 Eszter Matrai
Eszter Matrai Shaw Ting Kwok
Shaw Ting Kwok Ziying Tan
Ziying Tan Xiao Lin
Xiao Lin Wei-Kwan Lee1
Wei-Kwan Lee1 Paolo Martelli
Paolo Martelli Ákos Pogány
Ákos Pogány- 1Research Department, Ocean Park, Hong Kong, Hong Kong SAR, China
- 2Department of Psychiatry, Icahn School of Medicine at Mount Sinai, New York, NY, United States
- 3Department of Ethology, Eötvös Loránd University, Budapest, Hungary
Cognitive enrichments have gained popularity in the past two decades in both farm animal and zoological settings. In this study, we present a cognitive enrichment device that, for the first time, allowed testing for prosocial food-sharing actions in dolphins. The device was made of a PVC tube and two caps with rope handles; one handle was fixed to the poolside whereas the other was available for the dolphins. Its internal structure kept five fish in place while maintaining a closed status. The dolphin who pulled the rope handle could not access the fish but allowed another individual to access it. Once the handle was released, however, the device closed back. The device was tested with a male and a female group, separately. The males obtained approximately a third, while the females acquired none of the fish using the device. Moreover, the males were observed to open the device more frequently and longer in the presence of their group members and with consistent roles. These results provide the first support of male dolphins’ willingness to engage in food-sharing that could be considered as an altruistic action. It also provides basis for further investigations on more detailed and long-term monitoring of the emergence and consequences of such cooperative actions.
Introduction
Social animals engage in a variety of interactions with their peers that have different functions, including the reinforcement of social bonds through affiliative interactions (Connor et al., 2000) or the learning of important behaviors through play (Kuczaj and Yeater, 2007) or socio-sexual interactions for instance (Herzing, 1996). Social living provides opportunities for prosocial behaviors, where actions from a donor potentially benefit the recipient without preluding benefits to the donor (de Waal, 2008). Prosocial behaviors may manifest in a variety of contexts, such as care giving, affiliation, social teaching, cooperation and sharing (Jean-Loup, 2019), and these are all apparent in cetaceans. The most common form of caregiving is parental care, which in cetaceans is restricted to extensive female-only (i.e. maternal) care in a form or nursing (e.g. Herzing, 1997; Martin and Da Silva, 2018; MacLeod and D’Amico, 2006; New et al., 2013). In cetacean societies, individuals other than the parents may also contribute to the care of the young, in a form of alloparental associations (Weinpress and Herzing, 2015; Simard and Gowans, 2004; Augusto et al., 2017; Whitehead, 1996; Gero et al., 2009). Affiliative associations (i.e., selective social interactions with specific individuals, including spatial proximity and behavioral synchronization) are also characteristic of cetaceans. Synchronous swimming with one or more partners is often described as an affiliative behavior in bottlenose dolphins (Tursiops sp, Connor et al., 2006; Fellner et al., 2013; Clegg et al., 2017). These behaviors may involve tactile interactions (Sakai et al., 2016) and could facilitate social learnings of prey capture (Bender et al., 2009; Guinet and Bouvier, 1995). But probably the most extensively studied prosocial behavior in cetacean is cooperation.
Bottlenose dolphins (Tursiops sp.) in the wild have been recoded to cooperate to capture food or to gain and maintain access to mating partners. A wide range of ingenious cooperative hunting tactics have been observed with bottlenose dolphins in the wild such as mud ring feeding (Ramos et al., 2022), strand-feeding (Jiménez and Alava, 2015), cooperative prey herding (Benoit-Bird and Au, 2009), coordinated leader-wall hunting (Gazda et al., 2005) and even inter-specific cooperation with fishermen (Romeu et al., 2017). The diversity and flexibility of these strategies likely contributed to the successful, wide range distribution of bottlenose dolphins. Cooperation, as part of their mating strategies, has been demonstrated though male-alliance formation. In Sarasota, Florida, unrelated male dolphins form stable alliances; when working together, they can maintain access to a single female dolphin, fend off predators and doing so increase their reproductive fitness compared to males with no partners (Wells, 2014). Male dolphins (Tursiops sp.) in Shark Bay, Australia, engage in multi-level alliances, creating examples of the most complex social network outside humans (Krützen et al., 2003). These alliances can last for decades or even for lifetime (Connor and Krützen, 2015).
Cooperation and coordination have been tested with dolphins under human care, in a synchronous button pressing task (Jaakkola et al., 2018), in a coordinated rope pulling task (Yamamoto et al., 2019), and in the spontaneous cooperative use of a set of cognitive enrichment devices. Cognitive enrichments represent a special category of environmental enrichments. They allow the animals to practice their natural problem-solving skills, control some aspect of their environment and the use of these enrichments correlates with one or more validated measures of their wellbeing (Clark, 2013, 2017, Clark et al., 2013). A set of cognitive enrichment devices were designed to test cooperative problem-solving in dolphins. They were made of PVC pipes and caps with rope handles attached. They contained ice cubes and fish (Mallotus villosus) which could be obtained by the simultaneous pull of the rope handles. Dolphin pairs, trios and quartets successfully manipulated the cognitive enrichment (Kuczaj et al., 2015; Matrai et al., 2021a, 2021b, 2022a). During the use of these devices, aggression decreased, while play and affiliative, social interactions increased significantly (Matrai et al., 2022b). Once the devices were opened, the dolphins could have access and even share their content. In wild cetaceans, food-sharing has only been reported in a handful of studies. Kin-directed food-sharing was recorded in a population of fish-eating killer whales, mostly expressed by adult females towards maternal kin (Orcinus orca, Wright et al., 2016). Prey-transfer was recorded between adult and calf marine tucuxi dolphin (Sotalia fluviatilis), whereby the mother captured a fish, swam side-by-side with the calf and transferred the fish to the calf’s mouth (Spinelli et al., 2008). In a case study, two adult common bottlenose dolphins (Tursiops truncatus) in Costa Rica were documented sharing a fish; the adults were a female dolphin (who was accompanied by a calf) and a male dolphin (Fedorowicz et al., 2003). A group of rough-toothed dolphins (Steno bredanensis) were observed to carry a large mahi-mahi (Coryphaena hippurus) and pass it between each other (Brower and Curtsinger, 1979). In a more recent study, five rough-toothed dolphins repeatedly passed an unidentified species of jack (Caranx spp.). A dolphin took pieces of flesh off the fish and dropped it, and then another dolphin picked it up and performed a similar action (Ramos et al., 2020).
The proximate mechanisms responsible for these prosocial behaviors may differ based on the donor’s motivation. It can be driven by higher-level social skills such as empathy and altruism, but just as well by lower-level mechanisms, including self-rewarding for the donor, and simpler neurobiological mechanisms. Finally, neuropeptides may be sufficient to elicit these behaviors (Jean-Loup, 2019). While empathy and the neurobiological mechanisms are understudied in non-humans, neuropeptides have been reported to promote prosocial behaviors. Oxytocin was found to facilitate prosocial behavior and prevent social voidance in rats and mice (Lukas et al., 2011). Oxytocin also enhanced grooming and food-sharing in bats (Desmodus rotundus, Carter and Wilkinson, 2015). During cognitive bias testing with dolphins under human care, affiliative behavior correlated with more positive emotional states (Clegg et al., 2017). Altruism in cetaceans has long been suggested as a driver, however, controlled tests are lacking. The most intriguing potential altruistic events were recorded between different species in the wild. A humpback whale (Megaptera novaeangliae) was recorded to interfere with mammal-eating killer whales (Orcinus orca) to help pinnipeds and fish species (Pitman et al., 2017). In Hawaii, humpback whales (Megaptera novaeangliae) were observed engaging in unusual, possibly cooperative play behavior with bottlenose dolphins (Tursiops truncatus, Deakos et al., 2010). In addition, Sakai et al. (2016) and her colleagues monitored a wild Indo-Pacific bottlenose dolphin (Tursiops aduncus) that adopted and cared for a genetically distant neonate calf. Finally, wild dolphins that participated in a provisioning program offered fish and cephalopods to staff members multiple times. This “gift-giving” behavior occurred as part of a sequence such as playing as well as a discrete behavior (Holmes and Neil, 2012).
While food-sharing has been frequently documented in a range of species, there is only limited documentation in cetaceans, which is surprising considering the social complexity and richness of cooperative interactions recorded in the taxon. Investigating food-sharing under controlled conditions allows us to better understand the socio-cognitive abilities, emergence and maintenance of the social structure and social dynamics of wild dolphins. While the maintaining mechanism of prosocial behaviors is debated, their presence in non-human species have frequently been reported and has intrigued researchers for over half a century. Controlled tests in the aquarium helps us understand the complex social interactions of wild dolphins. Research in aquariums has included a test of spontaneous use of water fountains in a prosocial vs selfish manner and Giving Assistance Task in which a dolphin (Tursiops truncatus) could provide shower to another or do nothing. In both tasks the dolphins more frequently chose the prosocial option (Nakahara et al., 2017). However, when dolphins (Tursiops truncatus) were tested in an instrumental helping task, transferring a token or do nothing, they did not spontaneously help their partners (Pérez-Manrique and Gomila, 2019). In another study dolphins (Tursiops spp.) were tested with dynamic stimuli, suggesting that the dolphins were able to recognize prosocial patterns, and also directed high-arousal behaviors toward these displays (Johnson et al., 2018). Finally, when dolphins (Tursiops truncatus) dyads were involved in a Prosocial Choice Tests where they offered the a prosocial (both subject and recipient rewarded), selfish (only subject rewarded) and null options (none rewarded), Lalot et al. (2021) found prosocial and reciprocal tendencies.
In this study, we tested a specifically designed cognitive enrichment that allowed dolphins to engage in prosocial food-sharing actions. The operation of the device required one dolphin’s action of opening and keeping the device open (actor) until another individual retrieved its content (recipient). We monitored the following responses during the test: trial outcomes (number of fish retrieved), the roles (actor or recipient) assumed by the dolphins and any device-related behaviors of the dolphins. We hypothesized that sociability and social dynamics are reflected in the dolphins’ device-related actions. If the dolphins were to engage in prosocial food-sharing actions rather than each individual randomly interacting with the device, we expected to observe higher frequency or longer opening (tugging) events in the presence of other individual(s) than when acting alone.
Materials and methods
Subjects and housing
The experiment focused on social interactions of groups of Indo-Pacific bottlenose dolphins (Tursiops aduncus) living in Ocean Park Hong Kong (OPHK). The dolphins were housed in two facilities of OPHK, called Ocean Theatre (OT) and Marine Mammal Breeding and Research Centre (MMBRC). OT had six inter-connecting pools, the experiment was conducted in pool 1 and 2 (both oval shaped, 15 m L, 7.9 m W and 4 m D). MMBRC also had six interconnecting pools; the experiment was conducted in one of the four main pools (rectangular shaped pools 17-18 m L, 15-20 m W and 4 m D). Two dolphin groups were tested: five adult males (referred to as male group) and six females with a subadult male (referred to as female group). The two groups were housed separately, the male group lived in OT, the female group at MMBRC. The demographics of the dolphins are summarized in Table 1. Besides participating in the weekly research sessions, the dolphins were also involved in daily presentations, educational and encounter programs. The dolphins were fed four times daily with a diet including herring (Culpea sp.), capelin (Mallotus villosus), sardine (Culpeidae sp.), and squid (Coleoidae sp.). The research sessions were conducted during the free-swimming period between feedings. All dolphins previously participated in other cognitive group testing, so that they were familiar with the mutualistically cooperative version of the device (Matrai et al., 2021a, 2021b, 2022a). Relatedness of the interacting pairs was established based on the known pedigree of the dolphins (Table 1).
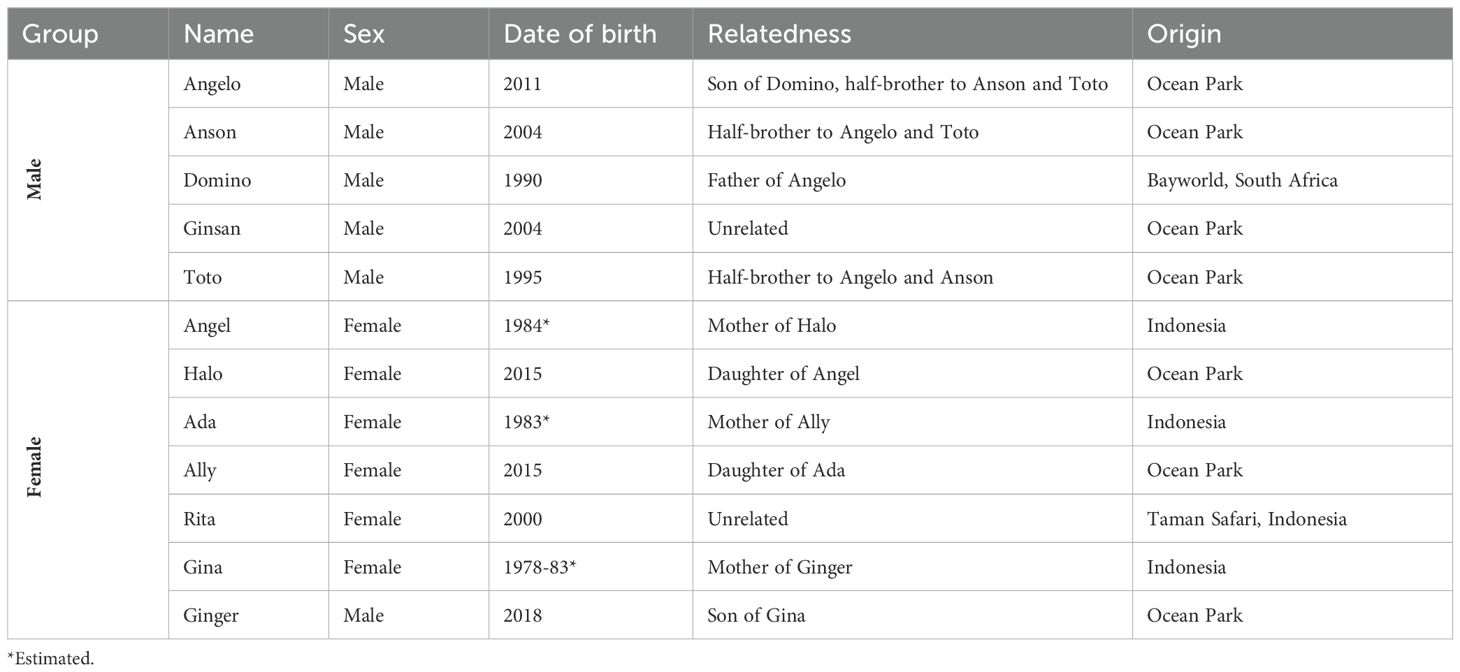
Table 1. Demographic data of the 12 dolphins.
Apparatus
The testing apparatus consisted of a white PVC tube (480 mm, 110 mm, 4 mm wall thickness) with two caps equipped with rope handles (254 mm L and 8 mm diameter rope). The device also had an inner structure with the purpose of positioning and holding five capelins in place (Figure 1). It consisted of two PVC plates securing a clear PVC sheet in between. The PVC plates had five evenly spaced holes (1 cm in diameter), where the PVC sheet was punctured in the shape of a cross for securing the capelins. The PVC plates were fitted inside the tube, fixed to one of the caps and flexibly attached to the other. At the flexible end the PVC plate was attached to the cap with a rubber band and with a 27 cm long safety rope underneath. The rubber band facilitated the opening and reclosing of the device, while the safety rope controlled the maximum length of opening. The device was attached to the poolside with the fixed end, while the flexible handle was available for the dolphins for interaction. Once a dolphin pulled on the handle the plate became accessible and the fish could be removed by another dolphin.

Figure 1. The dimensions and structure of the altruistic food-sharing cognitive enrichment device. The photo on the left shows the device when used by a pair of dolphins (photos were taken by the first author).
Procedure
The study was carried out between February-October 2018 with the male group and July-December 2022 with the female group. Experimental sessions were conducted twice weekly, during the dolphins’ free-swimming time, in between their regular programs. The experiment included three steps with a single device (Step 1), then double device (Step 2), and then again with a single device (Step 3) use. Each step included 10 repeated test sessions; thus 30 sessions were conducted with the male and the female groups, separately. Each session consisted of three repeated trials (total of 90 trial repeats/group). Independently from the dolphins’ response, trials had the same 4-min long duration. Each trial provided opportunity for the dolphins to retrieve five fish from a given device. The device was refilled before each trial, consequently (3x5) 15 fish/device could be obtained during each session. Overall, 600 fish were presented to each group during the experiment, 150-150 during single (Step 1 and 3) and 300 during double device (Step 2) settings. In step 2, the two devices were attached to the same poolside, 255 cm apart at OT and 470 cm at MMBRC (the difference in distance was due to the difference in the design of the pools).
The above-described three steps were designed to investigate the potential differences in fish retrieval and role-preference when more devices could be used (1 vs 2). We have introduced the second device due to the observed sharp differences between dolphins in Step 1; except for a single event, all fish were removed by one dolphin (see results section). We aimed to investigate if the observed difference was due to potential individual limitation in skills or role-preference. Thus, we introduced a second device (the two devices were referred to as device A and B). Finally, in Step 3 we aimed to test the persistence of roles (i.e., role-preference) after the second device was no longer available. The three steps were conducted in succession so that all Step 1 (single device) trial repeats were carried out first (over a period of months), followed by all Step 2 (double-device) test trials, and finally all Step 3 (single-device) trials in the end. The length of the trials (4 min) was selected as an optimal duration for avoiding deterioration of fish (see below). Prior to the sessions, all regular enrichment items were removed from the pool. Each session started with a recall at the poolside, opposite to the position of the device. The dolphins were fed before the session; thus, they were not hungry during the experiment. During this time only ice, jello and secondary reinforcement (e.g., rubbing) were used. This procedure ensured that all dolphins started from the same distance from the device. All dolphins in the group could access the device in each trial. Once the device was prepared, it was placed in the pool in a coordinated action with the dolphins receiving the ‘end of training’ signal. For 4 min the dolphins were allowed to swim freely and all of them had the opportunity to interact with the device(s), then they were recalled to their station. The device(s) was/were refilled with new fish and the same procedure was repeated two more times (three trials/session). Even if the fish was not removed by the dolphins, it was still replaced with new ones. This protocol was adapted to minimize the potential of the inserted fish getting too soft, breaking, or falling out of the device. The dolphins were fully fed before the experimental sessions and received no specific training regarding the operation of the device. No other experiments were performed during the testing of the new food-sharing device.
Data collection and analysis
It was not possible to record data blindly because our study involved focal animals. Two GoPro HERO4 Black (male group), two GoPro HERO6 Black (male and female groups) and two HERO9 (female group) action cameras were used for high quality, underwater video recording for the male and female groups. Moreover, two Sony FDR-AX100 4K Handycam recorders were used for in-air video data collection. The GoPros were mounted on custom-made PVC holders at the poolside.
Behavioral coding of the videos focused on the trial outcomes in terms of fish retrieval (categorical response: 0-5 fish acquired/trial) and on the dolphins’ device-related behavior. We also noted the identity of the participating dolphins and the roles played during fish retrieval. The dolphin pulling the handle and opening the device was considered the ‘actor’, while the dolphin who retrieved the fish was the ‘recipient’. The majority of the fish retrieval was performed in a simultaneous manner; the actor held the device open while the recipient fully removed the fish. However, on a few occasions (10% of the retrievals) the recipient grabbed the fish while the actor kept the device open but only completed the removal after the actor let the handle go. We still consider these events as food-sharing events as without the initial action of the actor the recipient could not acquire the fish.
The behavioral analysis focused on the device-related behaviors that were identified during post-session video analysis. The video recordings were reviewed and analyzed with BORIS (Friard and Gamba, 2016), using focal animal sampling (Altmann, 1974). The same observer re-coded the same video, each time following a different dolphin. The coded behaviors are listed in Table 2. Each video was behaviorally coded by multiple coders (2-3), with high inter-rater reliability index between observers (Cohen’s kappa > 0.7). From the five device-related behaviors only ‘tugging’ (pulling the device open, Table 2) provided opportunity for fish retrieval.

Table 2. List and operational definition of the device-related behaviours.
The difference between the two groups’ response to the device(s) was analyzed for the duration and frequency of their device-related behaviors (investigation, interaction, tug and observation). We summarized each individual’s device-related behaviors (all four behaviors) for each trial (90 trials for all 12 dolphins for frequency and duration data, separately). Statistical analyses were carried out in R statistical environment (version 4.2.2; R Core Team, 2022) and linear mixed effects models (LMMs; R package ‘lme4’, Bates et al., 2015), and cumulative link mixed models (CLMMs; R package ‘ordinal’, Christensen, 2022) were built to analyze the relationship between the sex of the dolphins and the duration or frequency of device-related behaviors, respectively. We included ID as a random effect, while initial models included sex (factor with two levels), steps (factor with three levels) and trials (factor with three levels) as fixed effects. The effects of explanatory variables were analyzed by likelihood ratio tests (LRT), and we provide χ2 and p-values of LRT of models with and without the explanatory variable. Assumptions of models were investigated prior to the analysis. Since residuals in the duration analysis could not be normalized, durations were re-coded into 30 sec interval categories, and we used cumulative link mixed models on these categories.
To further investigate the behavior of the actors, the recorded tugging events were parsed for tugging events that were overlapped by other dolphin’s device-related behaviors and tugging events with no other dolphin present. The former included tugging events that overlapped with another dolphin’s interaction, investigation, or observation. For example, while Anson was tugging the device, Angelo was inspecting the device. The tugging events (both with overlapping and without overlapping device-related behaviors) were analyzed for frequency and duration. The frequency analysis of the overlapped tugging events included two categories. Part of these tugging events were completely overlapped with other device-related events (referred to as complete overlap). In other cases, the overlapping behavior started later or ended earlier than the tugging event itself (referred to as partial overlap). In the duration analysis, the exact overlapping periods were calculated; thus, the duration data had no partial overlap portion.
The tugging events could have multiple overlapping device-related behaviors (for example, while Anson was tugging, Angelo was interacting with the device, and Domino was observing). These could also include more than one dolphin simultaneously engaging in the same device-related behavior that overlapped with a tugging event (for example, while Anson was tugging, Angelo and Domino were interacting with the device). Thus, we analyzed these overlapped tugging events pairwise. We calculated all tugging events with overlap for each dolphin for each device-related behavior. Thus, we had 7 data points for females and 5 for males for tugging overlapped with each device-related behaviors and without overlap. The calculations were made separately for frequency and duration data. The frequency data included both partial and complete overlapped events. The overlapped tugging events were then compared with the tugging events without overlap using paired, two-tailed t-tests.
The frequency and duration of these overlapped tugging events expressed by the various dolphin pairs were correlated with fish retrieval (0-5 fish/trial) of the same pairs (Pearson’s correlation). The five male dolphins provided 20 possible pairwise combinations for prosocial food-sharing and for overlapped tugging events (tugging-interaction, tugging-investigation and tugging-observation). The seven dolphins of the female group provided 42 pairwise combinations.
Results
Food-sharing considering sex differences and role separation
The male group was tested over three experimental steps (90 trials), with 600 fish possible to be acquired of which 192 (32%) were retrieved by one of the dolphins while another one held the device open. Out of the 192 fish, 56 were retrieved in the first step (37% of the maximum 150 fish), 95 in the second step (32% of the maximum 300 fish), and finally 41 in the last step (27% of the maximum 150 fish; Figure 2). The female group was also tested over three experimental steps; however, they were not documented to retrieve any fish in collaboration.
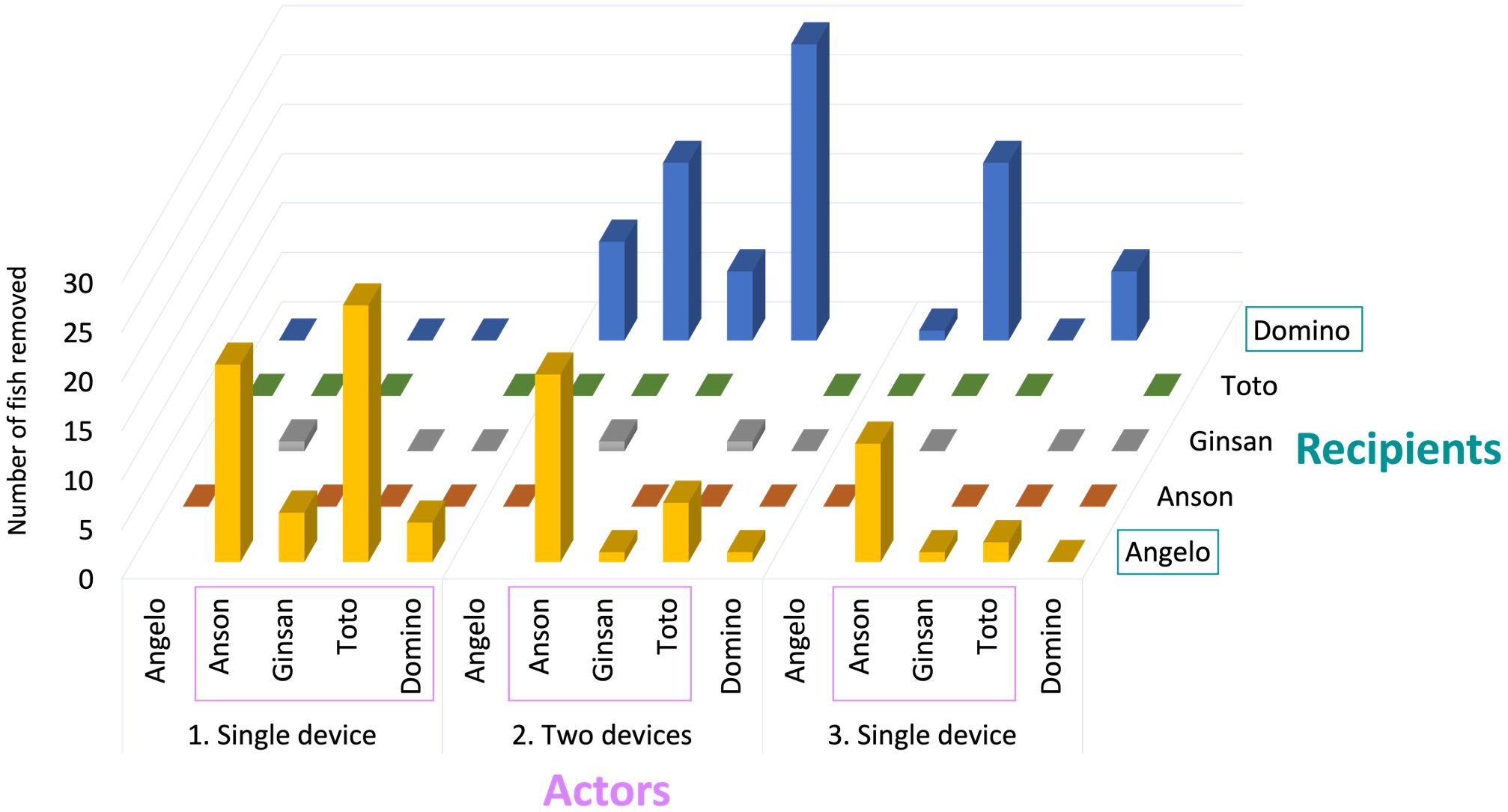
Figure 2. Total number of fish retrieved by male dolphin pairs with one dolphin assuming the role of ‘actor’ and the other the role of ‘recipient’ in prosocial food sharing. Results are provided for each of the three testing steps.
All five male dolphins participated in prosocial food-sharing events, albeit with great inter-individual variability in frequency. Angelo was the most involved, who participated in 108 fish retrievals, while Ginsan was the least involved: he participated only in the retrieval of 17 fish. The remaining 67 fish retrievals were distributed amongst Anson, Toto and Domino. In terms of roles, two of the five male dolphins mostly participated as recipients and three as actors (Figure 2). Only one dolphin, Domino, switched roles from actor in Step 1 to recipient in Step 2 and 3 (Figure 2).
The number of fish retrieval by food-sharing events were very similar between related (53%) and non-related (47%) males (Table 3). Reciprocity, i.e., reversing roles between an actor and recipient was only observed on a single occasion; during the first trial of the fifth session of Step 2: Angelo opened device A and Domino retrieved three fish in succession. About two minutes later Domino opened device B and Angelo retrieved one fish. While the first scenario (Angelo - actor and Domino – recipient) was observed multiple times during Step 2 and 3, the opposite (Domino - actor and Angelo – recipient) was only documented once during the entire experiment (Table 3 and Figure 2).

Table 3. Number of fish retrieved based on relatedness of dolphin pairs.
Despite our best effort of securing the fish in the device and optimizing duration of trials, a small portion of the fish (22 fish for the males and 23 for the females, < 4% of all fish used in the tests) became prematurely soft and came loose during the trials. This very low occurrence, nevertheless, suggests that our devices were robust and suitable to test prosocial food-sharing. Due to the low occurrence of fish loss and the fact that not all five fish were lost in a given trial, we did not exclude these trials from the analyses. While the majority of the fish were retrieved from the device one-by one, on two occasions Angelo managed to remove two fish at the same time (0.6%).
Device-related behaviors
Over the 90 trials, the five males engaged in a total of 19777 device-related actions while the seven females were documented to have 6461 such actions. The number of device-related actions/dolphin were six times more frequent in males than in females (mean ± SD device-related actions/dolphin: 3955 ± 2135 vs 923 ± 555, Figure 3). The frequency of device-related behaviors were different between the sexes (LMM, LRT of effect of sex χ21 = 10.555, P = 0.001; male→female: exp(β) = 0.373 [0.186; 0.751], t = -2.750, P = 0.021) and between testing steps involving different number of devices (LRT: χ22 = 77.698, P < 0.001; Step 1 (1 device)→Step 2 (2 devices): exp(β) = 1.831 [1.438; 2.332], t = 4.877, P < 0.001; Step 1 (1 device)→Step 3 (1 device): exp(β) = 0.536 [0.421; 0.683], t = -5.025, P < 0.001). We also found decreasing frequency of device-related behaviors with trials (LRT: χ22 = 22.456, P < 0.001; trial 1→trial 2: exp(β) = 0.937 [0.735; 1.193], t = -0.527, P = 0.598; trial 1→trial 3: exp(β) = 1.131 [0.888; 1.440], t = 0.991, P = 0.322). In addition, we found sex differences in the effects of experimental step and trial (LRT of sex x experimental step interaction: χ22 = 27.307, P < 0.001; LRT of sex x trial interaction: LRT: χ22 = 27.163, P < 0.001). For pairwise comparisons of estimated marginal means, see Supplementary Material (Supplementary Table 2).
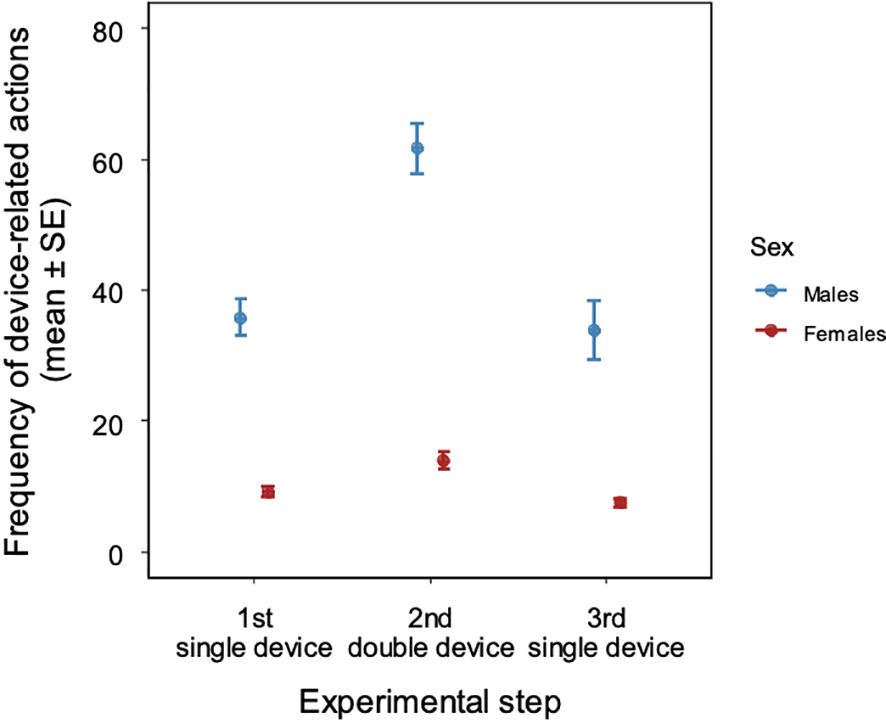
Figure 3. Frequency of device-related actions in male and female dolphins based on availability of prosocial food-sharing devices (‘experimental step’).
In the male group, tugging events were more frequently recorded with the dolphins that participated as actors during fish retrieval than those that participated as recipients (2351 vs 381 events). In the male group 2682, while in the female group 1120 tugging events were recorded. In the female group, the highest number of tugging events were recorded with the three calves (Ally, Halo and Ginger); this constituted 74% of all tugging events and 76% of the total tugging time in females. In the male group 86% of the tugging events were overlapped with at least one other device-related behavior (investigation, interaction or observation) by another dolphin (Table 4 and Figure 4). In the female group, 66% of the tugging events were recorded with overlapping device-related behaviors (Table 4 and Figure 4). When we looked at overlap with investigation, interaction and observation separately, we found that in the male group tugging events were significantly more frequently overlapped with other device-related behaviors. This was true for all three behaviors (Table 4 and Figure 4). In the female group tugging events with and without overlap showed no significant difference (Table 4 and Figure 4).
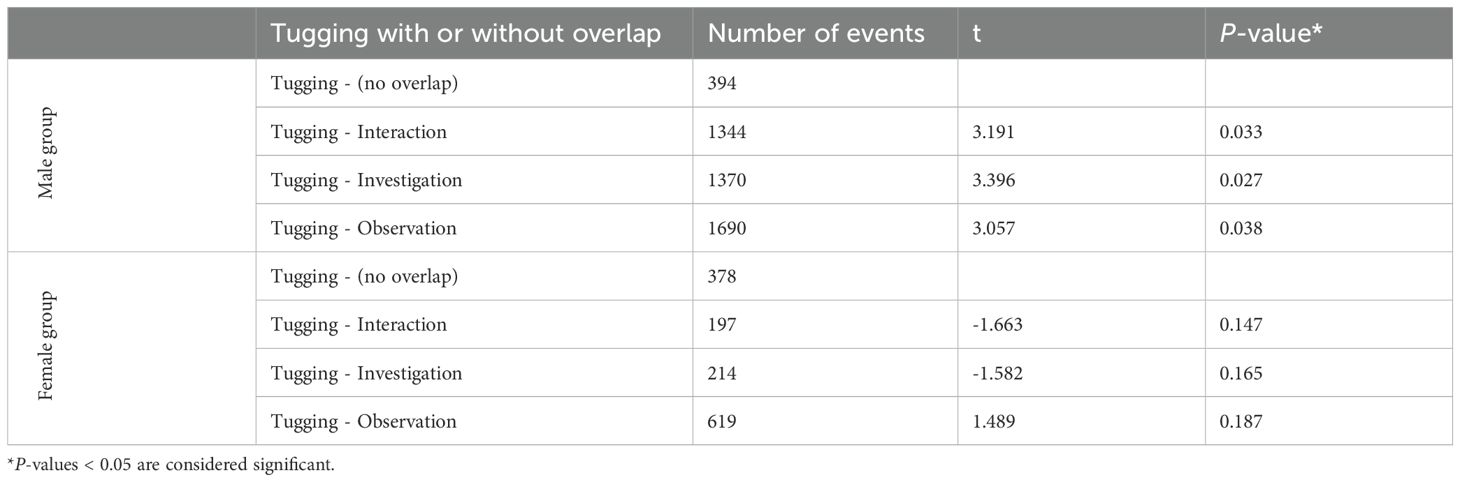
Table 4. Comparison of the frequency of tugging events without overlap vs with overlap (combined complete and partial overlap; paired, two-tailed t-test, df = 4 and df = 6 for males and females, respectively).

Figure 4. Illustration of the four device-related behaviors (left), their frequency recorded with the five male and the seven female dolphins over the 90 trials (middle) and the frequency of tugging events parsed for events that were overlapped with device-related behaviours and events without overlap (right). The asterisk indicate the level of significance, *P < 0.05.
The analysis of the duration of the device-related behaviors corresponded with the frequency analysis above. The males spent significantly more time with device-related behaviors than the females (mean ± SD duration/dolphin: 2 hrs 58 min ± 40 min or 49% ± 11% of testing time vs 1 hr 4 min ± 30 min or 18% ± 8% of testing time; Figure 5). The duration of device-related behaviors were different between the sexes (CLMM, LRT of effect of sex χ21 = 16.649, P < 0.001; male→female: exp(β) = 0.138 [0.055; 0.345], z = -4.243, P < 0.001) and between testing steps involving different number of devices (LRT for effect of the experimental step: χ22 = 41.552, P < 0.001; Step 1 (1 device)→Step 2 (2 devices): exp(β) = 2.370 [1.578; 3.559], z = 4.159, P < 0.001); Step 1 (1 device)→Step 3 (1 device): exp(β) = 0.536 [0.421; 0.683], z = -5.025, P < 0.001). We also found decreasing duration of device-related behaviors with trials (LRT for effect on trial: χ22 = 33.905, P < 0.001; trial 1→trial 2: exp(β) = 0.870 [0.582; 1.303], z = -0.674, P = 0.5; trial 1→trial 3: exp(β) = 0.906 [0.607; 1.355], z = -0.479, P = 0.632). In addition, we found sex differences in the effects of experimental step in relation to trial (LRT of sex x experimental step interaction: χ22 = 40.337, P < 0.001; LRT of sex x trial interaction: χ22 = 19.953, P < 0.001). For pairwise comparisons of estimated marginal means, see Supplementary Material (Supplementary Table 3).
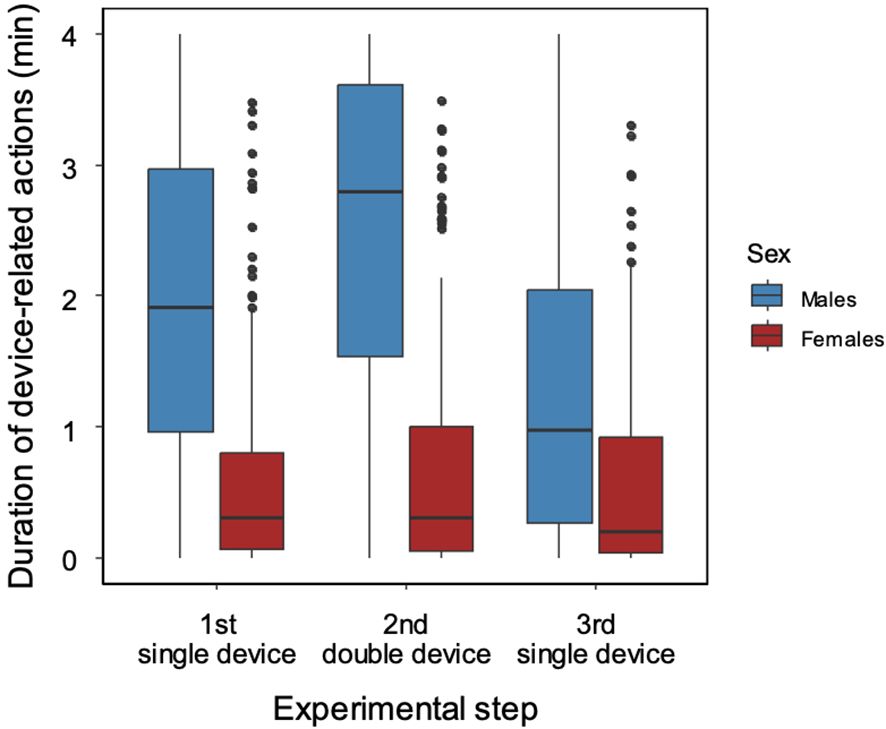
Figure 5. Duration of device-related actions in male and female dolphins based on availability of prosocial food-sharing devices (‘experimental step’).
Actors spent longer time tugging than recipients (56 min vs 19 min). The five males spent 1 hr 15 min tugging, of which 89% was recorded with overlapping device-related behaviors (investigation, interaction or observation). The seven females were observed to spend 38 min tugging, of which 71% overlapped. In the male group, the duration of tugging events was all significantly longer when overlapped with other device-related behaviors (Table 5 and Figure 6). In the female group tugging with or without overlapping with other device-related behaviors was not significantly different (Table 5 and Figure 6).
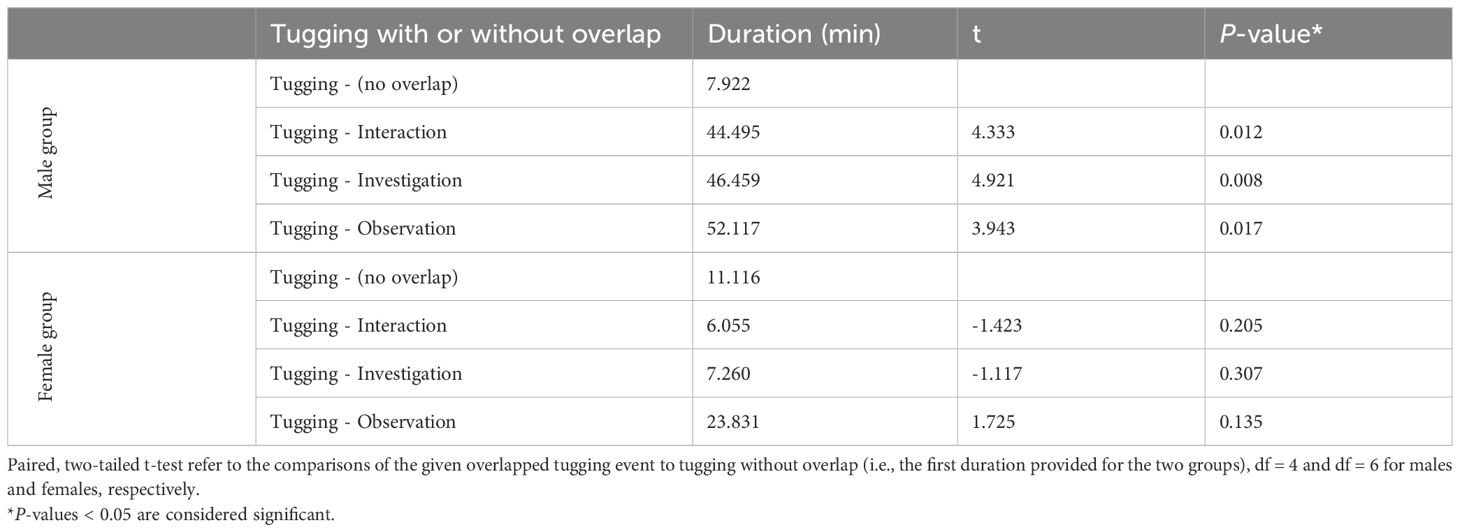
Table 5. Comparison of the duration of tugging events when these are performed in solo (no overlap) vs when overlapped by other one or more dolphin’s various device-related behaviours.
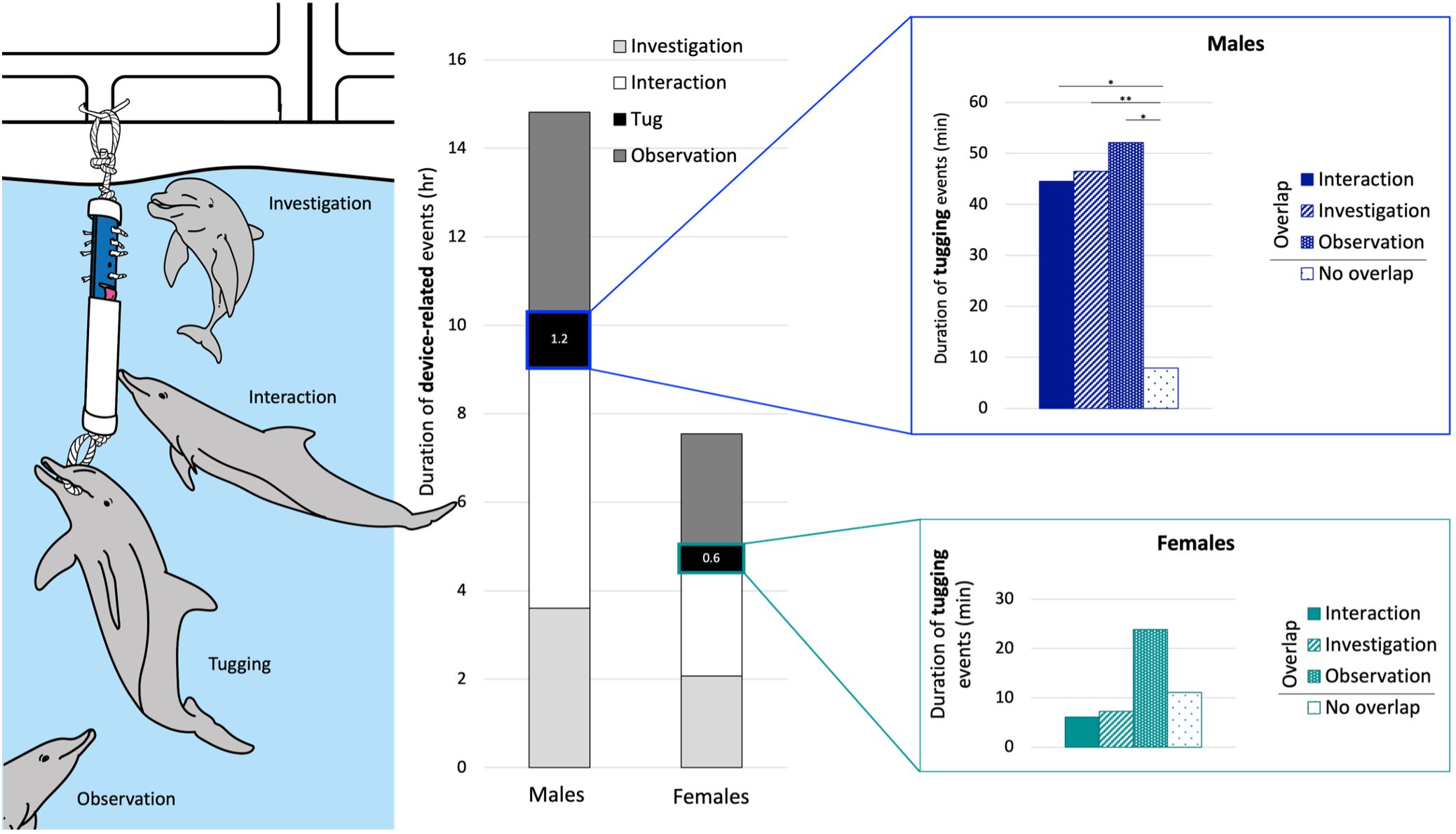
Figure 6. Illustration of the four device-related behaviors (left), their duration of the four device-related behaviors recorded with the five male and the seven female dolphins over the 90 trials (middle) and the duration of tugging events parsed for events that were overlapped with device-related behaviours and events without overlap (right). Asterisk indicate the level of significance, * P < 0.05 and ** P < 0.01.
Finally, strong correlation was found between overlapped tugging behaviors and fish retrieval in males. Dolphin pairs that engaged more frequently and/or longer in simultaneous tugging-interaction, tugging-investigation or tugging-observation were also more likely to participate in the fish removal (all Pearson’s r > 0.73, P < 0.001, Table 6 and Figure 7). Moreover, when overlapped tugging events were compared with each other, we also found a strong positive correlation between all three of these in males (Table 7 and Figure 7), but not in females (tugging-observation with the other two behaviors showed only moderate or low correlation; Table 7).
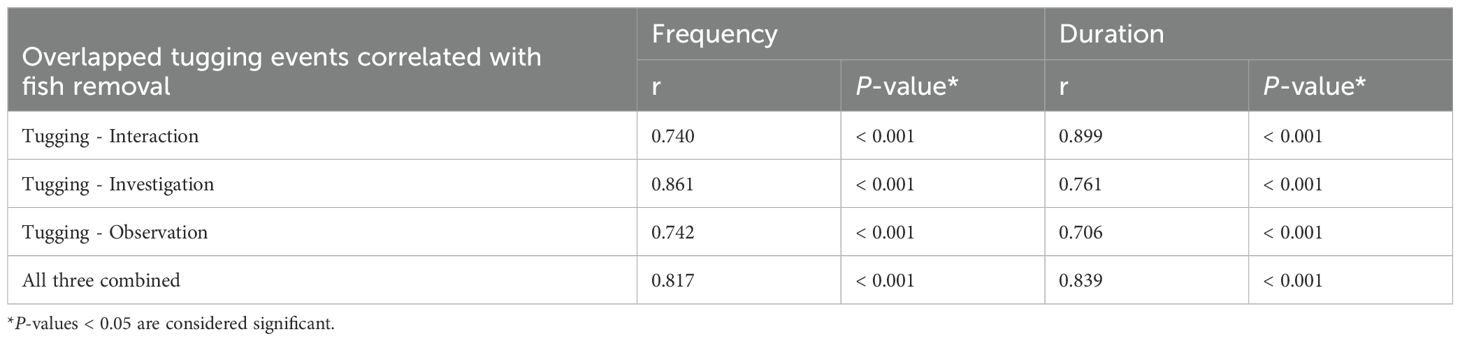
Table 6. Correlation (Pearson’s r) between simultaneous device-related behaviours of actors and recipients and fish retrieval in the 20 possible pairing of the five male dolphins.
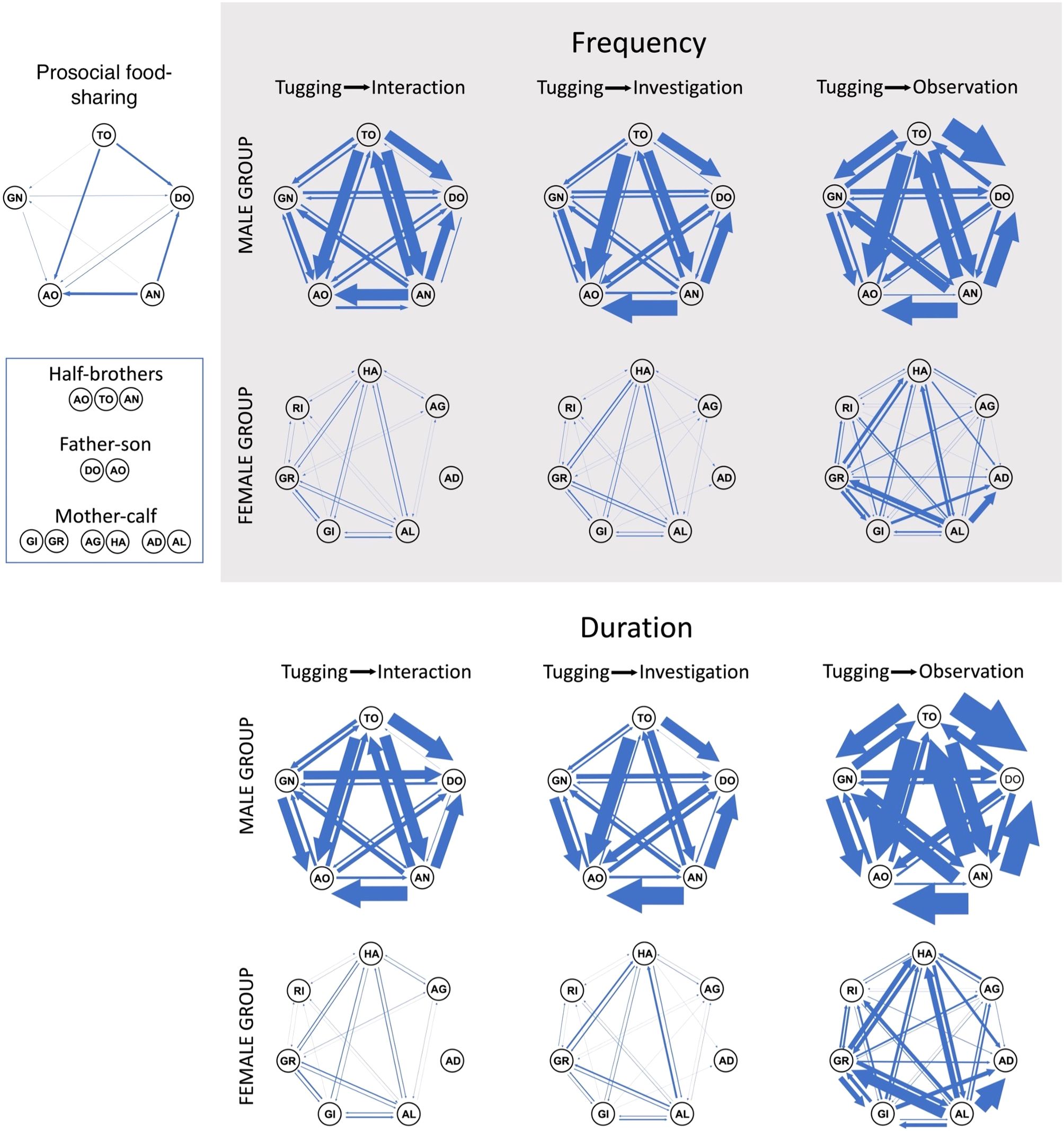
Figure 7. Sociograms depicting the fish retrieval and overlapped tugging events between the five male dolphins (AN, Anson; AO, Angelo; DO, Domino; GN, Ginsan; TO, Toto) and overlapped tugging events between the seven dolphins of the female group (GI, Gina; GR, Ginger; AG, Angel; HA, Halo; AD, Ada; AL, Ally; RI, Rita); the thickness of arrows represent interactions weighted by to the frequency or duration of the events. Arrow heads correspond to the direction of the actions (actor → recipient).
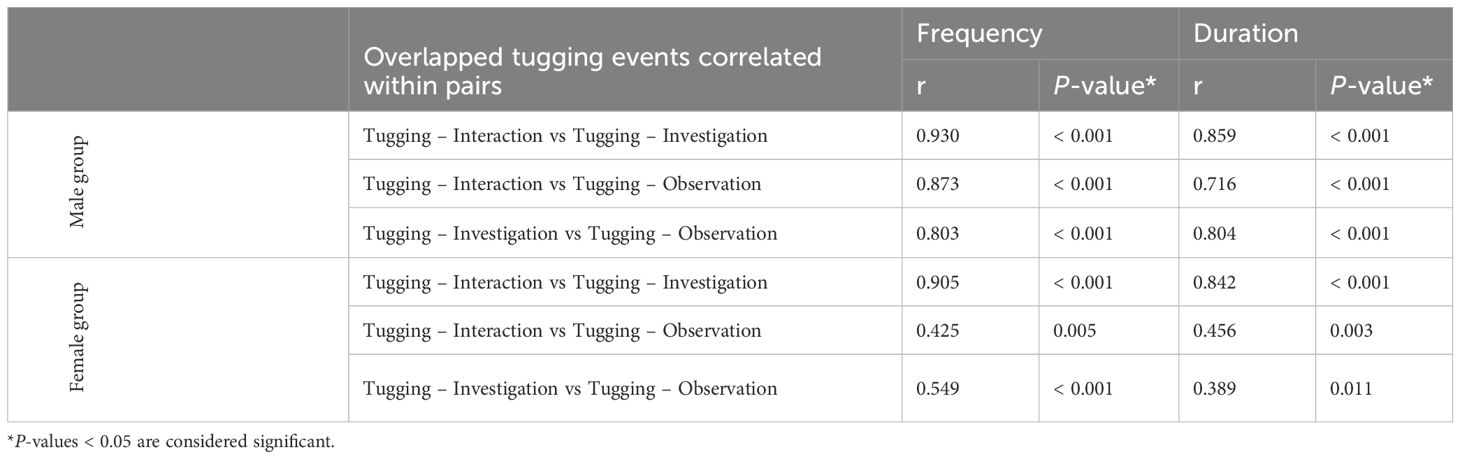
Table 7. Correlation (Pearson’s r) between different simultaneous (overlapping) device-related behaviour pairs in the 20 possible pairing of the five male and the 42 possible pairings of the seven female dolphins.
Discussion
Our experiment provided evidence for the first successful use of a cognitive enrichment device designed to allow prosocial food-sharing in dolphins. All five male but none of the female dolphins participated in the food-sharing, whereby one dolphin pulled the device open so that the other could retrieve and consume the fish. Although the male group size was smaller than the female group, male dolphins showed a higher level of participation in this task and demonstrated role separation. Males showed a higher level of tugging in the presence of other dolphin(s). In females, tugging showed no significant difference in the presence or absence of other dolphins. These sex differences were revealed by both frequency and duration data analysis. Finally, the involvement of the different male dolphins in the overlapped tugging events highly correlated with their food-sharing success. This correlation suggests an association between the two and confirms the prosocial use of this device.
Cognitive enrichments have gained popularity over the past decades due to their benefits in terms of welfare and science, as have been repeatedly confirmed (Morimura, 2006; Puppe et al., 2007; Manteuffel et al., 2009; Meyer et al., 2010; Clark, 2013; Clark et al., 2013; Zebunke et al., 2013; Clark, 2017; Matrai et al., 2022b). The design of cognitive enrichments for cetaceans mainly focused on problem-solving (Clark et al., 2013) and cooperation (Kuczaj et al., 2015; Matrai et al., 2020; 2021a; 2021b; 2022a). Dolphins (Tursiops aduncus) successfully learnt to manipulate these novel enrichments without receiving any specific training to operate them and invented a novel cooperative behavior referred to as cooperative play (Matrai et al., 2021a, 2021b, 2022a). However, in our previous studies, the cooperative manipulation of the devices allowed all collaborators to benefit directly from the action. Once the dolphins opened the device, its content became available to all participants. The dolphins shared the content of the devices, rendering the cooperative action mutualistic. Cetaceans in the wild might engage in cooperative actions in which the imminent benefits to the participants may be unequal, such as during group hunting (Gazda et al., 2005) or during alloparental associations (Gero et al., 2009). Thus, in this study, we aimed to test a cognitive enrichment device that was specifically designed for cooperative use where one dolphin, the actor, would temporarily forfeit access to the fish reinforcement and simultaneously provide access for another individual, the recipient, in a prosocial fashion. The nature of the dolphins’ actions was determined by their device-related behavior over the session time. The tugging events in the male group were significantly longer in the presence of other dolphins, suggesting prosocial tendencies.
Our findings, therefore, corroborate the prosocial nature of the food-sharing in males; only males (but not females) increased their tugging (both in frequency and duration) if the other dolphins were in the vicinity, independently from their distance of the device. In contrast, females only tugged somewhat longer (the difference was not statistically significant), while the other dolphin(s) were further away from the device and tended to reduce tugging (not statistically significantly) when the other(s) was(were) as close or closer to the device than themselves suggesting selfish, rather than cooperative, interest. Moreover, most of the tugging in the female group was recorded with the three subadult dolphins (Ally, Ginger and Halo) and most of the observations were also recorded by the three of them watching their mothers or each other. Observational learning plays an important role in dolphins’ lives (Yeater and Kuczaj, 2010; Kuczaj et al., 2012). For example, dolphin calves were documented to learn specific foraging strategies by observing their mothers (Boran and Heimlich, 1999; Sargeant and Mann, 2009). In another study on social learning in dolphins, Kuczaj et al. (2012) found that dolphin calves were selective in choosing who to observe and/or imitate. They were more often recorded to watch the play behavior of other calves than adults (including both related and non-related individuals). Thus, the observation events of the female group in our study were more likely to reflect on the subadults’ predisposition towards watching others as part of learning skills rather than their interest towards prosocial food-sharing. The difference in the potential intentionality of the two groups was further supported by the strong correlation between all overlapped tugging events in males but not in females.
Both fish retrieval and device-related actions showed a marked difference towards the male group. Only males obtained fish and were more likely to interact with the device in the presence of other males. The difference in cooperative actions between the male and the female group is in line with our previous findings; males were also more likely to open the mutual cooperative enrichment devices than females (Matrai et al., 2021a). Moreover, while females were not at all, males were recorded to engage in cooperative play up to 90% of the testing time (Matrai et al., 2021b). When participation in enrichment sessions was investigated between the sexes, Eskelinen et al. (2015) also reported a greater level of participation in males. In addition, when a novel visual enrichment was introduced, males were reported to have a more extended response (Winship and Eskelinen, 2018). In other studies, however, females were reported to exhibit a higher rate of interactions with novel enrichments (Lopes et al., 2016). These findings suggest a difference in inter-sexual predisposition towards enrichments, which may partially explain the difference in the dolphins’ device-related behaviors. However, the sharp contrast in the two groups’ tendency to cooperate (with complete absence of cooperative play in females, Matrai et al., 2021a) indicates that the difference lies not in whether to play or not to play but in how to play. Studies have shown that the presence of a younger individual may influence the mother’s behavior, as the juveniles tend to absorb more attention (Clay and de Waal, 2013), hence decreasing affiliative behaviors among adult females (Fultz et al., 2022) and altering the social structure of the group (Kirk and Wascher, 2018). However, in our study the three subadult dolphins were all older (five- and seven-year-old), independent dolphins. In addition, using of the cooperative enrichments, we documented interactions between all individuals in the group (unpublished data). Thus, we do not believe that the presence of the younger animals reduced the adult females’ device-related behaviors. A possible functional explanation is that these sex differences may reflect on the differences in the natural alliance formation tendencies of the two sexes (Connor et al., 2000; Connor, 2007). Association between females in the wild have been reported with other females with dependent calves and was described as a loosely connected network (Whitehead and Mann, 2000; Frere et al., 2010) rather than long-term alliances that were observed in males (Connor et al., 2000; Connor, 2007). Thus, the consistently recurring sex differences in various experimental test paradigms corroborate our current and previous findings and are in line with observations in the wild.
Role separation could be investigated with the active engagement in prosocial food-sharing of all five male dolphins. Of the five males, only one, Domino, switched roles from actor to recipient, while Angelo remained recipient, Anson and Toto stayed actors. The observed role separation could be due to differences in individual preferences for a given action (pulling the handle or taking the fish) or due to the coordinated division of ‘labor’. While in Step 1, differences in skills could also contribute to role separation (it is possible that not all five males learnt to engage with the device in both roles), Step 2 and 3 proved that experience was not a likely influencing factor. In Step 2, all five males demonstrated their abilities to participate as actors and four of them engaged in both roles. However, in Step 3, with a single device again, the dolphins reverted to nearly exclusively participating in one role only. Moreover, the strong correlations between the fish retrieval and the frequency and duration of overlapped tugging events suggest that role separation was consistent and maintained over time. Role separation in cooperative actions in the wild was documented during group hunting; Gazda et al. (2005) reported division of labor in cooperative hunting by a single dolphin acting as a ‘driver’ herding the fish towards the rest of the group, the ‘non-drivers’. The mud ring feeding strategy provides another example of role specialization in cooperating hunting. The ‘ring-maker’ individual circles around the fish, beating his tail to create the mud ring, while the other group members assist by herding the fish (Engleby and Powell, 2019). Role specialization has also been reported during travelling; Lewis et al. (2011) observed that certain dolphins of the group consistently lead the group during travelling (Lewis et al., 2011). Role separation in cooperative hunting and leadership was observed to be persistent. The drivers of the two groups did not change during the observation period (Gazda et al., 2005). The dolphins (Tursiops aduncus) in the Lower Florida Keys all spend little time leading group movements, but a few individuals assumed consistent leadership (Lewis et al., 2011). While the role of fidelity in cooperative hunting is not yet fully understood, Lewis et al. (2013) found a positive correlation between leader-follower associations and relatedness.
Food-sharing, when associated with real fitness consequences, can be considered altruistic. In our experiment, the actor dolphin did not have access to fish, only the recipient. However, the associated fitness costs/benefits in terms of resources are arguably negligible. Therefore, future studies should investigate this possible explanation by testing food-sharing when its associated costs and benefits are more expressed. The proportion of food-sharing between kin and non-kin was near equal. Although are relatively low sample size in terms of related vs non-related dyads did not allow to test the role of relatedness, the similar number of fish retrieved make relatedness an unlikely explanation behind the observed prosocial behavior. Domino (father) and Angelo (son) were the only pair in which both dolphins took both roles and opened the device for each other. Their actions were mainly separated over the three testing steps and represented a relatively small percentage of the fish they each retrieved. Thus, it is difficult to determine if the dolphins expressed reciprocity. The frequency and duration of overlapped tugging events for each other showed a slight preference over other dolphins; however, there was a similar trend observed with the remaining three dolphins, as they also tugged for Angelo and Domino the most. Therefore, these patterns reflect more likely on the preferred roles of Domino and Angelo.
Age is also unlikely to strongly affect the dolphins’ role preference, as the two recipients, Angelo and Domino, were the youngest and the oldest animals in the group. While personality traits were not investigated in this study, it may provide an explanation for role preference. For instance, as Domino was the father of Angelo, he might have passed on certain personality traits to his son, and this may explain their disposition towards the recipient role. In line with this, dolphins under human care have been reported to have distinct and well-defined personalities (Highfill and Kuczaj, 2007, 2010). Furthermore, differences in personality have been reported to contribute to dyadic bond formation (Moreno, 2017) and influence the dolphins’ actions in a cooperative task (Bagley et al., 2020). Finally, avoidance of punishment and manipulative strategies, especially in social organizations that include dominance hierarchy (Cant and Johnstone, 2006; Clutton-Brock and Parker, 1995), have been observed as other potential drivers of apparently altruistic behaviors. In contrast to this explanation, while we acknowledge that it cannot be ruled out, we observed no aggressive behaviors (chasing, biting, or pushing) during the experiment. In fact, in a long-term (three years) evaluation of the use of the cognitive enrichments, also including the food-sharing device of this study, the dolphins were found to be more affiliative with each other in non-research periods of the days when research sessions were conducted (Matrai et al., 2022b).
We acknowledge the limitations of our study, specifically, the involvement of only two same-sex groups and no mix-sex combination, and the relatively low sample size that does not allow us to exclude all alternative explanations behind the observed patterns of prosocial food-sharing. Given the limited literature on the topic, we argue that our findings provide a basis for future studies, investigating if sex differences recorded in our study would be corroborated in other captive groups and whether males would cooperate with females if they had them as available partners. Prosocial actions have been observed in wild societies, however, the proximate mechanism of these actions is not always clear. While cetaceans possess the cognitive capacity and social structure favorable for prosocial actions, controlled tests are lacking. Cognitive enrichment studies with controlled group settings and repeated trials can offer insights and bring scientists closer to understanding prosocial actions in the wild.
In conclusion, our study has successfully demonstrated the first application of a prosocial cognitive enrichment in dolphins under human care, using a novel food-sharing device. Both males and females engaged in device-related actions, but it was only in males that prosocial food-sharing was notably observed. These preliminary findings, particularly the unique role separation observed, provide the foundation for further investigations into the long-term nature, function, and emergence of socio-cognitive skills and cooperation and may contribute to our understanding of social behaviors and dynamics in cetaceans.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Animal Welfare and Ethics Committee of Ocean Park Hong Kong. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
EM: Conceptualization, Data curation, Formal analysis, Investigation, Supervision, Visualization, Writing – original draft, Writing – review & editing. SK: Data curation, Investigation, Methodology, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing. ZT: Data curation, Investigation, Methodology, Project administration, Validation, Writing – original draft, Writing – review & editing. XL: Formal analysis, Methodology, Software, Writing – original draft, Writing – review & editing. WL: Data curation, Investigation, Project administration, Validation, Writing – original draft, Writing – review & editing. PM: Conceptualization, Supervision, Visualization, Writing – original draft, Writing – review & editing. ÁP: Conceptualization, Formal analysis, Investigation, Methodology, Software, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
We would like to thank the trainers of the Marine Mammal Department of Ocean Park for their help and support during the experimental session and the interns and volunteers that made this research possible. Special thanks to WeiKiat Quek, for his immense help in the data analysis. We would also like to thank the trainers for their valuable contribution in conducting the experimental sessions. We thank the two reviewers for their constructive suggestions to improve a previous version of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2024.1479989/full#supplementary-material
References
Altmann J. (1974). Observational studies of behavior: sampling methods. Behaviour 49, 227–267. doi: 10.1163/156853974X00534
Augusto J. F., Frasier T. R., Whitehead H. (2017). Characterizing alloparental care in the pilot whale (Globicephala melas) population that summers off Cape Breton, Nova Scotia, Canada. Mar. Mammal Sci. 33 (2), 440–456. doi: 10.1111/mms.12377
Bagley K. C., Winship K., Bolton T., Foerder P. (2020). Personality and Affiliation in a Cooperative Task for Bottlenose Dolphin (Tursiops truncatus) Dyads. Int. J. Comp. Psychol. 33. Available at: https://escholarship.org/uc/item/5n43p6d7.
Bates D., Mächler M., Bolker B., Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Bender C. E., Herzing D. L., Bjorklund D. F. (2009). Evidence of teaching in Atlantic spotted dolphins (Stenella frontalis) by mother dolphins foraging in the presence of their calves. Anim. Cogn. 12, 43–53. doi: 10.1007/s10071-008-0169-9
Benoit-Bird K. J., Au W. W. (2009). Cooperative prey herding by the pelagic dolphin, Stenella longirostris. J. Acoust. Sociecty America 125, 125–137. doi: 10.1121/1.2967480
Boran J., Heimlich S. (1999). “Social learning in cetaceans: hunting, hearing and hierarchies,”. in Mammalian social learning. Eds. Box H., Gibson K. (Cambridge: Cambridge University Press), 282–307.
Brower K., Curtsinger W. R. (1979). “The blue water,” in Wake of the whale (Dutton Adult, New York, NY), 128–153.
Cant M. A., Johnstone R. A. (2006). Self-serving punishment and the evolution of cooperation. J. Evol. Biol. 19, 1383–1385. doi: 10.1111/j.1420-9101.2006.01151.x
Carter G. G., Wilkinson G. S. (2015). Intranasal oxytocin increases social grooming and food sharing in the common vampire bat Desmodus rotundus. Hormones Behav. 75, 150–153. doi: 10.1016/j.yhbeh.2015.10.006
Christensen R. H. B. (2022). Regression Models for Ordinal Data [R package ordinal version 2022.11-16].
Clark F. E. (2013). Marine mammal cognition and captive care: A proposal for cognitive enrichment in zoos and aquariums. J. Zoo Aquarium Res. 1, 1–6. doi: 10.19227/jzar.v1i1.19
Clark F. E. (2017). Cognitive enrichment and welfare: Current approaches and future directions. Anim. Behav. Cogn. 4, 52–71. doi: 10.12966/abc.05.02.2017
Clark F. E., Davies S. L., Madigan A. W., Warner A. J., Kuczaj S. A. (2013). Cognitive enrichment for bottlenose dolphins (Tursiops truncatus): evaluation of a novel underwater maze device. Zoo Biol. 32, 608–619. doi: 10.1002/zoo.21096
Clay Z., de Waal F. B. M. (2013). Bonobos respond to distress in others: consolation across the age spectrum. PloS One 8, e55206. doi: 10.1371/journal.pone.0055206
Clegg I. L., Rödel H. G., Delfour F. (2017). Bottlenose dolphins engaging in more social affiliative behaviour judge ambiguous cues more optimistically. Behav. Brain Res. 30, 115–122. doi: 10.1016/j.bbr.2017.01.026
Clutton-Brock T. H., Parker G. A. (1995). Punishment in animal societies. Nature 373, 209–216. doi: 10.1038/373209a0
Connor R. C. (2007). Dolphin social intelligence: complex alliance relationships in bottlenose dolphins and a consideration of selective environments for extreme brain size evolution in mammals. Philos. Trans. R. Soc. B: Biol. Sci. 362, 587–602. doi: 10.1098/rstb.2006.1997
Connor R. C., Krützen M. (2015). Male dolphin alliances in Shark Bay: changing perspectives in a 30-year study. Anim. Behav. 103, 223–235. doi: 10.1016/j.anbehav.2015.02.019
Connor R. C., Smolker R., Bejder L. (2006). Synchrony, social behaviour and alliance affiliation in Indian Ocean bottlenose dolphins, Tursiops aduncus. Anim. Behav. 72, 1371–1378. doi: 10.1016/j.anbehav.2006.03.014
Connor R. C., Wells R. S., Mann J., Read A. J. (2000). “The bottlenose dolphin: social relationships in a fission-fusion society,” in Cetacean Societies: Field Studies of Dolphins and Whales. Eds. Mann J., Connor R. C., Tyack P. L., Whitehead H. (University of Chicago Press, Chicago, IL), pp 91–pp126.
Deakos M. H., Branstetter B. K., Mazzuca L., Fertl D., Mobley J. R. (2010). Two Unusual Interactions Between a Bottlenose Dolphin (Tursiops truncatus) and a Humpback Whale (Megaptera novaeangliae) in Hawaiian Waters. Aquat. Mammals 36, 121–128. doi: 10.1578/AM.36.2.2010.121
de Waal F. B. M. (2008). Putting the altruism back into altruism: the evolution of empathy. Annu. Rev. Psychol. 59, 279–300. doi: 10.1146/annurev.psych.59.103006.093625
Engleby L. K., Powell J. R. (2019). Detailed observations and mechanisms of mud ring feeding by common bottlenose dolphins (Tursiops truncatus truncatus) in Florida Bay, Florida, U.S.A. Mar. Mammal Sci. 35, 1162–1172. doi: 10.1111/mms.12583
Eskelinen H. C., Winship K. A., Borger-Turner J. L. (2015). Sex, age, and individual differences in bottlenose dolphins (Tursiops truncatus) in response to environmental enrichment. Anim. Behav. Cogn. 2, 241–253. doi: 10.12966/abc.08.04.2015
Fedorowicz S. M., Beard D. A., Connor R. C. (2003). Food sharing in wild bottlenose dolphins. Aquat. Mammals 29, 355–359. doi: 10.1578/01675420360736523
Fellner W., Bauer G. B., Stamper S. A., Losch B. A., Dahood A. (2013). The development of synchronous movement by bottlenose dolphins (Tursiops truncatus). Mar. Mammal Sci. 29, E203–E225. doi: 10.1111/j.1748-7692.2012.00609.x
Frere C. H., Krützen M., Mann J., Watson-Capps J. J., Tsai Y. J., Patterson E. M., et al. (2010). Home range overlap, matrilineal and biparental kinship drive female associations in bottlenose dolphins. Anim. Behav. 80, 481–486. doi: 10.1016/j.anbehav.2017.04.011
Friard O., Gamba M. (2016). BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 7, 1325–1330. doi: 10.1111/2041-210X.12584
Fultz A., Yanagi A., Breaux S., Beaupre L. (2022). Aggressive, submissive, and affiliative behavior in sanctuary chimpanzees (Pan troglodytes) during social integration. Animals 12, 2421. doi: 10.3390/ani12182421
Gazda S. K., Connor R. C., Edgar R. K., Cox F. (2005). A division of labour with role specialization in group-hunting bottlenose dolphins (Tursiops truncatus) off Cedar Key, Florida. Proc. R. Soc. B: Biol. Sci. 272, 135–140. doi: 10.1098/rspb.2004.2937
Gero S., Engelhaupt D., Rendell L., Whitehead H. (2009). Who cares? Between-group variation in alloparental caregiving in sperm whales. Behav. Ecol. 20, 838–843. doi: 10.1093/beheco/arp068
Guinet C., Bouvier J. (1995). Development of intentional stranding hunting techniques in killer whale (Orcinus orca) calves at Crozet Archipelago. Can. J. Zool. 73, 27–33. doi: 10.1139/z95-004
Herzing D. L. (1996). Vocalizations and Associated Underwater Behavior of Free-ranging Atlantic Spotted Dolphins, Stenella frontalis and Bottlenose Dolphins, Tursiops truncatus. Aquat. Mammals 22, 61–79. doi: 10.12966/abc.02.02.2015
Herzing D. L. (1997). The life history of free-ranging Atlantic spotted dolphins (Stenella frontalis): age classes, color phases, and female reproduction. Mar. Mamm. Sci. 13, 576–595. doi: 10.1111/j.1748-7692.1997.tb00085.x
Highfill L. E., Kuczaj S. A. (2007). Do bottlenose dolphins (Tursiops truncatus) have distinct and stable personalities? Aquat. Mamm. 33, 380–398. doi: 10.1578/AM.33.3.2007.380
Highfill L. E., Kuczaj S. A. II (2010). How studies of wild and captive dolphins contribute to our understanding of individual differences and personality. Int. J. Comp. Psychol. 23. Available at: https://escholarship.org/uc/item/0xb2s7g3.
Holmes B. J., Neil D. T. (2012). Gift giving” by wild bottlenose dolphins (Tursiops sp.) to humans at a wild dolphin provisioning program, Tangalooma, Australia. Anthrozoos: A Multidiscip. J. Interact. People Anim. 25, 397–413. doi: 10.2752/175303712X13479798785698
Jaakkola K., Guarino E., Donegan K., King S. L. (2018). Bottlenose dolphins can understand their partner’s role in a cooperative task. Proc. R. Soc. B. 285, 20180948. doi: 10.1098/rspb.2018.0948
Jean-Loup R. (2019). Be kind to others: Prosocial behaviours and their implications for animal welfare. Appl. Anim. Behav. Sci. 210, 113–123. doi: 10.1016/j.applanim.2018.10.015
Jiménez P. J., Alava J. J. (2015). Strand-feeding by coastal bottlenose dolphins (Tursiops truncatus) in the Gulf of Guayaquil, Ecuador. Latin Am. J. Aquat. Mammals 10, 33–37. doi: 10.5597/lajam00191
Johnson C. M., Sullivan J., Jensen J., Buck C., Trexel J., St. Leger J. (2018). Prosocial predictions by bottlenose dolphins (Tursiops spp.) based on motion patterns in visual stimuli. psychol. Sci. 29, 1405–1413. doi: 10.1177/095679761877107
Kirk J., Wascher C. A. (2018). Temporal modification of social interactions in response to changing group demographics and offspring maturation in African lions (Panthera leo). Behav. Process. 157, 519–527. doi: 10.1016/j.beproc.2018.06.006
Krützen M., Sherwin W. B., Connor R. C., Barre L. M., Van de Casteele T., Mann J., et al. (2003). Contrasting relatedness patterns in bottlenose dolphins (Tursiops sp.) with different alliance strategies. Proc. R. Soc. B: Biol. Sci. 270, 497–502. doi: 10.1098/rspb.2002.2229
Kuczaj I. S. A., Winship K. A., Eskelinen H. C. (2015). Can bottlenose dolphins (Tursiops truncatus) cooperate when solving a novel task? Anim. Cogn. 18, 543–550. doi: 10.1007/s10071-014-0822-4
Kuczaj S. A. II, Yeater D. B. (2007). Observations of rough-toothed dolphins (Steno bredanensis) off Utila, Honduras. J. Mar. Biol. Assoc. United Kingdom 87, 141–148. doi: 10.1017/S0025315407054999
Kuczaj S. A. II, Yeater D., Highfill L. (2012). How selective is social learning in dolphins? Int. J. Of Comp. Psychol. 25, 221–236. doi: 10.46867/ijcp.2012.25.03.02
Lalot M., Delfour F., Mercera B., Bovet D. (2021). Prosociality and reciprocity in bottlenose dolphins (Tursiops truncatus). Anim. Cogn. 24, 1075–1086. doi: 10.1007/s10071-021-01499-z
Lewis J. S., Wartzok. D., Heithaus M. R. (2011). Highly dynamic fission–fusion species can exhibit leadership when traveling. Behav. Ecol. Sociobio. 65, 1061–1069. doi: 10.1007/s00265-010-1113-y
Lewis J. S., Wartzok D., Heithaus M., Krutzen M. (2013). Could relatedness help explain why individuals lead in bottlenose dolphin groups? PloS One 8, e58162. doi: 10.1371/journal.pone.0058162
Lopes M., Borger-Turner J. L., Eskelinen H. C., Kuczaj S. A. II (2016). The influence of age, sex, and social affiliation on the responses of bottlenose dolphins (Tursiops truncatus) to a novel stimulus over time. Anim. Behav. Cogn. 3, 32–45. doi: 10.12966/abc.02.03.2016
Lukas M., Toth I., Reber S., Slattery D. A., Veenema A. H., Neumann I. D. (2011). The neuropeptide oxytocin facilitates pro-social behavior and prevents social avoidance in rats and mice. Neuropsychopharmacol 36, 2159–2168. doi: 10.1038/npp.2011.95
MacLeod C. D., D’Amico A. (2006). A review of beaked whale behaviour and ecology in relation to assessing and mitigating impacts of anthropogenic noise. J. Cetacean Res. Manag. 7, 211–221. doi: 10.47536/jcrm.v7i3.731
Manteuffel G., Langbein J., Puppe B. (2009). From operant learning to cognitive enrichment in farm animal housing: bases and applicability. Anim. Welf. 18, 87–95. doi: 10.1017/S0962728600000105
Martin A. R., Da Silva V. M. F. (2018). Reproductive parameters of the Amazon river dolphin or boto, Inia geoffrensis (Cetacea: Iniidae); an evolutionary outlier bucks no trends. Biol. J. Linn. Soc. 123, 666–676. doi: 10.1093/biolinnean/bly005
Matrai E., Gendron S. M., Boos M., Pogány Á. (2022b). Cognitive group testing promotes affiliative behaviors in dolphins. J. Appl. Anim. Welf. Sci. 27 (1), 165–179. doi: 10.1080/10888705.2022.2149267
Matrai E., Kwok S. T., Boos M., Pogany A. (2021a). Cognitive enrichment device provides evidence for intersexual differences in collaborative actions in Indo-Pacific bottlenose dolphins (Tursiops aduncus). Anim. Cogn. 24, 1215–1225. doi: 10.1007/s10071-021-01510-7
Matrai E., Kwok S. T., Boos M., Pogány Á. (2021b). Group size, partner choice and collaborative actions in male Indo-Pacific bottlenose dolphins (Tursiops aduncus). Anim. Cogn. 25, 179–193. doi: 10.1007/s10071-021-01541-0
Matrai E., Kwok S. T., Boos M., Pogany A. (2022a). Testing use of the first multi-partner cognitive enrichment devices by a group of male bottlenose dolphins. Anim. Cogn. 25, 961–973. doi: 10.1007/s10071-022-01605-9
Matrai E., Ng A. K. W., Chan M. M. H., Gendron S. M., Dudzinski K. M. (2020). Testing use of a potential cognitive enrichment device by an Indo-Pacific bottlenose dolphin (Tursiops aduncus). Zoo Biol. 39, 156–167. doi: 10.1002/zoo.21536
Meyer S., Puppe B., Langbein J. (2010). Cognitive enrichment in zoo and farm animals - implications for animal behaviour and welfare. Berliner und Munchener tierarztliche Wochenschrift 123, 446–456.
Moreno K. R. (2017). Does personality similarity in bottlenose dolphin pairs influence dyadic bond characteristics? Dissertatuin. USA: Department of Psychology, The University of Southern Mississippi.
Morimura N. (2006). “Cognitive enrichment in chimpanzees: an approach of welfare entailing an animal’s entire resources,” in Cognitive development in chimpanzees. Eds. Matsuzawa T., Tomonaga M., Tanaka M. (Springer, Tokyo), pp 368–pp 391.
Nakahara F., Komaba M., Sato R., Ikeda H., Komaba K., Kawakubo A. (2017). Spontaneous prosocial choice by captive bottlenose dolphins, Tursiops truncatus. Behav. Process. 135, 8–11. doi: 10.1016/j.beproc.2016.11.009
New L. F., Moretti D. J., Hooker S. K., Costa D. P., Simmons S. E. (2013). Using energetic models to investigate the survival and reproduction of beaked whales (family Ziphiidae). PloS One 8, e68725. doi: 10.1371/journal.pone.0068725
Pérez-Manrique A., Gomila A. (2019). Bottlenose dolphins do not behave prosocially in an instrumental helping task. Behav. Process. 164, 54–58. doi: 10.1016/j.beproc.2019.04.014
Pitman R. L., Deecke V. B., Gabriele C. M., Srinivasan M., Black N., Denkinger J., et al. (2017). Humpback whales interfering when mammal-eating killer whales attack other species: Mobbing behavior and interspecific altruism? Mar. Mammal Sci. 33, 7–58. doi: 10.1111/mms.12343
Puppe B., Ernst K., Schön P. C., Manteuffel G. (2007). Cognitive enrichment affects behavioural reactivity in domestic pigs. Appl. Anim. Behav. Sci. 105, 75–86. doi: 10.1016/j.applanim.2006.05.016
Ramos E. A., Kiszka J. J., Pouey-Santalou V., Ramirez Barragan R., Garcia Chavez A. J., Audley K. (2020). Food sharing in rough-toothed dolphins off southwestern Mexico. Mar. Mammal Sci. 37, 352–360. doi: 10.1111/mms.12727
Ramos E. A., Santoya L., Verde J., Walker Z., Castelblanco-Martínez N., Kiszka J. J., et al. (2022). Lords of the Rings: Mud ring feeding by bottlenose dolphins in a Caribbean estuary revealed from sea, air, and space. Mar. Mammal Sci. 38, 364–373. doi: 10.1111/mms.12854
R Core Team (2022). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing).
Romeu B., Cantor M., Bezamat C., Simões-Lopes P. C., Daura-Jorge F. G. (2017). Bottlenose dolphins that forage with artisanal fishermen whistle differently. Ethology 123, 906–915. doi: 10.1111/eth.12665
Sakai M., Kita Y. F., Kogi K., Shinohara M., Morisaka T., Shiina T., et al. (2016). A wild Indo-Pacific bottlenose dolphin adopts a socially and genetically distant neonate. Sci. Rep. 6, 23902. doi: 10.1038/srep23902
Sargeant B., Mann J. (2009). Developmental evidence for foraging traditions in wild bottlenose dolphins. Anim. Behav. 78, 715–721. doi: 10.1016/j.anbehav.2009.05.037
Simard P., Gowans S. (2004). Two calves in echelon: an alloparental association in Atlantic white-sided dolphins (Lagenorhynchus acutus)? Aquat. Mammals 30, 330–334. doi: 10.1578/AM.30.2.2004.330
Spinelli L. H. P., de Jesus A. H., do Nascimento L. F., Yamamoto M. E. (2008). Prey-transfer in the marine tucuxi dolphin, Sotalia fluviatilis, on the Brazilian coast. Mar. Biodivers. Records 1, e25. doi: 10.1017/S1755267206002351
Weinpress M. R., Herzing D. L. (2015). Maternal and alloparental discipline in Atlantic spotted dolphins (Stenella frontalis). Anim. Behav. Cogn. 2, 348–364. doi: 10.12966/abc.11.04.2015
Wells R. S. (2014). “Social structure and life history of bottlenose dolphins near Sarasota Bay, Florida:insights from four decades and five generations,” in Primates and cetaceans: field research and conservation of complex mammalian societies. Eds. Yamagiwa J., Karczmarski L. (Springer, Tokyo).
Whitehead H. (1996). Babysitting, dive synchrony, and indications of alloparental care in sperm whales. Behav. Ecol. Sociobiol. 38, 237–244. doi: 10.1007/s002650050238
Whitehead H., Mann J. (2000). “Female reproductive strategies of cetaceans: life histories and calf care,” in Cetacean societies: field studies of dolphins and whales. Eds. Mann J., Connor R. C., Tyack P. L., Whitehead H. (University of Chicago Press, Chicago), pp 219–pp 246.
Winship K. A., Eskelinen H. C. (2018). Behavioral responses of two species of dolphins to novel video footage: An exploration of sex differences. Zoo Biol. 37, 399–407. doi: 10.1002/zoo.21444
Wright B. M., Stredulinsky E. H., Ellis G. M., Ford J. K. (2016). Kin-directed food sharing promotes lifetime natal philopatry of both sexes in a population of fish-eating killer whales, Orcinus orca. Anim. Behav. 115, 81–95. doi: 10.1016/j.anbehav.2016.02.025
Yamamoto C., Kashiwagi N., Otsuka M., Sakai M., Tomonaga M. (2019). Cooperation in bottlenose dolphins: bidirectional coordination in a rope-pulling task. PeerJ 7, e7826. doi: 10.7717/peerj.7826
Yeater D., Kuczaj S. A. II (2010). Observational learning in wild and captive dolphins. Int. J. Comp. Psychol. 23, 379–385. doi: 10.46867/IJCP.2010.23.03.04
Keywords: Indo-Pacific bottlenose dolphin, cooperation, altruism, environmental enrichment, sex differences, dolphin behavior
Citation: Matrai E, Kwok ST, Tan Z, Lin X, Lee W-K, Martelli P and Pogány Á (2024) Cognitive enrichment for testing prosocial food-sharing in dolphin groups. Front. Ecol. Evol. 12:1479989. doi: 10.3389/fevo.2024.1479989
Received: 13 August 2024; Accepted: 28 October 2024;
Published: 14 November 2024.
Edited by:
Anindita Bhadra, Indian Institute of Science Education and Research Kolkata, IndiaReviewed by:
Briseida Dogo Resende, University of São Paulo, BrazilAnna Zanoli, University of Turin, Italy
Copyright © 2024 Matrai, Kwok, Tan, Lin, Lee, Martelli and Pogány. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eszter Matrai, ZXN6dGVyLm1hdHJhaUBvY2VhbnBhcmsuY29tLmhr