 Di Huang
Di Huang Yihua Ren1,2,3,4,5*
Yihua Ren1,2,3,4,5*- 1Institute of Tibet Plateau Ecology, Tibet Agricultural and Animal Husbandry University, Nyingchi, China
- 2National Forest Ecosystem Observation and Research Station of Tibet, Nyingchi, China
- 3Key Laboratory of Forest Ecology in Tibet Plateau, Ministry of Education, Nyingchi, China
- 4Key Laboratory of Alpine Vegetation Ecological Security in Tibet, Nyingchi, China
- 5Nyingchi Ecological Monitoring Station, Nyingchi, China
- 6Animal Science College, Tibet Agriculture and Animal Husbandry University, Nyingchi, China
Introduction: In the face of escalating climate change, conserving fir forests in southeastern Tibet is paramount. These forests uphold Tibetan biodiversity and support crucial ecological services.
Methods: This study used the typical plot method to investigate the survival status and development trends of three Chinese endemic Abies species, namely, Abies georgei var. smithii, Abies chayuensis, and Abies delavayi var. motuoensis, from the diameter structure, static life table, and survival curve.
Results: The results revealed that 1) the density of Abies georgei var. smithii, Abies chayuensis, and Abies delavayi var. motuoensis in the community is 408, 320, and 103 trees per hectare, respectively. The structure of the diameter class of the three species of fir trees is a pyramid shape. As the diameter class increases, the number of each population fluctuates to varying degrees, and there is a lack of many medium- and large-diameter trees. 2) The survival curve of Abies georgei var. smithii tends to be similar to the Deevey type II curves, whereas the survival curves of Abies chayuensis and Abies delavayi var. motuoensis tend to be similar to the Deevey type III curves. Various population structures are growth-oriented but highly sensitive to external random disturbances, with mortality and disappearance rates multiple times, indicating poor population development. Survival analysis reveals that all populations begin to decline after the middle age range and that the age structure is relatively unstable.
Discussion: The results of this study contribute to revealing the structural characteristics and dynamic patterns of various spruce populations, providing a theoretical basis for the management and sustainable utilization of fir forests in southeastern Tibet.
1 Introduction
The structural characteristics and population dynamics of plant populations are among the core areas of plant ecology research (Lei et al., 2022). Combining static life tables, survival analysis, and quantitative analysis of population dynamics can reflect the fit between plants and the environment, as well as plant survival strategies and situations of disturbance (He et al., 2024). Understanding the response strategies of populations to the current environment, elucidating the ecological processes of populations, and displaying the dynamic characteristics of population numbers are highly important (Li and Zhang, 2015). Abies is the second largest genus in the Pinaceae family, with approximately 50 species worldwide. It is intermittently distributed in high-altitude areas in temperate and subtropical high mountainous regions in the Northern Hemisphere and is strongly shade-tolerant. It can adapt to warm and cold climate conditions. According to the Flora of China (Fu et al., 1999), 28 species and 7 varieties of Abies trees are naturally occurring in China, making it the country with the richest variety of Abies plants in the world. Tibet has one of the key state-owned forest areas in China, and its strategic ecological position is particularly critical. Nyingchi city, located in southeastern Tibet, is the core area of Tibet’s forests. This area has superior geographical and climatic conditions, diverse forests, and abundant natural resources. As one of the main constructive species of Southeast Tibet’s forests, Abies often exists in extensive contiguous pure forests and mixed forests in submountains and valleys, playing an essential role in maintaining the ecological balance, protecting water sources, and promoting biodiversity.
In recent years, due to global warming, human activities, natural disasters, and other reasons, the main forest layer of Abies fir forest in southeast Tibet has declined, the forest structure has become unbalanced, the function of the system has degraded, and the natural area of distribution has been shrinking daily, posing a significant threat to the construction of ecological security system in Tibet (Lu et al., 2010). As rare species unique to China, the Abies georgei var. smithii, Abies chayuensis, and Abies delavayi var. motuoensis not only play an irreplaceable role in biodiversity conservation, but also play a crucial role in maintaining regional ecological balance and promoting ecosystem service functions. Their unique ecological habits, growth strategies, and adaptive characteristics are of great significance for understanding the southeastern Tibet region’s ecological processes and climate change responses. Therefore, protection of the natural fir forest in southeastern Tibet is particularly urgent. Many scholars have conducted a series of studies on the single largest pure forest of Abies georgei var. smithii in southeastern Tibet, and these studies have focused on the microbial community structure (Fu et al., 2023), functional traits of seedlings (Zhang et al., 2022), community spatial pattern (Zhu et al., 2022), and other aspects. These studies help us understand this population’s ecological characteristics, growth habits, and distribution patterns, which are valuable for population restoration and species protection. However, Abies fabri forests are widely distributed in southeastern Tibet, and existing research results cannot be used to accurately and comprehensively ascertain the survival status and trend of development of Abies fabri forests in southeastern Tibet.
On the basis of this insufficiency of previous results, in this study, three Chinese endemic fir populations, namely, Abies georgei var. smithii, Abies chayuensis, and Abies delavayi var. motuoensis, were investigated via traditional plot survey methods. From the perspective of population structure and dynamics, the following questions were explored: (1) is the age structure of the main fir populations in southeastern Tibet stable, and is the future population structure trending toward decline or growth? (2) Is there any difficulty in natural regeneration under the forest? Moreover, the mechanisms of population renewal and stability maintenance were explored to provide a theoretical basis and data for managing and protecting the Abies forest ecosystem in the region.
2 Materials and methods
2.1 Study site
The distribution area of Abies fabri forest in southeastern Tibet was determined through the Plan for the Protection and Construction of Ecological Security Barriers in Tibet approved by the State Council in 2009 and field surveys. The research area consisted of the three main distribution areas of Abies fabri in southeastern Tibet: Bayi District, Zayu County, and Motuo County in Nyingchi city. These three regions exhibit significant differences and representativeness in ecological types, geographical locations, and climate characteristics. The study area has distinct dry and wet seasons with abundant precipitation, mainly from June to September. The average annual sunshine duration is approximately 2000 hours. The terrain is high in the north and low in the south. The soil is mainly acidic brown soil.
Bayi District (93°27′–95°17′ E, 29°21′–30°15′ N) is part of the Nyenchen Tanggula Mountains, with an average elevation of 3000 meters. This area is within a subalpine temperate semihumid climate zone (Lu et al., 2021), with an average annual precipitation of 654 mm and an average annual temperature of 8.5°C. The tree layer of the research plot is a single Abies georgi var. smithii forest. The species in the shrub layer mainly include Rhododendron faucium, Acer caudamum, Lonicera inconspicua, etc. The herbaceous plants in the shrub layer mainly include Parasenecio quinquelobus, Duchesnea indica, and Streptopus obtumatatus.
Zayu County (97°27′33” E, 28°56′46” N) is part of the high mountain canyon area of southeastern Tibet in the western section of the Hengduan Mountains, with an average elevation of 2800 m and a humid subtropical monsoon climate. The average annual temperature is 12°C, and the average annual precipitation is 801.1 mm (Cai et al., 2024). The main plants in the tree layer of the study site were Abies chayuensis, Pinus armandii, and Betula utilis, while the shrub layer vegetation mainly included Ribes pulchellum, Acer stackophyllum, and Deuteria purpurascens. The herbaceous layer plants included mainly Apios delavayi var. Pteridiirorum, Lepisorus contortus and Dryopteris sinofrillosa.
Motuo County (93°46′ E–96°05′E, 27°34′N–29°56′ N) is at the southern foot of the eastern section of the Himalayas, with an average altitude of 1200 m, and is part of the subtropical humid climate monsoon region. The annual average temperature range is 16–18°C, and the annual rainfall is more than 2358 mm (Si et al., 2024). The main tree layer plants in the research plot were Abies delavayi var. motuoensis and Tsuga dumosa, while the shrub layer mainly consisted of Heptapleurum mitistellatum, Edgeworthia Gardner, Maesa cavenervis, etc. The herbaceous layer mainly consisted of Laportea bulbifera, Sarcopyramis napalensis, Oxalis corniculata, etc.
2.2 Methods
2.2.1 Vegetation sampling
The typical vegetated experimental sites with minor human intervention were selected through field investigations based on their habitat conditions and community status. In November 2018, a 100 m × 100 m permanent sample plot was set up at an altitude of 3800 m on the east slope of Mount Sejila in Bayi District. From July to October 2021, three fixed plots of 50 m × 50 m were set up in the natural fir forest near Cheyingou in Zayu County and near the 50 K marker on Zhamo Road in Motuo County. The basic information of the sample plots was recorded (Table 1). Each plot was divided into 25 survey units of 20 m × 20 m in Bayi District and 4 survey units of 25 m × 25 m in Zayu County and Motuo County. The adjacent grid method was used to survey each tree (base diameters ≥ 0.1 cm) in the forest, recording information including the species name, crown width, base diameter, breast height diameter, height, and coordinates. At the same time, 54 small plots of 1 m × 1 m were uniformly set up within the fixed sample plots in Bayi District; 20 small plots of 1 m × 1 m per plot were established in Zayu County and Motuo County, and herbaceous information, including the species name, height, and coverage, was recorded through sampling.
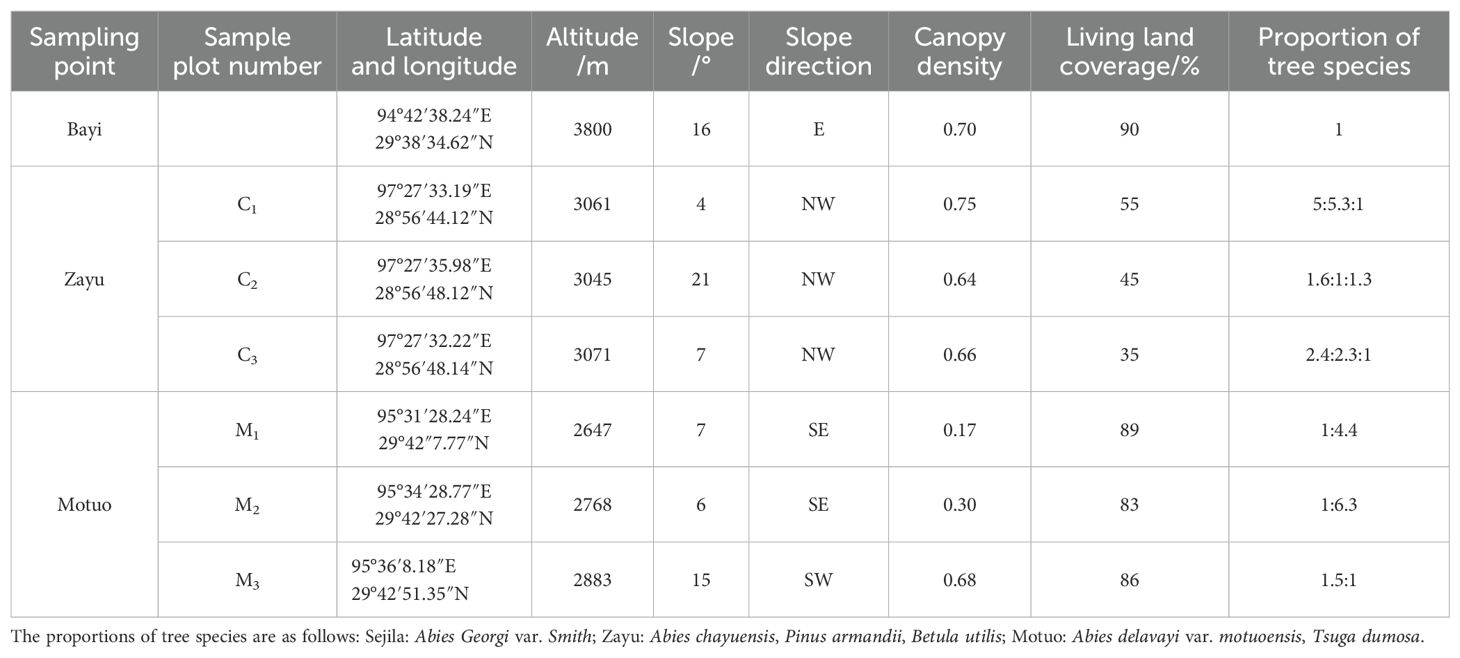
Table 1. Sample plot information.
2.2.2 Diameter classification
On the basis of the research objectives and specific conditions of the study area, we referred to previous methods of diameter classification for fir forests (Ren et al., 2021a). First, trees with a breast height diameter of less than 10 cm were uniformly classified into three classes on the basis of their base diameter (class 1: 0.1 cm ≤ D< 3.0 cm; class 2: 3.0 cm ≤ D< 6.0 cm; class 3: D ≥ 6.0 cm). Second, for plants with a diameter at breast height ≥ 10 cm, the standard was uniformly based on the diameter at breast height, with a step size of 10 cm for classification. This method was used to divide the populations of Abies georgei var. smithii, Abies chayuensis, and Abies delavayi var. motuoensis into 13, 24, and 20 diameter classes, respectively. It is difficult to determine their age for fir populations with longer growth cycles. Based on previous methods for dividing fir age classes (Wang et al., 2019; Ren et al., 2021a), adopting the widely used method of spatial extrapolation of time, namely, the diameter class method instead of the age class method (Shen et al., 2018; Wang et al., 2021). This method is based on the positive correlation between diameter class and age during tree growth. It indirectly infers the age stage of trees by measuring their base diameter or breast height diameter. As the number of individuals with a diameter at breast height ≥ 80 cm in the study area was relatively small, all diameter classes > 11 were classified under the XIth age class to analyze the life table characteristics of various classes. The specific criteria for the classification of diameter and age structures are shown in Table 2.
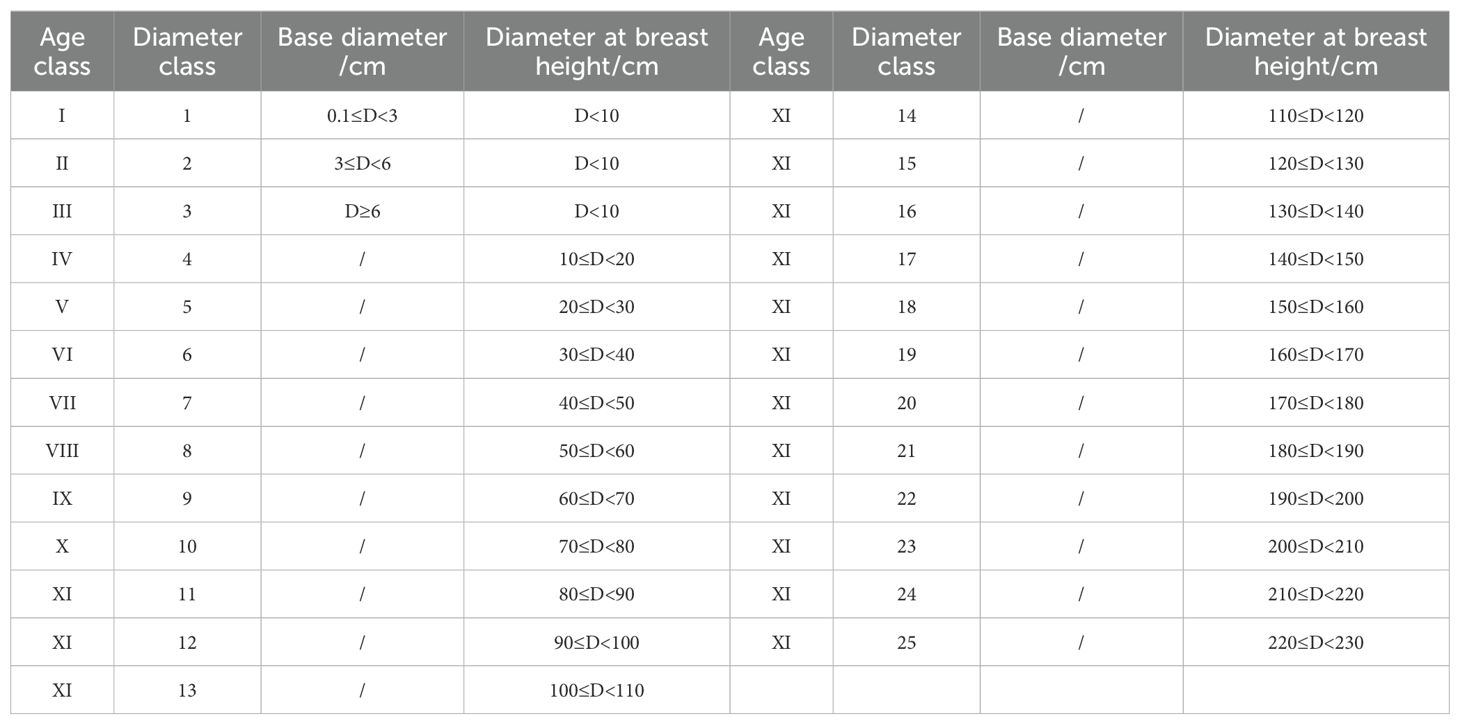
Table 2. Classification of diameter and age classes.
2.2.3 Static life table compilation
To compile the static life table, the method proposed by Wu et al. (2000) was adopted. Table 2 is the population life table of the rare and endangered plant, Nothotsuga longibracteata, along with its age classification. The main parameters in the table are as follows: the number of existing individuals in the first age class (a0); the current number of individuals within the x age class (ax); the standardized number of surviving individuals at the beginning of the x-age class (lx); the standardized number of deaths between adjacent age classes from x to x+1 (dx); the mortality rate (qx) between adjacent age classes from x to x+1; the number of surviving individuals between adjacent age classes from x to x+1 (Lx); the total number of individuals from the x age class to the x age class (Tx); the life expectancy (ex) of individuals entering the x age class; and the disappearance rate (degree of loss) (Kx). The specific formulas are as follows:
The survival curve of the tree population was drawn with the age class on the horizontal axis and lnlx on the vertical axis. Deevey divides survival curves into three types: Deevey-I is convex, indicating low mortality during early life and high mortality after reaching physiological lifespan; Deevey-II is linear, indicating roughly equal mortality at all stages; and Deevey-III is concave, indicating high mortality during early life. The type of survival curve for arbour populations is verified based on the exponential function Nx= N0e-bx and power function Nx= N0x-b models (Deevey, 1947; Hett and Loucks, 1976).
2.2.4 Method of dynamic quantification of the population size
The dynamic quantitative method for analyzing the population and community structure proposed by Chen Xiaode (Chen, 1998) was used to quantitatively describe the dynamics of the population structure of the main trees in a forest. The parameters involved and their meanings are as follows: the index of dynamic change (Vn) in the number of individuals in adjacent age classes; the number of individuals in each age class (Sn); the total age class (k); the dynamic index of quantitative changes in the population structure (Vpi), without considering external interference; and the dynamic index of quantitative changes in the population structure under random interference (V’pi); the risk rate of the largest interfering factor that threatens the survival or development of a population(P), only when the risk rate (P) is at its maximum value does it have the most significant impact on the population dynamics, V’pi. When Vn, Vpi, and V’pi are positive, negative, or 0, they represent the population’s dynamic growth, decline, and stable state or adjacent age classes. The specific formulas are as follows:
2.2.5 Analysis of population survival
The population survival rate (Sx), cumulative mortality rate (Fx), death density (fx), and hazard rate (λx) were calculated, and the survival rate, cumulative mortality rate, death density, and hazard rate curves (Wang et al., 2020; Zhang et al., 2022) were plotted via the following formulas:
In the formula, Pi represents the survival rate, and hi represents the age class width.
3 Results
3.1 Diameter structure of tree species
To explore the ecological status and trends of the major fir populations in southeastern Tibet, we initially conducted a plot survey method to analyze the diameter class structure of Abies georgei var. smithii, Abies chayuensis, and Abies delavayi var. motuoensis. The sample plot in Bayi District consists of a pure Abies georgei var. smithii forest with 408 trees per hectare. The sample plot in Zayu County has 745 trees per hectare, including Abies chayuensis, Pinus armandii, and Betula utilis. The sample plot in Motuo County has only 321 trees per hectare, including Abies delavayi var. motuoensis and Tsuga dumosa. Abies trees account for 100%, 43%, and 27% of the individuals surveyed in their respective plots, totaling 715 trees per hectare.
The statistical results of the diameter classes of the three species of Abies trees (Figure 1) reveal that the diameter class structure of Abies georgei var. smithii has a typical inverted “J” shape, and the diameters are in 13 diameter classes. The number of individuals in the first diameter class of the population is the highest, accounting for 21.08% of the total population. As the diameter increases, the number of individuals at the 1st to 3rd diameter classes continuously decreases significantly, and the number of individuals at the 3rd to 9th diameter classes fluctuates slightly. Among them, the number of individuals at the 7th and 9th diameter classes is greater than that at the 6th and 8th diameter class; after the 9th diameter class, the number of individuals sharply decreases, and the number of plants with diameters at breast heights< 10 cm accounts for 45.34% of the total population.
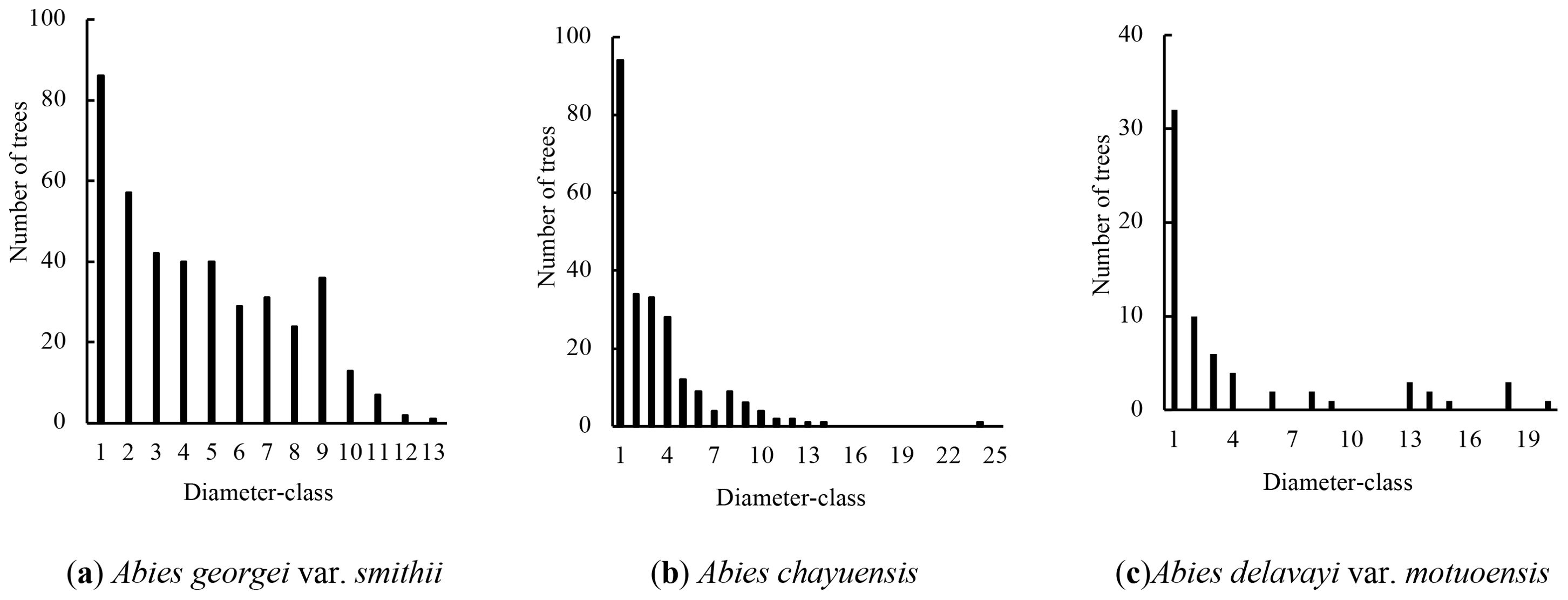
Figure 1. Radial structure. (A) Abies georgei var. smithii, (B) Abies chayuensis, (C) Abies delavayi var. motuoensis.
The number of individuals in the first diameter class of Abies chayuensis is the highest, accounting for 39.17% of the overall population. After the first diameter class, the number of individuals sharply decreases, and the number fluctuates irregularly with increasing diameter class. Among them, the number of individuals in the 8th diameter class is greater than that in the 7th diameter class, and the number of plants with DBH ≥ 10 cm accounts for only 32.91%, with the 15th to 23rd diameter classes missing.
Abies delavayi var. motuoensis is distributed mainly in the 1st to 2nd diameter classes, accounting for 62.69% of the total population. In the 2nd to 3rd diameter classes, the number of individuals in the population sharply decreases. Starting from the 3rd diameter class, as the diameter class increases, the population size distribution in the diameter class shows irregular fluctuations, with losses in the 5th, 7th, 10-12th, 16th, 17th, and 19th diameter classes.
Overall, the three populations of Abies have a pyramid-shaped diameter structure, with abundant reserves of seedlings and young trees. However, the proportion of medium- and large-diameter individuals is relatively small. The diameter structure fluctuates significantly, with varying degrees of diameter loss.
3.2 Tree population viability
3.2.1 static life table and quantitative analysis of population dynamics
To investigate the development trends, natural understory regeneration status, and responses to environmental disturbances of three Abies species populations in southeastern Tibet, this study employed a plot survey method to construct static life tables. It utilized Deevey’s theory and the Hett-Loucks test to analyze survival curves and calculated dynamic indices of quantity changes across age classes, aiming to comprehensively evaluate the population characteristics and dynamics. Because the community is a natural forest, the standardized mortality rate during the interval between some adjacent age classes has negative values. To ensure the authenticity of the data, no smoothing treatment was performed on the data. The static life tables of the three Abies tree populations (Table 3) reveal that the overall life expectancy values of the Abies georgei var. smithii and Abies chayuensis populations first increase but then decrease with age. After reaching a high value in age class IV, the life expectancy of the Abies delavayi var. motuoensis population significantly fluctuates because of the small number of surviving individuals in each age class. Therefore, the trend of change at this stage should be random.
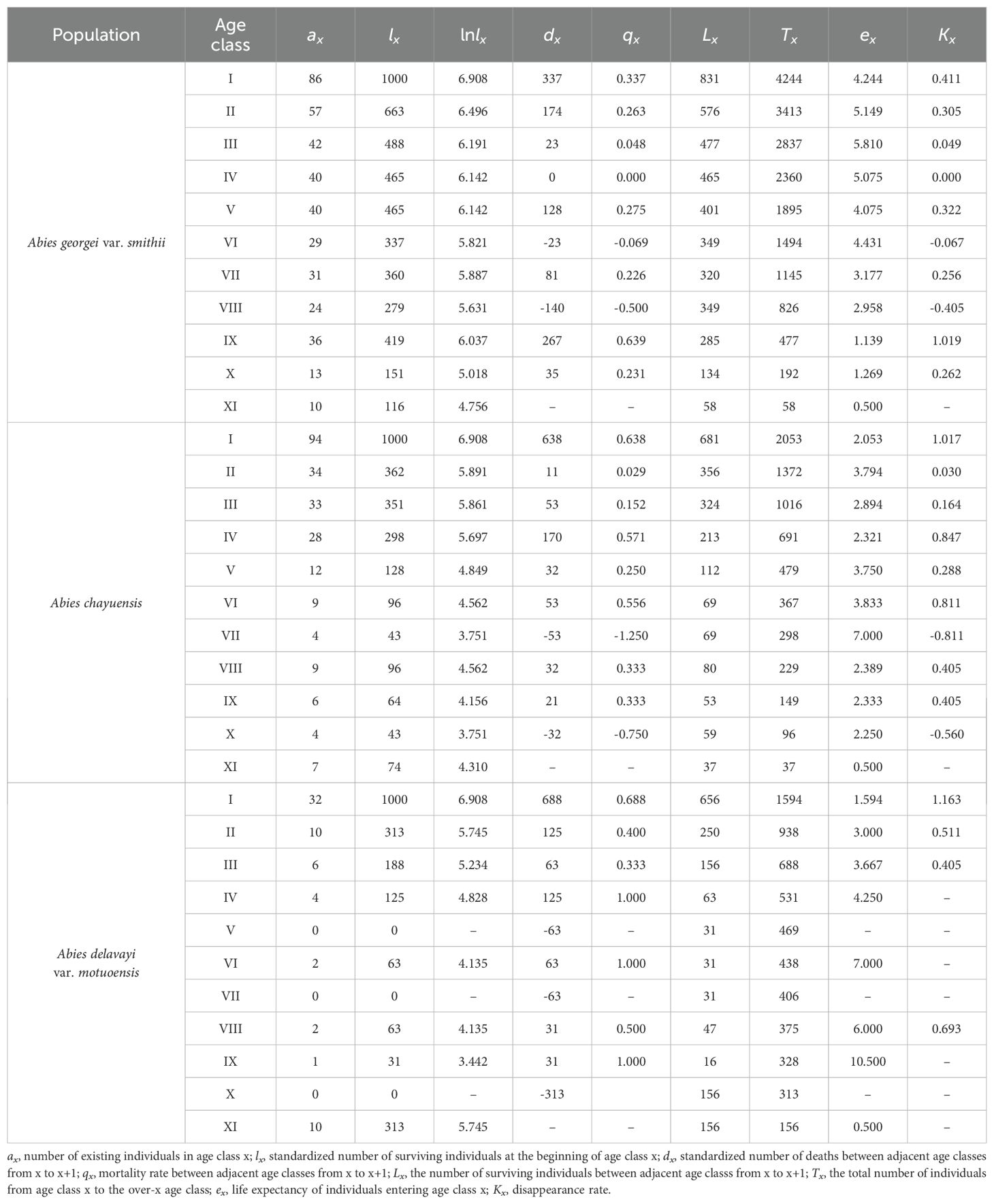
Table 3. Static life table of tree species.
An analysis of the survival curves of the three populations (Figure 2) reveals that the standardized logarithm of the number of surviving individuals in the three Abies genus tree populations reaches its peak in the first age class and then rapidly decreases in the second age class. As age increases, the rate of decline in the Abies georgei var. smithii and Abies chayuensis populations slows down in age classes II-XI, accompanied by inevitable fluctuations. Among them, the Abies chayuensis curve has a more significant fluctuating amplitude than the Abies georgei var. smithii curve does. The standardized survival logarithm of Abies delavayi var. motuoensis sharply decreases in age grades I-IV, and the curve shows irregular fluctuations after age grade IV.
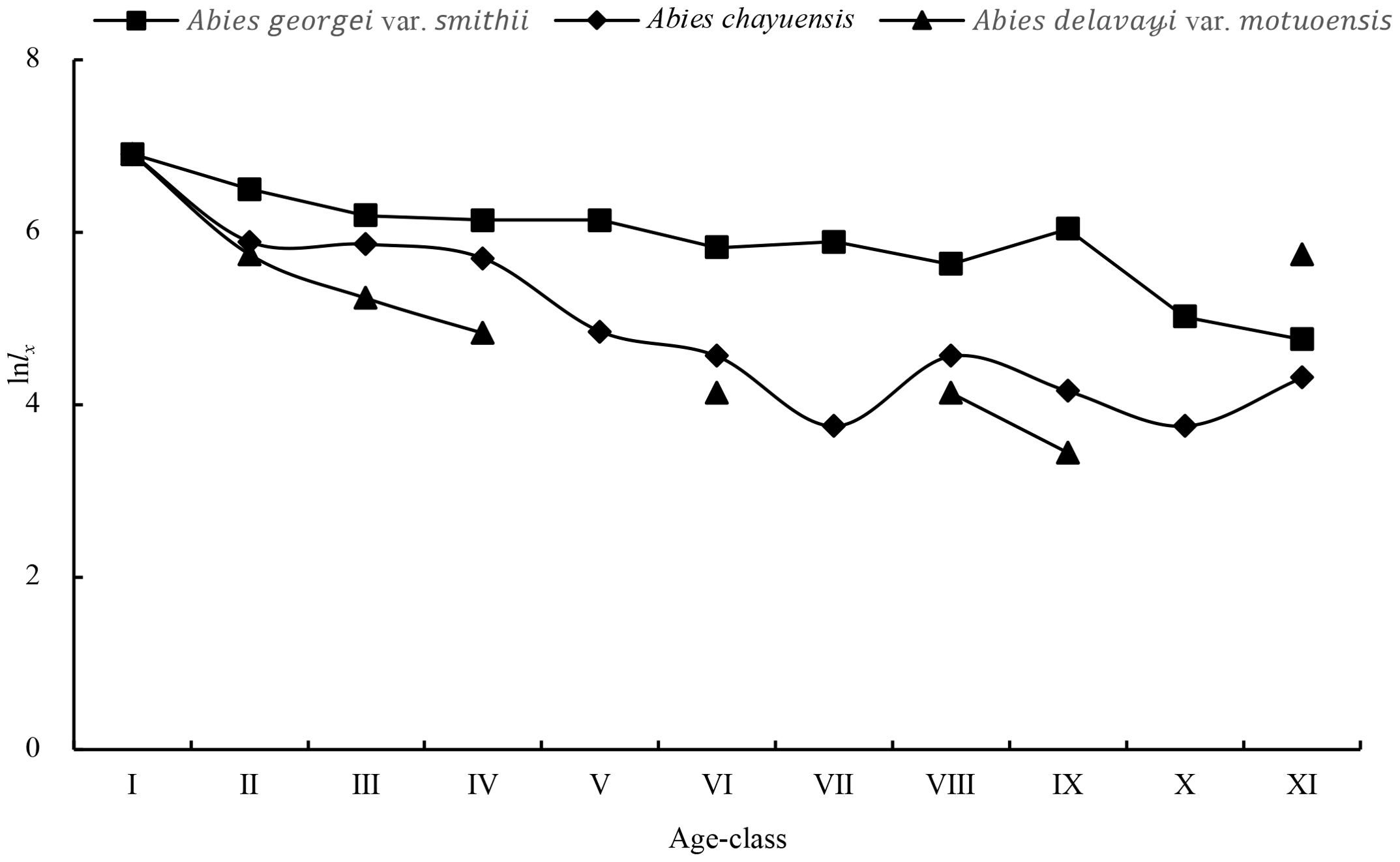
Figure 2. Survival curves.
In accordance with Deevey’s theory and Hdett and Loucks’ testing methods, the survival curves of 3 spruce tree populations were fitted and tested. A comparison of the R2 values (Table 4) reveals that the survival curve of the Abies georgei var. smithii population tends to be more linear (Deevey type II), indicating that the mortality rate of this population was similar at different age stages. The survival curves of the Abies chayuensis and Abies delavayi var. motuoensis populations tend to be concave (Deevey type III), indicating that high mortality rates are concentrated in the juvenile stage.
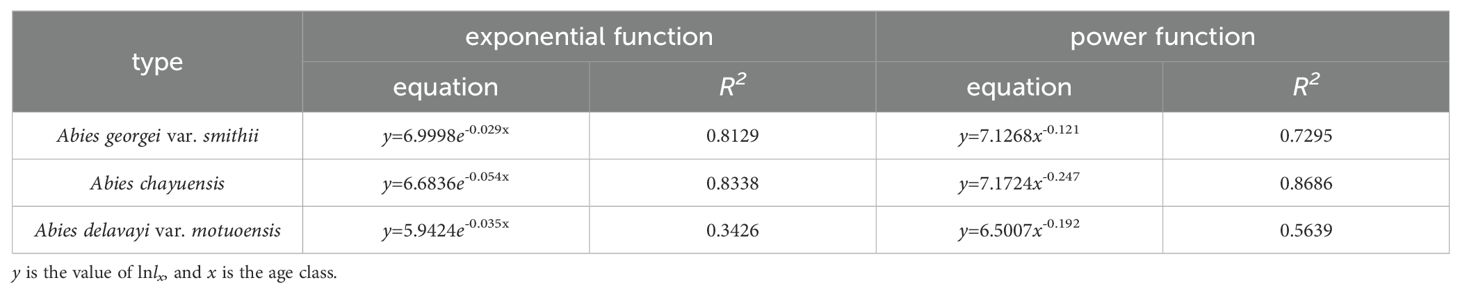
Table 4. Fitting equations for survival curves.
According to the analysis of the dynamic index of the changes in the number between adjacent age classes of the 3 Abies species (Table 5), except for V6 and V8< 0, V4 = 0, the remaining age classes of the Abies georgei var. smithii population are all > 0. Except for V7 and V10< 0, the remaining age classes of the Abies chayuensis population are all > 0. Except for V5, V7, and V10<0, the remaining age classes of the Abies delavayi var. motuoensis population are all > 0. Various classes exhibit varying degrees of decline after reaching the middle age range. The indices of dynamic change, Vpi, of the three populations are all greater than 0, indicating that each population shows a growth pattern at the current stage. Although the indices of dynamic change, V’pi, of various classes under external environmental interference are all greater than 0, the values are relatively low, indicating that various classes have slow growth and are susceptible to external environmental influences. The extreme value Pmax of the risk of random interference is more significant than zero, indicating that various classes are sensitive and unstable in the face of environmental interference.
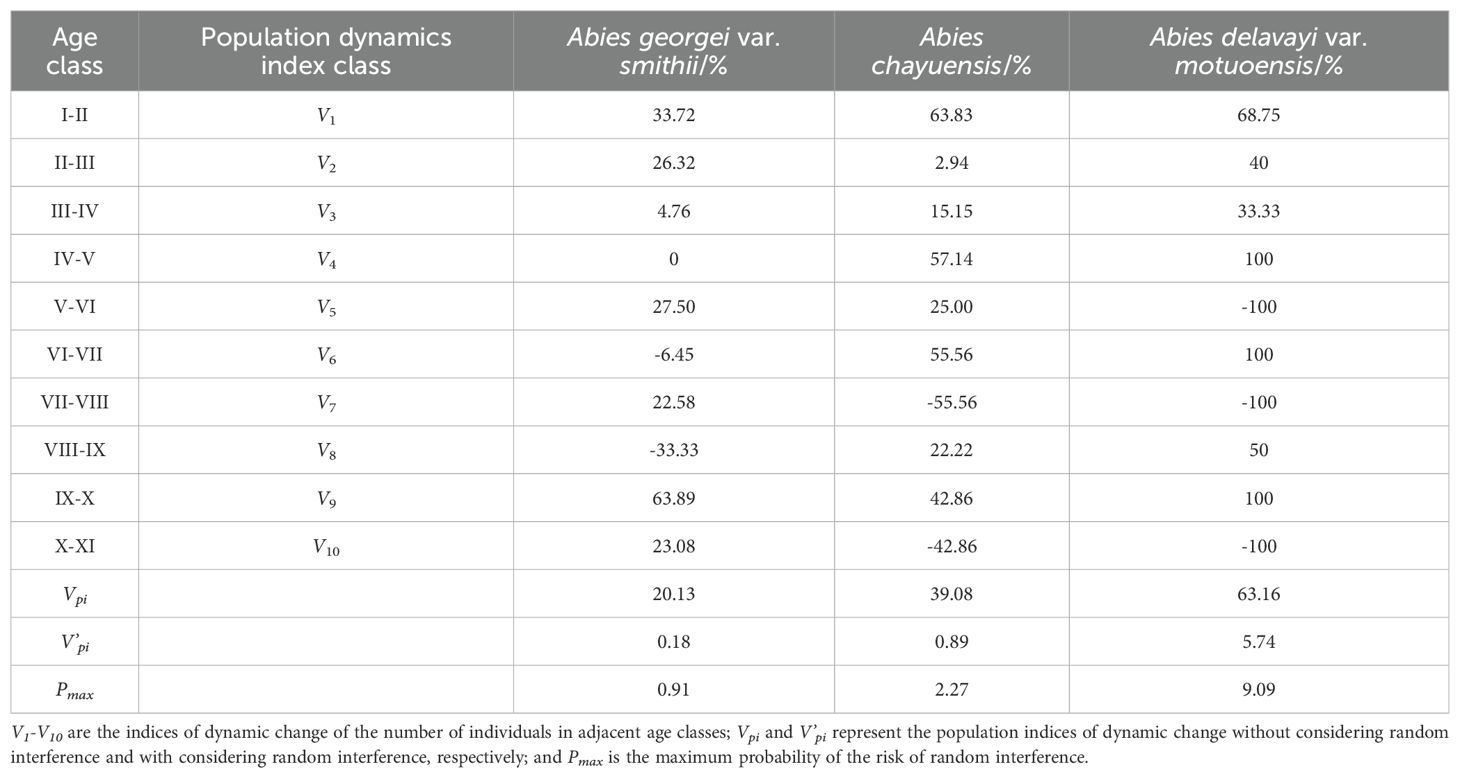
Table 5. Index of dynamic change in the three populations.
In summary, although the three Abies tree populations show an overall growth trend, they all exhibit slow growth characteristics, vulnerability to environmental disturbances. In particular, the mortality rate is high at the young age stage, and there are varying degrees of decline after the middle age class.
3.2.2 Mortality and disappearance rate curves
To comprehensively evaluate the dynamic stability of the various Abies species populations, static life table analysis was employed to fit mortality and disappearance rate curves. The three Abies populations’ mortality and disappearance rate curves (Figure 3) exhibit generally consistent trends in their variations. The mortality and disappearance rate curves of the Abies georgei var. smithii population reach high values in age class I, with some fluctuations in age classes I-VIII. The overall trend is decreasing, reaching a valley in age class VIII, rapidly increasing from age classes VIII-IX, reaching a peak in age class IX, and then rapidly decreasing again from age classes IX-X. The mortality and disappearance rate curves of the Abies chayuensis population show irregular fluctuations in age classes I to VI, with relatively high values appearing in age classes I, IV, and VI. They sharply decrease in age classes VI to VII, reach a valley at age VII, and gradually slow before rapidly decreasing in age classes VII to IX.
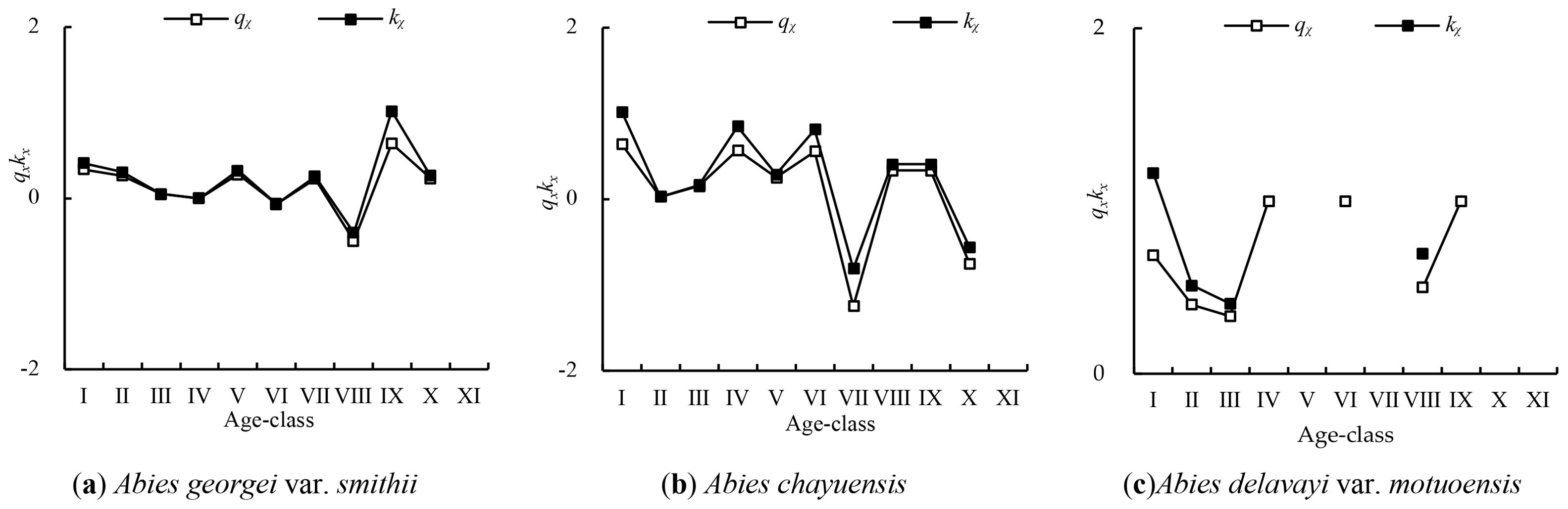
Figure 3. Mortality and disappearance rate curves. (A) Abies georgei var. smithii, (B) Abies chayuensis, (C) Abies delavayi var. motuoensis..
The mortality and disappearance rate curves of the Abies delavayi var. motuoensis population generally show a fluctuating trend of first decreasing and then increasing with increasing age in age classes I-VIII. All age classes reach relatively high mortality and disappearance rates. In this study, the number of individuals in the 11–25 cm diameter class of various populations is classified as age class XI, and the mortality and disappearance rates of the population in the X age class are relatively low. Therefore, the number of individuals in age classes IX-X for various populations shows a significant decrease. Overall, the mortality and disappearance rate curves of the 3 Abies tree populations repeatedly reach high values.
3.2.3 Analysis of tree population survival
This study constructed survival functions to evaluate the survival status and dynamic characteristics of Abies populations across age classes. It analyzed key indicators, aiming to reveal the survival capabilities of these two fir species. Owing to the lack of multiple classes, survival analysis was not conducted for Abies delavayi var. motuoensis. The survival functions of the Abies georgei var. smithii and Abies chayuensis populations were analyzed. The results (Figure 4) reveal that, owing to the negative mortality rates of these two populations, the survival rate and cumulative mortality rate curves do not show a monotonic upward or downward trend, and the two curves are complementary. The survival rate and cumulative mortality rate of the Abies georgei var. smithii population reaches a balanced state around age class II. Then, they tend to ease in age class V, indicating that the population enters a declining period afterward. After age class VIII, the survival rate sharply decreases, whereas the cumulative mortality rate sharply increases. The overall trend of the population mortality density and risk rate sharply decrease, followed by a gradual increase, with the peak mortality period mainly concentrated in age classes I and IX. The cumulative mortality rate of the Abies chayuensis population has always been higher than the survival rate, so there is no intersection between the two curves, and it tends to ease in age class VI, indicating that the population is beginning to show a trend toward decline and development. The mortality density of the population fluctuates greatly in the age classes I−III and age classes VI−X and is relatively flat in the age classes III−VI. The peak mortality period is mainly concentrated in age classes I and VIII. The population risk rate has undergone approximately four stages: a sharp decline in age classes I-II, a gradual decline in age classes II-VI, a sharp decline in age classes VI-VII, and a slow increase in age classes VII-X.
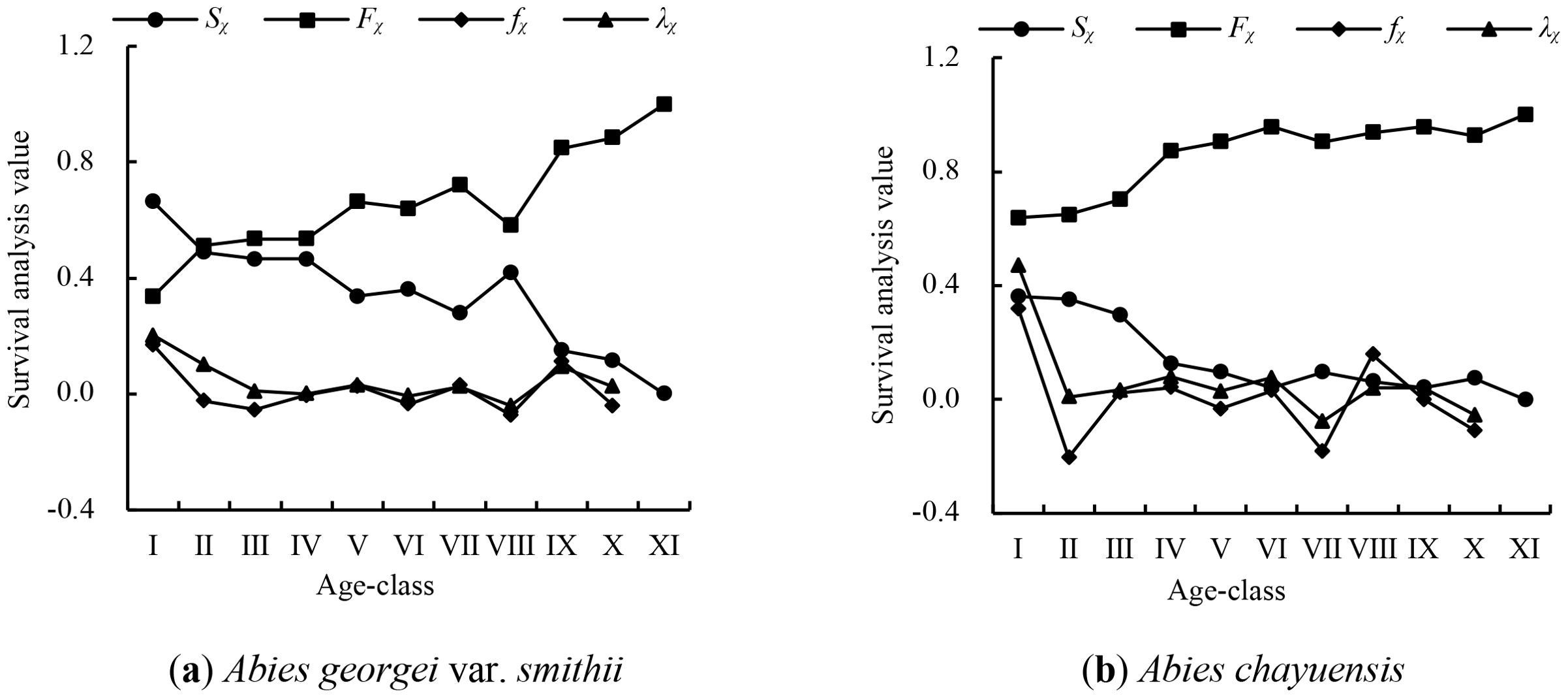
Figure 4. Survival analysis of the tree population. (A) Abies georgei var. smithii, (B) Abies chayuensis.
Overall, both spruce populations enter a decline period in the middle stage, and the mortality density and risk rate fluctuate greatly in the early and middle−late stages, which is consistent with the results of the static life table and survival curve studies. These findings indicate that the survival ability of seedlings in various populations is poor, the survival rate is low, the age structure is relatively unstable.
4 Discussions
4.1 Structural characteristics of trees
Under the background of the increasingly deteriorating global environment, this study conducted an in-depth analysis of the fir populations in southeastern Tibet, aiming to reveal their structural characteristics and development dynamics. The results showed that although the fir populations had increased in number, the scarcity of medium- and large-diameter individuals and their vulnerability to environmental disturbances highlighted the ecological fragility of the fir forests, indicating that their survival status was not optimistic. The radial structure of a population reflects not only its current growth and development status but also its response patterns to past environmental changes and future trends in community structure. It is one of the critical indicators reflecting a community’s succession process and stability maintenance status (Li et al., 2023). In this study, the diameter structure of the three Abies populations is a pyramid shape, and the population is growing, which is consistent with the results of the dynamic index analysis. However, as the diameter increases, the number of various populations shows varying degrees of fluctuation, indicating that although the three populations have high seed germination rates, with many seedlings, young plants, and sufficient reserves, their ability to sustain development is weak, and the number of medium- and large-diameter trees is relatively small.
There may be related to differences in the living habits and ecological adaptability of fir populations. 1) In actual investigations, it was found that the main trunk of fir populations in the Zayu and Motuo plots was thick and robust, but the heart rot was severe. This phenomenon limits the absorption and transportation of nutrients by trees. It weakens their ability to resist external environmental pressures, such as extreme weather conditions and invading pests and diseases. The deterioration of this health condition directly affects the development of tree crown width. It increases the occurrence of breakage, which may also hurt the regeneration of seedlings. In contrast, Abies georgei var. smithii exhibits a more robust performance, which may be related to the higher altitude of Mount Sejila. Low-temperature environments are more conducive to this population’s natural regeneration and growth. 2) In this study, the average coverage of the moss layer in the Abies georgei var. smithii forest is as high as 90%. Studies have shown that although the moss layer can improve the microenvironment and facilitate seed germination, it can form a mechanical barrier between the seedling roots and the soil, blocking the absorption of nutrients by seedlings from the soil, resulting in a large number of seedling deaths and inhibiting understory regeneration (Wang et al., 2016). Although the moss layer in the plots of Abies chayuensis and Abies delavayi var. motuoensis is underdeveloped, there are well-developed understory herbaceous plants. There may be intense interspecific competition between understory herbaceous vegetation and seedlings, which in turn affects the individual development of tree seedlings. 3) Previous studies have shown that fir tree species are affected by seed dispersal and interspecific competition and are often distributed in clusters during the seedling stage (Ren et al., 2021b). As seedlings and young trees continue to develop, individuals’ demand for environmental resources gradually increases. The dual competition within and between species produces strong “self-thinning and other thinning” effects, as well as asymmetric competition for nutrients and spatial resources among large trees in the main forest layer (Liu et al., 2016), which eliminates many new seedlings and makes it difficult to achieve good step-by-step replenishment between the medium- and large-diameter classes.
In addition, this study revealed significant fluctuations in the number of medium- and large-diameter spruce populations within the community, especially in the Zayu and Motuo populations, where large breaks in the diameter occurred, suggesting that there may have been more disturbances in the forest in the past. These disturbances may have included natural factors, such as abnormal climates, pests and diseases, geological disasters, and human factors, such as indiscriminate logging. In addition, spruce populations are sensitive to the environment. Therefore, when the living environment is disturbed, the structure of each spruce population experiences drastic fluctuations. The maximum risk of random disturbances in this study also shows that various populations are highly sensitive to high-altitude environments. However, further in-depth and systematic analysis is needed in conjunction with long-term monitoring to determine the specific disturbances that cause fluctuations in the population structure. Through a thorough analysis of the Abies populations in southeastern Tibet, this study has not only uncovered the critical issues in their structural characteristics and development dynamics, such as the scarcity of medium- and large-diameter individuals and their ecological vulnerability, but also explored multiple factors influencing population development, including competition from understory vegetation, poor seed setting rates in mature and overmature forests, and intraspecific competition. These discoveries deepen our understanding of the complexity and dynamics of alpine forest ecosystems and underscore the significance of Abies populations as ecological indicator species in the context of global environmental change.
4.2 Dynamic characteristics of the tree population
The use of static life table parameters to analyze plant population dynamics can provide insights into the survival status of a population and its relationship with its habitat. It also reflects the position and role of the population in the community, which is highly important for maintaining community stability and sustainable development (Wang et al., 2004; Zhang et al., 2022). Numerous studies have shown that the seedling stage is the most essential and sensitive stage in forest regeneration (Li et al., 2022). Limited resources and space often limit population renewal (Zhang et al., 2022). By fitting of the survival curve, the survival curve of the Abies georgei var. smithii population tends to be a Deevey type II curve, indicating that the population has a relatively stable mortality rate. Combined with the static life table, the life expectancy values of the Abies georgei var. smithii population tended to first increase but then decrease with increasing age. Both the mortality and disappearance rates were high in the young and old age classes. This may be due to the high density and weak adaptability of young plants to the environment. they are prone to high mortality rates, resulting in a smaller average lifespan. To some extent, this reflects that the large-scale death of young plant individuals is a bottleneck for plant population renewal and recovery. As the age class increases, a small number of individuals enter the middle age class, and their growth vitality increases, making them more competitive. Therefore, their mortality rate gradually decreases, and their life expectancy gradually increases. However, as the life cycle of the population continues, the physiological age of individual spruce trees gradually increases, their morphology and physiological functions significantly decline, and their ability to resist external interference significantly decreases, resulting in a rapid decrease in the life expectancy of elderly individuals. Consistent with the results of Yao et al. (2020) on the population of Pinus armandii in southeastern Tibet, similar reports have also been published on different species in other regions (Wu et al., 2021; Gong et al., 2024).
In contrast, the survival curve of the Abies chayuensis tends to be a Deevey III type curve, indicating a high mortality rate among young adults in this population. The static life table shows that this population’s expected life value, mortality rate, and disappearance rate are similar to those of the Abies georgei var. smithii. However, higher mortality and disappearance rates also occur in the middle age class, which may be related to the environment of the mixed forest where the Abies chayuensis is located. The multi-dimensional competitive environment not only intensifies the tension in resource allocation but may also affect the growth rate and health status of Abies chayuensis. Especially as the Abies chayuensis grows to the mid-age stage, competition with high-density Pinus armandii species becomes particularly fierce, leading to an increase in population mortality and disappearance rates, resulting in significant dynamic changes in the mortality and disappearance rates of the Chayu spruce population at different stages of life. However, the Abies delavayi var. motuoensis is facing more complex ecological challenges. The survival curve of this population tends to be Deevey III type curve, and the expected life value of the population exhibited irregular dynamic fluctuations throughout the entire life cycle, which was consistent with the mortality and disappearance rate curves. There may be three reasons for these results. First, the average canopy closure in the Abies delavayi var. motuoensis forest was only 0.38, and the gaps were relatively developed, resulting in a dense and tall herbaceous layer under the forest. For example, the height of ferns can reach 2 m, which severely inhibits the natural regeneration of fir trees in forests. Moreover, the growth of fir seedlings is slow, and it takes at least 10 years to reach a height of 2 m, which means that during this period, seedlings or small trees have to compete fiercely with the herbaceous layer, severely inhibiting the growth and development of fir trees. Second, owing to the unique high-temperature and high-humidity environment in the Motuo region, the trunks of middle and older spruce trees in Motuo have decayed, reducing their ability to interfere with external forces. In addition, because this species is mixed with Tsuga dumosa and is located within the same ecological niche, there is significant interspecific competition. With the succession of the community, interspecific and intraspecific competition intensifies, and the number of Abies delavayi var. motuoensis gradually decreases. Eventually, this population may be eliminated. According to the analysis of the four survival functions, the survival rates and cumulative mortality rates of the populations of Abies georgi var. smith and Abies chayuensis begin to decline from the middle age stage, which is consistent with the results of the diameter structure and static life table analyses. Through the analysis of life table parameters and survival curves of different Abies populations in southeastern Tibet, we have not only deepened our understanding of the population dynamics and their interaction mechanisms with the environment but also revealed the common challenges faced during the regeneration process of alpine forests, such as the high mortality rate of young individuals, the limitation of forest canopy closure on regeneration, and the complexity of interspecific competition. These findings are crucial for formulating effective forest conservation and management strategies, especially in global climate change, to ensure alpine forest ecosystems’ stability and sustainable development.
4.3 Limitation and development
This study has preliminarily revealed the population dynamics of Abies species in southeastern Tibet and their macro-environmental factors. Still, the results lack universality due to limited samples and specific regions. To enhance the comprehensiveness of the research, it is necessary to expand the sample size and geographical coverage in the future, conduct comparative studies across ecological regions, and clarify the differentiated impacts of environmental factors on population dynamics. Meanwhile, as the micro-physiological mechanisms such as population renewal, seedling survival, and growth are still unclear, it is essential to strengthen multidisciplinary cooperation and integrate the strengths of ecology, genetics, and molecular biology to explore the development, reproduction, and response mechanisms of Abies species, thereby providing a more solid scientific basis for their conservation and management.
5 Conclusion
This study focused on the in-depth analysis of the population structure and dynamics of fir forests in southeast Tibet against the backdrop of intensified global climate change. By adopting the typical plot survey method and integrating ecological techniques such as diameter class structure analysis, static life table compilation, and survival analysis, we aimed to reveal the survival status, population development trends, and their response mechanisms to external disturbances of these three Chinese endemic fir species in their natural environment. The main findings are summarized as follows:
1. The populations of Abies georgei var. smithii, Abies chayuensis, and Abies delavayi var. motuoensis are all growth-type populations, and they compose a relatively high proportion of small-diameter species. However, their stability is poor, and their ability to resist environmental disturbances is low. The community has sufficient seedling reserves to meet the current development requirements of the community. Nevertheless, few trees have grown into medium- and large-sized trees, and the population is unstable.
2. Rapidly growing Abies georgei var. smithii tends to be Deevey type II. In contrast, the populations of Abies chayuensis and Abies delavayi var. motuoensis tend to be Deevey type III. The mortality and disappearance rate curves of the three populations repeatedly reach high values, showing a fluctuating trend, all of which begin to decline after the middle age class. The age structure is relatively unstable, indicating that the three populations are highly unstable and have poor development due to considerable environmental screening and competitive pressure.
In summary, after a certain period, the lack of new growth, intraspecies competition, and habitat deterioration are the reasons for the decline of various classes in the natural fir forest in southeastern Tibet. In future protection and management, it is necessary to strengthen the nurturing and management of seedlings and young trees, such as by selectively cutting down excess individuals, such as adjusting the coverage of the moss layer in Mount Sejila through moderate manual intervention, reducing its mechanical barriers to the root system of Abies georgei var. smithii seedlings, and promoting their healthy growth. Sparse the understory herbaceous layer that hurts seedlings in the areas of Zayu County and Motuo County to improve the survival rate of young seedlings of Abies chayuensis and Abies delavayi var. motuoensis and provide more living space for small-diameter trees in the understory to enter the canopy layer to maintain population survival. It is also necessary to strengthen monitoring and prevention of heart rot disease, adopt reasonable nurturing measures such as pruning and thinning, reduce the occurrence of heart rot, and improve the health level of the population.
To gain a more comprehensive understanding of the dynamic changes in the Abies population, future efforts should broaden the scope of sample plots to encompass a wider range of ecological regions and diverse types of fir forests. Through long-term, systematic monitoring and surveys, we aim to accumulate a wealth of data. Building upon macroecological studies, modern technological approaches, including molecular biology and genetics, will be utilized to further explore the microscopic mechanisms of how environmental factors influence the regeneration and growth of fir populations. This will reveal the physiological and ecological responses as well as adaptation strategies employed by fir populations in the face of environmental changes, thereby providing a crucial theoretical basis for the sustainable utilization and conservation of fir forests in southeastern Tibet.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
DH: Conceptualization, Formal analysis, Methodology, Writing – original draft. YR: Funding acquisition, Supervision, Writing – review & editing, Project administration, Resources. SY: Data curation, Writing – review & editing, Investigation. YZ: Validation, Writing – review & editing. GZ: Validation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by National Natural Science Foundation of China: 32371651.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Cai J. A., Ming D. P., Zhao W. Y., Ling X., Zhang Y., Zhang X. X. (2024). Integrated remote sensing-based hazard identification and disaster-causing mechanisms of landslides in Zayu County. Remote Sens. Natural Resources. 36, 128–136. doi: 10.6046/zrzyyg.2023313
Chen X. D. (1998). A study on the method of quantitative analysis for plant population and community structural dynamics. Acta Ecologica Sin. 02, 104–107.
Deevey J. E. S. (1947). Life tables for natural populations of animals. Q. Rev. Biol. 22, 283–314. doi: 10.1086/395888
Fu F. W., Li J. R., Li Y. Y., Chen W. S., Ding H. H., Xiao S. Y. (2023). Simulating the effect of climate change on soil microbial community in an Abies georgei var. smithii forest. Front. Microbiol. 14, 1189859. doi: 10.3389/fmicb.2023.1189859
Fu L. G., Li N., Mill R. R. (1999). “Pinaceae,” in Flora of China, vol. 4 . Eds. Wu Z. Y., Raven P. H. (Science Press, Beijing), 11–52.
Gong Y. M., Xie G. L., Liu S., Zhu N. H. (2024). Population structure and dynamic characteristics of Iron-heart Cunninghamia lanceolata. J. Cent. South Univ. Forestry Technol. 44, 138–145. doi: 10.14067/j.cnki.1673-923x.2024.02.015
He J. N., Ning C. W., Zhang W. T., Halik Ü., Shen Z. H. (2024). Altitudinal patterns of population structure dynamics of Picea schrenkiana in the northern slope of the eastern Tianshan Mountains. Acta Ecologica Sinica. 44, 5389–5398. doi: 10.20103/j.stxb.202304210824
Hett J. M., Loucks O. L. (1976). Age structure models of balsam fir and eastern hemlock. J. Ecol. 64, 1029–1044. doi: 10.2307/2258822
Lei Y., He X. N., Wang J. M., Liu J. T., Tan X. W., Zhao B., et al. (2022). Population structure and dynamic characteristics of Phoebe zhennan S. Lee in karst areas of Chongqing. Acta Ecologica Sinica. 42, 4903–4911.doi: 10.5846/stxb202106171607
Li M. M., Liu P. C., Kong W. M., Ma F. L., Li S. F., Wang M. (2022). Population structure and dynamic characteristics of the endangered Pseudotsuga forrestii Craib. Acta Ecologica Sin. 42, 5504–5515.
Li W., Zhang G. F. (2015). Population structure and spatial pattern of the endemic and endangered subtropical tree Parrotia subaequalis (Hamamelidaceae). Flora-Morphology Distribution Funct. Ecol. Plants. 212, 10–18. doi: 10.1016/j.flora.2015.02.002
Li Y. W., Ren P., Wang Y. F. (2023). Structure and spatial distribution patterns of natural Pinus densata Mast populations in Milin County of Southeast Xizang, China. Chin. J. Appl. Environ. Biol. 29, 1109–1116. doi: 10.19675/j.cnki.1006-687x.2022.06029
Liu S., Hou M. M., Liao J. X., Wang J. W., Xiao C., Fan X. H. (2016). Local biotic neighborhood and habitat heterogeneity drive tree seedling survival dynamics in a temperate forest. Chin. J. Appl. Environ. Biol. 22, 639–645. doi: 10.3724/SP.J.1145.2015.11029
Lu J., Pan G., Luo D. Q., Zheng W. L., Guo Q. Q., Qin Y. L. (2010). Analysis to the life table of endangered population Abies Georgei. Res. Soil Water Conserv. 17, 212–216+221.
Lu J., Wang G., Ma J., Yu Q., Gao T., Niu T. (2021). Forest ecological network construction and structure evaluation based on complex network theory in Bayi district of Tibet. Trans. Chin. Soc. Agric. Machinery. 52, 152–158.
Ren Y. H., Luo D. Q., Fang J. P., Lu J. (2021a). Structure and dynamics of Abies georgei var. smithii population in eastern of the Sejila Mountains. J. Northwest A&F Univ. (Nat.Sci.Ed.) 49, 59–68. doi: 10.13207/j.cnki.jnwafu.2021.07.007
Ren Y. H., Zhou Y. Z., Hou L., Fang J. P., Luo D. Q. (2021b). Spatial distribution patterns of standing trees at different ages in Abies georgei var. smithii forests in Sejila Mountain. Acta Ecologica Sin. 41, 5417–5424. doi: 10.5846/stxb202005301406
Shen Z. Q., Lu J., Hua M., Tang X. Q., Qu X. L., Xue J. L., et al. (2018). Population structure and spatial pattern analysis of Quercus aquifolioides on Sejila Mountain, Tibet, China. J. forestry Res. 29, 405–414. doi: 10.1007/s11676-017-0444-1
Si M. X., Chen L., Ma H. P., Wang B., Gao Z. Y., Fan B. B. (2024). Flora of liverworts in typical area of Motuo, Xizang. J. For. Environment. 44, 207–216. doi: 10.13324/j.cnki.jfcf.2024.02.012
Wang Y. T., Huang Z. H., Wang J., Zhang T., Cui G. F. (2021). The population structure and dynamic characterisics of Phelodendron amurensein in Yanshan Mountains. Acta Ecologica Sinica. 41, 2826–2834.
Wang F., Huo Y. C., Zhao Y., Cao X. W., Liu J. Q., Yang Y. H., et al. (2019). Population structure and dynamics of Original Abies faxoniana Rehd- Rhododendron simsii Planch in High-mountain Timberline of Southern Gansu Province. Bull. Botanical Res. 39, 664–672.
Wang T., Liang Y., Ren H. B., Yu D., Ni J., Ma K. P. (2004). Age structure of Picea schrenkiana forest along an altitudinal gradient in the central Tianshan Mountains, northwestern China. For. Ecol. Manage. 196, 267–274. doi: 10.1016/j.foreco.2004.02.063
Wang R. H., Ren Z. D., Jin X. Q., Baima Y. Z., Pan G. (2016). Study on the seed characteristics and natural regeneration of Abies georgei var. smithii. J. Fujian Forestry Sci. Technol. 43, 49–52+75. doi: 10.13428/j.cnki.fjlk.2016.04.010
Wang J., Yao L., Ai X. R., Zhu J., Liu S. B. (2020). Structure and dynamic characteristics of Betula luminifera populations in different regions of Southwest Hubei Province, China. Chin. J. Appl. Ecol. 31, 357–365. doi: 10.13287/j.1001-9332.202002.010
Wu C. Z., Hong W., Xie J. S., Wu J. L. (2000). Analysis of the life table of the rare and endangered plant Nothotsuga longibracteata population. Chin. J. Appl. Ecol. 03, 333–336. doi: 10.13287/j.1001-9332.2000.0085
Wu Q. C., Zang F. Q., Li C. C., Ma Y., Gao Y., Zheng Y. Q., et al. (2021). Population stucture and dyamics of endangered Populus wulianensis. Acta Ecologica Sin. 41, 5016–5025.
Yao H. F., Lu J., Wang C., Chen K. (2020). Population structure and quantitative dynamics of Pinus armandii in Bomi Gang Township Nature Reserve. For. Grassland Resour. Res. 05, 108–115+130. doi: 10.13466/j.cnki.lyzygl.2020.05.016
Zhang J. F., Ge S. S., Liang J. H., Li J. Q. (2022). Population structure and dynamic characterisics of Tilia amurensis in broad-eaved Korean pine mixed forest in Changbai Mountain. Acta Ecologica Sin. 42, 5381–5390.
Zhang J. T., Pan Z. L., Tian Y. H., Hou J. L., Peng M. C., Duan H. X., et al. (2022). Age structure and dynamics of Pinus yunnanensis population in Yunlong Tianchi Nature Reserve. Acta Ecologica Sin. 42, 9091–9099.
Zhang X. P., Yu L. Z., Yang X. Y., Huang J. Q., Yin Y. (2022). Population structure and dynamics of Pinus koraiensis seedlings regenerated from seeds in a montane region of eastern Liaoning Province, China. Chin. J. Appl. Ecol. 33, 289–296. doi: 10.13287/j.1001-9332.202202.001
Zhang X. S., Zhao N. N., Zhou C. N., Lu J., Wang X. T. (2022). Seedling age of Abies georgei var. smithii reveals functional trait coordination in high-altitude habitats in southeast tibet. Front. Ecol. Evolution. 10, 955663. doi: 10.3389/fevo.2022.955663
Keywords: typical plot method, diameter structure, static life table, fir forests, Southeastern Tibet
Citation: Huang D, Ren Y, Yang S, Zhao Y and Zhao G (2024) Population structure and dynamic characteristics of three endemic Chinese Abies species in southeastern Tibet, China. Front. Ecol. Evol. 12:1487984. doi: 10.3389/fevo.2024.1487984
Received: 30 August 2024; Accepted: 21 November 2024;
Published: 10 December 2024.
Edited by:
Ming Jiang, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Zhijun Lu, Chinese Academy of Sciences (CAS), ChinaZongzheng Chai, Guizhou University, China
Copyright © 2024 Huang, Ren, Yang, Zhao and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yihua Ren, eHpfcnloQDE2My5jb20=