 Florencia Paola Coronel1
Florencia Paola Coronel1 Gisela Loretán1,2,3Sebastián Sanchez2,4Natalia Silva2,4Gualberto Pacheco-Sierra5Florencia Brancolini2,6Danilo Aichino7Facundo Vargas8Pablo Scarabotti2,3Luis Espínola2,3
Gisela Loretán1,2,3Sebastián Sanchez2,4Natalia Silva2,4Gualberto Pacheco-Sierra5Florencia Brancolini2,6Danilo Aichino7Facundo Vargas8Pablo Scarabotti2,3Luis Espínola2,3 Carla Simone Pavanelli9
Carla Simone Pavanelli9 Eva Carolina Rueda1,2*
Eva Carolina Rueda1,2*- 1Laboratorio de Genética, Facultad de Humanidades y Ciencias, Universidad Nacional del Litoral (FHUC-UNL), Santa Fe, Argentina
- 2Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET), Buenos Aires, Argentina
- 3Instituto Nacional de Limnología (INALI), Santa Fe, Argentina
- 4Instituto de Ictiología del Nordeste, Facultad de Ciencias Veterinarias (UNNE), Corrientes, Argentina
- 5Unidad de Biología de la Conservación, PCTY, UMDI Sisal – Facultad de Ciencias, Universidad Nacional Autónoma de México, Mexico, Mexico
- 6Instituto de Investigación e Ingeniería Ambiental, Universidad Nacional de San Martín (UNSAM), Buenos Aires, Argentina
- 7Proyecto de Biología Pesquera regional, Instituto de Biología Subtropical CONICET-Universidad Nacional de Misiones (UNAM), Misiones, Argentina
- 8Dirección de Fauna y Áreas Naturales Protegidas, Resistencia, Chaco, Argentina
- 9Ictiologia e Aquicultura, Núcleo de Pesquisas em Limnologia, Maringá, Universidade Estaduale de Maringá, Maringá, Brazil
Introduction: Megaleporinus obtusidens, also known as "boga," is a freshwater teleost fish species widely distributed across South America with significant commercial value in Argentina and Brazil. Fishers, particularly those in the lower Paraná River, frequently classify the fish they catch as M. obtusidens. Despite various approaches, including cytogenetics, molecular data, and morphological analysis, the taxonomic classification of the boga fish remains debated among researchers, resulting in discrepancies in the literature.
Methods: This study aimed to assess the diversity of specimens captured in the La Plata River Basin, initially classified as Megaleporinus obtusidens by fishers, using DNA barcoding. The mitochondrial COI marker was employed to explore species assignment and genetic diversity in Megaleporinus, a genus with high commercial exploitation in the southern area of the La Plata Basin.
Results: Our findings indicate that some boga samples, previously identified as M. obtusidens, are actually M. piavussu. We also observed significant differences in the geographic distribution, diversity, and genetic structure between the two species.
Discussion: These results highlight the importance of studying the ecology of each species separately. Proper resource management, based on accurate species identification, is critical for the conservation of ichthyofauna in the region.
Introduction
The family Anostomidae (Characiformes) is a group of freshwater fishes distributed throughout the Neotropical region in the La Plata basin, which includes the Paraná, Paraguay and Uruguay rivers (Agostinho and Zalewski, 1995; Cussac et al., 2009; Dagosta and de Pinna, 2017; Almeida et al., 2021; Scarabotti et al., 2021). The genus Megaleporinus is widespread in South America and is the most diverse in this family. Ramirez et al. (2017b) deeply revised the genus with complementary methods and identified 10 nominal species based on morphological analyses. However, the strongly supported phylogenetic analyses carried out introduced 16 lineages distributed among these 10 valid species. The authors found high genetic divergences among basins within four of the nominal species, including Leporinus obtusidens (Gery, 1977; Martins et al., 2003; Ramirez et al., 2017b). The genus Megaleporinus includes species that are sometimes difficult to classify; M. obtusidens (Valenciennes, 1837) shows significant morphological similarity to M. piavussu (Britski et al., 2012), which was considered endemic to Upper Paraná in Brazil until 2012 (Britski et al., 2012).
The floodplain of the Paraná River presents regular annual flood pulses, which play a crucial role in maintaining its rich fish biodiversity and supporting various artisanal and sport fisheries (Rabuffetti et al., 2020). In Argentina, the Paraná-Plata river basin harbors 580 fish species, being one of the most biologically diverse areas in the country (Maiztegui et al., 2022; Mirande and Koerber, 2020; Mirande and Koerber, 2015; Abell et al., 2008). The fish resources of the basin, especially species with migratory behavior, are an important source of protein for local riverine communities and provide essential support for artisanal fisheries (Liotta, 2020; Arrieta et al., 2023). All commercial catches of this genus in the Argentine stretch of the Paraná River are classified as a single species, Megaleporinus obtusidens (boga), which represents the third most important taxon in terms of fish exports in Argentina (Martins et al., 2003; Iwaszkiw and Firpo Lacoste, 2011; Scarabotti et al., 2021; Arrieta et al., 2023).
Genetic techniques based on mitochondrial DNA barcoding analysis have been used to reveal distinct genetic lineages corresponding to specific geographic distributions in many freshwater migratory fish species that co-occur with boga, such as Salminus brasiliensis (golden dorado) and Prochilodus lineatus (sábalo or curimbatá) (Cardoso et al., 2018, 2021; Melo et al., 2018; Rosso et al., 2018). The effectiveness of these techniques has been consistently demonstrated through the identification and resolution of numerous taxonomic issues in both marine and freshwater fish species. DNA barcoding has been particularly successful in differentiating species within closely related taxa, with a success rate of over 90% (Rubinoff and Holland, 2005; Lara et al., 2010; de Carvalho et al., 2011; Pereira et al., 2013; Rossini et al., 2016; Berbel-Filho et al., 2018; Cardoso et al., 2018; Nascimento et al., 2023). Previous molecular studies on the genus Megaleporinus have suggested that the current taxonomy may be obscuring isolated populations with significant intraspecific genetic divergence. These results underscore the need for further molecular studies to enhance our understanding of the taxonomy and population characteristics of these species (Avelino et al., 2015; Díaz et al., 2016; Ramirez et al., 2016; Pires et al., 2017; Ramirez et al., 2017b).
Given the economic and sporting significance of the species, our study aimed to identify the species of the genus Megaleporinus based on specimens caught by fishers. Fishers usually refer to them as “boga” and classify them as M. obtusidens based on morphological traits. To this end, molecular techniques based on DNA barcoding were employed to gain insights into the genetic diversity and population structure of these fish species. The potential presence of two species suggests the need to manage them separately.
Results
The 88 boga fish specimens were preliminary analyzed in the field (sampling sites are indicated in Figure 1; Table 1) using classical taxonomy. All the samples were classified as M. obtusidens (specimens of M. obtusidens and M. piavussu are shown in Figure 2). A total of 593-base pair fragments of the mitochondrial cytochrome c oxidase I (COI) gene from those 88 new boga samples were obtained. Additionally, 28 sequences were retrieved from the GenBank database, as detailed in the Supplementary Table 1.
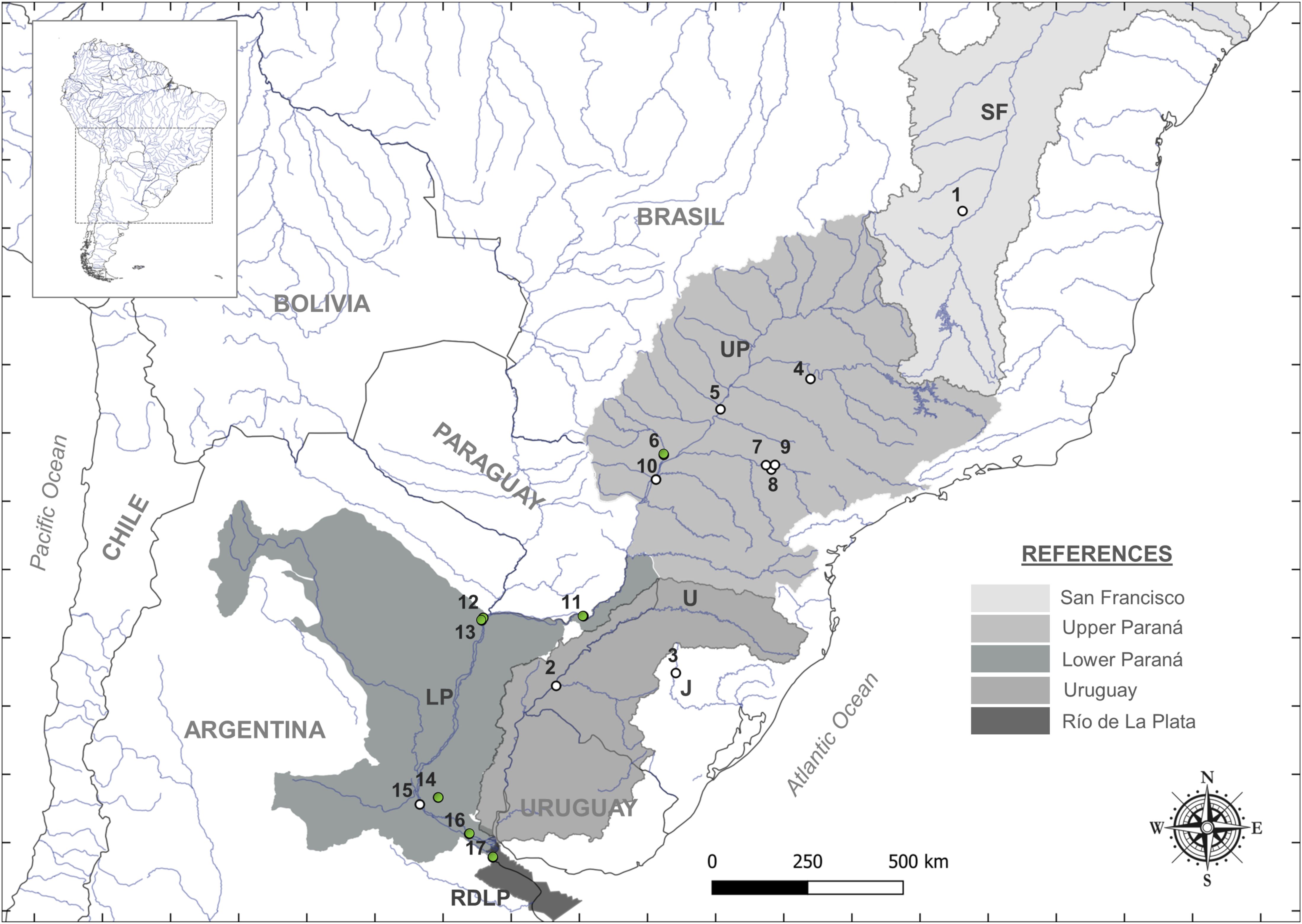
Figure 1. Map of the study area showing the locations and basins of the sampling sites. Rivers are indicated as follows: SF, San Fernando; U, Uruguay; J, Jacuí; UP, Upper Paraná; LoP, Lower Paraná; LP, La Plata. Sampling sites [1–17] correspond to those listed in Table 1. The grey shading represents the different basins sampled.
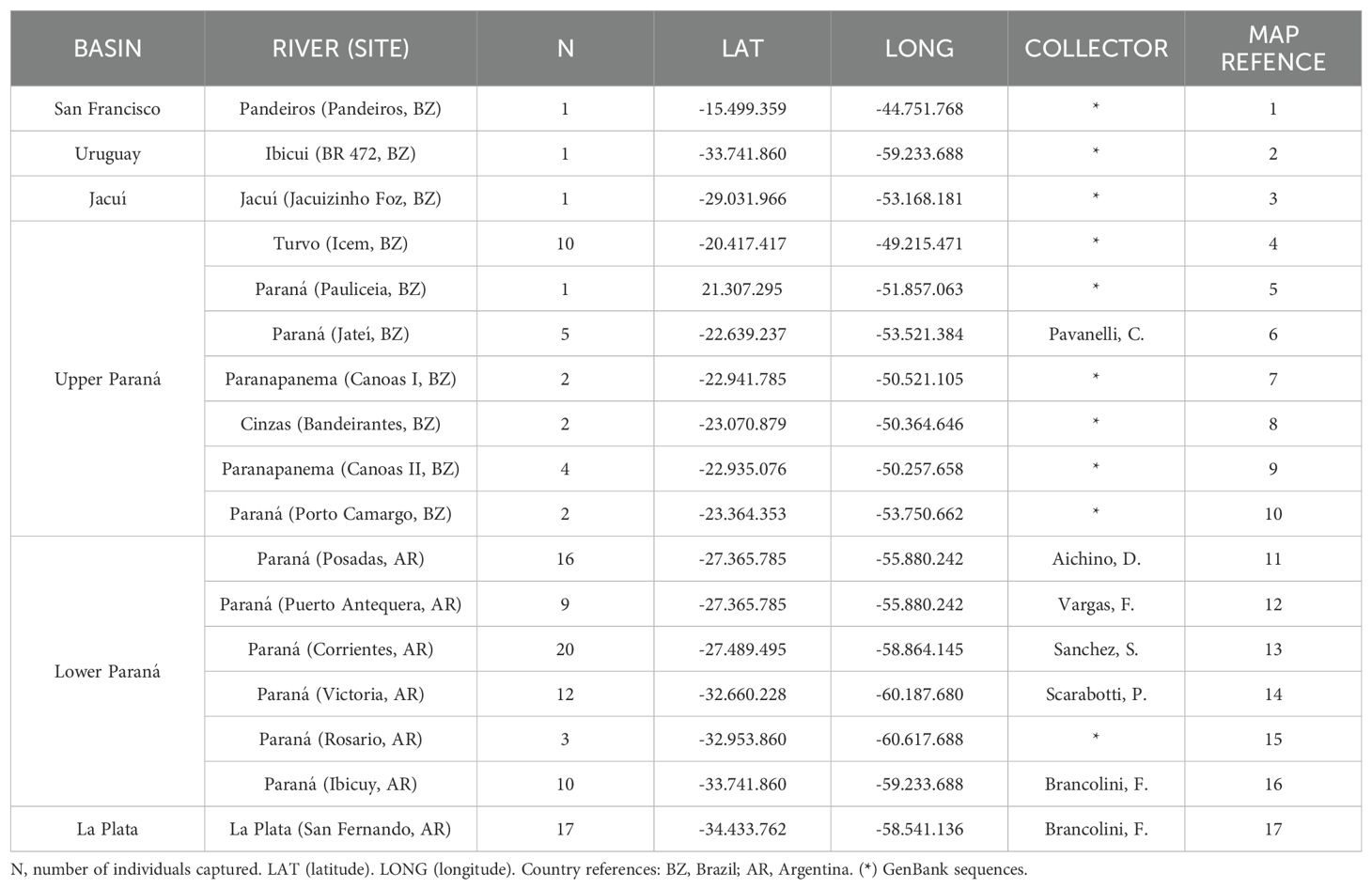
Table 1. Description and map code of sampling sites.

Figure 2. Representative images of the two study species: M. obtusidens (top) and M. piavussu (bottom). Photographs were taken prior to sampling in Corrientes, Argentina (map reference: site 13).
A total of 82 polymorphic sites were identified among the 116 sequences, which allowed for the definition of 16 haplotypes (Table 2). The newly obtained sequences were deposited in GenBank (Accession Numbers PQ586696 - PQ586780, PQ591847-PQ591849; Supplementary Table 1).

Table 2. Diversity indexes of M. obtusidens and M. piavussu.
COI isolates obtained from the samples were analyzed individually with the BLAST algorithm. 59 of the new isolates. They showed 95%-99.8% range coincidence with the M. piavussu COI sequences available in the GenBank database. The results from the phylogenetic rooted tree (Figure 3; Supplementary Figures A, B) clearly distinguished two clades corresponding to the two species.
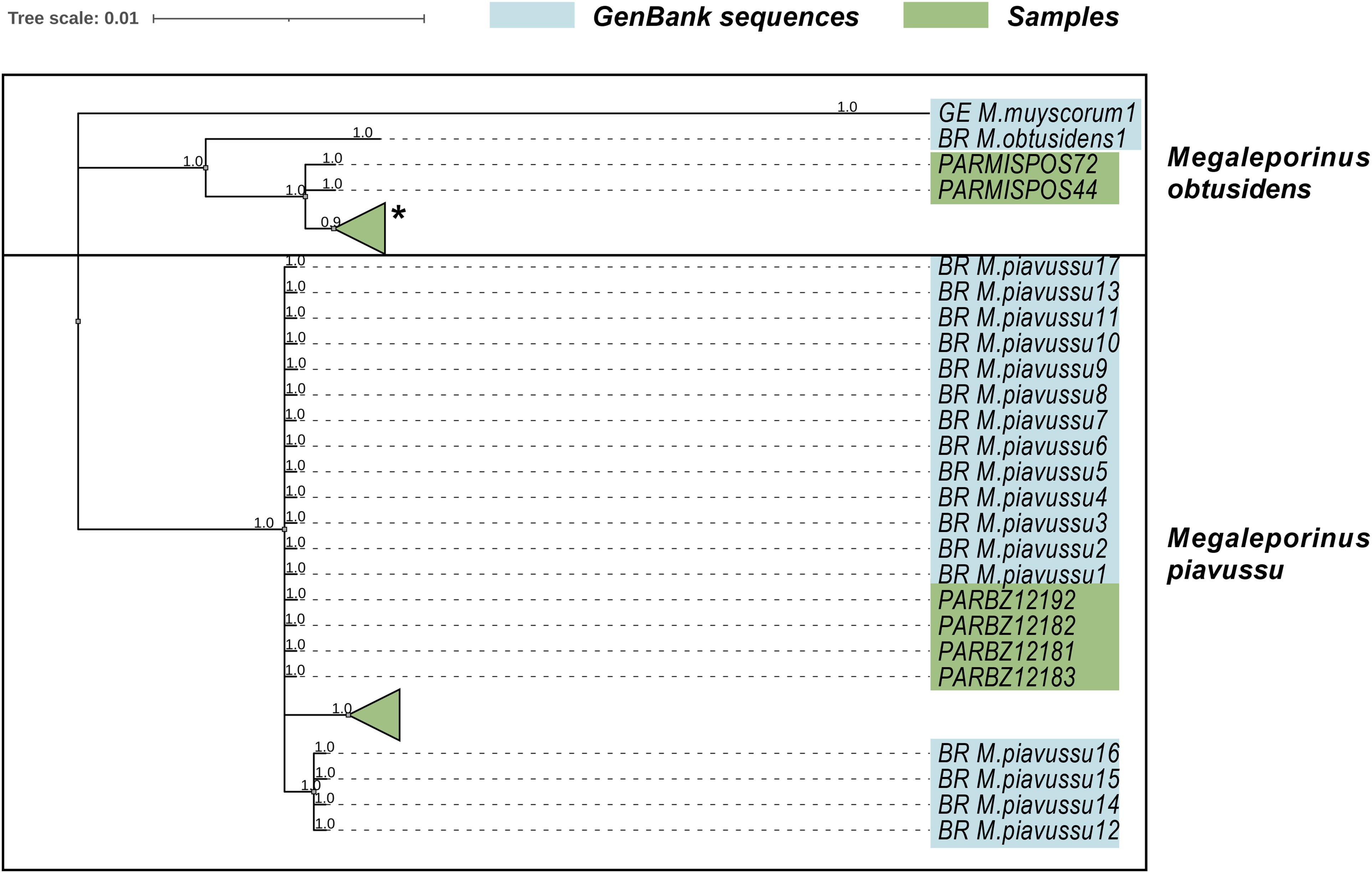
Figure 3. Collapsed Bayesian tree showing the distribution of M. obtusidens and M. piavussu sequences. The full extended trees are available in SI (Supplementary Figures A, B). The (*) symbol indicates that the collapsed tree includes sequences from both GenBank and the sampling conducted for this study.
The haplotype network (Supplementary Figure C) showed that haplotypes Hap2 and Hap3 had the highest frequencies and that haplotypes Hap3 and Hap14 were shared between M. obtusidens and M. piavussu. The greatest haplotype diversity was observed in the lower Paraná, and most haplotypes are unique to M. piavussu. The haplotype network illustrates the presence of these two primary haplogroups, which are formed by different basins and the preponderance of the identified haplotypes was derived from the lower Paraná basin.
The M. obtusidens populations exhibited no evidence of population structure along the Paraná River. In contrast, the M. piavussu lineages are distinguished into two groups according to the geographic location. Clusters were identified for the Upper Paraná and Lower Paraná-Río de la Plata regions (Figure 3). These findings suggested the potential existence of population genetic structure within this species, which was further supported by the results of the hierarchical AMOVA test (Supplementary Table 2). The AMOVA statistics indicate that most of the variance observed in M. piavussu comes from the among-group component (basin). We confirmed this unequal lineage distribution by examining the haplotype distribution; we observed that M. obtusidens exhibits a uniform haplotype distribution along the entire length of the Paraná River. In contrast, genetic structure is apparent in M. piavussu, delineated by the geographic distribution of the lineages (Figure 4).
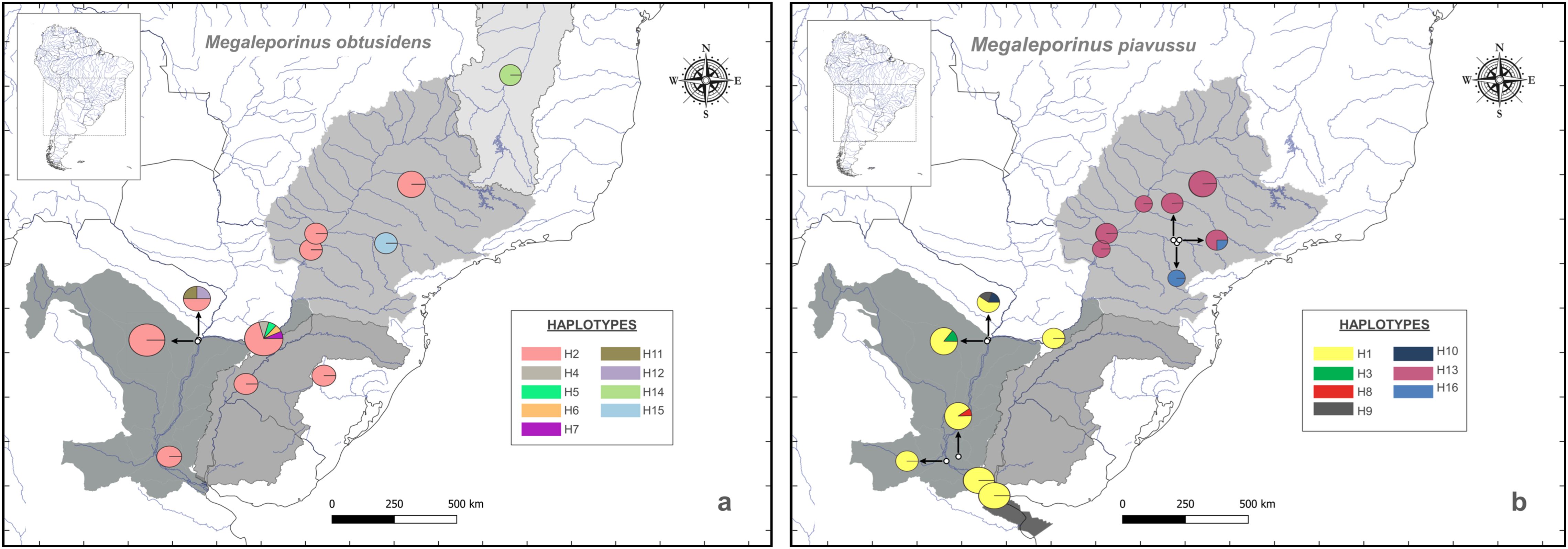
Figure 4. Haplotype distribution in the Paraná River basin. Circle size represents the frequency of each haplotype at the respective capture sites. (A, B) illustrate the haplotypes of M. obtusidens and M. piavussu, respectively. The haplotype network based on COI sequences is provided in Supplementary Figure C.
In the Río de la Plata River and in the lower section of the Paraná River (corresponding to the Ibicuy site), only specimens of M. piavussu were identified with COI. It should be noted that fishers had identified them as M. obtusidens through morphological assessments.
Discussion
The use of molecular methods for the identification and description of species of the Anostomidae family was previously reported in the literature (Chiari and Maria Koelblinger Sodré, 2001; Ferreira et al., 2017; Silva-Santos et al., 2018; Utsunomia et al., 2019). DNA barcoding is one of the most widely used strategies for species identification within the genus Megaleporinus (Avelino et al., 2015; Ramirez et al., 2016, 2017a, 2017b; Nascimento et al., 2023). Several barcoding studies have been conducted on other freshwater neotropical fish around the world, resulting in new insights into the field of taxonomy and conservation biodiversity (Ribolli et al., 2012; Rosso et al., 2018; Cardoso et al., 2018; Melo et al., 2018; Oliveira Carvalho et al., 2024; Zafar et al., 2024).
The present study was designed to identify Megaleporinus species that occur in the Paraná River based on specimens initially identified as M. obtusidens through morphological characterization. The findings of this study demonstrate that the examined specimens belong to two discrete species: M. piavussu and M. obtusidens. Furthermore, most of the specimens were subsequently identified as M. piavussu through DNA barcoding. These results are consistent with those of previous analyses, which highlighted the significant challenge in identifying Megaleporinus species, due to their striking morphological similarities (Ramirez et al., 2016, 2017a, 2017b). These authors postulate that the most effective method for differentiating M. obtusidens from M. piavussu is through the examination of two crucial characteristics: number of perforated scales in the lateral line and position of the mouth. However, these features are not easily distinguished in the field, hindering the accurate identification of these species. Misclassification leads to inadequate legislation and ineffective policies for the conservation of freshwater fish (Ahmed et al., 2022).
According to Abrial et al. (2019, 2021), the hydrological attributes of the Paraná River, such as frequency, timing, duration and intensity, have undergone significant changes since the 1970s. These alterations have led to substantial changes in surface hydrological connectivity patterns. These findings suggest that the captured fishes belong to different species, and that the current legislative framework governing the exploitation of boga does not recognize this diversity. The Argentine legislation does not regulate fishing activities or the amount allowed for export of M. piavussu; in addition, there is also a knowledge gap about the species that is currently fished and exported (Filippo and Alvarez, 2008; Deinet et al., 2020; Baigún and Minotti, 2021).
In conclusion, the misidentification of boga species in the Paraná River has led to the exploitation of a commercial fishery resource that remains incompletely understood. To guarantee the conservation of these species, it is imperative that research efforts are increased and improved.
Two species, two stories
The broad distribution observed in the Megaleporinus species reflects the extensive distances covered by other migratory species, such as P. lineatus and S. brasiliensis, which play a key economic role in Argentina (Iwaszkiw and Firpo Lacoste, 2011; Deinet et al., 2020; Baigún and Minotti, 2021). The results of the analysis of genetic structure based on COI marker demonstrate that M. piavussu, exhibits two distinct clades clearly differentiated between the Upper and Lower Paraná River populations (Figure 4; Table 2; Supplementary Figure C). This result lends support to the hypothesis formulated by Ramirez et al. (2017) postulating that there are discrete genetic lineages within the M. piavussu species, as observed in the present study. The first record of M. piavussu in the Upper Paraná River was provided by Britski et al. (2012) and subsequently documented in the Lower Paraná (Benitez and Aichino, 2020). However, a review of the literature revealed cases of misclassification of M. piavussu as M. obtusidens (Avelino et al., 2015; Díaz et al., 2016). Therefore, it can be speculated that M. piavussu may have been present in the Lower Paraná River before 2012. Similar findings were documented in other migratory fish species. Studies conducted on S. brasiliensis indicated the existence of discrete clusters that correspond to the Upper and Lower Paraná distribution regions of the species (García‐Machado et al., 2022; Rosso et al., 2018). This suggests the existence of genetic differentiation and the presence of discrete population groups (genetic clusters).
Our results suggest that M. piavussu displays genetic structure, which may be attributed to the challenges faced when attempting to cross both artificial and natural geographic barriers. These barriers constrain the upward and downward movement of the ichthyofauna and can function as environmental filters that exert selective influence on the functional characteristics of the fish fauna. Indeed, the limited dispersal ability of M. piavussu may contribute to the differentiation observed among populations, as in S. brasiliensis (Da Silva et al., 2015; Casimiro et al., 2017; Chanchay Castro, 2019). This assumption is supported by a previous study, which evaluated the effectiveness of a fish ladder in the Porto Primavera Dam in the Upper Paraná River (Gutfreund et al., 2018). Other findings indicated that only three of the 116 M. piavussu specimens were able to successfully pass across the dam. These results show the difficulties encountered by this species in surmounting physical obstacles such as dams, hindering the dispersal and gene flow among disparate populations (Pope et al., 2018).
Furthermore, fish species that have morphological characteristics that allow them to undertake long-distance migrations (de Assumpção et al., 2012) may exhibit shorter migrations when suitable spawning and feeding areas are accessible in downstream sections (Oliveira et al., 2018). The proximity of favorable habitats can significantly influence the migratory behavior of fishes, leading to adaptations in their migration patterns. These characteristics may have contributed to the differentiation observed between individuals occurring in the Upper Paraná River and those occurring in the Lower Paraná River, as previously reported (Ramirez et al., 2017). Nevertheless, further studies are required to gain a comprehensive understanding of the evolutionary processes that could explain these differentiations.
The results obtained in this study make two important contributions: (1) The specimens caught in the Paraná River and identified as “boga” could actually be M. obtusidens or M. piavussu; (2) Analyses of genetic diversity and population structure using COI show differences between species, suggesting that M. obtusidens and M. piavussu may have had different evolutionary histories.
In conclusion, we present valuable information for the conservation of biodiversity and the management of boga fisheries in Argentina and Brazil. To ensure effective regulation and sustainable exploitation of this resource, morphological, ecological, and evolutionary analyses should be performed to accurately identify species, understand their ecological roles, and assess their evolutionary history. However, most importantly, it is essential to ascertain the extent and intricacy of the various migratory species regarding their distribution, dispersion, and evolutionary diversification. Their histories converge in a singular system, namely the Paraná River.
Materials and methods
Study area
The Paraná River basin is approximately 4,000 km long and is the third most important in South America in terms of flow and drained area (Bonetto et al., 1986; Latrubesse, 2008; Pereira et al., 2013; Rosso et al., 2018). It is composed of two distinct ichthyo-faunistic provinces (ecoregions corresponding to the Upper and Lower Paraná River), which were connected when the Itaipú reservoir was built. The Upper Paraná River is in the Brazilian territory and extends from the confluence of the Paranaíba and Grande rivers to Sete Quedas Falls. Before the construction of the reservoir, the falls formed a natural barrier dividing the ecoregions (Da Graça and Pavanelli, 2007; Sivasundar et al., 2001). The Lower Paraná River flows downstream the falls, forming an extensive floodplain. The confluence of the Lower Paraná and the Uruguay rivers forms the estuary known as Río de La Plata (Agostinho and Zalewski, 1995; Sverlij et al., 2013). Figure 1 shows the location of the study area.
Fish and tissue sampling
We collected fish samples from Paraná and Río de La Plata rivers; then we analyzed and compared them with sequences from the GenBank database corresponding to the San Francisco, Paraná, and Uruguay rivers. Samples were collected from seven sites along the Paraná River (Figure 1; Table 1), corresponding to the locations of Jateí (Mato Grosso do Sul) in Brazil, and Posadas (Misiones), Puerto Antequera (Chaco), Corrientes (Corrientes), Victoria (Entre Ríos), Ibicuy (Entre Ríos), and San Fernando (Buenos Aires) in Argentina from monitoring or research activities from local government. Occasionally, tissues were collected with the assistance of fisherman. The collected specimens were anesthetized by immersion in 1% benzocaine in water and euthanized by benzocaine excess. A portion of muscle (0.5 mm x 0.5 cm) was carefully stored in 1.5 mL microtubes and preserved in 95% ethanol for further laboratory processing. Initially, the specimens were identified as M. obtusidens. All the sampling process was performed with the appropriate scientific fishing license and covered by provincial and national authorities; and complied with Law 12212 of Santa Fe Province, Law 5628 and Decree 422/2010, Art. 35, for Chaco Province, and with the Authorization of the Directorate of Natural Resources for the Corrientes survey. Furthermore, the genetic analyses incorporated sequences specifically corresponding to the San Francisco, Paraná, and Uruguay rivers obtained from the GenBank database.
DNA extraction, amplification and sequencing
Genomic DNA extraction was performed in the Genetics Laboratory (FHUC‐UNL, Santa Fe, Argentina) using the salt-extraction protocol (Aljanabi and Martinez, 1997). A fragment amplification of the mitochondrial cytochrome c oxidase subunit I gene (COI) was performed using the polymerase chain reaction (PCR) by standardized barcoding protocols (Hebert et al., 2004; Ivanova et al., 2007; Ratnasingham and Hebert, 2007). We selected the following primers: forward (F) FishF2_t1 5’-TGTAAAACGACGGCCAGTCGACTAATCATAAAGATATCGGCAC-3’ and reverse (R) FishR2_t1 5’-CAGGAAACAGCTATGACACTTCAGGGTGACCGAAGAATCAGAA-3’.
PCRs were performed using DNA (50-100 ng), reaction buffer (1X), MgCl2 (2 mM), dNTP (40 µM), forward and reverse primers (0.2 mM), Taq polymerase (0.75 U), and demineralized water in a final volume of 25 μL. The amplification conditions included an initial step of 4 min at 95°C, and 40 cycles of 1 min at 94°C, 1 min at 40°C, and 1 min at 72°C, with two final extensions: first at 72°C for 10 min and then at 60°C for 20 min. The amplicons of COI were sequenced by Macrogen Inc. (www.macrogen.com) using Sanger sequencing. The isolates were uploaded to Genbank with AN.
Molecular data analysis
First, using the default algorithm implemented in BLAST (Basic local alignment search tool, https://blast.ncbi.nlm.nih.gov/Blast.cgi), we compared the sequenced amplicons with the information present in GenBank database to test if they matched Megaleporinus sequences. These preliminary check of species is important because the genus Megaleporinus has been revised and the morphological characters may not be enough for classification (Ramirez et al., 2017). Then we compared our sequences with the ones of Megaleporinus retrieved from GenBank (Ramirez et al., 2017a, 2017). We included them in the subsequent analyses. All DNA sequences were combined into a single alignment to perform bioinformatic analyses. The alignment was carried out using MAFFT v.7 software (Katoh and Standley, 2013). The best-fit model of nucleotide substitution was estimated using jModelTest v.2.1.7 software (Posada, 2008) with default parameters. The HKY+I model was chosen for subsequent phylogenetic inference. Phylogenetic tree construction and final consensus tree edition were performed using the software packages MrBayes v.3.2.7 (Ronquist et al., 2012), FigTree v.1.3.1 (Rambaut, 2009), and iTOL (Letunic and Bork, 2021). MrBayes analyses were carried out with 1,000,000 initial generations and 3,000,000 subsequent generations of Markov chains and a bootstrap value of 1000.
Genetic diversity was assessed using the haplotype number, nucleotide diversity (π), and haplotype diversity (Hd) estimators in DnaSP v.5 (Librado and Rozas, 2009). These statistics allow for the description of haplotype distribution under an infinite site model. Median-Joining algorithm was implemented in haplotype networks built with NETWORK v 10.2.0.0 software (Bandelt et al., 1999; Rohl, 2000). PopArt software (Leigh et al., 2015) and PhyloGeoViz v.2.4.5 (Tsai, 2011) was used to graph and analyze the haplotype network including spatial distribution. Maps were constructed using QGIS software v.3.4.11 (Moyroud and Portet, 2018). Finally, to measure the genetic differentiation among sub-basins, we used hierarchical AMOVA test and FST index, calculated by Arlequin v. 3.5.1.2 (Excoffier and Lischer, 2010) for each species.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because we obtained the samples from the local government from the different provinces of Argentina, which conducted a series of monitoring and research activities. Brazil samples were provided from Nupeliá collection.
Author contributions
FC: Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. GL: Formal analysis, Investigation, Methodology, Writing – review & editing, Data curation, Writing – original draft. SS: Funding acquisition, Methodology, Resources, Project administration, Writing – review & editing. NS: Data curation, Resources, Methodology, Writing – review & editing. GP-S: Conceptualization, Methodology, Writing – review & editing. FB: Writing – review & editing, Data curation. DA: Methodology, Writing – review & editing. FV: Methodology, Writing – review & editing. PS: Data curation, Methodology, Writing – review & editing. LE: Conceptualization, Writing – review & editing. CP: Data curation, Writing – review & editing. ER: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was financially supported by project PIP 2014 (113). We thank Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET). Entidad Binacional Yacyretá. Convenio EBY-UNNE. Acta 27 also supported this research.
Acknowledgments
We thank PhD Guillermo Orti for his valuable contribution to this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2024.1519907/full#supplementary-material
References
Abell R., Thieme M. L., Revenga C., Bryer M., Kottelat M., Bogutskaya N., et al. (2008). Freshwater ecoregions of the world: a new map of biogeographic units for freshwater biodiversity conservation. BioScience 58, 403–414. doi: 10.1641/B580507
Abrial E., Espínola L. A., Rabuffetti A. P., Amsler M. L., Wantzen K. M. (2019). Interannual flow variability in a large subtropical–temperate floodplain: A challenge for fish reproduction. Can. J. Fish. Aquat. Sci. 76, 390–400. doi: 10.1139/CJFAS-2017-0579
Abrial E., Lorenzón R. E., Rabuffetti A. P., Blettler M. C., Espínola L. A. (2021). Hydroecological implication of long-term flow variations in the middle Paraná river floodplain. J. Hydrol. 603, 126957. doi: 10.1016/j.jhydrol.2021.126957
Agostinho A. A., Zalewski M. (1995). “The dependence of fish community structure and dynamics on floodplain and riparian ecotone zone in Parana River, Brazil,” in The Importance of Aquatic-Terrestrial Ecotones for Freshwater Fish. Developments in Hydrobiology, vol 105, eds. F. Schiemer, M. Zalewski, J. E. Thorpe. (Dordrecht: Springer). doi: 10.1007/978-94-017-3360-1_13
Ahmed S. F., Kumar P. S., Kabir M., Zuhara F. T., Mehjabin A., Tasannum N., et al. (2022). Threats, challenges and sustainable conservation strategies for freshwater biodiversity. Environ. Res. 214, 113808. doi: 10.1016/j.envres.2022.113808
Aljanabi S. M., Martinez I. (1997). Universal and rapid salt-extraction of high-quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 25, 4692–4693. doi: 10.1093/nar/25.22.4692
Almeida M. S., Birindelli J. L. O., Assega F. M., Barros M. C., Fraga E. C. (2021). New records of the occurrence of Megaleporinus macrocephalus (Characiformes, Anostomidae) from the basins of the Itapecuru and Mearim rivers in Maranhão, Northeastern Brazil. Braz. J. Biol. 82, e232868. doi: 10.1590/1519-6984.232868
Arrieta P., Balboni L., Fuchs D., Liotta J., Picotti G., Somoza D., et al. (2023). Proyecto “Evaluación Biológica y Pesquera de especies de interés deportivo y comercial en la Cuenca del Río de la Plata en Argentina”. Available online at: https://www.magyp.gob.ar/sitio/areas/pesca_continental/informes/baja/index.ph (Accessed December 16, 2024).
Avelino G. S., Britski H. A., Foresti F., Oliveira C. (2015). Molecular identification of Leporinus from the south portion of South America. DNA Barcodes 3, 98–109. doi: 10.1515/dna-2015-0013
Baigún C. R. M., Minotti P. G. (2021). Conserving the Paraguay-Paraná fluvial corridor in the XXI century: conflicts, threats, and challenges. Sustainability 13, 5198. doi: 10.3390/su13095198
Bandelt H.-J., Forster P., Röhl A. (1999). Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 16, 37–48. doi: 10.1093/oxfordjournals.molbev.a026036
Benitez M. F., Aichino D. R. (2020). First record of Megaleporinus piavussu (Characiformes: Anostomidae) from Argentina. Ichthyol. contrib. Peces Criollos. 64, 1–4. Available at: http://hdl.handle.net/11336/114685 (Accessed March 16, 2020).
Berbel-Filho W. M., Ramos T. P. A., Jacobina U. P., Maia D. J. G., Torres R. A., Lima S. M. Q. (2018). Updated checklist and DNA barcode-based species delimitations reveal taxonomic uncertainties among freshwater fishes from the mid-north-eastern Caatinga ecoregion, north-eastern Brazil. J. Fish Biol. 93, 311–323. doi: 10.1111/jfb.13758
Bonetto A. A., Neiff J. J., Di Persia D. H. (1986). The Paraná River system (Dordrecht: Springer), 541–598. doi: 10.1007/978-94-017-3290-1_11
Britski H. A., Birindelli J. L. O., Garavello J. C. (2012). A new species of Leporinus Agassiz 1829 from the upper Rio Paraná basin (Characiformes, Anostomidae) with redescription of L. elongatus Valenciennes 1850 and L. obtusidens (Valenciennes, 1837). Papeis Avulsos Zool. 52, 441–475. doi: 10.1590/S0031-10492012021700001
Cardoso Y. P., Jardim de Queiroz L., Bahechar I. A., Posadas P. E., Montoya-Burgos J. I. (2021). Multilocus phylogeny and historical biogeography of Hypostomus shed light on the processes of fish diversification in La Plata Basin. Sci. Rep. 11, 5073. doi: 10.1038/s41598-021-83464-x
Cardoso Y. P., Rosso J. J., Mabragaña E., Gonzalez-Castro M., Delpiani M., Avigliano E., et al. (2018). A continental-wide molecular approach unraveling mtDNA diversity and geographic distribution of the Neotropical genus Hoplias. PloS One 13, e0202024. doi: 10.1371/journal.pone.0202024
Casimiro A. C. R., Garcia D. A. Z., Costa A. D. A., Britton J. R., Orsi M. L. (2017). Impoundments facilitate a biological invasion: Dispersal and establishment of non-native armoured catfish Loricariichthys platymetopon (Isbrückler & Nijssen 1979) in a neotropical river. Limnologica 62, 34–37. doi: 10.1016/j.limno.2016.11.001
Chanchay Castro J. R. (2019). Estructura Genética del Dorado (Salminus brasiliensis) en el área de influencia del Itaipú Binacional. Disertación de maestría, Programa de Posgrado en Biodiversidad Neotropical, Universidad Federal de la Integración Latinoamericana (UNILA), Foz do Iguaçu. Available at: http://repositorioslatinoamericanos.uChile.cl/handle/2250/3843692 (Accessed December 16, 2024).
Chiari L., Maria Koelblinger Sodré L. (2001). Study of eight species of the Anostomidae family (Pisces, Characiformes) by RAPD analysis. Acta Sci. Biol. Sci. 23, 445–451.
Cussac V. E., Fernández D. A., Gómez S. E., López H. L. (2009). Fishes of southern South America: A story driven by temperature. Fish Physiol. Biochem. 35, 29–42. doi: 10.1007/S10695-008-9217-2
Dagosta F. C. P., de Pinna M. (2017). Biogeography of Amazonian fishes: Deconstructing river basins as biogeographic units. Neotrop. Ichthyol. 15, e170034. doi: 10.1590/1982-0224-20170034
Da Graça W. J., Pavanelli C. S. (2007). Peixes da planície de inundação do alto rio Paraná e áreas adjacentes. Maringá, Brasil: Eduem.
Da Silva P. S., Makrakis M. C., Miranda L. E., Makrakis S., Assumpção L., Paula S., et al. (2015). Importance of reservoir tributaries to spawning of migratory fish in the upper Paraná River. River Res. Appl. 31, 313–322. doi: 10.1002/rra.v31.3
de Assumpção L., Makrakis M. C., Makrakis S., Piana P. A., da Silva P. S., de Lima A. F., et al. (2012). Morphological differentiation among migratory fish species from the Paraná River basin. Biota Neotropica 12, 41–49.
de Carvalho D. C., Oliveira D. A. A., Pompeu P. S., Leal C. G., Oliveira C., Hanner R. (2011). Deep barcode divergence in Brazilian freshwater fishes: the case of the São Francisco River basin. Mitochondr. DNA 22, 80–86. doi: 10.3109/19401736.2011.588214
Deinet S., Scott-Gatty K., Rotton H., Twardek W. M., Marconi V., McRae L., et al. (2020).The living planet index (LPI) for migratory freshwater fish: Technical report. Available online at: https://research.rug.nl/files/130829826/LPI_report_2020.pdf (Accessed December 16, 2024).
Díaz J., Villanova G. V., Brancolini F., Del Pazo F., Posner V. M., Grimberg A., et al. (2016). First DNA barcode reference library for the identification of South American freshwater fish from the lower Paraná river. PloS One 11, e0157419. doi: 10.1371/journal.pone.0157419
Excoffier L., Lischer H. E. L. (2010). Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 10, 564–567. doi: 10.1111/j.1755-0998.2010.02847.x
Ferreira D. G., Souza-Shibatta L., Shibatta O. A., Sofia S. H., Carlsson J., Dias J. H. P., et al. (2017). Genetic structure and diversity of migratory freshwater fish in a fragmented Neotropical river system. Rev. Fish Biol. Fish. 27, 209–231. doi: 10.1007/s11160-016-9441-2
Filippo P. F., Alvarez M. (2008). Consultora “Revisión y análisis del marco legal regulatorio de la pesca marítima y continental”. Available online at: http://ceiba.agro.uba.ar/cgi-bin/koha/opac-detail.pl?biblionumber=17346 (Accessed December 16, 2024).
García‐Machado E., Laporte M., Normandeau E., Hernández C., Côté G., Paradis Y., et al. (2022). Fish community shifts along a strong fluvial environmental gradient revealed by eDNA metabarcoding. Environmental DNA 4, 117–134.
Gutfreund C., Makrakis S., Castro-Santos T., Celestino L. F., Dias J. H. P., Makrakis M. C. (2018). Effectiveness of a fish ladder for two Neotropical migratory species in the Paraná River. Mar. Freshw. Res. 69, 1848–1856. doi: 10.1071/MF18129
Hebert P. D. N., Penton E. H., Burns J. M., Janzen D. H., Hallwachs W. (2004). Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. United States America 101, 14812–14817. doi: 10.1073/pnas.0406166101
Ivanova N. V., Zemlak T. S., Hanner R. H., Hebert P. D. N. (2007). Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 7, 544–548. doi: 10.1111/j.1471-8286.2007.01748.x
Iwaszkiw J. M., Firpo Lacoste F. (2011). La pesca artesanal en la Cuenca del Plata (Argentina) y sus implicancias en la conservación de la biodiversidad. Rev. Del Museo Argentino Cienc. Naturales 13, 21–25.
Katoh K., Standley D. M. (2013). {MAFFT} Multiple sequence alignment software version 7: improvement in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Lara A., Ponce de León J. L., Rodriguez R., Casane D., Cote G., Bernatchez L., et al. (2010). DNA barcoding of Cuban freshwater fishes: evidence for cryptic species and taxonomic conflicts. Mol. Ecol. Resour. 10, 421–430. doi: 10.1111/j.1755-0998.2009.02785.x
Latrubesse E. M. (2008). Patterns of anabranching channels: The ultimate end-member adjustment of mega rivers. Geomorphology 101, 130–145. doi: 10.1016/j.geomorph.2008.05.035
Leigh J. W., Bryant D., Nakagawa S. (2015). POPART: full-feature software for haplotype network construction. Methods Ecol. Evol. 6, 1110–1116. doi: 10.1111/mee3.2015.6.issue-9
Letunic I., Bork P. (2021). Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 49, W293–W296. doi: 10.1093/nar/gkab301
Librado P., Rozas J. (2009). DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25, 1451–1452. doi: 10.1093/bioinformatics/btp187
Liotta J. (2020). Analysis of artisanal fishing effort in San Nicolás, Paraná River delta, Argentina. Pan-Am J. Aquat Sci. 15, 218–225.
Maiztegui T., Paracampo A. H., Liotta J., Cabanellas E., Bonetto C., Colautti D. C. (2022). Freshwater fishes of the Río de la Plata: current assemblage structure. Neotrop. Ichthyol. 20, e210159. doi: 10.1590/1982-0224-2021-0159
Martins C., Wasko A. P., Oliveira C., Foresti F. (2003). Mitochondrial DNA variation in wild populations of Leporinus elongatus from the Paraná River basin. Genet. Mol. Biol. 26, 33–38. Available at: www.sbg.org.br (Accessed December 16, 2024).
Melo B. F., Dorini B. F., Foresti F., Oliveira C. (2018). Little divergence among mitochondrial lineages of prochilodus (Teleostei, characiformes). Front. Genet. 9. doi: 10.3389/fgene.2018.00107
Mirande J. M., Koerber S. (2020). Checklist of the Freshwater Fishes of Argentina (2nd ed.). Ichthyological Contributions of Peces Criollos 72, 1–81.
Mirande J. M., Koerber S. (2015). On some species of Astyanax reported erroneously from Argentina. Ichthyological Contributions of PecesCriollos 35, 1–8.
Moyroud N., Portet F. (2018). Introduction to QGIS. QGIS and generic tools. Wiley 1, 1–17. doi: 10.1002/9781119457091.CH1
Nascimento M. H. S., Aragão D. G., Silva J. L. N., Lima R. C., Birindelli J. L. O., Fraga E. C., et al. (2023). The DNA barcode reveals cryptic diversity and a new record for the genus Leporinus (Characiformes, Anostomidae) in the hydrographic basins of central northern Brazil. PeerJ 11, e15184. doi: 10.7717/peerj.15184
Oliveira A. G., Baumgartner M. T., Gomes L. C., Dias R. M., Agostinho A. A. (2018). Long-term effects of flow regulation by dams simplify fish functional diversity. Freshw. Biol. 63, 293–305. doi: 10.1111/fwb.2018.63.issue-3
Oliveira Carvalho C., Pazirgiannidi M., Ravelomanana T., Andriambelomanana F., Schrøder-Nielsen A., Stuart Ready J., et al. (2024). Multi-method survey rediscovers critically endangered species and strengthens Madagascar’s freshwater fish conservation. Sci. Rep. 14, 20427. doi: 10.1038/s41598-024-71398-z
Pereira L. H. G., Hanner R., Foresti F., Oliveira C. (2013). Can DNA barcoding accurately discriminate megadiverse Neotropical freshwater fish fauna? BMC Genet. 14, 1–14.
Pires A. A., Ramirez J. L., Galetti P. M., Troy W. P., Freitas P. D. (2017). Molecular analysis reveals hidden diversity in Zungaro (Siluriformes: Pimelodidade): a genus of giant South American catfish. Genetica 145, 335–340. doi: 10.1007/s10709-017-9968-8
Pope E., Baumgartner L. J., Boys C., Gilligan D. M., Silva L., Pflugrath B. D., et al. (2018). “A perspective on the need for assessing the fish transfer risk associated with pumped hydropower schemes,” in 2018 International Conference on River Connectivity incorporating the First International Symposium on Hydropower and Fish Management: Fish Passage 2018.
Posada D. (2008). jModelTest: phylogenetic model averaging. Mol. Biol. Evol. 25, 1253–1256. doi: 10.1093/molbev/msn083
Rabuffetti A. P., Espínola L. A., Abrial E., Amsler M. L., Blettler M. C., Eurich M. F., et al. (2020). Commercial fisheries in a mega unregulated floodplain river: Assessment of the most favourable hydrological conditions for its preservation. J. Fish Biol. 96, 59–73. doi: 10.1111/jfb.14184
Rambaut A. (2009). FigTree. Tree figure drawing tool. Available online at: http://Tree.Bio.Ed.Ac.Uk/Software/Figtree/ (Accessed December 16, 2024).
Ramirez J. L., Birindelli J. L., Carvalho D. C., Affonso P. R. A. M., Venere P. C., Ortega H., et al. (2017a). Revealing hidden diversity of the underestimated neotropical ichthyofauna: DNA barcoding in the recently described genus Megaleporinus (Characiformes: Anostomidae). Front. Genet. 8, 149. doi: 10.3389/fgene.2017.00149
Ramirez J. L., Birindelli J. L. O., Galetti P. M. (2017b). A new genus of Anostomidae (Ostariophysi: Characiformes): Diversity, phylogeny and biogeography based on cytogenetic, molecular and morphological data. Mol. Phylogenet. Evol. 107, 308–323. doi: 10.1016/j.ympev.2016.11.012
Ramirez J. L., Carvalho-Costa L. F., Venere P. C., Carvalho D. C., Troy W. P., Galetti P. M. Jr. (2016). Testing monophyly of the freshwater fish Leporinus (Characiformes, Anostomidae) through molecular analysis. J. Fish Biol. 88, 1204–1214. doi: 10.1111/jfb.2016.88.issue-3
Ratnasingham S., Hebert P. D. (2007). BOLD: The Barcode of Life Data System (http://www. barcodinglife. org). Mol. Ecol. Notes 7, 355–364. doi: 10.1111/j.1471-8286.2007.01678.x
Ribolli J., de Melo C. M. R., Zaniboni-Filho E. (2012). Genetic characterization of the neotropical catfish Pimelodus maculatus (Pimelodidae, Siluriformes) in the Upper Uruguay River. Genet. Mol. Biol. 35, 761–769. doi: 10.1590/S1415-47572012005000060
Rohl A. (2000). Network: a program package for calculating phylogenetic networks, Version 4.1 (Hamburg: Mathematisches Seminar, University of Hamburg). Available at: http://Www.Fluxus-Engineering.Com (Accessed December 16, 2024).
Ronquist F., Teslenko M., van der Mark P., Ayres D. L., Darling A., Höhna S., et al. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. System. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Rossini B. C., Oliveira C. A. M., de Melo F. A. G., Bertaco V. de A., de Astarloa J. M. D., Rosso J. J., et al. (2016). Highlighting Astyanax species diversity through DNA barcoding. PloS One 11, e0167203. doi: 10.1371/journal.pone.0167203
Rosso J. J., Rueda E. C., Sanchez S., Bruno M. C., Casciotta J., Aguilera G., et al. (2018). Basin-scale distribution and haplotype partitioning in different genetic lineages of the Neotropical migratory fish Salminus brasiliensis. Aquat. Conserv.: Mar. Freshw. Ecosyst. 28, 444–456. doi: 10.1002/aqc.2830
Rubinoff D., Holland B. S. (2005). Between two extremes: mitochondrial DNA is neither the panacea nor the nemesis of phylogenetic and taxonomic inference. System. Biol. 54, 952–961. doi: 10.1080/10635150500234674
Scarabotti P. A., Lucifora L. O., Espínola L. A., Rabuffetti A. P., Liotta J., Mantinian J. E., et al. (2021). Long-term trends of fishery landings and target fish populations in the lower La Plata basin. Neotrop. Ichthyol. 19, e210013. doi: 10.1590/1982-0224-2021-0013
Silva-Santos R., Ramirez J. L., Galetti P. M. Jr., Freitas P. D. (2018). Molecular evidence of a hidden complex scenario in Leporinus cf. friderici. Front. Genet. 9, 47. doi: 10.3389/fgene.2018.00047
Sivasundar A., Bermingham E., Ortí G. (2001). Population structure and biogeography of migratory freshwater fishes (Prochilodus: Characiformes) in major South American rivers. Mol. Ecol. 10, 407–417. doi: 10.1046/j.1365-294X.2001.01194.x
Sverlij S., Liotta J., Minotti P., Brancolini F., Baigún C., Lacoste F. F. (2013). “Los peces del corredor fluvial Paraná-Paraguay,” in Inventario de Los Humedales de Argentina: Sistemas de Paisajes de Humedales Del Corredor Fluvial Paraná Paraguay (Secretaría de Ambiente y Desarrollo Sustentable de la Nación, Buenos Aires), 341–356.
Tsai Y. E. (2011). PhyloGeoViz: A web-based program that visualizes genetic data on maps. Mol. Ecol. Resour. 11, 557–561. doi: 10.1111/j.1755-0998.2010.02964.x
Utsunomia R., Silva D. M. Z., de A., Ruiz-Ruano F. J., Goes C. A. G., Melo S., et al. (2019). Satellitome landscape analysis of Megaleporinus macrocephalus (Teleostei, Anostomidae) reveals intense accumulation of satellite sequences on the heteromorphic sex chromosome. Sci. Rep. 9, 5856. doi: 10.1038/s41598-019-42383-8
Valenciennes A. (1837). "Poissons [plates]" in Voyage dans l'Amérique méridionale. 1834‑42., ed. A. d'Orbigny. (Pitois-Levrault, Paris. Pls.), 1–16.
Keywords: boga, freshwater fish, COI, genetic diversity, conservation
Citation: Coronel FP, Loretán G, Sanchez S, Silva N, Pacheco-Sierra G, Brancolini F, Aichino D, Vargas F, Scarabotti P, Espínola L, Pavanelli CS and Rueda EC (2025) DNA barcoding reveals an unexpected distribution of two Megaleporinus species in the La Plata Basin system. Front. Ecol. Evol. 12:1519907. doi: 10.3389/fevo.2024.1519907
Received: 30 October 2024; Accepted: 06 December 2024;
Published: 07 January 2025.
Edited by:
Karine Frehner Kavalco, Universidade Federal de Viçosa, BrazilReviewed by:
Pablo Bolaños-Villegas, University of Costa Rica, Costa RicaLuiz Roberto Malabarba, Federal University of Rio Grande do Sul, Brazil
Francisco de Menezes Cavalcante Sassi, Federal University of São Carlos, Brazil
Copyright © 2025 Coronel, Loretán, Sanchez, Silva, Pacheco-Sierra, Brancolini, Aichino, Vargas, Scarabotti, Espínola, Pavanelli and Rueda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eva Carolina Rueda, ZXZhLmNhcm9saW5hLnJ1ZWRhQGdtYWlsLmNvbQ==