
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
CORRECTION article
Front. Ecol. Evol. , 30 January 2024
Sec. Conservation and Restoration Ecology
Volume 12 - 2024 | https://doi.org/10.3389/fevo.2024.1357830
 Stuart H. Munsch1*
Stuart H. Munsch1* Mike McHenry2
Mike McHenry2 Martin C. Liermann3Todd R. Bennett3John McMillan4Raymond Moses2
Martin C. Liermann3Todd R. Bennett3John McMillan4Raymond Moses2 George R. Pess3
George R. Pess3A Corrigendum on
Dam removal enables diverse juvenile life histories to emerge in threatened salmonids repopulating a heterogeneous landscape
by Munsch SH, McHenry M, Liermann MC, Bennett TR, McMillan J, Moses R and Pess GR (2023) Front. Ecol. Evol. 11:1188921. doi: 10.3389/fevo.2023.1188921
When calculating smolt abundance estimates from rotary screw trap data, an error in one of the analysis steps resulted in incorrect values. While this error affected most of the abundance estimates to some degree, the patterns described in the manuscript remain largely unchanged with the exception of steelhead smolts. None of the size data was affected.
The error in the values was introduced in a processing step when calculating daily out-migrants based on catch and efficiency trial data. For a small proportion of days, traps were fishing but not checked. We used an R function to redistribute the accumulated fish observed on the first day that checking was re-started. So, for example, if a trap was not checked on day 4 or 5, but then checked on day 6, the function would assign 1/3rd of the observed catch on day 6 to each of days 4, 5, and 6. We inadvertently applied this function to multiple species at one time (an array instead of a vector), which resulted in copying catches from one species to another in a haphazard way. This had minimal effect on the more abundant species, but dramatically changed estimates for steelhead.
Additionally, since the corrected data indicate significantly more steelhead in Indian Creek than Little River, we have added a paragraph to the Discussion to elaborate on this result.
The thesis of this article is that salmonid life history diversity is emerging following dam removal from two tributaries with different environments – the tributaries’ salmonid species, age structures, growth regimes, and outmigration timings appear to be distinct. Correcting our abundance estimates – largely affecting only steelhead – does not alter this original finding. For this reason, we state that the errors do not change the scientific conclusions of the article.
In the published article, there were errors in Figures 2–6 and 8 as published. We reported incorrect fish counts. The corrected Figures 2–6 and 8 appear below.
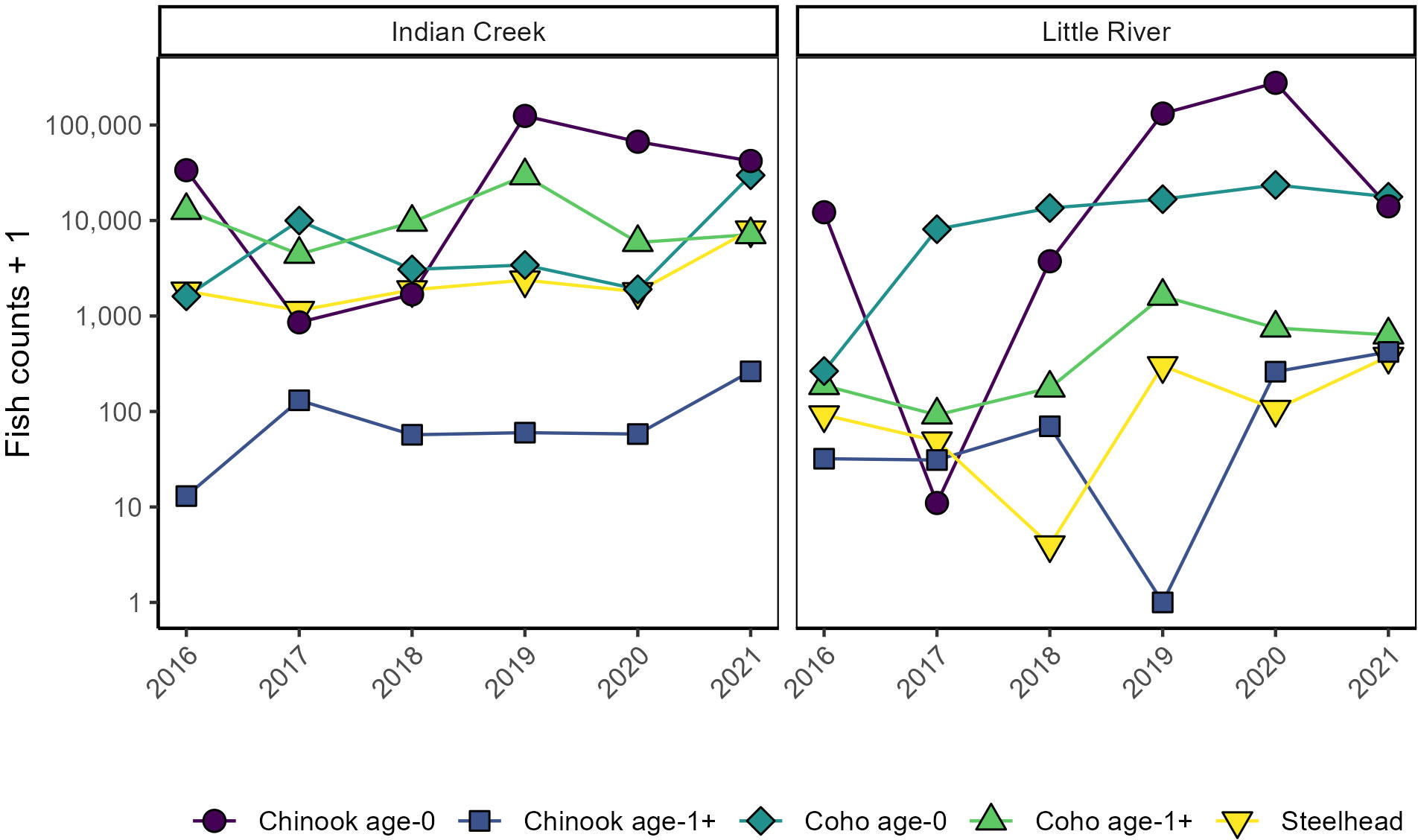
Figure 2 Assemblage composition compared between tributaries and among years. Y axes are log transformed to improve visibility of smaller counts.
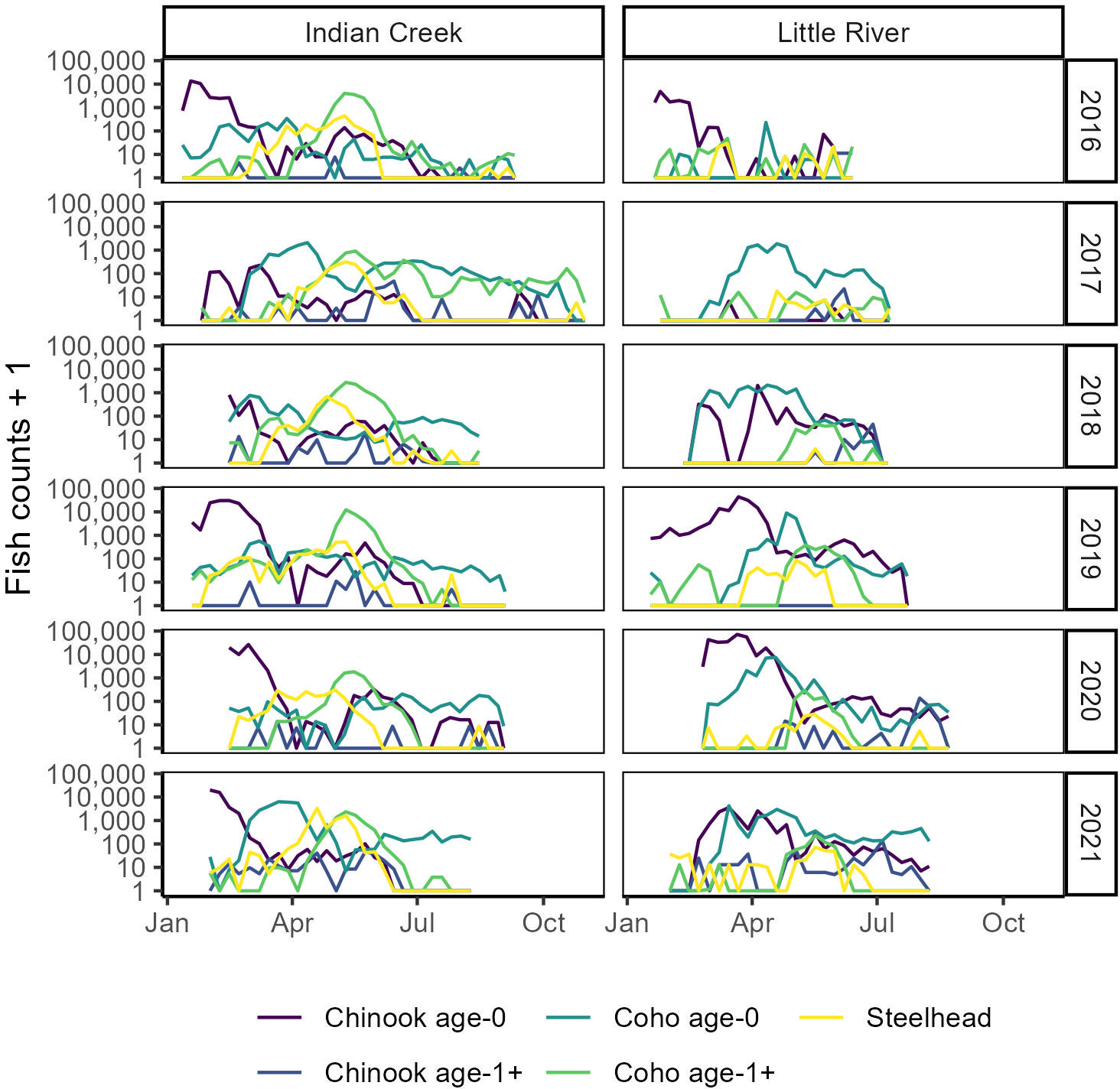
Figure 3 Timing and assemblage composition compared between tributaries and among years. Within-year counts are summed by week. Y axes are log transformed to improve visibility of smaller counts.
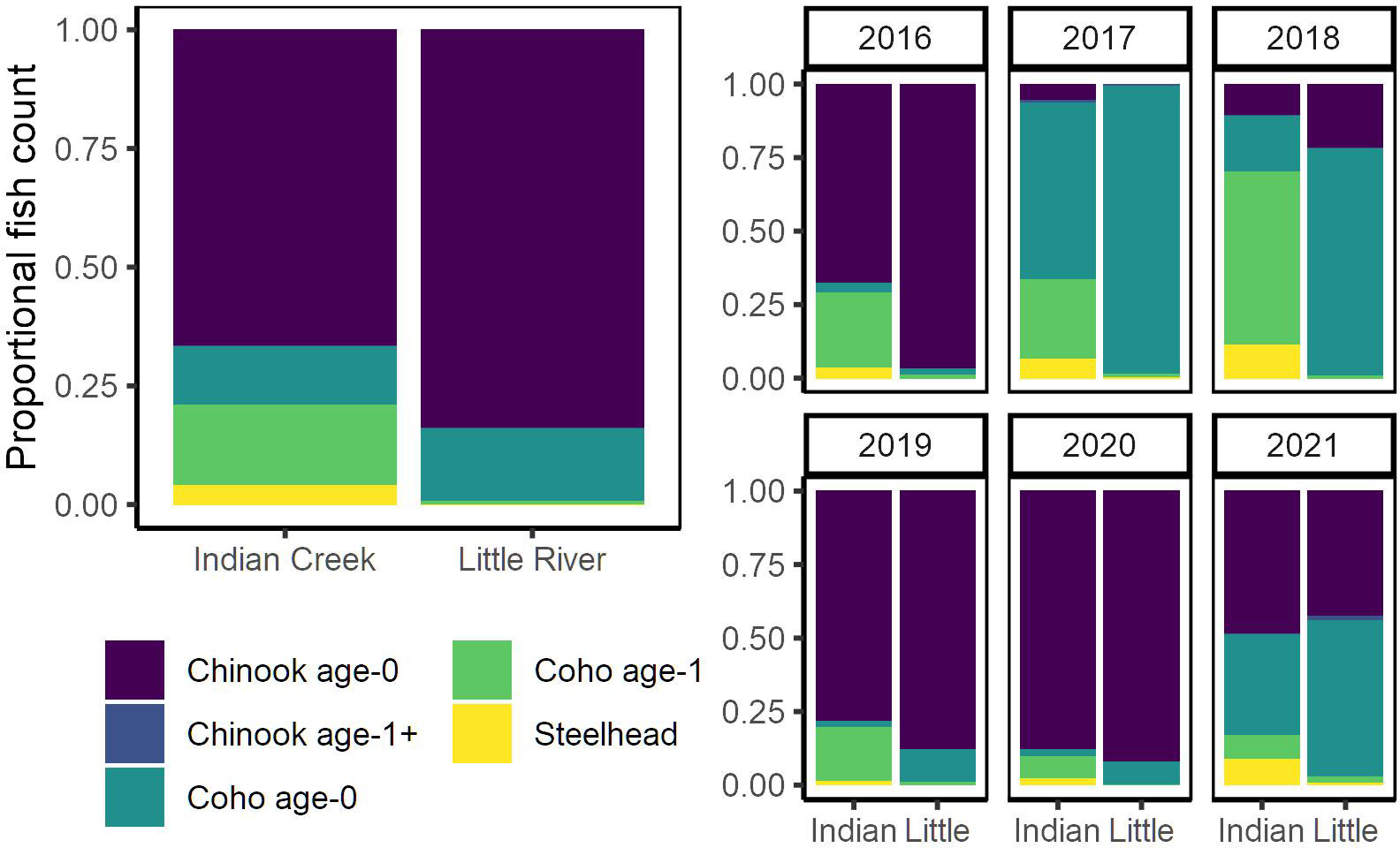
Figure 4 Proportional composition of salmonid assemblage compared between Indian Creek and Little River for all years (left) and individual years (right).
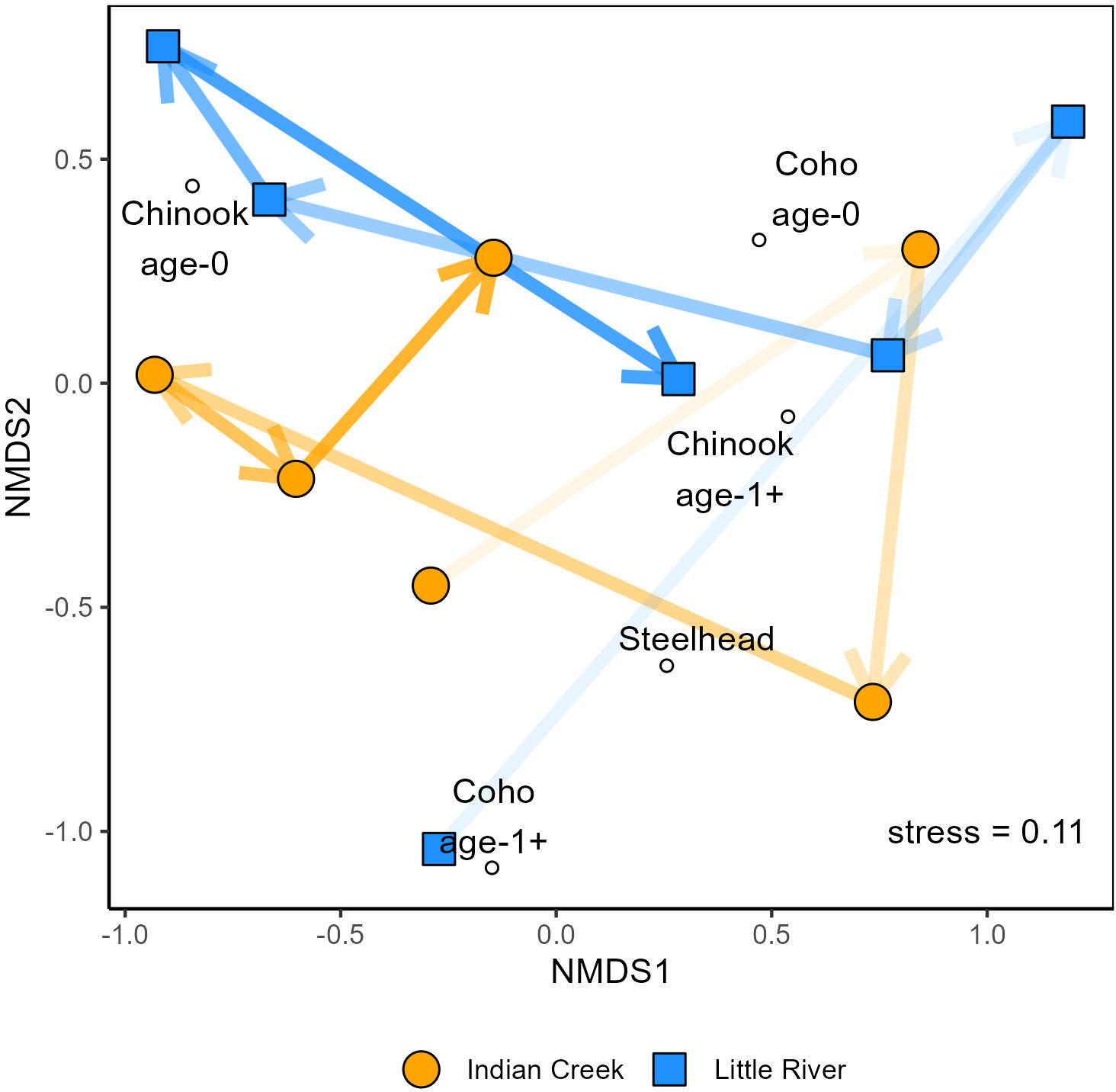
Figure 5 NMDS comparing salmonid assemblages between Indian Creek and Little River. Lines and arrows track the tributaries’ changes from 2016–2021 and darken across time.
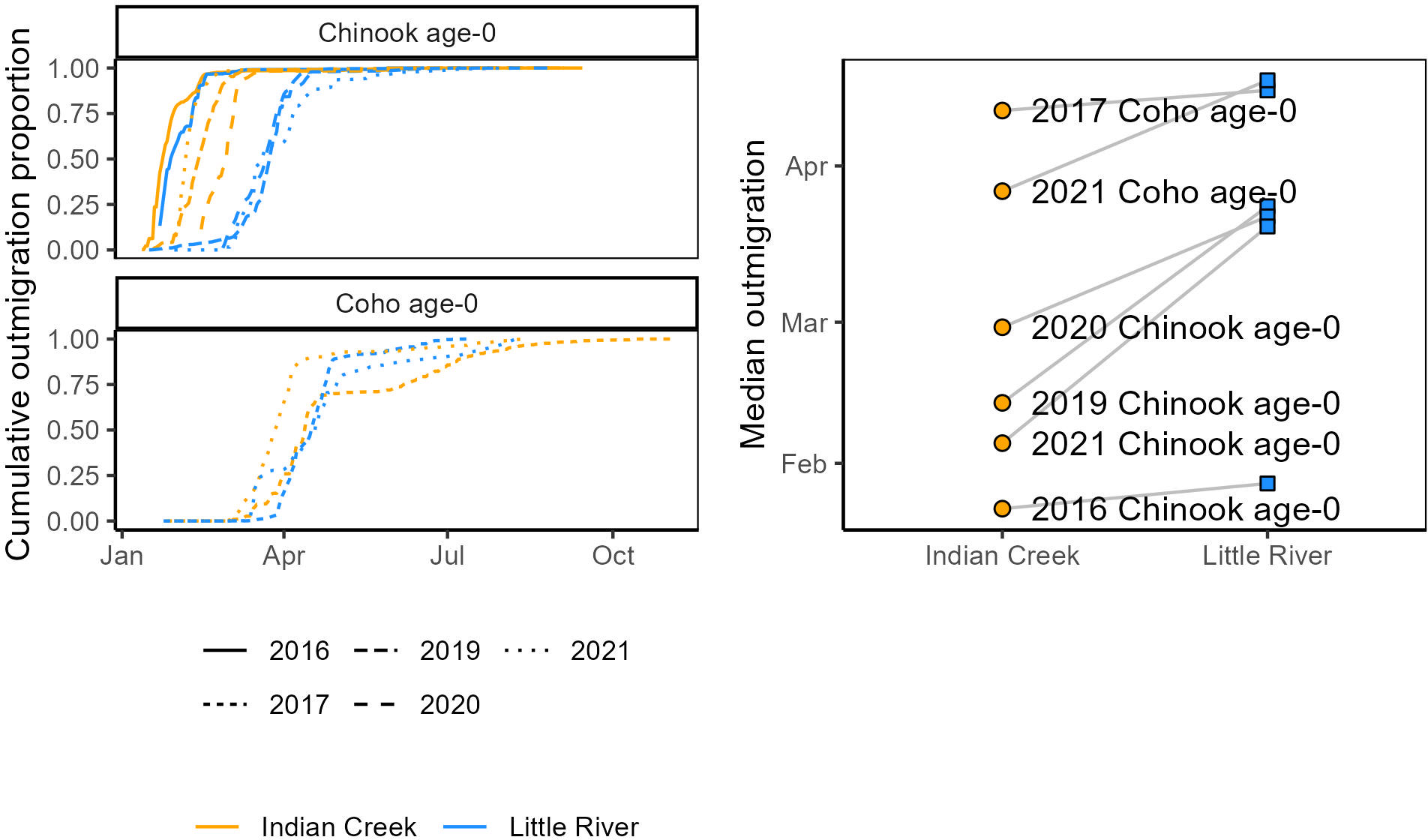
Figure 6 Salmonid outmigration timing compared between Indian Creek and Little River. Left: cumulative outmigrations as a proportion of total outmigrations. Right: median outmigration date, with dashed lines connecting observations of the same salmonid type and year.
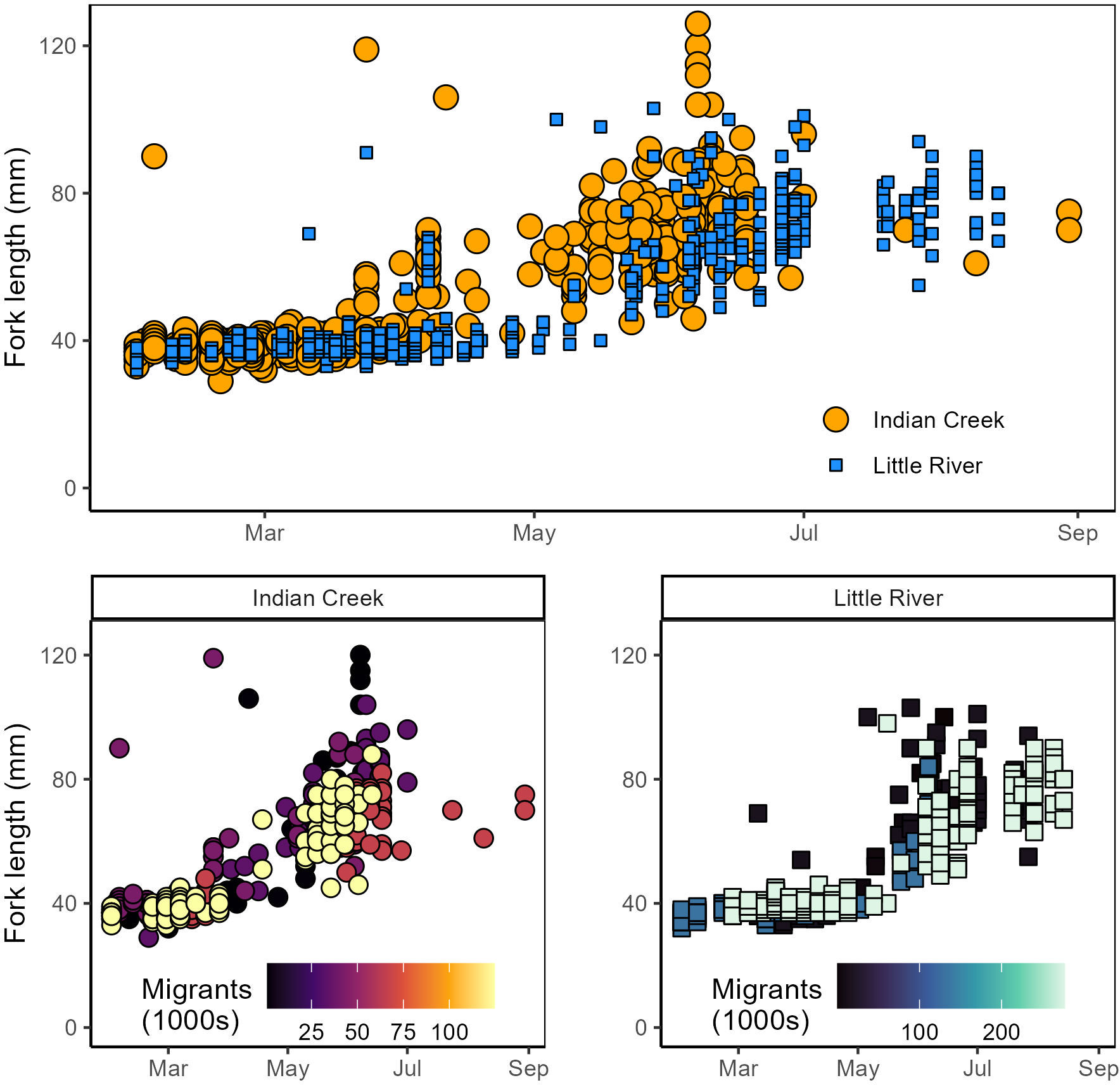
Figure 8 Top: Chinook salmon lengths compared between Indian Creek and Little River. Points describing Indian Creek fish lengths are larger and plotted behind Little River to improve visual comparison between tributaries. Bottom: Chinook salmon lengths compared within tributaries among years with different abundances of Chinook salmon outmigrants.
Notably, we have log-transformed the axes in Figures 2 and 3 so that small counts are more visible. Because log-transformations are inappropriate for stacked bar plots, we have reformatted these plots to be dot and line charts.
Additionally, because the corrected steelhead abundances no longer meet our original criteria for making robust comparisons between the two tributaries (i.e., instances when combinations of years, species, and identifiable age classes in both tributaries were in the top 33rd percentile of abundances relative to each combination of year, species, age class, and tributary), Figure 6 examines the outmigration timing of some different combinations of years and species, and no longer includes steelhead.
In the published article, there were errors in Tables 1 and 2 as published. We reported incorrect fish counts, which resulted in incorrect statistical output. The corrected Tables 1 and 2 appear below.
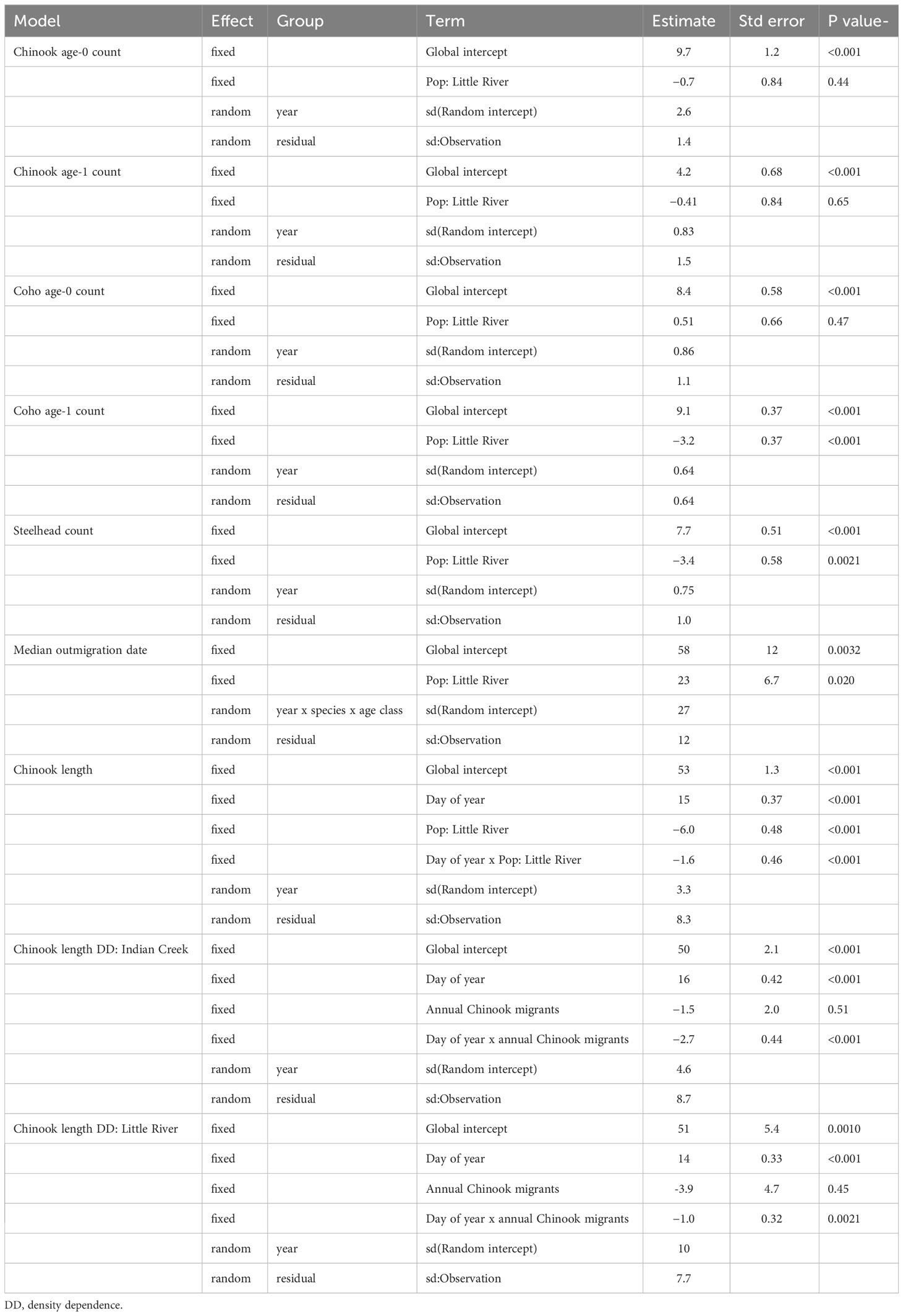
Table 1 Summary statistics of mixed effects models quantifying salmonid abundance, outmigrations timing, and length.
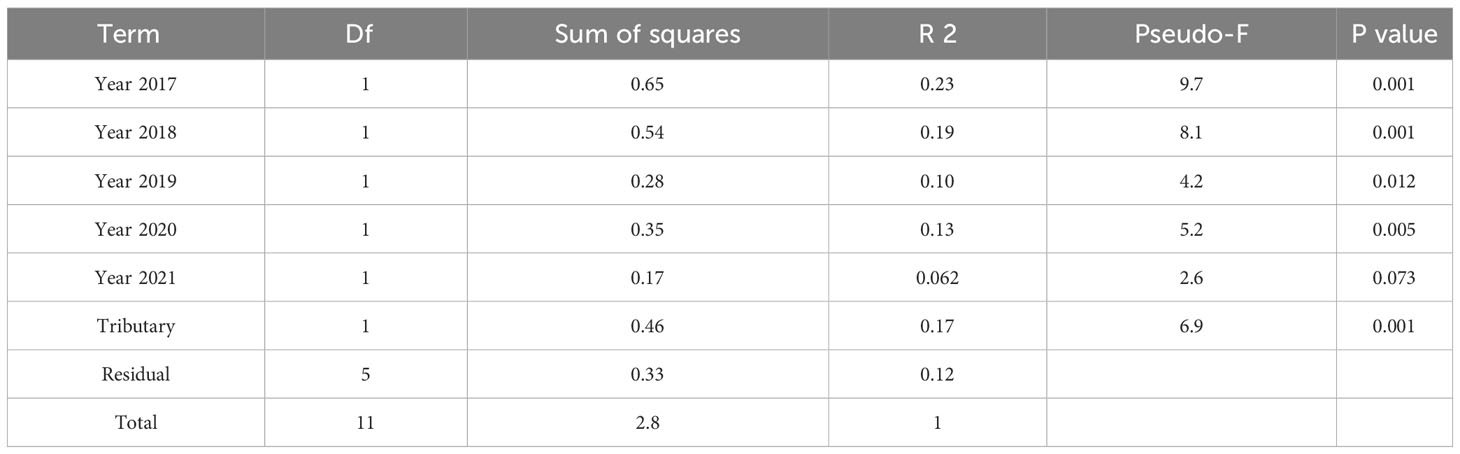
Table 2 Summary statistics of PERMANOVA comparing salmonid assemblages between tributaries and among years.
A correction has been made to the Abstract. These sentences previous stated:
“The warmer, low-gradient tributary produced more age-1+ coho salmon while the colder, steeper tributary produced a notably high abundance of steelhead smolts in 2020. Additionally, salmonids exiting the warmer tributary were older and possibly larger for their age class, emigrated ~25 days earlier, and included age-0 Chinook salmon that were larger.”
The corrected sentence appears below:
“The warmer, low-gradient tributary produced more age-1+ coho salmon and steelhead. Additionally, salmonids exiting the warmer tributary were older and possibly larger for their age class, emigrated ~23 days earlier, and included age-0 Chinook salmon that were larger.”
Corrections have been made to the Methods, Analyses, paragraph one. These sentences previously stated:
“where log(abundance) or median migration date μ was a function of an intercept β0, an effect β1 of tributary X1, and random intercepts ac.”
“Also for the outmigration timing model, we only compared combinations of years, species, and identifiable age classes when abundances in both tributaries’ traps were in the top 33rd percentile of abundances (13,555 individuals) relative to each combination of year, species, age class, and tributary to focus on comparisons with more robust sample sizes.”
The corrected sentences appear below:
“where log(abundance+1) or median migration date μ was a function of an intercept β0, an effect β1 of tributary X1, and random intercepts ac.”
“Also for the outmigration timing model, we only compared combinations of years, species, and identifiable age classes when abundances in both tributaries’ traps were in the top 33rd percentile of abundances (5,804 individuals) relative to each combination of year, species, age class, and tributary to focus on comparisons with more robust sample sizes.”
A correction has been made to the Results, paragraphs one, two, three, four, and seven. These sentences previously stated:
“Indian Creek and Little River supported a diversity of abundances, outmigration timings, and ages across Chinook salmon, coho salmon, and steelhead (Figures 2–4). Salmonids outmigrated from January to November, mostly within late winter to early summer. Chinook salmon generally migrated earliest, followed by steelhead and a relatively protracted outmigration of coho salmon. In 2019 and 2021, pulses of steelhead outmigrated from Indian Creek early in the outmigration window. Abundance and composition also varied among years, with greater abundances in 2019–2021 than 2016–2018, coho salmon dominating the assemblage in 2017 and 2018, Chinook salmon dominating the assemblage in 2016, 2019, and 2020, and notably abundant steelhead in Little River in 2020. Juveniles were more abundant in Indian Creek than Little River except in 2020. Chinook salmon were dominated by small individuals presumably age-0 whereas coho salmon and steelhead included a wider range of sizes that presumably reflected multiple age classes. Overall, while distinct patterns in assemblages were present between tributaries (discussed below), there was also considerable variation among years, with each year and tributary supporting different assemblages. Altogether, this variation meant that the timing, shape, and number of migration peaks was markedly different among years and between tributaries.”
“Indian Creek and Little River supported distinct salmonid assemblages (Figures 2–5). Indian Creek supported greater abundances of coho salmon, especially age 1+ individuals. Indeed, age-1 coho salmon were significantly more abundant in Indian Creek than Little River (Table 1, p = 0.002) while abundances of other salmonids did not significantly differ between tributaries (Table 1, p ≥ 0.1). Little River in 2020 notably produced the most abundant steelhead cohort, nearly quadruple the next greatest annual number observed in either tributary. In both tributaries, age-0 Chinook salmon were numerically dominant.”
“The tributaries’ assemblages differed significantly (p = 0.006, Table 2) but also shared significant temporal patterns (Figure 5, Table 2). Indeed, both tributaries’ assemblage trajectories followed a counterclockwise pattern in NMDS space from 2016–2021 (Figure 5). Perhaps the most striking variation shared by the tributaries over time was in Chinook salmon abundances, which were relatively high in 2016 and 2019–2021. In addition to this, steelhead were relatively abundant in both tributaries in 2020–2021 while coho salmon dominated assemblages in 2017 and 2018. Finally, both tributaries supported greater total salmonid abundances in each year of 2019–2021 than each year of 2016–2018.”
“Salmonids outmigrated ~25 days earlier from Indian Creek than Little River (Figure 6, Table 1, p = 0.01). Notably, monitoring in Indian Creek in some years appeared to begin after annual outmigrations had begun, suggesting the difference in median outmigration dates between the tributaries was likely an underestimate. For all species combined, the outmigration timing was also more protracted in Indian Creek compared to a more pulsed and shorter migration period in Little River, which was often due to its abundance of later-migrating age-0 coho salmon.”
“Age at outmigration also varied between tributaries. Both tributaries supported multiple outmigrant age classes, but Indian Creek supported proportionally older (or markedly larger – see Discussion) coho salmon and steelhead (Figure 7). Ages (or possibly only sizes, in the case of larger individuals) of coho salmon were especially different, with Little River supporting predominantly age-0 outmigrants whereas age-0 outmigrants comprised only ~50% of outmigrants in Indian Creek.”
“Some salmonid lengths differed between tributaries and were constrained by competition. Lengths of coho salmon (tributary parameter 95% CI posteriors: −0.57–0.55; Supplement) and steelhead (−0.83–0.71; Supplement) were not detectably different between tributaries. Notably, the complex age structure and multi-modal length distributions of these species may have made differences between tributaries harder to detect. Chinook salmon, however, were significantly smaller in Little River than Indian Creek (p < 0.001; Table 1) and these differences widened as winter progressed through summer (p < 0.001; Table 1) (Figure 8).”
“The model detecting this relationship indicated that salmon were 6 mm smaller in Little River on April 5, the average day of year that Chinook salmon were measured. Effects of competition on salmon length (i.e., density dependence) were also evident in Chinook salmon. Specifically, Chinook salmon in both tributaries were smaller later in the year during years when tributaries’ total Chinook outmigrants were greater (Figure 8, bottom). Statistical evidence for this was greater in Indian Creek (p < 0.001; Table 1) than Little River (p = 0.01; Table 1), and visual patterns suggested that density dependent effects on length were particularly apparent in Indian Creek during May–July (Figure 8, bottom left).”
The corrected sentences appear below:
“Indian Creek and Little River supported a diversity of abundances, outmigration timings, and ages across Chinook salmon, coho salmon, and steelhead (Figures 2–4). Salmonids outmigrated from January to November, mostly within late winter to early summer. Chinook salmon generally migrated earliest, followed by a relatively protracted outmigration of coho salmon that began before and ended after steelhead outmigrations. Abundance and composition also varied among years, with greater abundances in 2019–2021 than 2016–2018, coho salmon dominating the assemblage in 2017 and 2018, and Chinook salmon dominating the assemblage in 2016, 2019, and 2020. Juveniles were generally more abundant in Indian Creek than Little River, except in 2020 when juveniles were much more abundant in Little River and in 2018 when abundances were approximately equal between tributaries. Chinook salmon were dominated by small individuals presumably age-0 whereas coho salmon and steelhead included a wider range of sizes that presumably reflected multiple age classes. Overall, while distinct patterns in assemblages were present between tributaries (discussed below), there was also considerable variation among years, with each year and tributary supporting different assemblages. Altogether, this variation meant that the timing, shape, and number of migration peaks was markedly different among years and between tributaries.”
“Indian Creek and Little River supported distinct salmonid assemblages (Figures 2–5). Indian Creek supported greater abundances of coho salmon, especially age 1+ individuals, and steelhead. Indeed, age-1 coho salmon and steelhead were significantly more abundant in Indian Creek than Little River (Table 1, p <0.001 [coho salmon], p = 0.0021 [steelhead]). Abundances of other salmonids did not significantly differ between tributaries (Table 1, p ≥ 0.44). In both tributaries, age-0 Chinook salmon were numerically dominant.”
“The tributaries’ assemblages differed significantly (Table 2, p = 0.001) but also shared significant temporal patterns (Figure 5, Table 2). Both tributaries’ assemblage trajectories generally went up and right, then down, then left, then right in NMDS space from 2016–2021 (Figure 5). Perhaps the most striking variation shared by the tributaries over time was in Chinook salmon abundances, which were relatively high in 2016 and 2019–2021. In addition to this, steelhead abundances generally increased over time while coho salmon dominated assemblages in 2017 and 2018. Finally, both tributaries supported greater total salmonid abundances in each year of 2019–2021 than each year of 2016–2018.”
“Salmonids outmigrated ~23 days earlier from Indian Creek than Little River (Figure 6, Table 1, p = 0.020). Notably, monitoring in Indian Creek in some years appeared to begin after annual outmigrations had begun, suggesting the difference in median outmigration dates between the tributaries was likely an underestimate. For all species combined, the outmigration timing was also more protracted in Indian Creek compared to a more pulsed and shorter migration period in Little River, which was often due to its abundance of later-migrating coho salmon.”
“Some salmonid lengths differed between tributaries and were constrained by competition. Lengths of coho salmon (tributary parameter 95% CI posteriors: −0.57–0.55; Supplement) and steelhead (−0.83–0.71; Supplement) were not detectably different between tributaries. Notably, the complex age structure and multi-modal length distributions of these species may have made differences between tributaries harder to detect. Chinook salmon, however, were significantly smaller in Little River than Indian Creek (p < 0.001; Table 1) and these differences widened as winter progressed through summer (p < 0.001; Table 1) (Figure 8).”
“The model detecting this relationship indicated that salmon were 6 mm smaller in Little River on April 5, the average day of year that Chinook salmon were measured. Effects of competition on salmon length (i.e., density dependence) were also evident in Chinook salmon. Specifically, Chinook salmon in both tributaries were smaller later in the year during years when tributaries’ total Chinook outmigrants were greater (Figure 8, bottom). There was statistical evidence for density-dependent effects on growth in Indian Creek (p < 0.001; Table 1) and Little River (p = 0.0021; Table 1), with effect size being greater in Indian Creek than Little River (parameter estimate: −2.7 vs. −1.0; Table 1). Visual patterns suggested that density dependent effects on length were particularly apparent during May–July (Figure 8, bottom).”
A correction has been made to the Discussion, paragraphs one, two, three, and four (now one, two, three, and five). These sentences previously stated:
“We quantified the demographics of juvenile salmonids outmigrating from a heterogeneous landscape made accessible by dam removal. The tributaries supported different species assemblages, outmigration timings, age structures, and population-level growth trajectories. Specifically, the warmer tributary produced salmonids that outmigrated ~25 days earlier, more age-1 coho salmon, some notably large or old coho salmon, and larger Chinook salmon given the date. Additionally, species composition and abundance varied substantially among years and between streams. Moreover, the colder, steeper tributary generally produced fewer juveniles, except in 2020 when abundances were dramatically higher than in other years, which uniquely included a large cohort of steelhead. The patterns we observed were potentially due to multiple processes: (a) different stream gradients and temperature regimes favoring different species, (b) warmer temperature regimes accelerating incubation and growth, thus shifting seasonal outmigration timing forward, (c) stochastic, patchy adult recruitment and juvenile production during early phases of salmon repopulating the landscape, and (d) hatcheries sustaining baseline abundances of domesticated Chinook salmon that are not locally-adapted to the tributaries and happen to perform better in one than the other. Overall, restoring connectivity to tributaries with different characteristics rapidly enabled species to express diverse life histories. Such biological diversity is known to emerge from diverse habitat mosaics and promote resilience (e.g., Schindler et al., 2010, Lisi et al., 2013).”
“Temperature, stream gradient, and lake presence are fundamental to salmonid habitat mosaics, vary across landscapes, and likely drove some differences in demographics between the tributaries. High abundances of coho salmon being produced by a warmer, low-gradient tributary with lake access and ample beaver wetlands as well as an abundant cohort of steelhead being produced by a colder, steeper tributary are consistent with these species’ known habitat preferences (Bisson et al., 1988, Bugert and Bjornn 1991). Additionally, temperature regimes vary across landscapes and influence all phases of salmonid life histories (e.g., Brett et al., 1969, Richter & Kolmes 2005, Lisi et al., 2013, Fitzgerald et al., 2021). Our estimate that salmonids outmigrate ~25 days earlier in the warmer tributary extends similar findings on coho fry (Liermann et al., 2017) to the full diversity of salmonid species and life stages abundant in these systems. Warm or increasing temperatures can cue earlier migrations in juvenile salmonids (Spence & Dick 2014 and references therein), which is consistent with our observations of earlier migrations in warmer Indian Creek. In warmer waters that accelerate metabolism, juveniles can incubate and – given sufficient food – grow faster, which may enable them to reach outmigration size thresholds earlier in warmer environments (Brett et al., 1969, Murray & McPhail 1988, Peven 1994, Cline et al., 2019). Put together, warmer waters and presumably sufficient prey may have enabled Chinook salmon inhabiting Indian Creek to grow faster, promoting earlier outmigration. Interestingly, salmonids in warmer Indian Creek included greater proportions of older (age-1+) coho salmon. Plausibly, threshold lengths may determine outmgiration timing more directly in individuals genetically predisposed to enter the sea at age-0 while migration timing in individuals predisposed to rear for a year depends less on individual length and thus temperature regime (unfortunately ago-1 coho were not abundant enough in both tributaries during the same years to robustly compare outmigration timings between them). Moreover, mosaics of habitat types, and the associated differences in the environmental conditions such as water temperature, depth, and velocity, can affect the life history diversity and age structure of coho salmon populations (Sethi et al., 2021, Jones et al., 2021). Coho salmon with access to lake environments (e.g., Lake Sutherland connected to Indian Creek) in higher latitude watersheds can utilize both lentic and lotic habitats, resulting in differences in life history, age structure, and freshwater migration patterns (Sethi et al., 2017, 2021, 2022). Conversely, there can be differences in life history and size at migration (yearling v. parr, v. fry migrants) and associated age structure (age 0 v. age 1 – spring, summer, or fall/winter) in systems that have freshwater, estuarine, and ocean habitats connected or disconnected (Jones et al., 2021). Overall, the distinct environments in Indian Creek and Little River appear to have driven distinct life histories in juvenile salmonids.”
“In addition to landscapes generating diversity, some assemblage patterns among years and between tributaries may be attributable to adult recruitment and hatchery processes. During this study’s timeframe, salmonids were becoming more abundant and widely distributed across the landscape (Duda et al., 2021, Pess et al., in review, this issue). Notably, Chinook salmon tend to spawn in mainstems unless high spawner abundances (and adequate flows) promote expansion into tributaries. This appeared to happen in 2018 and 2019, when Chinook salmon escapement was high (Pess et al., in review, this issue) and redd counts in Indian Creek and Little River were especially high, resulting in high juvenile abundances the following years. Indeed, some of the synchronized variation in juvenile assemblage composition between tributaries among years appeared to be attributable to variation in adult spawning the year prior. If Chinook salmon abundances continue to increase across the landscape, annual spawner distributions may more routinely expand into the tributaries, resulting in greater abundances the following years. However, an important nuance to understanding Chinook salmon in this system is that returning adults are, to date, overwhelmingly hatchery-origin and presumably descend from a hatchery lineage that originated in 1930 (Pess et al., in review, this issue). It therefore seems unlikely that juvenile Chinook salmon are locally adapted to Indian Creek or Little River. One interpretation of Chinook salmon outmigrating at smaller sizes from Little River may be that the current stock’s genetically-determined traits happen to align more with niches in Indian Creek than Little River. Over decades as natural productivity has an opportunity to eclipse hatchery productivity, it remains to be seen whether natural selection will produce locally-adapted populations that exploit localized environments and opportunities, potentially driving more divergence in life histories between tributaries.”
“Competition also appeared to influence juvenile lengths and interact with the tributaries’ different growth opportunities. Chinook salmon were smaller when conspecific abundances were higher, and such density dependent effects are common for salmonids in general (Grossman and Simon 2019). Notably, these effects were most evident in Indian Creek, which appeared to support faster growth than Little River. That fish appeared to grow faster but experience greater density-dependent constraints on growth in Indian Creek is consistent with Indian Creek’s warmer temperatures offering greater potential for growth, but possibly also greater potential for prey limitation by increasing salmonids’ metabolic rates. Indeed, whether warmer waters within tolerable ranges promote growth depends on the interaction of temperature, prey availability, and competition (Crozier et al., 2010). While examining for the effects of competition on coho salmon and steelhead was beyond the scope of our study because of their more complex residence times, similar dynamics may influence growth in these species as well.”
The corrected sentences appear below:
“We quantified the demographics of juvenile salmonids outmigrating from a heterogeneous landscape made accessible by dam removal. The tributaries supported different species assemblages, outmigration timings, age structures, and population-level growth trajectories. Specifically, the warmer tributary produced salmonids that outmigrated ~23 days earlier, more age-1 coho salmon, more steelhead, some notably large or old coho salmon, and larger Chinook salmon given the date. Additionally, species composition and abundance varied substantially among years and between streams. Moreover, the colder, steeper tributary generally produced fewer juveniles, except in 2020 when abundances were dramatically higher in the colder, steeper tributary than in other years and in 2018 when abundances were approximately equal between tributaries. The patterns we observed were potentially due to multiple processes: (a) different stream gradients and temperature regimes favoring different species, (b) warmer temperature regimes accelerating incubation and growth, thus shifting seasonal outmigration timing forward, (c) stochastic, patchy adult recruitment and juvenile production during early phases of salmon repopulating the landscape, and (d) hatcheries sustaining baseline abundances of domesticated Chinook salmon that are not locally-adapted to the tributaries and happen to perform better in one than the other. Overall, restoring connectivity to tributaries with different characteristics rapidly enabled species to express diverse life histories. Such biological diversity is known to emerge from diverse habitat mosaics and promote resilience (e.g., Schindler et al., 2010, Lisi et al., 2013).”
“Temperature, stream gradient, and lake presence are fundamental to salmonid habitat mosaics, vary across landscapes, and likely drove some differences in demographics between the tributaries. High abundances of coho salmon being produced by a warmer, low-gradient tributary with lake access and ample beaver wetlands was consistent with this species’ known habitat preferences (Bisson et al., 1988, Bugert and Bjornn 1991). Additionally, temperature regimes vary across landscapes and influence all phases of salmonid life histories (e.g., Brett et al., 1969, Richter & Kolmes 2005, Lisi et al., 2013, Fitzgerald et al., 2021). Our estimate that salmonids outmigrate ~23 days earlier in the warmer tributary extends similar findings on coho fry (Liermann et al., 2017) to Chinook salmon in these systems. Warm or increasing temperatures can cue earlier migrations in juvenile salmonids (Spence & Dick 2014 and references therein), which is consistent with our observations of earlier migrations in warmer Indian Creek. In warmer waters that accelerate metabolism, juveniles can incubate and – given sufficient food – grow faster, which may enable them to reach outmigration size thresholds earlier in warmer environments (Brett et al., 1969, Murray & McPhail 1988, Peven 1994, Cline et al., 2019). Put together, warmer waters and presumably sufficient prey may have enabled Chinook salmon inhabiting Indian Creek to grow faster, promoting earlier outmigration. Interestingly, salmonids in warmer Indian Creek included greater proportions of older (age-1+) coho salmon. Plausibly, threshold lengths may determine outmgiration timing more directly in individuals genetically predisposed to enter the sea at age-0 while migration timing in individuals predisposed to rear for a year depends less on individual length and thus temperature regime (unfortunately ago-1 coho were not abundant enough in both tributaries during the same years to robustly compare outmigration timings between them). Moreover, mosaics of habitat types, and the associated differences in the environmental conditions such as water temperature, depth, and velocity, can affect the life history diversity and age structure of coho salmon populations (Sethi et al., 2021, Jones et al., 2021). Coho salmon with access to lake environments (e.g., Lake Sutherland connected to Indian Creek) in higher latitude watersheds can utilize both lentic and lotic habitats, resulting in differences in life history, age structure, and freshwater migration patterns (Sethi et al., 2017, 2021, 2022). Conversely, there can be differences in life history and size at migration (yearling vs. parr, vs. fry migrants) and associated age structure (age 0 vs. age 1 – spring, summer, or fall/winter) in systems that have freshwater, estuarine, and ocean habitats connected or disconnected (Jones et al., 2021). Overall, the distinct environments in Indian Creek and Little River appear to have driven distinct life histories in juvenile salmonids.”
“We observed greater abundances of steelhead in Indian Creek than Little River, despite the latter’s steeper, colder environment. While steelhead generally predominate among species in higher gradient habitats (McMillan et al., 2013), this may reflect their evolutionary ability to hold in faster currents (Bisson et al., 1988) and their comparatively broad spatial distribution within a watershed (McMillan et al., 2013) rather than a tendency to avoid warmer, lower gradient areas. Indeed, steelhead as a species can certainly tolerate rearing areas that are warm for salmonids (Richter and Kolmes 2005, Sloat and Osterback 2012) because they have evolved a scope for activity that is maximized at relatively warmer temperatures (Dickson and Kramer 1971). And, as outlined above, warmer streams within tolerable temperature ranges offer greater potential for growth, given adequate prey availability. Additionally, Indian Creek is larger and therefore presumably provides greater habitat capacity and its lake head dampens flow variation that can cause scour and flooding. For these reasons, Indian Creek’s environment may be more conducive to steelhead production than Little River.”
“In addition to landscapes generating diversity, some assemblage patterns among years and between tributaries may be attributable to adult recruitment and hatchery processes. During this study’s timeframe, salmonids were becoming more abundant and widely distributed across the landscape (Duda et al., 2021, Pess et al., in review, this issue). Notably, Chinook salmon tend to spawn in mainstems unless high spawner abundances (and adequate flows) promote expansion into tributaries. This appeared to happen in 2018 and 2019, when Chinook salmon escapement was high (Pess et al., in review, this issue) and redd counts in Indian Creek and Little River were especially high, resulting in high juvenile abundances the following years. Indeed, some of the synchronized variation in juvenile assemblage composition between tributaries among years appeared to be attributable to variation in adult spawning the year prior. If Chinook salmon abundances continue to increase across the landscape, annual spawner distributions may more routinely expand into the tributaries, resulting in greater abundances the following years. However, an important nuance to understanding Chinook salmon in this system is that returning adults are, to date, overwhelmingly hatchery-origin and presumably descend from a hatchery lineage that originated in 1930 (Pess et al., in review, this issue). It therefore seems unlikely that juvenile Chinook salmon are locally adapted to Indian Creek or Little River. One interpretation of Chinook salmon outmigrating at smaller sizes from Little River may be that the current stock’s genetically-determined traits happen to align more with niches in Indian Creek than Little River. Over decades as natural productivity has an opportunity to eclipse hatchery productivity, it remains to be seen whether natural selection will produce locally-adapted populations that exploit localized environments and opportunities, potentially driving more divergence in life histories between tributaries.”
“Competition also appeared to influence juvenile lengths and interact with the tributaries’ different growth opportunities. Chinook salmon were smaller when conspecific abundances were higher, and such density dependent effects are common for salmonids in general (Grossman and Simon 2019). Notably, these effects were greater in Indian Creek, which appeared to support faster growth than Little River. That fish appeared to grow faster but experience greater density-dependent constraints on growth in Indian Creek is consistent with Indian Creek’s warmer temperatures offering greater potential for growth, but possibly also greater potential for prey limitation by increasing salmonids’ metabolic rates. Indeed, whether warmer waters within tolerable ranges promote growth depends on the interaction of temperature, prey availability, and competition (Crozier et al., 2010). While examining for the effects of competition on coho salmon and steelhead was beyond the scope of our study because of their more complex residence times, similar dynamics may influence growth in these species as well.”
A correction has been made to the Acknowledgments. These sentences previously stated:
“Funding was provided to the Lower Elwha Klallam Tribe by the Environmental Protection Agency/Puget Sound Partnership through the Northwest Indian Fisheries Commission (contract no. PA-01J64601-1). Funding was provided by the Lower Elwha Klallam Tribe to the National Oceanic and Atmospheric Administration – Northwest Fisheries Science Center for the contributions of George Pess, Todd Bennett, John McMillan, and Martin Liermann (contract no. PA-01J64601-1). We thank Vanessa Castle, Mel Elofson, Allyce Miller, Ernest Sampson, Justin Stapleton, and mcKenzie Taylor for fieldwork, and Joshua W. Chamberlin, Sarah A. Morley, two anonymous reviewers, and handling editor Jean-Marc Roussel for critiques that improved our manuscript.”
The corrected sentences appear below:
“Funding was provided to the Lower Elwha Klallam Tribe by the Environmental Protection Agency/Puget Sound Partnership through the Northwest Indian Fisheries Commission (contract no. PA-01J64601-1). Funding was provided by the Lower Elwha Klallam Tribe to the National Oceanic and Atmospheric Administration – Northwest Fisheries Science Center for the contributions of George Pess, Todd Bennett, John McMillan, and Martin Liermann (contract no. PA-01J64601-1). We thank Vanessa Castle, Mel Elofson, Allyce Miller, Ernest Sampson, Justin Stapleton, and mcKenzie Taylor for fieldwork, and Joshua W. Chamberlin, Sarah A. Morley, two anonymous reviewers, and handling editor Jean-Marc Roussel for critiques that improved our manuscript. We also thank John Mahan for alerting us to an error in abundance values in a previous version of the article.”
A correction has been made to the References. We have added entries for Dickson and Kramer (1971), McMillan et al. (2013), and Sloat and Osterback (2013):
Dickson, I. W., & Kramer, R. H. (1971). Factors influencing scope for activity and active and standard metabolism of rainbow trout (Salmo gairdneri). Journal of the Fisheries Board of Canada, 28(4), 587–596.
McMillan, J. R., Liermann, M. C., Starr, J., Pess, G. R., & Augerot, X. (2013). Using a stream network census of fish and habitat to assess models of juvenile salmonid distribution. Transactions of the American Fisheries Society, 142(4), 942–956.
Sloat, M. R., & Osterback, A. M. K. (2013). Maximum stream temperature and the occurrence, abundance, and behavior of steelhead trout (Oncorhynchus mykiss) in a southern California stream. Canadian Journal of Fisheries and Aquatic Sciences, 70(1), 64–73.
The authors apologize for these errors and state that they do not change the scientific conclusions of the article in any way. The original article has been updated.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Keywords: Elwha river, habitat mosaic, restoration, thermal regime, phenology
Citation: Munsch SH, McHenry M, Liermann MC, Bennett TR, McMillan J, Moses R and Pess GR (2024) Corrigendum: Dam removal enables diverse juvenile life histories to emerge in threatened salmonids repopulating a heterogeneous landscape. Front. Ecol. Evol. 12:1357830. doi: 10.3389/fevo.2024.1357830
Received: 18 December 2023; Accepted: 15 January 2024;
Published: 30 January 2024.
Edited and Reviewed by:
Jean-Marc Roussel, Institut Agro, IFREMER, FranceCopyright © 2024 Munsch, McHenry, Liermann, Bennett, McMillan, Moses and Pess. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stuart H. Munsch, U3R1YXJ0Lk11bnNjaEBOT0FBLmdvdg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.