 Karen R. Klinger
Karen R. Klinger Aster F. Hasle
Aster F. Hasle Karen S. Oberhauser
Karen S. Oberhauser
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 31 July 2024
Sec. Conservation and Restoration Ecology
Volume 12 - 2024 | https://doi.org/10.3389/fevo.2024.1444460
The eastern population of monarch butterflies (Danaus plexippus) has dramatically declined in the last few decades, which is largely attributed to a loss of milkweed habitat in agricultural areas. Residential land in metropolitan areas has the potential to provide a significant source of the milkweed needed to support the monarch population. To examine if and how urban milkweed patches can support monarch butterflies, we worked with community scientists in the Chicago metropolitan area to monitor monarch eggs and caterpillars in yards and community gardens. We hypothesized that the largest numbers of eggs (i.e. gardens that were most attractive to monarchs) would be observed in patches that were older, had a high abundance of milkweed, contained more than one species of milkweed including Asclepias syriaca (common milkweed) and/or A. incarnata (swamp milkweed), and had a large diversity of nectar plants. All patches were assessed at their peak egg count each year, which ranged from zero to 170 eggs in a given observation. To determine which characteristics were most influential to egg presence, we examined patches where eggs were present or absent during this peak observation. For abundance, we divided these peak observations into whether or not the egg counts were in the top 20% in a given year, excluding patches where eggs were absent, and assessed their patch characteristics. Our results show that patch age and presence of A. syriaca affected whether patches contained eggs or not. We also found that patches with the largest number of eggs observed tended to have A. syriaca, more milkweed plants, and a higher diversity of blooming plant species. The data we collected from community scientists in the Chicago area has enhanced our understanding of how urban gardeners can create effective breeding habitats for monarch butterflies. By planting Asclepias syriaca within its natural range, along with other native milkweed species and a diverse selection of flowering plants, individuals can create gardens that serve as excellent habitats for monarchs and other pollinators.
The eastern North American population of monarch butterflies (Danaus plexippus) has dramatically declined in the last few decades (Semmens et al., 2016). In the Midwest, a major contributor to this decline was the loss of milkweed plants (Asclepias spp.), the host plants for monarch larvae, in agricultural areas because of increased use in glyphosate herbicide (Pleasants and Oberhauser, 2013; Thogmartin et al., 2017). This loss was especially significant, because agricultural fields provided the largest area of habitat for milkweed in the monarch’s breeding range (Oberhauser et al., 2001). As a result, researchers and government officials have called for the planting of more milkweed across all sectors of the landscape (Thogmartin et al., 2017).
Our research (Johnston et al., 2019) has shown that in many metropolitan areas, single-family homes made up the second largest source of potential plantable space for milkweed after agricultural areas. Although smaller in size, their cumulative area means that even at low milkweed densities, they can provide significant habitat. Further, we found that residents were enthusiastic about supporting monarchs by planting milkweed. Because of the generally small size of most urban gardens, residents frequently have to figure out how to make the best use of their plantable space. While a variety of resources are available to guide gardeners (e.g., Redlinski and Diaz, 2021), many questions remain on how best to attract monarchs to urban yards.
Characteristics that are thought to influence where monarchs lay their eggs include the quality, configuration, and species of milkweed as well as the diversity of nearby nectar flowers. In the Midwest, monarchs lay the most eggs on Asclepias incarnata (swamp milkweed) and Asclepias syriaca (common milkweed) (Ladner and Altizer, 2005; Baker and Potter, 2018; Pocius et al., 2018a, Pocius et al., 2018b). In addition, patches with both milkweed and nearby nectar plants tend to contain more eggs, since nectar resources provide the necessary fuel needed by adult monarchs for flying, reproduction, and migration (Kral-O’Brien et al., 2020; Nestle et al., 2020).
We sought to examine which garden characteristics most influence where monarch butterflies lay their eggs. We conducted a community science project in which participants collected data on the number of monarch eggs and larvae observed on each species of milkweed present in a patch of plants (Hasle et al., In Press). Our expectations were that more monarch butterfly eggs would be observed in patches that were older, had a high abundance of milkweed, contained more than one species of milkweed including A. syriaca and/or A. incarnata, and had a large diversity of neighboring nectar plants.
The Field Museum’s monarch community science project ran from May through September from 2020 through 2022 in the Chicago metropolitan area. Each participant chose one or more sites to monitor: at their homes, public and community gardens, street medians, and nature preserves. At a given site, a participant could choose to monitor anywhere from one to six patches of plants that included at least one species of milkweed. Patches were separated by enough distance or non-habitat (e.g., concrete sidewalk) such that it was unlikely that caterpillars would move from one to another in search of food (Figure 1). Participants were not required to monitor every patch of milkweed at their site, however they were asked to observe all of the milkweed plants within the patches they were monitoring.

Figure 1 Garden site with four patches of plants. Photo credit: Abigail Derby Lewis.
As part of the initial site survey, participants provided information about the site and patches they planned to monitor. Site information included a unique site name, the address or location of the site, and what type of garden maintenance occurred. Patch information included when the patch was planted, whether more than half of the plants were located in pots, and the species of milkweed in the patch.
During weekly data collection, participants examined milkweed plants in their patch(es) and recorded the number of milkweed plants searched, the number of monarch eggs, and the number of each larval instar found for each milkweed species present. During monitoring, participants could also choose to record the number of adult monarch butterflies observed, other insects and pollinators seen, as well as the different species of plants that were blooming. In addition to weekly monitoring, participants were asked to provide the shape of their patch and measure the appropriate dimensions (e.g., length and width or radius) in inches during the month of July. A full description of the project methodology can be found in Hasle et al., In Press.
Our analysis primarily focused on the characteristics of individual milkweed patches located within sites. Each patch in a given year was treated as an independent record, because patches were monitored as separate units and had distinct characteristics that often changed each year. In order to account for shared characteristics, a site’s name was used to group patches that occurred at the same location.
Participants monitored their patches anywhere from one to 18 weeks, which meant that there was a greater chance of eggs being observed in patches that were monitored more frequently. We sought to limit the influence of the amount of monitoring on egg observations by focusing only on patches where there were at least three monitoring observations between June and August, the primary months when surveying occurred. From each patch’s monitoring records during this period, we identified the single observation when the maximum number of eggs were observed. This allowed us to examine the characteristics of patches at their highest egg production.
This approach is similar to that used by Stenoien et al. (2015), which examined the maximum egg density observed at a site during two-month periods for sites with at least four sampling events and more than 10 plants. Our analysis focused on the maximum number of eggs observed rather than maximum egg density, because a significant portion of patches had less than 10 milkweed plants and these smaller patches of milkweed are known to have abnormally high egg densities (Stenoien et al., 2015). If a patch had multiple observations with the same maximum number of eggs, then the monitoring observation with the maximum number of plants was used. If there were still multiple observations, the monitoring record with the latest date was used.
In addition to only including patches with at least three observations, this analysis only included records where both patch measurements and the number of blooming flowers observed were recorded. Patch size was calculated using the provided measurements for the patch’s dimensions and converted from square inches to square meters. If there was no number of blooming flowers provided during the selected observation, the number of blooming species for the closest week within a month was used. As a result, all patches had the same variables available for analysis.
The variables in this analysis included site name; survey year (2020, 2021, or 2022); age of the patch (which participants selected as either planted in the current year, one year ago, 2–5 years ago, or more than five years ago); milkweed species diversity (presence of one milkweed species or more than one milkweed species); the presence or absence of A. syriaca, A. incarnata, Asclepias tuberosa (butterfly milkweed), and Asclepias verticillata (whorled milkweed) (the four most common species of milkweed observed in patches); number of milkweed plants; number of blooming flower species; and patch size as recorded during the month of July. The values for the number of milkweed plants, number of blooming plant species, and patch size were divided into four similarly sized categories: Minimum–25th quartile (low), 25th–50th quartile (low-medium), 50th–75th quartile (medium-high), and 75th quartile–Maximum (high). By grouping these variables into categories instead of using their numeric values, we were able to lessen the impact of small errors in measurements, reduce the influence of outliers, and examine broader patterns.
We used Chi-square tests of independence and multiple regression to assess the relationships of monarch egg presence and egg abundance with the variables of interest. For all patches in the analysis, we determined the presence or absence of eggs during the selected observation. To examine abundance, we used the same selected observation to determine the patches that were or were not in the top 20% of egg observations in each survey year, excluding patches where eggs were absent. We chose not to use the values for the number of eggs observed, because the significant variation each year meant that an egg count could be considered high one year and low the next. All statistical analyses were performed with RStudio version 2023.06.1 with R version 4.2.2 (R Core Team 2023; RStudio Team 2023).
We used binomial generalized linear mixed models (GLMMs) with the lme4 package (Bates et al., 2015) to examine the patch characteristics that affect the presence of eggs and egg abundance (whether a patch was in the top 20% of egg observations). Both models included site name as a random effect to cluster patches into their respective sites. To assess the effect of survey year on the presence of eggs, we explored including it as both a random effect and as a fixed effect. Survey year was not included in the model for egg abundance, since it was accounted for in calculating the top 20% of observations. All other variables (age of the patch, milkweed species diversity, presence of A. syriaca, presence of A. incarnata, presence of A. tuberosa, presence of A. verticillata, number of milkweed plants, number of blooming flower species, and patch size) were included as fixed effects. To ensure that variables were uncorrelated, we calculated the variance inflation factor (VIF) of each independent variable in the model (VIF < 3). We calculated R2 values with the r2 function in the performance package (Lüdecke et al., 2020). Akaike Information Criteria (AIC) values were used to rank the top supported comparable models.
From 2020 to 2022, participants recorded 5,166 observations on 682 patches of milkweed. Between June and August, 450 patches recorded at least three monitoring observations and provided both patch size data and number of blooming flower species. During these three months, 134 patches were monitored from three to six times, 175 were monitored from seven to ten times, and 141 were monitored from 11–25 times. The categorical ranges for number of milkweed plants within patches included: 1–6 (low), 7–12 (low-medium), 13–26 (medium-high), and 27–498 (high). The categorical ranges for number of blooming plant species within patches included: 0–2 (low), 3–4 (low-medium), 5–6 (medium-high), and 7–29 (high). The categorical ranges for patch size included: 0.012–<0.85 square meters (low), 0.85–<2.7 square meters (low-medium), 2.7–<8.5 square meters (med-high), and 8.5–4,738 square meters (high).
Of the 450 patches included in this analysis, eggs were observed in 340 patches, with one to 170 eggs in the selected monitoring session in these 340 patches. Chi-square tests indicated that survey year (χ2 = 18, df = 2, p < 0.001), patch age (χ2 = 35, df = 3, p < 0.001), presence of A. syriaca (χ2 = 37, df = 1, p < 0.001), and number of milkweed plants (χ2 = 20, df = 2, p < 0.001) had significant relationships with the presence of eggs in a patch.
In the best-fit logistic regression model with mixed effects (Model 1) and in competing models, a patch age of 5 years or older and the presence of A. syriaca significantly increased the odds of eggs being present (Table 1). For all top models, the years 2021 and 2022 had a negative relationship with the presence of eggs. The presence of A. incarnata and the number of plants, especially those in the high category, increased the likelihood of eggs being present in a patch.
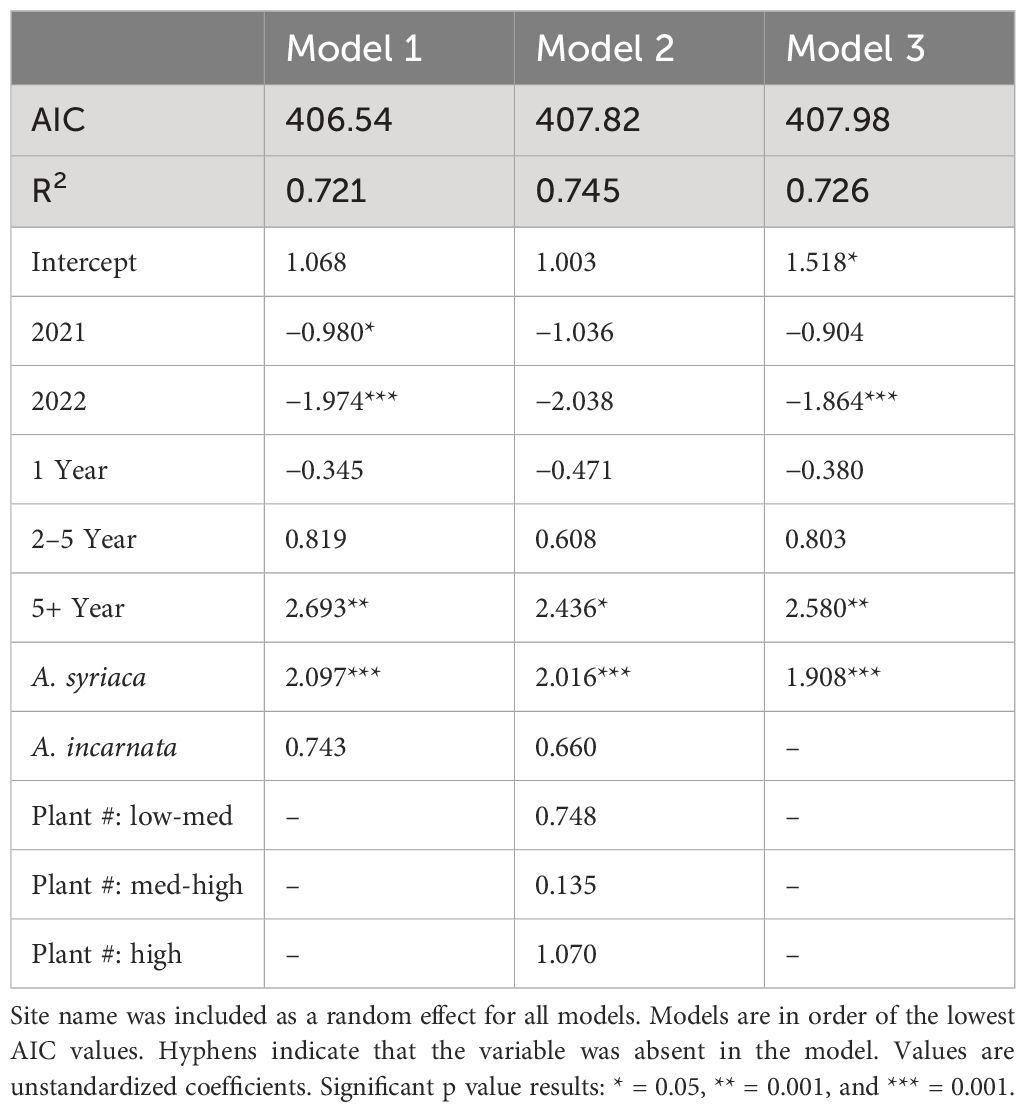
Table 1 The top three models comparing egg presence against predictor variables.
To examine patches that had a higher abundance of eggs, the 340 patches with at least one egg observation were divided into 2 groups for each year: patches with the top 20% of egg observations and patches in the bottom 80% of observations. A total of 74 patches were in the top 20%: 31 patches with 18 or more eggs in 2020, 25 patches with 30 or more eggs in 2021, and 18 patches with 10 or more eggs in 2022. Chi-square tests indicated that the presence of A. syriaca (χ2 = 15, df = 1, p < 0.001), the number of milkweed plants (χ2 = 36, df = 3, p < 0.001), and the number of blooming plant types (χ2 = 21, df = 3, p < 0.001), had significant relationships with the likelihood of a patch being in the top 20% of egg observations.
In the best-fit logistic regression model with mixed effects (Model 1), the presence of A. syriaca, more milkweed plants, and more blooming plant types increased the odds of patches being in the top 20% of egg observations (Table 2). The model’s intercept, which corresponded to no A. syriaca, a low number of plants, and a low number of blooming flower species decreased the odds of patches having the top 20% of egg observations.
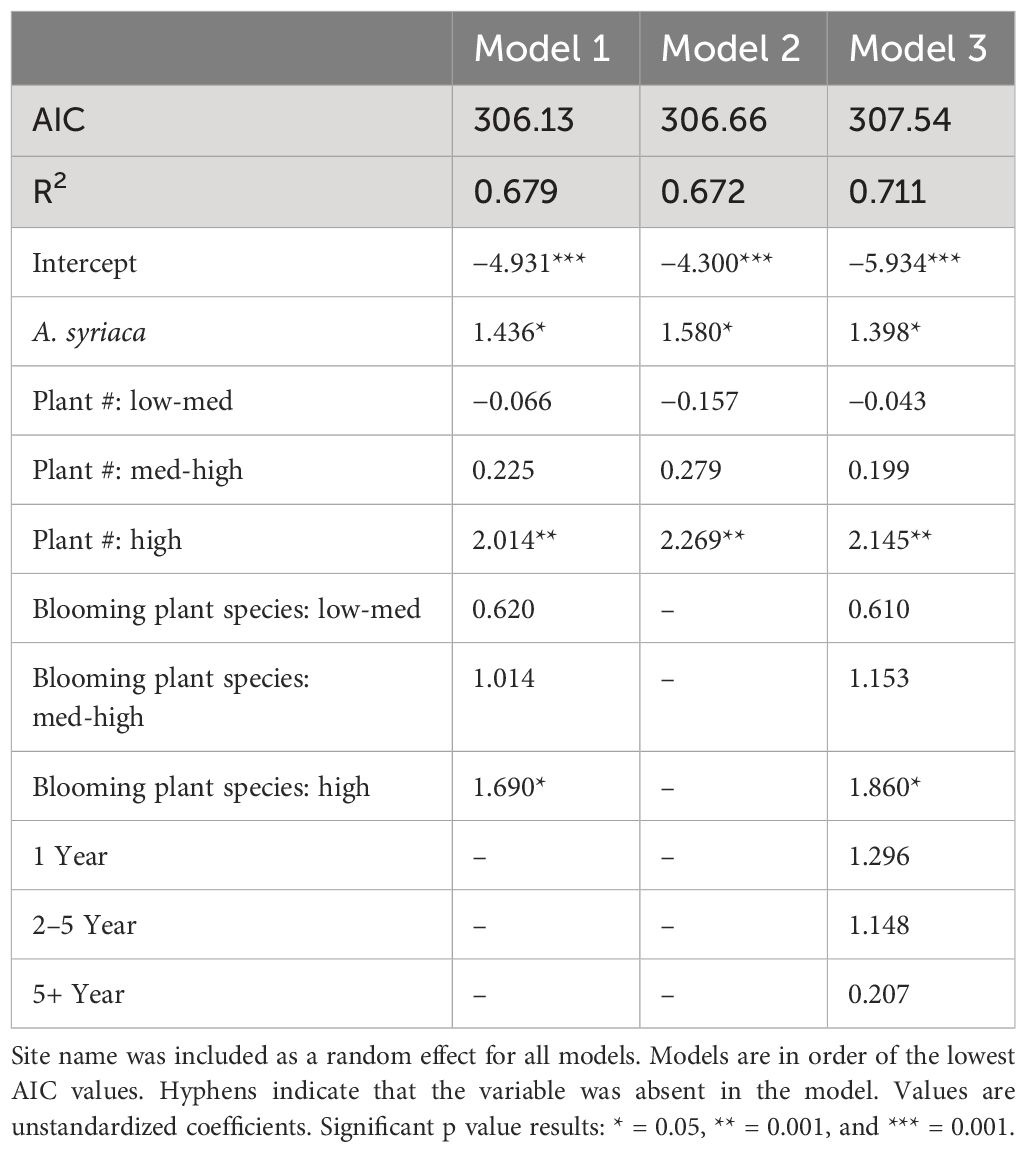
Table 2 The top three models comparing top 20% of egg observations against predictor variables.
We also found that as the number of milkweed plants in a patch increased, the average number of eggs observed increased, while the average egg density (eggs/plant) decreased (Table 3).
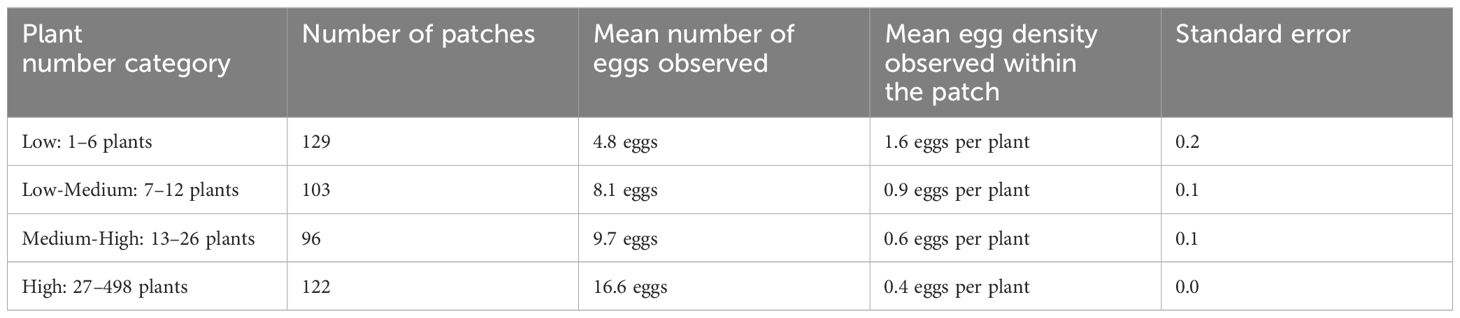
Table 3 Summary of egg observations by number of milkweed plants.
One of the top blooming flower species in participant’s patches was A. tuberosa, which was present during all three months of the study period (Table 4). A. syriaca was also observed blooming in a large number of patches during June and July. Because blooming flowering plants was an optional field for participants to fill in to the best of their knowledge, some participants provided the common name of a plant, while others included the genus and species. Further, since this data came from planted gardens, it is difficult to determine whether some of these flowering plants included cultivars, which can be difficult to distinguish. As a result, purple coneflower (Echinacea purpurea), coneflower (Echinacea spp., Rudbeckia spp.), and black-eyed Susan (Rudbeckia fulgida, Rudbeckia hirta) were all included in the top five most abundant plants in both July and August.
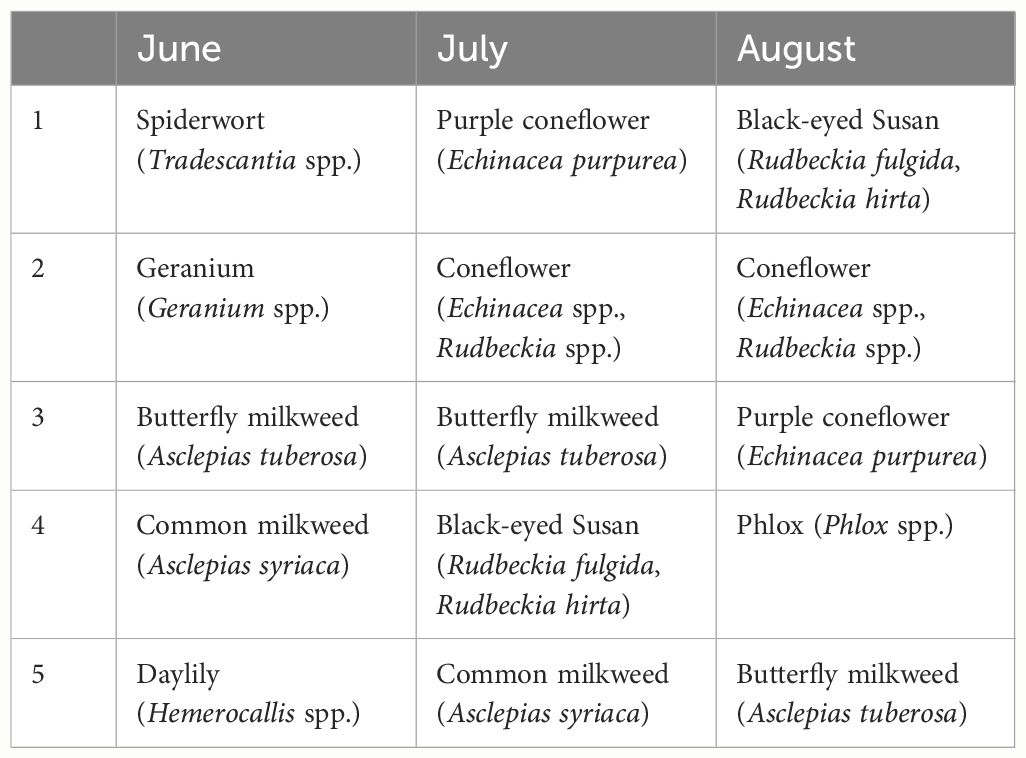
Table 4 The top five most abundant blooming plants during June, July, and August.
Residential gardens in metropolitan areas have the potential to provide significant habitat for monarch butterflies. Because urban gardeners often have limited space in which to plant milkweed and nectar plants, many questions arise on how they can best attract monarch butterflies to their yards. These questions include which and how many species of milkweed to plant, how many milkweed plants are needed, and the importance of having a diversity of other flowering plants nearby. We sought to examine which patch characteristics most influence the presence and abundance of monarch eggs in people’s milkweed gardens.
Based on previous research, we expected that the most successful patches would be older, have a high number of milkweed plants, would have A. syriaca or A. incarnata, would have at least two species of milkweed, and would have a large diversity of neighboring nectar plants. We found that the presence of eggs was most affected by the inclusion of A. syriaca and the age of planting. The occurrence of A. incarnata and the number of milkweed plants also positively affected whether eggs were observed. We also found that the presence of A. syriaca, the number of milkweed plants, and the number of blooming plant species were positively associated with a higher egg abundance.
Asclepias syriaca was present in 60% of patches and in 70% of sites. While A. syriaca may not be the most garden-friendly plant due to its ability to spread aggressively and the difficulty in removing it, it has a key role in many people’s milkweed gardens and contributes significant habitat for monarchs. Among patches that contained A. syriaca, 86% observed eggs, while 60% of patches without A. syriaca observed eggs. These patches are not unusual; A. syriaca is the most common host plant for North American monarchs (Agrawal et al., 2015), with one estimate (Malcolm et al., 1993) suggesting that over 90% of the monarchs that migrate to Mexico have fed on this species. Because of its prevalence in our study area, especially within the patches and sites that were largest in area and quantity of milkweed, we also found that the presence of A. syriaca was associated with other patch characteristics including patch age, the number of milkweed plants, and the size of the patch. Thus, it likely had a confounding effect in the regression models, although VIF values were acceptable.
Patches that were at least 5 years of age were significantly more likely to contain eggs than patches in any other age group. Of the 78 patches in this category, 95% contained eggs; this value was only 61% for patches planted in the current year. We found that patch age was associated with both patch size and the number of milkweed plants, which indicates that more established patches tended to have more area and more plants available for monarchs to lay their eggs on. While these patches may have been established at least 5 years ago, it is very likely that newer plants were added over time. Thus, more research is needed on the importance of sustaining older milkweed gardens.
Patches in our study came in all shapes and sizes, from a single milkweed plant to almost 500 milkweed plants. For this analysis, the number of milkweed plants in a patch was divided into four approximately even-sized categories: low (1–6 plants), low-medium (7–12 plants), medium-high (13–26 plants), and high (27–498 plants). While people with smaller milkweed plantings did observe monarch eggs, having a larger number of milkweed plants significantly increased the chances that more eggs were seen. As the number of plants in a patch increased, the average number of eggs per plant decreased, while the total number of eggs increased. This is consistent with Kasten et al. (2016) and Bruce et al. (2022), who found that even though more eggs were seen per plant in patches with fewer plants, patches with more plants tended to contain more eggs. We also found that the number of milkweed plants in a patch was positively correlated with the size of the patch, which may explain why it did not show up in the top models.
While the number of blooming flower species did not affect the presence or absence of eggs, it was a significant factor in determining which patches observed the most eggs. This seems to indicate that female monarchs will lay eggs when fewer blooming flower species are present, but they may stay longer and lay more eggs when a larger diversity of nectar resources is available. Thus, our data support the recommendation of having a diverse array of blooming flowers as part of monarch gardens. Since different species bloom at different times, planting a mix of species ensures that monarchs and other pollinators have a stable supply of nectar from spring to fall (Lukens et al., 2020; Tew et al., 2022). Finally, gardeners should remember that milkweed species are also good nectar sources, as both A. syriaca and A. tuberosa were included in the top five most abundant blooming plants between June and August.
It is important to note that this analysis is based on data collected by community scientists. While this enabled us to collect a significant amount of data on sites that would otherwise be difficult to access (Hasle et al., In Press), there are some concerns about data accuracy, data consistency, and bias which may have influenced our results. One of the most significant challenges with community science is the limited ability to ensure data accuracy. Participants were trained on how to identify eggs as well as how to record and submit their data during a two-hour (often-virtual) training session. While some incidents of data inaccuracies (from misidentification and mistakes in submissions) were caught and fixed, there are likely other errors that persisted in the data. Also, since participants chose when and where to monitor, there was some variation and bias in data collection and site selection. Participants monitored anywhere from once to 18 weeks. Because of the significant variation in the amount of data collected, this analysis focused on one observation per patch in a given year. As a result, we analyzed a small fraction of the data that was collected. The amount of time a person spent searching for eggs may also have influenced their ability to find eggs. For example, 82% of participants who monitored at least 11 times between June and August observed eggs, while 71% of participants who monitored three to six times observed eggs. Finally, because participants selected their sites to monitor, many likely chose gardens that were planted with monarchs and other pollinators in mind. Thus, many sites may have been of much higher quality than is typical of gardens in metropolitan areas.
Our protocol emphasized monitoring of individual patches rather than sites as a whole, which meant that a participant could choose not to monitor all of the milkweed plants at their site and they could choose to monitor their patches on different days and for a different number of weeks. A site’s patches could range from being close together to being on opposite sides of a property (e.g., front and back yard). Patches at the same site were also unique and could significantly differ in the number and diversity of milkweed species, number of milkweed plants, patch age, and patch size. We found that the number of eggs observed at each patch within a site was highly variable. Thus, for this analysis, we used the site’s name as a random effect to cluster patches into their respective sites rather than consolidating the data at the site level to avoid misrepresenting the collected data and losing the specificity observed at individual patches. However, analysis examining the data at the site-level found similar results.
The most significant characteristics in determining the success of a patch in attracting monarch butterfly eggs included the number of milkweed plants, the presence of A. syriaca (common milkweed), the diversity of nearby nectar flowers, and the age of the patch. While we found that gardens with more milkweed plants tended to contain more eggs, we discovered that even patches with a single milkweed plant had eggs present. Thus, even very small urban plantings can help to support monarchs. While A. syriaca was a significant factor in determining the presence and number of eggs observed, we recognize that it is not always the most desirable species to plant and encourage gardeners to plant the species of milkweed that best suits their needs. We also encourage people to plant a greater diversity of blooming flowers around their milkweed, which will ensure nectar availability throughout the season, allowing monarchs to stay in the area longer and potentially lay more eggs. Finally, we found that older milkweed patches were more likely to have eggs present and contained the most eggs. However, milkweed gardens of all ages can support monarchs.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
KK: Conceptualization, Data curation, Formal analysis, Methodology, Project administration, Writing – original draft, Writing – review & editing, Investigation, Visualization. AH: Conceptualization, Methodology, Project administration, Writing – review & editing, Investigation. KO: Methodology, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Funding for the research included support from the Maxwell/Hanrahan Foundation to support graduate students in a field-based internship for urban monarch and pollinator conservation.
We gratefully acknowledge our community science participants who made this project possible. Our thanks to our monarch community science coordinators, Adriana Fernandez, Airis Cervantes, and Lily Peppers who provided essential support in running the project each year. We would also like to thank Wendy Caldwell, Katie-Lyn Bunney, Julia Whidden, and Karen Tuerk for their guidance in aligning and ultimately transferring this project to the Monarch Larva Monitoring Project and their participation in our annual results presentations. Finally, we would like to thank our colleagues and funders for their support.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Agrawal A. A., Ali J. G., Rasmann S., Fishbein M. (2015).“Macroevolutionary trends in the defense of milkweeds against monarchs,”, in Monarchs in a changing world: biology and conservation of an iconic butterfly. Eds. Oberhauser K. S., Nail K. R., Altizer S. (Cornell University Press, Ithaca, NY), 47–59.
Baker A. M., Potter D. A. (2018). Colonization and usage of eight milkweed (Asclepias) species by monarch butterflies and bees in urban garden settings. J. Insect Conserv. 22, 405–418. doi: 10.1007/s10841-018-0069-5
Bates D., Mächler M., Bolkwer B., Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Soft. 67, 1–48. doi: 10.18637/jss.v067.i01
Bruce A. S., Thogmartin W. E., Trosen C., Oberhauser K., Gratton C. (2022). Landscape- and local-level variables affect monarchs in Midwest grasslands. Landsc. Ecol. 37, 93–108. doi: 10.1007/s10980-021-01341-4
Johnston M. K., Hasle E. M., Klinger K. R., Lambruschi M. P., Derby Lewis A., Stotz D. F., et al. (2019). Estimating milkweed abundance in metropolitan areas under existing and user-defined scenarios. Front. Ecol. Evol. 7. doi: 10.3389/fevo.2019.00210
Kasten K., Stenoien C., Caldwell W., Oberhauser K. S. (2016). Can roadside habitat lead monarchs on a route to recovery? J. Insect Conserv. 20, 1047–1057. doi: 10.1007/s10841-016-9938-y
Kral-O’Brien K. C., Hovick T. J., Limb R. F., Harmon J. P., Gillam E. H. (2020). Incorporating field behaviors into monarch surveys to promote informed conservation actions. J. Nat. Conserv. 53: 125761. doi: 10.1016/j.jnc.2019.125761
Ladner D. T., Altizer S. M. (2005). Oviposition preference and larval performance of North American monarch butterflies on four Asclepias species. Entomol. Exp. Appl. 116, 9–20. doi: 10.1111/j.1570-7458.2005.00308.x
Lüdecke D., Makowski D., Waggoner P., Patil I. (2020). Assessment of regression models performance (CRAN). doi: 10.5281/zenodo.3952174
Lukens L., Kasten K., Stenoien C., Cariveau A., Caldwell W., Oberhauser K. (2020). Monarch habitat in conservation grasslands. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.00013
Malcolm S. B., Cockrell B. J., Brower L. P. (1993). “Spring recolonization of eastern North America by the monarch butterfly: successive brood or single sweep migration?”,” in Biology and conservation of the monarch butterfly, vol. 38 . Eds. Malcolm S. B., Zalucki M. P. (Natural History Museum of Los Angeles County, Science Series, Los Angeles, CA), 253–267.
Nestle R., Daniels J. C., Dale A. G. (2020). Mixed-species gardens increase monarch oviposition without increasing top-down predation. Insects 11, 648. doi: 10.3390/insects11090648
Oberhauser K. S., Prysby M. D., Mattila H. R., Stanley-Horn D. E., Sears M. K., Dively G. P., et al. (2001). Temporal and spatial overlap between monarch larvae and corn pollen. Proc. Natl. Acad. Sci. U.S.A. 98, 11913–11918. doi: 10.1073/pnas.211234298
Pleasants J. M., Oberhauser K. S. (2013). Milkweed loss in agricultural fields because of herbicide use: effect on the monarch butterfly population. Insect Conserv. Divers. 6, 135–144. doi: 10.1111/j.1752-4598.2012.00196.x
Pocius V. M., Debinski D. M., Pleasants J. M., Bidne K. G., Hellmich R. L. (2018a). Monarch butterflies do not place all of their eggs in one basket: oviposition on nine Midwestern milkweed species. Ecosphere 9, e02064. doi: 10.1002/ecs2.2064
Pocius V. M., Pleasants J. M., Debinski D. M., Bidne K. G., Hellmich R. L., Bradbury S. P., et al. (2018b). Monarch butterflies show differential utilization of nine midwestern milkweed species. Front. Ecol. Evol. 6. doi: 10.3389/fevo.2018.00169
R Core Team. (2023). R: a language and environment for statistical computing. R Foundation for Statistical Computing. 6. Available at: https://www.R-project.org/.
Redlinski I., Diaz A. (2021). Creating monarch habitat in your Midwestern garden. Available online at: https://fieldguides.fieldmuseum.org/sites/default/files/rapid-color-guides-pdfs/878_usa_illinois_monarchhabitat_version2.pdf (Accessed May 16, 2022).
Semmens B. X., Semmens D. J., Thogmartin W. E., Wiederholt R., López-Hoffman L., Diffendorfer J. E., et al. (2016). Quasi-extinction risk and population targets for the Eastern, migratory population of monarch butterflies (Danaus plexippus). Sci. Rep. 6, 23265. doi: 10.1038/srep23265
Stenoien C., Nail K. R., Oberhauser K. S. (2015). Habitat productivity and temporal patterns of monarch butterfly egg densities in the Eastern United States. Ann. Entomol. Soc Am. 108, 670–679. doi: 10.1093/aesa/sav054
Tew N. E., Baldock K. C. R., Vaughan I. P., Bird S., Memmott J. (2022). Turnover in floral composition explains species diversity and temporal stability in the nectar supply of urban residential gardens. J. Appl. Ecol. 00, 1–11. doi: 10.1111/1365-2664.14094
Keywords: Asclepias, monarch butterfly, urban, garden, community science
Citation: Klinger KR, Hasle AF and Oberhauser KS (2024) Characteristics of urban milkweed gardens that influence monarch butterfly egg abundance. Front. Ecol. Evol. 12:1444460. doi: 10.3389/fevo.2024.1444460
Received: 05 June 2024; Accepted: 08 July 2024;
Published: 31 July 2024.
Edited by:
Jay E. Diffendorfer, United States Department of the Interior, United StatesReviewed by:
Christie Bahlai, Kent State University, United StatesCopyright © 2024 Klinger, Hasle and Oberhauser. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karen R. Klinger, a2tsaW5nZXJAZmllbGRtdXNldW0ub3Jn
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.