
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 13 January 2025
Sec. Biogeography and Macroecology
Volume 12 - 2024 | https://doi.org/10.3389/fevo.2024.1477480
 Tosif Fida1
Tosif Fida1 Alireza Mohammadi2
Alireza Mohammadi2 Kamran Almasieh3
Kamran Almasieh3 Luciano Bosso4*
Luciano Bosso4* Shams Ud Din1Urwah Shamas5Muhammad Ali Nawaz6
Shams Ud Din1Urwah Shamas5Muhammad Ali Nawaz6 Muhammad Kabir1*
Muhammad Kabir1*The main cause of the global threat to large carnivore populations living in high-altitude areas is human pressure. The Himalayan brown bear (Ursus arctos isabellinus, hereafter bear) is listed as a critically endangered species, and it dominates the higher altitudes of the North and Western Himalayan landscapes. Here, we used species distribution modelling and landscape connectivity to find bears’ suitable habitats and corridors in the Deosai National Park (DNP) and surrounding areas. Species distribution models detected a bear’s suitable habitat of 1,125 km2, of which ca. 73% was covered by DNP. The most crucial predictors in determining the bears’ distribution were elevation—particularly if it was between 3,500 and 4,500 meters above sea level—as well as the bears’ distance from rivers, cattle, and the DNP borders. We discovered robust connectivity among presence points in the core of the study area, particularly within the DNP boundaries, by using the electrical circuit theory models. Landscape connectivity values were indeed low in some areas outside and close to the DNP confines. Based on our findings, it is imperative to protect habitats and corridors that allow the bears to migrate between the suitable patches, particularly beyond the park’s boundaries, to improve bear conservation.
Graphical Abstract.
In the last two decades, anthropogenic pressures have affected the survival of several large carnivore species living in in high-altitude ecosystems (Sarkar et al., 2018; Mohammadi et al., 2021a; Almasieh et al., 2022; Dar et al., 2023; Singh et al., 2024). Unfortunately, the current Protected Areas (PAs) network fails to support viable populations of large carnivores in remote, high-altitude landscapes, because these animals have particular habitat requirements, great dispersal ability, and low population density (Rezaei et al., 2022a, b). The establishment of protected areas to guarantee the conservation of critical habitat patches and corridors is necessary for the long-term survival of carnivore species (Kaszta et al., 2019, 2020), especially in high-altitude ecosystems. Thus, every conservation measure for large carnivores needs a careful assessment of habitat suitability, landscape connectivity, and the PAs network to protect threatened species (Mohammadi et al., 2021b; 2022; Rezaei et al., 2022a; b; Ashrafzadeh et al., 2023).
Due to anthropogenic pressures, the brown bear (Ursus arctos) populations and distribution have declined by more than 50% since the mid-1800s (Servheen, 1990). The Himalayan brown bear (Ursus arctos isabellinus, hereafter bear), a sub-species of brown bear, represents an ancient lineage that stayed genetically isolated for a long time in the Central Asia mountain range (Galbreath et al., 2007), where it still lives dominating the higher altitudes of the North and Western Himalayan landscapes (e.g., Nawaz, 2007; Hosseini et al., 2022). A bear population of ca. 150–200 individuals can be found in the northern region of Pakistan in an area of ca. 150,000 km2 (Nawaz, 2007; Haq and Rahatullah, 2012). This bear population is mainly restricted to subalpine and alpine zones in Khunjerab, Deosai National Park (DNP), and neighbouring areas, such as in other protected areas placed in Astore and Nanga Parbat (Virk et al., 2003). Numerous factors are posing a threat to the bear population in Pakistan, where it is listed as critically endangered (Sheikh and Molur, 2005), including the expanding human population, farming and overgrazing, the extraction of ethno-botanical plants, the unsustainable use of natural resources, poaching, shoddy tourism, and the illegal trade in bear fat and pelts (Sheikh and Molur, 2005; Nawaz, 2007; Dixo et al., 2009; Ali et al., 2016; Mohammadi et al., 2018; Ali A. et al., 2021; Mohammadi and Fatemizadeh, 2021; Fida et al., 2024). Currently, the most important and stable bear population lives on the Deosai plateau, in the western Himalayan areas (Nawaz, 2007; Fida et al., 2024). Although bear habitat is generally falling, there are significant protection measures in place for the populations in DNP, particularly against anthropogenic pressure (Nawaz, 2008), which is contributing to the bear population’s growth (Abbas et al., 2022). Even though conservation efforts have contributed to an increase in bear populations in DNP areas since 1993, the previous population decline and habitat fragmentation have considerably reduced genetic variability in the existing bear population (Bellemain et al., 2007). Effective conservation measures on the Deosai plateau are therefore crucial for the brown bear’s existence in the future, not only in Pakistan, but also throughout its wide distribution (Nawaz, 2007).
To carry out an efficient conservation program, it is a priority to know the movement patterns of the bears (Ciucci et al., 2014; Kavčič et al., 2015) as well as the suitable habitats in Asia (Nawaz et al., 2014; Su et al., 2018; Ansari and Ghoddousi, 2018; Rehan et al., 2024; Ahmad et al., 2024). The assessment of the status of habitat suitability and landscape connectivity are essential tools for developing successful planning strategies (e.g., Velázquez et al., 2017; Kabir et al., 2017; Velázquez et al., 2022; Tang et al., 2023). These assessments consider the species’ geographic location and their critical function in promoting migration within the available habitat. Our study used a mix of Species Distribution Models (SDMs) and corridor network simulators to determine the habitat suitability and landscape connectivity for bears in DNP, respectively. Furthermore, we also explored the role and importance of the single variable in identifying the most important areas for bear distribution in DNP and neighbouring areas. We were not confined to evaluating simply the DNP region since we understand that the long-term viability of the Deosai and surrounding bear populations requires landscape-wide management techniques. Because some individuals appear to have home ranges larger than the park, the national park may not be sufficient to ensure long-term population survival.
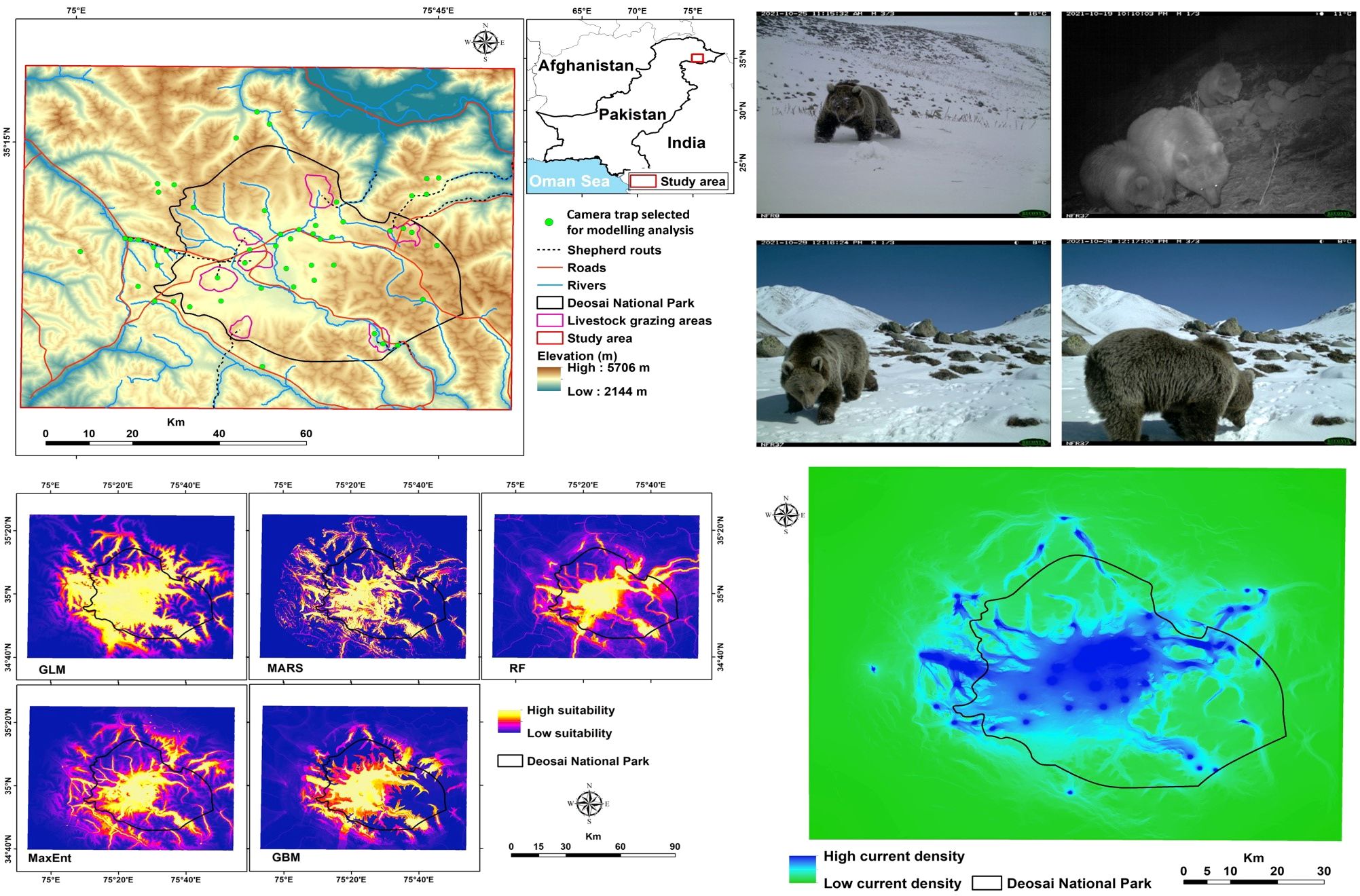
The DNP (75°27′E, 35°00′N) is a 3,584 km2 alpine plateau east of Nanga Parbat Peak, Northern Areas, Pakistan. Elevations range from 3,500 to 5,200 m above sea level, with about 60% of the area between 4,000 and 4,500 m above sea level (Nawaz et al., 2008; Figure 1).
Figure 1. Study area showing the Desoai National Park (black line) and Himalayan brown bear occurrences obtained from the camera traps (green circle).
Wetland regions and several tiny lakes are found on the DNP plateau. Their genesis is owing to snow and ice melting from the surrounding high mountains, as well as the existence of a permafrost table, which forms a thermokarst landscape (Karlsson et al., 2012). Deosai’s and surrounding highland habitat is known for its unpredictable, unstable, highly seasonal, and harsh environmental conditions. The typical daily temperatures range from -20°C to 12°C, and yearly precipitation ranges from 510 to 750 mm, largely as snow (Nawaz et al., 2008).
The vegetation in DNP is primarily herbaceous perennials, with a high number of cushion-forming and tufted species. Grasslands and areas with persistent snow or ice cover make up the majority of the land cover in high-elevation regions. Instead, the valley are mainly constituted by bogs and ponds with associated vegetation such as sedges, grasses, and plants like Aconitum violaceum, Swertia perfoliata, and Saxifraga hircus (Bellemain et al., 2007).
The latest results on bear population in DNP estimated ca. 58 – 77 bear specimens (Abbas et al., 2022). In addition to the bear, other mammalian species live in the DNP, such as the Himalayan red fox (Vulpes vulpes montana), the Mongolian wolf (Canis lupus chanco), the long-tailed marmot (Marmota caudata), the Himalayan ibex (Capra ibex sibrica), and 17 other smaller species (Nawaz, 2008; Ali H. et al., 2021). Many permanent human settlements exist inside and along the boundaries of the DNP, such as Sadpara, Shilla, Dappa, and Karabos. In addition to these permanent communities, it is common to find transhumance activity coming from the valleys (Nawaz, 2007).
Aside from the issues of overgrazing and soil degradation that affect DNP and its surrounding areas, the most pressing concern is the regions with water bodies and wetlands that are nourished by glacier/snow and permafrost melting throughout the summer. These regions, as well as their surrounds, are critical for flora and animals to live and thrive.
We collected bear occurrences by using camera traps in DNP and neighbouring areas during the field surveys from June 2021 to May 2022. This method was widely used to capture images of several bear species in their habitat (e.g., Ahmad et al., 2024; Lacy et al., 2024; Thakur et al., 2023; Fida et al., 2024). We used a motion-sensing digital camera equipped with infrared flash (HC500/PC900 model, Reconyx brand, Holmen, Wisconsin, USA) to capture bear pictures placed in 89 camera stations (e.g., Ahmad et al., 2024) with a capture rate value of 65.5%. Each camera was placed on a ground-mounted steel pole (about 50 cm tall), with a minimum distance of 1 km (mean = 1.824 km; maximum = 5.84 km) between camera stations (e.g., Gubbi et al., 2023; Wolfson et al., 2023; Melo-Dias et al., 2024). We used this distance value following a similar study carried out in the south of Iran with similar ecosystem conditions (Almasieh et al., 2016). The camera traps were positioned such that vegetation in the front could not activate them; they were designed to take pictures continuously at 1-second intervals when triggered (Bischof et al., 2014). The camera traps were normally left operational at each point for 10–40 days (mean = 25 days per station, Ahmad et al., 2022). To reduce the effect of spatial autocorrelation, we thinned all the species occurrences to ensure one presence record per environmental raster cell (e.g., Freeman et al., 2022). Finally we obtained 52 bear presence records (Figure 1) and used them in the bear’s spatial analyses. Although previous publications have reported bear presence records for this area, we prefer to use only our records for two main reasons: 1) they are based on a more recent and reliable camera trap survey (e.g., Fida et al., 2024); and 2) they cover the whole park without any spatial autocorrelation.
We selected the environmental predictors based on the bear ecology obtained from other studies carried out in similar areas (Nawaz et al., 2014; Almasieh et al., 2016, 2019; Mohammadi et al., 2021a). The elevation map was derived from a 30-meter Digital Elevation Model (DEM) created by the Shuttle Radar Topography Mission (SRTM, http://earthexplorer.usgs.gov). We generated the slope maps starting from the DEM map by using the Surface tool in ArcGIS v. 10.3 (http://www.esri.com/software/arcgis). We considered 23-class global land cover maps (GlobCover version 2.3; ESA, 2009) for our analysis, but, after an initial screening, we selected only those most preferred by the bears, such as: 1) closed to open grasslands (>15%); 2) bare areas (23.5%); and 3) permanent snow or ice (22.6%). We used a moving window with a radius of 5 km was in the Focal Statistics tool to create density maps for the cover types selected (Mohammadi et al., 2021a; Almasieh et al., 2023a). All the following distances were calculated in the Euclidean space between a point in the map and the corresponding predictor. Distance from DNP borders was added in consideration of the current level of bear protection. Distance from rivers was used for water resources (Ansari and Ghoddousi, 2018), and distance from roads and human settlements was used for human disturbances. Distances to areas used by livestock for grazing and livestock movement routes represented both human and livestock presence. The coordinate system utilized was WGS 1984 UTM zone 43, and all variables were used at a 30-m resolution (Supplementary Materials 2 Figures S1–S9).
We used the Variance Inflation Factor (VIF) to evaluate environmental predictor multicollinearity. VIF > 3 was set as the threshold (Zuur et al., 2010), and variables exceeding the threshold were excluded using the USDM package (Naimi et al., 2014) in R. 6.0 (R Core Team, 2019). All the predictors used in our modelling analysis were included in the Table 1.
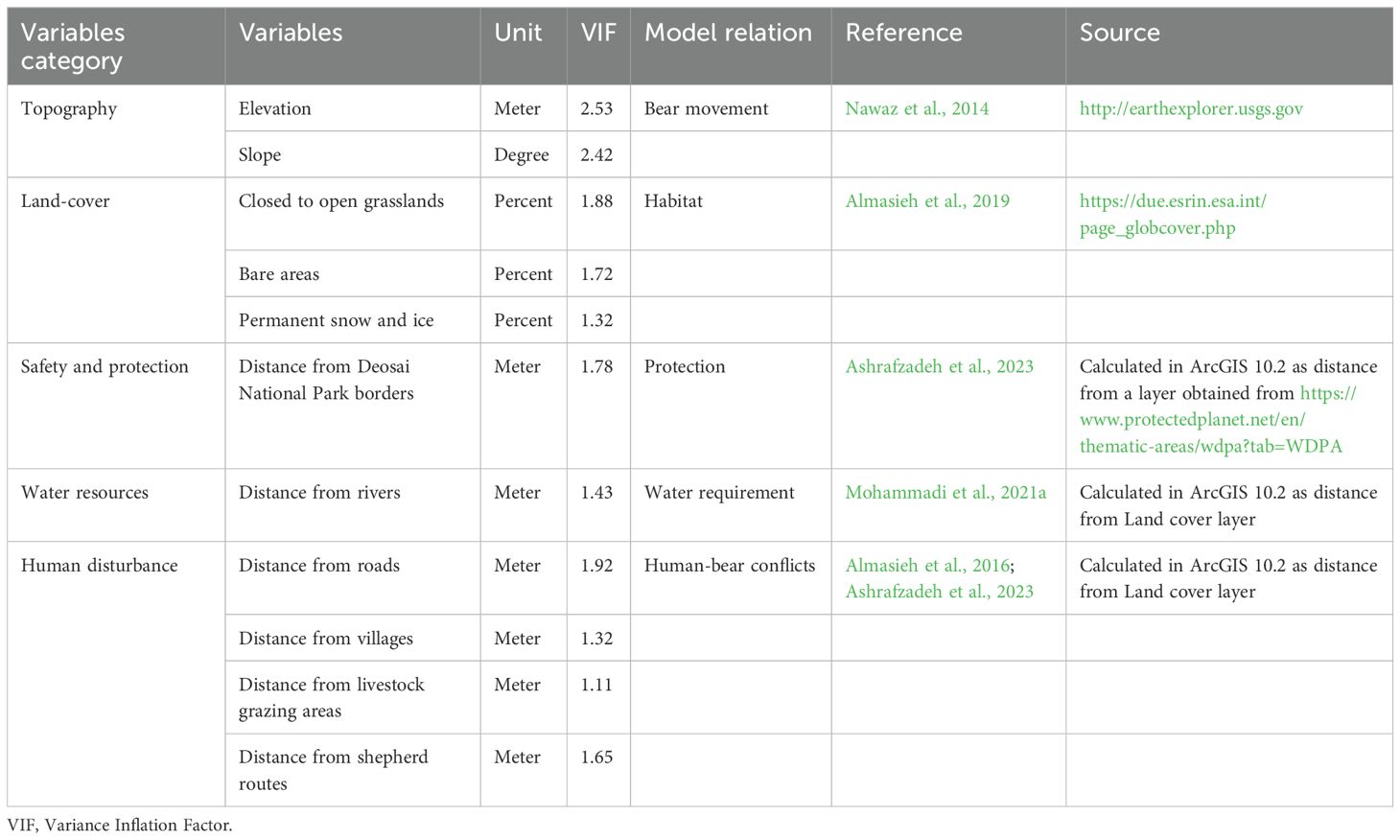
Table 1. Environmental variables used for species distribution modelling of the Himalayan brown bear in the Deosai National Park and neighbouring areas.
We explored the bear’s habitat suitability by using SDMs in an ensemble forecasting approach, as implemented in the R (v. 4.0.2) package “biomod2: Ensemble Platform for Species Distribution Modeling” (Thuiller et al., 2019; the R code is available in the Supplementary Material 1). We combined predictions from different model types and fitted multiple suitability models because it has been shown that this methodology improves model accuracy (Araújo and New, 2007; Almasieh et al., 2022; Mohammadi et al., 2022; Di Febbraro et al., 2023). We combined five algorithms into two main categories, such as: 1) the regression-based models—Generalized Linear Model (GLM) and Multivariate Adaptive Regression Splines (MARS); 2) the machine learning models—Maximum Entropy Model (MaxEnt), Random Forest (RF), and Generalized Boosting Model (GBM).
A 3:1 training-testing division was used to allocate occurrence points. A total of 500 pseudo-absence points were randomly generated outside a 1 km radius around occurrences. We chose this value of pseudo-absence beyond a radius of 1 km from occurrence since the ecosystem around the DNP was identical to that of the park itself (Barbet-Massin et al., 2012). Each model was run with 20 replications to increase reliability (Barbet-Massin et al., 2012; Kaboodvandpour et al., 2021). For each variable, we calculated the mean contribution to the model with the best performance by using biomod2, which also shows the response curves. We converted the continuous habitat suitability map generated by the ensemble model into a binary map through the 10th percentile threshold (Ahmadi et al., 2020; Almasieh et al., 2023b). This threshold (the value above which the model classifies correctly 90% of the training locations) is a conservative value that excludes outliers from the training presence points and is commonly used in species distribution modelling studies, especially when we have areas with similar environments (Ten Caten et al., 2023). Patches overlapping with the main presence records were assumed to be the core of the bear’s habitat suitability, and the total coverage by DNP was calculated.
The initial round of SDMs utilised all 10 models; then, we selected only models with an AUC > 0.9 and a True Statistic Skill (TSS) > 0.75 (Eskildsen et al., 2013; Almasieh and Cheraghi, 2022). As a threshold-independent measure of predictive accuracy, AUC provides an overall summary of the model’s diagnostic accuracy (Fielding and Bell, 1997). Following Swets (1988), the evaluation criteria for the AUC were classified as fail (0.5–0.6), poor (0.6–0.7), fair (0.7–0.8), good (0.8–0.9), and excellent (0.9–1). The TSS is a threshold-dependent measure that combines sensitivity (true positive rate) and specificity (true negative rate) to assess model performance. It is expressed as Sensitivity + Specificity – 1 (Allouche et al., 2006). The evaluation criteria for TSS were classified as poor (<0.4), good (0.4–0.75), and excellent (>0.75) (Landis and Koch, 1977).
Enhancing landscape connectivity is one of the most recommended anthropogenic pressure-adaptation strategies for protecting biodiversity as the global changes (e.g., Velázquez et al., 2017; Kabir et al., 2017; Velázquez et al., 2022; Tang et al., 2023). In order to generate a landscape connectivity map, we used the ensemble modelling output to produce a resistance layer following the methods proposed by Wan et al. (2019). This approach is based on the notion that animals select dispersal movement paths based on the same features that they use to select habitat. If this assumption is correct, habitat suitability would be the inverse of landscape resistance (i.e., areas with high habitat suitability would have low resistance to dispersal movement). The ensemble layer was rescaled in 0-1 mode by using the Rescale by Function tool in ArcGIS. A negative exponential function calculates resistance as follows: R=1000(-1×Ensemble Suitability), where R is the pixel resistance value (Mateo-Sánchez et al., 2015). Resistance values were linearly rescaled from 1 to 10, where 1 and 10 were minimum and maximum resistance, respectively (Wan et al., 2019).
We well know that there are numerous methods for evaluating landscape connectivity, including least-cost path analysis, resistant kernel models, and circuit-theory-models (Phillips et al., 2021). Of these methods, circuit-theory based models have shown consistently high performance in estimating connectivity, and there has been a notable uptick in their use in connectivity modelling (Phillips et al., 2021). Additionally, omnidirectional methods like Circuitscape demonstrated accurate estimations of connection corridors, particularly in situations where the animal uses random exploration of the underground —as is the case with bears during dispersion migrations, for instance (Zeller et al., 2020).
Electrical circuit theory models (McRae et al., 2008) simulate habitat connectivity based on the random walk approach and electrical circuits, with the current that imitates the organisms moving between focal nodes (occurrences or patches) based on voltage (movement probability) and resistance (permeability) (McRae et al., 2008; Roever et al., 2013). This approach identifies multiple potential movements, unlike least-cost methods, which identify only one route (Urban and Keitt, 2001; Urban et al., 2009), making it useful for modelling gene flow (McRae and Beier, 2007).
We carried out electrical circuit modelling using Circuitscape v4 (McRae and Shah, 2009) through an all-to-one approach that maps current density occurrences, consuming less RAM (McRae et al., 2008). The resistance map and bear occurrence records were used as inputs. We determined connections to eight neighbours per cell. Lower resistance indicates a higher movement density.
We found that five models (GLM, MaxEnt, RF, MARS, and GBM) showed high model validation values with AUC > 0.9 and TSS > 0.75, respectively. RF obtained the best performance (Table 2). We detected that elevation and distance from DNP, livestock, and rivers were the most important variables for predicting the potential distribution of bears (Table 3).
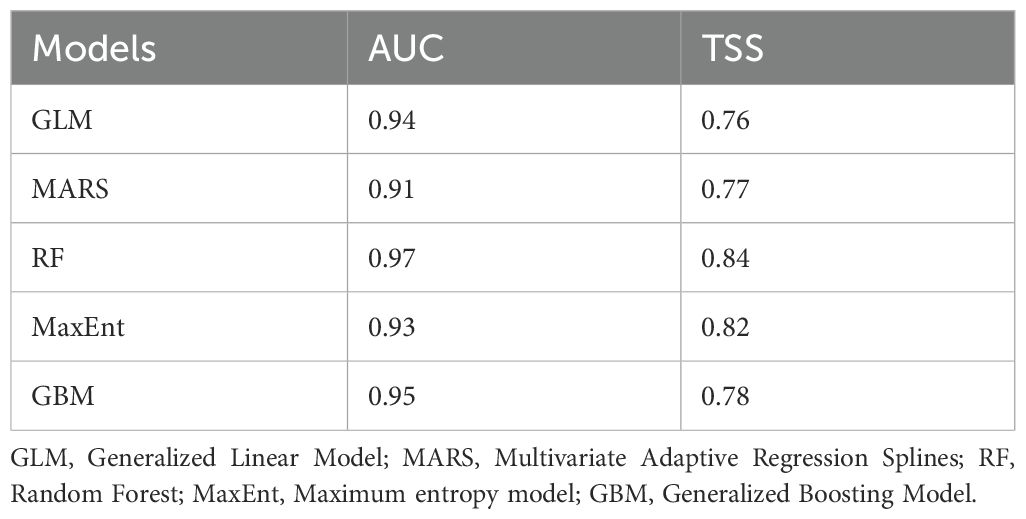
Table 2. Values of Area Under Curve (AUC) and True Skill Statistics (TSS) of the five algorithms used in the ensemble model approach for the Himalayan brown bear in the Deosai National Park and neighbouring areas.
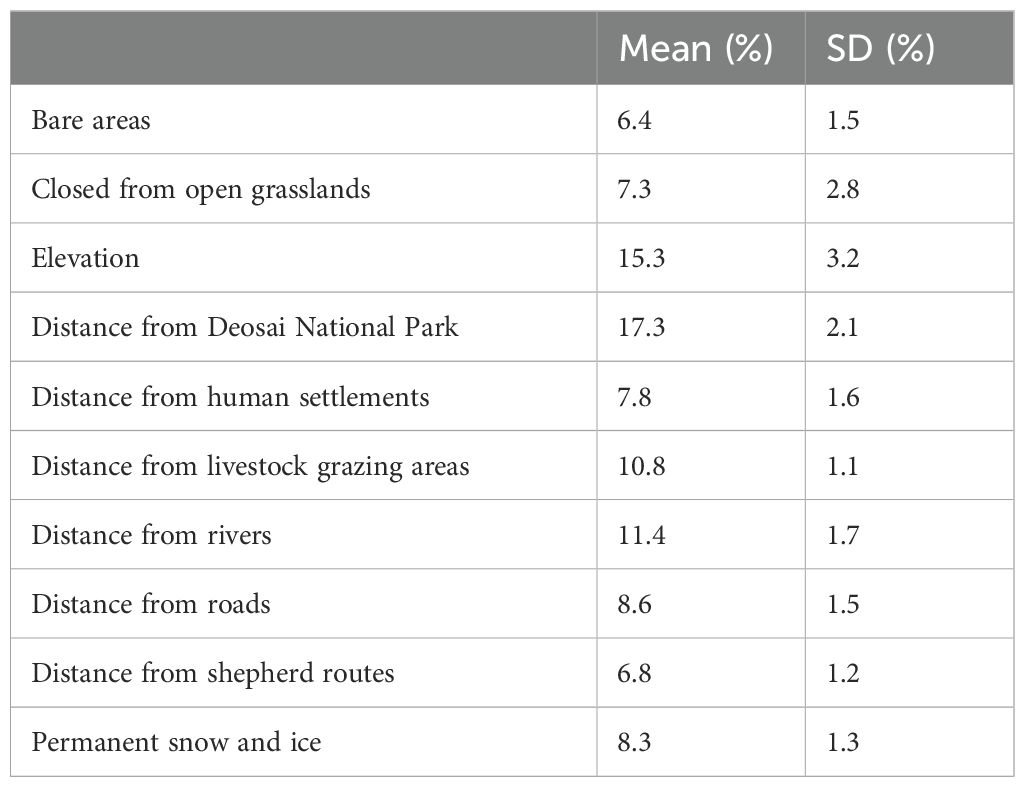
Table 3. Percentage of variable contributions (mean and standard deviation SD) included in the species distribution modelling for the Himalayan brown bear in the Deosai National Park and neighbouring areas.
Low habitat suitability values increased with distance from DNP borders (up to ~10 km). The bear’s probability of presence was consistent about 2 km from rivers. Up to around 20 kilometers from roadways, bear habitat suitability appeared to improve. Distance from livestock regions decreased gradually, unlike distance from livestock transportation routes, which decreased quickly before flattening at around 20 km (Figure 2). At least, high probabilities of bear presences were found between 3,750 and 4,250 meters above sea level.
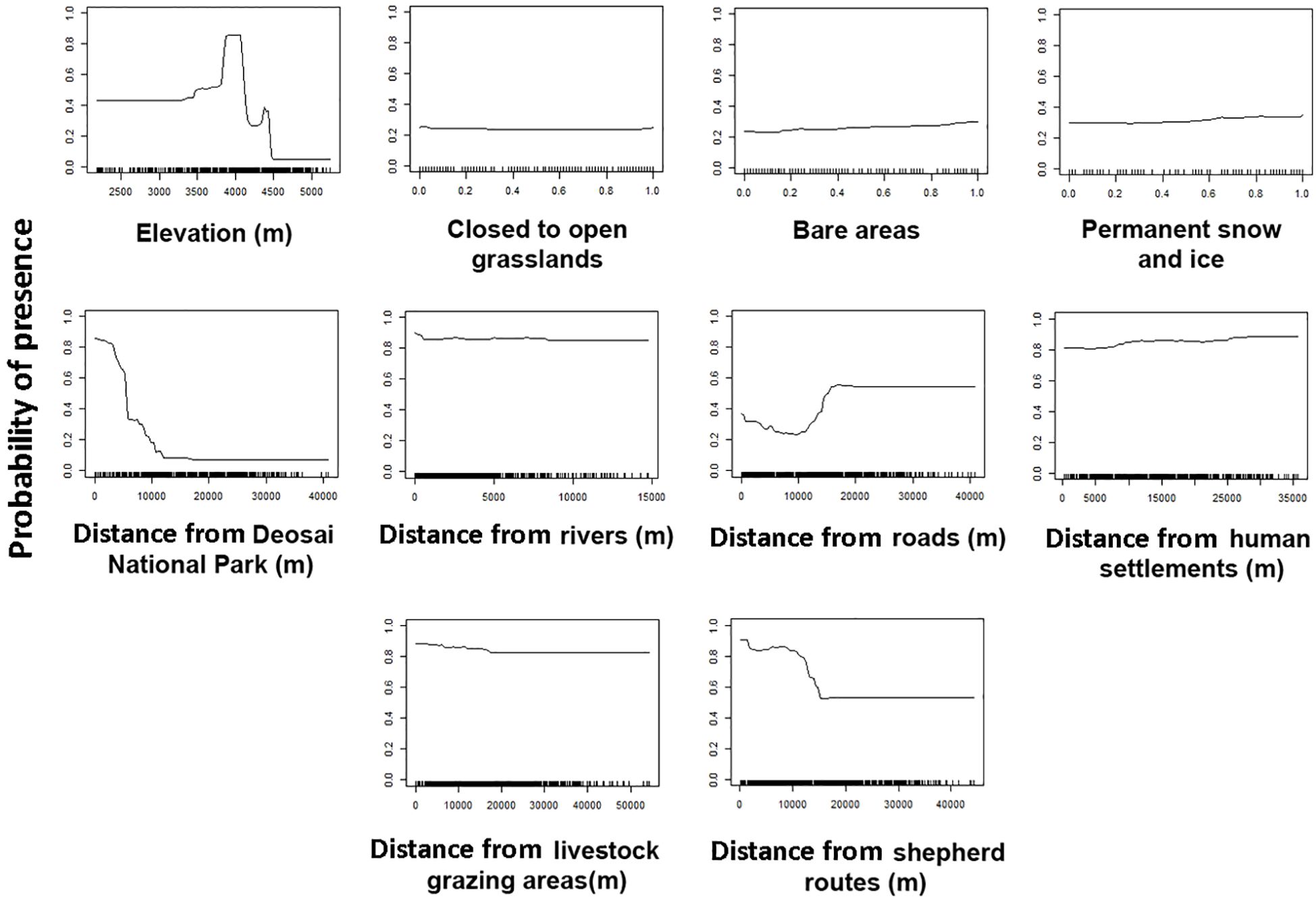
Figure 2. Response curves (from 0 to 1) of the environmental predictors used in species distribution modelling for the Himalayan brown bear in the Desoai National Park.
According to the ensemble suitability maps, the bear’s habitat suitability was highest in the central region of the research area (inside DNP), while there were also other suitable patches in nearby areas (Figure 3). These locations are crucial for encouraging bear migration over their whole range (Figure 3).
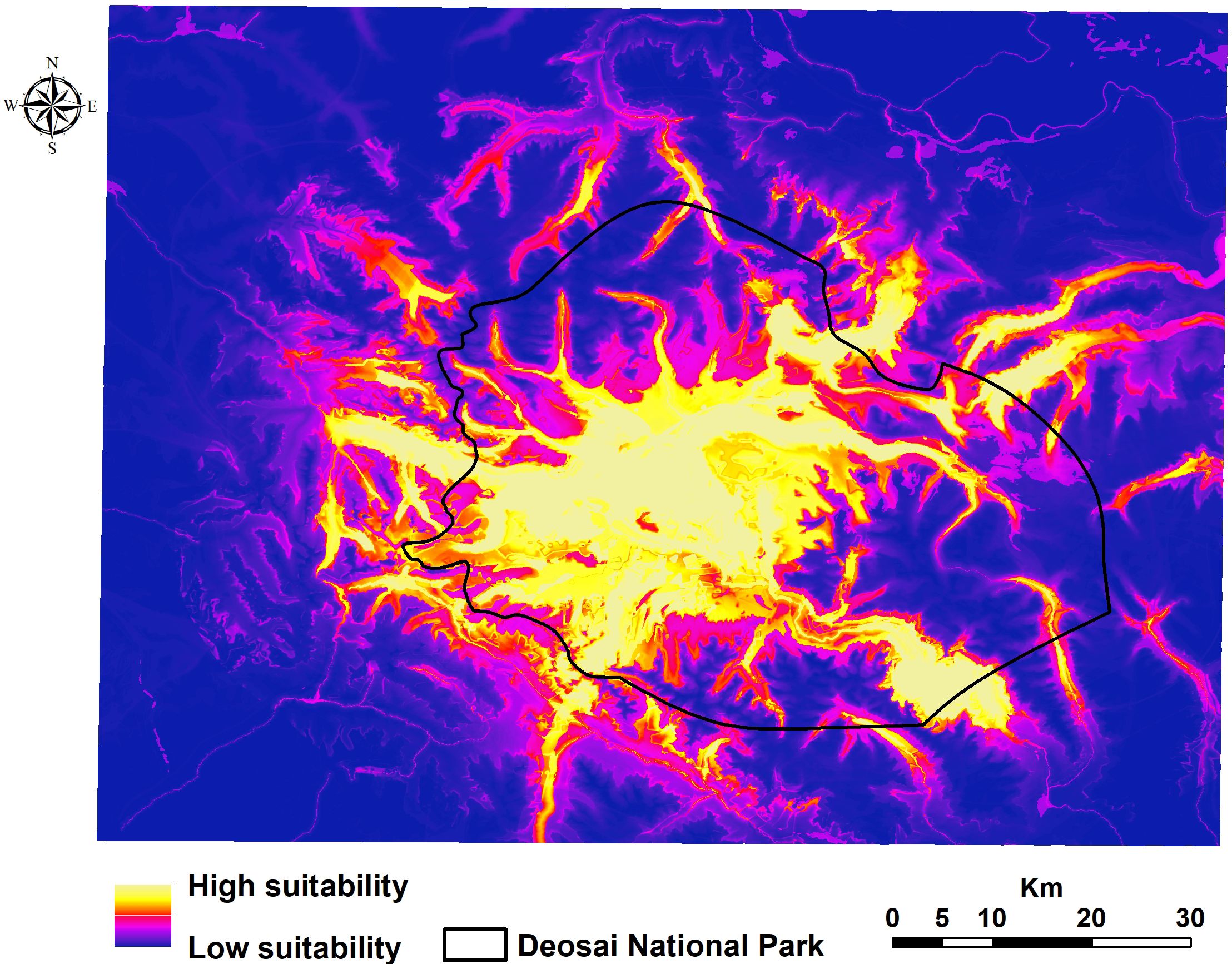
Figure 3. Potential distribution of the Himalayan brown bear in the Deosai National Park and neighbouring areas.
We found that the habitat patches covered 1,125 km2 of the entire study area, and ca. 73% of them were located inside the DNP. The core of the bear’s potential distribution fell mainly within DNP, with a total area of about 1,020 km2 (13% of the study area; 80% coverage by DNP, Figure 4). We detected some other little patches in areas close to the border of DNP (Figure 4). Our results showed that ca. 82% of livestock grazing areas occurred within habitat patches, and livestock grazing areas created about 11% of habitat patches for the bear. Total road and river densities within habitat patches were about 151 and 341 m/km2, respectively. There was robust connectivity among occurrence points of the bear in the central part of the study area within DNP (Figure 5), mainly due to the high number of bear records and low resistance values, i.e., a high value of probability of presence. Connectivity to several locations outside DNP close to the national park boundaries was discontinued (Figure 5).
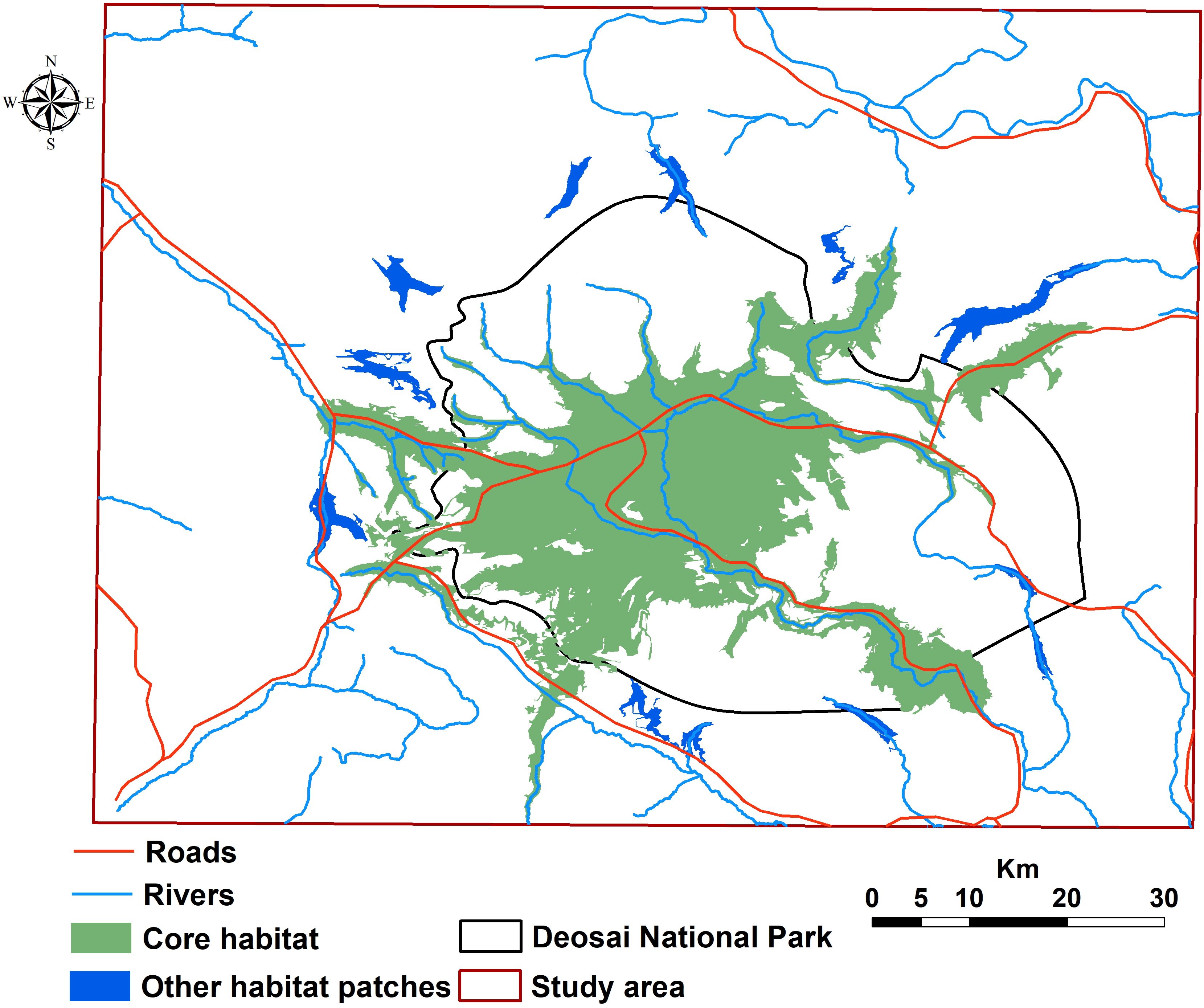
Figure 4. Habitat patches for the Himalayan brown bear in the Deosai National Park and neighbouring areas.
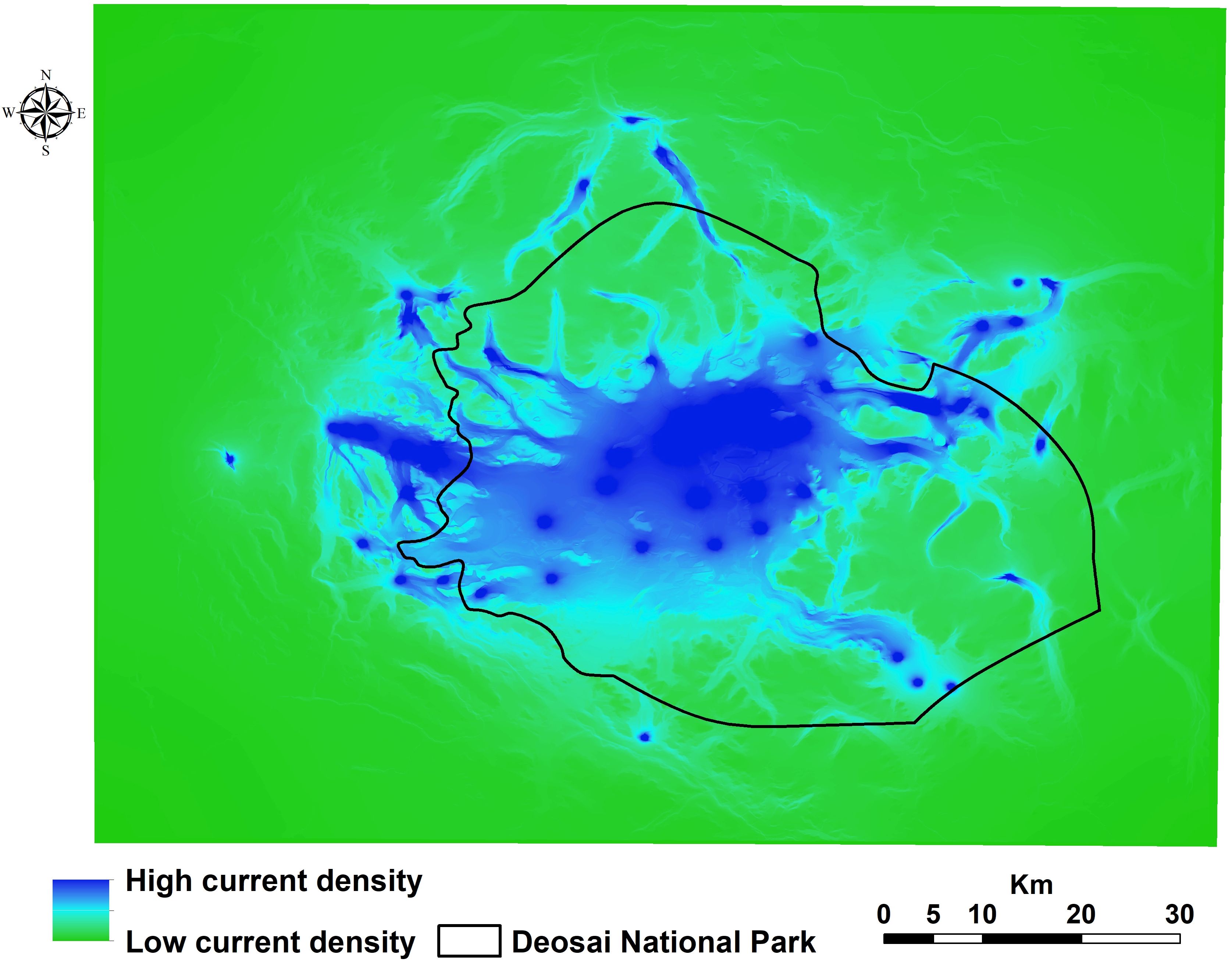
Figure 5. Landscape connectivity map for the Himalayan brown bear in the Deosai National Park and neighbouring areas.
We explored the current distribution and landscape connectivity of the bear in the DNP and surrounding areas to shed light on the factors that are needed for its presence and conservation. Our spatial analysis findings confirmed that the Himalayan and Karakorum-Pamir highlands are the center of the ecosystem where the most important bear populations in Pakistan live. Our results demonstrate the efficacy of DNP and conservation measures that are used inside the protected area’s borders but that may be extended into neighbouring areas.
Along the altitudinal gradients, we found that the bears preferred an elevation between 3,500 and 4,500 m a.s.l. (e.g., Dar et al., 2021). Here, the possibility of sighting the bear increases due to the presence of meadows, bare patches, and prolonged snow and ice. Instead, in other areas, the bear preferred to avoid high-elevation areas because food availability is lower than in forested habitats at low elevations (e.g., valley), increasing the human-bear interactions (Martin et al., 2012; Almasieh et al., 2019; Mohammadi et al., 2021a; Kumar et al., 2022). In the valleys, uncontrolled overgrazing and activities such as beekeeping in the highlands can increase the risk of conflict with local communities (Sharief et al., 2020; Kumar et al., 2022).
Our results confirmed that one of Pakistan’s best bear-protected areas is DNP, where bear populations are growing due to effective conservation initiatives, as reported by Abbas et al. (2022). A few patches of bear habitats located outside the boundary of the park support the survival of the bear population, promoting the dispersion of this species also in other areas of its range (Nellemann et al., 2007; Martin et al., 2010; Fernández et al., 2012; Ziółkowska et al., 2016; Penteriani et al., 2020). Our results showed that bears select riparian and riverine environments because the water resource was among the most influential variables in predicting the bear’s potential distribution, as confirmed by Ansari and Ghoddousi (2018).
Distances to human settlements and roads have an important influence on the habitat suitability of bears in the DNP. Bears stay away from human communities until they are forced to look for food. During the winter, they may come upon abandoned grain and agricultural storage facilities. Furthermore, our results confirmed that the human impact, as for example the presence of roads, contributes negatively to the prediction of the bear’s habitat (Mateo-Sánchez et al., 2014; Farashi et al., 2016; Coogan et al., 2018; Almasieh et al., 2019). If on one hand the roads are increasing human-bear interactions, on the other hand, the road favours connections with other bear populations that are scattered or unevenly distributed (Kaczensky et al., 2003; Bischof et al., 2017). Finally, it is important to remember that the roads represent one of the most important causes of death for bears (Herrero et al., 2005; Mohammadi et al., 2021a). The roads used by shepherds can be a facility for the bear’s seasonal migration and foraging for food (pers. obs. by authors). A large amount of a bear’s diet consists of vegetables (Nawaz, 2008; Nawaz et al., 2019). The field-based observation showed that agricultural cultivation and pastoral farming are the primary land use practices also in the zones close to the DNP borders. Large herds of domestic cattle grazing freely inside the bears’ primary habitat create competition for resources. The Himalayan area is strongly susceptible to the effects of human impacts, mainly on land use, negatively influencing the bear presence and distribution (Dar et al., 2021). Throughout their historical range, human land use practices have played a significant role in the bear decline (Zedrosser et al., 2011; Wolf and Ripple, 2017), negatively impacting bear habitat (Dixo et al., 2009), survival (Cushman et al., 2018; Kaszta et al., 2019), dispersal pattern (Cushman et al., 2014; Mateo-Sánchez et al., 2014), genetic diversity (Kaszta et al., 2020), and ecological connections in habitat patches (Scolozzi and Geneletti, 2012; Gao et al., 2013).
The bears migrate following the change in seasons, spending the summers on plateaus at higher elevations and the winters in the valleys, where they hibernate. Cattle grazing and fuelwood collection are two main human-caused disturbances of bear habitat that commonly occur in the valleys. Our findings revealed that DNP covered 73% of the bears’ habitat regions. However, other tiny, well-located, and suitable places were discovered along the DNP’s borders; these areas should be conserved since they exhibit a high degree of environmental similarities to those found in the park. On the one hand, we discovered a robust network between the bear-rich areas of the DNP, but on the other hand, because of anthropogenic activity, the areas outside the DNP were not strongly connected to one another (e.g., Fida et al., 2024). This situation means that bears in DNP may survive in isolated populations with minimal genetic diversity (Nawaz, 2008). Several studies have shown that maintaining core habitats and promoting migration across appropriate habitat patches are necessary to ensure the appropriate survival of vertebrate populations (Margules and Pressey, 2000; Mech and Hallett, 2001). In order to resolve human-bear conflicts, spatial analyses, such as landscape connectivity, are crucial to improving effective management plans to protect bear population and distribution (Lecis and Norris, 2003; Goursi et al., 2021; Mohammadi et al., 2021b; Rehan et al., 2024).
With a predicted loss of 56,501 km² (17.52%) by 2050, mostly due to human activity, Pakistan’s bear habitat is predicted to be seriously threatened (Su et al., 2018). A research-based conservation plan is required to monitor the bear population and the effects of the factors affecting its survival. One such approach would be to preserve the landscape corridors (Bellemain et al., 2007). Although there is a healthy bear population in the park, there may be serious risks related to ongoing construction projects, infrastructure, and human access to bears’ primary habitat. In order to support the unchecked growth of tourism in PAs, it should be banned to construct infrastructure in the DNP and its neighbouring areas that could disrupt bears’ natural habitat. The road that passes through DNP separates the bear’s habitat, and when summer traffic increases, the road becomes busier, causing habitat fragmentation and population isolation. We should establish safe movement corridors, underpasses, and tunnels to allow bears and other wildlife to freely roam across the appropriate ecosystems found along the major road. Our findings imply that the DNP limits should be expanded to include areas where appropriate habitat and pathways were discovered. The Pakistani government recently declared two locations within the bear range to be protected areas: Nanga Parbat National Park and Himalaya National Park. Preservation and restoration of species movement corridors should be the primary focus of conservation efforts to preserve genetic diversity and gene flow among bear populations (Straka et al., 2012).
To detect the bear distribution and landscape connectivity in Pakistan’s DNP and neighbouring areas, we used a combination of species distribution models and connectivity simulation. Our research shows that the core bear population and distribution are well protected in DNP; however, about 27% of the pathways used by bears were found outside of protected area boundaries. We discovered a gap in the protection of corridors by crossing these ecological elements with already-existing protected areas. To overcome this conservation gap, we propose establishing new protected areas strategically positioned to successfully defend the bear distribution and ecological corridors. The planning and design of PAs, as well as the evaluation of development plans, must incorporate this spatially explicit prioritisation in order to effectively balance the conflicting demands of development and sustainable conservation initiatives, as well as to guarantee the long-term survival of threatened species.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
TF: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AM: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. KA: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. LB: Supervision, Writing – original draft, Writing – review & editing, Methodology. SUD: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. US: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MN: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MK: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. LB was funded by the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4 - Call for tender No. 3138 of 16 December 2021, rectified by Decree n.3175 of 18 December 2021 of Italian Ministry of University and Research funded by the European Union – NextGenerationEU; Project code CN_00000033, Concession Decree No. 1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, CUP, H43C22000530001, Project title “National Biodiversity Future Center - NBFC”.
We would like to express our gratitude to the Editor Oana Moldovan and the two reviewers for the really positive feedback and helpful comments that supported this final version of the manuscript. We are thankful to the Forest, Wildlife & Environment Department Government of Gilgit-Baltistan for their assistance during the fieldwork activities and also appreciate Abuzar Ahmad (Game watcher, DNP) help during the camera trapping process. We also sincerely acknowledge the logistical support provided by the Snow Leopard Foundation in Pakistan.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2024.1477480/full#supplementary-material
Abbas S. N., Haider J., Younas M., Ali H. (2022). Status and Distribution of Himalayan Brown Bear in Deosai National Park and its Buffer Valleys, Gilgit-Baltistan (Gilgit-Baltistan, Pakistan: Forest, Wildlife and Parks Department, Government of Gilgit-Baltistan), 20 pp.
Ahmad F., Mori T., Rehan M., Bosso L., Kabir M. (2024). Applying a random encounter model to estimate the asiatic black bear (Ursus thibetanus) density from camera traps in the hindu raj mountains, Pakistan. Biology 13, 341. doi: 10.3390/biology13050341
Ahmad F., Nawaz M. A., Salim M., Rehan M., Farhadinia M., Bosso L., et al. (2022). Patterns of spatial distribution, diel activity and human-bear conflict of Ursus thibetanus in the Hindu Kush mountains, Pakistan. Global Ecol. Cons. 37, e02145. doi: 10.1016/j.gecco.2022.e02145
Ahmadi M., Farhadinia M. S., Cushman S. A., Hemami M.-R., Nezami Balouchi B., Jowkar H., et al. (2020). Species and space: a combined gap analysis to guide management planning of conservation areas. Landsc. Ecol. 35, 1505–1517. doi: 10.1007/s10980-020-01033-5
Ali H., Din J. U., Bosso L., Hameed S., Kabir M., Younas M., et al. (2021). Expanding or shrinking? range shifts in wild ungulates under climate change in Pamir-Karakoram mountains, Pakistan. PloS One 16, e0260031. doi: 10.1371/journal.pone.0260031
Ali A., Khattak M. N. K., Nawaz M. A., Hameed S. (2021). Conflicts involving brown bear and other large carnivores in the Kalam Valley, Swat, Pakistan. Pak. J. Zool. 53, 1889–1896. doi: 10.17582/journal.pjz/20200604180627
Ali U., Minhas R. A., Awan M. S., Ahmed K. B., Qamar Q. Z., Dar N. I. (2016). Human-grey wolf (Canis lupus linnaeus 1758) conflict in shounther valley, district neelum, azad jammu and kashmir, Pakistan. Pak. J. Zool. 48, 861–868.
Allouche O., Tsoar A., Kadmon R. (2006). Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J. App. Ecol. 43, 6,1223–6,1232. doi: 10.1111/j.1365-2664.2006.01214.x
Almasieh K., Cheraghi M. (2022). Habitat suitability, core habitats and diversity hotspots for the conservation of the mustelid species in Iran. Glob. Ecol. Conserv. 36, e02120. doi: 10.1016/j.gecco.2022.e02120
Almasieh K., Kaboli M., Beier P. (2016). Identifying habitat cores and corridors for the Iranian black bear in Iran. Ursus 27, 18–30. doi: 10.2192/URSUSD-15-00032.1
Almasieh K., Mohammadi A., Alvandi R. (2022). Identifying core habitats and corridors of a near threatened carnivore, striped hyaena (Hyaena hyaena) in southwestern Iran. Sci. Rep. 12, 3425. doi: 10.1038/s41598-022-07386-y
Almasieh K., Rouhi H., Hasti F. (2023a). Identifying core habitats and connectivity paths for the conservation of mouflon (Ovis gmelini) in Western Iran. Glob. Ecol. Conserv. 41, e02377. doi: 10.1016/j.gecco.2023.e02377
Almasieh K., Rouhi H., Kaboodvandpour S. (2019). Habitat suitability and connectivity for the brown bear (Ursus arctos) along the Iran-Iraq border. Eur. J. Wildl. Res. 65, 57. doi: 10.1007/s10344-019-1295-1
Almasieh K., Zamani N., Piri R. (2023b). An ensemble modeling approach to predict spatial risk patches of the Persian leopard-livestock conflicts in Lorestan Province, Iran. Environ. Sci. pollut. R 30, 93002–93013. doi: 10.1007/s11356-023-28963-2
Ansari M., Ghoddousi A. (2018). Water availability limits brown bear distribution at the southern edge of its global range. Ursus 29, 13–24. doi: 10.2192/URSUS-D-16-00017.1
Araújo M. B., New M. (2007). Ensemble forecasting of species distributions. Trends Ecol. Evol. 22, 42–47. doi: 10.1016/j.tree.2006.09.010
Ashrafzadeh M. R., Shahbazinasab K., Mohammadi A., Penteriani V. (2023). Determining the distribution factors of an endangered large carnivore: a case study of the brown bear Ursus arctos population in the Central Zagros Mountains, Southwest Iran. Glob. Ecol. Conserv. 46, e02590. doi: 10.1016/j.gecco.2023.e02590
Barbet-Massin M., Jiguet F., Albert C. H., Thuiller W. (2012). Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 3, 327–338. doi: 10.1111/j.2041-210X.2011.00172.x
Bellemain E., Nawaz M. A., Valentini A., Swenson J. E., Taberlet P. (2007). Genetic tracking of the brown bear in northern Pakistan and implications for conservation. Biol. Conserv. 134, 537–547. doi: 10.1016/j.biocon.2006.09.004
Bischof R., Hameed S., Ali H., Kabir M., Younas M., Shah K. A., et al. (2014). Using time-to-event analysis to complement hierarchical methods when assessing determinants of photographic detectability during camera trapping. Methods Ecol. Evol. 5, 44–53. doi: 10.1111/2041-210X.12115
Bischof R., Steyaert S. M., Kindberg J. (2017). Caught in the mesh: Roads and their network-scale impediment to animal movement. Ecography 40, 1369–1380. doi: 10.1111/ecog.02801
Ciucci P., Tosoni E., Di Domenico G., Quattrociocchi F., Boitani L. (2014). Seasonal and annual variation in the food habits of Apennine brown bears, central Italy. J. Mammal. 95, 572–586. doi: 10.1644/13-MAMM-A-218
Coogan S. C., Coops N. C., Janz D. M., Cattet M. R. L., Kearney S. P., Stenhouse G. B., et al. (2018). Towards grizzly bear population recovery in a modern landscape. J. Appl. Ecol. 56, 93–99. doi: 10.1111/1365-2664.13259
Cushman S. A., Elliot N. B., Bauer D., Kesch K., Bahaa-El-Din L., Bothwell H., et al. (2018). Prioritizing core areas, corridors and conflict hotspots for lion conservation in southern Africa. PloS One 13, e0196213. doi: 10.1371/journal.pone.0196213
Cushman S. A., Lewis J. S., Landguth E. L. (2014). Why did the bear cross the road? Comparing the performance of multiple resistance surfaces and connectivity modeling methods. Diversity 6, 844–854. doi: 10.3390/d6040844
Dar S. A., Singh S. K., Wan H. Y., Cushman S. A., Bashir T., Sathyakumar S. (2023). Future land use and climate change escalate connectivity loss for Himalayan brown bears. Anim. Conserv. 26, 199–215. doi: 10.1111/acv.12813
Dar S. A., Singh S. K., Wan H. Y., Kumar V., Cushman S. A., Sathyakumar S. (2021). Projected climate change threatens Himalayan brown bear habitat more than human land use. Anim. Conserv. 24, 659–676. doi: 10.1111/acv.12671
Di Febbraro M., Bosso L., Fasola M., Santicchia F., Aloise G., Lioy S., et al. (2023). Different facets of the same niche: Integrating citizen science and scientific survey data to predict biological invasion risk under multiple global change drivers. Glob. Chang. Biol. 29, 5509–5523. doi: 10.1111/gcb.16901
Dixo M., Metzger J. P., Morgante J. S., Zamudio K. R. (2009). Habitat fragmentation reduces genetic diversity and connectivity among toad populations in the Brazilian Atlantic Coastal Forest. Biol. Conserv. 142, 1560–1569. doi: 10.1016/j.biocon.2008.11.016
ESA (2009). GlobCover land-cover (global land cover map). Available online at: http://due.esrin.esa.int/page_globcover.php (Accessed 20 May 2022).
Eskildsen A., Roux P. C., Heikkinen R. K., Høye T. T., Kissling W. D., Pöyry J., et al. (2013). Testing species distribution models across space and time: high latitude butterflies and recent warming. Glob. Ecol. Biogeogr. 22, 1293–1303. doi: 10.1111/geb.12078
Farashi A., Parvian N., Shariati N. M. (2016). Land use and land cover change in protected areas: using remote sensing to survey suitable habitats of brown bear ursus arctos. Pol. J. Ecol. 64, 420–430. doi: 10.3161/15052249PJE2016.64.3.011
Fernández N., Selva N., Yuste C., Okarma H., Jakubiec Z. (2012). Brown bears at the edge: Modeling habitat constrains at the periphery of the Carpathian population. Biol. Conserv. 153, 134–142. doi: 10.1016/j.biocon.2012.04.013
Fida T., Ahmad F., Bosso L., Ali N., Ud Din S., Kabir M. (2024). Distribution, diel activity patterns and human-bear interactions of the Himalayan brown bear (Ursus arctos isabellinus) in the Deosai National Park, Pakistan. Mamm. Res. doi: 10.1007/s13364-024-00760-3
Fielding A. H., Bell J. F. (1997). A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 24, 38–49. doi: 10.1017/S0376892997000088
Freeman M. S., Dick J. T., Reid N. (2022). Dealing with non-equilibrium bias and survey effort in presence-only invasive Species Distribution Models (iSDM); Predicting the range of muntjac deer in Britain and Ireland. Ecol. Inform. 69, 101683. doi: 10.1016/j.ecoinf.2022.101683
Galbreath G. J., Groves C. P., Waits L. P. (2007). Genetic resolution of composition and phylogenetic placement of the isabelline bear. Ursus 18, 129–131. doi: 10.2192/1537-6176(2007)18[129:GROCAP]2.0.CO;2
Gao P., Kupfer J. A., Guo D., Lei T. L. (2013). Identifying functionally connected habitat compartments with a novel regionalization technique. Land. Ecol. 28, 1949–1959. doi: 10.1007/s10980-013-9938-1
Goursi U. H., Anwar M., Bosso L., Nawaz M. A., Kabir M. (2021). Spatial distribution of the threatened Asiatic black bear in northern Pakistan. Ursus 32e13), 1–5. doi: 10.2192/URSUS-D-19-00031.3
Gubbi S., Menon A., Suthar S., Prabhu K., Poornesha H. C. (2023). Home range estimate of sloth bear using noninvasive camera-trap data. Ursus 2023, 1–6. doi: 10.2192/URSU-D-22-00004
Haq F., Rahatullah H. A. (2012). Diversity and population status of Ursus arctos in Pakistan. I. J. B. 2, 12–20.
Herrero S., Smith T., DeBruyn T. D., Gunther K., Matt C. A. (2005). From the field: brown bear habituation to people—safety, risks, and benefits. Wild Soc Bull. 33, 362–373. doi: 10.2193/0091-7648(2005)33[362:FTFBBH]2.0.CO;2
Hosseini S. P., Amiri M., Senn J. (2022). The effect of environmental and human factors on the distribution of brown bear (Ursus arctos isabellinus) in Iran. Appl. Ecol. Environ. Res. 20, 153–170. doi: 10.15666/aeer/2001_153170
Kabir M., Hameed S., Ali H., Bosso L., Din J. U., Bischof R., et al. (2017). Habitat suitability and movement corridors of grey wolf (Canis lupus) in Northern Pakistan. PloS One 12, e0187027. doi: 10.1371/journal.pone.0187027
Kaboodvandpour S., Almasieh K., Zamani N. (2021). Habitat suitability and connectivity implications for the conservation of the Persian leopard along the Iran-Iraq border. Ecol. Evol. 11, 13464–13474. doi: 10.1002/ece3.8069
Kaczensky P., Knauer F., Krze B., Jonozovic M., Adamic M., Gossow H. (2003). The impact of high speed, high volume traffic axes on brown bears in Slovenia. Biol. Conserv. 111, 191–204. doi: 10.1016/S0006-3207(02)00273-2
Karlsson J. M., Lyon S. W., Destouni G. (2012). Thermokarst lake, hydrological flow and water balance indicators of permafrost change in Western Siberia. J. Hydrol. 464, 459–466. doi: 10.1016/j.jhydrol.2012.07.037
Kaszta Ż., Cushman S. A., Hearn A. J., Burnham D., Macdonald E. A., Goossens B., et al. (2019). Integrating Sunda clouded leopard (Neofelis diardi) conservation into development and restoration planning in Sabah (Borneo). Biol. Conserv. 235, 63–76. doi: 10.1016/j.biocon.2019.04.001
Kaszta Ż., Cushman S. A., Htun S., Naing H., Burnham D., Macdonald D. W. (2020). Simulating the impact of Belt and Road initiative and other major developments in Myanmar on an ambassador felid, the clouded leopard, Neofelis nebulosa. Landsc. Ecol. 35, 727–746. doi: 10.1007/s10980-020-00976-z
Kavčič I., Adamič M., Kaczensky P., Krofel M., Kobal M., Jerina K. (2015). Fast food bears: Brown bear diet in a human-dominated landscape with intensive supplemental feeding. Wild. Biol. 21, 1–8. doi: 10.2981/wlb.00013
Kumar V., Sharief A., Dutta R., Mukherjee T., Joshi B. D., Thakur M., et al. (2022). Living with a large predator: Assessing the root causes of Human–brown bear conflict and their spatial patterns in Lahaul valley, Himachal Pradesh. Ecol. Evol. 12, e9120. doi: 10.1002/ece3.9120
Lacy H., De Cuyper A., Dalerum F., Tosoni E., Clauss M., Ciucci P., et al. (2024). Estimating body condition of Apennine brown bears using subjective scoring based on camera trap photographs. Mamm. Res. 69, 355–364. doi: 10.1007/s13364-024-00755-0
Landis J. R., Koch G. G. (1977). An application of hierarchical kappa-type statistics in the assessment of majority agreement among multiple observers. Biometrics 33, 363–374. doi: 10.2307/2529465
Lecis R., Norris K. (2003). Geographical distribution of the endemic Sardinian brook salamander, Euproctus platycephalus, and implications for its conservation. Herpetol. J. 13, 125–133.
Margules C. R., Pressey R. L. (2000). Systematic conservation planning. Nature 405, 243–253. doi: 10.1038/35012251
Martin J., Basille M., Van Moorter B., Kindberg J., Allaine D., Swenson J. E. (2010). Coping with human disturbance: spatial and temporal tactics of the brown bear (Ursus arctos). Can. J. Zool. 88, 875–883. doi: 10.1139/Z10-053
Martin J., Revilla E., Quenette P. Y., Naves J., Allainé D., Swenson J. E. (2012). Brown bear habitat suitability in the Pyrenees: transferability across sites and linking scales to make the most of scarce data. J. Appl. Ecol. 49, 621–631. doi: 10.1111/j.1365-2664.2012.02139.x
Mateo-Sánchez M. C., Balkenhol N., Cushman S., Pérez T., Domínguez A., Saura S. (2015). A comparative framework to infer landscape effects on population genetic structure: Are habitat suitability models effective in explaining gene flow? Landsc. Ecol. 30, 1405–1420. doi: 10.1007/s10980-015-0194-4
Mateo-Sánchez M. C., Cushman S. A., Saura S. (2014). Scale dependence in habitat selection: the case of the endangered brown bear (Ursus arctos) in the Cantabrian Range (NW Spain). Int. J. Geogr. Inf. Sci. 28, 1531–1546. doi: 10.1080/13658816.2013.776684
McRae B. H., Beier P. (2007). Circuit theory predicts gene flow in plant and animal populations. Proc. Natl. Acad. Sci. 104, 19885–19890. doi: 10.1073/pnas.0706568104
McRae B. H., Dickson B. G., Keitt T. H., Shah V. B. (2008). Using Circuit theory to model connectivity in ecology, evolution and conservation. Ecology 89, 2712–2724. doi: 10.1890/07-1861.1
McRae B. H., Shah V. B. (2009). Circuitscape user’s guide (Santa: The university of California). Available online at: http://www.circuitscape.org (Accessed 10 Dec 2022).
Mech S. G., Hallett J. G. (2001). Evaluating the effectiveness of corridors: a genetic approach. Conserv. Biol. 15, 467–474. doi: 10.1046/j.1523-1739.2001.015002467.x
Melo-Dias M., Huatuco J. F., Arizapana-Almonacid M. A., Castañeda-Tinco M. I., Chanamé F., Passamani M. (2024). Lighting up mountain coexistence: Understanding the effects of environment and livestock on habitat use by Andean bear in a conflict zone in Peruvian Andes. J. Nat. Cons. 81, 126677. doi: 10.1016/j.jnc.2024.126677
Mohammadi A., Almasieh K., Clevenger A. P., Fatemizadeh F., Rezaei A., Jowkar H., et al. (2018). Road expansion: A challenge to conservation of mammals, with particular emphasis on the endangered Asiatic cheetah in Iran. J. Nat. Conserv. 43, 8–18. doi: 10.1016/j.jnc.2018.02.011
Mohammadi A., Almasieh K., Nayeri D., Adibi M. A., Wan H. Y. (2022). Comparison of habitat suitability and connectivity modelling for three carnivores of conservation concern in an Iranian montane landscape. Landsc. Ecol. 37, 411–430. doi: 10.1007/s10980-021-01386-5
Mohammadi A., Almasieh K., Nayeri D., Ataei F., Khani A., LópezBao J. V., et al. (2021a). Identifying priority core habitats and corridors for effective conservation of brown bears in Iran. Sci. Rep. 11, 1044. doi: 10.1038/s41598-020-79970-z
Mohammadi A., Fatemizadeh F. (2021). Quantifying landscape degradation following construction of a highway using landscape metrics in southern Iran. Front. Ecol. Evol. 9. doi: 10.3389/fevo.2021.721313
Mohammadi A., Lunnon C., Moll R. J., Tan C. K. W., Hobeali K., Behnoud P., et al. (2021b). Contrasting responses of large carnivores to land use management across an Asian montane landscape in Iran. Biodivers. Conserv. 30, 4023–4037. doi: 10.1007/s10531-021-02290-9
Naimi B., Hamm N. A. S., Groen T. A., Skidmore A. K., Toxopeus A. G. (2014). Where is positional uncertainty a problem for species distribution modeling? Ecography 37, 191–203. doi: 10.1111/j.1600-0587.2013.00205.x
Nawaz M. A. (2007). Status of the brown bear in Pakistan. Ursus 18, 89–100. doi: 10.2192/1537-6176(2007)18[89:SOTBBI]2.0.CO;2
Nawaz M. A. (2008). Ecology, genetics and conservation of Himalayan brown bears (Ås, Norway: Department of Ecology and Natural Resource Management, Norwegian University of Life Sciences).
Nawaz M. A., Martin J., Swenson J. E. (2014). Identifying key habitats to conserve the threatened brown bear in the Himalaya. Biol. Conserv. 170, 198–206. doi: 10.1016/j.biocon.2013.12.031
Nawaz M. A., Swenson J. E., Zakaria V. (2008). Pragmatic management increases a flagship species, the Himalayan brown bears, in Pakistan’s Deosai National Park. Biol. Cons. 141, 2230–2241. doi: 10.1016/j.biocon.2008.06.012
Nawaz M. A., Valentini A., Khan N. K., Miquel C., Taberlet P., Swenson J. E. (2019). Diet of the brown bear in Himalaya: Combining classical and molecular genetic techniques. PloS One 14, e0225698. doi: 10.1371/journal.pone.0225698
Nellemann C., Støen O. G., Kindberg J., Swenson J. E., Vistnes I., Ericsson G., et al. (2007). Terrain use by an expanding brown bear population in relation to age, recreational resorts and human settlements. Biol. Conserv. 138, 157–165. doi: 10.1016/j.biocon.2007.04.011
Penteriani V., Zarzo-Arias A., Delgado M. M., Dalerum F., Gurarie E., Peón Torre P., et al. (2020). Female brown bears use areas with infanticide risk in a spatially confined population. Ursus 2020, 1–9. doi: 10.2192/URSUS-D-18-00019R4
Phillips P., Clark M. M., Baral S., Koen E. L., Bowman J. (2021). Comparison of methods for estimating omnidirectional landscape connectivity. Landscape Ecology 36, 1647–1661. doi: 10.1007/s10980-021-01254-2
R Core Team (2019). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/ (Accessed 20 December 2021).
Rehan M., Hassan A., Zeb S., Ullah S., Ahmad F., Bohnett E., et al. (2024). Application of species distribution models to estimate and manage the Asiatic black bear (Ursus thibetanus) habitat in the Hindu Kush Mountains, Pakistan. Eur. J. Wildlife. Res. 70, 1–11. doi: 10.1007/s10344-024-01806-2
Rezaei S., Mohammadi A., Bencini R., Rooney T., Naderi M. (2022a). Identifying connectivity for two sympatric carnivores in human-dominated landscapes in central Iran. PloS One 17, e0269179. doi: 10.1371/journal.pone.0269179
Rezaei S., Mohammadi A., Malakoutikhah S., Khosravi R. (2022b). Combining multiscale niche modeling, landscape connectivity, and gap analysis to prioritize habitats for conservation of striped hyaena (Hyaena hyaena). PloS One 17, e0260807. doi: 10.1371/journal.pone.0260807
Roever C. L., van Aarde R. J., Leggett K. (2013). Functional connectivity within conservation networks: delineating corridors for African elephants. Biol. Conserv. 157, 128–135. doi: 10.1016/j.biocon.2012.06.025
Sarkar M. S., Segu H., Bhaskar J. V., Jakher R., Mohapatra S., Shalini K., et al. (2018). Ecological preferences of large carnivores in remote, high-altitude protected areas: insights from Buxa Tiger Reserve, India. Oryx 52, 66–77. doi: 10.1017/S0030605317000060
Scolozzi R., Geneletti D. (2012). A multi-scale qualitative approach to assess the impact of urbanization on natural habitats and their connectivity. Environ. Impact. Assess. Rev. 36, 9–22. doi: 10.1016/j.eiar.2012.03.001
Servheen C. (1990). The status and conservation of the bears of the world. Int. Conf. Bear. Res. Manage. Monogr. Ser. 2, 311–316.
Sharief A., Joshi B. D., Kumar V., Kumar M., Dutta R., Sharma C. M., et al. (2020). Identifying Himalayan brown bear (Ursus arctos isabellinus) conservation areas in Lahaul Valley, Himachal Pradesh. Glob. Ecol. Conserv. 21, e00900. doi: 10.1016/j.gecco.2019.e00900
Sheikh K. M., Molur S. (2005). Status and red list of Pakistan’s mammals, based on conservation assessment and management plan for mammals. IUCN. Pakistan 344, 1–311.
Singh A. P., De K., Uniyal V. P., Sathyakumar S. (2024). Livestock depredation by large carnivores in Western Himalayan region of Jammu and Kashmir: temporal adherence in predator’s choice. Trop. Ecol. 65, 26–31. doi: 10.1017/S0030605317000060
Straka M., Paule L., Ionescu O., Štofík J., Adamec M. (2012). Microsatellite diversity and structure of Carpathian brown bears (Ursus arctos): consequences of human caused fragmentation. Conserv. Genet. 13, 153–164. doi: 10.1007/s10592-011-0271-4
Su J., Aryal A., Hegab I. M., Shrestha U. B., Coogan S. C., Sathyakumar S., et al. (2018). Decreasing brown bear (Ursus arctos) habitat due to climate change in Central Asia and the Asian Highlands. Ecol. Evol. 8, 11887–11899. doi: 10.1002/ece3.4645
Swets J. A. (1988). Measuring the accuracy of diagnostic systems. Science 240, 1285–1293. doi: 10.1126/science.3287615
Tang T., Li J., Sun H., Velázquez J. (2023). Integrated approach considering seasonal variability and habitat uncertainty to map habitat for the prey of South China tiger. Ecol. Ind. 148, 110082. doi: 10.1016/j.ecolind.2023.110082
Ten Caten C., Lima-Ribeiro M. S., Yañez-Arenas C., Villalobos F., Díaz-Gamboa L., Terribile L. C. (2023). Robustness of Bergmann’s and Rapoport’s rules to different geographical range estimates in New World pit vipers. J. Biogeogr. 50, 365–379. doi: 10.1111/jbi.14538
Thakur S., Pal R., Kahera N. S., Sathyakumar S. (2023). Forced sympatry? Spatiotemporal interactions of ursids, the Himalayan brown bear and the Asiatic black bear, along a gradient of anthropic disturbances in Western Himalaya. J. Zool. 321, 59–74. doi: 10.1111/jzo.13090
Thuiller W., Georges D., Engler R., Breiner F. (2019). Biomod2: ensemble platform for species distribution modeling. R package version 3.3-7.1. Available online at: https://CRAN.R-project.org/package=biomod2 (Accessed 20 February 2022).
Urban D., Keitt T. (2001). Landscape connectivity: a graph-theoretic perspective. Ecology 82, 1205–1218. doi: 10.1890/0012-9658(2001)082[1205:LCAGTP]2.0.CO;2
Urban D. L., Minor E. S., Treml E. A., Schick R. S. (2009). Graph models of habitat mosaics. Ecol. Lett. 12, 260–273. doi: 10.1111/j.1461-0248.2008.01271.x
Velázquez J., Gülçin D., Vogt P., Rincón V., Hernando A., Gutiérrez J., et al. (2022). Planning restoration of connectivity and design of corridors for biodiversity conservation. Forests 13, 2132. doi: 10.3390/f13122132
Velázquez J., Gutiérrez J., Hernando A., García-Abril A. (2017). Evaluating landscape connectivity in fragmented habitats: Cantabrian capercaillie (Tetrao urogallus cantabricus) in northern Spain. For. Ecol. Manage. 389, 59–67. doi: 10.1016/j.foreco.2016.12.006
Virk A. T., Sheikh K. M., Marwat A. H. (2003). Northern Areas strategy for sustainable development backgroud paper: biodiversity Vol. 73 (Gilgit, PK: IUCN Northern Areas Programme).
Wan H. Y., Cushman S. A., Ganey J. L. (2019). Improving habitat and connectivity model predictions with multi-scale resource selection functions from two geographic areas. Landsc. Ecol. 34, 503–519. doi: 10.1007/s10980-019-00788-w
Wolf C., Ripple W. J. (2017). Range contractions of the world’s large carnivores. R. Soc Open Sci. 4, 170052. doi: 10.1098/rsos.170052
Wolfson D. W., Schlichting P. E., Boughton R. K., Miller R. S., VerCauteren K. C., Lewis J. S. (2023). Comparison of daily activity patterns across seasons using GPS telemetry and camera trap data for a widespread mammal. Ecosphere 14, e4728. doi: 10.1002/ecs2.4728
Zedrosser A., Steyaert S. M., Gossow H., Swenson J. E. (2011). Brown bear conservation and the ghost of persecution past. Biol. Cons. 144, 2163–2170. doi: 10.1016/j.biocon.2011.05.005
Zeller K. A., Wattles D. W., Destefano S. (2020). Evaluating methods for identifying large mammal road crossing locations: black bears as a case study. Landsc. Ecol. 35, 1799–1808. doi: 10.1007/s10980-020-01057-x
Ziółkowska E., Ostapowicz K., Radeloff V. C., Kuemmerle T., Sergiel A., Zwijacz-Kozica T., et al. (2016). Assessing differences in connectivity based on habitat versus movement models for brown bears in the Carpathians. Landsc. Ecol. 31, 1863–1882. doi: 10.1007/s10980-016-0368-8
Keywords: conservation, Himalayan brown bear, landscape connectivity, species distribution models, wildlife management
Citation: Fida T, Mohammadi A, Almasieh K, Bosso L, Ud Din S, Shamas U, Nawaz MA and Kabir M (2025) Species distribution modelling and landscape connectivity as tools to inform management and conservation for the critically endangered Himalayan brown bear (Ursus arctos isabellinus) in the Deosai National Park, Pakistan. Front. Ecol. Evol. 12:1477480. doi: 10.3389/fevo.2024.1477480
Received: 07 August 2024; Accepted: 12 December 2024;
Published: 13 January 2025.
Edited by:
Oana Moldovan, Romanian Academy, RomaniaReviewed by:
Francesco Bisi, University of Insubria, ItalyCopyright © 2025 Fida, Mohammadi, Almasieh, Bosso, Ud Din, Shamas, Nawaz and Kabir. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Muhammad Kabir, a2FiaXJfYWprQGhvdG1haWwuY29t; Luciano Bosso, bHVjaWFuby5ib3Nzb0BjbnIuaXQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.