 Víctor M. Escobedo
Víctor M. Escobedo Persy Gómez1
Persy Gómez1- 1Centro de Ecología Integrativa, Instituto de Ciencias Biológicas, Universidad de Talca, Talca, Chile
- 2Dirección de Investigación, Vicerrectoría Académica, Universidad de Talca, Talca, Chile
- 3Centro de Investigación en Estudios Avanzados del Maule (CIEAM), Universidad Católica del Maule, Talca, Chile
Coastal Maulino Forest, a biodiversity hotspot, is increasingly threatened by frequent and higher-severity wildfires. Endangered tree species, including Nothofagus spp., inhabit small, isolated native forest fragments surrounded by extensive Pinus radiata plantations, a non-native species that often colonises fire-affected areas. However, the seedling density of the native Chilean wineberry, Aristotelia chilensis, negatively correlates with the abundance of P. radiata seedlings in post-fire areas. This pattern emerged across areas burned at varying fire severities, sampled 8 and 24 months following the 2017 “Las Máquinas” megafire in Chile. The high proportion of plots lacking P. radiata seedlings, coupled with this negative relationship, suggests that A. chilensis may play a role in limiting P. radiata invasion. The negative relationship was most pronounced in areas with moderate fire severity, likely reflecting differences in shade tolerance between the species. While A. chilensis, a light-demanding species with some shade tolerance, can persist in partially shaded environments, P. radiata, a strictly light-demanding species, struggles under significant shade. In low-severity areas, no significant relationship was observed since the substantial native canopy remaining likely limits P. radiata establishment by shading. Conversely, in high-severity fire areas, the absence of a significant relationship might result from the detrimental effects on both species, including potential microbiome dependence for A. chilensis. Given the successful establishment of A. chilensis at low fire severity, enhancing its post-fire recruitment, particularly in moderately burned areas, could be a valuable strategy for mitigating P. radiata invasion and restoring fire-affected Mediterranean ecosystems.
Introduction
Wildfires pose a significant threat to biodiversity, disrupting ecosystem functions and threatening sensitive habitats worldwide. Their increased frequency and severity are attributed to various factors, including climate change and land-use modifications (McLauchlan et al., 2020). The Coastal Maulino Forest, a biodiversity hotspot in central Chile (Myers et al., 2000), has experienced more frequent and higher-severity wildfires in recent decades, driven by rising temperatures, a megadrought, and extensive forestry plantations of non-native species (González et al., 2018, 2023), some of which become invasive after fire disturbances. These fires are particularly concerning for the survival of endangered species such as Nothofagus alessandrii (Nothofagaceae) and N. glauca, which are confined to small, isolated forest fragments surrounded by vast Pinus radiata (Pinaceae) plantations. The invasive P. radiata, which covers approximately 60% of the country’s 2.5 million hectares of forest plantations (Bustamante and Simonetti, 2005; González et al., 2018), often rapidly colonises post-fire areas, further endangering these native species. The devastating 2017 “Las Máquinas” megafire burned over 200,000 ha of the Coastal Maulino Forest, underscoring the vulnerability of this ecosystem (Valencia et al., 2018). Despite ongoing active and passive restoration efforts in south-central Chile (Morales et al., 2021; Souza-Alonso et al., 2022), challenges persist, including the rapid arrival of post-fire pine regeneration that hinders restoration success (Gómez et al., 2019; González et al., 2020, 2023). This highlights the need for conservation and restoration practices tailored to this unique ecosystem.
Invasive species often display rapid resource utilisation, potentially outcompeting native species and promoting more frequent fire events. This can create a positive invasion-fire feedback loop (Contreras et al., 2011; Taylor et al., 2017). P. radiata is a light-demanding and shade-intolerant species known for its aggressive post-fire regeneration through serotinous cones, which release large amounts of viable wind-dispersed seeds after fire events (Franzese and Raffaele, 2017; Gómez and Garrido, 2018). Studies have shown a higher probability of fire ignition in areas dominated by P. radiata plantations compared to native forests in south-central Chile (Contreras et al., 2011; Gómez-González et al., 2019).
Previous research suggests limited success in controlling P. radiata invasion through overall native species diversity (Gómez et al., 2019; González et al., 2020). However, recent field studies provide evidence that the native wineberry species Aristotelia chilensis (Elaeocarpaceae) can efficiently recolonise burnt areas, often where P. radiata seedlings are sparse or absent (Promis et al., 2019; Becerra et al., 2022). A. chilensis is a semi-dioecious, fast-growing, light-demanding, fleshy-fruiting, bird-dispersed, evergreen small tree species. These traits allow it to not only colonise clearings but also persist after plantations replace native forests because it can exhibit some shade tolerance (Guerra et al., 2010; Salgado-Luarte and Gianoli, 2012). This rapid establishment and fast growth of A. chilensis would align with the concept of the “pre-emptive resource effect” – a mechanism where early colonising native species can outcompete invasive plants by monopolising essential resources (Byun et al., 2013; Byun and Lee, 2017; Delavaux et al., 2023). Additionally, studies suggest that P. radiata, being a shade-intolerant species, might struggle to establish into a darker understory dominated by A. chilensis and other native species (Gómez et al., 2019; Becerra and Simonetti, 2020). The efficient colonisation and fast growth of A. chilensis suggest that it has the potential to act as a native plant competitor against P. radiata invasion in fire-affected ecosystems.
Building upon competition-based biotic resistance (Elton, 1958) and the theory of limiting similarity, where native species can limit invasive plant establishment due to niche overlap, we hypothesised that P. radiata abundance would negatively correlate with increasing A. chilensis abundance. Specifically, we tested the relationship between the abundance of A. chilensis and P. radiata in plots affected by varying fire severity levels caused by the Las Máquinas mega-fire in the Coastal Maulino Forest. Additionally, we explored whether fire severity modulates this relationship. Moderate- or low-severity fires that increase light penetration while retaining understory vegetation could favour A. chilensis establishment, potentially strengthening its competitive effect on P. radiata (i.e., a negative relationship). In contrast, high-severity fires that create harsher conditions and potential soil disruption (i.e., depleting the soil microbiome) could hinder the establishment of both A. chilensis and P. radiata, obscuring any competitive effects. By elucidating these dynamics, we aim to provide valuable data to guide and enhance conservation and restoration efforts in fire-affected areas across the central Mediterranean region of Chile.
Materials and methods
Study site
The study was conducted at El Porvenir (35°42’ S, 72°22’ W), located at the northern edge of the Coastal Maulino Forest in central-south Chile (Gómez et al., 2022). El Porvenir is a fragment of native mesic forest type, surrounded by large stands of planted P. radiata and Eucalyptus globulus (Myrtaceae). The dominant tree species include Nothofagus glauca (Nothofagaceae), N. alessandrii, N. obliqua, Cryptocarya alba (Lauraceae), Aextoxicon punctatum (Aextoxicaceae), Gevuina avellana (Proteaceae), and A. chilensis. The study area has a Mediterranean climate with a mean annual precipitation of 918 mm and a mean annual temperature of 12.7°C (Becerra and Simonetti, 2020).
The total plant species richness at El Porvenir is 104 species, with Asteraceae and Fabaceae being the most representative families. Growth forms within the forest include herbs, shrubs, trees, and climbers. In the understory, in addition to A. chilensis, notable species include Solanum crispum, Carex phleoides, Francoa appendiculata, Polystichum chliensis, Escallonia illinita, Dioscorea brachybotria, Colletia ulicina, Pteris chilensis, Ageratina glechnophylla, Baccharis racemosa, Retanilla ephedra, Tropaeolum ciliatum, Boquila trifoliolata, Pseudognaphalium vira-vira, Epilobium ciliatum, Sanicula crassicaulis, Synammia feullei, Lathyrus cabrenianus, Baccharis concava, Dioscorea humifusa, Sophora macrocarpa, Senna stipulacea and Rhaphythamnus spinosus, among others.
In January 2017, the Las Máquinas megafire affected El Porvenir, which experienced fire severity ranging from low to high (see Valencia et al., 2018; Gómez et al., 2022). Following the fire, several species exhibited regeneration at different levels, with high seedling recruitment for the invasive P. radiata and the native A. chilensis (Gómez et al., 2022).
Fire severity assessment
Fire severity at the study site was estimated from plots established in March 2017 using the Composite Burn Index (CBI), following the protocol of Key and Benson (2006) (see Gómez et al., 2022). In twenty-three 625 m2 plots randomly established across El Porvenir, we evaluated five vertical strata (soil, herbs, shrubs, intermediate trees, and large trees) arranged in a hierarchical structure (De Santis and Chuvieco, 2007). At each stratum, visual attributes were assessed, including changes in colour due to fire, litter and surface fuels, canopy cover alterations, and vegetation mortality (De Santis and Chuvieco, 2009). These attributes were used to generate CBI values for each plot, which were then classified into three severity categories: low (minimal changes in cover and low vegetation mortality, n = 8), moderate (a mix of unchanged and highly altered cover, n = 10), and high (extensive to complete vegetation mortality, n = 5). The mean values of the attributes for each plot were used to compile a composite CBI score.
Plot establishment and seedling survey
To assess the potential role of A. chilensis in limiting P. radiata invasion, we used the previously established twenty-three plots across El Porvenir. As noted in the “Fire severity assessment” section, these plots were randomly established across the forest fragment, covering a range of distances from surrounding pine plantations. Although the distance from pine seed sources was not explicitly analysed, the random establishment ensured a representation of various distances, thereby minimising potential bias in plot location. In each plot, seedling surveys were conducted at 8 and 24 months following the Las Máquinas mega-fire (hereafter 2017 and 2019), meaning the same plots were revisited to assess changes over time. To estimate the density of A. chilensis and P. radiata seedlings, three 1 m2 sub-plots were randomly located within each plot to survey all A. chilensis and P. radiata seedlings that had regenerated (not resprouted) and were under 60 cm in height. The average number of seedlings per 3 m2 sampled area was 21.98 ± 20.34 for A. chilensis and 7.46 ± 12.78 for P. radiata. To confirm that the seedlings originated from seeds and not resprouts, we collected at least three random plant samples per species from each sub-plot for root system examination. While P. radiata is not known to resprout after fire, A. chilensis may exhibit resprouting behaviour (Gómez, 2019), which was accounted for during root examination.
Data analysis
We performed a negative binomial Generalised Linear Mixed-effects Model (NB GLMM) to analyse the relationship between the abundance of A. chilensis and P. radiata seedlings. This statistical method is suitable for counting data with overdispersion, a common characteristic of ecological data. Here, we account for the potential influence of sampling time at each plot by including time since the fire (2017 and 2019) as a random factor nested within fire severity. This nested structure considers the variation in fire severity across the landscape while acknowledging the potential influence of sampling time within each fire severity category (see above). Additionally, we conducted separate NB GLMM analyses for each fire severity level, including time sampling as a random factor.
Results and discussion
Our analysis revealed a negative relationship between Aristotelia chilensis and Pinus radiata abundance across the study site (χ2(1,46) = 8.0707, p < 0.01; Figure 1). Areas with higher numbers of A. chilensis seedlings had fewer P. radiata seedlings, suggesting a potential suppressive effect of native small tree species on the establishment of the invasive tree.
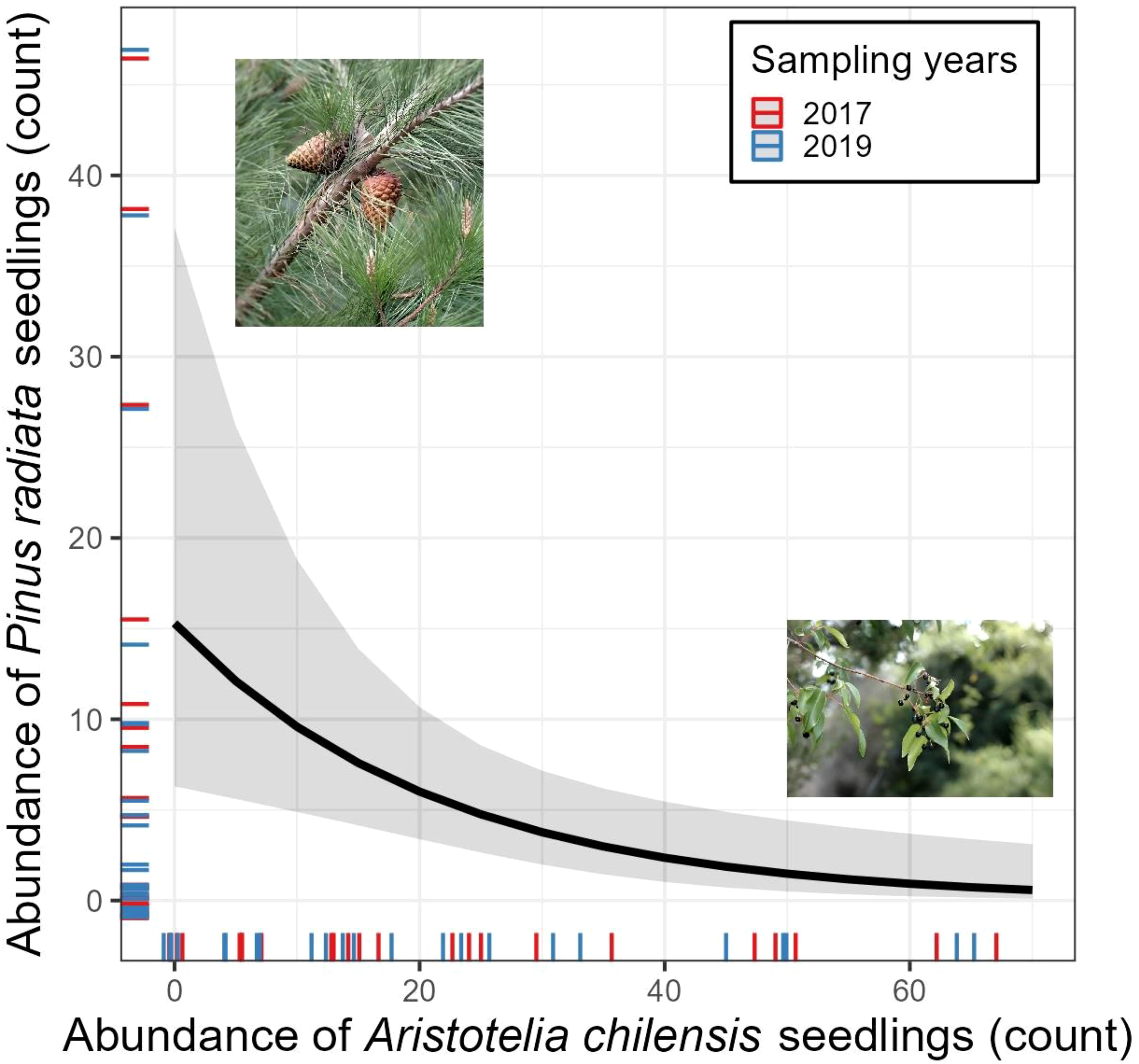
Figure 1. Model-predicted relationship between Aristotelia chilensis and Pinus radiata seedlings abundance for two sampling times (2017 and 2019). Line indicates a statistically significant negative relationship (p < 0.05) based on a negative binomial GLMM.
This negative relationship between A. chilensis and P. radiata was especially pronounced in areas with moderate fire severity (χ2(1,20) = 16.385, p < 0.01; Figure 2), whereas no significant relationship was observed in plots with high or low fire severity (Figure 2). These findings suggest that fire severity may mediate the interaction between A. chilensis and P. radiata, with moderate fire severity favouring the competitive advantage of native species. Fire severity is known to shape post-fire succession and ecosystem dynamics, making it crucial to understand these effects for both effective forest restoration and conservation management strategies.
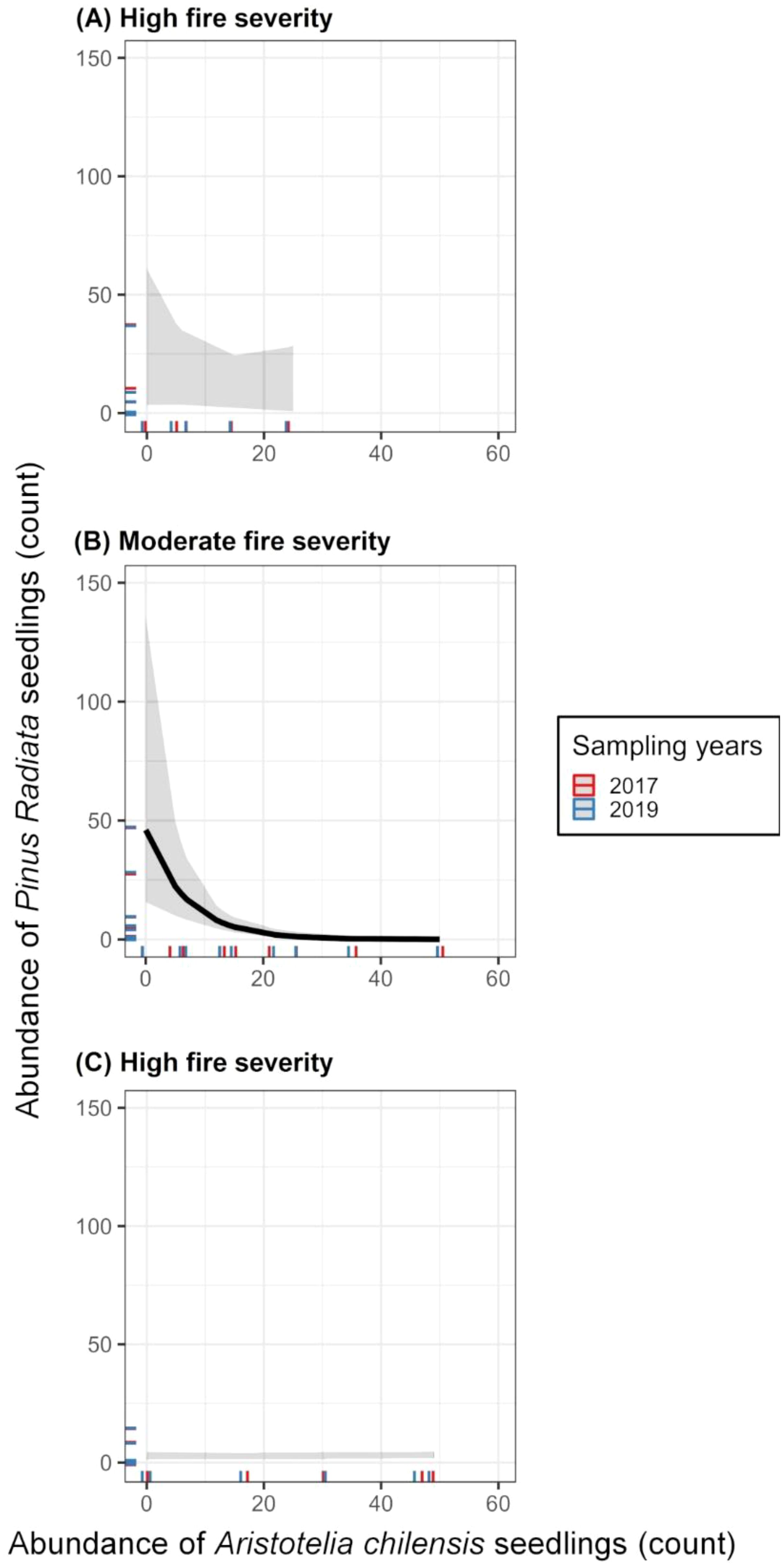
Figure 2. Model-predicted relationships between Aristotelia chilensis and Pinus radiata seedling abundance across fire severity levels [(A), high; (B), moderate; (C), low] for two sampling times (2017 and 2019). The solid line in the middle panel [(B), moderate-severity fire area] indicates a statistically significant negative relationship (p < 0.05) based on a negative binomial GLMM. Relationships were not statistically significant in high- or low-fired-severity areas.
In addition to A. chilensis, other native species, particularly the endangered Nothofagus alessandrii and N. glauca, are acknowledged components of the Coastal Maulino Forest. These species, such as A. chilensis, are part of the pre-fire native flora, and their presence may contribute to post-fire regeneration dynamics. However, both N. alessandrii and N. glauca face particular challenges in regenerating after fires, especially in areas with high-severity burns (Gómez et al., 2022). The severe disruption of soil conditions and the elimination of key microbial communities could limit their recovery (Certini, 2005; Warneke et al., 2023; Escobedo et al., 2024), further threatening the ecological integrity of these fragmented populations (Gómez et al., 2022; González et al., 2023). Given their importance, restoration efforts must consider the regeneration of fast-growing species like A. chilensis, which may support strategies for the recovery of slow-growing, fire-sensitive species such as N. alessandrii and N. glauca.
The observed negative relationship between A. chilensis and P. radiata suggests that the former’s presence, as a component of the pre-fire native flora, may influence P. radiata establishment in post-fire areas. As A. chilensis was already present in these ecosystems before the fires, its abundance at the time of the fire event likely influenced the available resources and habitat conditions for P. radiata establishment. Several mechanisms could explain this, including the priority effect by pre-empting resources and habitat filtering (Byun et al., 2013; Byun and Lee, 2017). Given that A. chilensis is a fast-growing, light-demanding species, it may pre-empt light, water, or nutrients in areas where it is abundant, hindering P. radiata establishment. Future studies quantifying resource availability and seedling performance in relation to A. chilensis density could provide stronger evidence for this hypothesis.
Moreover, fire can have profound and different effects on plant community assembly depending on its severity (McLauchlan et al., 2020). The environmental conditions created by moderate fire severity may be more favourable for the establishment of native compared to invasive species. Specifically, these fires create a more open canopy with increased light availability in the understory, typically forming a patchy mosaic of burned and unburned areas rather than eliminating the entire canopy. While A. chilensis is a light-demanding species with some degree of shade tolerance (Guerra et al., 2010; Salgado-Luarte and Gianoli, 2012), P. radiata is a strictly shade-intolerant species (Gómez et al., 2019). Thus, this variation in light availability could favour A. chilensis over P. radiata establishment in suitable microsites within the burned landscape. In low-severity fires with more remaining canopy cover, P. radiata pine showed very low establishment (only two plots with 9 and 15 seedlings), likely due to limited light availability for germination and seedling growth. In contrast, A. chilensis, which can exhibit some shade tolerance, could persist and thrive, with an average of 16 seedlings per plot and up to 67 in one case (Figure 2). Consequently, the lingering native canopy foliage after low-severity fires probably limits light availability, thus restricting the establishment of P. radiata seedlings.
High-severity fires present a vastly different scenario because nearly all vegetation is fire-consumed and heat sterilises the soil, eliminating vital microbes and severely disrupting biogeochemical processes. In these harsh conditions, the establishment of A. chilensis and P. radiata is hindered, resulting in the lack of relationship observed in Figure 2. In this line, A. chilensis’s lower establishment suggests a dependence on healthy soil microbes (Escobedo et al., 2024), which are eliminated by high-severity fires. P. radiata, meanwhile, sometimes showed higher abundance in these areas, potentially benefiting from the transiently increased nutrient availability and A. chilensis-free conditions since its establishment and survival are less reliant upon microbe communities (Escobedo et al., 2024). This resilience disparity in high-severity fire areas poses a significant threat to the already vulnerable Coastal Maulino Forest, especially for endangered species such as N. alessandrii and N. glauca that are less adapted to such extreme and changing conditions. Although the general trends observed in areas of high fire severity are consistent with our expectations, we acknowledge that the small number of high-severity plots (n=5) may limit the statistical power to detect finer-scale interactions between A. chilensis and P. radiata in these areas. This limitation should be addressed in future studies with a larger number of high-severity plots to confirm the patterns observed. Furthermore, more extensive sampling could improve our understanding of how fire severity impacts post-fire recovery dynamics in native and invasive species.
To expand on the implications of long-term competition dynamics at the Coastal Maulino Forest, it is important to consider that the competitive interactions between A. chilensis and P. radiata may shift over time. While our study captured early successional processes in the first two years post-fire, we speculate that as A. chilensis continues to establish and grow, it may increasingly outcompete P. radiata in the coming years due to its superior shade tolerance. Conversely, in areas where P. radiata manages to establish initially, its rapid growth may enable it to outcompete A. chilensis over time. Active management strategies that include the removal of P. radiata seedlings in the early stages, combined with the promotion of A. chilensis, could provide a highly effective restoration approach. Restoration techniques promoting A. chilensis recruitment—such as direct seedling planting with native soil microbiomes, boosting seed dispersal, and optimizing environmental factors like light and soil conditions—could be pivotal in encouraging its establishment post-fire. This dual strategy may not only suppress invasive P. radiata populations but also support the long-term recovery of native ecosystems, fostering the establishment of fire-sensitive species like N. alessandrii and N. glauca. Long-term monitoring of these interactions would be essential to confirm whether the trends observed in the short term persist or evolve in different fire severity scenarios.
While our study focused on the Las Máquinas 2017 megafire, the findings possess broader implications beyond the Coastal Maulino Forest, particularly in regions experiencing increased fire frequencies where P. radiata is invasive. Similar interactions between native and invasive species have emerged in other Mediterranean-type ecosystems, such as those in California (Dimitrakopoulos and Papaioannou, 2001; Alexander and D’Antonio, 2003) or Australia (Grice, 1997), and South Africa (Van Wilgen and Richardson, 1985; Holmes et al., 2000), where variable fire frequencies and species compositions influence competition dynamics. In areas such as Hawaii, Australia, and New Zealand (Mandle et al., 2011; Hynson et al., 2013), the spread of P. radiata and resultant fire-induced disturbances present parallel challenges. These comparisons underscore the necessity of factoring ecosystem-specific characteristics into our understanding of post-fire recovery. The competitive dynamics observed between native species such as A. chilensis and P. radiata could inform management strategies aimed at controlling pine invasions across diverse regions. Although our study concentrated on the Coastal Maulino Forest, the mechanisms of native species pre-empting resources and moderating invasive species establishment (e.g., competition for resources, shading effects) might be applicable to other ecosystems with similar invasion and fire dynamics (e.g., Grace, 1998; Brooks et al., 2004). Further research is essential to comprehensively understand how native species can be leveraged to manage P. radiata invasions effectively in varying fire-prone environments.
Conclusions
Our findings suggest that promoting the establishment of native species like A. chilensis, particularly in areas with moderate fire severity, could help limit the invasion of P. radiata. Thus, restoration strategies might consider A. chilensis as a targeted support species for the recovery of fire-sensitive species such as N. alessandrii and N. glauca, which are crucial for maintaining the ecological integrity of the Coastal Maulino Forest. Moreover, this study highlights potential applications beyond this region, offering insights into managing P. radiata invasions in other fire-prone ecosystems worldwide. Efforts focused on enhancing post-fire recruitment of A. chilensis, especially in areas of moderate fire severity, could offer a pathway to mitigating invasive tree spread and promoting ecosystem recovery in this unique biodiversity hotspot.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: https://doi.org/10.6084/m9.figshare.27640251.v1.
Author contributions
VE: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. PG: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Validation, Writing – review & editing. MM: Conceptualization, Resources, Supervision, Validation, Writing – review & editing. IA: Funding acquisition, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. PG was supported by the Global Botanic Garden Fund number 2022/022 (Botanic Gardens Conservation International, BGCI). IA-R was supported by ANID-FONDECYT grant 11240628. We would also like to thank the Parcelación Agrícola de Conservación El Secreto de Pilén for providing us with a cabin and the essential supplies necessary for our field activities.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alexander J. M., D’Antonio C. M. (2003). Control methods for the removal of french and scotch broom tested in coastal California. Ecol. Restor. 21, 191. doi: 10.3368/er.21.3.191
Becerra P. I., Figueroa C., Meza A. (2022). Dinámica post-incendio de la vegetación en la localidad de Rastrojos, Chile central. Gayana Bot. 79, 10–26. doi: 10.4067/S0717-66432022000100010
Becerra P. I., Simonetti J. A. (2020). Native and exotic plant species diversity in forest fragments and forestry plantations of a coastal landscape of central Chile. Bosque 41, 125–136. doi: 10.4067/S0717-92002020000200125
Brooks M. L., D’Antonio C. M., Richardson D. M., Grace J. B., Keeley J. E., Ditomaso J. M., et al. (2004). Effects of invasive alien plants on fire regimes. BioScience 54, 677. doi: 10.1641/0006-3568(2004)054[0677:EOIAPO]2.0.CO;2
Bustamante R. O., Simonetti J. A. (2005). Is Pinus radiata invading the native vegetation in central Chile? Demographic responses in a fragmented forest. Biol. Invasions 7, 243–249. doi: 10.1007/s10530-004-0740-5
Byun C., Lee E. J. (2017). Ecological application of biotic resistance to control the invasion of an invasive plant, Ageratina altissima. Ecol. Evol. 7, 2181–2192. doi: 10.1002/ece3.2799
Byun C., de Blois S., Brisson J. (2013). Plant functional group identity and diversity determine biotic resistance to invasion by an exotic grass. J. Ecol. 101, 128–139. doi: 10.1111/1365-2745.12016
Certini G. (2005). Effects of fire on properties of forest soils: a review. Oecologia 143, 1–10. doi: 10.1007/s00442-004-1788-8
Contreras T., Figueroa J., Abarca L., Castro S. (2011). Fire regimen and spread of plants naturalized in central Chile. Rev. Chil. Hist. Nat. 84, 307–323. doi: 10.4067/S0716-078X2011000300001
Delavaux C. S., Crowther T. W., Zohner C. M., Robmann N. M., Lauber T., van den Hoogen J., et al. (2023). Native diversity buffers against severity of non-native tree invasions. Nature 621, 773–781. doi: 10.1038/s41586-023-06440-7
De Santis A., Chuvieco E. (2007). Burn severity estimation from remotely sensed data: Performance of simulation versus empirical models. Remote Sens. Environ. 108, 422–435. doi: 10.1016/j.rse.2006.11.022
De Santis A., Chuvieco E. (2009). GeoCBI: A modified version of the Composite Burn Index for the initial assessment of the short-term burn severity from remotely sensed data. Remote Sens. Environ. 113, 554–562. doi: 10.1016/j.rse.2008.10.011
Dimitrakopoulos A. P., Papaioannou K. K. (2001). Flammability assessment of mediterranean forest fuels. Fire Technol. 37, 143–152. doi: 10.1023/A:1011641601076
Elton C. S. (1958). The ecology of invasions by animals and plants (Boston: Springer US). doi: 10.1007/978-1-4899-7214-9
Escobedo V. M., Acuña-Rodríguez I. S., García L., Torres-Diáz C., Atala C., Suazo M., et al. (2024). Native woody species depend on the soil microbiome to establish on burned soils, while non-native do not. J. Appl. Ecol. doi: 10.1111/1365-2664.14809
Franzese J., Raffaele E. (2017). Fire as a driver of pine invasions in the Southern Hemisphere: a review. Biol. Invasions 19, 2237–2246. doi: 10.1007/s10530-017-1435-z
Gómez P. (2019). “Diversidad florística en los bosques de ruil,” in Ruil (Nothofagus alessandrii): biodiversidad de sus bosques (Chile: Universidad de Talca), 41–93.
Gómez P., Espinoza S., Garrido P., Martín J. S., Ormazábal Y. (2022). Post-fire tree regeneration from seed of the endangered Nothofagus alessandrii Espinosa in the Maule region of central Chile. South. Forests: J. For. Sci. 84, 75–82. doi: 10.2989/20702620.2022.2039044
Gómez P., Garrido P. (2018). Regeneración masiva del árbol exotico Pinus radiata D. Don luego de un gran incendio en fragmentos de Nothofagus alessandrii (Ruil) en la Región del Maule, Chile central. Bioinvasiones 7, 30–38.
Gómez P., Murúa M., San Martín J., Goncalves E., Bustamante R. O. (2019). Maintaining close canopy cover prevents the invasion of Pinus radiata: Basic ecology to manage native forest invasibility. PloS One 14, e0210849. doi: 10.1371/journal.pone.0210849
Gómez-González S., González M. E., Paula S., Díaz-Hormazábal I., Lara A., Delgado-Baquerizo M. (2019). Temperature and agriculture are largely associated with fire activity in Central Chile across different temporal periods. For. Ecol. Manage. 433, 535–543. doi: 10.1016/j.foreco.2018.11.041
González M. E., Galleguillos M., Lopatin J., Leal C., Becerra-Rodas C., Lara A., et al. (2023). Surviving in a hostile landscape: Nothofagus alessandrii remnant forests threatened by mega-fires and exotic pine invasion in the coastal range of central Chile. Oryx 57, 228–238. doi: 10.1017/S0030605322000102
González M. E., Gómez-González S., Lara A., Garreaud R., Díaz-Hormazábal I. (2018). The 2010–2015 Megadrought and its influence on the fire regime in central and south-central Chile. Ecosphere 9, e02300. doi: 10.1002/ecs2.2300
González M. E., Sapiains R., Gómez-González S., Garreaud R., Miranda A., Galleguillos M., et al. (2020). Incendios forestales en Chile: causas, impactos y resiliencia (Chile: Centro de Ciencia del Clima y la Resiliencia). Available at: www.cr2.cl.
Grace J. B. (1998). Can prescribed fire save the endangered coastal prairie ecosystem from chinese tallow invasion? Endangered Species UPDATE 15, 70–91.
Grice A. C. (1997). Post-fire regrowth and survival of the invasive tropical shrubs Cryptostegia grandiflora and Ziziphus mauritiana. Aust. J. Ecol. 22, 49–55. doi: 10.1111/j.1442-9993.1997.tb00640.x
Guerra P. C., Becerra J., Gianoli E. (2010). Explaining differential herbivory in sun and shade: The case of Aristotelia Chilensis saplings. Arthropod-Plant Interact. 4, 229–235. doi: 10.1007/s11829-010-9099-y
Holmes P. M., Richardson D. M., Van Wilgen B. W., Gelderblom C. (2000). Recovery of South African fynbos vegetation following alien woody plant clearing and fire: implications for restoration. Austral Ecol. 25, 631–639. doi: 10.1111/j.1442-9993.2000.tb00069.x
Hynson N. A., Merckx V. S. F. T., Perry B. A., Treseder K. K. (2013). Identities and distributions of the co-invading ectomycorrhizal fungal symbionts of exotic pines in the Hawaiian Islands. Biol. Invasions 15, 2373–2385. doi: 10.1007/s10530-013-0458-3
Key C. H., Benson N. C. (2006). “Landscape assessment (LA) sampling and analysis methods,” in FIREMON: Fire effects monitoring and inventory system, (U.S. Department of Agriculture (Forest Service, Rocky Mountain Research Station, Fort Collins, CO, USA), 1–55.
Mandle L., Bufford J. L., Schmidt I. B., Daehler C. C. (2011). Woody exotic plant invasions and fire: reciprocal impacts and consequences for native ecosystems. Biol. Invasions 13, 1815–1827. doi: 10.1007/s10530-011-0001-3
McLauchlan K. K., Higuera P. E., Miesel J., Rogers B. M., Schweitzer J., Shuman J. K., et al. (2020). Fire as a fundamental ecological process: research advances and frontiers. J. Ecol. 108, 2047–2069. doi: 10.1111/1365-2745.13403
Morales N. S., Fernández I. C., Duran L. P., Venegas-González A. (2021). Community-driven post-fire restoration initiatives in Central Chile: when good intentions are not enough. Restor. Ecol. 29, e13389. doi: 10.1111/rec.13389
Myers N., Mittermeier R. A., Mittermeier C. G., da Fonseca G. A. B., Kent J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Promis A., Olivares S., Acuña S., Cruz G. (2019). Respuesta temprana de la regeneración de plantas leñosas después del incendio forestal denominado “Las Máquinas” en la Región del Maule, Chile. Gayana Bot. 76, 257–262. doi: 10.4067/S0717-66432019000200257
Salgado-Luarte C., Gianoli E. (2012). Herbivores modify selection on plant functional traits in a temperate rainforest understory. Am. Nat. 180, E42–E53. doi: 10.1086/666612
Souza-Alonso P., Saiz G., García R. A., Pauchard A., Ferreira A., Merino A. (2022). Post-fire ecological restoration in Latin American forest ecosystems: insights and lessons from the last two decades. For. Ecol. Manage. 509, 120083. doi: 10.1016/j.foreco.2022.120083
Taylor K., Maxwell B., McWethy D., Pauchard A., Nunez M., Whitlock C. (2017). Pinus contorta invasions increase wildfire fuel loads and may create a positive feedback with fire. Ecology 98, 678–687. doi: 10.1002/ecy.1673
Valencia D., Saavedra J., Brull J., Santelices R. (2018). Severidad del daño causado por los incendios forestales en los bosques remanentes de Nothofagus alessandrii Espinosa en la Región del Maule de Chile. Gayana. Botánica 75, 531–534. doi: 10.4067/S0717-66432018000100531
Van Wilgen B. W., Richardson D. M. (1985). Factors influencing burning by prescription in mountain fynbos catchment Aareas. South Afr. Forestry J. 134, 22–32. doi: 10.1080/00382167.1985.9629590
Keywords: invasion resistance, fire severity, Coastal Maulino Forest, post-fire establishment, soil microbiome
Citation: Escobedo VM, Gómez P, Molina-Montenegro MA and Acuña-Rodríguez IS (2024) Post-fire negative relationship between a native tree and an invasive pine at the Coastal Maulino Forest in Central Chile. Front. Ecol. Evol. 12:1494548. doi: 10.3389/fevo.2024.1494548
Received: 11 September 2024; Accepted: 30 October 2024;
Published: 11 December 2024.
Edited by:
Emilio Badalamenti, University of Palermo, ItalyReviewed by:
Alison Ainsworth, United States Department of the Interior, United StatesHelena Freitas, University of Coimbra, Portugal
Copyright © 2024 Escobedo, Gómez, Molina-Montenegro and Acuña-Rodríguez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Víctor M. Escobedo, dmljdG9yLmVzY29iZWRvQHV0YWxjYS5jbA==; Ian S. Acuña-Rodríguez, aWFuLmFjdW5hQHV0YWxjYS5jbA==