. L., Gil-Santana H. R., et al. (2021). Bionomy of mosquitoes in bamboo internodes in an atlantic forest remnant of the state of rio de janeiro, Brazil. J. Am. Mosq. Control Assoc. 37, 208–215. doi: 10.2987/21-7044
Bradshaw W. E., Holzapfel C. M. (1989). Life-historical consequences of density-dependent selection in the pitcher-plant mosquito. Wyeomyia smithii. Am. Nat. 133, 869–887. doi: 10.1086/284957
Cardoso C. A. A., Lourenço-de-Oliveira R., Codeço C. T., Motta M. A. (2015). Mosquitoes in bromeliads at ground level of the Brazilian atlantic forest: the relationship between mosquito fauna, water volume, and plant type. Ann. Entomological Soc. America 108, 449–458. doi: 10.1093/aesa/sav040
Ceretti-Júnior W., Medeiros-Sousa A. R., Multini L. C., Urbinatti P. R., Vendrami D. P., Natal D., et al. (2014). Immature mosquitoes in bamboo internodes in municipal parks, city of são paulo, Brazil. J. Am. Mosq. Control Assoc. 30, 268–274. doi: 10.2987/14-6403R.1
Clements A. N. (1992). The biology of mosquitoes. Development, nutrition and reproduction (Chapman and Hall, London: CABI), 536p.
Consoli R. A. G. B., de Oliveira R. L. (1994). Principais mosquitos de importância sanitária no Brasil (Rio de Janeiro, RJ: Fiocruz), 225 p.
Cunha S. P., Alves J. R. C., Lima M. M., Duarte J. R., de Barros L. C., da Silva J. L., et al. (2002). Presence of Aedes aEgypti in Bromeliaceae and plant breeding places in Brazil. Rev. Saúde Pública 36, 244–245. doi: 10.1590/S0034-89102002000200018
de Albuquerque C. M., Melo-Santos M. A. V., Bezerra M. A. S., Barbosa R. M., Silva D. F., da Silva E. (2000). Primeiro registro de Aedes albopictus em área da Mata Atlântica, Recife, PE, Brasil. Rev. Saúde Pública 34, 314–315. doi: 10.1590/S0034-89102000000300017
Fish D. (1983). “Phytotelmata flora and fauna,” in Phytotelmata: terrestrial plants as host for aquatic insect communities. Eds. Frank J. H., Lounibos L. P. (Plexus Press, Medford, New Jersey), 101–128.
Forattini O. P. (2002). Culicidologia médica: identificação, biologia e epidemiologia (São Paulo, SP: Depertamento de Epidemiologia, Universidade de São Paulo), p. 860.
Forattini O. P., Marques G. R. A. M., Kakitani I., de Brito M., Sallum M. A. M. (1998). Significado epidemiológico dos criadouros de Aedes albopictus em bromélias. Rev. Saúde Pública 32, 186–188. doi: 10.1590/S0034-89101998000200014
Frank J. H. (1983). “Bromeliad phytotelmata and their biota, especially mosquitoes,” in Phytotelmata: terrestrial plants as hosts for aquatic insect communities (Plexus Press, Medford, New Jersey), 101–128.
Gould E. A., Higgs S. (2008). Impact of climate change and other factors on emerging arbovirus diseases. Trans. R. Soc. Trop. Med. Hygiene 103, 109–121. doi: 10.1016/j.trstmh.2008.07.025
Greeney H. F. (2001). The insects of plant-held waters: a review and bibliography. J. Trop. Ecol. 17, 241–260. doi: 10.1017/S026646740100116X
Hammer Ø., Harper D. A. T., Ryan P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4, 1–9.
Jenkins D. W., Carpenter S. J. (1946). Ecology of the tree hole breeding mosquitoes of nearctic north america. Ecol. Monogr. 16, 31–47. doi: 10.2307/1943573
Jocque M., Fiers F., Romero M., Martens K. (2013). Crustacea in phytotelmata: A global overview. J. Crustacean Biol. 33, 451–460. doi: 10.1163/1937240X-00002161
Juliano S. A. (2009). Species interactions among larval mosquitoes: context dependence across habitat gradients. Annu. Rev. Entomology 54, 37–56. doi: 10.1146/annurev.ento.54.110807.090611
Kariyawasam C. S., Wegiriya H. C. E. (2022). Species diversity and coexistence of mosquito larvae breeding in phytotelmata microhabitats; a cross-sectional study from Kalutara district, Western Province, Sri Lanka. Ceylon J. Sci. 51, 63–72. doi: 10.4038/cjs.v51i1.7980
Kitching R. L. (2000). Food webs and container habitats: the natural history and ecology of phytotelmata (Cambridge, New York: Cambridge University Press), 431.
Kitching R. L. (2001). Food webs in phytotelmata: “bottom-up” and “top-down” explanations for community structure. Annu. Rev. Entomology 46, 729–760. doi: 10.1146/annurev.ento.46.1.729
Köppen W. (1948). Climatologia: con um estúdio de los climas de la Tierra (Ciudad de México: Fondo de Cultura Economica), p. 474
Kumm H. W. (1933). Mosquitos breeding in Bromeliads, at Bahia, Brazil. Bull. Entomological Res. 24, 561–573. doi: 10.1017/S0007485300035513
Lane J. (1953). Neotropical Culicidae. 2nd edition (São Paulo, Brazil: Editora Univ. Sao Paulo), p. 1112
Leite M. P. C., Dias R., Leite P. J., Silva S. O. F., Gil-Santana H. R., Barbosa R. P., et al. (2024). Bamboo (Poales, poaceae): an important maintainer of immature mosquitoes (Diptera: culicidae) in a biodiversity hotspot in the city of rio de janeiro, Brazil. Life 13, 351. doi: 10.3390/life14030351
Lopes J. (2002). “Mosquitos (Diptera: Culicidae) da Região do Baixo Tibagi e suas adaptações a ambientes antropogênicos: causas e conseqüências,” in A bacia do rio tibagi. Eds. Medri M. E., Bianchini E., Shibatta O. A., Pimenta J. A. (Londrina: EDUEL), 327–351.
Lourenço-de-Oliveira R., Castro M. G., Braks M. A. H., Lounibos L. P. (2004). The invasion of urban forest by dengue vectors in Rio de Janeiro. J. Vector Ecol. 29, 94–100.
Macdonald W. W., Traub R. (1960). An introduction to the ecology of the mosquitoes of the lowland dipterocarp forest of Selangor, Malaya. Malaysian Parasites XXXV-XLIX. Studies from the Institute of Medical Research, Federation of Malaya, 29, 79–110.
Maguire J. B. (1971). Phytotelmata: biota and community structure determination in plant-held waters. Annu. Rev. Ecol. Systematics 2, 439–464. doi: 10.1146/annurev.es.02.110171.002255
Marques G. R. A. M., Forattini O. P. (2008). Culicídeos em bromélias: diversidade de fauna segundo influência antrópica, litoral de São Paulo. Rev. Saúde Pública 42, 979–985. doi: 10.1590/S0034-89102008000600001
Marteis L. S., Natal D., Sallum M. A. M., Medeiros-Sousa A. R., Corte R. L. (2017). Mosquitoes of the Caatinga: 2. Species from periodic sampling of bromeliads and tree holes in a dry Brazilian forest. Acta Tropica 171, 114–123. doi: 10.1016/j.actatropica.2017.03
Medeiros-Sousa A. R., Ceretti-Júnior W., de Carvalho G. C., Nardi M. S., Araujo A. B., Vendrami D. P., et al. (2015). Diversity and abundance of mosquitoes (Diptera: Culicidae) in an urban park: larval habitats and temporal variation. Acta Tropica 150, 200–2009. doi: 10.1016/j.actatropica.2015.08.002
Mittermeier R. A., Gil P. R., Hoffmann M., Pilgrim J., Brooks T., Mittermeier C. G., et al. (2004). Hotspots revisited. Earth’s biologically richest and most endangered terrestrial ecoregions (Washington, DC: Cemex.: Conservation International), 392.
Mordecai E. A., Caldwell J. M., Grossman M. K., Lippi C. A., Johnson L. R., Neira M., et al. (2019). Thermal biology of mosquito-borne disease. Ecol. Lett. 22, 1690–1708. doi: 10.1111/ele.13335
Müller G. A., Marchi M. J., Marcondes C. B. (2014). Mosquito immature in bamboo internodes in eastern Santa Catarina, South Brazil (Diptera: Culicidae). Biotemas 27, 151–154. doi: 10.5007/2175-7925.2014v27n1p151
Müller G. A., Marcondes C. B. (2007). Immature mosquitoes (Diptera: Culicidae) on the bromeliad Nidularium innocentii in ombrophilous dense forest of Santa Catarina Island, Florianópolis, Santa Catarina State, southern Brazil. Biotemas 20, 27–31.
Myers N., Mittermeier R. A., Mittermeier C. G., da Fonseca G. A. B., Kent J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Nascimento-Pereira A. C., Neves M. S. A. S., Guimares A. R., Motta M. D. A., Loureno-de-Oliveira R. (2021). Wyeomyia shannoni Lane amp; Cerqueira, 1942, a taxonomic puzzle (Diptera: Culicidae): synonymy, genus transfer, homonymy, and description of a new species of Sabethes Robineau-Desvoidy, 1827. Zootaxa 5082, 259–277. doi: 10.11646/zootaxa.5082.3.4
Olano V., Carrasquilla G., Mendez F. (1997). Transmisión de la malaria urbana en Buenaventura, Colombia: aspectos entomológicos. Rev. Panamerica Salud Publica 1, 287–294. doi: 10.1590/S1020-49891997000400005
Ortega-Morales A. I., Moreno-García M., González-Acosta C., Correa-Morales F. (2018). Mosquito Surveillance in Mexico: The Use of Ovitraps for Aedes aEgypti, Ae. albopictus, and Non-Target Species. Florida Entomologist 101, 623–626. doi: 10.1653/024.101.0425
Ramos G. J. P., do Nascimento Moura C. W. (2019). Algae and cyanobacteria in phytotelmata: diversity, ecological aspects, and conservation. Biodiversity Conserv. 28, 1667–1697. doi: 10.1007/s10531-019-01771-2
Reis M., Müller G. A., Marcondes C. B. (2010). Inventário de mosquitos (Diptera: Culicidae) da Unidade de Conservação Ambiental Desterro, Ilha de Santa Catarina, Sul do Brasil. Biota Neotropica 10, 333–337. doi: 10.1590/S1676-06032010000300031
Roberts D. R., Hsi B. P. (1979). An index of species abundance for use with mosquito surveillance data. Environm. Entomol. 8, 1007–1013.
Shannon C. E. (1948). A mathematical theory of communication. Bell System Tech. J. 27, 379–423. doi: 10.1002/j.1538-7305.1948.tb01338.x
Silva A. M. da, Nunes V., Lopes J. (2004). Culicídeos associados a entrenós de bambu e bromélias, com ênfase em Aedes (Stegomyia) albopictus (Diptera, Culicidae) na Mata Atlântica, Paraná, Brasil. Iheringia Série Zoologia 94, 63–66. doi: 10.1590/S0073-47212004000100011
Silva M. B. da, Ribeiro M. J. R., Costa P. T. S. (2007). Comunidade planctônica presente no fitotelmata de bromélias (bromeliaceae) em ambiente urbano de feira de Santana, Bahia, Brasil. Sitientibus Série Ciências Biológicas 7, 125–127. doi: 10.13102/scb8138
Stahlhöfer B. D., Wegener M. K., Muller G. A. (2022). Mosquitoes (diptera: culicidae) in bamboo internodes in the northwestern region of the state of Rio Grande do Sul, Brazil. J. Trop. Pathol. 51, 165–170. doi: 10.5216/rpt.v51i2.70392
Sunahara T., Mogi M. (1998). Distribution and turnover of a mosquito (Tripteroides bambusa) metapopulation among bamboo groves. Ecol. Res. 13, 291–299. doi: 10.1046/j.1440-1703.1998.00265.x
Sunahara T., Mogi M. (2002). Variability of intra- and interspecific competitions of bamboo stump mosquito larvae over small and large spatial scales. Oikos 97, 87–96. doi: 10.1034/j.1600-0706.2002.970109.x
Torreias Sr da S., Ferreira-Keppler R. L., Godoy B. S., Hamada N. (2010). Mosquitoes (Diptera, Culicidae) inhabiting foliar tanks of Guzmania brasiliensis Ule (Bromeliaceae) in central Amazonia, Brazil. Rev. Bras. Entomologia 64, 618–623. doi: 10.1590/S0085-56262010000400013
Vasconcelos P. F. da C. (2003). Febre amarela. Rev. Da Sociedade Bras. Medicina Trop. 36, 275–293. doi: 10.1590/S0037-86822003000200012
Veloso H. P., Rangel Filho A. L. R., Lima J. C. A. (1991). Classificação da vegetação brasileira, adaptada a um sistema universal (Rio de Janeiro: Ministério da Economia, Fazenda e Planejamento, Fundação Instituto Brasileiro de Geografia e Estatística, Diretoria de Geociências, Departamento de Recursos Naturais e Estudos Ambientais), 123 p.
Keywords: immature stages, reproduction, culicids, behavior, biodiversity
Citation: de Almeida NM, Dias RF, Alves DCV, Machado SL, Silva JdS, de Mello CF and Alencar J (2025) Mosquito fauna (Diptera: Culicidae) in phytotelmata environments of the Guapiaçu Ecological Reserve, Cachoeiras de Macacu, Rio de Janeiro, Brazil. Front. Ecol. Evol. 13:1525202. doi: 10.3389/fevo.2025.1525202
Received: 12 November 2024; Accepted: 22 January 2025;
Published: 11 February 2025.
Edited by:
Matteo Marcantonio, Université catholique de Louvain, BelgiumReviewed by:
Enrico Alejandro Ruiz, National Polytechnic Institute, MexicoDaniela De Angeli Dutra, Stanford University, United States
Copyright © 2025 de Almeida, Dias, Alves, Machado, Silva, de Mello and Alencar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jeronimo Alencar, amFsZW5jYXJAaW9jLmZpb2NydXouYnI=