
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 04 September 2024
Sec. Biogeography and Macroecology
Volume 12 - 2024 | https://doi.org/10.3389/fevo.2024.1455329
This article is part of the Research TopicBiodiversity of Antarctic and Subantarctic EcosystemsView all 10 articles
 Quentin Jossart1,2*†
Quentin Jossart1,2*† Yann Lelièvre1†
Yann Lelièvre1† Andreas Kelch3,4
Andreas Kelch3,4 Blanca Figuerola5†Camille V. E. Moreau2†Davide Di Franco3†
Blanca Figuerola5†Camille V. E. Moreau2†Davide Di Franco3† Jamie Maxwell6†Marie L. Verheye7,8†
Jamie Maxwell6†Marie L. Verheye7,8† Melanie Mackenzie9†
Melanie Mackenzie9† Rachel Downey10†
Rachel Downey10† Sebastián Rosenfeld11,12†Stéphane Hourdez13†
Sebastián Rosenfeld11,12†Stéphane Hourdez13† Thomas Saucède1†
Thomas Saucède1†Sub-Antarctic islands are expected to show a high degree of endemicity due to their remoteness. However, biogeographic affinities in the sub-Antarctic remain poorly understood, especially in the marine realm. Sub-Antarctic islands being at the crossroads between Antarctic and cold temperate regions, biodiversity characterization and biogeographic analyses are a priority for monitoring and rapidly assessing variations associated with environmental changes. One underexplored sub-Antarctic area is Crozet, a protected archipelago located halfway between Antarctica and South Africa. In this study, we investigated the shallow-water Crozet macrofaunal diversity, distribution patterns and biogeographic affinities based on the examination of fieldwork specimens via a thorough morphological identification and a genetic characterisation. The resulting dataset provides an important baseline for further studies and conservation strategies, compiling the first genetic and taxonomic database for the Crozet archipelago. In total, 100 morphotypes were found, belonging to nine different phyla, among which arthropods (32), molluscs (18) and echinoderms (17) were the richest. Forty-seven morphotypes were identified to the species level, among which 20 were reported in Crozet for the first time. This confirms that Crozet is a poorly known region, even compared to other sub-Antarctic areas. A large proportion of species (62%) had circum Southern Ocean or circum sub-Antarctic distributions. These species were mostly shared with Kerguelen (72%), the Magellan Province (64%), and Prince Edward Islands (64%), confirming the patterns found in macroalgae and specific macrofaunal groups. However, this large-distribution statement needs to be counterbalanced by the detection (genetic data) of more restricted distributions than expected in four study cases (the tanaid Apseudes spectabilis, the nudibranch Doris kerguelenensis, the polychaete Neanthes kerguelensis and the chiton Hemiarthrum setulosum). Considering that most morphotypes had no genetic data available from other regions, the proportion of morphotypes with restricted distribution is likely to increase alongside future investigations. In addition, we also found a few cases of unrecognized diversity that might lead to the descriptions of new species, some likely to be endemic to Crozet (e.g., within the polychaete genus Harmothoe and the bryozoan genus Antarctothoa). Altogether, this stresses the need to maintain conservation efforts in Crozet and pursue integrative investigations in order to highlight and protect its unusual diversity.
In the Southern Ocean, the sub-Antarctic region is located between the subtropical and the Antarctic polar marine fronts, comprising a series of remote archipelagos and small islands (De Broyer et al., 2014). Due to the geographic remoteness of sub-Antarctic islands, a high degree of endemicity can be expected in both terrestrial and marine taxa (Frenot et al., 2001). Several studies from the last decades have highlighted contrasting patterns of distribution in various taxonomic groups, from single island endemicity to species exhibiting broad distributions (e.g., Poulin et al., 2014; Figuerola et al., 2017, Figuerola et al., 2018; Féral et al., 2021; Vantomme et al., 2023). Overall, biodiversity and biogeographic affinities of the different sub-Antarctic areas remain poorly understood, with a contrast of knowledge existing among sub-Antarctic islands (Griffiths and Waller, 2016; Féral et al., 2021). It is thus particularly important to better characterize their biodiversity and biogeographical affinities to improve our understanding of marine life dynamics, especially considering their key location at the crossroads between high Antarctic and cold temperate areas (Hemmings and Stephens, 2010; De Broyer et al., 2014). It is also central to conservation considerations, given that high endemicity can mean increased sensitivity to global change and risk of biodiversity loss (Griffiths, 2010; Park et al., 2014; Jossart et al., 2019; Féral et al., 2021). In particular, an improved biogeographic baseline would thus detect distribution shifts that are expected in response to current and future environmental changes (Saucède et al., 2017; Guillaumot et al., 2018). In response to these conservation concerns, some sub-Antarctic islands have already been given the status of Marine Protected Areas (Crozet MPA, Heard Island and McDonald Islands MPA, Kerguelen MPA, Prince Edward Islands MPA; Brooks et al., 2020) or have received a significant extension of their marine parks (e.g., Macquarie Island Marine Park in 2023, parksaustralia.gov.au).
Crozet’s benthic marine biodiversity is notably underexplored. This five-island archipelago is located in the Indian Ocean sector of the Southern Ocean (45°48’S–46°26’S; 50°14’E–52°15’E) and is about 2,300 km away from both Antarctica and South Africa. Crozet originated from three main cycles of volcanism, among which the first one initiated nine million years ago (Chevallier and Nougier, 1981). The archipelago is strongly influenced by the Antarctic Circumpolar Current (ACC), which is expected to increase its connectivity with other sub-Antarctic areas (Pollard et al., 2007; De Broyer et al., 2014). Previous terrestrial studies have been undertaken on Crozet terrestrial biodiversity (e.g., Frenot et al., 2001; Convey, 2007; Hullé and Vernon, 2021), however, marine benthic ecosystems have received far less attention (Canteras and Arnaud, 1985; Sicinski and Gillet, 2002; Griffiths and Waller, 2016; Lelièvre et al., 2023). The waters around Crozet are part of the National Nature Reserve of the French Southern Territories MPA and recognized as a UNESCO World Heritage site (whc.unesco.org/en/list/1603). Marine coastal areas have also additionally received an IUCN “enhanced protection” status (Féral et al., 2021). In this context of conservation initiatives, the French polar institute project #1044 Proteker was implemented to monitor nearshore marine benthic communities of the French Southern Territories (Kerguelen, Saint-Paul & Amsterdam, and Crozet). The main goal of this ongoing project is to establish a baseline for monitoring biodiversity dynamics and assess the impact of climate change on coastal marine ecosystems (Féral et al., 2016). Since its initiation, samples collected from Crozet during the Proteker campaigns have been invaluable for the investigation of biogeographic patterns in selected taxa (e.g., González-Wevar et al., 2021; Rosenfeld et al., 2023). At the community level, using underwater video-imagery, Lelièvre et al. (2023, 2024) investigated the taxonomic and functional diversities of shallow benthic communities associated with hard substrates. The authors highlighted high taxonomic diversity and low functional richness, evenness, and redundancy. This suggests a potential vulnerability to current and future environmental changes, which, again, stresses the need for further investigations based on additional biological samples (Mason et al., 2005; Lelièvre et al., 2023).
In our study, we investigated the diversity and biogeography of macrofauna (>1 mm) at two sites (Baie du Marin and Crique du Sphinx) around Possession island (“Ile de la Possession”, 46°25’S; 51°45’E), the largest island of the Crozet archipelago. Based on the examination of specimens collected in a recent fieldwork campaign, we combined a thorough morphological identification by taxonomic experts with a genetic characterisation based on the COI barcode region. The joint use of morphological and genetic data has been demonstrated to be effective in species identification of various taxonomic groups (e.g., Gostel and Kress, 2022; Jossart et al., 2023). In addition, comparing recorded biogeographic patterns (relying on morphology-based taxonomy) with genetic data was also important in the revision and clarification of species distributions (Christiansen et al., 2018; Jossart et al., 2019; Vantomme et al., 2023). In Lelièvre et al. (2023), Crozet faunal communities exhibited a high-level diversity, notably in echinoderms. Regarding its geographic situation and oceanic features (strong influence by the ACC), we expect benthic marine communities of Crozet to be composed of both endemic and circum sub-Antarctic species, with high-level faunal affinities to both Kerguelen and Prince Edward Islands. In fact, while remoteness can explain endemicity after rare colonisation and settlement events, a certain homogenisation can also be expected due to the role of the ACC in larval or even adult (kelp rafting) transport (Moore et al., 2018; Fraser et al., 2022). By compiling the first genetic and taxonomic database including specimen images and occurrence records for Crozet, this study’s newly generated dataset provides an important identification tool and baseline for further studies and conservation purposes.
Specimen sampling occurred during November 2021 as part of the Proteker 9 campaign onboard the R/V Marion Dufresne II. Two geographically close sites (< 2 km), Baie du Marin and Crique du Sphinx, were investigated on the east coast of Possession Island (“Ile de la Possession”, Crozet archipelago). This investigation was part of an impact study on benthic communities after the recent installation of the International Monitoring System (IMS) hydroacoustic station HA04 in the area (Lelièvre et al., 2023). Biological samples were opportunistically collected by hand during six dives at depths ranging from 4 to 20 m. Specimens were then preserved in 96% ethanol for subsequent analyses.
Each specimen was examined under a stereomicroscope (Leica LK300 LED), assigned to a morphotype (or putative species) and a preliminary identification was obtained using available taxonomic resources (e.g., Féral et al., 2019; Lelièvre et al., 2023). Each morphotype was also photographed using the same stereomicroscope and an Olympus OMD-EM1 camera with a 60 mm macro lens. Preliminary identifications of morphotypes were then confirmed or refined by taxonomic experts (Supplementary Material 1). Macro photographs were sent to taxonomists as well as voucher specimens when required in the identification process.
The barcode region of the cytochrome c oxidase subunit I (COI; 658 base pairs) was obtained for each morphotype (one to twenty individuals sequenced per morphotype). DNA extractions were undertaken on a small piece of tissue, following the salting-out protocol of Sunnucks and Hales (1996). For COI amplification, we used a PCR mix of 12.5 µL Accustart Toughmix (QuantaBio), 10 µL ultrapure water, 0.5 µL each primer (10 µM) and 1.5 µL DNA extract. PCR conditions consisted of 40 cycles for the following temperature steps: 30 s at 94°C (denaturation), 30 s at 45–49°C (annealing, see details below) and 45 s at 72°C (elongation). These cycles were preceded by 3 min at 94°C and followed by 3 min at 72°C. Amplifications occurred using either universal or taxon-specific primers. Bryozoans, chordates, cnidarians, nemerteans and sponges were amplified using the universal primer pair “F-LCO1490 + R-HCO2198” at an annealing temperature of 45°C (Folmer et al., 1994). Amphipods and pycnogonids were amplified with the same primers at an annealing temperature of 49°C. Isopods and tanaids were amplified using the primer pair “F-CrustCOIF + R-HCO2198” at an annealing temperature of 47°C (Teske et al., 2006). Echinoderms were amplified using the primer pair “F-LCOech1aF1+ R-HCO2198” at an annealing temperature of 45°C (Folmer et al., 1994; Layton et al., 2016). Annelids were amplified using either the primer pair “F-LCO1490+ R-HCO2198” and the primer pair “F-polyHCO + R-polyLCO” at an annealing temperature of 47°C (Carr et al., 2011). Molluscs were amplified using either the primer pair “F-LCO1490 + R-HCO2198” (annealing temperature: 45°C) or the primer pair “F-COI-mol & R-COI-mol” (annealing temperature: 49°C; Owada et al., 2013). The sequences of each of the aforementioned primer were F-LCO1490: GGTCAACAAATCATAAAGATATTGG; R-HCO2198: TAAACTTCAGGGTGACCAAAAAATCA; F-CrustCOIF: TCAACAAATCAYAAAGAYATTGG; F-LCOech1aF1: TTTTTTCTACTAAACACAAGGATATTGG; F-polyHCO: TAMACTTCWGGGTGACCAAARAATCA; R-polyLCO: GAYTATWTTCAACAAATCATAAAGATATTGG; F-COI-mol: TCWACAAATCAYAAAGATATTGG; R-COI-mol: ACYTCMGGRTGMCCAAAAAATCA. The purification step and Sanger sequencing were carried out at the sequencing centre of Azenta/Genewiz Germany. Sequences were edited and checked for stop codons in Geneious Prime 2023.2.1 (Kearse et al., 2012). Barcodes were then aligned (Muscle algorithm), compared among them within classes/orders (e.g., amphipods) and to the ones available on GenBank and Barcode of Life (BOLD) databases (Ratnasingham and Hebert, 2007). Combining morphological and genetic data, a final identification was then determined at the most accurate taxonomic level possible. In addition to the COI sequences aforementioned, additional genetic barcodes from non-Crozet areas were obtained from either scientific publications or public BOLD records. Combining all the barcodes for each taxon, alignments were then realized in Geneious Prime 2023.2.1. Templeton Crandall Singh (TCS) haplotype networks were then constructed in PopART 1.7 (Leigh and Bryant, 2015) and mean pairwise genetic distances (uncorrected p-distances) were calculated between clades in the software MEGA X (Kumar et al., 2018). We then used a genetic distance threshold of 2% (among clades of the haplotype network) to consider that an initial morphotype represents more than one putative species. This threshold is based on the most common interspecific genetic distances observed in the groups of interest [crustaceans: Vieira et al., 2016, molluscs: Layton et al., 2014, polychaetes: Carr et al., 2011, sea stars: Ward et al., 2008 and Moreau et al., 2021)]. Species delimitation methods, such as ASAP and ABGD, were not used considering their low suitability for datasets with limited number of sequences (Puillandre et al., 2012, Puillandre et al., 2021) A public Barcode of Life (BOLD) project (HAOIV: Shallow benthic communities of Crozet archipelago) was created on the platform boldsystems.org. Macro pictures were uploaded for each specimen as well as COI barcodes, when successfully obtained (Table 1).
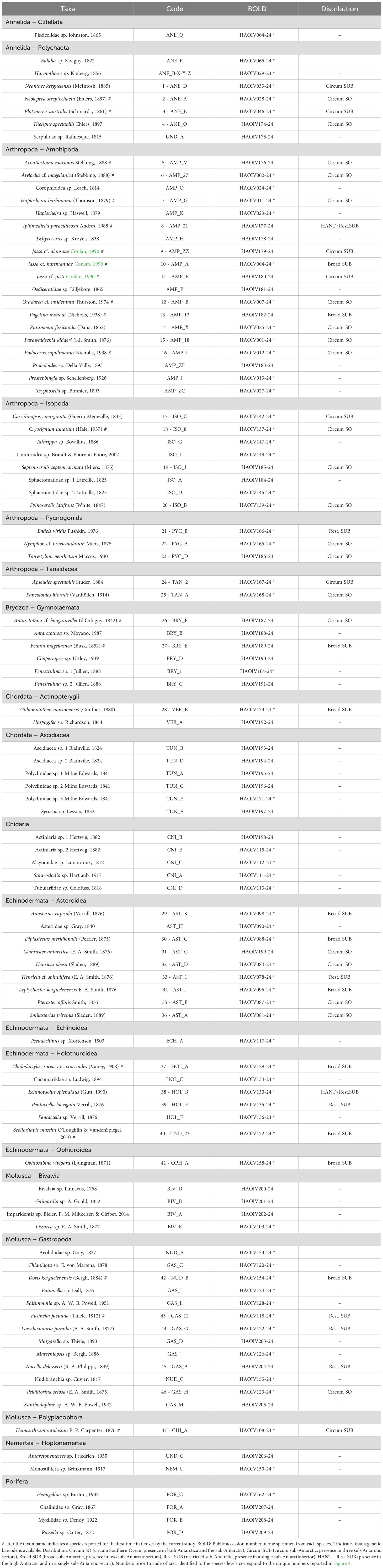
Table 1. List of the 100 taxa found in the studied area, ranked by alphabetical order of phyla (then by class or order).
The biogeographic distribution of taxa identified up to the species level was analysed based on the following bioregionalization of the Southern Ocean: Magellan Province, South Georgia/South Sandwich Islands, Bouvet (= Atlantic Sector of the Southern Ocean); Prince Edward Islands (Prince Edward and Marion islands), Crozet, Kerguelen, Heard Island (= Indian Ocean Sector of the Southern Ocean); Macquarie/sub-Antarctic New Zealand region (= Pacific sector of the Southern Ocean); West Antarctica; East Antarctica). Species occurrence data were either based on: 1) occurrences mentioned in scientific publications; 2) GBIF occurrences (Global Biodiversity Information Facility; https://www.gbif.org); 3) additional unpublished occurrences obtained from direct contacts with taxonomic experts. If a contradiction arose, occurrences obtained from direct contacts with taxonomists or dedicated publications had precedence over GBIF records. COI barcodes were also used to refine the species distribution of four species (e.g., the expected presence of Hemiarthrum setulosum P. P. Carpenter, 1876 in Antarctica was not considered as it appeared that Antarctic specimens were highly divergent from the ones of Crozet, see results). Four different distribution patterns were then considered: 1) restricted sub-Antarctic distribution (i.e., recorded in a single sector of the sub-Antarctic); 2) broad sub-Antarctic distribution (i.e., recorded in two sectors of the sub-Antarctic); 3) circum sub-Antarctic (i.e., recorded in three sector of the sub-Antarctic); 4) circum Southern Ocean (recorded both in the Antarctic and in either two or three sub-Antarctic sectors). Based on identified species occurrence data, a similarity network was produced using Gephi 0.10.1 (Bastian et al., 2009). In the network, nodes (circles; n = 56) represent both geographic regions and species that are connected by edges (lines; n = 241). The graph type was undirected, and no edge merging strategy was used. The layout algorithm “ForceAtlas2” (Jacomy et al., 2014) was then used to spatialize the network. The following input settings were used: tolerance: 0.05, approximate repulsion: checked, approximation: 1.2, scaling: 50, stronger gravity: checked, gravity: 0.01. Considering the low number of occurrences at Bouvet, this bioregion was not taken into account in the similarity network analysis.
In total, 1050 specimens representing 100 morphotypes were collected in the study area (Table 1), and 173 individual genetic barcodes were obtained from 67 distinct morphotypes, indicating a barcoding success rate of 67%. The 100 morphotypes belonged to nine different phyla, among which Arthropoda (32 morphotypes) was the richest, followed by Mollusca (18), Echinodermata (17), Annelida (8), Chordata (8), Bryozoa (6), Cnidaria (5), Porifera (4) and Nemertea (2) (Figure 1). In Arthropoda, the diversity was split between amphipods (19 morphotypes), isopods (8 morphotypes), pycnogonids (3 morphotypes) and tanaids (2 morphotypes) (Figure 1). The phylum Mollusca was composed of gastropods (13 morphotypes), bivalves (4 morphotypes) and chitons (1 morphotype) (Figure 1). Within the Echinodermata, asteroids were the most speciose class (9 morphotypes), followed by holothuroids (6 morphotypes), echinoids (1 morphotype), and ophiuroids (1 morphotype) (Figure 1). From the 100 different morphotypes, 47 were identified to the species level (Table 1) among which 20 species were reported at Crozet for the first time (Table 1): three holothuroid species (Cladodactyla crocea var. croceoides (Vaney, 1908), Echinopsolus splendidus (Gutt, 1990), Scoliorhapis massini O’Loughlin & VandenSpiegel, 2010); two polychaete species (Neoleprea streptochaeta (Ehlers, 1897), Platynereis australis (Schmarda, 1861)); two bryozoan species (Antarctothoa cf. bougainvillei (d’Orbigny, 1842), Beania magellanica (Busk, 1852)); two gastropod species (Doris kerguelenensis (Bergh, 1884); Fusinella jucunda (Thiele, 1912)); one chiton species (Hemiarthrum setulosum); ten amphipod species (Acontiostoma marionis Stebbing, 1888, Atyloella cf. magellanica (Stebbing, 1888), Haplocheira barbimana (Thomson, 1879), Iphimediella paracuticoxa Andres, 1988, Jassa cf. alonsoae Conlan, 1990, Jassa cf. hartmannae Conlan, 1990, Jassa cf. justi Conlan, 1990, Oradarea cf. unidentata Thurston, 1974, Pagetina monodi (Nicholls, 1938), Podocerus capillimanus Nicholls, 1938); and one isopod species (Cryosignum lunatum (Hale, 1937)).
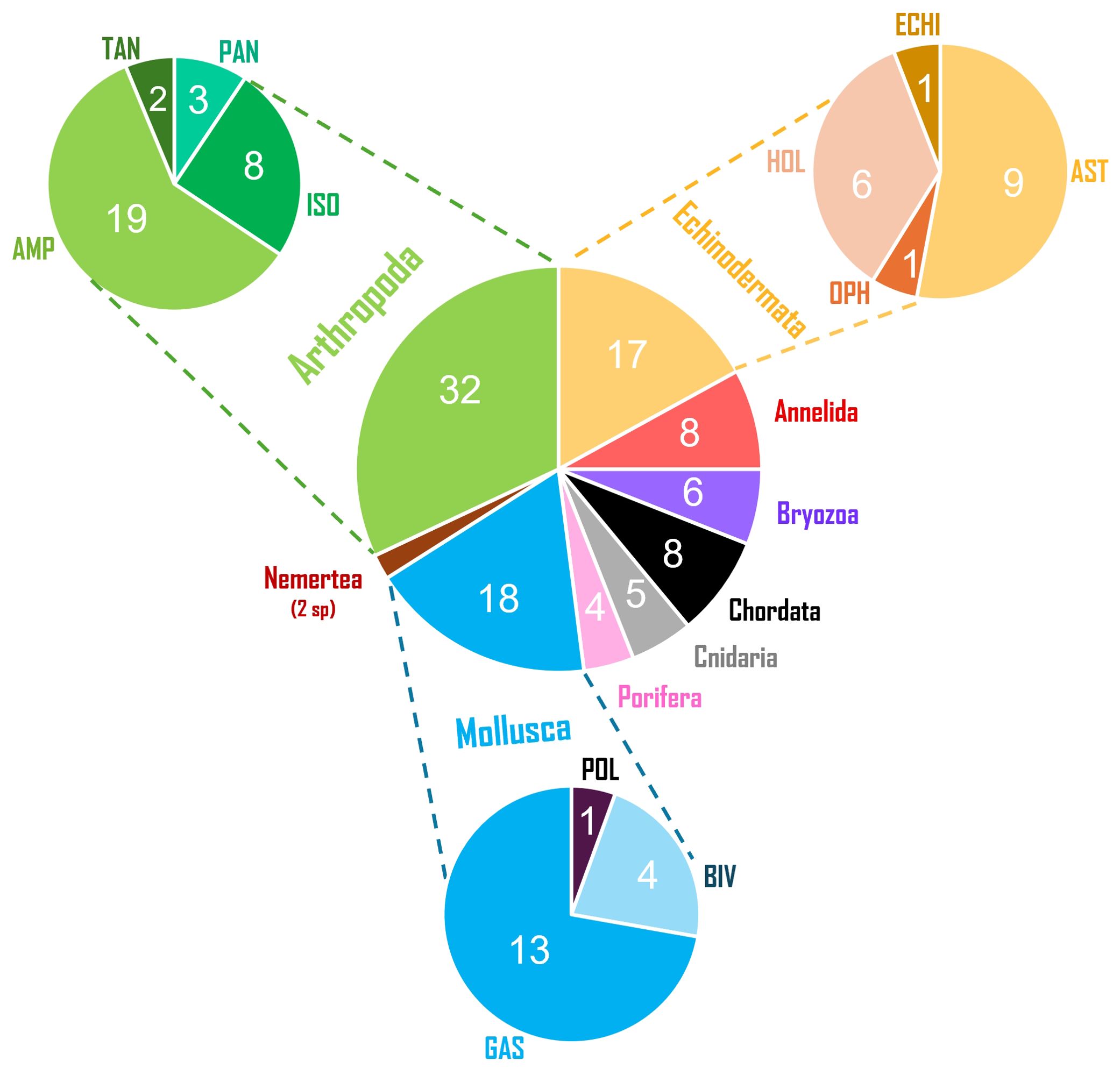
Figure 1. Overall taxonomic diversity for the 100 morphotypes found in the current Crozet investigation. Centre chart depicts phyla, top-left chart depicts arthropod orders (AMP, Amphipoda; ISO, Isopoda; TAN, Tanaidacea; PAN, Pantopoda), top-right chart depicts echinoderm classes (AST, Asteroidea; ECHI, Echinoidea; HOL, Holothuroidea; OPH, Ophiuroidea) and bottom chart depicts mollusc classes (BIV, Bivalvia; GAS, Gastropoda; POL, Polyplacophora). Numbers in each slice indicate the number of morphotypes for the specific group.
Out of the 47 taxa identified to the species level, 35 were successfully barcoded in our study. Previous reference databases with COI sequence data were available for only 14 species (Miya et al., 2016; Moreau et al., 2021; Sands et al., 2021). The four following phylogeographic patterns were observed (Figure 2): (1) a circum Southern Ocean distribution such as in the sea stars Pteraster affinis Smith, 1876 and Smilasterias triremis (Sladen, 1889), and the pycnogonid Nymphon brevicaudatum Miers, 1875; (2) a circum sub-Antarctic distribution such as in the amphipod Parawaldeckia kidderi (S.I. Smith, 1876) and the brittle star Ophiosabine vivipara (Ljungman, 1871); (3) a broad sub-Antarctic distribution such as for the fish species Gobionotothen marionensis (Günther, 1880), and the sea stars Anasterias rupicola (Verrill, 1876) and Diplasterias meridionalis (Perrier, 1875); and (4) a restricted sub-Antarctic distribution such as in Henricia cf. spinulifera (E. A. Smith, 1876).
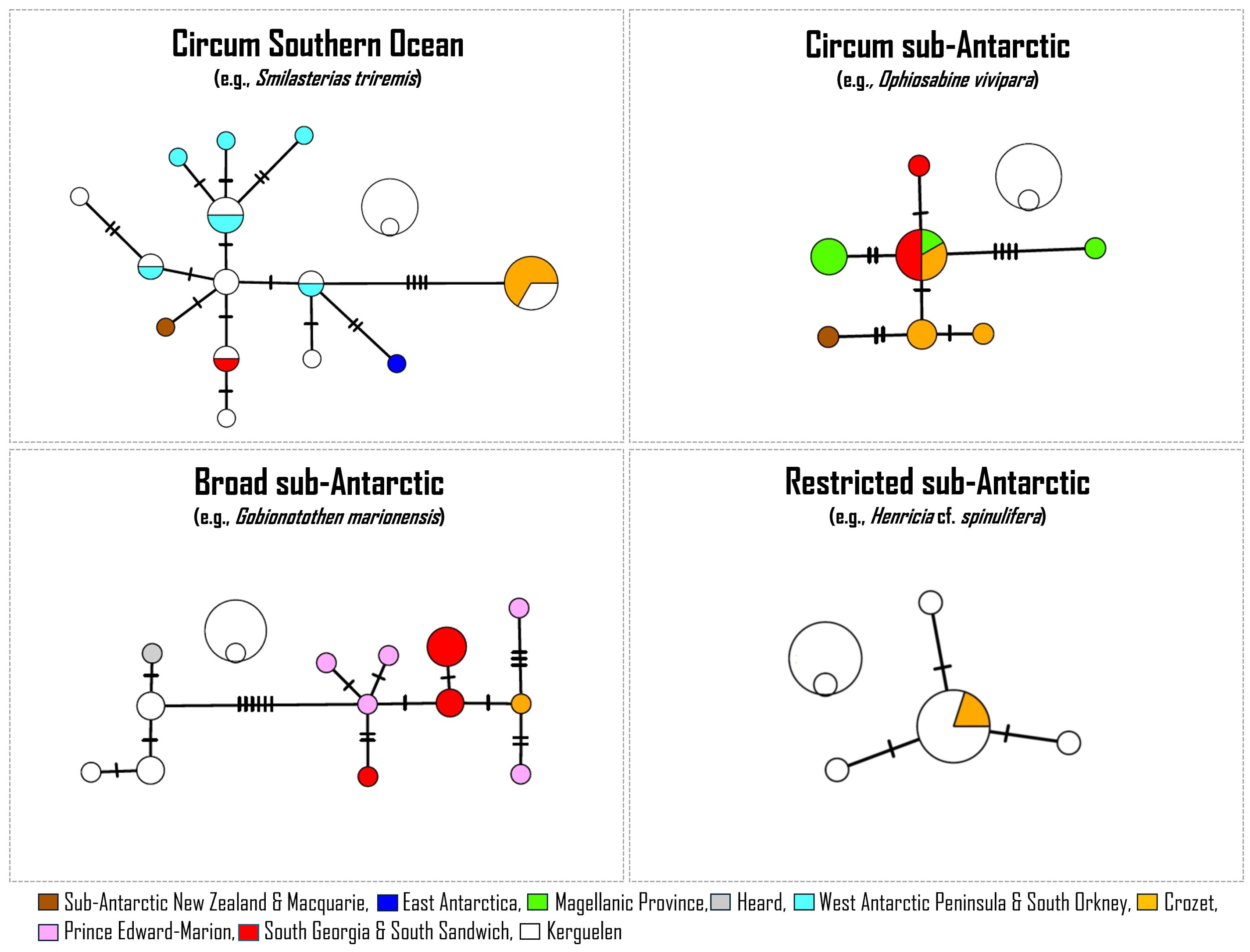
Figure 2. Haplotype networks illustrating the most common biogeographic and phylogeographic patterns. Sizes of imbricated circles close to each network denote the number of specimens exhibiting the haplotype (outer circle: 10 specimens, inner circle: 1 specimen). All the Crozet sequences are from the current study. Non-Crozet sequences are from Miya et al., 2016 (Gobionotothen marionensis), Moreau et al., 2021 (Henricia cf. spinulifera, Smilasterias triremis) and Sands et al., 2021 (Ophiosabine vivipara).
Based on COI data, some unrecognized species diversity was found in six taxa (genetic distance among clades > 2%, cf. Materials and Methods; Figure 3; Supplementary Material 2). In the chiton Hemiarthrum setulosum P. P. Carpenter, 1876, specimens showed an important genetic distance from sequences known from the Antarctic Peninsula (11.19%). Analysed specimens of the nudibranch Doris kerguelenensis are found within a clade with specimens from South Georgia (genetic distance of 2.82% with the closest related clade; Figure 3). In the tanaid Apseudes spectabilis Studer, 1884, the single Crozet specimen showed a p-distance of 2.32% with a specimen from Kerguelen. For the polychaete Neanthes kerguelensis (McIntosh, 1885), Crozet specimens clustered with specimens from South Georgia and Kerguelen and are well-differentiated from two distinct Antarctic groups (3.93% et 4.82%, respectively; Figure 3). In addition, potential species complexes were found in the polychaete Harmothoe Kinberg, 1856 and the pycnogonid Nymphon brevicaudatum (Supplementary Material 2). Finally, while the level of genetic divergence was lower than the defined threshold (1.65%), the Crozet specimens of the sea star Leptychaster kerguelenensis interestingly clustered into a clade sharing no haplotype with other sub-Antarctic and Antarctic specimens.
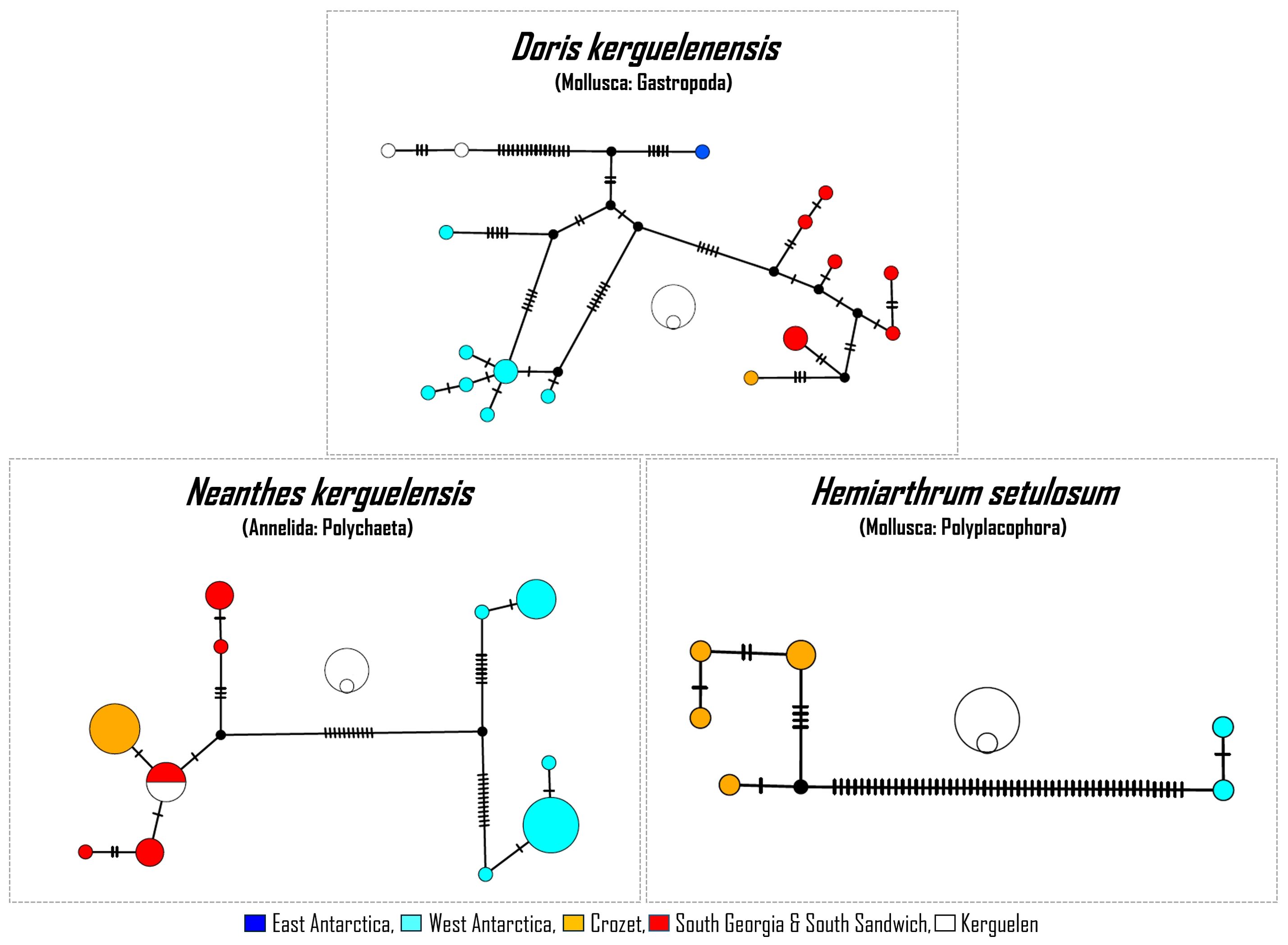
Figure 3. Haplotype networks illustrating three cases of unrecognized species diversity. Sizes of imbricated circles close to each network denote the number of specimens exhibiting the haplotype (outer circle: 10 specimens, inner circle: 1 specimen). All the Crozet sequences are from the current study. Non-Crozet sequences are from Maroni et al., 2022 (Doris kerguelenensis), Leiva et al. (2022) (Neanthes kerguelensis), Irisarri et al. (2014) and Jossart et al. (2023) (Hemiarthrum setulosum).
After updating distribution information for four species based on available genetic barcodes (cf. results above), we found that nearly half of the 47 morphotypes identified to the species level were reported to have a circum Southern Ocean distribution (47%), 15% to have a circum sub-Antarctic distribution, 21% to have a broad sub-Antarctic distribution, and 13% to have a distribution restricted to one sector of the Southern Ocean (Table 1; Supplementary Material 3). Two species (the holothurian Echinopsolus splendidus (Gutt, 1990) and the amphipod Iphimediella paracuticoxa Andres, 1988) also showed a peculiar distribution across the polar front, being present in the high Antarctic and a single sub-Antarctic sector (4%). As highlighted in the similarity network (Figure 4), 34 of these 47 taxa were also found to occur in Kerguelen, 30 in the Magellan Province, 30 in Prince Edward (Marion) Islands, 25 in Macquarie/sub-Antarctic New Zealand region, 22 in South Georgia/South Sandwich Islands, 23 in West Antarctica, 16 in Heard Island, and 13 in East Antarctica. The faunal similarity with Kerguelen was notably marked in most echinoderms (10/13), molluscs (4/6), arthropods (14/21) and in all annelids (4/4) (Supplementary Material 3). This faunal similarity pattern was also observed when comparing with Prince Edwards Islands and the Magellan province (Figure 4; Supplementary Material 3). The similarity of Crozet with Macquarie and sub-Antarctic New Zealand was associated to different shared taxa: it was very high in amphipods (11/12) and in arthropods in general (16/21), but far less marked in echinoderms (5/13) and molluscs (1/6) (Supplementary Material 3).
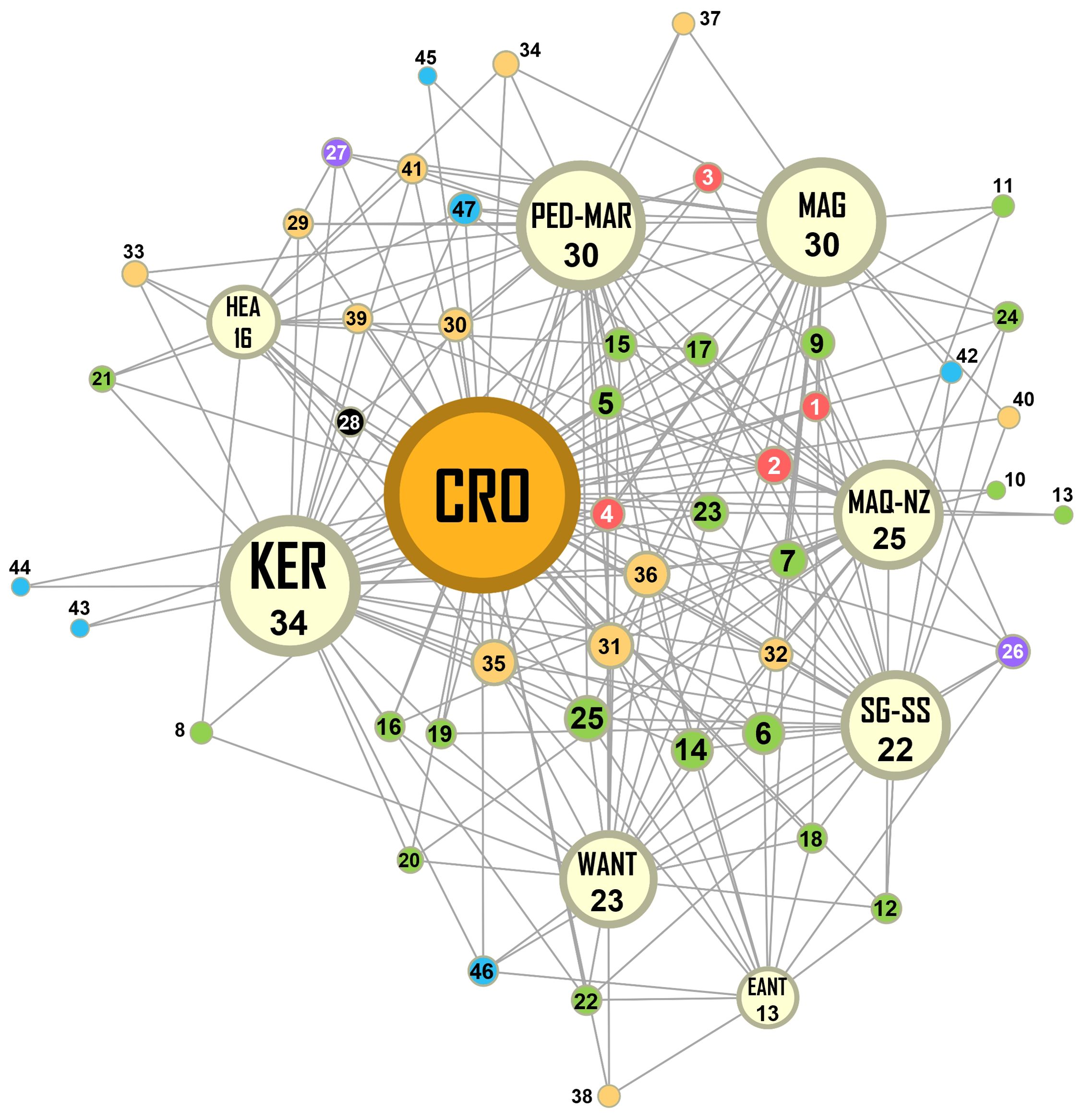
Figure 4. Similarity network produced with Gephi 0.10.1. Bioregions (except Crozet in orange) are represented by pale-yellow circles (CRO, Crozet; EANT, East Antarctica; HEA, Heard Island; KER, Kerguelen; MAG, Magellan Province; MAQ-NZ, Macquarie/sub-Antarctic New Zealand region; PED-MAR, Prince Edward (Marion) Islands; SG-SG, South Georgia/South Sandwich Islands; WANT, West Antarctica). The size of each circle is proportional to the total number of species shared by this bioregion with Crozet (indicated under the bioregion acronym). The smaller coloured circles indicate the 47 taxa identified up to the species level: their colours denoting their phyla (black: Chordata; blue: Mollusca; green: Arthropoda; purple: Bryozoa; red: Annelida; yellow: Echinodermata), their sizes denoting the number of bioregions where the species is found; their unique numbers indicating the species referenced in Table 1 (e.g., “45” refers to the gastropod Nacella delesserti (R. A. Philippi, 1849) that occurs in two bioregions).
The current study has highlighted the presence of one hundred macrofaunal species in the shallow waters of Crozet. Among these 100 species, we reported 20 species for the first time in Crozet. In addition, 17 taxa identified to the family level (2), genus level (5) or species level (10) were already inventoried by Lelièvre et al. (2023), based on imagery transects of the same area. This included two annelids: Serpulidae Rafinesque, 1815 and Thelepus spectabilis Ehlers, 1897; the isopod Spinoserolis latifrons (White, 1847); six sea stars: Anasterias rupicola, Diplasterias meridionalis, Glabraster antarctica (E. A. Smith, 1876), Henricia sp. Gray, 1840, Leptychaster kerguelenensis E. A. Smith, 1876 and Smilasterias triremis; the sea urchin Pseudechinus sp. Mortensen, 1903, two holothuroids: Echinopsolus splendidus (Gutt, 1990) and Pentactella sp. Verrill, 1876; the brittle star Ophiosabine vivipara; the bivalve Gaimardia sp. Gould, 1852; the nudibranch Aeolidiidae sp. Gray, 1827; the gastropods Margarella sp. Thiele, 1893 and Nacella delesserti; and the sponge Hemigellius sp. Burton, 1932. The 100 taxa belonged to nine different phyla, with 75% belonging to either arthropods (32 species), molluscs (18), echinoderms (17) or annelids (8; Figure 1). At lower taxonomic levels, amphipods and gastropods were particularly represented (31% of the species; Figure 1). The high-level species richness found in echinoderms is similar to the value previously reported by Lelièvre et al. (2023) (17 versus 14 species). However, our results contrast with the former study regarding the diversity of amphipods (19 versus 0) and gastropods (13 versus 3 species). Such contrast among sampling and imagery, are related to the higher detection of smaller specimens in physical inventory, which is not surprising and already highlighted in other studies (e.g., Hanafi-Portier et al., 2021).
Interestingly, several taxa found during our investigation might be species that are new to science. First, some specimens could not be assigned to known species by taxonomic experts based on morphology. This was notably the case in amphipods (Prostebbingia sp. Schellenberg, 1926), bryozoans (Antarctothoa sp. Moyano, 1987, Fenestrulina sp. Jullien, 1888) and echinoids (Pseudechinus sp). Second, the genetic analysis revealed an unexpected diversity in some taxa. This included species that exhibited more restricted distribution than initially expected (see details below) as well as taxa for which we found more putative species than formerly inventoried in Crozet. For instance, three species of the polychaete genus Harmothoe were previously reported around Crozet (Harmothoe crosetensis (McIntosh, 1885), Harmothoe magellanica (McIntosh, 1885), and Harmothoe spinosa Kinberg, 1856). However, our genetic analyses indicated that none of the studied specimens belong to either Harmothoe crosetensis or Harmothoe magellanica (based on specimens from Cowart et al., 2022) alongside a pattern of species complex (Supplementary Material 2). The exact number of species within this complex remains to be determined, but it is likely that more species than initially expected occur in Crozet. Altogether, our results have significant taxonomic implications for our knowledge of the diversity of various taxa, and definitively call for additional investigations, which are out of the scope of the current study.
Among the 47 taxa identified to species level, we found that nearly half of the species (47%) were reported to have a circum Southern Ocean distribution. The rest had a circum sub-Antarctic (15%), broad sub-Antarctic (21%), restricted sub-Antarctic (13%) distributions or a peculiar distribution across the polar front (4%, see results). These 47 species were mostly found in three other bioregions: Kerguelen (72%), the Magellan Province (64%), and Prince Edward Islands (64%), and notably associated with high-level sharing of echinoderms (9 to 10 species shared). Such biogeographic affinity of Crozet with other sub-Antarctic areas has been highlighted from historical and recent studies (e.g., Hedgepth, 1969; Griffiths et al., 2009). More specifically, the high affinity with Kerguelen was recently highlighted for marine macroalgae (Féral et al., 2021) while not found for deeper water polychaetes (Sicinski and Gillet, 2002). The high-level similarity with Prince Edward Islands was already found in sea stars at both species and genus levels (Moreau et al., 2017). Interestingly, some species are also reported a few latitudinal degrees above the subtropical front, and therefore outside the Southern Ocean (Supplementary Material 3). For example, several species occur in Tristan da Cunha archipelago (37°S; e.g., the pycnogonid Tanystylum neorhetum Marcus, 1940; the amphipod Parawaldeckia kidderi) or around Australia and New Zealand (e.g., the polychaete Platynereis australis, the amphipod Jassa cf. justi) (Conlan, 1990; Hutchings and Reid, 1991). Some other species are also expected to occur in both hemispheres such as the sea star Pteraster affinis (confirmed by genetic data) or the tanaid Apseudes spectabilis (Larsen and Shimomura, 2006; Jossart et al., 2021). Overall, the investigated species therefore include a high proportion of widely distributed species, for which the known biogeographic distribution was confirmed by our genetic results for eight species in the current study (the sea stars A. rupicola, D. meridionalis, P. affinis and S. triremis, the pycnogonid N. brevicaudatum, the amphipod P. kidderi, the brittle star O. vivipara, and the fish G. marionensis). In contrast, it is very likely that more species with restricted distribution will be found as additional specimens and genetic data are obtained. This situation is illustrated in the present study by four study cases. The tanaid Apseudes spectabilis, the nudibranch Doris kerguelenensis, the polychaete Neanthes kerguelensis, and the chiton Hemiarthrum setulosum showed restricted distributions. The tanaid A. spectabilis, while expected to have a circum sub-Antarctic distribution (Schmidt and Brandt, 2001), also showed a significant genetic divergence (2.5%) between specimens from Crozet and those from Kerguelen. The nudibranch D. kerguelenensis was shown to be composed of 60 putative species exhibiting contrasting distribution ranges (Maroni et al., 2022; Maroni and Wilson, 2022). The Crozet specimen belonged to a clade that might be distributed in two sub-Antarctic sectors (Atlantic and Indian oceans; Figure 3). The Crozet specimen of polychaete N. kerguelensis belongs to a sub-Antarctic clade and our results are in line with Leiva et al. (2022), highlighting an important divergence from an Antarctic clade (Figure 3). Finally, the chiton H. setulosum, the single species of the genus Hemiarthrum, was expected to have a circum Southern Ocean distribution (type locality: Kerguelen). Present genetic results indicate a high genetic distance between the specimens from Crozet and those from Antarctica (11.19%; Irisarri et al., 2014; Jossart et al., 2023; Figure 3). From now, it would be relevant to further explore whether this species is largely distributed in the sub-Antarctic, notably in the Macquarie-New Zealand region where another Hemiarthrum species was previously recognized (H. hamiltonorum Iredale & Hull, 1932; Sirenko, 2006).
Our study shows the importance of integrative inventories to fill the gap of diversity knowledge existing in remote ecosystems that are facing important environmental changes. We found one hundred taxa, amongst which 20 were reported for the first time in the Crozet archipelago. This emphasizes the fact that shallow coastal waters around Crozet are an under-investigated area, even compared to most other sub-Antarctic areas. Regarding biogeographic patterns, a greater proportion of broadly distributed species was found but this statement needs to be counterbalanced by the detection (based on genetic data) of more restricted distributions than expected for several species. It is likely that future sampling will increase the proportion of species with restricted distribution. We also found a few cases of unrecognized diversity that might lead to the future descriptions of new species, some likely to have a restricted distribution or even be endemic to Crozet. Combined with the high diversity observed, this stresses the need to protect the biologically unusual Crozet archipelago. Additional sampling and barcoding efforts are necessary to better unravel marine faunal diversity and affinities with other sub-Antarctic islands and the overall Southern Ocean in the future. While this was not possible in the current study (due to the limited data available), bathymetry should notably be taken into account in such further investigations. In fact, for several taxonomic groups, species initially reported to occur over broad depth ranges has been showed to rather be distinct species with more restricted depth ranges (Barnes and Kuklinski, 2010; Neal et al., 2018; Moreau et al., 2019). Finally, another perspective includes the sharing of our data for conservation purposes. While the publicly available data will benefit to large scale projects related to conservation (genetic or occurrence data), they will also be directly shared with nature reserve managers of the French Southern Territories. In addition, the creation of a field guide of the most common species investigated is an ongoing work that would help in the monitoring of this area in the future.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
The animal study was approved by TAAF-UB convention #2258 – French Polar Institute project #1044 Proteker – Project #2021-0882 “Nearshore Cable Inspection and Environmental Survey at IMS Hydroacoustic Station HA04 Crozet, France”. The study was conducted in accordance with the local legislation and institutional requirements.
QJ: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Writing – original draft, Writing – review & editing. YL: Conceptualization, Data curation, Formal analysis, Investigation, Writing – original draft, Writing – review & editing. AK: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. BF: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. CM: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. DD: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. JM: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. MV: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. MM: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. RD: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. SR: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. SH: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. TS: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by TAAF-UB convention #2258 and European Union HORIZON-MSCA-2021 program under the Marie Skłodowska-Curie postdoctoral grant No. 101066993 (BioRISC project). This is a contribution to the French Polar Institute project #1044 Proteker and to the project #2021-0882 “Nearshore Cable Inspection and Environmental Survey at IMS Hydroacoustic Station HA04 Crozet, France”. BF has received funding by a Ramon y Cajal grant (RYC2022-036268-I) funded by MICIU/AEI/10.13039/501100011033 and FSE+. With the institutional support of the ‘Severo Ochoa Centre of Excellence’ accreditation (CEX2019-000928-S). JM is funded by the Irish Research Council (GOIPG/2019/4020). SR would like to thank the INACH project DG_10-22, the ANID– Millenium Science Intitative program – ICN2021_002 and the project from the Cape Horn International Center (CHIC) Project ANID/BASAL FB210018.
We would like to thank Adrien Force, Emilie Steimetz, Irina Bundeleva, Jérôme Thomas and Maria Teixeira for their help and advice. We would like to thank Magdalena Blazewicz, Nicolas Lavesque and Marcos Tatián for their help in the identification process. We also thank the captains and crew of the R/V Marion Dufresne II who contributed to the material collected as well as the staff and divers of the French Polar Institute and TAAF (S. Motreuil, C. Marschal, L. Le Gall, M. Gueíneí, L. Wauters, Y. Sabatheí, S. Seímelin, and M.-F. Bernard).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2024.1455329/full#supplementary-material
Barnes D. K., Kuklinski P. (2010). Bryozoans of the Weddell Sea continental shelf, slope and abyss: did marine life colonize the Antarctic shelf from deep water, outlying islands or in situ refugia following glaciations? J. Biogeography 37, 1648–1656. doi: 10.1111/j.1365-2699.2010.02320.x
Bastian M., Heymann S., Jacomy M. (2009). “Gephi: an open source software for exploring and manipulating networks,” in International AAAI Conference on Weblogs and Social Media, Vol. 3. doi: 10.1609/icwsm.v3i1.13937
Brooks C. M., Chown S. L., Douglass L. L., Raymond B. P., Shaw J. D., Sylvester Z. T., et al. (2020). Progress towards a representative network of Southern Ocean protected areas. PLoS One 15, e0231361. doi: 10.1371/journal.pone.0231361
Canteras J., Arnaud P. M. (1985). Les gastéropodes prosobranches des Îles Kerguelen et Crozet (Sud de l’Ocean Indien) comparaisons écologiques et particularités biologiques. Comité Natl. Français Des. Recherches Antarctiques 57, 1–169.
Carr C. M., Hardy S. M., Brown T. M., Macdonald T. A., Hebert P. D. (2011). A tri-oceanic perspective: DNA barcoding reveals geographic structure and cryptic diversity in Canadian polychaetes. PLoS One 6, e22232. doi: 10.1371/journal.pone.0022232
Chevallier L., Nougier J. (1981). Première étude volcanostructurale de l’ile de la Possession, iles Crozet (TAAF), Ocean Indien austral. C.R. Acad. Sci. Paris 292, 363–368.
Christiansen H., Dettai A., Heindler F. M., Collins M. A., Duhamel G., Hautecoeur M., et al. (2018). Diversity of mesopelagic fishes in the Southern Ocean-a phylogeographic perspective using DNA barcoding. Front. Ecol. Evol. 6. doi: 10.3389/fevo.2018.00120
Conlan K. E. (1990). Revision of the crustacean amphipod genus Jassa Leach (Corophioidea: Ischyroceridae). Can. J. Zoology 68, 2031–2075. doi: 10.1139/z90-288
Convey P. (2007). Influences on and origins of terrestrial biodiversity of the sub-Antarctic islands. Papers Proc. R. Soc. Tasmania 141, 83–93. doi: 10.26749/rstpp.141.1.83
Cowart D. A., Schiaparelli S., Alvaro M. C., Cecchetto M., Le Port A. S., Jollivet D., et al. (2022). Origin, diversity, and biogeography of Antarctic scale worms (Polychaeta: Polynoidae): a wide-scale barcoding approach. Ecol. Evol. 12, e9093. doi: 10.1002/ece3.9093
De Broyer C., Koubbi P., Griffiths H. J., Raymond B., Udekem d’Acoz C., Van de Putte A. P., et al. (2014). Biogeographic Atlas of the Southern Ocean (Cambridge: Scientific Committee on Antarctic Research). 498p.
Féral J. P., Poulin E., De Ridder C., Saucède T. (2019). “A field guide to coastal echinoderms of the Kerguelen Islands,” in Proceedings of the 16th International Echinoderm Conference, Nagoya, Japan, 2018 May 28-June 1, Vol. 15, Zoosymposia. doi: 10.11646/zoosymposia.15.1.6
Féral J. P., Saucède T., Poulin E., Marschal C., Marty G., Roca J. C., et al. (2016). PROTEKER: implementation of a submarine observatory at the Kerguelen islands (Southern Ocean). Underwater Technol. 34, 3–10. doi: 10.3723/ut.34.003
Féral J. P., Verlaque M., Rosenfeld S., Poulin E., Chenuil A., Saucède T. (2021). The marine vegetation of the Kerguelen Islands: history of scientific campaigns, inventory of the flora and first analysis of its biogeographical affinities. Cryptogamie Algologie 42, 173–216. doi: 10.5252/cryptogamie-algologie2021v42a12
Figuerola B., Barnes D. K. A., Brickle P., Brewin P. D. (2017). Bryozoan diversity around the Falkland and South Georgia Islands: Overcoming Antarctic barriers. Mar. Environ. Res. 126, 81–94. doi: 10.1016/j.marenvres.2017.02.005
Figuerola B., Gordon D. P., Cristobo J. (2018). New deep Cheilostomata (Bryozoa) species from the Southwestern Atlantic: shedding light in the dark. Zootaxa 4375, 211–249. doi: 10.11646/zootaxa.4375.2.3
Folmer O., Black M., Hoeh W., Lutz R., Vrijenhoek R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 3, 294–299.
Fraser C. I., Dutoit L., Morrison A. K., Pardo L. M., Smith S. D., Pearman W. S., et al. (2022). Southern Hemisphere coasts are biologically connected by frequent, long-distance rafting events. Curr. Biol. 32, 3154–3160. doi: 10.1016/j.cub.2022.05.035
Frenot Y., Gloaguen J. C., Massé L., Lebouvier M. (2001). Human activities, ecosystem disturbance and plant invasions in subantarctic Crozet, Kerguelen and Amsterdam Islands. Biol. Conserv. 101, 33–50. doi: 10.1016/S0006-3207(01)00052-0
González-Wevar C. A., Segovia N. I., Rosenfeld S., Noll D., Maturana C. S., Hüne M., et al. (2021). Contrasting biogeographical patterns in Margarella (Gastropoda: Calliostomatidae: Margarellinae) across the Antarctic polar front. Mol. Phylogenet. Evol. 156, 107039. doi: 10.1016/j.ympev.2020.107039
Gostel M. R., Kress W. J. (2022). The expanding role of DNA barcodes: Indispensable tools for ecology, evolution, and conservation. Diversity 14, 213. doi: 10.3390/d14030213
Griffiths H. J. (2010). Antarctic marine biodiversity–what do we know about the distribution of life in the Southern Ocean? PLoS One 5, e11683. doi: 10.1371/journal.pone.0011683
Griffiths H. J., Barnes D. K., Linse K. (2009). Towards a generalized biogeography of the Southern Ocean benthos. J. Biogeography 36, 162–177. doi: 10.1111/j.1365-2699.2008.01979.x
Griffiths H. J., Waller C. L. (2016). The first comprehensive description of the biodiversity and biogeography of Antarctic and Sub-Antarctic intertidal communities. J. Biogeography 43, 1143–1155. doi: 10.1111/jbi.12708
Guillaumot C., Fabri-Ruiz S., Martin A., Eléaume M., Danis B., Féral J. P., et al. (2018). Benthic species of the Kerguelen Plateau show contrasting distribution shifts in response to environmental changes. Ecol. Evol. 8, 6210–6225. doi: 10.1002/ece3.4091
Hanafi-Portier M., Samadi S., Corbari L., Chan T. Y., Chen W. J., Chen J. N., et al. (2021). When imagery and physical sampling work together: Toward an integrative methodology of deep-sea image-based megafauna identification. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.749078
Hedgepth J. W. (1969). “Introduction to antarctic zoogeography,” in Distribution of selected groups of marine invertebrates in waters south of 35°S. (New York, United States) 1–29.
Hemmings A. D., Stephens T. (2010). The extended continental shelves of sub-Antarctic Islands: implications for Antarctic governance. Polar Rec. 46, 312–327. doi: 10.1017/S0032247409990532
Hullé M., Vernon P. (2021). Terrestrial macro-arthropods of the sub-Antarctic islands of Possession (Crozet Archipelago) and Kerguelen: Inventory of native and non-native species. Zoosystema 43, 549–561. doi: 10.5252/zoosystema2021v43a22
Hutchings P., Reid A. (1991). The nereididae (Polychaeta) from Australia-leonnates, platynereis and solomononereis. Records Aust. Museum 43, 47–62. doi: 10.3853/j.0067-1975.43.1991.40
Irisarri I., Eernisse D. J., Zardoya R. (2014). Molecular phylogeny of Acanthochitonina (Mollusca: Polyplacophora: Chitonida): three new mitochondrial genomes, rearranged gene orders and systematics. J. Natural History 48, 2825–2853. doi: 10.1080/00222933.2014.963721
Jacomy M., Venturini T., Heymann S., Bastian M. (2014). ForceAtlas2, a continuous graph layout algorithm for handy network visualization designed for the Gephi software. PLoS One 9, e98679. doi: 10.1371/journal.pone.0098679
Jossart Q., Bauman D., Moreau C. V., Saucède T., Christiansen H., Brasier M. J., et al. (2023). A pioneer morphological and genetic study of the intertidal fauna of the Gerlache Strait (Antarctic Peninsula). Environ. Monit. Assess. 195, 514. doi: 10.1007/s10661-023-11066-3
Jossart Q., Kochzius M., Danis B., Saucède T., Moreau C. V. (2021). Diversity of the Pterasteridae (Asteroidea) in the Southern Ocean: a molecular and morphological approach. Zoological J. Linn. Soc. 192, 105–116. doi: 10.1093/zoolinnean/zlaa097
Jossart Q., Sands C. J., Sewell M. A. (2019). Dwarf brooder versus giant broadcaster: combining genetic and reproductive data to unravel cryptic diversity in an Antarctic brittle star. Heredity 123, 622–633. doi: 10.1038/s41437-019-0228-9
Kearse M., Moir R., Wilson A., Stones-Havas S., Drummond A. (2012). Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649. doi: 10.1093/bioinformatics/bts199
Kumar S., Stecher G., Li M., Knyaz C., Tamura K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547. doi: 10.1093/molbev/msy096
Larsen K., Shimomura M. (2006). Tanaidacea (crustacea: peracarida) from Japan. I. Apseudomorpha from the east China Sea, Seto inland Sea, and nansei islands. Zootaxa 1341, 29–48. doi: 10.11646/zootaxa.1341.1.2
Layton K. K., Corstorphine E. A., Hebert P. D. (2016). Exploring Canadian echinoderm diversity through DNA barcodes. PLoS One 11, e0166118. doi: 10.1371/journal.pone.0166118
Layton K. K., Martel A. L., Hebert P. D. (2014). Patterns of DNA barcode variation in Canadian marine molluscs. PLoS One 9, e95003. doi: 10.1371/journal.pone.0095003
Leigh J. W., Bryant D. (2015). POPART: full-feature software for haplotype network construction. Methods Ecol. Evol. 6, 1110–1116. doi: 10.1111/2041-210X.12410
Leiva C., Riesgo A., Combosch D., Arias M. B., Giribet G., Downey R., et al. (2022). Guiding marine protected area network design with comparative phylogeography and population genomics: An exemplary case from the Southern Ocean. Diversity Distributions 28, 1891–1907. doi: 10.1111/ddi.13590
Lelièvre Y., Motreuil S., Specq L., Marschal C., Dubois P., Wauters L., et al. (2024). MEDUSA: Marine benthic Ecological Data from Underwater imagery Surveys of sub-Antarctic Crozet environments. Sci. Data 11, 613. doi: 10.1038/s41597-024-03460-4
Lelièvre Y., Specq L., Lamy T., Boyé A., Downey R. V., Saucède T. (2023). Taxonomic and functional diversity of subtidal benthic communities associated with hard substrates at Crozet archipelago (sub-Antarctic, Southern Ocean). Front. Mar. Sci. 10. doi: 10.3389/fmars.2023.1291038
Maroni P. J., Baker B. J., Moran A. L., Woods H. A., Avila C., Johnstone G. J., et al. (2022). One Antarctic slug to confuse them all: the underestimated diversity of Doris kerguelenensis. Invertebrate Systematics 36, 419–435. doi: 10.1071/IS21073
Maroni P. J., Wilson N. G. (2022). Multiple Doris “kerguelenensis”(Nudibranchia) species span the Antarctic Polar Front. Ecol. Evol. 12, e9333. doi: 10.1002/ece3.9333
Mason N. W., Mouillot D., Lee W. G., Wilson J. B. (2005). Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos 111, 112–118. doi: 10.1111/j.0030-1299.2005.13886.x
Miya T., Gon O., Mwale M., Poulin E. (2016). Molecular systematics and taxonomic status of three latitudinally widespread nototheniid (Perciformes: Notothenioidei) fishes from the Southern Ocean. Zootaxa 4061, 381–396. doi: 10.11646/zootaxa.4061.4.4
Moore J. M., Carvajal J. I., Rouse G. W., Wilson N. G. (2018). The Antarctic Circumpolar Current isolates and connects: Structured circumpolarity in the sea star Glabraster Antarctica. Ecol. Evol. 8, 10621–10633. doi: 10.1002/ece3.4551
Moreau C., Danis B., Jossart Q., Eléaume M., Sands C., Achaz G., et al. (2019). Is reproductive strategy a key factor in understanding the evolutionary history of Southern Ocean Asteroidea (Echinodermata)? Ecology and Evolution 9, 8465-8478. doi: 10.1002/ece3.5280
Moreau C., Jossart Q., Danis B., Eléaume M., Christiansen H., Guillaumot C., et al. (2021). The high diversity of Southern Ocean sea stars (Asteroidea) reveals original evolutionary pathways. Prog. oceanography 190, 102472. doi: 10.1016/j.pocean.2020.102472
Moreau C., Saucède T., Jossart Q., Agüera A., Brayard A., Danis B. (2017). Reproductive strategy as a piece of the biogeographic puzzle: a case study using Antarctic sea stars (Echinodermata, Asteroidea). J. Biogeography 44, 848–860. doi: 10.1111/jbi.12965
Neal L., Linse K., Brasier M. J., Sherlock E., Glover A. G. (2018). Comparative marine biodiversity and depth zonation in the Southern Ocean: evidence from a new large polychaete dataset from Scotia and Amundsen seas. Mar. Biodiversity 48, 581–601. doi: 10.1007/s12526-017-0735-y
Owada M., Kasai Y., Sato S. I., Hong J. S. (2013). Shell morphology and genetic characteristics of Corbula (Potamocorbula) spp. (Bivalvia: Corbulidae) from the waters adjacent to Japan, Korea and San Francisco Bay. Aquat. Biol. 19, 297–305. doi: 10.3354/ab00539
Park Y. H., Durand I., Kestenare E., Rougier G., Zhou M., d'Ovidio F., et al. (2014). Polar Front around the Kerguelen Islands: An up-to-date determination and associated circulation of surface/subsurface waters. J. Geophysical Research: Oceans 119, 6575–6592. doi: 10.1002/2014JC010061
Pollard R. T., Venables H. J., Read J. F., Allen J. T. (2007). Large-scale circulation around the Crozet Plateau controls an annual phytoplankton bloom in the Crozet Basin. Deep Sea Res. Part II: Topical Stud. Oceanography 54, 1915–1929. doi: 10.1016/j.dsr2.2007.06.012
Poulin E., González-Wevar C., Díaz A., Gérard K., Hüne M. (2014). Divergence between Antarctic and South American marine invertebrates: What molecular biology tells us about Scotia Arc geodynamics and the intensification of the Antarctic Circumpolar Current. Global Planetary Change 123, 392–399. doi: 10.1016/j.gloplacha.2014.07.017
Puillandre N., Brouillet S., Achaz G. (2021). ASAP: assemble species by automatic partitioning. Mol. Ecol. Resour. 21, 609–620. doi: 10.1111/1755-0998.13281
Puillandre N., Lambert A., Brouillet S., Achaz G. J. M. E. (2012). ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 21, 1864–1877. doi: 10.1111/j.1365-294X.2011.05239.x
Ratnasingham S., Hebert P. D. (2007). BOLD: The Barcode of Life Data System (http:// www. barcodinglife.org). Mol. Ecol. Notes 7, 355–364. doi: 10.1111/j.1471-8286.2007.01678.x
Rosenfeld S., Segovia N. I., Maturana C. S., Aldea C., Saucède T., Brickle P., et al. (2023). A revision of the higher latitude periwinkle species Laevilitorina caliginosa sensu lato. Zoological J. Linn. Soc., zlad171. doi: 10.1093/zoolinnean/zlad171
Sands C. J., O’Hara T. D., Martín-Ledo R. (2021). Pragmatic assignment of species groups based on primary species hypotheses: the case of a dominant component of the Southern Ocean benthic fauna. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.723328
Saucède T., Guillaumot C., Michel L., Fabri-Ruiz S., Bazin A., Cabessut M., et al. (2017). “Modelling species response to climate change in sub-Antarctic islands: echinoids as a case study for the Kerguelen Plateau,” in The Kerguelen Plateau: Marine Ecosystems and Fisheries. (Paris, France) 95–116.
Schmidt A., Brandt A. (2001). The tanaidacean fauna of the Beagle Channel (southern Chile) and its relationship to the fauna of the Antarctic continental shelf. Antarctic Sci. 13, 420–429. doi: 10.1017/S095410200100058X
Sicinski J., Gillet P. (2002). Biogeographical affinities of polychaetes from Îles Crozet. Antarctic Sci. 14, 353–363. doi: 10.1017/S0954102002000172
Sirenko B. (2006). Report on the present state of our knowledge with regard to the chitons (Mollusca: Polyplacophora) of the Magellan Strait and Falkland Islands (< Special Number> the 2nd International Chiton Symposium). Venus (Journal Malacological Soc. Japan) 65, 81–89. doi: 10.18941/venus.65.1-2_81
Sunnucks P., Hales D. F. (1996). Numerous transposed sequences of mitochondrial cytochrome oxidase I-II in aphids of the genus Sitobion (Hemiptera: Aphididae). Mol. Biol. Evol. 13, 510–524. doi: 10.1093/oxfordjournals.molbev.a025612
Teske P. R., McQuaid C. D., Froneman P. W., Barker N. P. (2006). Impacts of marine biogeographic boundaries on phylogeographic patterns of three South African estuarine crustaceans. Mar. Ecol. Prog. Ser. 314, 283–293. doi: 10.3354/meps314283
Vantomme L., Jossart Q., Gérard K., Danis B., Moreau C. (2023). Preliminary assessment of sea star (Echinodermata, Asteroidea) diversity in the coastal magellanic Region (South Chile) and their geographical distribution. Diversity 15, 1129. doi: 10.3390/d15111129
Vieira P. E., Gomes N., Lobo J., Borges L., Queiroga H., Costa F. (2016). A DNA barcode reference library for the superorder Peracarida (Crustacea) from the Southern European Atlantic coast. Front. Mar. Sci. 128. Conference Abstract: XIX Iberian Symposium on Marine Biology Studies.
Keywords: biodiversity, Southern Ocean, taxonomy, barcoding, marine
Citation: Jossart Q, Lelièvre Y, Kelch A, Figuerola B, Moreau CVE, Di Franco D, Maxwell J, Verheye ML, Mackenzie M, Downey R, Rosenfeld S, Hourdez S and Saucède T (2024) A first glimpse into the biogeographic affinities of the shallow benthic communities from the sub-Antarctic Crozet archipelago. Front. Ecol. Evol. 12:1455329. doi: 10.3389/fevo.2024.1455329
Received: 26 June 2024; Accepted: 14 August 2024;
Published: 04 September 2024.
Edited by:
Hamish Spencer, University of Otago, New ZealandReviewed by:
Santiago E. A. Pineda Metz, Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research (AWI), GermanyCopyright © 2024 Jossart, Lelièvre, Kelch, Figuerola, Moreau, Di Franco, Maxwell, Verheye, Mackenzie, Downey, Rosenfeld, Hourdez and Saucède. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Quentin Jossart, cWpvc3NhcnRAZ21haWwuY29t
†ORCID: Quentin Jossart, 0000-0002-2280-243X
Yann Lelièvre, 0000-0003-3508-418X
Blanca Figuerola, 0000-0003-4731-9337
Camille V.E. Moreau, orcid.org/0000-0002-0981-7442
Davide Di Franco, orcid.org/0000-0002-4595-839X
Jamie Maxwell, orcid.org/0000-0001-5705-2811
Marie L. Verheye, orcid.org/0000-0001-8702-9292
Melanie Mackenzie, orcid.org/0000-0002-0030-7032
Rachel Downey, orcid.org/0000-0001-9275-8879
Sebastián Rosenfeld, orcid.org/0000-0002-4363-8018
Stéphane Hourdez, orcid.org/0000-0001-6418-3887
Thomas Saucède, orcid.org/0000-0001-6056-4447
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.