 Ke Xin Sun
Ke Xin Sun Yan Jia Zhang2
Yan Jia Zhang2- 1College of Landscape Architecture, Changchun University, Changchun, China
- 2College of Communication, Changchun Humanities and Sciences College, Changchun, China
Introduction: Gradual pollen presentation is a reproductive strategy for plants to improve the efficiency and accuracy of pollen export and acceptance as well as to promote outcrossing. Vaccinium corymbosum Berkeley has a typical mechanism of gradual pollen presentation.
Methods: In this study, we used indoor and outdoor observations to investigate the floral characteristics, pollination characteristics and breeding system of this species in open ground and greenhouse.
Results: The results showed that the single white, inverted bell-shaped flower of V. corymbosum was able to effectively avoid the negative effects of rainfall, nectar dilution and pollen removal by wind, and improved pollen viability and the duration of stigma receptivity. The setting rate of ‘Northland’ is higher than open ground by artificial pollination in greenhouse, therefore, greenhouses are more suitable for ‘Northland’ reproduction. The gradual pollen presentation prolongs dispersal time, promotes pollinator participation in the pollination process, increases staminate fitness, and provides for nectar feeding by pollinators as well as pollen export. Bombus spp. and A. m. ligustica were effective pollinators in the open ground and greenhouse, and the pollination efficiency of Bombus spp. was higher than that of A. m. ligustica, and greenhouse pollination is more efficient than open ground. The coincidence of the peak period of pollinator visits with the period of highest pollen viability and optimal pollination of stigmas, as well as the high nectar production, could maximize the frequency of pollinator visits and increase the chances of successful pollination as well as the pollination efficiency.
Discussion: Therefore, it is important to study the floral characteristics of V. corymbosum Berkeley and the mechanism of gradual pollen presentation in order to improve the pollination efficiency and promote the success of outcrossing, which can provide a theoretical basis for the breeding of V. corymbosum.
1 Introduction
Floral characteristics of plants include flower composition and flower opening pattern (Zhang et al., 2024). Flower composition includes flower structure, color, morphology, and nectar production; flower opening pattern refers to the number of flowers open at a certain time and the spatial arrangement of flowers on the inflorescence, i.e., the flower expression characteristics at the group level (Xie et al., 2023; Cao et al., 2022). Studies on plant floral characteristics have focused on flower color, petal structure, petal morphology, flowering time, and pollen release mode, which are important in attracting pollinators, improving pollen dispersal efficiency, inducing pollinators to provide reward, pollinators’ flower visiting behavior, and gradual presentation of pollen (Ji et al., 2023; Chai et al., 2017). And attraction to pollinators and pollinators response to floral characteristics, pollination behavior is closely related to the comprehensive characteristics of flowers, which to some extent determines the efficiency of pollination (Yin et al., 2016; Huang and Guo, 2000; Guan et al., 2024; Xiang, 2019).
Environment has a wide range of effects on plant growth, development and reproduction as well as morphological characteristics (Miao, 2018). floral characteristics of most plants often differ markedly from habitat to habitat (Arroyo and Dafni, 1995; Fabbro and Körner, 2004). Plant floral characteristics (petal size, flower color, odor, nectar volume, etc.) change in response to the environment and are considered an adaptation to environmental factors as well as pollinators (Wang, 2021). Temperature, humidity, oxygen concentration and soil conditions in greenhouses affect plant growth, development and yield (Arie, 2004; Masayyuki, 2013; Chen, 2020). Therefore, studying the role of different environments on the growth, development and reproduction of plants can improve the yield and quality of plants.
Gradual pollen presentation by which pollen is released from anthers, flowers or inflorescences one by one by contracting and squeezing the anthers so that pollinators move out of the pollen one by one (Halibunuer et al., 2022; Caster et al., 2008). The mechanism of pollen presentation is influenced not only by the plant’s floral characteristics, but also by the type and efficiency of the pollinator (Wang and Tan, 2011). The gradual presentation of pollen by plants attracts more pollinators to visit, reduces the amount of pollen removed at one time, thus avoiding interference between anthers, reducing pollen limitation, and increasing staminate reproduction and fitness (Halibunuer et al., 2022; Wang and Tan, 2011). In outcrossing pollinators with short flower lifespan, low frequency of flower visits and low pollinator efficiency, gradual pollen presentation is the best strategy to reduce pollen limitation, increase staminate fitness and outcrossing, and safeguard pistillate reproduction (Homson et al., 1994; Gargan and Peruzz, 2021). Therefore, gradual pollen presentation is important in promoting outcrossing pollination, avoiding anther wastage and improving pollination efficiency.
Breeding system refers to all the sexual characteristics that affect the genetic composition of offspring, including flower morphology, opening pattern, flower longevity, pollinator type, frequency of pollinators, and breeding system (Li et al., 2024; Barretts and Harder, 1996). Different types of plant breeding systems vary. Studies on plant breeding systems have mainly focused on Leguminosae, Cruciferae, etc. (Wang et al., 2024), and the types of breeding systems include self-crossing, outcrossing, mixed mating, etc. (Zhang et al., 2024), the breeding system of Hesperis oreophila is autogamy affinity (Dong et al., 2022). Domestic research on plant breeding systems mainly focuses on the examination and the role in plant conservation (Wang et al., 2024). Breeding system is the link between plant adaptation and species formation mechanism, which is important for plant genetic diversity and breeding diversity (Lei et al., 2019; Chen et al., 2019). As one of the five healthy fruits, ‘Northland’ has a strong health function, but also has important economic value. It is suitable for planting in northeast China, and its taste is widely welcomed by people. Therefore, the study of the breeding system and its reproductive success is of great significance to population dynamics and evolution, and can provide a theoretical basis and practical foundation for the efficient reproduction of ‘Northland’.
Vaccinium corymbosum Berkeley in the Ericaceae, is widely distributed in Heilongjiang, Neimenggu, Jilin Changbai Mountain and other regions (Li and Hu, 2013). Ericaceae have special floral characteristics, breeding systems, pollination, and most of them are outcrossing, which can increase the fruiting rate, some varieties are self-fertile, and their degree of reproduction varies from species to species (Yu et al., 2022). ‘Northland’ is common name for the Vaccinium corymbosum Berkeley, ‘Northland’ is represented below. At present, the research on blueberry in this country and abroad mainly focuses on its cultivation technology and breeding, and the research on floral characteristics and breeding system, especially in the comparison of open ground and greenhouse, has not been reported yet. Therefore, in this study, we chose open ground and greenhouse, to follow up and compare the reproductive ecological characteristics of ‘Northland’, aiming at exploring the following issues: (1) anther dispersal patterns during single-flower opening of ‘Northland’, (2) mutual adaptive mechanisms between floral characteristics and pollinators, (3) influence of nectar presentation strategies and pollen dispersal patterns on the process of pollinator visiting, (4) differences between ‘Northland’ breeding systems.
2 Materials and methods
2.1 Study site
Vaccinium corymbosum Berkeley study area in Jian City, Tonghua City, Jilin Province, China (126°18’ E, 41°12’ N), and the open ground and greenhouse were experimented in the same topography, landscape and at the same altitude of 108-1337 m to ensure that the soil condition, climate, and other environmental factors were consistent with the population area of about 500 m2. The overall terrain of the study area is flat, and it belongs to the continental climate in the northern temperate zone, with a large diurnal temperature difference. There is a large temperature difference, with an average annual rainfall of 600-700 mm, an average annual temperature of 5.6°C (Figure 1), an average annual frost-free period of 150 ± 10 d, and an average annual greenhouse temperature of 23°C and relative humidity of 56%. Experimental materials were selected from 100 plants of ‘Northland’ 5-years-old the open ground and greenhouse, respectively.
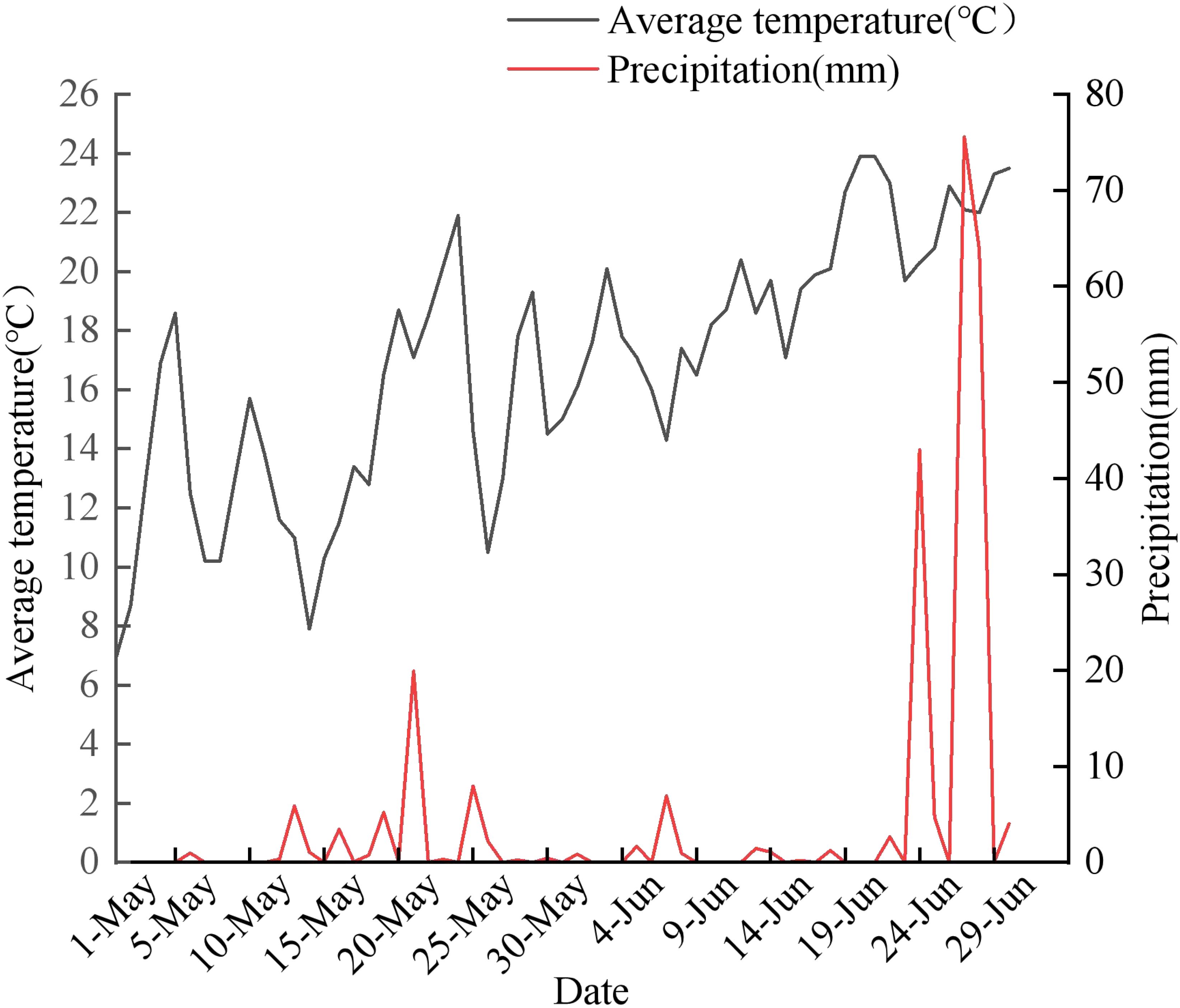
Figure 1. Mean daily temperature and rainfall during flowering of ‘Northland’ in open ground.
2.2 Methods
2.2.1 Floral characteristics and flowering process
In the open ground and greenhouse, 30 flower buds were randomly selected, respectively, observed to record the flowering dynamics. Corolla, filament, anther and stigma lengths were measured at different flowering periods using electronic digital calipers in the laboratory, and anther morphology, corolla morphology, anther color and stigma color were recorded. The flowering time of single flowers was counted and the process of single flower flowering was recorded.
2.2.2 Pollen gradual presentation, pollen grain morphology and number of pollen grains
In the open ground and greenhouse, 240 flower buds were selected, bagged, and observed up to 4 periods of single flower opening, and 30 anthers were removed from the bag at each period, put into clean penicillin vials. After the pollen was fully dispersed, 1 mL of 1% sucrose solution was dropped into each vial, and the vials were filled to 3 mL with a lid and shaken, so that the pollen appeared to be in suspension. A volume of 20 μL of microsampler was used to absorb the suspension on a haematocrit plate with a coverslip, and the number of pollen grains was counted manually under a low-power microscope 100x. The number of pollen grains (N) in a single anther was calculated by taking the average of six repetitions (N = (average number of pollen grains in the field of view × 10,000 × 2)/30), and the above method was used to count the number of pollen grains remaining in the anthers at four periods of the opening process of a single flower, and the process of anther dehiscence and the morphology of the pollen grains were observed and recorded by using a JSM-6510 (Japan) scanning electron microscope (Yang et al., 2019; Dafni et al., 2005).
2.2.3 Pollen viability, stigma receptivity and nectar accumulation
At the flowering periods in the open ground and greenhouse, we collected flowers of ‘Northland’ respectively selected 10 single flowers from the two habitats in each period for the experiment. We took the anthers to the slide, put 1~2 drops of 0.5% TTC solution, so that the pollen was dispersed in the solution. The slides were put in an incubator at 120°C for 20 min and then observed under a light microscope (BM2000). The pollen viability was red, the pollen with weak viability was light red, and the flowers without viability or sterility were colorless (Pan et al., 2012).
In addition, 10 single flowers of ‘Northland’ were collected from four different periods, and the stigmas were removed, placed on concave slides, and dropped into the reaction solution “benzidine-hydrogen” (1% benzidine: 3% hydrogen peroxide: water = 4:11:22). The stigma is blue with a large number of bubbles around it, which is highly receptivity; the stigma is light blue with a small number of bubbles around it, which is relatively weak receptivity; there is no color change of the stigma and no bubbles around it, which is not receptivity (He et al., 2023).
During the opening of single flowers of ‘Northland’, the nectar of single flowers from four different periods (pre-flowering period had been bagged) within the two habitats was sucked up with a point sampling capillary (Nantong Surui Experimental Equipment Company) with an inner diameter of 0.3 mm and a length of 100 mm, 15 single flowers were taken respectively from the two habitats in each period. When the nectar was exhausted, the height of the nectar in the sampling capillary was recorded at different periods. The nectar content (%) of single flowers was calculated using the volume equation: V = πr2h (Liao et al., 2024).
2.2.4 Pollination characteristics
2.2.4.1 Visiting behavior and pollinator identification
May 30 - June 4 (early flowering) and June 5 - June 11 (peak flowering), 2023, 5 plants were randomly selected continuous observations in the open ground. April 25 - April 30 (early flowering) and May 1 - May 6 (peak flowering), 2023, 5 plants were randomly selected continuous observations in the greenhouse, and continuous observations were made from 7:00 to 19:00 in the open ground and greenhouse. The visiting behavior of pollinators on plants, the number of visiting, the inflorescences, and the residence time on a single flower were recorded, and the frequency of visiting by pollinators was calculated (times·flowering-1·h-1). In order to avoid the disturbance of pollinators by observers, all observations were made at a distance of 1 m or more by a person.
There were 10 inflorescences of ‘Northland’ were randomly selected in the open ground and greenhouse respectively. After a single effective visiting, the pollinators were caught with a net and put into a gas bottle made of ethyl acetate to kill them, 5 pollinators of each species were selected for species identification, and the remaining insects were washed several times with 75% alcohol. It was placed under an Olympus BH-2 optical microscope to observe whether the pollinators were carrying pollen of ‘Northland’ to determine whether they were pollinators or not, and Mr. Chen Liu-sheng from Guangdong Forestry Research Institute identified the pollinator specimens for species identification.
2.2.4.2 Pollination efficiency
In the open ground and greenhouse, 10 ‘Northland’ plants were randomly selected respectively, and on each plant, 5 branches were randomly selected, and in the middle of each branch, one inflorescence was selected, and 10 closed buds were retained on each inflorescence. The inflorescences were destaminated, bagged and tagged, and more than 80% of the buds on the inflorescences opened after one week. After a fully open single flower was first visited by a pollinator, the stigma was removed and placed on a slide evenly covered with Vaseline (Stigma maturity evaluation based on morphology and color), rolled evenly, and the sampling was repeated 20 times. Immediately brought back to the laboratory for microscopic examination, the number of pollen on the slides was counted through an Olympus BH-2 light microscope, and the amount of pollen dropped on the stigma after pollinator was counted (Han et al., 2008). In order to detect the number of pollen grains taken away by pollinators after a single visit, the same treatment was performed as above, and the flowers that had not opened were destaminated and bagged, and the bag was removed after the stigma unfolded at the pistillate stage of flowering, and the stigmas were removed and crushed immediately after a single visit by pollinators, and the number of pollen grains remaining in the anthers was counted. The anthers were stained with Senka stain to 2 mL, and 5 μL of the anthers were observed under an Olympus BH-2 light microscope to count the number of pollen grains. The stained part was considered to be the pollen grains brought by the pollinator from the rest of the flowers, and the unstained part was the remaining pollen number. The number of pollen grains taken away by the pollinators in a single visit = the original number of pollen grains - the number of pollen grains remaining after a single visit (Conner et al., 1995).
Pollination efficiency (%) = (Number of pollen dropped on the stigma by pollinators in a single visit/Number of pollen taken away from by pollinators in a single visit) × 100%.
2.2.5 Correlation analysis between nectar accumulation, pollen grains number and visiting frequency
The correlations between nectar accumulation, pollen grains number and visiting frequency in the early and peak flowering of ‘Northland’ were analyzed in open ground and greenhouse.
2.2.6 Pollination experiment
The type of breeding system was detected by an artificial bag pollination experiment, in which 60 plants with essentially the same development were randomly selected in the open ground and greenhouse, and 4 single flowers were selected on each plant for treatment. (1) Natural Pollination: randomly selected single flowers in peak flowering in their natural state and counted their fruiting rate after they had set fruit; (2) Non-emasculated bagging: randomly selected its flowers are not open that is, the bud stage of the plant (stamens are not pollinated) for bagging treatment, to be fruiting after the statistics of its fruiting rate, to observe the degree of self-crossing affinity; (3) The same plant cross-pollination: artificially removing staminates, pollinating different single flowers of the same plant, bagging, counting the fruiting rate after fruiting, and observing the degree of acceptance of the same plant cross-pollination; (4) Different plants cross-pollination: artificially remove the staminate, pollinate different plants with different single flowers, and then count the fruiting rate after fruiting to observe the acceptance of different plants cross-pollination. Each treatment was set up with 60 replications.
2.2.7 Fruit quality
A total of 20 plants were randomly selected from open ground and greenhouse, three fruits were randomly selected from different fruiting parts. A total of 180 fruits were selected for determination of fruit shape index: the longitudinal and transverse diameters of the fruits were measured using electronic digital calipers (0.01 mm). Total sugar content (%) was determined by anthrone method. Titratable acid content (TA, %) was determined by acid-base neutralization titration, VC (VC, mg/100 g) was determined by 2,6-dichloroindophenol titration (Cao et al., 2007).
2.3 Data analysis
Data were organized using Excel 2018, and the data of corolla, filament, anther and stigma length, pollen number, pollen viability, visiting frequency, nectar secretion, and pollination experiment were statistically analyzed and compared differently using SPSS 19.0 statistical software. Differences in pollen grain number, pollen viability, visiting frequency, nectar secretion, and pollination experiments were compared using independent samples t-test (p < 0.05) in the two habitats, and differences in pollination experiments were compared using one-way ANOVA (p < 0.05). Experimental data are expressed as mean ± standard error. All graphs were produced using Origin 2021 software.
3 Results
3.1 Floral characteristics and flowering process
In the open ground and greenhouse, ‘Northland’ had a single white inverted bell-shaped flower with 10 stamens and 1 pistil. The single-flower opening process was divided into four periods based on the size of the corolla opening (Figure 2), Phase I-corolla closed, Phase II-corolla slightly opened 1-3 mm (measured by corolla opening diameter), Phase III - corolla fully opened, 4-8 mm, and Phase IV-corolla with filament anthers detached together. The four periods is shown in differences in anther morphology, corolla morphology, anther color, and stigma color; the duration of single-flower opening was 13 ± 2 and 10 ± 2 d, with single-flower opening in the open ground being prolonged by about 3 d compared with the greenhouse; the corolla depth, filament, anther and stigma lengths within the greenhouse population were greater than those of the open ground population, with significant differences between filament and anther lengths within the two populations (p < 0.05) and no significant differences between corolla and stigma lengths (p > 0.05) (Table 1).
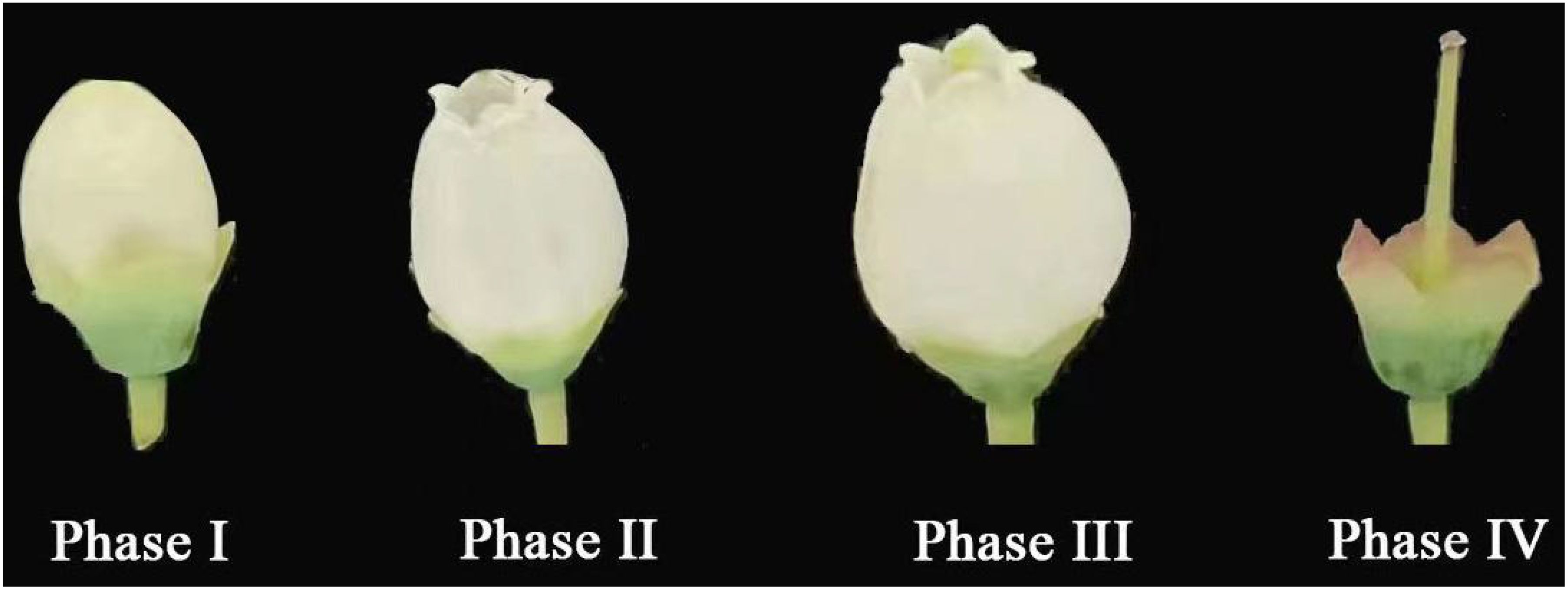
Figure 2. Single flower blooming dynamics opening of ‘Northland’. Phase I: corolla closed; Phase II: corolla slightly opened (1-3 mm); Phase III: corolla fully opened (4-8 mm); Phase IV: corolla detached.
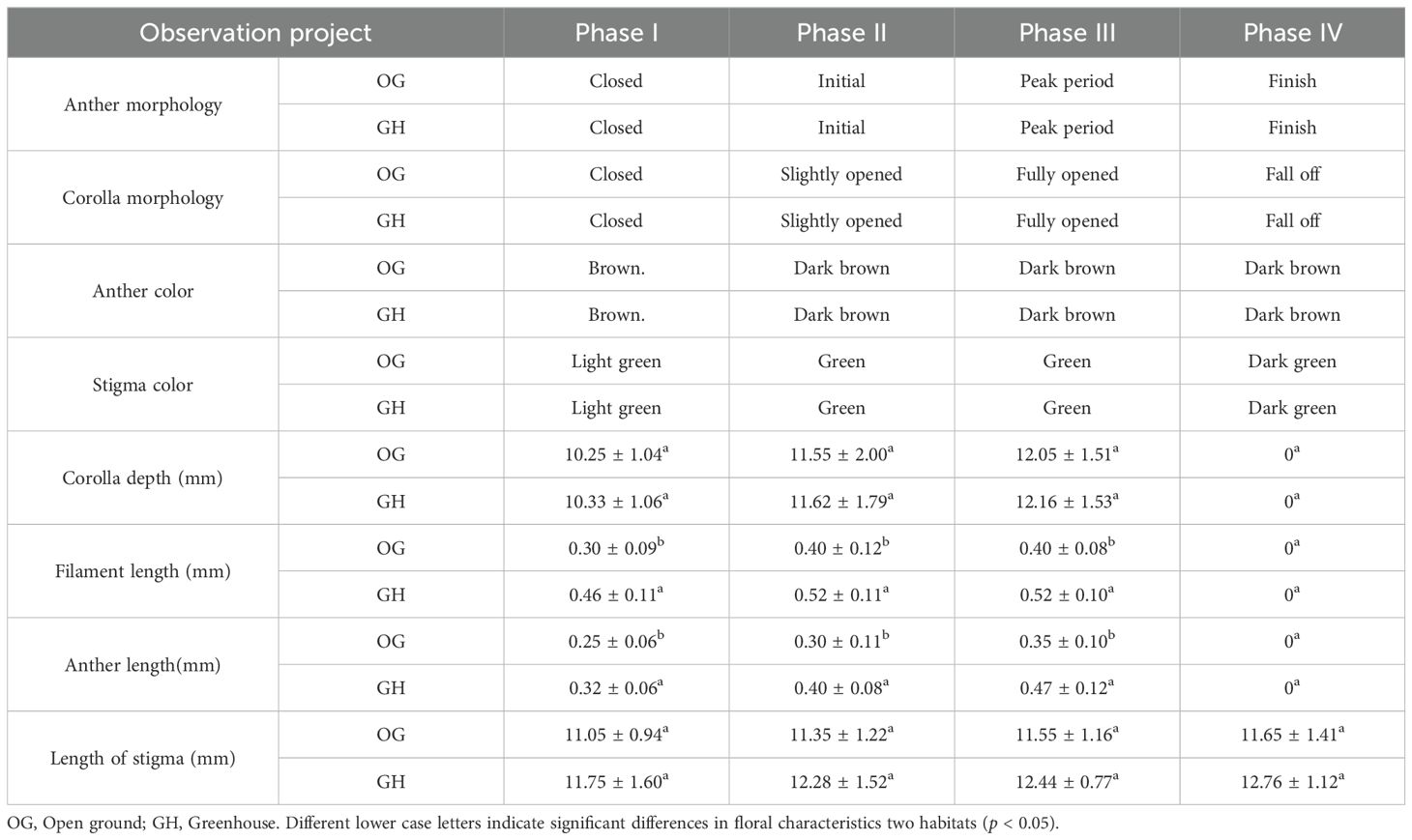
Table 1. Single flower characteristic of ‘Northland’ in two habitats.
3.2 Pollen gradual presentation and pollen grains number
The number of pollen grains in single flower of ‘Northland’ was 19000 ± 2550 and 24000 ± 1821 when the anthers were not dispersed, in the open ground and greenhouse. The number of pollen grains per flower remaining in the anthers at pre-flowering and peak flowering was highly significantly different (p < 0.01) in the open ground and greenhouse, and the number of pollen grains remaining in the anthers at the early flowering period was significantly different (p < 0.05) (Figure 3), and the number of pollen grains remaining in the anthers at each period was 12,600 ± 597 grains, 13,500 ± 716 grains, 6560 ± 216 grains and 7830 ± 155 grains. The number of pollen grains in inflorescence remaining in the anthers was significantly different (p < 0.05) (Figure 3), and the number of pollen grains remaining in the anthers at each period was 2680 ± 155 grains, 3900 ± 347 grains, 1560 ± 131 grains, 2470 ± 182 grains, 840 ± 54 grains and 1260 ± 176 grains. During flowering, the anthers were porous dehiscence, and the pollen grains inside the anthers were gradually released through the split anther top holes above the anthers, and the pollen dispersal of the anthers was in the form of gradual pollen presentation. Scanning electron microscopy showed that the anther top of ‘Northland’ formed two tubular structures connected with four dispensaries, and the epidermal cells of the anther were papillate. The stigma surface had a large number of papilla cells. The pollen surface was irregularly crinkled and corrugated, and the pollen grains were tetrads of complex pollen (Figure 4).
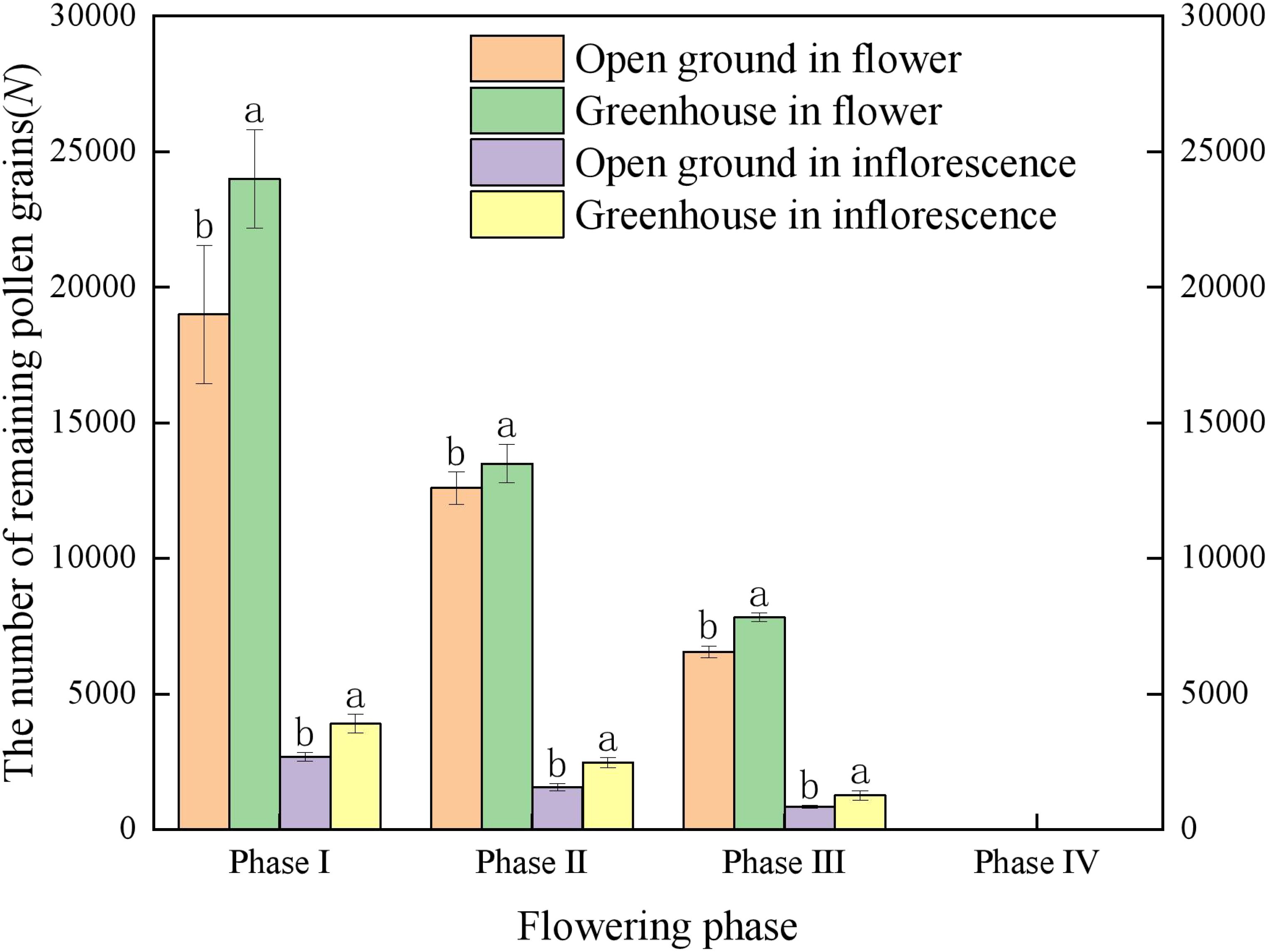
Figure 3. Number of remaining pollen grains at four periods during single flower and inflorescence opening in ‘Northland’. Different lowercase letters indicate different habitats in the same period. environment at the same period of time Significant differences exist between different habitats at the same time period (p < 0.05).
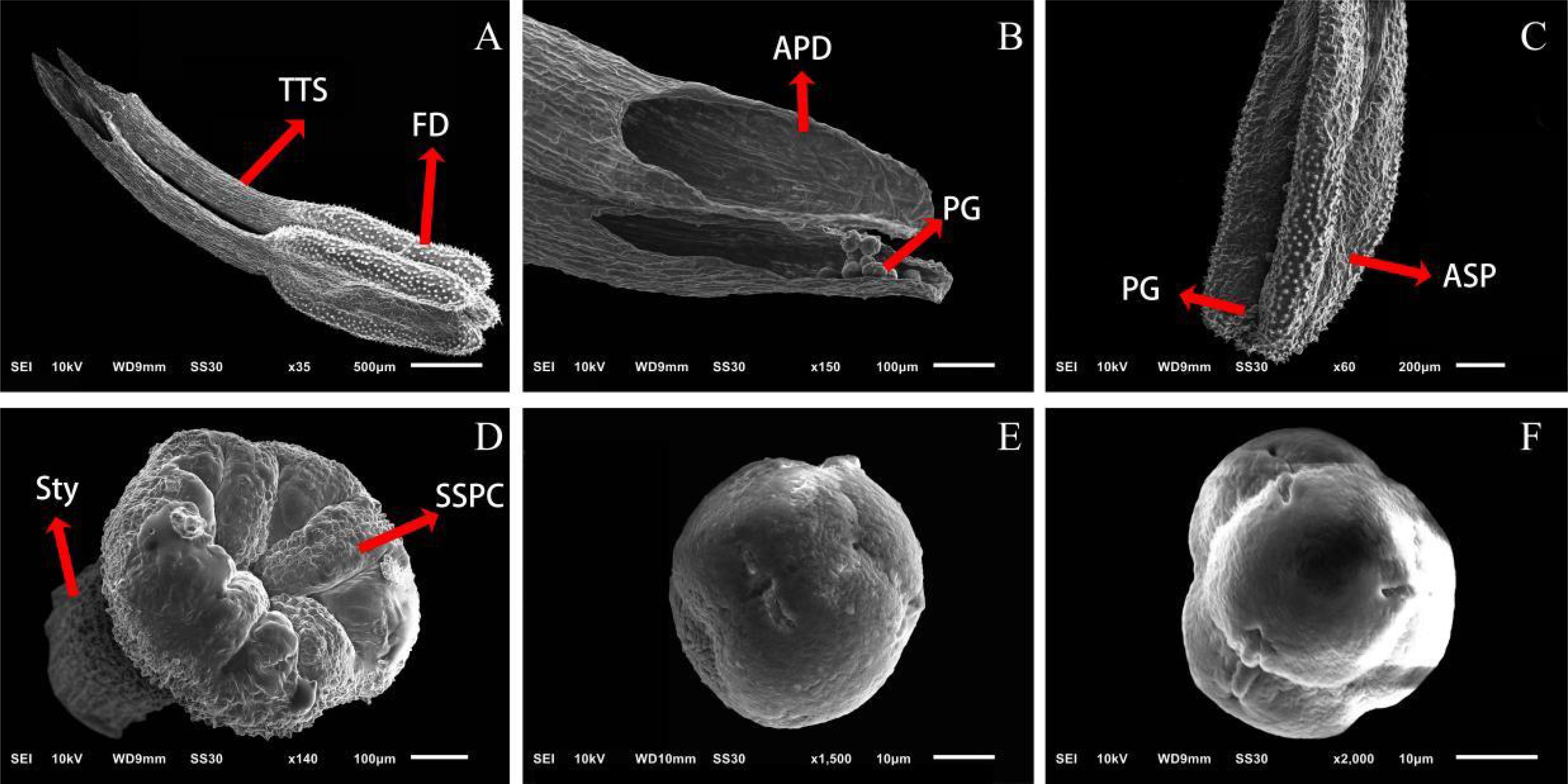
Figure 4. Anther, stigma, and pollen grain morphology of ‘Northland’. (A–C) Anther. TTS, Two tubular structures; FD, Four dispensaries; APD, Anthers porous dehiscence; PG, Pollen grains; ASP, Anther surface papillate; (D) Stigma. SSPC, Stigma surface papilla cells; Sty, style; (E) Frontal view of ‘Northland’ pollen grains; (F) Polar view of ‘Northland’ pollen grain.
3.3 Pollen viability, stigma receptivity, and nectar accumulation
In the open ground and greenhouse, ‘Northland’ had the highest pollen viability at the early flowering, 86 ± 2.9% and 75 ± 2.7% (Figure 5); and there was a highly significant difference in pollen viability between open ground and greenhouse over the three time periods (p < 0.01). The stigma receptivity in the open ground and greenhouse was shown to be stronger in the pre-flowering period, strongest in the early flowering period, reaching a relative maximum, decreasing in the peak flowering period, and weakest at the end of the flowering period in all four periods. With in the open ground and greenhouse, the nectar accumulation of ‘Northland’ was the highest at the peak of flowering at 22 ± 2.5 μL and 18 ± 2.2 μL, respectively, with significant differences between nectar accumulation at the pre-flowering stage (p < 0.05), and highly significant differences between nectar accumulation at the early flowering stage and at the end of flowering (p < 0.01), and no difference existed during the peak period (p > 0.05).
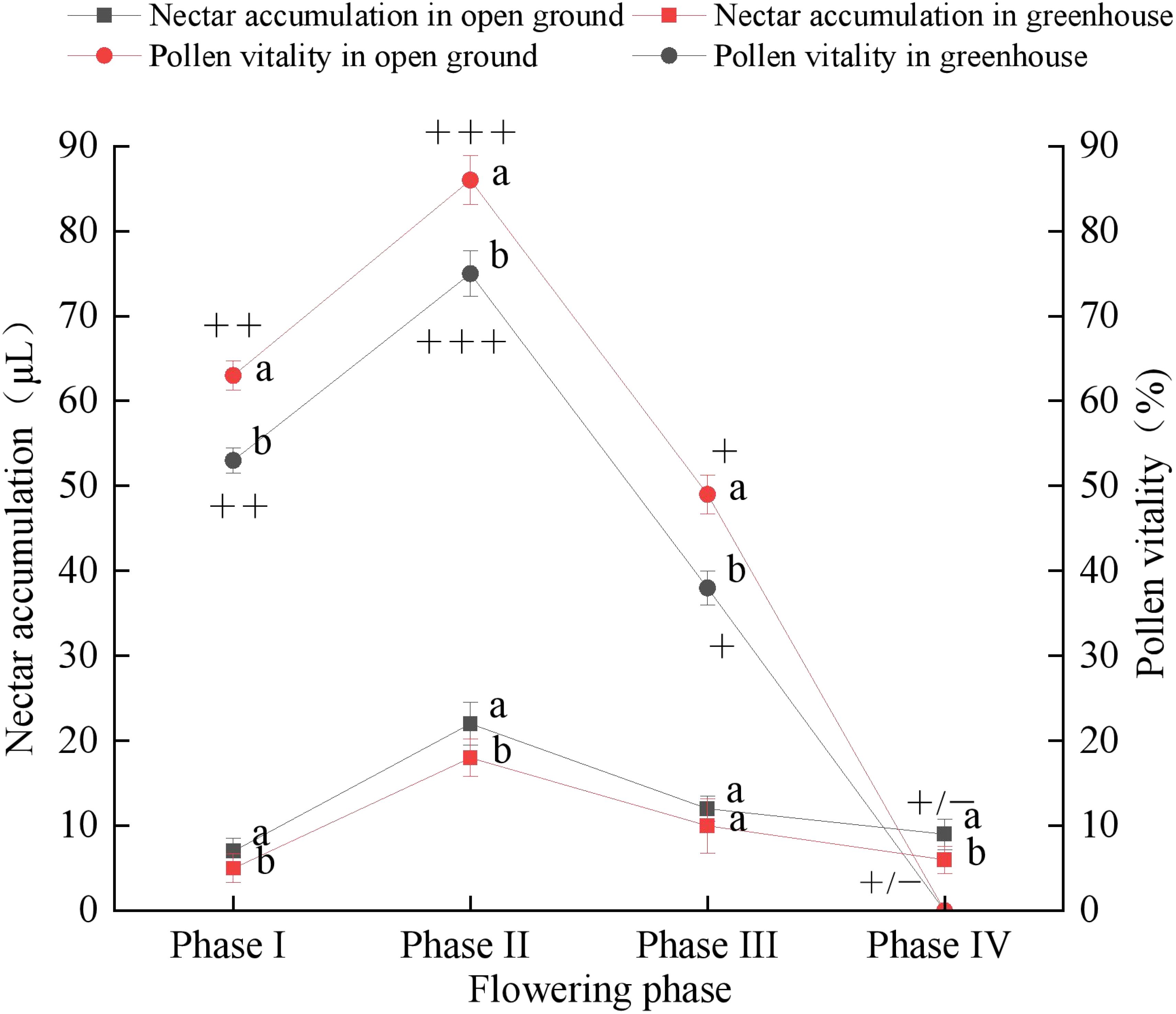
Figure 5. Nectar accumulation and Pollen vitality of ‘Northland’ in four different flowering phase in open ground and greenhouse. Strength of the stigma of ‘Northland’ in terms of its granulability, strongest (+++); Strong (+ + +); Weak (+); Weak (+/-). Different letters indicate significant differences in pollen viability as well as nectar content in different habitats during the same period (p < 0.05).
3.4 Pollination characteristics
3.4.1 Pollinator and visiting behavior
The species and number of pollinators were the same in the open ground and the greenhouse (Table 2). After observing all the pollinator specimens through a microscope, the effective pollinators of ‘Northland’ were Bombus spp. and A. m. ligustica. Syrphid fly, Syrphidae sp., Tetramorium sp., Myrmica sp. and Harmonia axyridis are ineffective pollinators. In the open ground and greenhouse, when Bombus visit flowers, they usually grasp the flowers with their forelegs and then shake the flowers while shaking their wings, which shakes the pollen onto the stigma of the pistil as well as the abdomen of the body, completing the pollination, and then send the pollen to the pollen-carrying foot from the abdomen to carry the pollen away. Some of the released pollen grains adhere to the downy hairs of the Bombus spp. through the vibration of the wings, and act as a cross-pollinated when collecting the next flower. When A. m. ligustica visit flowers, they first extend their head into the corolla, and the back or ventral surface of the cephalothorax touches the stigma to complete pollination, and the forefeet collect pollen onto the hindfeet to complete the pollination process. In the open ground and greenhouse, Bombus spp. stayed on single inflorescences for 25 ± 5 s and 27 ± 3 s, and on single flowers for 12 ± 2 s and 15 ± 3 s. A. m. ligustica stayed on single inflorescences for 20 ± 5 s and 23 ± 5 s, and on single flowers for 8 ± 5 s and 11 ± 3 s.
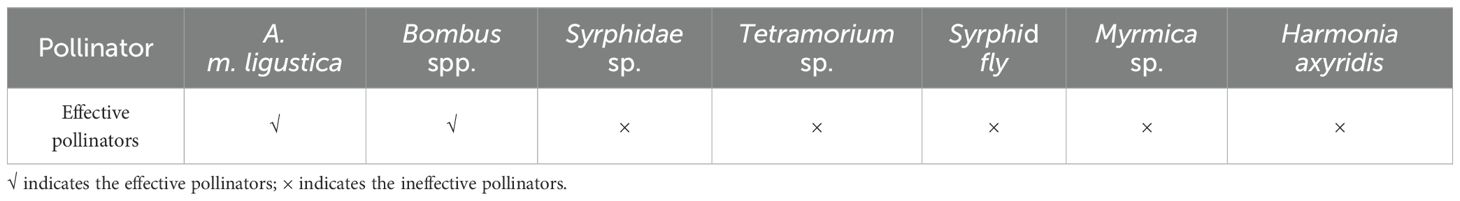
Table 2. Statistical of visiting insects of ‘Northland’.
During the 24 d of observation of visiting frequency, the highest average daily temperature period in the open ground was from 12:00 to 15:00, with an average temperature of 27.5°C and an average wind speed of 12 - 14 km/h. Under the open ground conditions, with early flowering Bombus spp. had the highest visiting frequency at 14:00-15:00 every day, with a maximum of 89 ± 3 visits; A. m. ligustica had the highest visiting frequency at 12:00 - 13:00 every day, with a maximum of 30 ± 3 visits; with peak flowering Bombus spp. had the highest visiting frequency at 14:00-15:00 every day, with a maximum of 135 ± 5 visits; A. m. ligustica had the highest visiting frequency at 13:00 - 14:00 every day, with a maximum of 46 ± 4 visits. Under greenhouse conditions, with early flowering Bombus spp. had the highest visiting frequency at 13:00-14:00 every day, with a maximum of 42 ± 3 visits; A. m. ligustica had the highest visiting frequency at 14:00 - 15:00 every day, with a maximum of 18 ± 2 visits; with peak flowering Bombus spp. visited flowers most frequently from 13:00 to 14:00, with a maximum of 60 ± 2 visits. At 14:00 - 15:00 daily, A. m. ligustica visited flowers at their peak, with a maximum of 20 ± 2 visits, and Bombus spp. visited flowers more frequently than A. m. ligustica. In the open ground and greenhouse with early flowering, the visiting frequency by Bombus spp. was highly significant (p < 0.01) at 13:00 - 14:00, 14:00 - 15:00. The visiting frequency by A. m. ligustica was highly significantly different (p < 0.01) at 12:00 - 13:00, 13:00 - 14:00, 14:00 - 15:00. In the open ground and greenhouse with peak flowering, the visiting frequency by Bombus spp. was highly significant (p < 0.01) at 12:00 - 13:00, 13:00 - 14:00, 14:00 - 15:00. The visiting frequency by A. m. ligustica was highly significantly different (p < 0.01) at 11:00 - 12:00, 12:00 - 13:00, 13:00 - 14:00 (Figure 6).
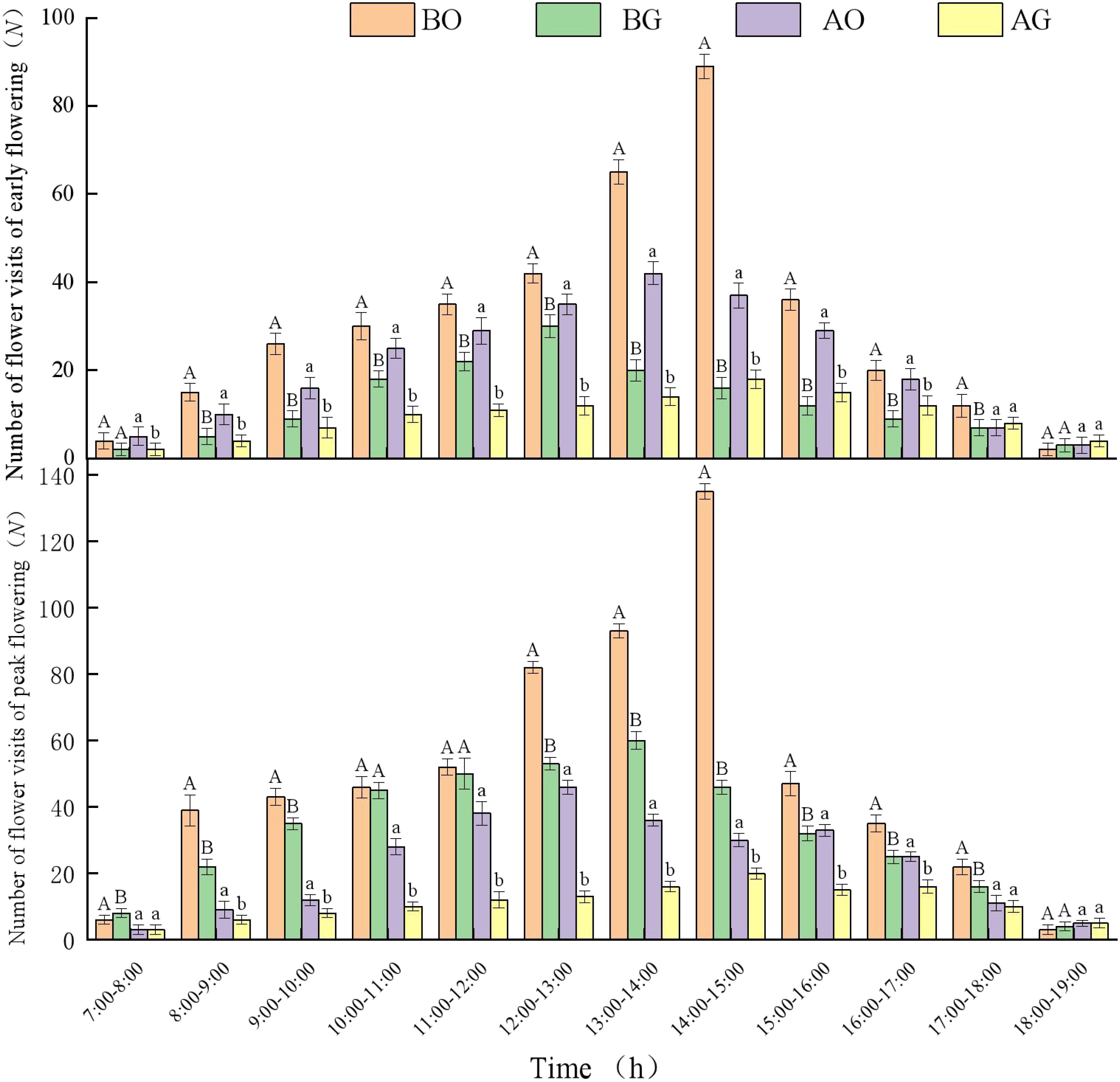
Figure 6. The daily change of the frequency of effective flower visits of ‘Northland’ visitors of early flowering and peak flowering in open ground and greenhouse. BO, Bombus spp. in open ground; BG, Bombus spp. in greenhouse; AO, A. m. ligustica in open ground; AG, A. m. ligustica in greenhouse. Different uppercase letters indicate significant differences in the frequency of visiting by Bombus spp. in different habitats at the same time period; different lowercase letters indicate significant differences in the frequency of visiting by A. m. ligustica in different habitats at the same time period (p < 0.05).
3.4.2 Pollination efficiency of pollinators
In the open ground and greenhouse, the number of pollen grains carried away by Bombus spp. after a single visit to was 2050 ± 127 and 2810 ± 136, while the number of pollen grains carried away by A. m. ligustica after a single visit to was 1680 ± 131 grains and 1951 ± 118 grains (Table 3); the number of pollen grains dropped on the stigma after a single visit to the flowers by Bombus spp. was 228 ± 33 grains and 358 ± 19 grains, and the number of pollen grains dropped on the stigma after a single visit to A. m. ligustica was 153 ± 13 grains and 196 ± 20 grains. There was a significant difference in the number of pollen grains removed by Bombus spp.after a single visit between the two habitats (p < 0.05). The pollination efficiency of Bombus spp. was 11.17 ± 0.02% and 12.76 ± 0.01% in the open ground and greenhouse; and the pollination efficiency of A. m. ligustica was 9.15 ± 0.01% and 10.08 ± 0.01%, which made Bombus spp. more efficient pollinators than A. m. ligustica, and thus Bombus spp. were the most effective pollinators.
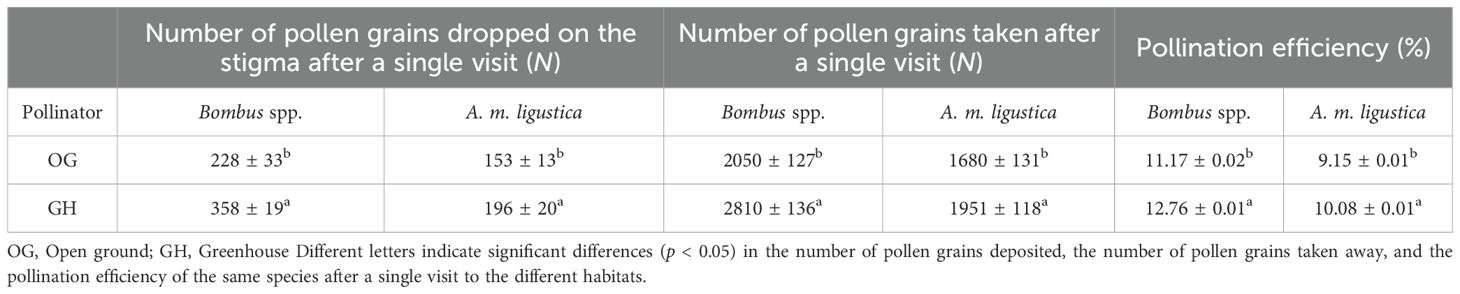
Table 3. Pollination efficiency of two pollinators in two habitats.
3.5 Correlation analysis between nectar accumulation, pollen grains number and visiting frequency
The results of the study showed that nectar accumulation showed highly significant correlation (p < 0.01) with the visiting frequency under open ground and greenhouse at the early flowering, pollen grains number and visiting frequency showed significant correlation (p < 0.05) under greenhouse; nectar accumulation showed significant correlation (p < 0.05) with the visiting frequency under open ground and greenhouse at the peak flowering, nectar accumulation and pollen grains number under open ground (p < 0.05), under the open ground and greenhouse, nectar accumulation and pollen grains number showed significant correlation (p < 0.05), under the open ground and greenhouse, nectar accumulation and pollen grains number showed significant correlation (p < 0.01), under the open ground and greenhouse, nectar accumulation and pollen grains number showed highly significant correlation(p < 0.01) (Table 4).
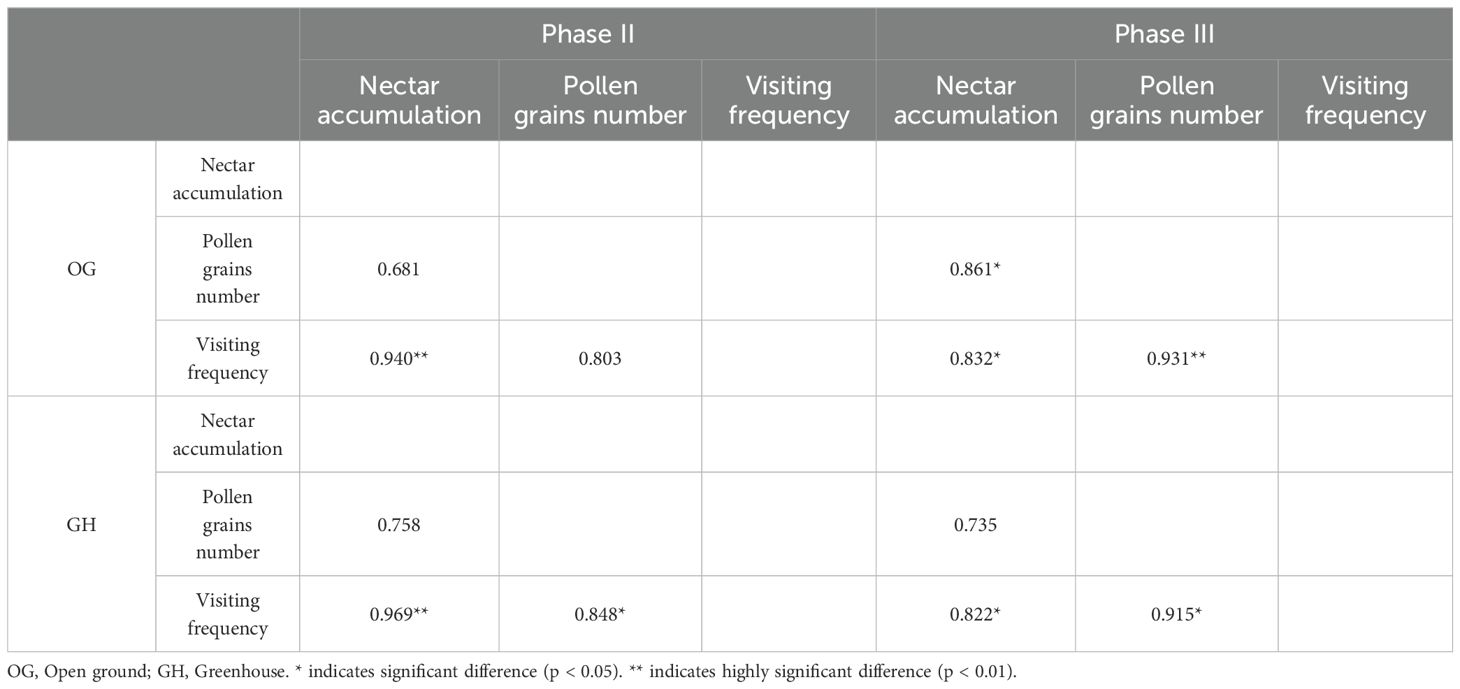
Table 4. Correlation analysis between nectar accumulation, pollen grains number and visiting frequency in two habitats.
3.6 Artificial pollination experiment
In the open ground and greenhouse, there were significant differences (p < 0.05) between the fruiting rates of natural pollination and different plants cross-pollination, significant differences (p < 0.05) between the same plant cross-pollination and different plants cross-pollination, the fruiting rates of different plants cross-pollination were significantly higher than that of the same plant cross-pollination, and there was a highly significant difference (p < 0.01) between non-emasculated bagging and different plants cross-pollination (Table 5). Therefore, the type of breeding system was the same in the open ground and greenhouse, with predominantly outcrossing pollination, partial self-pollination, and the need for pollinators.
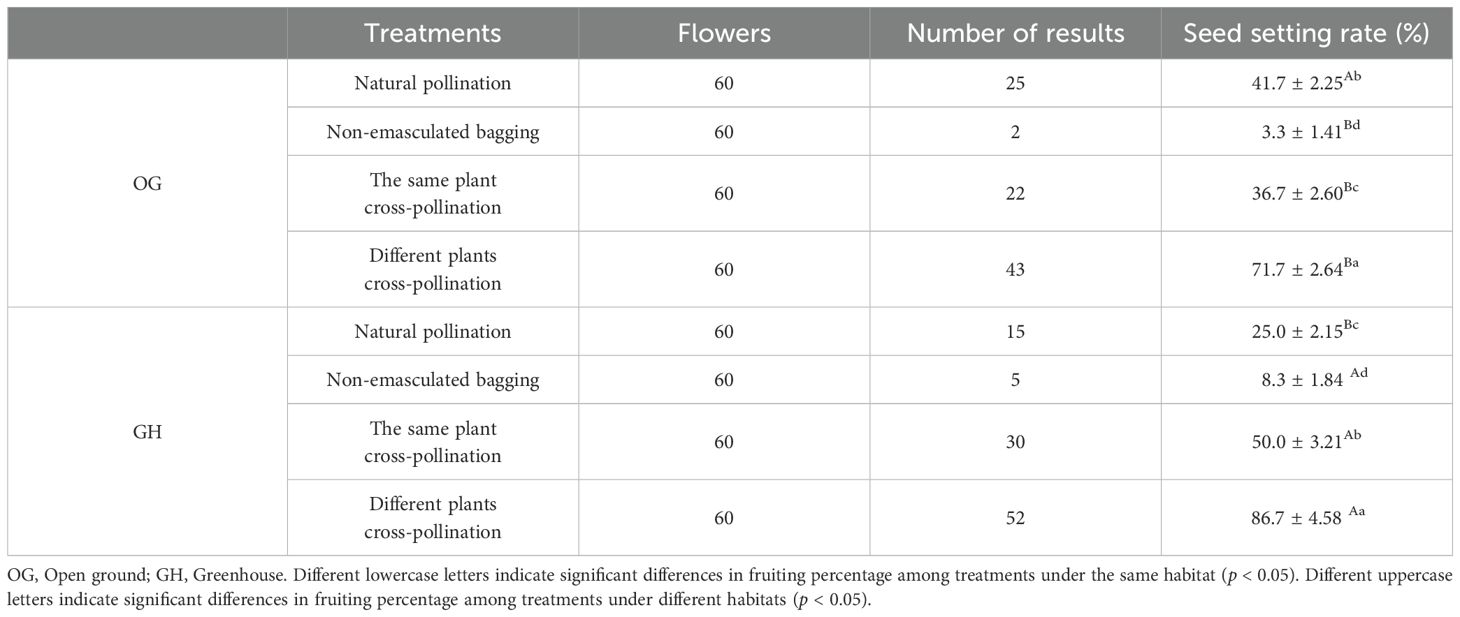
Table 5. Artificial bagging pollination results of ‘Northland’ in two habitats.
3.7 Fruit quality
Fruit size and seed thousand-grain weight at different locations on the branches within different populations showed the same pattern of middle > top > base. In the open ground and greenhouse, there were no significant differences in fruit longitudinal diameter at the top and middle of branches (p > 0.05), significant differences in fruit longitudinal diameter at the base of branches (p < 0.05), and no significant differences in fruit transverse diameter at the top, middle and base of branches (p > 0.05). There was no significant difference in the thousand seed weight of fruit seeds at the top (p > 0.05), significant differences in the thousand seed weight of fruit seeds at the middle and base of branches (p < 0.05) in the open ground and greenhouse (Table 6).
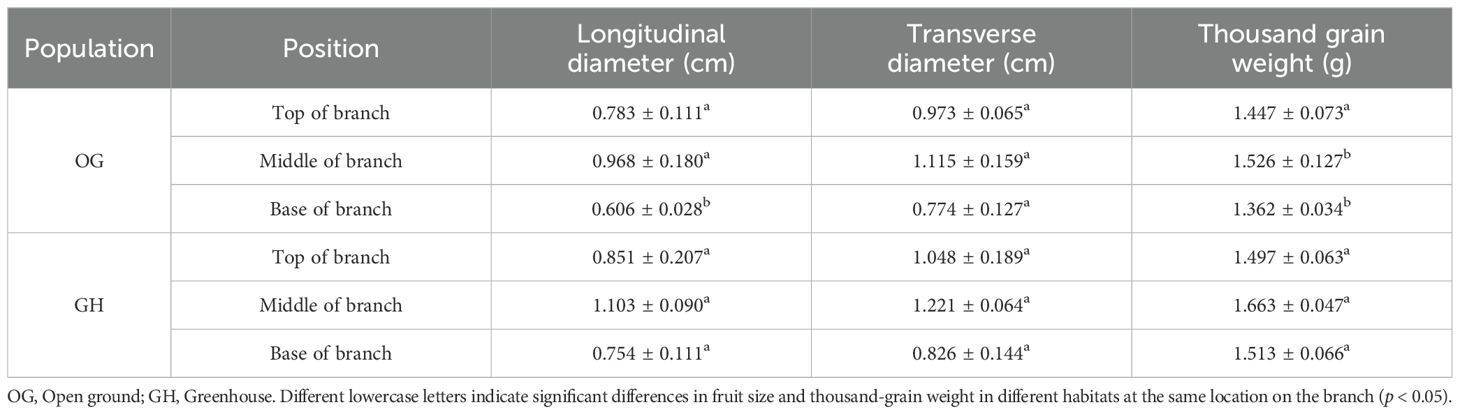
Table 6. Comparison of fruit size and thousand grain weight of seeds at different positions of branches of different populations of ‘Northland’.
The study showed that the sugar content was 9.62 ± 0.14% in the open ground and 8.75 ± 0.22% in the greenhouse, which was significantly higher than that of the greenhouse population (p < 0.05); the acid content was 2.35 ± 0.18 g/kg in the open ground and 3.11 ± 0.14 g/kg in the greenhouse, which was significantly lower than that of the greenhouse(p < 0.05); and the VC content was 2.52 ± 0.21 mg/100g and 2.37 ± 0.17 mg/100g in open ground and greenhouse with no significant difference (p > 0.05) (Table 7).
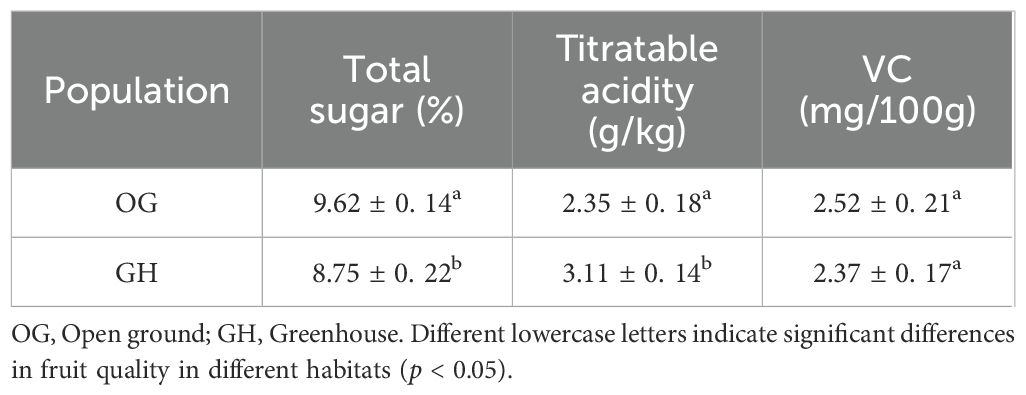
Table 7. Comparison of fruit quality in two different environments of ‘Northland’.
4 Discussion
4.1 Gradual pollen presentation in ‘Northland’
In this study, we found that in both open ground and greenhouse, the pollen dispersal of anthers in ‘Northland’ was a gradual mechanism, by contracting and squeezing the anthers, so that the pollen was released sequentially through the anther porous dehiscence and was removed by the pollinator one by one. This anther dehiscence can effectively avoid pollinators to take away too much pollen at a time, reduce pollen loss, so that pollinators can only get a small amount of pollen in a visiting, prolonging the pollen presentation time, improve pollination efficiency, and further promote the success of plant reproduction. Gradual pollen presentation is one of the typical floral characteristics of plants to improve staminate fitness. The interaction between the process of gradual pollen presentation and its pollinators enables accurate and efficient transfer of pollen to different plants stigmas, which not only improves staminate fitness, but also guarantees the success of different plants cross-pollination (Yang, 2016; De et al., 2015; He and Liu, 2023). Pollinator plant Bomareaacutifolia Snow A A et al. improves pollen transfer rate by gradual presentation of six stamens opening sequentially (Snow and Grove, 1995); in pollinator-deficient environments Gageanigra Zheng R Q et al. prolongs pollen dispersal time and reduces pollen wastage by gradual presentation (Snow, 1986; Zheng et al., 2016a). Pollen presentation is mainly divided into gradual pollen presentation and simultaneous pollen presentation (Thomson et al., 1994). In pollinator-scarce conditions, plants prefer simultaneous pollen presentation, while in pollinator-rich environments, plants strictly limit pollen output (gradual pollen presentation) so that only a small amount of pollen is available to each pollinator and more pollinators are involved in the pollination, thus increasing staminate fitness (Thomson et al., 1994, Thomson et al., 2000). Gradual pollen presentation not only improves the efficiency of pollen transfer, but also increases the plant’s ability to adapt to environmental changes to ensure the success of pistillate reproduction (Howell et al., 1993; Lu and Tan, 2007; He et al., 2024).
4.2 The effect of floral characteristics on pollination characteristics among two habitats of ‘Northland’
The floral characteristics include flower opening characteristics, pollen viability, stigma receptivity, nectar secretion, etc (Wang and Tan, 2011; Yang et al., 2019). The opening characteristics include flower structure, flower color, flower shape, etc. (Ji et al., 2023; He et al., 2010; Jing et al., 2023). Flowering patterns influence pollen dispersal patterns by attracting pollinators, which in turn affects the mating system of the plant (Tang and Han, 2007). It has been pointed out that the morphological characteristics of flower appearance of different plants are related to their reproductive and developmental characteristics as well as to the pollination of flowers (Ruan and Jiang, 2006). In this study, we showed that the single flowers of ‘Northland’ are inverted bell-shaped, which not only prevents rain from washing away the stamens and pistils and prolongs pollen viability and stigma pollination, but also reduces the negative effects of nectar dilution by rain and pollen removal by wind, and ensures the success of pollination. The flower characters of plants not only change according to the pollination status of pollinators, but are also closely related to environmental conditions (light, temperature, and humidity, etc.). Most plant populations often show more differences in floral characteristics across habitats (Ma, 2014; Tang, 2016; Huang et al., 2019). For example, the flower structures of Iris ruthenica show differences between under the forest and forest edge habitats (Wang et al., 2021). In this study, we measured each basic structure of the flower parts of ‘Northland’, and the corolla, anthers, filament length, and stigmas were longer in the greenhouse than in the open ground, but no significant differences existed, and the authors hypothesized that the greenhouse was more suitable for its growth due to its stable temperature and humidity factors.
The variability in the length of plant stigma receptivity and the duration of pollen viability, as well as the length of the overlap period, not only affects the pollination success rate of pollinators, but also influences the rate of self-crossing and outcrossing (Quan, 2011; Tang and Huang, 2012; Zheng et al., 2016b). Pollen viability is not only affected by genetic factors, but also by the external natural environment (Wei et al., 2006). Research showed that the floral characteristics of plants and their pollen viability affect the reproductive development of plants, and also indicated that the receptivity of the stigma is also strongly related to its reproductive development (Guan et al., 2009; Wan et al., 2011; Xiao, 2005). The pollen viability of ‘Northland’ differed significantly between the open ground and greenhouse, but the trend was the same, the pollen viability was highest at the early flowering and began to decline at the peak of flowering. Increasing pollen viability after flowering ensured that pollinators transferred the higher-viability pollen to the stigma after visiting to maximize the efficiency of staminate resource utilization, which was consistent with the results of the stigma-receptivity test. The stigma receptivity is strongest at the early flowering, and the stigma receptivity becomes weakened as the flowering time increases; pollen viability interacts with stigma receptivity at the early flowering. At the same time, ‘Northland’ secretes a large amount of nectar when it blooms, and when insects contact the stigma, pollination is completed and the chances of successful fertilization are guaranteed, which ensures that pollen adheres to the villi of pollinators at the peak of their viability, and promotes the pollination success rate of ‘Northland’.
4.3 Adaptive significance of different inter-environmental breeding systems in ‘Northland’
Breeding systems are an expression of the interactions between the genetic mechanisms within the plant and the external environment, and their combination of pollinators and pollination behavior is a major factor in influencing the genetic composition and fitness of reproductive offspring, and plays an important role in determining the evolutionary pathways and characterizing variation in plants (He et al., 2023; Gran, 1881). The study of plant breeding systems helps to understand the responses of plants to environmental changes and is a necessary prerequisite for plant breeding research (Murúa et al., 2014; Wang et al., 2010). The visiting frequency of ‘Northland’ in the two habitats was correlated with the ambient temperature of the time period of the day, which was consistent with the results of the study on Iris ruthenica Wang Z Z et al. (Wang et al., 2021). The study showed in the open ground and greenhouse, the frequency of visiting increased as the temperature increased, and was highest when the daily temperature reached the highest. As the temperature decreased, the frequency of visiting also decreased, and the frequency of visiting by pollinators showed a tendency of increasing first to reach the highest and then decreasing gradually.
Plant breeding systems are an important part of plant life history and an important factor in plant evolution (Chen et al., 2023). The breeding system of plants is influenced by their own genetic factors and the external natural environment (He and Liu, 2003). Sexually reproducing plant breeding systems are categorized into self-crossing, outcrossing and various concurrent mating systems (Zhang et al., 2024). In nature, outcrossing is favorable to increase the genetic diversity of plants (Wang et al., 2024). According to the results of artificial pollination experiments, the type of breeding system of ‘Northland’ in the two habitats was partially self-crossing, outcrossing-dominated and required pollinators. Since the results of the different plants cross-pollination treatment were higher than the other treatment groups, the natural pollination and the same plant cross-pollination fruiting rates were lower than those different plants cross-pollination. Under natural conditions, the setting rate of open ground is higher than that of greenhouse, and under artificial pollination, the setting rate of greenhouse is higher than that of open ground, indicating that artificial pollination can be carried out in greenhouse to improve the setting rate.
5 Conclusions
The results of the study show that pollen dispersal of ‘Northland’ anthers is gradual pollen presentation mechanism, the anther dehiscence by pore cleavage, which increase pollination frequency, improves the pollination efficiency and promotes outcrossing. The setting rate of ‘Northland’ is higher than open ground by artificial pollination in greenhouse. Effective pollinators are artificially cultivated in greenhouses to increase fruiting rates and promote reproductive success. Therefore, the greenhouse growing environment is more suitable for ‘Northland’ than the open ground. This study could provide a basis for efficient reproduction of ‘Northland’ in the future, which is important for the reproduction of its species.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
KS: Conceptualization, Data curation, Formal Analysis, Software, Writing – original draft. YZ: Data curation, Investigation, Methodology, Writing – review & editing. CZ: Formal Analysis, Validation, Visualization, Writing – review & editing. SY: Funding acquisition, Project administration, Resources, Supervision, Writing – review & editing. HJ: Validation, Visualization, Writing – review & editing. RC: Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the “Science and Technology Development Plan Project of Science and Technology Department of Jilin Province (YDZJ202201ZYTS468)”, the “Science Research Project of the Jilin Provincial Education Department (JJKH20230664KJ)”, the “Changda Researcher Climing Plan of Changchun University (ZKP202031)”.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Arroyo J., Dafni A. (1995). Variations in habitat season flower traits and pollinators in dimorphic NarcissustazettaL.(Amaryllidaceae) inIsrael. New Phytol. 129, 135–145. doi: 10.1111/j.1469-8137.1995.tb03017.x
Barretts S. C., Harder L. D. (1996). Ecology and evolution of plantmating. Trends Ecol. Evol. 11, 73–79. doi: 10.1016/0169-5347(96)81046-9
Cao J. K., Jiang W. B., Zhao Y. M. (2007). Experiment Guidance of Postharvest Physiology and Biochemistry of Fruits and Vegetables, 56–128.
Cao M. H., Zhao Q. Y., Wei C. M. (2022). Flowering traits and breeding system of three species of Bromeliads. Plant Sci. 40, 291–301.
Caster O. S., Silveir A. P., Navarr O. L. (2008). How flower bi-ology and breeding system affect the reproductive success of the narrow endemic Polygala vayredae Costa(Polygalaceae). Bot. J. Linn. Soc 157, 67–81. doi: 10.1111/j.1095-8339.2008.00784.x
Chai Y. X., Cai M. Y., Jin X. L. (2017). A preliminary study on the pollination biology of purple-flowered hilaria. Guangxi Plant 37, 1322–1329. doi: 10.11931/guihaia.gxzw201702027
Chen X. B. (2020). Research on nitrogen fertiliser distribution technology for raspberry greenhouse cultivation. J. Shenyang Agric.l Univ., 1–57. doi: 10.27327/d.cnki.gshnu.2020.000985
Chen Y. J., Wang M. L., Tang H. (2023). Flowering phenology and breeding system of the endemic plant in Guangxi. Seed. 42, 21–27. doi: 10.16590/j.cnki.1001-4705.2023.02.021
Chen L. Y., Xiao X., Xiao J. (2019). Flower and reproductive characteristics of different large-flowered taxa of Epimedium. J. Plant Res. 39, 808–816. doi: 10.7525/j.issn.1673-5102.2019.06.002
Conner J. K., Davis R., Rush S. (1995). The effect of wild radish floral morphology on pollination efficiency by four taxa of pollinators. Oecologia 104, 234–245. doi: 10.1007/BF00328588
Dafni A., Kevan P. G., Husband B. C. (2005). Practical Pollination Biology (Cambridge: Enviroquest Ltd.).
De Y., Wu J. M., Mu H. B. (2015). Studies on the mating system of three species of forage grasses of the genus Phyllostachys (Elymus). J. Plant Genet. Resour. 16, 903–906. doi: 10.7525/j.issn.1673-5102.2019.06.002
Dong M. Y., Wu M., Tong G. J., Wang J. X., Chen X. N., Zhao X. Y., et al. (2022). Studies on flowering characteristics and breeding system of Hesperis oreophila. Yuanyi Xuebao (Acta Hortic. Sinica) 50, 607–619.
Fabbro T., Körner C. (2004). Altitudinal differences in flower trait sand reproductiv eallocation. Flora. 199, 70–81. doi: 10.1078/0367-2530-00128
Gargan O. D., Peruzz I. L. (2021). Comparing flower biology in five species of Gagea (Liliaceae) from Southern Italy. Flora Mediterranea. 31, 131–144. doi: ;10.7320/FlMedit31SI.131
Guan W. L., Li Y. F., Chen X. (2009). Iris flower structure and flowering pollination biology. Acta Hortic. Sin. 4, 1485–1490.
Guan C., Sun Y., Yang X. J. (2024). Study on the blooming and breeding characteristics of eggplant. Mol. Plant Breed., 1–16. Available online at: https://kns.cnki.net/kcms/detail/46.1068.S.20220818.1548.004.html.
Halibunuer, Reyilamu M., Aishajiang A. (2022). Studies on pollen presentation pattern and mating system in Ningxia wolfberry. Nord. J. Bot. 42, 1042–1050. doi: 10.17521/cjpe.2021.0463
Han Y., Dai C., Yang C. F. (2008). Anther appendages of Incarvillea trigger a pollen-dispensing mechanism. Ann. Bot. 102 (3), 473–479. doi: 10.1093/aob/mcn102
He Y. P., Liu J. Q. (2003). Recent progress and review of plant breeding system research. J. Plant Ecol. 27, 151–163. doi: 10.17521/cjpe.2003.0024
He Y. P., Liu J. Q. (2023). Areviewon recent advance sin the studies of plant breeding system. Acta Phytoecol. Sin. 27 (2), 151–163. doi: 10.17521/cjpe.2003.0024
He R., Yan F., Zhu R. Y. (2024). Research on the flowering and breeding system of Tangut Ruixiang. Acta Agrestia Sin. 32, 188–197. doi: 10.11733/j.issn.1007-0435.2024.01.020
He L., Yu H., Fan Y. H. (2010). Advances in plant reproductive biology. J. @ Mountain Agric. Biol. 29, 456–460.
He M., Zhang Z. F., Zang S. T. (2023). Flowering characteristics and breeding system of Adonis calendula. J. Northeast For. Univ. 51 (03), 54–59. doi: 10.13759/j.cnki.dlxb.2023.03.011
Homson J. D., Rigney L. P., Karoly K. M. (1994). Pollen viability, vigor, and competitive nessin Erythronium grandiflorum (Liliaceae). Am. J. Bot. 81, 1257–1266. doi: 10.1002/j.1537-2197.1994.tb11447.x
Howell G. J., Slater A. T., Knox R. B. (1993). Secondary pollen presentation in angiosperms and its biological significance. Aust. J. Bot. 41, 417. doi: 10.1071/BT9930417
Huang M. (2019). Preliminary study on flowering phenology and reproductive characteristics of Acacia nigra. Fujian Agric. For. Univ., 1–58.
Huang S. Q., Guo Y. H. (2000). Progress of pollination biology. Sci. Bull. 45, 225–237. doi: 10.3321/j.issn:0023-074X.2000.03.001
Ji S., Yang L., Yan S. J., Ye J. W., Liao J. W. (2023). Progress in the study of interactions between plant floral characteristics and pollinators. World Forestry Res. 36 (02), 26–31. doi: 10.13348/j.cnki.sjlyyj.2023.0009.y
Jing Y. B., Xiao M. Z., Yi K. G. (2023). Floral characteristics and breeding system of Hemerocallis minor. J. China Agric. Univ. 28, 146–152.
Lei X., Qu W. L., Sha Y. (2019). Preliminary study on comprehensive characteristics of flower parts and breeding system of tamarind. China Trop. Agriculture. 4, 64–68. doi: 10.3969/j.issn.1673-0658.2019.04.015
Li J. X., Hu Z. H. (2013). Research progress of blueberry anthocyanins. J. Nucl. Agric. Sci. 27, 817–822. doi: 10.11869/hnxb.2013.06.0817
Li Q., Ma H. Z., Jinaitihan M. (2024). Study on flower syndrome and breeding system of the spring flowering plant Iris membranaceus. Guangxi Plant 44 (04), 756–765. Available online at: https://link.cnki.net/urlid/46.1068.S.20240202.1609.006.
Liao R., Ling X. Y., Wu Y. H., et al. (2024). Studies on floral characters and butterfly wing pollination in Lycoris aurea. Guangxi Plant 10, 1–11. doi: 10.11931/guihaia.gxzw202308058
Lu T., Tan D. Y. (2007). Evolutionary significance of pollen presentation time series in animal pollinated plants. Biodiversity. 15, 673–679. doi: 10.3321/j.issn:1005-0094.2007.06.015
Ma J. Q. (2014). Studies on the flowering biology of five species of lilies of the genus Lilium, , Master’s thesis. Jilin Agric. Univ, Changchun of Jilin province, 38–40.
Masayyuki N. (2013). New development and challenges in plant factory. J. Jpn. Oil Chem. Soc 13, 267–273. doi: 10.5650/oleoscience.13.267
Miao L. (2018). Effects of light and pollination environment on growth and reproductive characteristics of Sagittaria trifolia L. Hubei Univ., 1–38.
Murúa M., Cistern A. J., Rosend E. B. (2014). Pollination ecology and breeding system of two Calceolaria species in Chile. Rev. Chil Hist Nat. 87 (1), 7. doi: 10.1186/0717-6317-87-7
Pan C. C., Liu L. D., Zhao H. L., Hou Y. L., Zhang L., Wang L. J. (2012). Pollination biology of Elaeagnus angustifolia in middle reaches of the Heihe River in Northwest China, Zhongguo Shamo. J. Desert Res. 32, 780–783.
Ruan C. J., Jiang G. B. (2006). Adaptive significance of herkogamy and flower behavior. J. Plant Ecol. 30, 210–220. doi: 10.17521/cjpe.2006.0030
Snow A. A. (1986). Pollination dynamics in Epilobiumcanum (On-(agraceae): Consequences for game tophytic selection. Am. J. Bot. 73, 139–151. doi: 10.1002/j.1537-2197.1986.tb09689.x
Snow A. A., Grove K. (1995). Protandryaneuter phaseuni sexual umbelsinaherma phroditicneo tropical vine(Bo- (mareaacutifolia, Alstroemeriaceae). Am. J. Bot. 82, 741–744. doi: 10.1002/j.1537-2197.1995.tb15685.x
Tang X. (2016). Breeding system of the desert relict plant Naked Fruitwood and its pollination characteristics. Xinjiang Agric. Univ., 1–45. doi: CNKI:CDMD:2.1016.239731
Tang L. L., Han B. (2007). Effects of flowering pattern on pollinator behavior and pollen dispersal. Biodiversity. 15, 680–686. doi: 10.3321/j.issn:1005-0094.2007.06.016
Tang X. X., Huang S. Q. (2012). Research progress of flower color diversity and variation. Plant Divers. 34, 239–247. doi: 10.3724/SP.J.1143.2012.12030
Thomson J. D., Rigney L. P., Karoly K. M. (1994). Pollen viability, vigor, and competitive ability in erythronium grandiflorum (Liliaceae). Am. J. Bot. 81, 1257–1266. doi: 10.1002/j.1537-2197.1994.tb11447.x
Thomson J. D., Wilson P., Valenzuela M. (2000). Pollen presentation and pollination syndromes,with special reference to Penstemon. Plant Spec. Biol. 15, 11–29. doi: 10.1046/j.1442-1984.2000.00026.x
Wan G. J., Yang Z. L., Yan G. X. (2011). Advances in the studies of endangered plants' breeding system. J. Northwest A F Univ. 39, 207–213.
Wang Z. Z. (2021). Reproductive characteristics of purple-bracted iris in forest edge and understory habitats. Xinjiang Agric. Univ., 1–39. doi: 10.27431/d.cnki.gxnyu.2021.000269
Wang Z. Z., Amanula Y., Yu Q. H. (2021). Flowering and reproductive characteristics of the purple-bracted iris. J. Xinjiang Agric. Univ. 44, 347–354. doi: 10.3969/j.issn.0517-6611.2022.12.015
Wang Y. J., Fang X., Zhong Z. C. (2010). Effects of different habitats on floral and fruit characteristics of iris. J. Ecol. 8, 25–38. doi: CNKI:SUN:STXB.0.2010-17-012
Wang N., Peng T., Pei Y. (2024). Floral characteristics and breeding system of Gegu radish. Mol. Plant Breed. 10, 1–15. Available online at: https://link.cnki.net/urlid/46.1068.S.20240202.1609.006.
Wang L., Tan D. Y. (2011). Flower syndrome and secondary pollen presentation in Codonopsis clematidea. Biodiversity. 19, 24–33. doi: 10.3724/SP.J.1003.2011.10167
Wei J. H., Huang L. Q., Chen S. L., Cheng H. Z., Chu Q. L. (2006). Study on stigma, pollen viability and self-compatibility of Platycodon grandiflorum. China J. Chin. Mater. Med. 31 (5), 366–368.
Xiang G. J. (2019). Evolution of flower characters and pollination biology of Lonicera japonica in the Qinling region. Wuhan Univ., 1–59. doi: 10.27379/d.cnki.gwhdu.2019.000098
Xiao Y. A. (2005). Reproductive Ecology and Photosynthetic Adaptation of the Endangered Long-stalked Dicotyledonous Wood (Chongqing City, China: Southwest Normal Univ).
Xie M., Ma Y. P., Cao Y. R., Liu D. T., Li Z. H., Ma H. (2023). Studies on the floral characteristics and breeding system of a very small population of wild plants of Rhododendron boswellii. J. Yunnan Agric. Univ. (Nat. Sci.). 38 (01), 95–103. doi: 10.12101/j.issn.1004-390X(n).202207033
Yang S. L. (2016). An investigation on the reproductive ecological characteristics of horned fennel. Shihezi Univ., 1–36. doi: CNKI:CDMD:2.1016.779117
Yang S. L., Chu G. M., Shi X., Wang S. M. (2019). Elaborated pollen packaging and dispensing mechanism induced by petal architecture from a Papaveraceae species. PeerJ. 7, e7066. doi: 10.7717/peerj.7066
Yin D. S., Wei X. H., Shen H. L. (2016). Research on the comprehensive characteristics of flower parts and breeding system of bellflower fruit. J. Beijing For. Univ. 38, 67–73. doi: 10.13332/j.1000-1522.20150340
Yu Y. L., Liu Y. W., Yang S. L. (2022). Flowering and reproductive characteristics of Bluecrop. J. Anhui Agric. Sci. 50, 57–59+99. doi: 10.3969/j.issn.0517-6611.2022.12.015
Zhang H. F., Xiao X. F., Mai J. X., Han Z. Z., Ma Q., Huang Y. Q., et al. (2024). Analysis of flower characters, flowering phenology and breeding mode of Plumbago officinalis. Mol. Plant Breed. 10, 1–13. Available online at: https://kns-cnki-net.webvpn.ccu.edu.cn/kcms/detail/46.1068.s.20230901.1853.020.html.
Zheng R. Q., Hou Z. Z., Zhang A. Q. (2016a). Pollen of the short-lived early spring plant Black Scale Top Ice flower Presentation timing and its adaptation. J. Bot. 51, 594–600. doi: 10.11983/CBB15194
Keywords: Vaccinium corymbosum Berkeley, gradual pollen presentation, floral syndrome, breeding system, outcrossing
Citation: Sun KX, Zhang YJ, Zhou CZ, Yang SL, Jiang HX and Cui RR (2024) Effect of gradual pollen presentation on pollination efficiency and reproduction of Vaccinium corymbosum Berkeley in two habitats. Front. Ecol. Evol. 12:1476848. doi: 10.3389/fevo.2024.1476848
Received: 06 August 2024; Accepted: 25 September 2024;
Published: 14 October 2024.
Edited by:
Massimo Nepi, University of Siena, ItalyReviewed by:
Rosa Ranalli, University of Milano-Bicocca, ItalyAlexsandro Bezerra-Silva, State University of Feira de Santana, Brazil
Copyright © 2024 Sun, Zhang, Zhou, Yang, Jiang and Cui. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shan Lin Yang, eWFuZ3NsQGNjdS5lZHUuY24=