 Giovanna Sandretti-Silva1,2*
Giovanna Sandretti-Silva1,2* Leandro Corrêa2
Leandro Corrêa2 Mariana Amirati1
Mariana Amirati1 Ivan Rodrigo Abrão Laurino3
Ivan Rodrigo Abrão Laurino3 Fernando Camargo Passos4
Fernando Camargo Passos4 Marcio R. Pie5Marcos R. Bornschein1
Marcio R. Pie5Marcos R. Bornschein1- 1Instituto de Biociências, Departamento de Ciências Biológicas e Ambientais, Universidade Estadual Paulista (UNESP), São Vicente, São Paulo, Brazil
- 2Mater Natura – Instituto de Estudos Ambientais, Curitiba, Paraná, Brazil
- 3Instituto Oceanográfico da Universidade de São Paulo (IO-USP), São Paulo, São Paulo, Brazil
- 4Departamento de Zoologia, Universidade Federal do Paraná (UFPR), Curitiba, Paraná, Brazil
- 5Biology Department, Edge Hill University, Ormskirk, Lancashire, United Kingdom
Introduction: The “win-stay, lose-switch” (WSLS) strategy used by birds involves decisions to maintain or alter the characteristics of the subsequent nest according to the outcome of the previous one. In salt marshes, nest failure occurs due to flooding, predation, or tipping, creating a trade-off for nest placement: it should be low enough to avoid predators from above and tipping by the wind, and high enough to avoid flooding. In salt marshes of Southern Brazil, predation from above is carried out by rails that also prey on nests from below, promoting nondirectional pressure capable of neutralizing bird responses. We aim to test the WSLS strategy and to assess its adaptive significance for Formicivora acutirostris, the only thamnophilid endemic to salt marshes. Our general premise is that the adoption of the WSLS strategy would vary in response to the fate of the previous nest, considering also the environment type.
Methods: We evaluated the fate, environment type, height, altitude, and thickness of nests of F. acutirostris in southern Brazil, between 2006 and 2023. We assessed the effects of the nest attributes, tested the adoption of the WSLS strategy, and explored its adaptive significance using generalized linear mixed models. We also examined the influence of factors such as nest environment, nesting timing, and pair age on nest fate and parental behavior.
Results: We studied 98 renesting cases. Flooded nests were at a lower height than predated, tipped, and successful nests, and tipped nests were thicker than flooded and successful nests. Nest heights differed among environment types. The species adopted the WSLS strategy by increasing nest height after flooding, but we do not support its adaptive significance. No additional factors influenced the behavior of the species.
Discussion: Although the adoption of the WSLS strategy by F. acutirostris was confirmed regarding the increase in nest height after flooding, it did not result in a significant increase in reproductive success, suggesting that other pressures, such as predation and tipping, are limiting the adaptive potential of this strategy. We emphasize the threat of the high reproductive failure for the conservation of F. acutirostris and proposes reducing predation pressure as an important conservation strategy.
1 Introduction
Birds’ breeding decisions and the reasons behind them are directly related to their success (Martin and Roper, 1988; Filliater et al., 1994; Chalfoun and Martin, 2009; Parejo and Avilés, 2010; Benvenuti et al., 2018); thus, understanding it is crucial for conservation (Campomizzi et al., 2012). Essentially, bird parents have the challenge of selecting nest characteristics that minimize risks and increase their reproductive success (Forstmeier and Weiss, 2004; Jiang et al., 2017). Animals can address behavioral tasks in several forms (MaBoudi et al., 2020), and one simple strategy involves adopting behavioral plasticity to maintain or change the characteristics of the current nest using information from the previous one (Refsnider and Janzen, 2012; Benvenuti et al., 2018; Bressler et al., 2020). This behavior requires the association of nest characteristics with the risks and advantages of those characteristics through a mechanism of reward or disappointment (Marzluff, 1988; Freund et al., 2017). Such plasticity characterizes a strategy known as “win-stay, lose-switch” (WSLS; following Freund et al. [2017]) and has been detected in several bird species (Marzluff, 1988; Chalfoun and Martin, 2010; Beckmann and McDonald, 2016; Hunter et al., 2016; Benvenuti et al., 2018; Ma et al., 2019). However, this nesting behavior is not universal in birds and some studies have shown that there are species with unconditional strategies (Hendricks, 1991; Grand and Flint, 1996; Howlett and Stutchbury, 1996; Kershner et al., 2004).
In salt marshes, habitats dominated by herbaceous plants adapted to the salinity and tidal flooding (Tiner, 2013), breeding challenges for birds are particularly complex due to several environmental factors and conflicting pressures. The sparse and simple vegetation can make nests more vulnerable to being tipped by the wind (Burger, 1985; Reinert, 2008) and more exposed to abundant predators from above (Greenberg et al., 2006). In these environments, the regular tidal effect also increases reproductive failure by flooding the nests (Reinert, 2006, 2008). Thus, in salt marshes, there is a trade-off for birds between placing the nest low enough in the dense vegetation to avoid aerial predation and tipping by the wind, and placing it high enough to avoid flooding (Greenberg et al., 2006). This height trade-off presents an ideal opportunity to evaluate the WSLS strategy, and the two studies that evaluated it in salt marsh-dwelling birds from temperate regions have found that they do indeed adopt the strategy for flooding and predation (Hunter et al., 2016; Benvenuti et al., 2018).
Thamnophilids are generally found in forests and avoid open areas (Zimmer and Isler, 2003), such as marshes. In southern Brazil, however, there is a particular salt marsh habitat called “subtropical salt marsh” (Bornschein et al., 2017), to which the threatened Parana Antwren (Formicivora acutirostris) is endemic (Bornschein et al., 2024; Ordinance MMA 148, of 7 June 2022). That species and the Marsh Antwren, F. paludicola (Buzzetti et al., 2013), from southeastern Brazil, are the only members of the species-rich thamnophilid family (c. 240 species; Winkler et al. [2020]) that exclusively inhabit marshes (Buzzetti et al., 2013; Winkler et al., 2020).
Our objective is to test the WSLS strategy in F. acutirostris. Nest flooding, predation, and tipping are the main causes of nest failure for this species (Reinert, 2008). However, in the subtropical salt marshes inhabited by F. acutirostris, predation pressure comes less from visual aerial predators like raptors, which are rare in the area (Guerra, 2023), and more from the Blackish Rail, Pardirallus nigricans, and the Black Rat, Rattus rattus (GS-S and MRB, personal observation), which can access the nest from above or below, depending on its height and the environment type. Therefore, the non-directional predator pressure could increase complexity or eventually neutralize possible bird reward or disappointment responses following predation. Based on previous field knowledge regarding environment types of salt marshes and their available strata for nest placement and local pressures on them (Figure 1), our general premise is that the species adopts the strategy by changing or maintaining the attributes based on previous nest fate. This adoption would involve bird responses in relation to nest height after losses due to flooding and tipping, nest position after losses to predation, and nest thickness after tipping. In addition, it would vary according to the type of the nest environment, with birds preferring to remain in the same type when nesting in favorable environments, regardless of the fate of the previous nest (Figure 2).
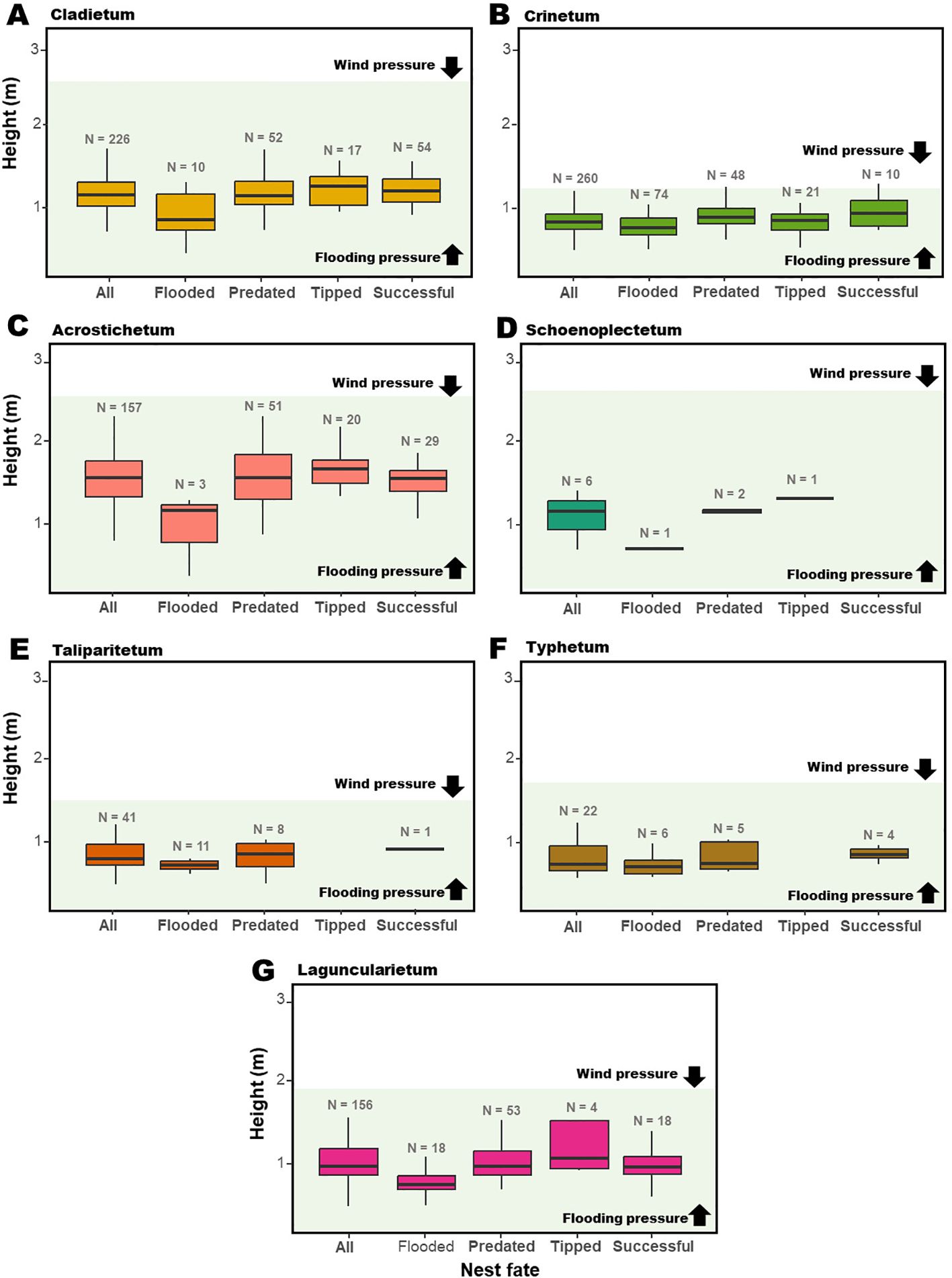
Figure 1. Nest height and fate of Formicivora acutirostris in (A) Cladietum, (B) Crinetum, (C) Acrostichetum, (D) Schoenoplectetum, (E) Taliparitietum, (F) Typhetum, and (G) Laguncularietum in Guaratuba Bay, Paraná, southern Brazil, from 2006 to 2024. The light grey background indicates the height of the environment, based on the mean of the height of the dominant plant species. The boxplot represents the interquartile range (IQR) of nest heights, the line inside the box indicates the median, and the whiskers show the range of heights within 1.5 times the IQR from the quartiles.
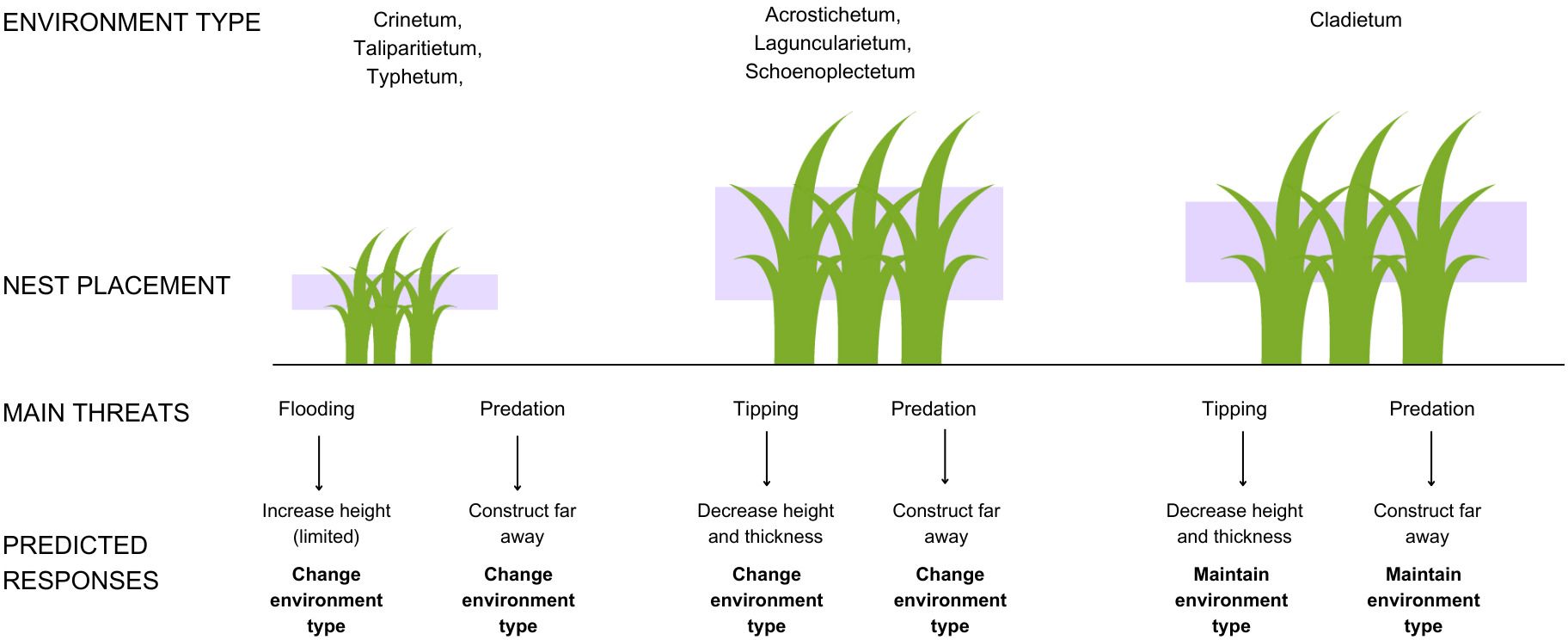
Figure 2. Predicted responses for the premise regarding the adoption of the “win-stay, lose-switch” strategy by Formicivora acutirostris, for all nest fates and environments, based on Reinert, 2008 and Sandretti-Silva et al. (2024). The best response for each environment and nest fate is in bold. Designed by Freepik.
2 Methods
2.1 Target species
Formicivora acutirostris, a threatened species in Brazil (Ordinance MMA 148, of 7 June 2022), is a territorial and socially monogamous bird that forms long-term pairs, sharing reproductive and defensive activities throughout the year (Sobotka, 2011; Reinert et al., 2012; Bornschein et al., 2015). The breeding season extends from August to February (Reinert et al., 2012); the species builds cup-shaped nests attached to vegetation, at a height between 30 and 220 cm above the ground (Reinert et al., 2012), and may renest up to eight times during a breeding season (Reinert, 2008). Formicivora acutirostris lays two eggs (Reinert et al., 2012), incubates them for approximately 16 days (Reinert, 2008), and the nestling stage extends for approximately 10 days (Reinert, 2008).
2.2 Study area
The study was conducted in the Guaratuba Bay, Guaratuba Ramsar Site (see https://rsis.ramsar.org/ris/2317), municipality of Guaratuba, on the southern coast of Paraná, southern Brazil (Figure 3). We worked at the confluence of the Claro and São João rivers (“Continente”; c. 25.873°S, 48.761°W; 8.7 ha) and on Jundiaquara Island (“Jundiaquara”; c. 25.873°S, 48.759°W; 11.5 ha), located upstream on the São João river, and in part of the Folharada Island (“Folharada”; c. 25.866°S, 48.723°W; 16.3 ha), located downstream on this river. The studied areas contain “estuarine marshes” (sensu Doody, 2001), “tidal marshes” (Reinert et al., 2007), “subtropical salt marshes” (Bornschein et al., 2017), or pioneer formation of fluviomarine influence (sensu Veloso et al., 1991; IBGE, 1992). The environment experiences two daily floodings according to a mixed semi-diurnal tide (Lee and Chang, 2019).
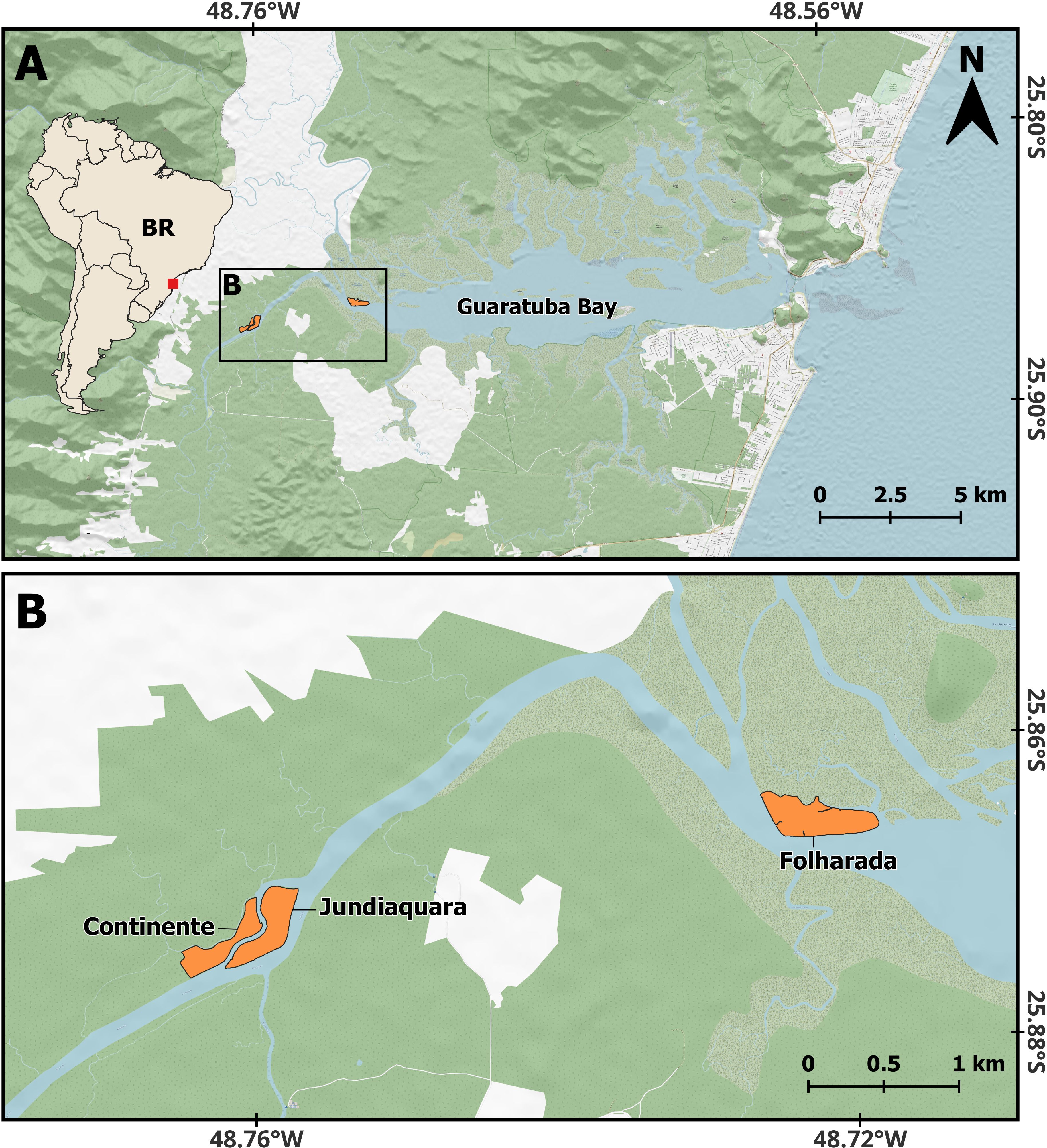
Figure 3. (A) General view and (B) closest view of the study areas for Formicivora acutirostris at the Guaratuba Bay, Paraná, southern Brazil. Background images: Geomorphometric Database of Brazil (TOPODATA), National Water and Sanitation Agency (ANA), OpenStreetMap contributors.
The dominant plant species are crinum lily (Crinum americanum) and California bulrush (Schoenoplectus californicus). Locally, the following species can be abundant: the herbaceous southern cattail (Typha domingensis), swamp sawgrass (Cladium mariscus), giant leather fern (Acrostichum danaeifolium), the bush Talipariti pernambucensis, pond apple (Annona glabra), Brazil beauty-leaf (Calophyllum brasiliense), and white mangrove (Laguncularia racemosa) (Reinert et al., 2007; Bornschein et al., 2017; Favretto et al., 2022).
2.3 Nest search and monitoring
Fieldwork was conducted between January 2006 and March 2023. Until May 2008, it was conducted daily during the breeding season and for 10 days per month outside of it. From June 2008 to March 2023, fieldwork was conducted for 3–8 days per month throughout the year. We worked in the field as a group of two to five people, usually three, from dawn to approximately 1 p.m. and for a further two to three-and-a-half hours in the afternoon, before dusk.
All territorial birds were banded (with a metal numbered band and three colored plastic bands) in unique color codes for individual recognition with binoculars in order to determine the owners of the nests and the nest sequences. During banding, each individual was assigned with an identity code (referred to as “ID number”) and its minimum age was estimated based on the plumage [for details on juvenile and adult plumage patterns, see Reinert (2008)]. To reduce data variability, we estimated the birth date as either October 1 (a period with many early breeding season births) or February 1 (a period with the latest births in the season), in accordance with Bornschein et al. (2015). We assigned the closest birth date that best matched the observed plumage pattern at the time of the banding (Bornschein et al., 2015). Every year, we calculated the resulting age of the individuals by summing a year.
Target nests were systematically searched in 14–41 territories per year during the breeding seasons (August to February), making a total of 521 monitored territories. The nests were located mainly by observing the behavior of individual birds (e.g., carrying plant material or food and alarm vocalizations) that we followed up to discover nests. When finding nests at different stages, we calculated their initiation, as the date of nest construction completion and the start of egg laying, based on mean reproductive times studied by Reinert (2008). The nests were numbered and marked with a ribbon tied on the vegetation to facilitate the correct identification in the returns. We took the geographic coordinates of each nest and information on its height (as lip to the ground), thickness, and environment type. The thickness, which represents the robustness of the nest, may be a factor related to losses due to rips and holes (Palomino et al., 1998; Suárez et al., 2005) and was estimated as the average of the differences between (1) internal and external diameters and (2) nest height and depth of the incubation chamber.
We classified the environment type in which the nest was constructed, based on the dominant plant species. We named them by adding the suffix “etum” to the genus of the dominant species, following Braun-Blanquet (1979), as follows: (1) “Crinetum” (for the domain of the herbaceous C. americanum), (2) “Schoenoplectetum” (for the domain of the herbaceous S. californicus), (3) “Cladietum” (for the domain of the herbaceous C. mariscus), (4) “Acrostichetum” (for the domain of the herbaceous A. danaeifolium), (5) “Typhetum” (for the domain of the herbaceous T. domingensis), (6) “Taliparitetum” (for the domain of the bush T. pernambucensis), and (7) “Laguncularietum” (for the domain of the tree L. racemosa).
We calculated nest altitude based on an altimetric mapping, made with the Real Time Kinematic (RTK) technique in 2014 (when more than 4,000 altitude points were taken with 1 mm precision). The altitude points were distributed in grids of 20 m × 20 m or 5 m × 5 m, the latter for regions with terrain slope. A map was generated with contour lines for every 10 cm of altitude. The altitude of each nest was taken to be that of the altitude of the point that were closest to the nest, but within the same contour line as the nest.
Monitored nests were considered as having success or failure (= nest fate), and failure was attributed to predation, flooding, or tipping. We consider the nest to be successful if at least one fledgling leaves it. We attributed failure to predation when observed or inferred attacks on nests that led to loss of eggs, hatchlings, or the entire nest. We inferred predation if we observed evidence in the nests such as bottom or side holes, eggshell bottoms, or completely ripped nest material, with no signs of friction in the plant fibers. We attributed nest failure to flooding if we observed flooded nests, floating eggs around the nests, drowned chicks inside the nests, wet nests (on non-rainy days), and the presence of debris inside the nests. Finally, we attributed nest failure to tipping if we observed or inferred fallen nests due to the observation of friction in the plant support. We observed many empty nests that should have been active, but it was not possible to attribute the cause of the loss, such as nests that may have been lost due to strong winds or predator approach. In these cases, we attributed the cause of failure as undetermined and removed these data from the analyses.
After determining success or failure, we only counted subsequent nest attempts by the same pair (renesting case) that were carried out to completion (i.e., a complete cup), since there are changes in the height and thickness of nests during construction. We concluded that we were dealing with a case of renesting only when we were certain that no third nest had been built between the previous and subsequent nests.
2.4 Statistical analysis
We compared the characteristics of the previous and the subsequent nests (renesting) to evaluate the changes adopted by the individuals. We calculated the differences in nest attributes by subtracting the height (change in nest height), altitude (change in nest altitude), and thickness (change in nest thickness) of the nests (change in nest attribute = second nest attribute − previous nest attribute). Additionally, considering the position and environment of the nests, we calculated the distance between subsequent nests and assessed whether there was change in environment type.
We conducted three different analyses to assess the adoption of the WSLS strategy by F. acutirostris, as well as its adaptive significance. Initially, we tested the relationship between the nest attributes and their fates (analysis 1). Next, we evaluated the relationship between changes in nest characteristics in renesting cases and the fate of the previous nest (analysis 2) to determine whether the species makes systematic changes (Figure 2). Finally, we tested the relationship between the changes in nest characteristics and the success or failure of the second nest (analysis 3) to assess the adaptive significance of these changes. In addition to the previously mentioned nest attributes (analysis 1) and changes in these attributes (analyses 2 and 3) to test our main hypotheses, we also included potential factors associated with nest fate or nesting behavior, such as seasonality (indicated by month of nesting initiation), environment type (simplified according to the sample size and objective), and parental age (Table 1) in the models. We did not include these potential factors in analysis 3 due to the low sample size resulting from the model’s specificity (see below). For the same reason of sample size, thickness and altitude were not included in the models of this analysis. We used generalized linear mixed models (GLMMs) with different functions (see below) from the lme4 1.1-35.1 package (Bates et al., 2024) in R (version 4.3.3; R Core Team, 2024).
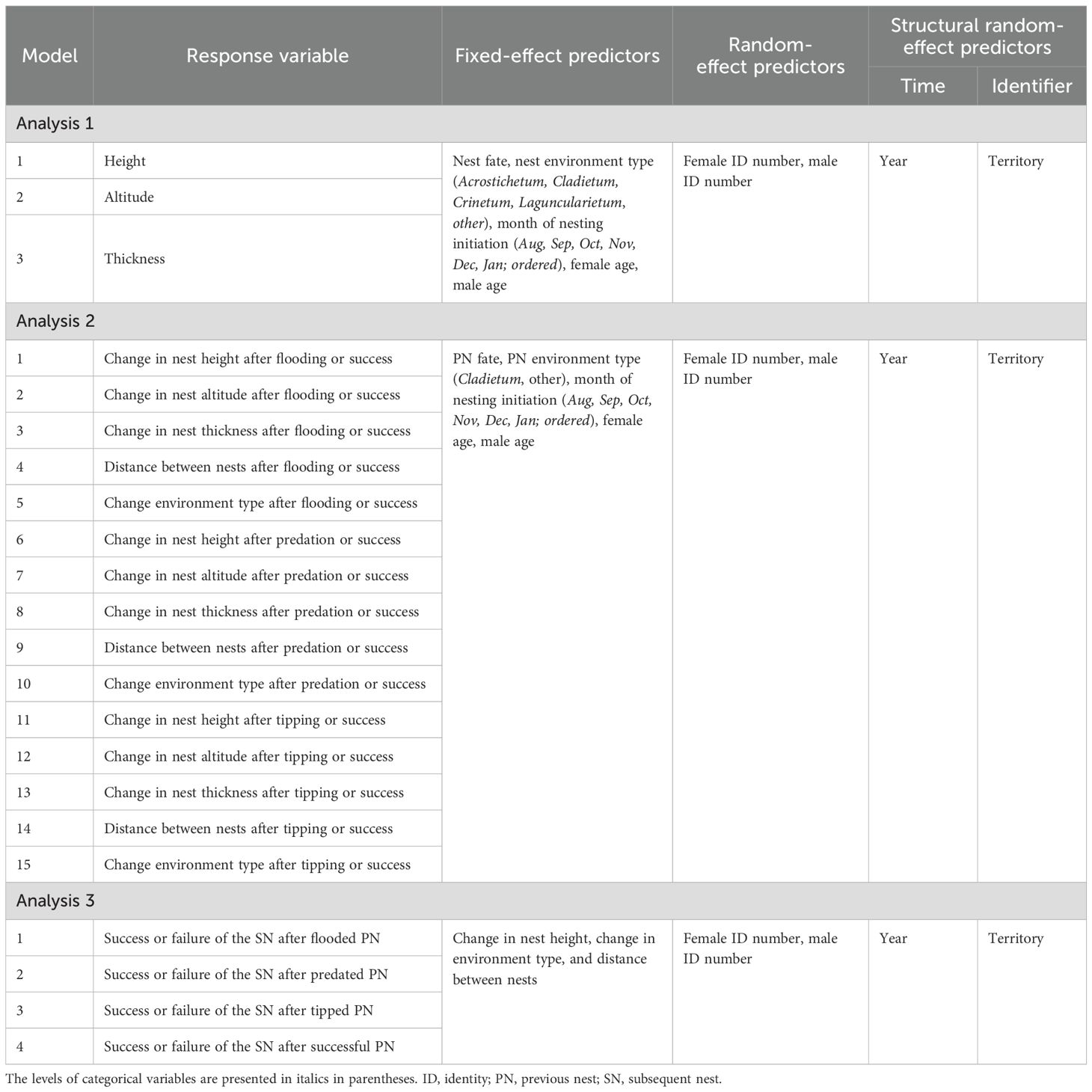
Table 1. Structure of the generalized linear mixed models for testing the adoption of the “win-stay, lose-switch” strategy by Formicivora acutirostris and its potential adaptive significance.
For analysis 1, we constructed three models, one for each nest attribute (height, altitude, and thickness), analyzing each one for the different nest fates (Table 1). For analysis 2, we constructed 15 models, one model for each of the five changes in nest attributes (nest height, altitude, thickness, distance between nests, and environment type) across the three comparisons between success (“win”) or failure (“lose” = flooded, predated, or tipped) of the previous nest (Table 1). For analysis 3, we built four models, one for each fate of the first nest (flooded, predated, tipped, and successful nets), comparing what led to the success or failure of the subsequent nest (Table 1).
We included random variables in all models. Territory and year were treated as structural random predictors, meaning they were retained in all models (i.e., not removed during model selection; see below) to account for the identifier and time of repeated measures or longitudinal samples. Additionally, in all models, female ID number and male ID number were also included as random predictors, but they could be removed during model selection (see below) if this improved the model fit. The complete model structure can be found in Table 1.
We disregarded the models with a sample size smaller than twice the number of predictor variables included. To avoid over-testing due to multiple parallel analyses, we adjusted the p-values for each of the three analyses separately. For this, we used the p.adjust function from the stats 4.3.3 R package (R Core Team, 2024), applying the Bonferroni method.
After selecting the best complete model distribution based on AIC values and convergence success, we proceeded with variable selection. We evaluated the importance ranking of the predictors using the MERF function from the LongituRF 0.9 package (Capitaine, 2020). We designated territory as the identifier and year as the time trajectory—for this reason, they are treated as structural variables in our GLMMs, as they contributed to the longitudinal structure of the models and did not have their importance obtained. The model was configured without any stochastic process, two predictor variables per split, and with other default settings. For random-effect predictors, importance was measured by summing their estimates, and for fixed-effect predictors, we assessed importance based on the percent increase in mean squared error (%IncMSE; Supplementary Tables 1–3). We sequentially removed the least significant random-effect predictors until their best combination based on AIC is identified, considering equally adjusted models with ΔAIC ≤ 2 (Bolker, 2008). Next, we applied the same method of removing the least important fixed predictors to obtain the best model.
Model validation was conducted by verifying normality and homoscedasticity of residuals using the residual, fitted, bptest, and shapiro.test functions of the stats (R Core Team, 2024) and lmtest 0.9-40 (Hothorn et al., 2022) R packages for normal and log-normal models. For binomial models, we validated the results by analyzing the receiver operating characteristic (ROC) curve and calculating the area under the curve (AUC) using the predict and roc functions of the stats (R Core Team, 2024) and pROC 1.18.5 (Robin et al., 2023) R packages. If our best models did not validate, we removed outliers, tested alternative distributions, or selected more complete models of the best distribution. In order to verify the differences in nest fate for analysis 1, we applied pairwise comparison tests using the emmeans and pairs function from the emmeans 1.10.4 (Lenth, 2024) and multcomp 1.4.26 (Hothorn et al., 2024) R packages.
3 Results
We located 963 nests, of which 785 were built up to completion (the rest had been abandoned during the construction phase). We monitored 98 renesting cases involving a total of 166 nests, of which 150 proceeded to the egg-laying stage, and we could not monitor the remaining 16 nests after they became a cup. In half of the cases, we studied only one annual renesting by each pair (n = 49 cases), but the pairs carried out up to six successive renestings tracked by us (= 1.46). The renesting cases included a total of 90 individuals in 50 pair combinations. The nests were built by females aged 0.5 to 14.5 years (x = 3.88 years; SD = 2.93) and by males aged 0.5 to 13.5 years (x = 4.24 years; SD = 3.42). The largest number of renesting events occurred after the predation of the previous nest (n = 42), followed by renesting after flooding (n = 22), after tipping (n = 15), and after the success of the previous nest (n = 19). The main cause of nest failure was predation (n = 66), followed by flooding (n = 34) and tipping (n = 27). The overall nest success rate was 23.49%. The renesting attempts occurred at 3.4 to 116.3 m from the first nest ( = 33.38 m, n = 93).
3.1 The relationship between nest attributes and fates (analysis 1)
The attributes of the nests of F. acutirostris were related to their fate (Figure 4; Table 2). The height of the nests varied according to nest fate (Figure 4A), with flooded nests being at lower heights than predated (pairwise, p < 0.001), tipped (pairwise, p < 0.001), and successful nests (pairwise, p < 0.001). However, there was no difference in the height of successful nests versus predated (pairwise, p = 0.986) and tipped nests (pairwise, p = 1.000), and in the height of predated versus tipped nests (pairwise, p = 0.996). Nest height also varied between environments, with nests in Cladietum being at higher heights than those in Crinetum (pairwise, p = 0.001), Taliparitetum, Typhetum, and Schoenoplectetum (“other”; pairwise, p < 0.001); at lower heights than nests in Acrostichetum (pairwise, p = 0.002); and at similar heights to nests in Laguncularietum (pairwise, p = 0.195). Nests in Acrostichetum were at higher heights than those nests in Crinetum (pairwise, p < 0.001), in Laguncularietum (pairwise, p < 0.001), and in Taliparitetum, Typhetum, and Schoenoplectetum (“other”; pairwise, p < 0.001). There was no difference in the height of nests in Crinetum versus nests in Laguncularietum (pairwise, p = 0.560); nests in Crinetum versus nests in Taliparitetum, Typhetum, and Schoenoplectetum (“other”; pairwise, p = 0.856); or nests in Laguncularietum versus nests in Taliparitetum, Typhetum, and Schoenoplectetum (“other”; pairwise, p = 0.116).
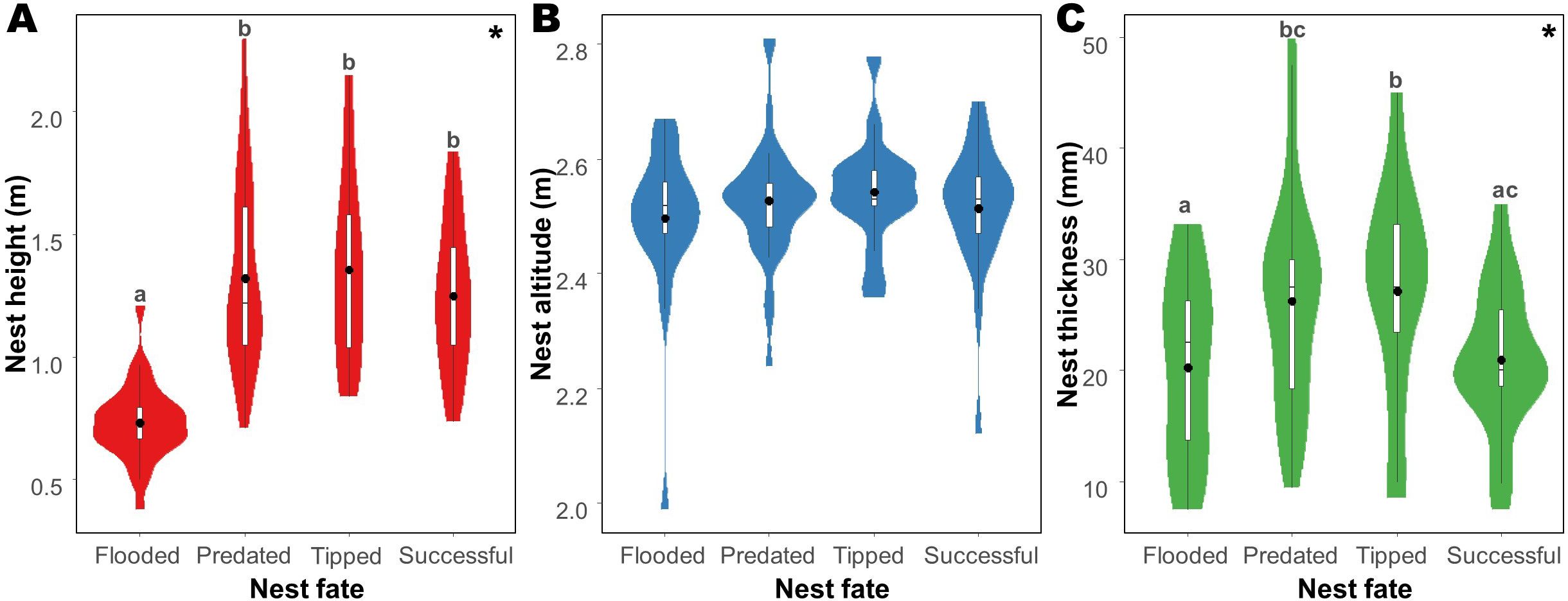
Figure 4. Attributes of Formicivora acutirostris nest in Guaratuba Bay, Paraná, southern Brazil, for all nest fates. Nest (A) height, (B) altitude, and (C) thickness. The violin plots provide a visual representation of the data distribution (width of the violin) and the boxplot provides the following summary statistics: mean value (black points), interquartile range (IQR) of data (boxplot), median (line inside the box), and the range of heights within 1.5 times the IQR from the quartiles (whiskers). Asterisks (*) indicate significance, and the lowercase letters (“a”, “b”, and “c”) show significant differences between groups.
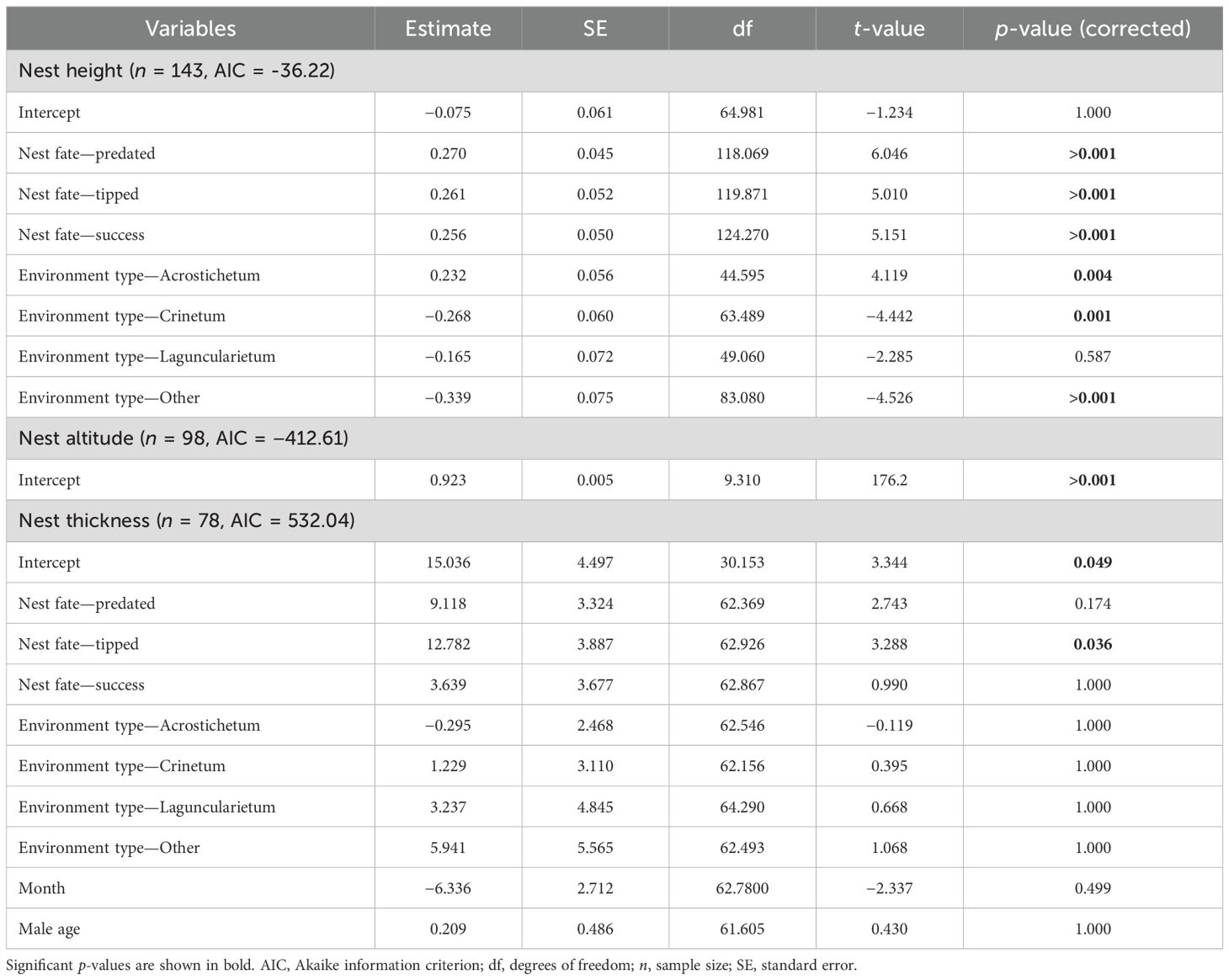
Table 2. Generalized linear mixed models for testing relationship between the nest fates and nest attributes (analysis 1) of Formicivora acutirostris.
Nest thickness was related to nest fate (Figure 4C; Table 3), with predated and tipped being thicker than flooded (pairwise, p = 0.046 and p = 0.012), and tipped nests being thicker than successful nests (pairwise, p = 0.043). Predated nests were equally thicker than tipped (pairwise, p = 0.664) and successful nests (pairwise, p = 0.274); and flooded nests were also equally thicker than successful nests (pairwise, z = 0.251; p = 0.774). We did not observe a relationship between nest altitude and fate (Figure 4B; Table 3).
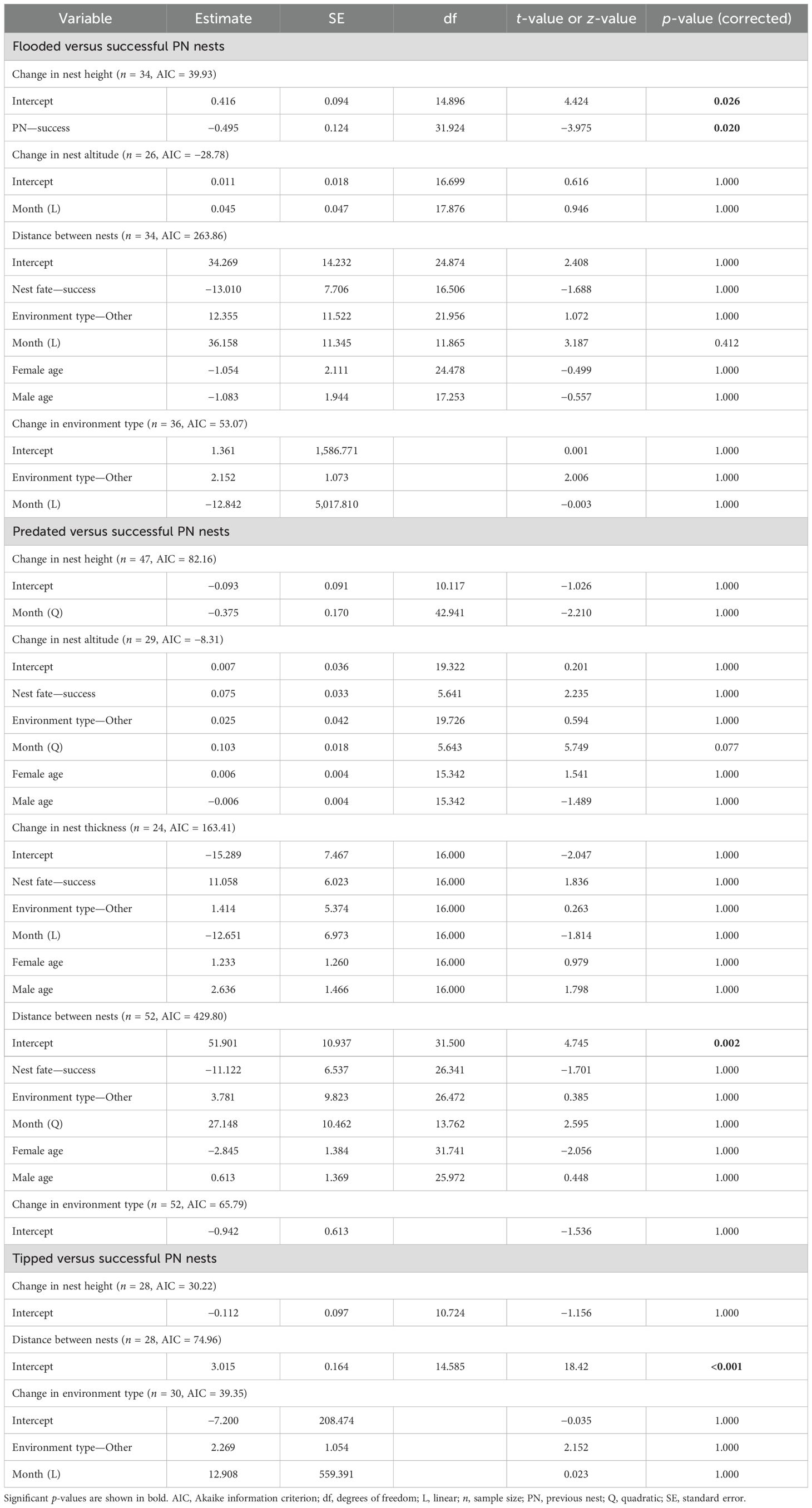
Table 3. Generalized linear mixed models for testing the relationship between previous nest fates and changes in characteristics of the nests in renesting cases (analysis 2) of Formicivora acutirostris.
3.2 Testing the WSLS strategy (analysis 2)
Changes in subsequent nest characteristics were related to the fate of the first nest (Figure 5; Table 3). After losing the nests due to flooding, the species constructed the second nest higher, making significant changes in nest height compared to chances in height after experiencing success (GLMM, p = 0.020; Figure 5A). The changes in nest height following a successful nest were similar to those after predation and tipping (Figures 5B, C; Table 3). Similarly, the changes in nest altitude after experiencing success were similar to those after experiencing flooding and predation of the nest (Figures 5D, E; Table 3), and changes in nest thickness after a successful nest was similar to those after a predated nest (Figure 5H; Table 3). The distances between nests after experiencing success were similar to those after experiencing flooding, predation, and tipping (Figures 5J–L; Table 3), as well as the changes in nest environment (Figures 5M–O; Table 3). No other factors significantly influenced the changes in nests’ attributes adopted by the species (Table 3). Because of the small sample size, it was not possible to analyze changes in nest altitude after tipping (n = 16), as well as changes in nest thickness after flooding (n = 141) and tipping (n = 2.).
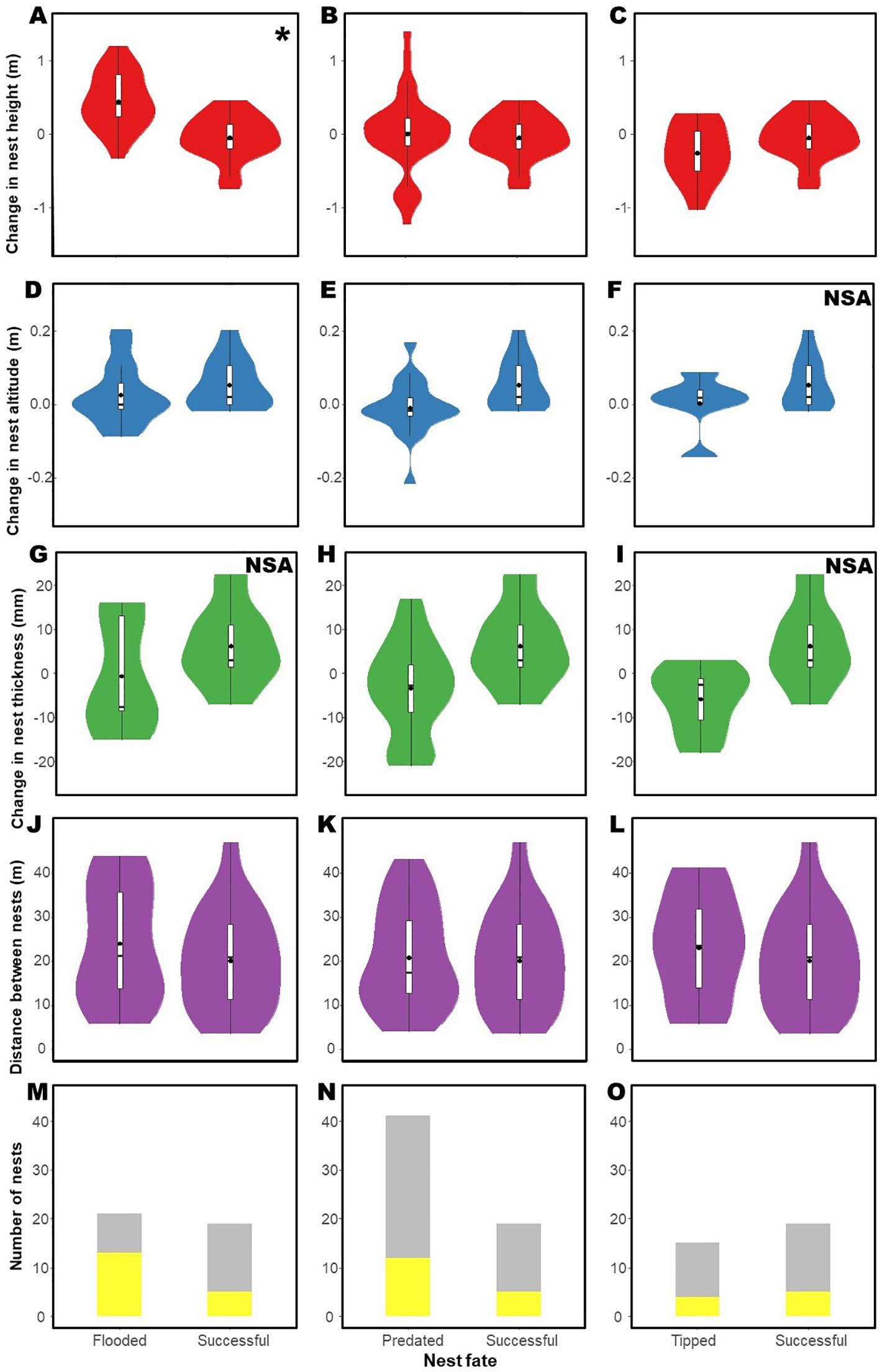
Figure 5. Changes in characteristics of the nests in renesting cases of Formicivora acutirostris in Guaratuba Bay, Paraná, southern Brazil, after failure or success of the previous nest, independent of the fate of the subsequent nest. Changes in nest height after (A) flooding versus success, (B) predation versus success, and (C) tipping versus success. Changes in nest altitude after (D) flooding versus success, (E) predation versus success, and (F) tipping versus success. Changes in nest thickness after (G) flooding versus success, (H) predation versus success, and (I) tipping versus success. Changes in distance between posterior nests after (J) flooding versus success, (K) predation versus success, and (L) tipping versus success. Changes in nest environment selected after (M) flooding versus success, (N) predation versus success, and (O) tipping versus success. The cases highlighted in yellow indicate changes in the environment, while those in gray represent environmental maintenance. The violin plots provide a visual representation of the data distribution (width of the violin) and the boxplot provides the following summary statistics: mean value (black points), interquartile range (IQR) of data (boxplot), median (line inside the box), and the range of heights within 1.5 times the IQR from the quartiles (whiskers). Asterisks (*) indicate significance. NSA, not statistically assessed.
3.3 Testing the adaptive significance of the changes (analysis 3)
Changes in nest height, distance between nests, and in the environment type did not influence the success rate of subsequent nests following flooding, predation, or success of the previous nest (Figure 6; Table 4). Analysis 3 could not be conducted following previous nest tipping due to the small sample size (n = 11).
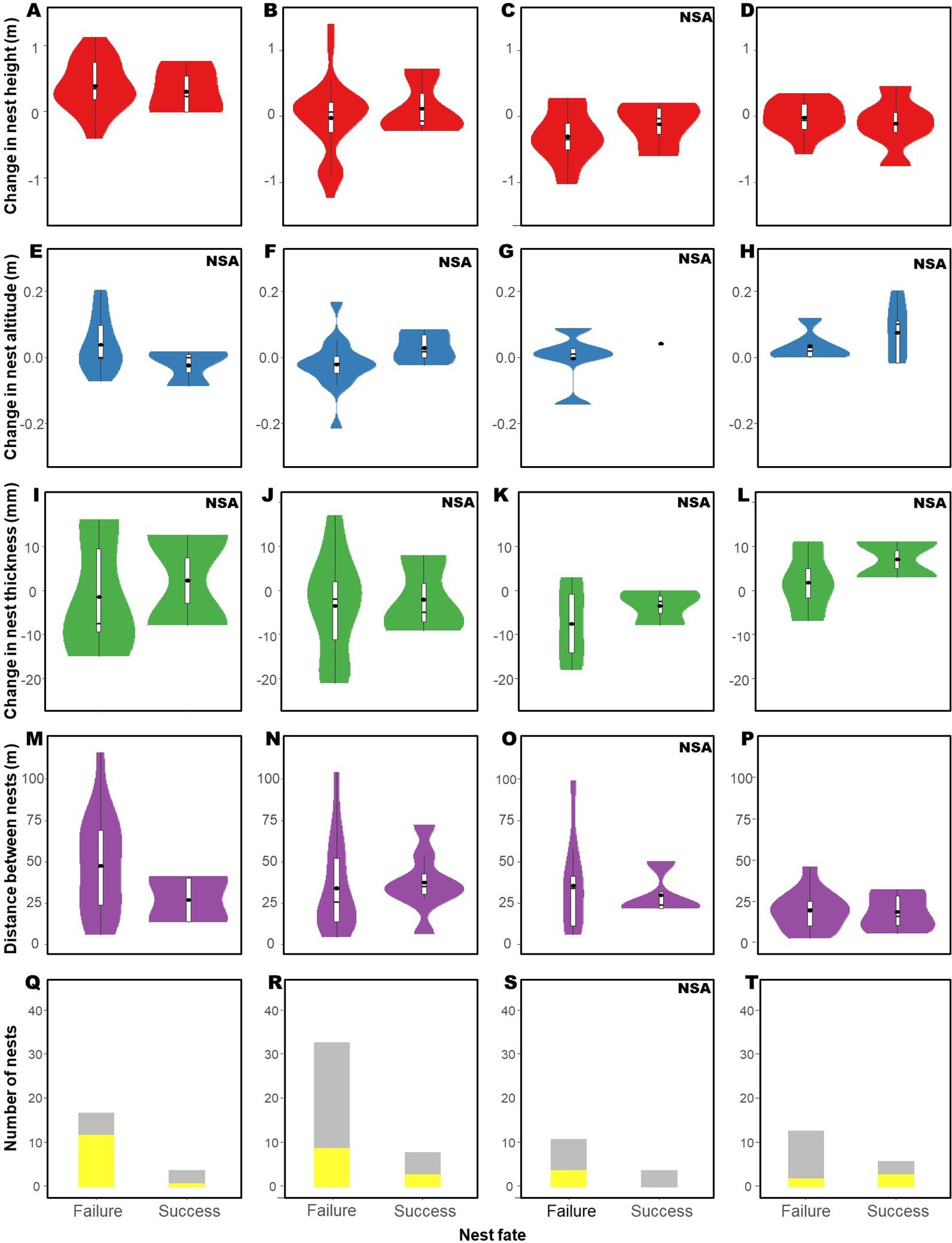
Figure 6. Changes in characteristics of the nests in renesting cases of Formicivora acutirostris in Guaratuba Bay, Paraná, southern Brazil, after all previous nest fates, presented for success and success of the subsequent nest separately. Changes in nest height after (A) flooding, (B) predation, (C) tipping, and (D) success. Changes in nest altitude after (E) flooding, (F) predation, (G) tipping, and (H) success. Changes in nest thickness after (I) flooding, (J) predation, (K) tipping, and (L) success. Changes in distance between nests after (M) flooding, (N) predation, (O) tipping, and (P) success. Changes in nest environment after (Q) flooding, (R) predation, (S) tipping, and (T) success. The cases highlighted in yellow indicate changes in the environment, while those in gray represent environmental type maintenance. The violin plots provide a visual representation of the data distribution (width of the violin) and the boxplot provides the following summary statistics: mean value (black points), interquartile range (IQR) of data (boxplot), median (line inside the box), and the range of heights within 1.5 times the IQR from the quartiles (whiskers). NSA, not statistically assessed.
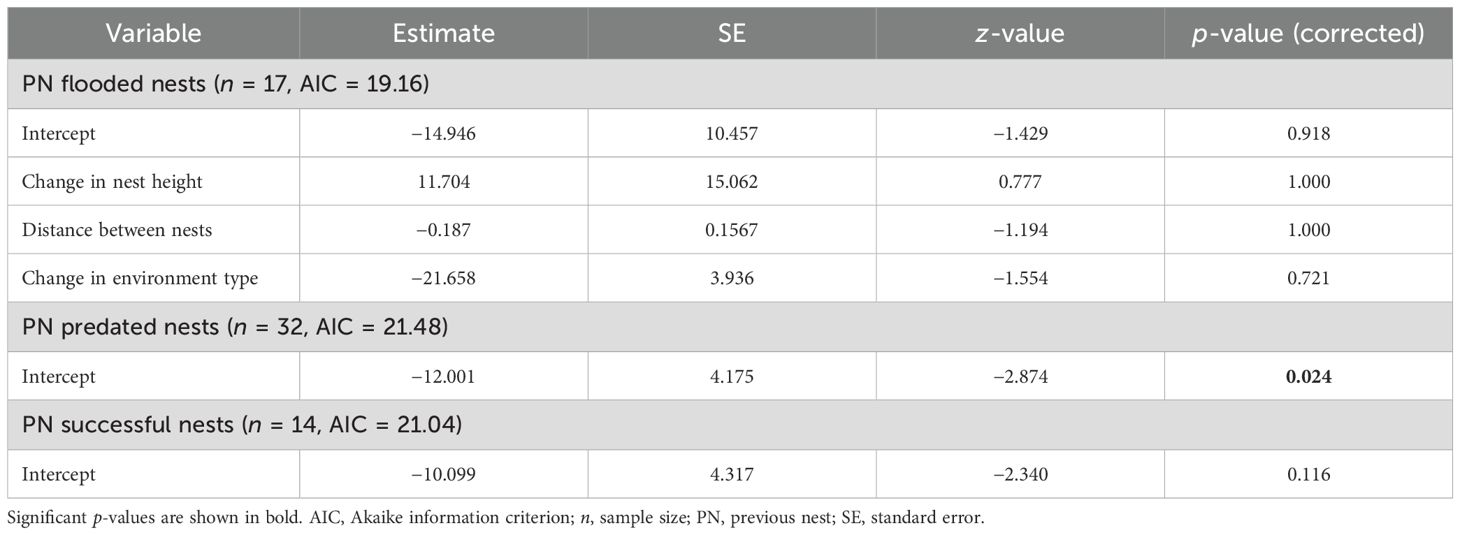
Table 4. Generalized linear mixed models for testing the relationship between changes in characteristics of the nests in renesting cases and the success of the subsequent nest attempt (analysis 3) of the Formicivora acutirostris.
4 Discussion
We found that F. acutirostris shows some behavioral plasticity according to previous nesting experience. The species makes greater changes in nest height, a key attribute for nesting in salt marshes (Figure 4; Table 2), to avoid flooding (Figure 5; Table 3). However, the species does not make greater changes in nest attributes after predation and tipping. This result partially supports the adoption of the WSLS strategy by the species and our general premise. However, this strategy does not influence the success rate of the subsequent nest (Figure 6; Table 4).
Tidal flooding is an important factor for the dynamics of salt marsh birds because it constantly threatens their productivity by drowning chicks and eggs, and by displacing eggs, which are carried away by the water (Reinert, 2006; Greenberg et al., 2006; Reinert, 2008; Reinert et al., 2012; this study). Marsh-dwelling bird species have thus evolved some ways to respond to flooding (Reinert, 2006; Shriver et al., 2007; Hunter et al., 2016; Benvenuti et al., 2018). The increase in nest height after nest flooding detected for F. acutirostris in the present study corroborates the results found for other species (Hunter et al., 2016; Benvenuti et al., 2018) and highlights the importance of nest height for estuarine birds (Greenberg et al., 2006; Hunter et al., 2016; Benvenuti et al., 2018). However, the adjustment in nest height in response to flooding did not increase the reproductive success of the subsequent nest of F. acutirostris, probably because of its exposure to other pressures, such as predation and tipping.
In general, predation is the main reason for nest failure and an important driver of evolution in birds (Oniki, 1979; Winkler, 2016). Therefore, many species adopt the WSLS strategy against this threat by systematically building nests at higher, or lower, heights (Marzluff, 1988; Chalfoun and Martin, 2010; Hunter et al., 2016; Benvenuti et al., 2018; Ma et al., 2019). For F. acutirostris, the nest height was not a significant attribute associated with predation, as it was similar between predated, tipped, and successful nests (Figure 4A). This might be due to a balanced effect of predators from above and below that results in non-directional predation pressure. Rallids forage for food all the time (Taylor, 1996), and P. nigricans could predate the nest from below, when moving in the lower strata of the vegetation, or from above, when moving in the intermediate strata of the vegetation (sensu Bornschein et al., 2022). Rattus rattus, more abundant downstream on Folharada, could also access the nests through the mangrove branches, both from above and below (GS-S and MRB, personal observation). In fact, we detected nests with small holes in the bottom of the nest cup and nests with nesting material pulled upwards, and we also observed an 8-day-old nestling stuck alive pinned by wing feathers on leaves beside the nest with a curved tarsus and some removed remiges, indicating predation from above and below.
The thickness of the nests proved to be an important factor influencing their fate, with thicker nests being more frequently tipped (Figure 4C; Table 2). Despite the suggestion that more nest material improves nest stability (Collias and Collias, 1984), this may be the opposite in the vegetation of salt marshes, where more nesting material could imply heavier nests and the need for attachment to a greater number of plant stems, favoring the tipping of the nest as a result of the movement of these stems due to the action of the wind. This fate will probably become more frequent due to increased wind speed caused by the climate change (Jong et al., 2019); thus, it would be valuable for the species to adopt the strategy of reducing thickness after tipping, a tendency that has been observed but not statistically assessed by us (Figure 5I). Although we could not statistically assess the adaptive significance of the changes in nest thickness, the graphic suggests that they do not impact the reproductive success (Figure 6).
We expected to observe the adoption of greater distances between nests after predation by F. acutirostris, as observed in other studies (Chalfoun and Martin, 2010; Benvenuti et al., 2018). This could benefit the species by helping it to find environments with different predator compositions, vegetation susceptibility, and nest site concealment (Chalfoun and Martin, 2010; Benvenuti et al., 2018). The absence of adopting a distance strategy by F. acutirostris could be related to the active foraging behavior (see Taylor, 1996) and abundance (Guerra, 2023) of its main predator, which is likely present throughout the territories of F. acutirostris, regardless of position, interfering with the reward system of the adopted changes. Furthermore, the absence of adopting a strategy for distance may occur due to an eventual choice of micro-habitat characteristics that were not explored in the present study and that are independent of nest position, since the influence of micro-habitat appears to be quite significant for the reproduction of the species (Sandretti-Silva, 2024). In contrast, large distances in renesting are not expected after success, as well as after flooding, as the species aims to maintain favorable characteristics and minimize the time spent making new choices to ensure a rapid nesting attempt since tidal conditions are also a temporal threat (Chalfoun and Martin, 2010; Benvenuti et al., 2018).
Although the environment types exhibit different rates of nesting success for F. acutirostris (Figure 1), the species does not systematically choose distinct environments following failures, nor does it consistently remain in the same environments after success (Figure 5). This may be related to the quantity (ha) and quality (vegetation density) of each environment type for each pair at the time of the change, which was not accounted for in our models. Vegetation characteristics in the species’ territories are important factors influencing the production of independent juveniles due to the safety of the nesting site and exposure to predators and bad weather (Sandretti-Silva, 2024).
5 Conclusion
The biotic (environment) and abiotic (high tides and altitudes) heterogeneity of F. acutirostris habitats are propitious to the occurrence of behavioral plasticity in nest construction by the species, which is crucial for marsh-dwelling species to deal with the strong pressures in dynamic environments (Benvenuti et al., 2018). Simultaneously, the interplay of nest flooding, predation, and tipping creates a complex challenge for the reproductive success of F. acutirostris. Our results highlight that the WSLS strategy is adopted by this species in relation to flooding but is insufficient for increasing its reproductive success, likely because it exposes the nests to other pressures, leading to uncertain long-term viability (Sandretti-Silva et al., 2024).
This adaptability of F. acutirostris is particularly important in response to flooding and could also be important in response to tipping events, which are becoming more frequent due to climate change, potentially affecting the reproductive success of birds in salt marshes (Jong et al., 2019; Ma et al., 2019, this study). The timing of this adaptation is also crucial, as climatic events tend to vary faster than species’ responses (Meyer and Pie, 2022), making the adoption of conservation measures essential. Given that direct mitigation of flooding and tipping due to high tides and strong winds is not feasible, conservation strategies must focus on reducing predation pressure. Managing predator populations could improve reproductive success not only by directly reducing nest predation but also by enhancing the effectiveness of the already adopted WSLS strategy. We recommend management aimed at reducing populations of R. rattus and P. nigricans. Since this bird is native to Brazilian fauna, though not endangered, obtaining permits from the relevant environmental agencies may be challenging. Therefore, a pilot project in the studied areas could yield consistent short-term results, leading to approval and implementation of long-term predator management.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
GS-S: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Validation, Visualization, Writing – original draft, Writing – review & editing. LC: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. MA: Investigation, Writing – review & editing. IRAL: Formal analysis, Methodology, Visualization, Writing – review & editing. FCP: Writing – review & editing, Validation. MRP: Formal analysis, Investigation, Methodology, Supervision, Writing – review & editing. MRB: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES; Edital Pró-Reitoria de Pós-Graduação PROPG 38/2024; financial code #9324), Fundação Grupo Boticário de Proteção à Natureza (FGBPN; projects #0682/20052; #0740/20071; #0908_20112; #BL0001_20111, #0004_2012, and #1110_20172), Fundo Brasileiro para a Biodiversidade (FUNBIO), 1a. Vara Federal de Paranaguá (process #50005063420184047008), the National Council for Scientific and Technological Development (CNPq; process #314038/2021-3), and the São Paulo Research Foundation (FAPESP; processes #2022/04847-7 and #2023/ 09718-3).
Acknowledgments
Bianca Luiza Reinert helped in several aspects to bring the work to reality, including data collection, project management, and funding acquisition. Juliana M. Berbert, Darren O’Connell, Carla Fontana, Miguel Marini, André Guaraldo, and Marco A. Pizo made suggestions that improved the quality of the study. Ricardo Belmonte-Lopes, Daiane D. Sobotka, Cláudia Golec, Mario Arthur Favretto, Tiago Machado de Souza, Larissa Teixeira, Bruno Guerra, Tamiris Pereira Lima, Maria Fernanda Ferreira Rivas, Cecı́lia Camargo Rocha, Gabriela Villalobo Nascimento, and Ailton Degues helped with data collection. The content of the manuscript have previously appeared online as part of a dissertation (Sandretti-Silva, 2024).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2024.1497317/full#supplementary-material
References
Bates D., Maechler M., Bolker B., Walker S. (2024). Lme4: Linear mixed-effects models using ‘Eigen’ and S4 (R package, version 1.1-35.1). Available at: https://cran.r-project.org/package=lme4.
Beckmann C., McDonald P. G. (2016). Placement of re-nests following predation: are birds managing risks? Emu 116, 9–13. doi: 10.1071/MU15064
Benvenuti B., Walsh J., O’Brien K. M., Kovach A. I. (2018). Plasticity in nesting adaptations of a tidal marsh endemic bird. Ecol. Evol. 8, 10780–10783. doi: 10.1002/ece3.4528
Bolker B. (2008). Ecological models and data in R (Princeton and Oxford: Princeton University Press).
Bornschein M. R., Pizo M. A., Sobotka D. D., Belmonte-Lopes R., Golec C., MaChado-de-Souza T., et al. (2015). Longevity records and signs of aging in Marsh Antwren Formicivora acutirostris (Thamnophilidae). Wilson J. Ornithol. 127, 98–102. doi: 10.1676/14-074.1
Bornschein M. R., Reinert B. L., MaChado-de-Souza T., Golec C., Whitney B. M., Favretto M. A. (2017). Abundance, occurrence, and seasonality of the Subtropical Doradito (Pseudocolopteryx acutipennis) on the coast of Brazil. Wilson J. Ornithol. 129, 199–206. doi: 10.1676/1559-4491-129.1.199
Bornschein M. R., Sandretti-Silva G., Sobotka D. D., Corrêa L., Reinert B. L., Vannucchi F. S., et al. (2024). Revision of the conservation status and assessment of the Green Status of the Parana Antwren Formicivora acutirostris, with management proposals. Bird Conserv. Int. 34, e21. doi: 10.1017/S095927092400008X
Bornschein M. R., Teixeira L., Guerra B., de M., Melchiori B. L., Reinert B. L., et al. (2022). Appearance of a population of the mangrove rail Rallus longirostris (Rallidae) in salt marshes invaded by the exotic tanner grass Urochloa arrecta (Poaceae) and its disappearance after plant management. Wetlands 42, 124. doi: 10.1007/s13157-022-01642-7
Braun-Blanquet J. (1979). Fitosociologia: bases para el estúdio de las comunidades vegetales (Madrid: Blume Ediciones).
Bressler S. A., Diamant E. S., Tingley M. W., Yeh P. J. (2020). Nests in the cities: adaptive and non-adaptive phenotypic plasticity and convergence in an urban bird. PNAS 287, 20202122. doi: 10.1098/rspb.2020.2122
Burger J. (1985). “Habitat selection in temperate marsh-nesting,” in Habitat selection in birds. Ed. Cody M. L. (Academic Press Inc, Los Angeles, CA), 253–278.
Buzzetti D. R. C., Belmonte-Lopes R., Reinert B. L., Silveira L. F., Bornschein M. R. (2013). A new species of Formicivora Swainson 1824 (Thamnophilidae) from the state of São Paulo, Brazil. Rev. Bras. Ornitol. 21, 269–291.
Campomizzi A. J., Morrison M. L., DeWoody J. A., Farrell S. L., Wilkins N. (2012). Win-stay, lose-switch and public information strategies for patch fidelity of songbirds with rare extra-pair paternity. Sci. Rep. 2, 294. doi: 10.1038/srep00294
Capitaine L. (2020). LongituRF: random forests for longitudinal data (R package, version 0.9). Available at: https://cran.r-project.org/package=LongituRF.
Chalfoun A. D., Martin T. E. (2009). Habitat structure mediates predation risk for sedentary prey: experimental tests of alternative hypotheses. J. Anim. Ecol. 78, 497–503. doi: 10.1111/j.1365-2656.2008.01506.x
Chalfoun A. D., Martin T. E. (2010). Facultative nest patch shifts in response to nest predation risk in the Brewer’s sparrow: a “win-stay, lose-switch” strategy? Oecologia 163, 885–892. doi: 10.1007/s00442-010-1679-0
Collias N. E., Collias E. C. (1984). Nest building and bird behavior (Princeton: Princeton University Press).
Doody J. P. (2001). Coastal conservation and management: an ecological perspective (Boston, Dordrecht and London: Kluwer Academic Publishers).
Favretto M. A., MaChado-de-Souza T., Golec C., Reinert B. L., Bornschein M. R. (2022). Habitat selection in Many-colored Rush Tyrant (Tachuris rubrigastra) and Wren-like Rushbird (Phleocryptes melanops) in the subtropical salt marshes of Brazil. Stud. Neotrop. Fauna Environ. 59, 276–288. doi: 10.1080/01650521.2022.2101351
Filliater T. S., Breitwisch R., Nealen P. M. (1994). Predation on northern cardinal nests: does choice of nest site matter? Condor 96, 761–768. doi: 10.2307/1369479
Forstmeier W., Weiss I. (2004). Adaptive plasticity in nest-site selection in response to changing predation risk. Oikos 104, 487–499. doi: 10.2307/1369479
Freund M., Bahat O., Motro U. (2017). Nest-site fidelity in Griffon Vultures: a case of win–stay/lose–shift? Isr. J. Ecol. Evol. 63, 39–43. doi: 10.1163/22244662-06301007
Grand J. B., Flint P. L. (1996). Renesting ecology of northern pintails on the Yukon-Kuskokwim Delta, Alaska. Condor 98, 820–824. doi: 10.2307/1369862
Greenberg R., Elphick C., Nordby J. C., Gjerdrum C., Spautz H., Shriver G., et al. (2006). Flooding and predation: trade-offs in the nesting ecology of tidal-marsh sparrows. Stud. Avian Biol. 32, 96–109.
Guerra B. de M. (2023). Abundância e biomassa de aves em brejos salinos no Sul do Brasil. Universidade Estadual Paulista, São Vicente (SP).
Hendricks P. (1991). Site fidelity and renesting of female American pipits. J. Field Ornithol. 62, 338–342.
Hothorn T., Bretz F., Westfall P. (2024). Multcomp: Simultaneous inference in general parametric models (R package, version 1.4-26). Available at: https://cran.r-project.org/package=multcomp.
Hothorn T., Zeileis A., Farebrother R. W., Cummins C. (2022). lmtest: Testing linear regression models (R package, version 0.9-40). Available at: https://cran.r-project.org/package=lmtest.
Howlett J. S., Stutchbury B. J. (1996). Nest concealment and predation in hooded warblers: experimental removal of nest cover. Auk 113, 1–9. doi: 10.2307/4088930
Hunter E. A., Nibbelink N. P., Cooper R. J. (2016). Threat predictability influences seaside sparrow nest site selection when facing trade-offs from predation and flooding. Anim. Behav. 120, 135–142. doi: 10.1016/j.anbehav.2016.08.001
IBGE (1992). Manual técnico da vegetação brasileira (Rio de Janeiro: Fundação Instituto Brasileiro de Geografia e Estatística).
Jiang A., Jiang D., Zhou F., Goodale E. (2017). Nest-site selection and breeding ecology of streaked wren-babbler (Napothera brevicaudata) in a tropical limestone forest of southern China. Avian Res. 8, 28. doi: 10.1186/s40657-017-0086-1
Jong P., Barreto T. B., Tanajura C. A. S., Kouloukoui D., Oliveira-Esquerre K. P., Kiperstok A., et al. (2019). Estimating the impact of climate change on wind and solar energy in Brazil using a South American regional climate model. Renew. Energy 141, 390–401. doi: 10.1016/j.renene.2019.03.086
Kershner E. L., Walk J. W., Warner R. E. (2004). Breeding-season decisions, renesting, and annual fecundity of female eastern meadowlarks (Sturnella magna) in southeastern Illinois. Auk 121, 796–805. doi: 10.1093/auk/121.3.796
Lee S. H., Chang Y. S. (2019). Classification of the global tidal types based on auto-correlation analysis. Ocean Sci. 52, 279–286. doi: 10.1007/s12601-019-0009-7
Lenth R. V. (2024). emmeans: Estimated marginal means, aka least-squares means (R package, version 1.10.4). Available at: https://cran.r-project.org/package=emmeans.
Ma L., Zhang J., Liu J., Yang C., Liang W., Møller A. P. (2019). Adaptation or ecological trap? Altered nest-site selection by Reed Parrotbills after an extreme flood. Avian Res. 10, 2. doi: 10.1186/s40657-019-0141-1
MaBoudi H., Solvi C., Chittka L. (2020). Bumblebees learn a relational rule but switch to a win-stay/lose-switch heuristic after extensive training. Front. Behav. Neurosci. 14. doi: 10.3389/fnbeh.2020.00137
Martin T. E., Roper J. J. (1988). Nest predation and nest-site selection of a western population of the hermit thrush. Condor 90, 51–57. doi: 10.2307/1368432
Marzluff J. M. (1988). Do Pinyon Jays alter nest placement based on prior experience? Anim. Behav. 36, 1–10. doi: 10.1016/S0003-3472(88)80244-6
Meyer A. L. S., Pie M. R. (2022). Climate change estimates surpass rates of climatic niche evolution in primates. Int. J. Primatol. 43, 40–56. doi: 10.1007/s10764-021-00253-z
Oniki Y. (1979). Is nesting success of birds low in the tropics? Biotropica 11, 60–69. doi: 10.2307/2388174
Palomino J. J., Martín-Vivaldi M., Soler M., Soler J. J. (1998). Functional significance of nest size variation in the rufous bush robin Cercotrichas galactotes. Ardea 86, 177–185.
Parejo D., Avilés J. M. (2010). Predation risk determines breeding territory choice in a mediterranean cavity-nesting bird community. Oecologia 165, 185–191. doi: 10.1007/s00442-010-1723-0
R Core Team (2024). R: A language and environment for statistical computing (Vienna: R Foundation for Statistical Computing).
Refsnider J. M., Janzen F. J. (2012). Behavioral plasticity may compensate for climate change in a long-lived reptile with temperature-dependent sex determination. Biol. Conserv. 152, 90–95. doi: 10.1016/j.biocon.2012.03.019
Reinert S. E. (2006). Avian nesting response to tidal-marsh flooding: literature review and a case for adaption in the Red-winged blackbird. Stud. Avian Biol. 32, 77–95.
Reinert B. L. (2008). Ecologia e comportamento do bicudinho-do-brejo (Stymphalornis acutirostris Bornschein, Reinert e Teixeira 1995 – Aves, Thamnophilidae). Universidade Estadual Paulista, Rio Claro (SP).
Reinert B. L., Belmonte-Lopes R., Bornschein M. R., Sobotka D. D., Corrêa L., Pie M. R., et al. (2012). Nest and eggs of the Marsh Antwren (Stymphalornis acutirostris): the only marsh-dwelling Thamnophilid. Wilson J. Ornithol. 124, 286–291. doi: 10.1676/11-099.1
Reinert B. L., Bornschein M. R., Firkowski C. (2007). Distribuição, tamanho populacional, hábitat e conservação do bicudinho-do-brejo Stymphalornis acutirostris Bornschein, Reinert e Teixeira 1995 (Thamnophilidae). Rev. Bras. Ornitol. 15, 493–519.
Robin X., Turck N., Hainard A., Tiberti N., Lisacek F., Sanchez J.-C., et al. (2023). pROC: Display and analyze ROC curves (R package, version 1.18.5). Available at: https://cran.r-project.org/package=pROC.
Sandretti-Silva G. (2024). Possível aprendizado em ninhadas sucessivas e influência de variações ambientais na dinâmica populacional de uma ave de brejos salinos do bioma Floresta Atlântica. Universidade Estadual Paulista, São Vicente (SP).
Sandretti-Silva G., Vannucchi F. S., Teixeira L., Tan T. Y., Mori G. M., Reinert B. L., et al. (2024). Short-term extinction predicted by population viability analysis for a Neotropical salt marsh endemic bird. Environ. Monit. Asses. 196, 520. doi: 10.1007/s10661-024-12618-x
Shriver W. G., Vickery P. D., Hodgman T. P., James P. G. (2007). Flood tides affect breeding ecology of two sympatric Sharp-tailed Sparrows. Auk 142, 552–560. doi: 10.1093/auk/124.2.552
Sobotka D. D. (2011). Comportamento de casais e estudo de paternidade em uma parcela da população de bicudinhos-do-brejo (Stymphalornis acutirostris – Thamnophilidae – Aves), em Guaratuba, Paraná, Brasil. Universidade Federal do Paraná, Curitiba (PR.
Suárez F., Morales M. B., Minguez I., Herranz J. (2005). Seasonal variation in nest mass and dimensions in an open-cup ground-nesting shrub-steppe passerine: the tawny pipit Anthus campestris. Ardeola 52, 43–51.
Taylor P. B. (1996). “Family Rallidae (Rails, gallinules and coots),” in Handbook of the birds of the World, Volume 3, Hoatzin to auk. Eds. Hoyo J.d., Elliott A., Christie D. A. (Lynx Edicions, Barcelona), 138–242.
Tiner R. W. (2013). Tidal wetlands primer: an introduction to their ecology, natural history, status, and conservation (Amherst and Boston: University of Massachusetts Press).
Veloso H. P., Rangel-Filho A. L., Lima J. C. A. (1991). Classificação da vegetação brasileira, adaptada a um sistema universal (Rio de Janeiro: Fundação Instituto Brasileiro de Geografia e Estatística).
Winkler D. W. (2016). “Breeding biology of birds,” in Handbook of bird biology. Eds. Lovette I. J., Fitzpatrick J. W. (Cornell Laboratory of Ornithology, Oxford).
Winkler D. W., Billerman S. M., Lovette I. J. (2020). “Typical antbirds (Thamnophilidae),” in Handbook of bird biology. Eds. Lovette I. J., Fitzpatrick J. W. (Cornell Laboratory of Ornithology, Oxford).
Keywords: conservation, flooding, Formicivora acutirostris, marsh stratification, predation, reproductive success
Citation: Sandretti-Silva G, Corrêa L, Amirati M, Laurino IRA, Passos FC, Pie MR and Bornschein MR (2024) The win-stay, lose-switch renesting strategy of a territorial bird endemic to subtropical salt marshes. Front. Ecol. Evol. 12:1497317. doi: 10.3389/fevo.2024.1497317
Received: 16 September 2024; Accepted: 07 November 2024;
Published: 09 December 2024.
Edited by:
Anindita Bhadra, Indian Institute of Science Education and Research Kolkata, IndiaReviewed by:
Juliana M. Berbert, Federal University of ABC, BrazilDarren O’Connell, University College Dublin, Ireland
Copyright © 2024 Sandretti-Silva, Corrêa, Amirati, Laurino, Passos, Pie and Bornschein. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giovanna Sandretti-Silva, Z2lvdmFubmEuc2FuZHJldHRpQHVuZXNwLmJy