 Muhammad Arif Asadi1,2,3*
Muhammad Arif Asadi1,2,3* Alvu Sya’ban Al-kareem1Riqki Yoga Aprilianto1
Alvu Sya’ban Al-kareem1Riqki Yoga Aprilianto1 Aida Sartimbul1,4
Aida Sartimbul1,4 Ade Yamindago1,2Dhira Khurniawan Saputra1,2
Ade Yamindago1,2Dhira Khurniawan Saputra1,2 Achmad Riyadi1
Achmad Riyadi1- 1Marine Science Department, Faculty of Fisheries and Marine Science, Brawijaya University, Malang, East Java, Indonesia
- 2Coastal Resilience and Climate Change Adaptation (CORECT) Research Group, Brawijaya University, Malang, East Java, Indonesia
- 3Eksplorasi Sumberdaya Perikanan dan Kelautan (ESPK) Laboratory, Faculty of Fisheries and Marine Science, Brawijaya University, Malang, East Java, Indonesia
- 4Marine Resources Exploration and Management (MEXMA) Research Group, Brawijaya University, Malang, East Java, Indonesia
Mangroves on the islands along the Java Sea play a crucial role in supporting biodiversity, protecting coastlines, and sequestering carbon. However, there is a lack of information on mangrove structure and biomass in the area. This study aimed to assess the mangrove structures and biomass on Desa Daun, located on the Bawean Islands, and Kemujan Island, located on the Karimunjawa Islands, respectively. The study utilized a combination of line transect and quadrant plot methods with a size plot of 10x10 m. We established 7 line transects and 28 quadrant plots in the mangrove forest of Desa Daun and similarly set up 7 line transects and 21 quadrant plots on Kemujan Island. Within each quadrant, the mangrove species were identified, and their diameters at breast height (DBH) were recorded. To assess the relative significance of each species, the importance value index (IVI) was calculated, and allometric equations were applied to estimate biomass and carbon storage in the trees. PlanetScope imagery was also used to quantify the carbon stock of mangroves on those islands. In Desa Daun, 13 mangrove species were identified, while 12 species were recorded on Kemujan Island. The species Nypa fruticans and Excoecaria agallocha exhibited the highest IVI in Desa Daun and Kemujan Island, respectively. Notably, Rhizophora apiculata demonstrated the greatest biomass in both locations, with measurements of 43.08 Mg B/ha in Desa Daun and 102.22 Mg B/ha in Kemujan Island. Overall, mangrove stands on Desa Daun stored biomass of 113.60 Mg B/ha and carbon of 50.98 Mg C/ha, whereas mangrove stands on Kemujan Island stored biomass of 295.04 Mg B/ha and carbon of 131.95 Mg C/ha. Mangrove stands of the whole Bawean Islands sequestered 22,941 Mg C, while Karimunjawa Islands hold much higher C stocks, 79,451 Mg C. Despite being smaller in size, the Karimunjawa Islands have more extensive mangrove coverage compared to the Bawean Islands due to their geomorphological characteristics, which in turn influence their mangrove structures and carbon stocks. However, the loss of mangroves in the Karimunjawa Islands due to aquaculture expansions could increase carbon loss in the islands.
Introduction
Mangrove forests are thriving at the interface between land and sea along tropical and subtropical latitudes. They serve valuable economic benefits and ecological services, providing food and wood resources as well as nursery grounds for a wide variety of fish, crustaceans, and other invertebrates. Mangroves also regulate nutrient cycling, deposit sediment, and protect coastal areas and communities from coastal erosion, tsunamis, and tropical storms (Hanggara et al., 2021; Nozarpour et al., 2023; Ruslan et al., 2022; Trégarot et al., 2021). As a large sink of carbon, mangrove forests play a crucial role in mitigating the adverse impact of climate change. Globally, mangrove ecosystems account for 14% of carbon sequestration by the ocean (Kandasamy et al., 2021; Nyanga, 2020).
Mangroves occupy a global area of 147,000 km2, roughly the same area as Bangladesh (Leal and Spalding, 2022). Indonesia has the most extensive mangrove forests, about 22% of the global total (Giri et al., 2011; Sasmito et al., 2023). Other countries with significant mangrove coverage—Brazil, Nigeria, Mexico, and Australia—together host about 23% of global mangrove areas (Leal and Spalding, 2022). However, the world’s mangrove coverage had been declining by 1.58% annually between 2007 and 2016 (Hagger et al., 2022). It is estimated that 38% of global mangrove loss is due to shrimp culture expansion, and fuelwood and timber production are responsible for 26% of mangrove extent loss worldwide (Nguyen et al., 2022; Ramarokoto et al., 2024). In Indonesia, around 800,000 ha of mangroves have disappeared over the past 30 years (Sasmito et al., 2023).
Mangrove forests could, in as short as 100 years, at least functionally disappear due to their rapid pace of destruction (Moity et al., 2019). The loss of mangroves has severely impacted their ecosystem biomass. Soil carbon and tree biomass will be lost by at least 25% following the mangrove destruction (Sasmito et al., 2019). Concerning individual mangrove species loss is also crucial, since even perfect mangrove habitats are species-poor when compared to other tropical plant ecosystems (Canty et al., 2022). Mangrove areas with declining mangrove diversity and continuing loss of mangrove species will have environmental consequences and devastating economic impacts for coastal communities (Lovelock et al., 2022; Yamamoto, 2023). Therefore, mangrove structure and biomass inventory are essential tools not only for preserving mangrove ecosystems but also for supporting biodiversity, protecting coastal areas, and contributing to climate change mitigation and adaptation efforts.
The total mangrove extant in the Java Islands is relatively small compared to the eastern Indonesia region (Basyuni et al., 2022), and most of them are increasingly fragmented, which poses a greater risk of exploitation (Asadi et al., 2019; Asy’Ari et al., 2022). Mangrove forests also exist on some groups of islands along the Java Sea, like the Karimunjawa Islands in Central Java, the Bawean Islands in East Java, and the Thousand Islands of the Capital Region of Jakarta. However, most mangrove structures and biomass studies are on the main island of Java (Asadi et al., 2019; Slamet et al., 2020; Soeprobowati et al., 2024; Trissanti et al., 2022). Mangroves on small islands typically have lower species diversity and are more vulnerable to changes due to their limited habitat area and higher salinity levels. In contrast, mangroves on large islands benefit from greater habitat diversity, freshwater input, and species richness (Sealey, 2024; Veitayaki et al., 2017; Visschers et al., 2022). However, there was no study that compared the mangrove structure and biomass of the Bawean Islands and Karimunjawa Islands and estimated the total mangrove carbon of the whole islands along the Java Sea. The differences in geomorphological characteristics of both areas may influence their mangrove structure and biomass. Therefore, in this study, a field assessment of the forest structure and biomass of Kemujan Island in the Karimunjawa Islands and Desa Daun Mangrove Forest in the Bawean Island was conducted in order to understand the mangrove species structure and biomass of both areas.
Materials and methods
Study sites
The field data collection was carried out in two locations: Daun village and Kemujan Island. The first research site, Daun village, is located on Bawean Island, 135 km north of the city of Gresik. The island has an area of 197 km2, a population of more than 80,000 people, and is under the administration of Gresik Regency, East Java, Indonesia. This site is the main mangrove area of the island, situated at a latitude of 5°50’44”S and a longitude of 112°42’21”E, and managed by POKWASMAS Hijau Daun. The main mangrove species is Rhizopora apiculata, and in its surrounding riverine area, Nypa fruticans dominates mangrove stands. The total area of mangrove stands on the site is estimated at around 108 ha. Furthermore, based on satellite data, the whole Bawean Islands (including some small satellite islands nearby) have mangrove coverage of 450 ha.
The second location, Kemujan Island, is part of the Karimunjawa islands, approximately 100 km north of the town of Jepara. The island has an area of 15 km2 and a population of around 3,000 people. The whole Karimunjawa Islands have an area of 70 km2 and a population of 9700 people and are under the administration of Jepara Regency, Central Java, Indonesia. Kemujan Island is situated at a latitude of 5°49’24”S and a longitude of 110°27’56”E and is under the management of Karimunjawa National Park. Kemujan Island is separated from the main island, Karimunjawa Island, by a narrow river, 2–3 m wide. The mangrove area of Kemujan Island amounts to 232 ha, and the mangrove formation of the whole Karimunjawa Islands has an estimated area of 602 ha. The maps of research areas are presented in Figures 1, 2.

Figure 1. The sampling stations on Desa Daun, Bawean Island.
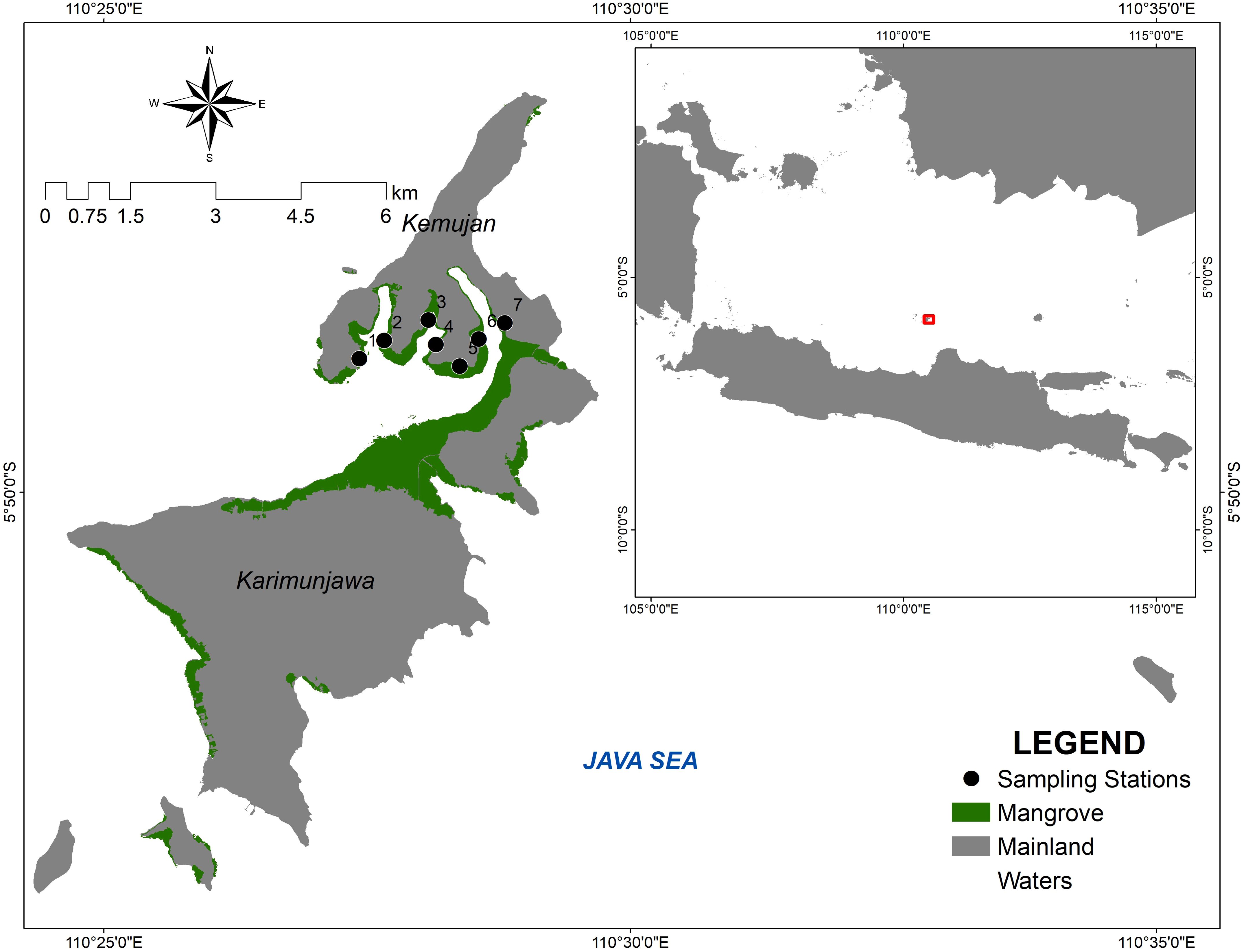
Figure 2. The sampling stations on Kemujan Island.
Data collection
The field data collection in Kemujan was performed in April and May 2021, at the end of the rainy season, while in Desa Daun, it was in June 2022, during the dry season. The optimal greeness of mangroves on small islands occurs in the rainy season, with maximum greeness occurring at the end of the rainy season and the beginning of the dry season (Prihantono et al., 2022). Small islands, in contrast to larger ones, typically lack a large river that supplies freshwater to the mangroves. As a result, groundwater and precipitation provide the island’s mangroves with freshwater (Hayes et al., 2019).
This study employed a non-destructive approach by integrating line transect and quadrat sampling techniques at both research sites. At each station, a 100-meter line transect was utilized, with a 10 x 10 meter quadrat placed along the line, maintaining a distance of 10 meters between each quadrat. There were seven stations established in each location, with four quadrats implemented at each station in Desa Daun and three quadrats set up at each station in Kemujan Island, resulting in a total of 28 quadrats in Desa Daun and 21 quadrats in Kemujan Island.
Desa Daun and Kemujan Island are the main mangrove areas and have the largest mangrove coverage on their respective islands. Within each sampling plot, all trees with at least 2.5 cm in diameter were identified and measured at breast height (DBH). Measuring the DBH in mangroves helps estimate biomass, which is crucial for understanding their carbon storage capacity and contribution to mitigating climate change (Zanvo et al., 2023). Additionally, DBH measurements provide insights into forest structure, aiding in the assessment of forest health and biodiversity (Trégarot et al., 2021; Wang et al., 2023). For mangroves of the genus Rhizophora, DBH was measured at 30 cm above the highest proof root, while other mangrove species were measured at DBH, approximately 130 cm above the ground (Abino et al., 2014; Asadi and Pambudi, 2020). Meanwhile, for Nypa fruticans, the stem diameter was measured for biomass measurement according to the model developed by Rahman et al. (2020). The model uses the average stem diameter and a base diameter of N. fruticans, and for the stem diameter, the correlation coefficient was 96.4%.
Data analysis
The analysis of mangrove vegetation was derived from the data of each individual mangrove stand of all mangrove species and its DBH. The data was used to determine the density, importance value index (IVI), and mangrove biomass and carbon. Mangrove density is calculated by dividing the number of mangrove trees by the area of the plot they occupy, measured in trees per hectare (Hanggara et al., 2021). A mangrove species’ basal area has a major impact on its IVI since it affects the calculation of the IVI’s relative dominance component (Sharma et al., 2023). IVI incorporates each species’ relative dominance, relative frequency, and relative density within the ecosystem. It provides a thorough overview of the ecological importance and structural function of many mangrove species, highlighting those that are most vital to the integrity of the ecosystem (Efriyeldi et al., 2023; Mahgoub, 2023). Meanwhile, the DBH of mangrove trees is a key indicator of carbon storage capacity, with bigger values generally indicating higher levels of carbon sequestered in the trees (Zaman et al., 2023; Zanvo et al., 2023). The IVI is measured by the sum of relative density (RDi), relative frequency (RFi), and relative dominance (RDo) (Asadi et al., 2018; Ismail et al., 2021).
In order to calculate mangrove biomass, common allometric equations developed by Komiyama et al. (2005) were used to estimate the above-ground biomass and below-ground biomass () of most mangrove species. For Sonneratia ovata, the allometric equation of above-ground biomass was according to Kusmana et al. (2018). Meanwhile, the mangrove biomass of Avicennia marina was calculated using the following allometric equation, , (Comley and McGuinness, 2005; Nam et al., 2016). The biomass calculation of Nypa fruticans was based on stem diameter, and the allometric equation was developed by Rahman et al. (2020). Furthermore, PlanetScope images using the Normalized Difference Vegetation Index (NDVI) were used to estimate the total area of mangroves and C-stocks of the whole Bawean Islands and Karimunjawa Islands.
Results
Structures of mangrove vegetation
A total of 680 and 339 mangrove trees were measured from the mangrove forest of Daun Village, Bawean Island, and Kemujan Island, respectively. Mangrove trees in the former area had an average DBH of 6.7 ± 2.92 cm and a total density of 2428,57 trees per ha. In contrast, the mangroves in the later area had much higher DBH and lower density, 11.75 ± 3.16 cm and 1614.29 trees per ha, respectively. Out of 13 mangrove species in Hijau Daun, Bawean Island, Nypa fruticans had the highest relative density (35.59%), while Rhizophora apiculata shared the highest relative dominance (28.95%). Therefore, both species had the highest IVI values, 62,67% and 66,02% for N. fruticans and R. apiculata, respectively.
Furthermore, in Kemujan Island, out of 12 mangrove species, Ceriops tagal, Excoecaria agallocha, Rhizophora apiculata, Xylocarpus granatum had the highest and same relative frequency values (12.94%). However, IVI value of E. agallocha stood over 70%, while C. tagal, R. apiculata, and X. granatum only had IVI values of 32.32%, 49,43%, and 28.88%, respectively. On the other hand, E. agallocha in Daun village, Bawean island had the lowest IVI value (1.71%). The ecological parameter values of each species are presented in Table 1.
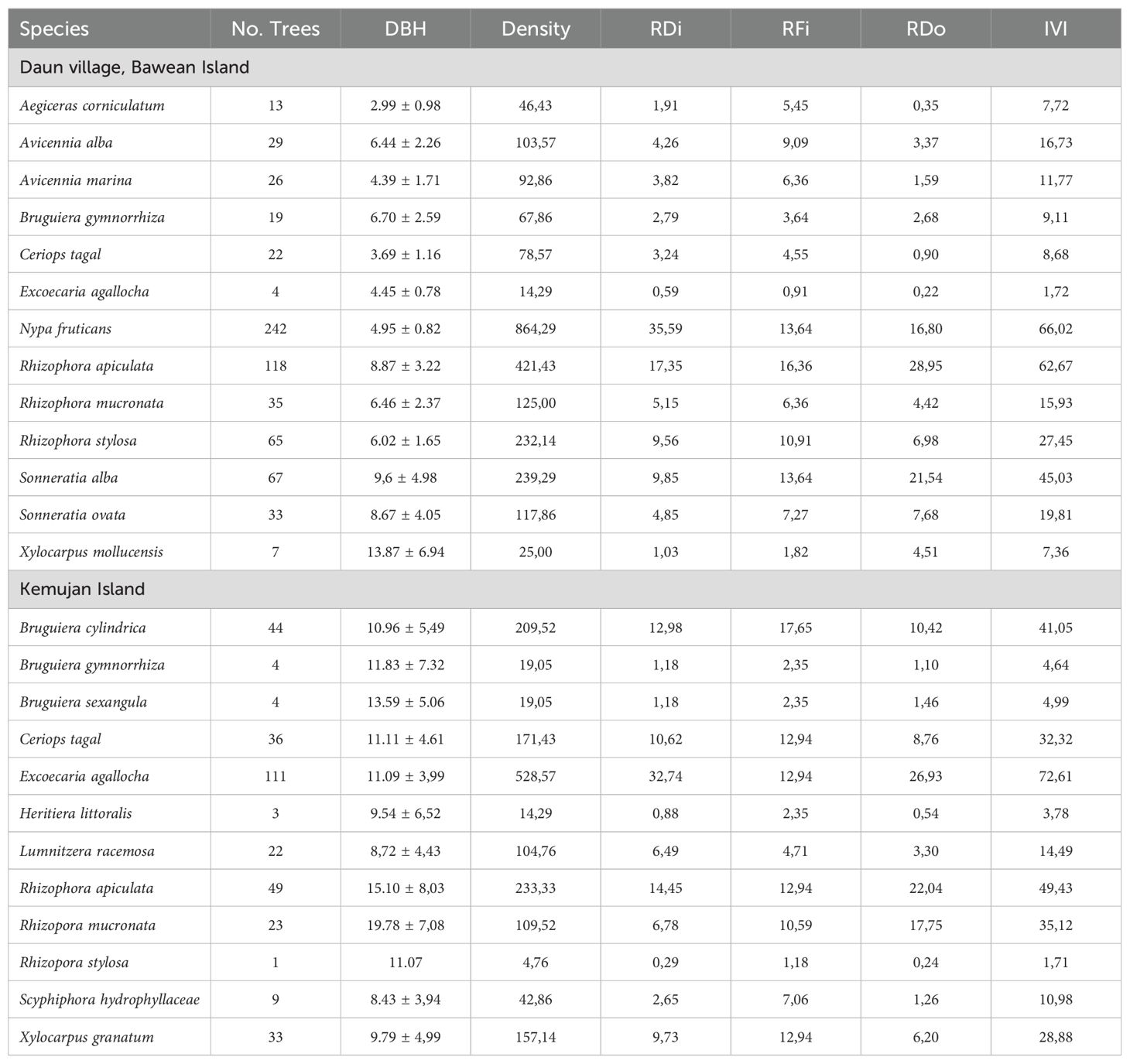
Table 1. Diameter breast height (DBH, cm), density (trees per ha), relative density (RDi, %), relative frequency (RFi, %), relative dominance (RDo, %), and importance value index (IVI, %) of mangroves from Daun Village, Bawean Island and Kemujan Island.
Biomass and C-stocks of mangroves
In the mangrove forest of Daun village, R. apiculata had the highest tree biomass and carbon (43.08 Mg B/ha and 19.19 Mg C/ha, respectively), followed by S. alba (22.38 Mg B/ha and 19.19 Mg C/2ha, respectively), and N. fruticans (15.45 Mg B/ha and 7.26 Mg C/2ha, respectively). A. corniculatum, E. agallocha, and C. tagal had the lowest tree biomass and carbon with values less than 1 Mg B/ha and 1 Mg C/ha, respectively. In total, the mangrove vegetation of Daun village contained biomass of 113.60 Mg B/ha and carbon of 50.98 Mg C/2ha (Figure 3).
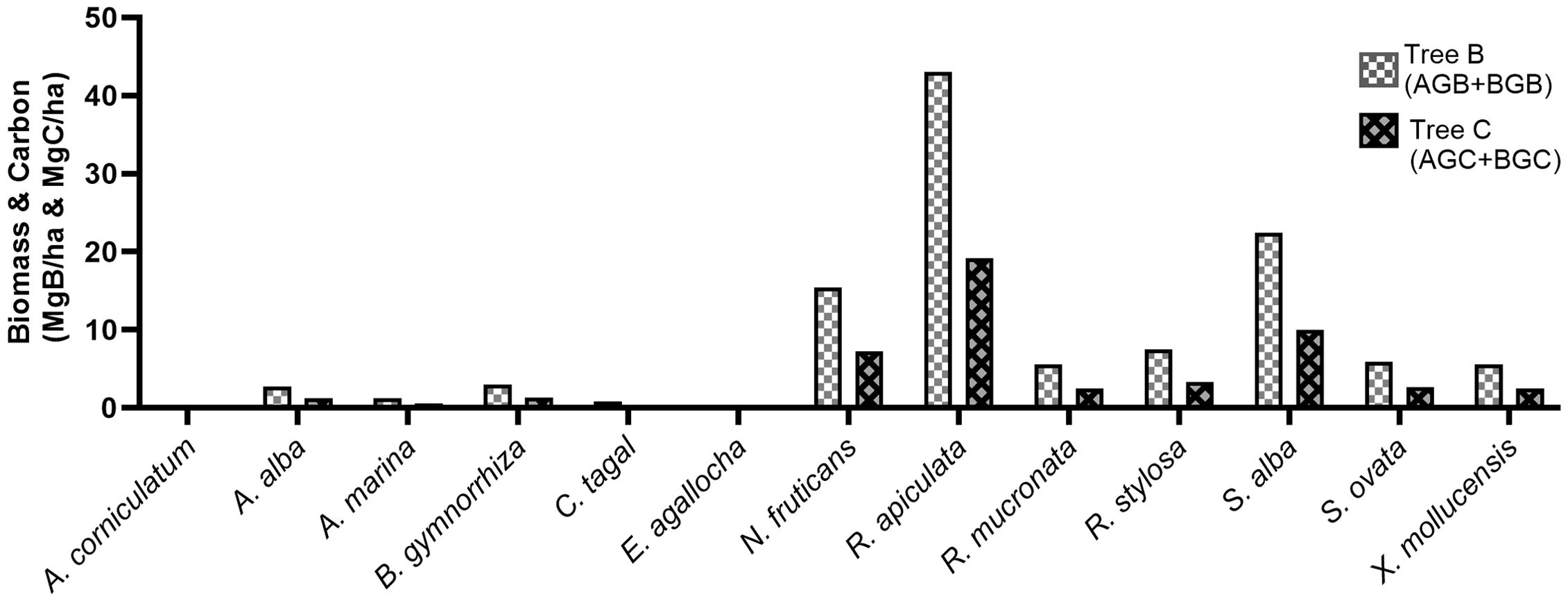
Figure 3. Tree biomass (AGB+BGB) and carbon of mangroves in the mangrove forest of Daun village, Bawean Island.
In Kemujan Island, R. apiculata also had the highest tree biomass and carbon (102.22 Mg B/ha and 45.82 Mg C/ha, respectively), while R. mucronata had the second largest tree biomass and carbon (67.02 Mg B/ha and 30.05 Mg C/ha, respectively). Mangrove with the highest density, E. agallocha, had the third largest tree biomass and carbon in the research area (30.93 Mg B/ha and 13.75 Mg C/ha, respectively). Other mangrove species with significant tree biomass were B. cylindrica, Ceriops tagal, and X. granatum (29.83 Mg B/ha, 26.83 Mg B/ha, and 16.21 Mg B/ha, respectively). Overall, Kemujan Island contained tree biomass and carbon of 295.04 Mg B/ha and 131.95 Mg C/ha, respectively (Figure 4).
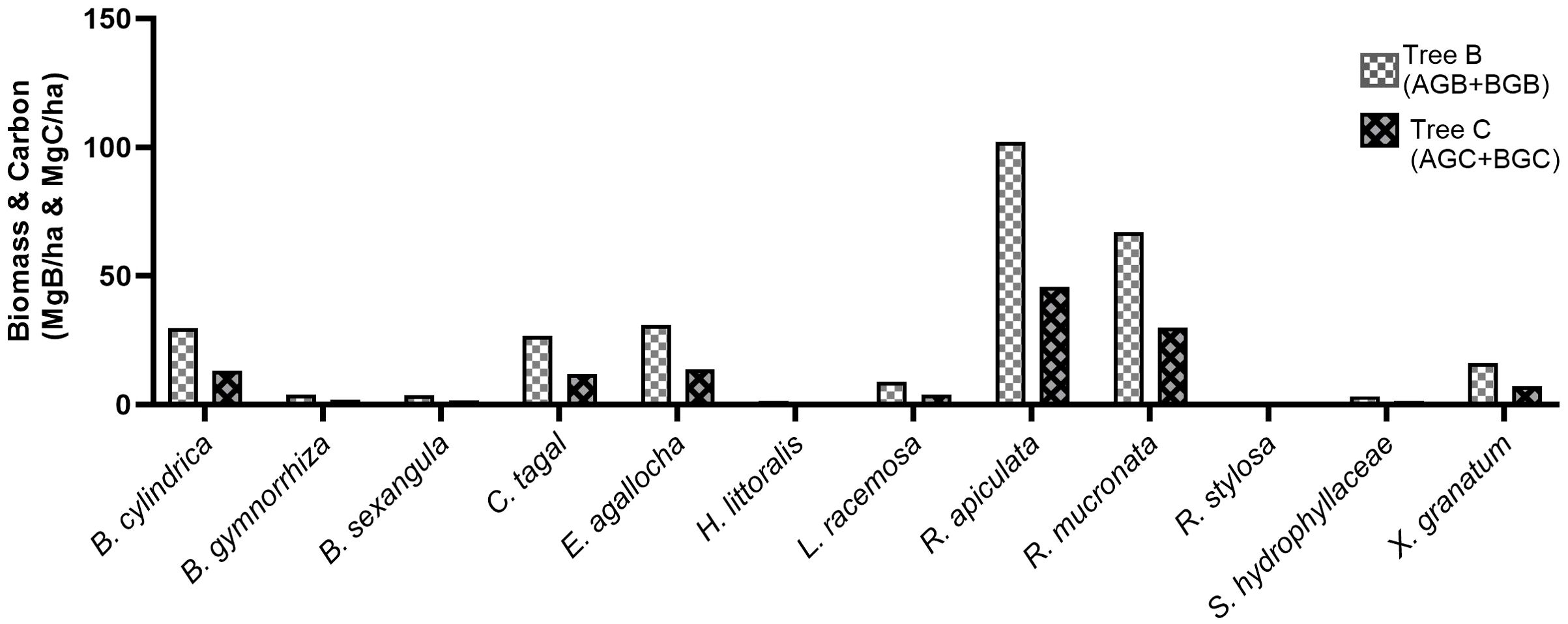
Figure 4. Tree biomass (AGB+BGB) and carbon of mangroves in Kemujan Island.
Correlation between IVI and biomass
In general, the linear regression model of mangrove biomass and IVI showed a stronger correlation in Desa Daun (R-squared = 0.72, p< 0.05) than in Kemujan Island (R-squared = 0.44, p< 0.05). Sequentially, mangrove species with the highest biomass in Desa Daun were R. apiculata (43.08 Mg B/ha), S. alba (22.38 Mg B/ha), and N. fruticans (15.45 Mg B/ha), whereas N. fruticans showed the highest IVI value (66.02%), followed by R. apiculata and S. alba (62.67% and 45.03%, respectively). Furthermore, on Kemujan Island, mangrove species with the highest IVI (72.61%) only had a biomass of 30.93 Mg B/ha. Whereas the species with the largest biomass, R. apiculata (102.22 Mg B/ha), only had an IVI of 49.43% (Figure 5).
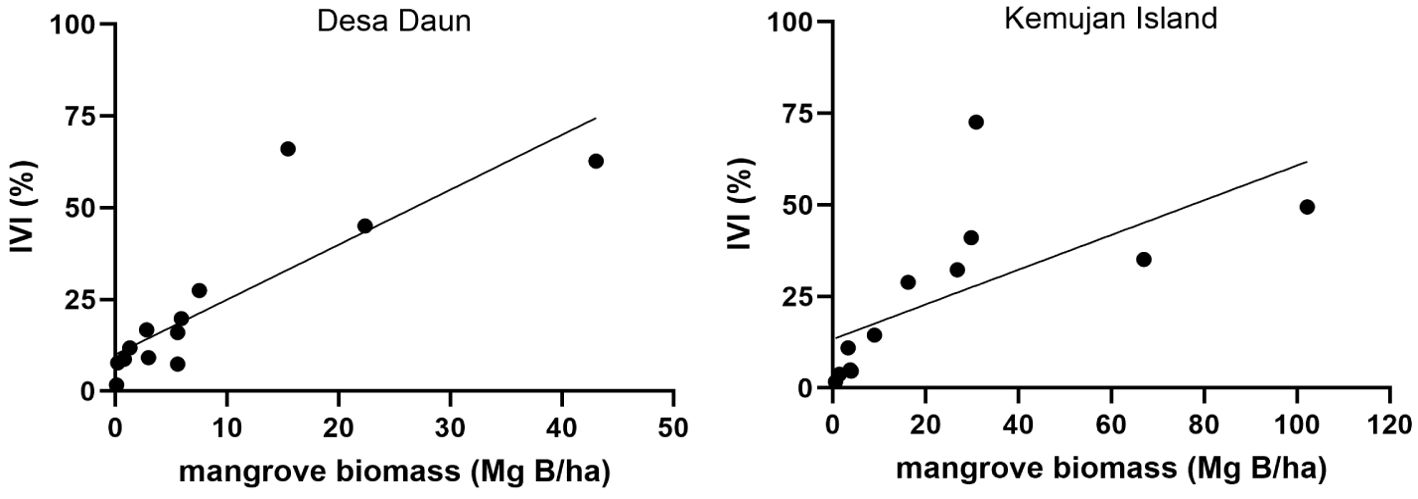
Figure 5. Pearson correlation between mangrove biomass (Mg B/ha) and importance value index (IVI) in Desa Daun (left) and Kemujan Island (right).
Discussion
Island characteristics and mangrove coverages
The Bawean Islands and Karimunjawa Islands are both located less than 100 miles north of the coast of Java Island, which formed as sea levels rose at the end of the last ice age. With a distance of around 250 km and different island characteristics, both islands have significantly different shoreline characteristics, which, therefore, influence their mangrove coverage landscapes. Based on Basemap RBI from the Geospatial Information Agency of Indonesia, the Bawean Islands (including some small satellite islands nearby) have a much larger total area (199.71 km2) than the Karimunjawa Islands (46.10 km2). However, the Karimunjawa Islands have a significantly longer total coastline than the Bawean Islands (130.77 km and 105.14 km, respectively). Most of Bawean Island is hilly, with some narrow coasts. Meanwhile, Karimunjawa Islands coasts are typically characterized by relatively low elevation and gentle slopes, and bays are notable features observed on Kemujan Island. Therefore, the coverage of mangrove in the whole Bawean Islands was significantly lower than that of the Karimunjawa Islands (450 ha and 602 ha, respectively).
Another chain of islands along the Java Sea is the Thousand Islands, which are located in the north of Jakarta Capital City. A significant portion of this region has been classified as a low-lying area, characterized by elevations varying from 0 to 31.88 meters, with a mean elevation of 3.9 meters above sea level. The island chain consists of 116 small islands and has a total land area of 8.7 km2 (Setiawati et al., 2023). Based on ArcGIS, the mangrove coverage of the whole thousand islands is 137 ha.
Structures of mangrove vegetation
The mangrove forests located in Desa Daun on Bawean Island and Kemujan Island exhibit a greater diversity of mangrove species compared to the majority of mangrove forests found on the mainland of Java. For instance, the mangrove ecosystem in Pasarbanggi, situated along the coastline of Rembang in Central Java, contains only five species, while the mangrove forest in Teluk Pangpang, Banyuwangi, East Java, comprises seven species (Hidayah et al., 2024; Soeprobowati et al., 2024). Additionally, the restored mangrove forests along the Karawang and Subang Coasts in West Java support eight species (Suwanto et al., 2021).Meanwhile, the mangrove forest at the Bengawan Solo Estuary in Ujungpangkah, Gresik Regency, is home to six species (Ali et al., 2021). Covering an area of 1,554 hectares, the Ujungpangkah mangrove forest is the largest in Gresik Regency and ranks among the largest mangrove forests in East Java (Saputra et al., 2022). In contrast, the mangrove forest in Desa Daun on Bawean Island, also situated in Gresik Regency, boasts remarkable diversity, with a total of 13 species within just 108 hectares of mangrove habitat.
In Desa Daun of the Bawean Islands, the highest percentage of measured mangrove was from the species Nypa fruticans (36%, 242 trees); therefore, it had the highest relative density and IVI (35.59% and 66.02%, respectively). N. fruticans is the only species of the genus Nypa and one of the most ancient extant mangrove species. Unlike most mangrove species that have aerating roots, this species relies on the leaf bases as a “giant pneumatophore” to supply oxygen to the underground tissues. This highly specialized phenotypic feature is valuable for the species for long-term plant adaptation mechanisms to harsh conditions (Wu et al., 2024). In Desa Daun mangrove forest, N. fruticans flourish along its estuary and always submerge under the water; therefore, such an adaptation mechanism is extremely important for the species to sustain.
The tall-stilt mangrove, R. apiculata, was among the dominant mangrove species in both locations, with relative high dominance values (28.95% and 22.04% in Desa Daun and Kemujan Island, respectively); therefore, R. apiculata was among the species with the highest IVI value. R. apiculata also dominated the mangrove landscape on the mainland of Java. In Labuhan mangrove forest, Lamongan, East Java, R. apiculata had 31.15% relative dominance and 80.46% IVI (Asadi et al., 2018). In North and South Andaman Islands, India, R. apiculata also had high IVI values of 83.83% and 106.01%, respectively (Kiruba-Sankar et al., 2018). This species is found throughout its continuous distribution range, which stretches from India to the western Pacific and northern Australia. Although it may grow in a variety of soil types, species of the genus Rhizophora prefer the fine mud sediments found in river estuaries downstream (Acharya et al., 2021; Guo et al., 2016; Kiruba-Sankar et al., 2018).
The most noticeable species composition in both locations was the abundance of E. agallocha. In Desa Daun, there were only a few trees of this species (4 individuals in total), whereas it was plentiful in stations 5, 6, and 7 of the Kemujan Islands (111 trees in total). The stations were located at higher elevations, back away from the sea, where salinity was lower. The species is often found in areas with a high input of freshwater or on the landward margins of mangrove forests. Therefore, it is also a common species in the Mekong River Delta, Vietnam (Besset et al., 2019). In contrast to the majority of mangrove species, it lacks the specialized aerial roots known as pneumatophores (Pi et al., 2009).
Mangrove forest biomass & correlation between IVI and biomass
Although Kemujan Island had a much lower mangrove density than that of the Desa Daun mangrove forest, the mangrove biomass of Kemujan Island was almost threefold higher than that of Desa Daun. It was mainly due to the fact that the average mangrove stand on Kemujan Island had almost double the DBH value. The measurement of DBH is critical in estimating carbon content, as allometric equations to estimate mangrove biomass are based on DBH measurements (Komiyama et al., 2005). Generally, there is a strong positive correlation between DBH and biomass. Mangrove trees tend to have higher biomass as they go broader (greater DBH). Bigger trees produce more biomass because they have a thicker stem and more leaves, branches, and roots.
In comparison with some other areas, the biomass and C stocks of Kemujan Island were higher than in Labuhan mangrove forest, Lamongan, East Java, in which the forest holds mangrove biomass of 168.05 Mg B/ha and carbon of 74.70 Mg C/ha (Asadi et al., 2018). The mangrove vegetation of Kerala, India, had almost equal stocks of biomass with the Desa Daun mangrove forest, in which the area held 117 Mg B/ha (Harishma et al., 2020). Meanwhile, mangrove stands in Baluran National Park had much higher biomass (533 Mg B/ha). In the park, there were some mangrove trees that had DBH over 50 cm, which in turn increased overall mangrove biomass in the area (Asadi and Pambudi, 2020).
In both locations, R. apiculata shared the highest biomass and carbon due to its higher number of mangrove stands and higher DBH values than most other mangrove species. In Labuhan mangrove forest, Lamongan, R. apiculata contributed almost half of the above-ground biomass in the forest (Asadi et al., 2018). In contrast, in Baluran National Park, R. apiculata had a much smaller contribution to overall mangrove biomass. R. stylosa and C. tagal had more stands and higher DBH values than R. apiculata in the park; therefore, they contributed more biomass (Asadi and Pambudi, 2020).
Furthermore, the importance value index (IVI) and biomass do not always have a very strong correlation. On Kemujan Island, the highest IVI value of E. agallocha was not correlated with the highest biomass of the species. In comparison with other mangrove species, E. agallocha has a significantly lower wood density (0.45 g/cm3) (Kauffman and Donato, 2012). Therefore, it contributes to lower mangrove biomass. However, in most mangrove forests, IVI is highly correlated with biomass. For example, in natural and planted mangroves in Sikka, East Nusa Tenggara, Indonesia, the positive correlations were very high (R-squared 0.92) (Matatula et al., 2021).
Carbon stocks of mangroves on islands along the Java Sea using PlanetScope images
Based on PlanetScope images acquired in July 2023, the mean carbon stocks of mangrove stands in the whole Bawean Islands were 53.76 Mg C/ha (Figure 6), which was slightly higher than the field data on Desa Daun in this study (50.98 Mg C/ha). It is indicated that mangrove forests in the Bawean Islands are better preserved, which might be due to the conservation effort from POKWASMAS Hijau Daun. Besides, a large percentage of Bawean people live at higher latitudes and work as farmers; therefore, its coastline and mangrove ecosystem have fewer anthropogenic impacts. With a total mangrove area of 449.64 ha, the Bawean Islands could store as much as 24,176.64 tons of mangrove stand carbon (Figure 6).
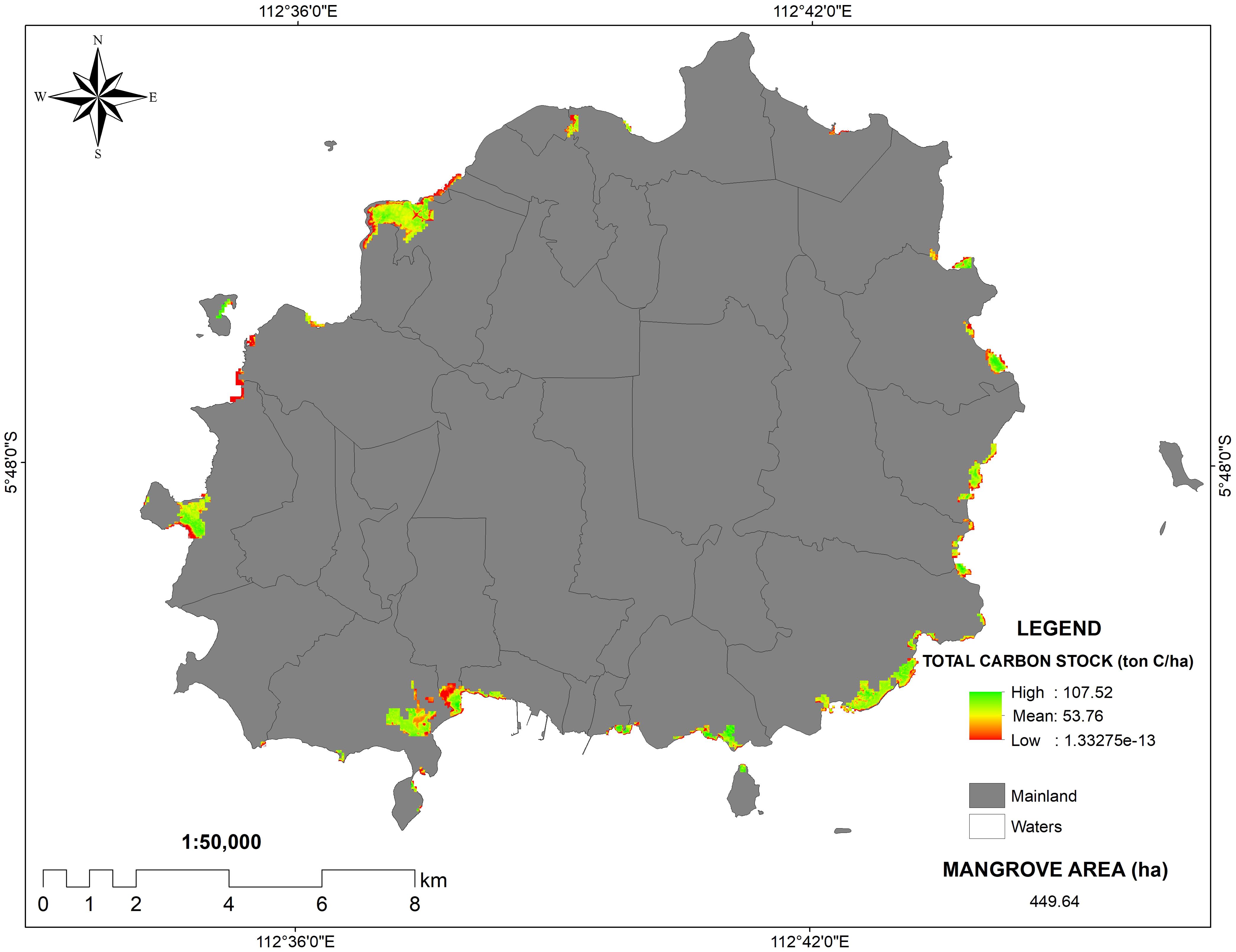
Figure 6. PlanetScope images of carbon stocks in Bawean Island and its surrounding satellite island.
On the contrary, despite the field data showing a higher carbon content on Kemujan Island (131.95 Mg C/ha), PlanetScope images revealed that the average C of the whole Karimunjawa Islands was 68.73 Mg C/ha. It might be due to the fact that field data was taken on Kemujan Island, which was shown to have a higher C-stock than the average of mangrove areas on Karimunjawa Islands. However, the expansion of shrimp farming aquaculture in recent years has led to the clearing of mangrove forests. The aquaculture expansion might be concentrated in the areas between Kemujan Island and Karimunjawa Island, which are shown to have lower C-stock concentrations based on the PlanetScope images. Despite the anthropogenic disruption, mangrove stands in the Karimunjawa Islands still hold a high C-stock. With a total area of 602.13 ha, they can store carbon as much as 41,384.39 tons (Figure 7).
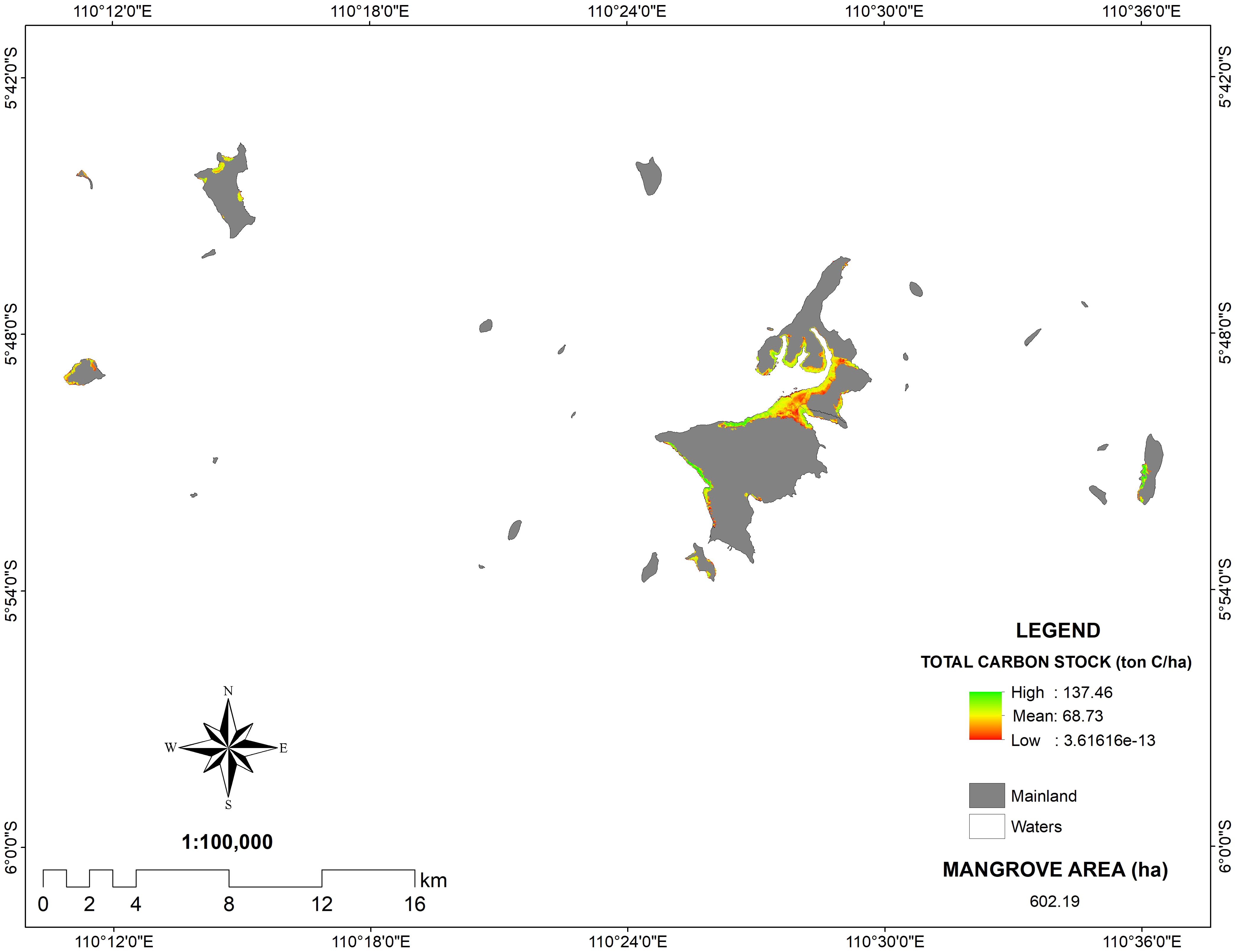
Figure 7. PlanetScope images of carbon stocks in Karimunjawa Islands.
Furthermore, the extensive mangrove ecosystems flourishing along the Java Sea are found in the Thousand Islands, situated 45 kilometers north of West Jakarta. According to PlanetScope imagery, the average carbon stocks of these islands amount to 66.66 Mg C/ha. With a total mangrove area of 137 hectares, the mangrove forests in this island chain sequester approximately 9,132.42 tons of carbon (Figure 8). Mangrove forests in the island chains along the Java Sea are critical for carbon sequestration, capturing and storing large amounts of carbon in their biomass. This process helps mitigate climate change by reducing atmospheric carbon dioxide levels. Conserving these ecosystems ensures the preservation of this carbon storage capacity while also protecting biodiversity and coastal resilience.
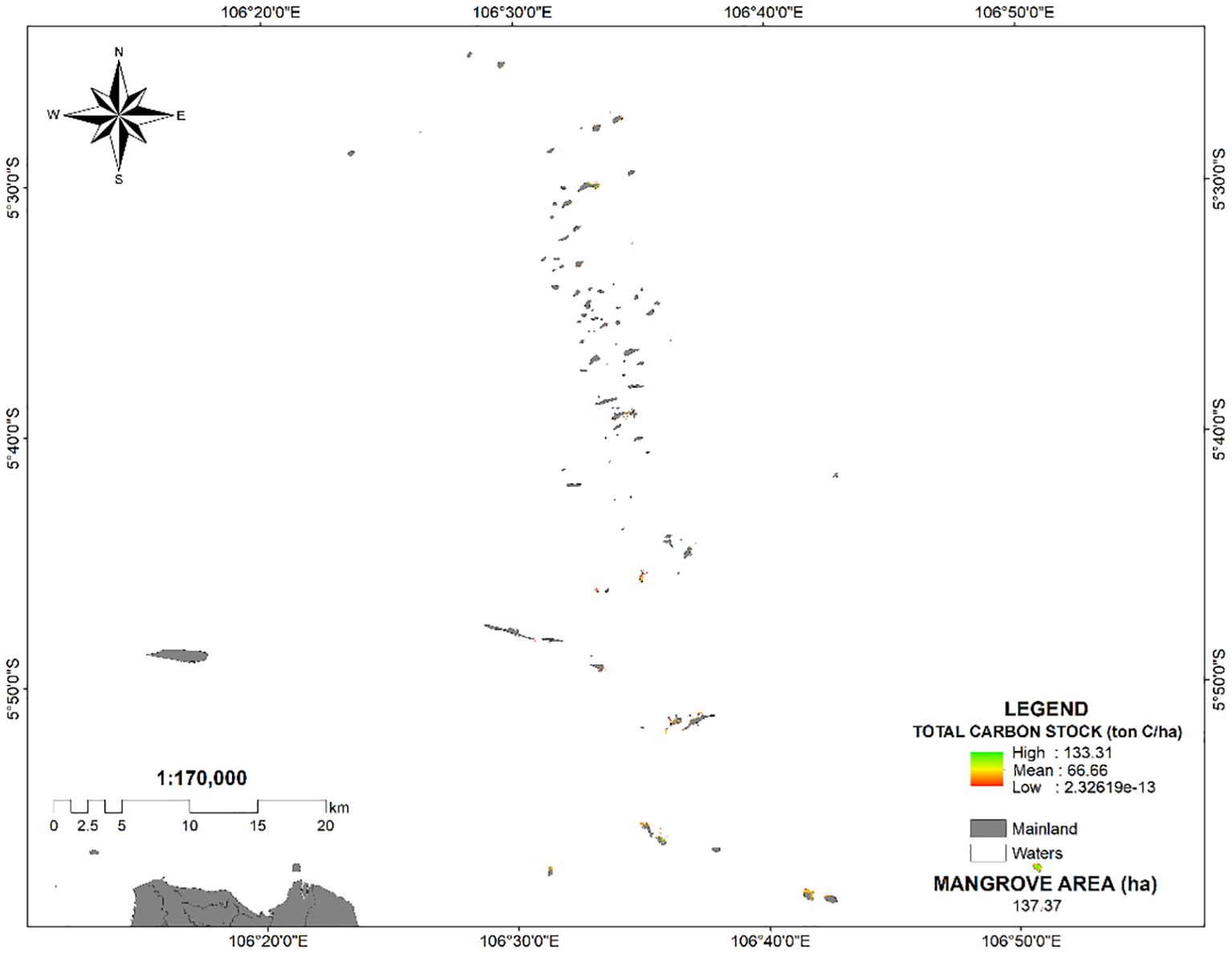
Figure 8. PlanetScope images of carbon stocks of The Thousand Islands.
Conclusion
In this study, we examine the biodiversity and carbon sequestration of two major islands along the Java Sea, the Karimunjawa Islands and the Bawean Islands. PlanetScope imagery was also used to calculate mangrove carbon on both islands and the whole island chain along the Java Sea. Both field data and the PlanetScope image revealed that the mangrove forests of the Karimunjawa Islands hold higher C-stocks than those of the Bawean Islands, despite the later islands having higher mangrove densities. The lower DBH and higher dominance of mangroves with lower wood densities resulted in lower C-stocks of mangroves in the Bawean Islands. Furthermore, the higher biomass of R. apiculata is associated with the higher role of the species in storing carbon in both areas. Research on mangrove structure and biomass on islands along the Java Sea can inform targeted conservation strategies, enhance biodiversity preservation, optimize carbon sequestration, and improve sustainable resource management, thereby strengthening the ecology of these islands.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
MA: Data curation, Formal analysis, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing. AA: Data curation, Writing – original draft, Writing – review & editing. RA: Data curation, Writing – original draft, Writing – review & editing. AS: Writing – original draft, Writing – review & editing. AY: Validation, Writing – original draft, Writing – review & editing. DS: Conceptualization, Methodology, Writing – original draft, Writing – review & editing. AR: Project administration, Software, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and authorship. Publication of this article was financially supported by CIFOR-ICRAF.
Acknowledgments
We would like to acknowledge the financial support from CIFOR-ICRAF. The authors also thank POKWASMAS Hijau Daun, Bawean, for facilitating research in Desa Daun, Bawean Island. Additionally, we extend our appreciation to Balai Taman Nasional Karimunjawa (BTNKJ) for providing us with a with a research permit. A special thank you to Bapak Kuswadi from BTNKJ SPTN 1 Kemujan Island for valuable guidelines during field data collection.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abino A. C., Castillo J. A. A., Lee Y. J. (2014). Assessment of species diversity, biomass and carbon sequestration potential of a natural mangrove stand in Samar, the Philippines. For. Sci. Technol. 10, 2–8. doi: 10.1080/21580103.2013.814593
Acharya S., Patra D. K., Mahalik G., Mohapatra P. K. (2021). Quantitative ecological study of rhizophoraceae mangroves of bhitarkanika wildlife sanctuary regions of odisha coast, India. Proc. Natl. Acad. Sci. India. Section. B.: Biol. Sci. 91, 897–908. doi: 10.1007/s40011-021-01295-2
Ali M., Sulistiono S., Imran Z., Simanjuntak C. (2021). The potential development of ecotourism based on mangrove ecosystem in Ujung Pangkah of Gresik Regency, East Java Province, Indonesia. IOP. Conf. Series.: Earth Environ. Sci. 800, 12054. doi: 10.1088/1755-1315/800/1/012054
Asadi M. A., Pambudi G. S. (2020). Diversity and biomass of mangrove forest within Baluran National Park, Indonesia. Aquacult. Aquarium. Conserv. Legislation. 13, 19–27.
Asadi M. A., Rahardani A., Semedi B., Soegianto A. (2019). Carbon storage of mangrove ecosystems in Pasuruan and Probolinggo Regency, East Java, Indonesia. Ecol. Environ. Conserv. 25, S162–S167. doi: 10.13140/RG.2.2.12317.67045
Asadi M. A., Yona D., Saputro S. E. (2018). Species diversity, biomass, and carbon stock assessments of mangrove forest in labuhan, Indonesia. IOP. Conf. Series.: Earth Environ. Sci. 151, 12009. doi: 10.1088/1755-1315/151/1/012009
Asy’Ari R., Rahmawati A., Sa’diyya N., Gustawan A., Setiawan Y., Zamani N., et al. (2022). Mapping mangrove forest distribution on Banten, Jakarta, and West Java Ecotone Zone from Sentinel-2-derived indices using cloud computing based Random Forest. Jurnal. Pengelolaan. Sumberdaya. Alam. Dan. Lingkungan. (Journal. Natural Resour. Environ. Management). 12, 97–111. doi: 10.29244/jpsl.12.1.97-111
Basyuni M., Sasmito S. D., Analuddin K., Ulqodry T. Z., Saragi-Sasmito M. F., Eddy S., et al. (2022). “Mangrove biodiversity, conservation and roles for livelihoods in Indonesia,” in Mangroves: Biodiversity, Livelihoods and Conservation. Eds. Das S. C., Pullaiah, Ashton E. C. (Singapore: Springer Nature Singapore), 397–445. doi: 10.1007/978-981-19-0519-3_16
Besset M., Gratiot N., Anthony E. J., Bouchette F., Goichot M., Marchesiello P. (2019). Mangroves and shoreline erosion in the Mekong River delta, Viet Nam. Estuarine. Coast. Shelf. Sci. 226, 106263. doi: 10.1016/j.ecss.2019.106263
Canty S. W. J., Kennedy J. P., Fox G., Matterson K., González V. L., Núñez-Vallecillo M. L., et al. (2022). Mangrove diversity is more than fringe deep. Sci. Rep. 12, 1695. doi: 10.1038/s41598-022-05847-y
Comley B. W. T., McGuinness K. A. (2005). Above- and below-ground biomass, and allometry, of four common northern Australian mangroves. Aust. J. Bot. 53, 431–436. doi: 10.1071/BT04162
Efriyeldi E., Syahrial S., Effendi I., Almanar I. P., Syakti A. D. (2023). The mangrove ecosystem in a harbor-impacted city in Dumai, Indonesia: A conservation status. Regional. Stud. Mar. Sci. 65, 103092. doi: 10.1016/j.rsma.2023.103092
Giri C., Ochieng E., Tieszen L. L., Zhu Z., Singh A., Loveland T., et al. (2011). Status and distribution of mangrove forests of the world using earth observation satellite data. Global Ecol. Biogeogr. 20, 154–159. doi: 10.1111/j.1466-8238.2010.00584.x
Guo Z., Huang Y., Chen Y., Duke N. C., Zhong C., Shi S. (2016). Genetic discontinuities in a dominant mangrove Rhizophora apiculata (Rhizophoraceae) in the Indo-Malesian region. J. Biogeogr. 43, 1856–1868. doi: 10.1111/jbi.12770
Hagger V., Worthington T. A., Lovelock C. E., Adame M. F., Amano T., Brown B. M., et al. (2022). Drivers of global mangrove loss and gain in social-ecological systems. Nat. Commun. 13, 6373. doi: 10.1038/s41467-022-33962-x
Hanggara B. B., Murdiyarso D., Ginting Y., Widha Y. L., Panjaitan G. Y., Lubis A. A. (2021). Effects of diverse mangrove management practices on forest structure, carbon dynamics and sedimentation in North Sumatra, Indonesia. Estuarine. Coast. Shelf. Sci. 259, 107467. doi: 10.1016/j.ecss.2021.107467
Harishma K. M., Sandeep S., Sreekumar V. B. (2020). Biomass and carbon stocks in mangrove ecosystems of Kerala, southwest coast of India. Ecol. Processes. 9, 31. doi: 10.1186/s13717-020-00227-8
Hayes M. A., Jesse A., Welti N., Tabet B., Lockington D., Lovelock C. E. (2019). Groundwater enhances above-ground growth in mangroves. J. Ecol. 107, 1120–1128. doi: 10.1111/1365-2745.13105
Hidayah Z., Rachman H. A., Wiyanto D. B. (2024). Assessment of spatio-temporal dynamics of mangrove forest in Teluk Pangpang, Banyuwangi, East Java, Indonesia. Biodivers. J. Biol. Diversity 25, 3138–3150. doi: 10.13057/biodiv/d250736
Ismail, Sulistiono S., Hariyadi S., Madduppa H. (2021). “Diversity, density, and Importance Value Index of mangroves in the Segara Anakan lagoon and its surrounding area, Cilacap Regency, Indonesia,” in IOP Conference Series Earth and Environmental Science, Vol. 744. doi: 10.1088/1755-1315/744/1/012034
Kandasamy K., Rajendran N., Balakrishnan B., Thiruganasambandam R., Narayanasamy R. (2021). Carbon sequestration and storage in planted mangrove stands of Avicennia marina. Regional. Stud. Mar. Sci. 43, 101701. doi: 10.1016/j.rsma.2021.101701
Kauffman J., Donato D. (2012). Protocols for the measurement, monitoring and reporting of structure, biomass and carbon stocks in mangrove forests (Bogor, Indonesia: CIFOR).
Kiruba-Sankar R., Krishnan P., Dam Roy S., Raymond Jani Angel J., Goutham-Bharathi M. P., Lohith Kumar K., et al. (2018). Structural complexity and tree species composition of mangrove forests of the Andaman Islands, India. J. Coast. Conserv. 22, 217–234. doi: 10.1007/s11852-017-0588-3
Komiyama A., Poungparn S., Kato S. (2005). Common allometric equations for estimating the tree weight of mangroves. J. Trop. Ecol. 21, 471–477. doi: 10.1017/S0266467405002476
Kusmana C., Hidayat T., Tiryana T., Rusdiana O., Istomo (2018). Allometric models for above- and below-ground biomass of Sonneratia spp. Global Ecol. Conserv. 15, e00417. doi: 10.1016/j.gecco.2018.e00417
Lovelock C. E., Barbier E., Duarte C. M. (2022). Tackling the mangrove restoration challenge. PloS Biol. 20, e3001836. doi: 10.1371/journal.pbio.3001836
Mahgoub A. M. M. A. (2023). Ecological importance and sociological characters of fourteen widespread weeds and their contribution to the observed (dis)similarity between weed communities. Heliyon 9, e13638. doi: 10.1016/j.heliyon.2023.e13638
Matatula J., Afandi A., Wirabuana P. (2021). A comparison of stand structure, species diversity and aboveground biomass between natural and planted mangroves in Sikka, East Nusa Tenggara, Indonesia. Biodivers. J. Biol. Diversity 22, 1098–1103. doi: 10.13057/biodiv/d220303
Moity N., Delgado B., Salinas-de-León P. (2019). Mangroves in the Galapagos islands: Distribution and dynamics. PloS One 14, e0209313. doi: 10.1371/journal.pone.0209313
Nam V. N., Sasmito S. D., Murdiyarso D., Purbopuspito J., MacKenzie R. A. (2016). Carbon stocks in artificially and naturally regenerated mangrove ecosystems in the Mekong Delta. Wetlands. Ecol. Manage. 24, 231–244. doi: 10.1007/s11273-015-9479-2
Nguyen H., Chu L., Harper R. J., Dell B., Hoang H. (2022). Mangrove-shrimp farming: A triple-win approach for communities in the Mekong River Delta. Ocean. Coast. Manage. 221, 106082. doi: 10.1016/j.ocecoaman.2022.106082
Nozarpour R., Shojaei M. G., Naderloo R., Nasi F. (2023). Crustaceans functional diversity in mangroves and adjacent mudflats of the Persian Gulf and Gulf of Oman. Mar. Environ. Res. 186, 105919. doi: 10.1016/j.marenvres.2023.105919
Nyanga C. (2020). “The role of mangroves forests in decarbonizing the atmosphere,” in Carbon-Based Material for Environmental Protection and Remediation (p. Ch. 9). Eds. Bartoli M., Frediani M., Rosi L. (London, UK: IntechOpen). doi: 10.5772/intechopen.92249
Pi N., Tam N. F. Y., Wu Y., Wong M. H. (2009). Root anatomy and spatial pattern of radial oxygen loss of eight true mangrove species. Aquat. Bot. 90, 222–230. doi: 10.1016/j.aquabot.2008.10.002
Prihantono J., Nakamura T., Nadaoka K., Wirasatriya A., Adi N. S. (2022). Rainfall variability and tidal inundation influences on mangrove greenness in karimunjawa national park, Indonesia. Sustainability 14 (14), 8948. doi: 10.3390/su14148948
Rahman R., Effendi H., Rusmana I., Ali M. (2020). Metode Pengukuran dan Model Pendugaan Biomassa Nypa Fruticans di Sungai Tallo, Makassar-Indonesia. Grouper 11, 25. doi: 10.30736/grouper.v11i1.65
Ramarokoto S., Raharijaona L. J. R., Ravonjimalala R. H., Randriamalala J. R. (2024). Relative importance of local and regional/global drivers of mangrove degradation and deforestation in Madagascar. Regional. Environ. Change 24, 84. doi: 10.1007/s10113-024-02247-y
Ruslan N. F. N., Goh H. C., Hattam C., Edwards-Jones A., Moh H. H. (2022). Mangrove ecosystem services: Contribution to the well-being of the coastal communities in Klang Islands. Mar. Policy 144, 105222. doi: 10.1016/j.marpol.2022.105222
Saputra D. K., Semedi B., Yamindago A., Dewi C. S. U., Asadi M. A., Isdianto A., et al. (2022). Characteristics of mangrove fisheries in essential ecosystem area ujungpangkah, Indonesia. J. Environ. Manage. Tourism. 13 (59), 812–820. doi: 10.14505/Jemt.V13.3(59).20. Vol 13 No 3 (2022): JEMT, Volume XIII, Issue 3(59), Summer 2022DO.
Sasmito S. D., Basyuni M., Kridalaksana A., Saragi-Sasmito M. F., Lovelock C. E., Murdiyarso D. (2023). Challenges and opportunities for achieving Sustainable Development Goals through restoration of Indonesia’s mangroves. Nat. Ecol. Evol. 7, 62–70. doi: 10.1038/s41559-022-01926-5
Sasmito S. D., Taillardat P., Clendenning J. N., Cameron C., Friess D. A., Murdiyarso D., et al. (2019). Effect of land-use and land-cover change on mangrove blue carbon: A systematic review. Global Change Biol. 25, 4291–4302. doi: 10.1111/gcb.14774
Sealey K. S. (2024). Assessing coastal vulnerability and climate resilience for Caribbean small island states using ecological principles. Int. J. Disaster. Risk Reduction. 105, 104410. doi: 10.1016/j.ijdrr.2024.104410
Setiawati M. D., Chatterjee U., Djamil Y. S., Alifatri L. O., Nandika M. R., Rachman H. A., et al. (2023). Seribu islands in the megacities of Jakarta on the frontlines of the climate crisis. Front. Environ. Sci. 11. doi: 10.3389/fenvs.2023.1280268
Sharma A., Patel S. K., Singh G. S. (2023). Variation in species composition, structural diversity, and regeneration along disturbances in tropical dry forest of northern India. J. Asia-Pac. Biodivers. 16, 83–95. doi: 10.1016/j.japb.2022.11.004
Slamet N. S., Dargusch P., Aziz A. A., Wadley D. (2020). Mangrove vulnerability and potential carbon stock loss from land reclamation in Jakarta Bay, Indonesia. Ocean. Coast. Manage. 195, 105283. doi: 10.1016/j.ocecoaman.2020.105283
Soeprobowati T. R., Sularto R. B., Hadiyanto H., Puryono S., Rahim A., Jumari J., et al. (2024). The carbon stock potential of the restored mangrove ecosystem of Pasarbanggi, Rembang, Central Java. Mar. Environ. Res. 193, 106257. doi: 10.1016/j.marenvres.2023.106257
Suwanto A., Takarina N., Koestoer R. H., Frimawaty E. (2021). Diversity, biomass, covers, and NDVI of restored mangrove forests in Karawang and Subang Coasts, West Java, Indonesia: Restored mangrove biomass and diversity. Biodivers. J. Biol. Diversity 22, 4115–4122. doi: 10.13057/biodiv/d220960
Trégarot E., Caillaud A., Cornet C. C., Taureau F., Catry T., Cragg S. M., et al. (2021). Mangrove ecological services at the forefront of coastal change in the French overseas territories. Sci. Total. Environ. 763, 143004. doi: 10.1016/j.scitotenv.2020.143004
Trissanti V. N., Amalo L. F., Handayani L. D. W., Nugroho D., Yuliani A. R., Mulyana D. (2022). The estimation of biomass and carbon stocks in mangrove forest ecosystem of Karawang Regency, West Java. IOP. Conf. Series.: Earth Environ. Sci. 1109, 12099. doi: 10.1088/1755-1315/1109/1/012099
Veitayaki J., Waqalevu V., Varea R., Rollings N. (2017). “Mangroves in small island development states in the pacific: an overview of a highly important and seriously threatened resource,” in Participatory Mangrove Management in a Changing Climate: Perspectives from the Asia-Pacific. Eds. DasGupta R., Shaw R. (Japan: Springer Japan), 303–327. doi: 10.1007/978-4-431-56481-2_19
Visschers L. L. B., Santos C. D., Franco A. M. A. (2022). Accelerated migration of mangroves indicate large-scale saltwater intrusion in Amazon coastal wetlands. Sci. Total. Environ. 836, 155679. doi: 10.1016/j.scitotenv.2022.155679
Wang J., Wang Y., Zhang Z., Wang W., Jiang L. (2023). Enhanced awareness of height-diameter allometry in response to climate, soil, and competition in secondary forests. For. Ecol. Manage. 548, 121386. doi: 10.1016/j.foreco.2023.121386
Wu W., Feng X., Wang N., Shao S., Liu M., Si F., et al. (2024). Genomic analysis of Nypa fruticans elucidates its intertidal adaptations and early palm evolution. J. Integr. Plant Biol. 66, 824–843. doi: 10.1111/jipb.13625
Yamamoto Y. (2023). Living under ecosystem degradation: Evidence from the mangrove–fishery linkage in Indonesia. J. Environ. Econ. Manage. 118, 102788. doi: 10.1016/j.jeem.2023.102788
Zaman M., Rahman M., Ahmed S., Zuidema P. A. (2023). What drives carbon stocks in a mangrove forest? The role of stand structure, species diversity and functional traits. Estuarine. Coast. Shelf. Sci. 295, 108556. doi: 10.1016/j.ecss.2023.108556
Keywords: Bawean Islands, blue carbon, Excoecaria agallocha, Karimunjawa Islands, mangrove structure and biomass, Rhizophora apiculata
Citation: Asadi MA, Al-kareem AS, Aprilianto RY, Sartimbul A, Yamindago A, Saputra DK and Riyadi A (2024) Assessment of mangrove structures and biomass on islands along the Java Sea: a case study on Bawean Islands and Karimunjawa Islands. Front. Ecol. Evol. 12:1422749. doi: 10.3389/fevo.2024.1422749
Received: 24 April 2024; Accepted: 28 August 2024;
Published: 17 September 2024.
Edited by:
Mahmoud El-Sharkawy, Tanta University, EgyptReviewed by:
Adrian Damora, Syiah Kuala University, IndonesiaAaron Froilan Raganas, University of the Philippines Mindanao, Philippines
Copyright © 2024 Asadi, Al-kareem, Aprilianto, Sartimbul, Yamindago, Saputra and Riyadi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Muhammad Arif Asadi, YXNhZGlAdWIuYWMuaWQ=