{kind=link}

95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 15 July 2024
Sec. Urban Ecology
Volume 12 - 2024 | https://doi.org/10.3389/fevo.2024.1352625
 Terézia Jauschová1,2
Terézia Jauschová1,2 Lenka Sarvašová1*
Lenka Sarvašová1* Peter Zach1Miroslav Saniga1Zdenka Martinková3
Peter Zach1Miroslav Saniga1Zdenka Martinková3 Alois Honěk3
Alois Honěk3 Jiří Skuhrovec3
Jiří Skuhrovec3 Milada Holecová4
Milada Holecová4 Ján Kulfan1
Ján Kulfan1The abundance and species composition of adult ladybird communities (Coleoptera: Coccinellidae) were investigated on two congeneric trees, native Norway spruce (Picea abies) and the introduced blue spruce (Picea pungens), at four locations in Slovakia (central Europe). For two years (2021–2022), coccinellid adults were sampled using a standard method involving beating branches at monthly intervals from April to November. Although the species composition of the communities on both spruce species was similar, the abundance of the entire coccinellid community as well as the abundance of individual species was significantly greater on Norway spruce than on blue spruce. With the current decline of Norway spruce as a result of several negative factors, blue spruce has emerged as a suitable substitute host plant for coccinellid communities in urban areas.
Norway spruce (Picea abies (L.) H. Karst) is a coniferous species with a wide distribution range across central Europe, not only in naturally colonised mountain areas but also in large areas where it has been intensively planted in the past two hundred years. Due to ongoing climate change, Norway spruce, a species highly sensitive to summer droughts and warm temperatures, is currently experiencing dieback (Kolář et al., 2017; Boczoń et al., 2018; Krejza et al., 2021) and frequent pest outbreaks (i.e., bark beetles), leading to the periodic decline of large, forested areas (Jactel et al., 2012; Marini et al., 2017; Netherer et al., 2019; Kamińska et al., 2021; Nardi et al., 2022). Stress factors such as air pollution, heat, water deficiency, and drought have even more intensive impacts on trees in urban environments (Lüttge and Buckeridge, 2023), resulting in the disappearance of many Norway spruce trees from urban plantings (Anonymous, 2023).
With the dieback of Norway spruce stands, insect communities associated with this host plant, including communities of coccinellids (Coleoptera, Coccinellidae), have also become threatened. Coccinellids play a crucial role in limiting the abundance of phytophagous insects (Hodek, 1973; Majerus, 2016), and their absence puts host plants in urban stands at greater risk. The preservation of diverse coccinellid fauna in towns and cities has both ecological and cultural significance (Soares et al., 2022). Norway spruce is a coniferous tree in which coccinellid communities have rarely been studied (Reddersen and Jensen, 2002; Selyemová et al., 2007; Kenis et al., 2020). This tree species is reported as a host of diverse coccinellid communities (Selyemová et al., 2007; Kenis et al., 2020). In central Europe, the coccinellid communities on Norway spruce include species similar to those of communities on other trees, mainly the dominant native Coccinella septempunctata Linnaeus, 1758 and invasive Harmonia axyridis (Pallas, 1773) (Stathas et al., 2011; Latibari et al., 2016; Kempkens and Gruppe, 2018; Kenis et al., 2020; Zach et al., 2020; Farrow et al., 2022), as well as coccinellids typically occurring on conifers such as Aphidecta obliterata (Linnaeus, 1758) (Burmeister et al., 2007; Timms et al., 2008; Banfield-Zanin and Leather, 2016) and Exochomus quadripustulatus (Linnaeus, 1758) (Stathas et al., 2011; Holecová et al., 2018; Nedvěd, 2020).
Therefore, in our study, we aimed to determine whether there are suitable substitute conifer species capable of replacing the role of the disappearing Norway spruce in the survival of these coccinellid communities. A suitable substitute species for Norway spruce, at least in urban habitats, may be the introduced blue spruce (Picea pungens Engelm). This species, which is native to North America (Fechner, 1990), can withstand drought better than other spruce species and is resistant to high insolation (Fechner, 1990). Since blue spruce is also resilient to high levels of air pollution (Qin et al., 2014; Bukharina et al., 2016), after its introduction to Europe, it was planted as a replacement for Norway spruce stands damaged by industrial emissions (Slodičák and Novák, 2008; Kula et al., 2016). Due to its aesthetic qualities, blue spruce can replace Norway spruce in park stands. This tree species has been widely planted in urban parks for approximately the past 160 years (Pagan and Randuška, 1988; Kulfan et al., 2010), and its entomofauna can therefore be studied in most cities in central Europe. Blue spruce is known to be a host for numerous insect groups in Europe: xylophagous beetles (Coleoptera) (Kršiak et al., 2009; Kula et al., 2012, 2013), ladybirds (Coleoptera) (Jauschová et al., 2024), moths (Lepidoptera) (Kulfan et al., 2010), sawflies (Hymenoptera, Symphyta) (Kula et al., 2016) and aphids (Homoptera) (Fryč, 2016; Wojciechowski et al., 2016).
Can blue spruce function as a host for insect communities hosted by the native Norway spruce? To answer this question, we investigated and compared the diversity of coccinellid communities on Norway spruce and blue spruce grown together in urban areas. Given their close congeneric relationships, we expected similar coccinellid communities on both spruce species in urban environments. For our study, over a two-year period, we sampled adult coccinellid communities (including Scymninae and Coccidulinae subfamilies) monthly on both spruce species in four Slovak towns representing diverse climatic regions. We then evaluated the qualitative and quantitative differences in the adult coccinellid communities occurring concurrently during the growing season on both spruce species.
Coccinellids were sampled from urban plantings (parks, alleys, cemeteries, housing estates, etc.) in four towns in Slovakia, where Norway spruce and blue spruce are planted together in sufficient numbers: Žilina (49.19805 N, 18.73806E, 363 m a.s.l.), Sliač (48.61125 N, 19.15655E, 302 m a.s.l.), Zvolen (48.57933 N, 19.14886E, 321 m a.s.l.) and Levice (48.21743 N, 18.60141E, 160 m a.s.l.). The towns are located in different geographical regions: northern (Žilina), central (Sliač, Zvolen) and southern (Levice) (Figure 1). At the sites, other deciduous trees (Tilia spp., Acer spp., Quercus spp.), conifers (Pinus spp., Pseudotsuga menziesii, Abies spp., Taxus spp.) and shrubs (Forsythia × intermedia, Juniperus spp., Sambucus nigra, Ligustrum spp.) occurred widely. The ground surface was covered with mown lawn and ornamental plants and, to a lesser extent, with asphalt and concrete.
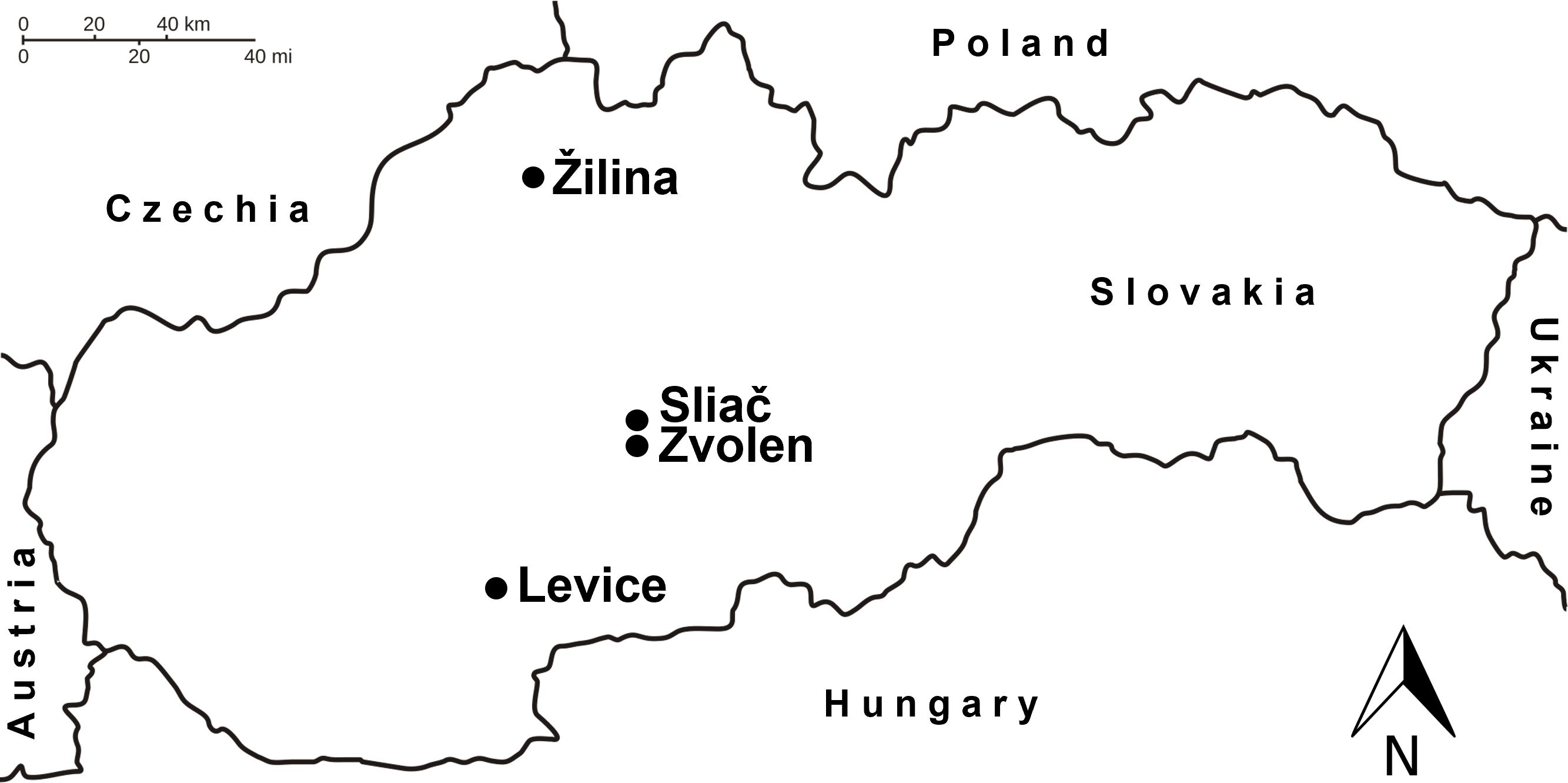
Figure 1 Locations of towns where sampling was carried out (•). Source: Adapted from https://commons.wikimedia.org/wiki/File:Slovensko_hranice.svg (public domain).
At each location, sampling was carried out at monthly intervals from April to November (8 total sampling sessions per year) in two consecutive years, 2021 and 2022 (16 total sampling sessions for the entire research period at each site). In each month, beetles from all locations were collected within 5-day intervals. Coccinellids were beaten from lower branches (1–3 m above the ground) with wooden sticks into a circular tray with a diameter of 1 m (Roy et al., 2011; Roy and Brown, 2018). All sampled trees were taller than 5 m and older than 15 years. At each sampling session, ten different Norway spruce trees and ten different blue spruce trees were sampled for ladybirds. A total of 10 branches were beaten per sampled tree. The samples were collected around the entire circumference of the tree crown. The maximal distance among the sampled trees was 100 m. Adult coccinellids were identified at the species level according to Nedvěd (2020). Coccinellid species were classified as “conifer specialists” according to their preference for coniferous trees following Koch (1989), Holecová et al. (2018) and Nedvěd (2020). The representation of “conifer specialists” was calculated as the relative abundance of “conifer specialists” among all ladybirds.
Differences in ladybird communities on Norway spruce and blue spruce were examined separately at individual sites. Qualitative differences were determined by the strength of the correlation of the relative abundance of individual species of coccinellids (the quantity of individuals of a species expressed as a proportion of the total number of coccinellid individuals) on Norway spruce and blue spruce. The calculations were performed for (i) all species for the entire research period at individual sites, (ii) species of the tribe Coccinellini (aphidophagous species), (iii) species belonging to all other tribes (non-Coccinellini, species readily accepting food other than aphids) and (iv) the 7 most abundant species [Exochomus quadripustulatus, Aphidecta obliterata, Harmonia quadripunctata (Pontoppidan, 1763), Rhyzobius chrysomeloides (Herbst, 1792), Adalia conglomerata (Linnaeus, 1758), Coccinella septempunctata, Harmonia axyridis]. Quantitative differences were investigated in a randomised block pattern without replicates, where samples collected at one location and date formed blocks and the spruce species represented the treatment. If the distribution of the experimental data passed the normality test, we used the pairwise multiple comparison procedure with the Holm-Sidak method. If the distribution of the experimental data failed the test of normality or equal variances, we used the Friedman repeated measures analysis of variance on ranks. This method was used because the abundance of ladybirds changed significantly during the growing season, and we were interested in the overall difference in the abundance of coccinellids on Norway spruce and blue spruce. All analyses were performed using SigmaStat 3.5 (Systat Software Inc, 2006).
The community structure of coccinellids on both spruce species in all four towns was similar. The correlation of the relative abundance of coccinellid species on Norway spruce and blue spruce was high at all locations: Levice (R2 = 0.991; P ≤ 0.001), Sliač (R2 = 0.920; P ≤ 0.001), Žilina (R2 = 0.903; P ≤ 0.001) and Zvolen (R2 = 0.975; P ≤ 0.001). The correlation of the relative abundance of species across all sampling sites was high (R2 = 0.986; P ≤ 0.001), which further indicated that both species host the same coccinellid communities (Figure 2).
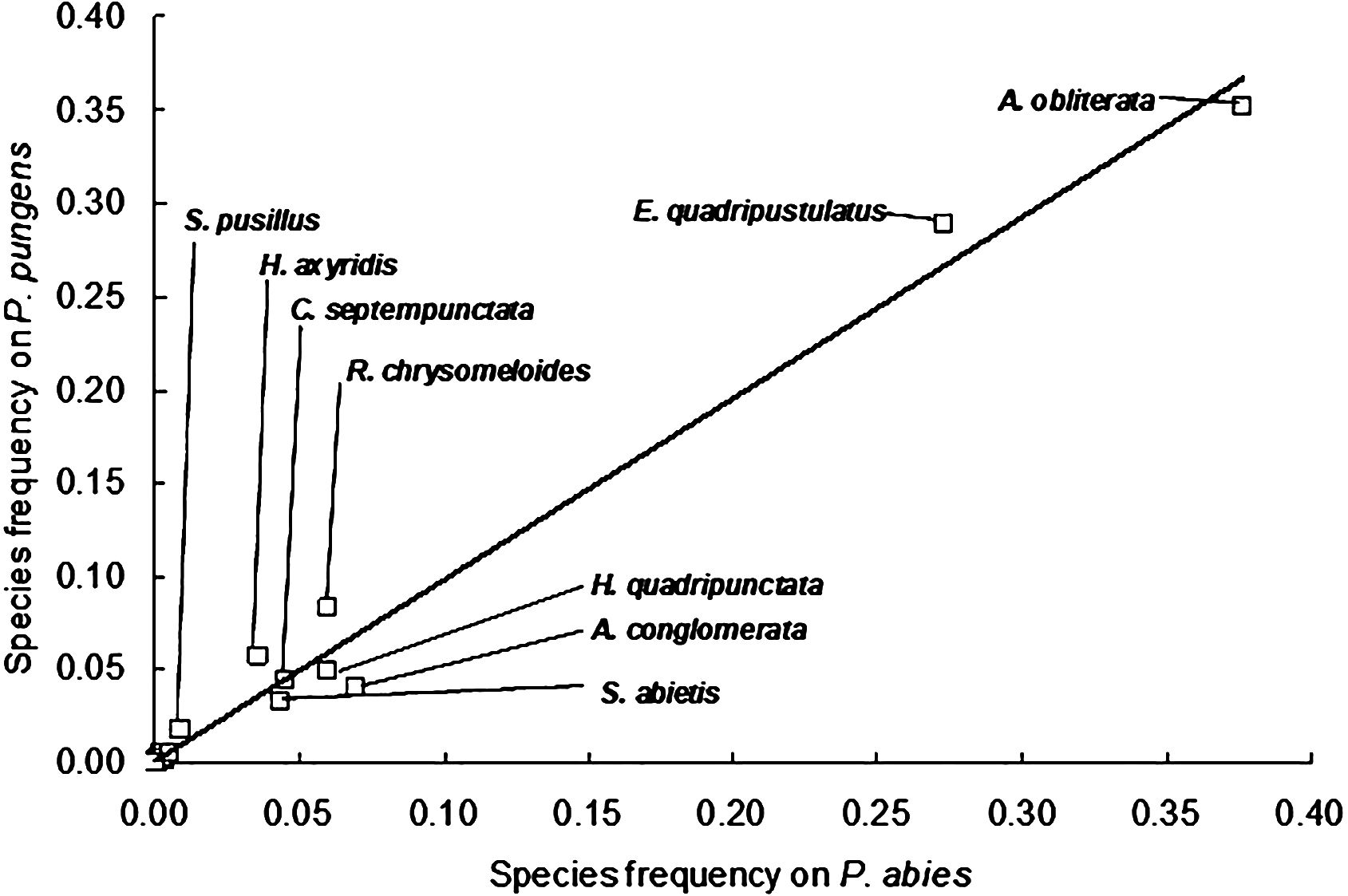
Figure 2 Frequency of coccinellid species on blue spruce plotted against the frequency of coccinellid species on Norway spruce. Data for the total sample of coccinellids from the Levice, Sliač, Žilina and Zvolen locations (average values from frequencies at individual locations). The black line indicates the same species frequency on Norway spruce and blue spruce.
In contrast to the species composition of the coccinellid community, we found significant differences in the abundance of coccinellids on Norway spruce and blue spruce (Table 1, Appendix 1). At each site, there was a significantly greater abundance of ladybirds on Norway spruce than on blue spruce.
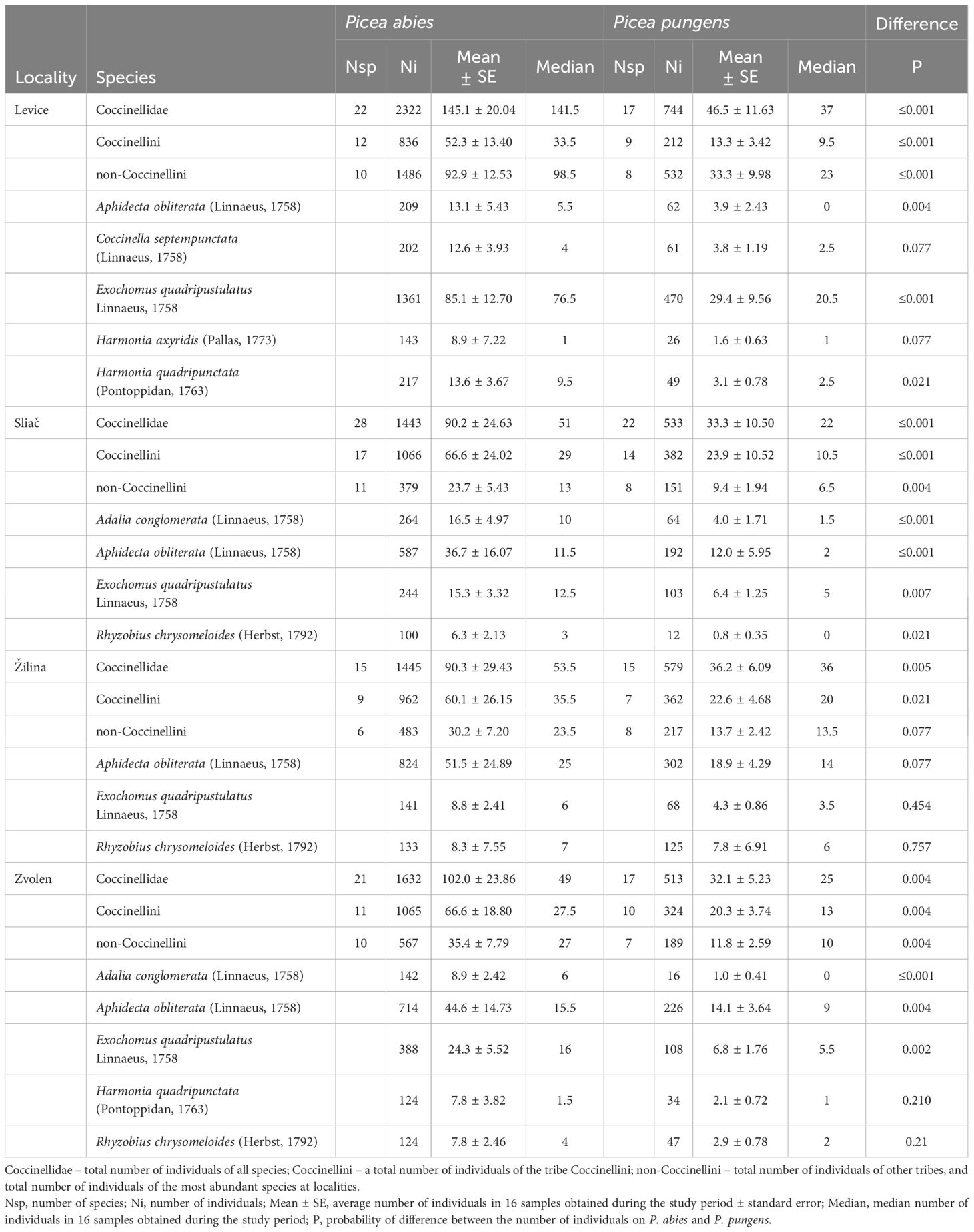
Table 1 Comparison of the coccinellid abundance on Norway spruce (Picea abies) and Blue spruce (Picea pungens) in locations in Slovakia.
The abundance of coccinellids was significantly greater on Norway spruce at the whole-community level, as was that of the coccinellids of the tribe Coccinellini and of other tribes, with the exception of that at the Žilina location. Some of the seven most abundant species (E. quadripustulatus, A. obliterata, H. quadripunctata, R. chrysomeloides, A. conglomerata, C. septempunctata, and H. axyridis) were significantly more abundant on Norway spruce than on blue spruce at individual locations (Table 1). The total abundance of the coccinellid community (both years and all locations) was 2.88-fold greater on Norway spruce than on blue spruce.
Additionally, the total number of species found at individual sites on Norway spruce (mean 21.5 ± 2.66 species) was greater than that found on blue spruce (mean 17.8 ± 1.49 species). However, this difference was only marginally significant (F=8.133, P=0.065) due to the small number of locations sampled. For the whole research period at all locations, a total of 34 coccinellid species were recorded on Norway spruce, and 27 species were recorded on blue spruce. Among them, Exochomus quadripustulatus and Aphidecta obliterata were eudominant (> 10%) on both spruce species. The invasive harlequin ladybird (Harmonia axyridis) was slightly more dominant on blue spruce (5.45%) than on Norway spruce (3.86%). Conifer specialist coccinellids strongly dominated communities on both spruce species [Norway spruce 87.66% (10 species), blue spruce 84.76% (9 species)].
The absence of qualitative differences in coccinellid communities between Norway spruce and blue spruce was mainly determined by the similar relative abundance of dominant species and the low relative abundance of rare species.
As a result, the relative abundances of coccinellid species on Norway spruce and blue spruce were similar across locations, although the dominant species across locations differed (Table 1), with the exception of Aphidecta obliterata and Exochomus quadripustulatus, which were abundant across all locations. Since the relative abundance of rare species was very low, it did not affect the correlation between the relative abundance of rare coccinellid species and that of either spruce species.
The species of spruce did not affect the relative abundance of coccinellid species in the total population that were established on the species. Additionally, Norway spruce and blue spruce have different species-specific properties that affect the attraction or repulsion of particular coccinellid species.
The invasive eurytopic coccinellid Harmonia axyridis (Roy et al., 2016) is known to dominate coccinellid communities on broadleaved trees (Honek et al., 2015; Viglášová et al., 2017). This ladybird species occurs on Norway spruce as well but usually in lower abundances (Kenis et al., 2020), which is in accordance with our results. Among conifers, Harmonia axyridis seems to prefer Scots pine (Pinus sylvestris) (Holecová et al., 2018; Kenis et al., 2020).
While no qualitative differences were found in the coccinellid communities on either spruce species, there were significant quantitative differences. All coccinellids (the community of all species), species of the tribe Coccinellini, species of other tribes (with the exception of the Žilina location) and some of abundant species were significantly more abundant on Norway spruce than on blue spruce. We can speculate about the reasons for these differences in the abundance of coccinellids between both spruce species. Possible causes include trophic factors, such as differences in the abundance or species composition of the arthropod complex serving as the prey for coccinellids. The aphid communities on Norway spruce and blue spruce seem to consist of similar aphid species (see, for example, Fryč, 2016; Wojciechowski et al., 2016). Trophic differences can be quantitative – the abundance of aphids on blue spruce seems to be lower than that on Norway spruce (pers. observation TJ, PZ). Other groups of phytophagous insects, as possible alternative food sources for coccinellids (Hodek and Evans, 2012; Majerus, 2016), seem to occupy both spruce species similarly, such as the larvae of moths (Kulfan et al., 2010) and sawflies (Kula et al., 2016).
Another biotic difference is the quality of the host plant (Goßner and Ammer, 2006; Bezemer et al., 2014). Differences in plant architecture, structural heterogeneity and plant surface, e.g., foliage shape and density, may aid or hinder coccinellid movement and searching and foraging behaviour (Shah, 1982; Grevstad and Klepetka, 1992; Pervez and Yadav, 2018). Little is known about whether there are differences in needle chemicals between blue spruce and Norway spruce (Soukupová et al., 2001). A comparison of the needles of Norway spruce and blue spruce with regard to their resistance to emissions revealed similar concentrations of chlorophyll and carotenoids but showed differences in phenolic compound accumulation between the two spruce species (Soukupová et al., 2001). The effects of chemical differences in the needles of both spruce species on coccinellids and their prey are not known.
How would coccinellid communities change if Norway spruce trees completely disappeared? For example, we can consider the replacement of the spruce by deciduous trees in the Děčínský Sněžník locality (Czech Republic, 50.79 N, 14.11E, 700 m a.s.l.), which has resulted in a shift in the composition of insect communities (Kula and Tryner, 2003a, b), including those of aphidophagous syrphids (Diptera) (Kula, 1997). A similar shift in the species composition of the local coccinellid fauna could also occur in the urban habitats in which this study was conducted.
Unlike the native Norway spruce, blue spruce is expected to be able to withstand unfavourable environmental conditions (Fechner, 1990; Qin et al., 2014; Bukharina et al., 2016). Our results suggest that in the absence of Norway spruce in urban environments, the coccinellid community inhabiting congeneric blue spruce will likely survive with no changes in composition, i.e., a decrease in species richness will not occur. In our study, the abundance of most coccinellid species was greater on Norway spruce than on blue spruce when coccinellids had the option to select Norway spruce and selection did not require migration to distant habitats. When such an option is not available, an increase in the abundance of coccinellids on blue spruce can be expected since the community of coccinellids adapted to conifers may find refuge there. However, confirmation of this assumption requires a more detailed study.
In conclusion, both native Norway spruce and introduced blue spruce host similar coccinellid communities in urban areas. Blue spruce was introduced to Slovakia more than 150 years ago (Pagan and Randuška, 1988), and in the time period since its introduction, it has recruited a coccinellid community with a species composition similar to that of the native Norway spruce. Closely related nonnative blue spruce seems to be a useful substitute host for spruce-associated insect fauna in the case of Norway spruce extinction in urban areas. A long-term study of coccinellid communities on spruces would be beneficial since the abundance of this insect is known to fluctuate interannually (this study; Selyemová et al., 2007).
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
The manuscript presents research on animals that do not require ethical approval for their study.
TJ: Conceptualization, Data curation, Investigation, Methodology, Writing – original draft, Writing – review & editing. LS: Conceptualization, Project administration, Writing – original draft, Writing – review & editing. PZ: Conceptualization, Investigation, Methodology, Supervision, Writing – original draft. MS: Funding acquisition, Investigation, Project administration, Writing – review & editing. ZM: Funding acquisition, Methodology, Project administration, Writing – review & editing. AH: Software, Supervision, Validation, Writing – original draft, Writing – review & editing. JS: Formal analysis, Methodology, Software, Writing – review & editing. MH: Funding acquisition, Investigation, Project administration, Writing – review & editing. JK: Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by a grant VEGA 2/0022/23 from the Scientific Grant Agency of Ministry of Education, Science, Research and Sport of the Slovak Republic (TJ, LS, JK, MS, MH), a grant for PhD students DoktoGrant APP0404 from the Slovak Academy of Sciences (TJ), a grant BAS-SAS-2022-02 (TJ, JK) and the Ministry of Agriculture of the Czech Republic, institutional support MZE-RO0423 (AH, ZM, and JS).
We thank Milan Mikuš for technical assistance with field research.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Anonymous (2023). In the town Zvolen, more trees in the parks and on the streets have to be cut down as a result of the drought and the bark beetle infestation, spruces are suffering, other trees, especially birches are also weakened. Available online at: https://www.lesmedium.sk/o-com-sa-pise/vo-zvolene-musia-vyrubat-dalsie-stromy-v-parkoch-a-na-uliciach-v-dosledku-posobenia-sucha-a-lykozruta-trpia-smreky-oslabene-su-aj-dalsie-dreviny-najma-brezy (Accessed November 15, 2023).
Banfield-Zanin J. A., Leather S. R. (2016). Prey-mediated effects of drought on the consumption rates of coccinellid predators of Elatobium abietinum. Insects 4), 49. doi: 10.3390/insects7040049
Bezemer T. M., Harvey J. A., Cronin J. T. (2014). Response of native insect communities to invasive plants. Ann. Rev. Entomol. 59, 119–141. doi: 10.1146/annurev-ento-011613-162104
Boczoń A., Kowalska A., Ksepko M., Sokołowski K. (2018). Climate warming and drought in the Bialowieza Forest from 1950–2015 and their impact on the dieback of Norway Spruce stands. Water 10, 1502. doi: 10.3390/w10111502
Bukharina I. L., Vedernikov K. E., Pashkova A. S. (2016). Morphophysiologic traits of spruce trees in conditions of Izhevsk. Contemp. Probl. Ecol. 9, 853–862. doi: 10.1134/S1995425516070027
Burmeister J., Goßner M., Gruppe A. (2007). Insektengemeinschaften im Kronenraum von Koniferenarten im Forstlichen Versuchsgarten Grafrath. Nachr. Bayer. Entomol. 56, 19–29.
Farrow R. A., Roy H. E., Brown P. M. J. (2022). Ladybird communities in rural woodlands: Does an invader dominate? Front. Conserv. Sci. 3. doi: 10.3389/fcosc.2022.759046
Fechner G. H. (1990). “Picea pungens Engelm. blue spruce,” in Silvics of north america 1: conifers. Eds. Burns R. M., Honkala B. H. (Forest Service United States Departement of Agriculture, Wshington DC), 238–249.
Fryč D. (2016). Mšice a mšičky na lesních dřevinách. [Aphids and Phylloxerans on Forest Trees (Opava: Ústřední kontrolní a zkušební ústav zemědělský).
Goßner M., Ammer U. (2006). The effects of Douglas-fir on tree-specific arthropod communities in mixed species stands with European beech and Norway spruce. Eur. J. For. Res. 125, 221–235. doi: 10.1007/s10342-006-0113-y
Grevstad F. S., Klepetka B. W. (1992). The influence of plant architecture on the foraging efficiencies of a suite of ladybird beetles feeding on aphids. Oecologia 92, 399–404. doi: 10.1007/BF00317466
Hodek I. (1973). Biology of the coccinellidae (Dordrecht: Springer Netherlands). doi: 10.1007/978-94-010-2712-0
Hodek I., Evans E.W. (2012). “Food relationships”, in Ecology and behaviour of the ladybird beetles (Coccinellidae), eds Hodek I., Honek A., Van Emden H.F. (Oxford: Blackwell Publishing Ltd.), 141–274.
Holecová M., Zach P., Hollá K., Šebestová M., Klesniaková M., Šestáková A., et al. (2018). Overwintering of ladybirds (Coleoptera: Coccinellidae) on Scots pine in central Europe. Eur. J. Entomol. 115, 658–667. doi: 10.14411/eje.2018.065
Honek A., Martinkova Z., Dixon A. F. (2015). Detecting seasonal variation in composition of adult Coccinellidae communities. Ecol. Entomol. 40, 543–552. doi: 10.1111/een.12225
Jactel H., Petit J., Desprez-Loustau M. L., Delzon S., Piou D., Battisti A., et al. (2012). Drought effects on damage by forest insects and pathogens: a meta-analysis. Glob. Change Biol. 18, 267–276. doi: 10.1111/j.1365-2486.2011.02512.x
Jauschová T., Sarvašová L., Saniga M., Langraf V., Holecová M., Honěk A., et al. (2024). Ladybird (Coleoptera, Coccinellidae) communities on nonnative blue spruce in central Europe. Folia Oecol. 51, 18–28. doi: 10.2478/foecol-2024-0002
Kamińska A., Lisiewicz M., Kraszewski B., Stereńczak K. (2021). Mass outbreaks and factors related to the spatial dynamics of spruce bark beetle (Ips typographus) dieback considering diverse management regimes in the Białowieża forest. For. Ecol. Manage. 498, 119530. doi: 10.1016/j.foreco.2021.119530
Kempkens V., Gruppe A. (2018). “Coccinellid communities in christmas tree cultures (Coleoptera, Coccinellidae),” in Mitteilungen der Deutschen Gesellschaft fur Allgemeine und Angewandte Entomologie, vol. 21. , 93–97.
Kenis M., Nacambo S., Van Vlaenderen J., Zindel R., Eschen R. (2020). Long term monitoring in Switzerland reveals that Adalia bipunctata strongly declines in response to Harmonia axyridis invasion. Insects 11, 883. doi: 10.3390/insects11120883
Koch K. (1989). “Coccinellidae”, in Die Käfer Mitteleuropas. Ökologie [The beetles of Central Europe. Ecology]. Band 2. (Krefeld: Goecke & Evers), 237–253. (In German).
Kolář T., Čermák P., Trnka M., Žid T., Rybníček M. (2017). Temporal changes in the climate sensitivity of Norway spruce and European beech along an elevation gradient in Central Europe. Agric. For. Meteorol. 239, 24–33. doi: 10.1016/j.agrformet.2017.02.028
Krejza J., Cienciala E., Světlík J., Bellan M., Noyer E., Horáček P., et al. (2021). Evidence of climate-induced stress of Norway spruce along elevation gradient preceding the current dieback in Central Europe. Trees 35, 103–119. doi: 10.1007/s00468-020-02022-6
Kršiak B., Zach P., Kulfan J., Dvořáčková K. (2009). Je smrek pichľavý (Picea pungens) atraktívny pre xylofilné chrobáky (Coleoptera) aj mimo oblasť prirodzeného výskytu? [Is blue spruce (Picea pungens) attractive for xylophilous beetles (Coleoptera) also beyond the area of its origin?]. Entomofauna Carpathica 21, 18–21. (In Slovak).
Kula E. (1997). Hoverflies (Dipt.: Syrphidae) of spruce forest in different health condition. Entomophaga 42, 133–138.
Kula E., Holuša J., Roller L., Úradník M. (2016). Allochthonous blue spruce in Central Europe serves as a host for many native species of sawflies (Hymenoptera, Symphyta). J. Hymenopt. Res. 51, 159–169. doi: 10.3897/jhr.51.9219
Kula E., Kajfosz R., Polívka J. (2013). Attractiveness of Picea pungens to the bark beetle species Ips amitinus (Eichh.) and Pityogenes chalcographus (L.). J. For. Sci. 59, 493–502.
Kula E., Kajfosz R., Polívka J. (2012). Dendroctonus micans (Kug.) a kambioxylofÁgnÍ fauna smrku pichlavého (Picea pungens Engelm.) ve střednÍ Evropě(Krušné hory, Česká republika) [Dendroctonus micans (Kug.) and cambioxylophagous fauna of blue spruce (Picea pungens Engelm.) in the Central Europe (Ore Mts., Czech Republic).]. Zprávy lesnického výzkumu 57, 378–386. (In Czech).
Kula E., Tryner P. (2003a). Hymenoptera (Aculeata) in spruce stands of air-polluted area of Northern Bohemia. J. For. Sci. 49, 200–207. doi: 10.17221/4694-JFS
Kula E., Tryner P. (2003b). Hymenoptera (Aculeata) in birch stands of air-polluted area of Northern Bohemia. J. For. Sci. 49, 148–158. doi: 10.17221/4690-JFS
Kulfan J., Kula E., Zach P., Dvořáčková K., Patočka J., Kršiak B. (2010). Caterpillar assemblages on introduced blue spruce: differences from native Norway spruce. Allg. Forst. Jagdztg 181, 188–194.
Latibari M. H., Moravvej G., Namaghi H. S., Khormizi M. Z. (2016). Coccinellid biodiversity on the coniferous trees Thuja orientalis and Pinus mugo in urban landscape of Mashhad, Razavi Khorasan Province, Iran. Egypt. 26, 419–421.
Lüttge U., Buckeridge M. (2023). Trees: structure and function and the challenges of urbanization. Trees 37, 1–8. doi: 10.1007/s00468-020-01964-1
Majerus M. E. N. (2016). A natural history of ladybird beetles (Cambridge: Cambridge University Press). doi: 10.1017/CBO9781316336960
Marini L., Økland B., Jönsson A. M., Bentz B., Carroll A., Forster B., et al. (2017). Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography 40, 1426–1435. doi: 10.1111/ecog.02769
Nardi D., Finozzi V., Battisti A. (2022). Massive windfalls boost an ongoing spruce bark beetle outbreak in the Southern Alps. Ital. For. Mont. 77, 1, 23–34. doi: 10.36253/ifm-1617
Nedvěd O. (2020). Brouci čeledi slunéčkovití (Coccinellidae) střední Evropy/Ladybird beetles (Coccinellidae) of Central Europe (Praha: Academia).
Netherer S., Panassiti B., Pennerstorfer J., Matthews B. (2019). Acute drought is an important driver of bark beetle infestation in Austrian Norway spruce stands. Front. For. Glob. Change 2, 39. doi: 10.3389/ffgc.2019.00039
Pagan J., Randuška D. (1988). Atlas drevín 2. - Cudzokrajné dreviny. [The Atlas of Woody Plants 2. – Introduced Woody Plants (Bratislava: Obzor).
Pervez A., Yadav M. (2018). Foraging behaviour of predaceous ladybird beetles: a review. Eur. J. Environ. Sci. 8, 102–108. doi: 10.14712/23361964.2018.14
Qin X., Sun N., Ma L., Chang Y., Mu L. (2014). Anatomical and physiological responses of Colorado blue spruce to vehicle exhausts. Environ. Sci. pollut. Res. 21, 11094–11098. doi: 10.1007/s11356-014-3015-4
Reddersen J., Jensen T. S. (2002). The Syrphidae, Coccinellidae, Neuroptera (s. lat.) of a large Danish spruce forest. Entomol. Medd. 70, 113–127.
Roy H. E., Brown P. M. J., Adriaens T., Berkvens N., Borges I., Clusella-Trullas S., et al. (2016). The harlequin ladybird, Harmonia axyridis: global perspectives on invasion history and ecology. Biol. Invasions 18, 997–1044. doi: 10.1007/s10530-016-1077-6
Roy H. E., Brown P. M. J., Frost R., Poland R. (2011). Ladybirds (Coccinellidae) of Britain and Ireland. Wallingford: Biological Records Centre.
Roy H. E., Brown P. M. J. (2018). Field guide to the ladybirds of Great Britain and Ireland. London: Bloomsbury Publishing.
Selyemová D., Zach P., Némethová D., Kulfan J., Úradník M., Holecová M., et al. (2007). Assemblage structure and altitudinal distribution of lady beetles (Coleoptera, Coccinellidae) in the mountain spruce forests of Poľana Mountains, the West Carpathians. Biologia 62, 610–616.
Shah M. A. (1982). The influence of plant surfaces on the searching behaviour of coccinellid larvae. Entomol. Exp. Appl. 31, 377–380. doi: 10.1111/j.1570-7458.1982.tb03163.x
Slodičák M., Novák J. (2008). Nutrients in the aboveground biomass of substitute tree species stand with respect to thinning–blue spruce (Picea pungens Engelm.). J. For. Sci. 54, 85–91. doi: 10.17221/3032-JFS
Soares A. O., Haelewaters D., Ameixa O. M. C. C., Borges I., Brown P. M. J., Cardoso P., et al (2022). A roadmap for ladybird conservation and recovery. Conserv. Biol. 37, e13965. doi: 10.1111/cobi.13965
Soukupová J., Rock B. N., Albrechtová J. (2001). Comparative study of two spruce species in a polluted mountainous region. New Phytol. 150, 133–145. doi: 10.1046/j.1469-8137.2001.00066.x
Stathas G. J., Eliopoulos P. A., Salmas I. C., Kozar F. (2011). Data on ecology of some Hemiptera species recorded in the forest of Taygetus Mountain, Peloponnesus, Greece. Phytoparasitica 39, 377–383. doi: 10.1007/s12600-011-0173-5
Timms J. E., Oliver T. H., Straw N. A., Leather S. R. (2008). The effect of host plant on the coccinellid functional response: Is the conifer specialist Aphidecta obliterata (L.) (Coleoptera: Coccinellidae) better adapted to spruce than the generalist Adalia bipunctata (L.) (Coleoptera: Coccinellidae)? Biol. Control. 47, 273–281. doi: 10.1016/j.biocontrol.2008.08.009
Viglášová S., Nedvěd O., Zach P., Kulfan J., Parák M., Honěk A., et al. (2017). Species assemblages of ladybirds including the harlequin ladybird Harmonia axyridis: a comparison at large spatial scale in urban habitats. BioControl 62, 409–421. doi: 10.1007/s10526-017-9793-0
Wojciechowski W., Depa Ł., Halgoš J., Matečný I., Lukáš J., Kanturski M. (2016). Aphids of Slovakia: distributional catalogue, checklist, key and list of host plants (Bratislava: Comenius University in Bratislava. Faculty of Natural Sciences).
Zach P., Holecová M., Brabec M., Hollá K., Šebestová M., Martinková, et al. (2020). Scots pine forest in Central Europe as a habitat for Harmonia axyridis: temporal and spatial patterns in the population of an alien ladybird. Folia Oecol. 47, 81–88. doi: 10.2478/foecol-2020-0010
Total numbers of individuals of coccinellid species recorded at all locations in this study on Norway spruce (Picea abies).
Adalia bipunctata (Linnaeus) 1; Adalia conglomerata (Linnaeus) 426; Adalia decempunctata (Linnaeus) 12; Anatis ocellata (Linnaeus) 9; Aphidecta obliterata (Linnaeus) 2334; Calvia decemguttata (Linnaeus) 3; Calvia quatuordecimguttata (Linnaeus) 2; Chilocorus bipustulatus Linnaeus 9; Clitostethus arcuatus (Rossi) 1; Coccinula quatuordecimpustulata (Linnaeus) 1; Coccinella quinquepunctata Linnaeus 1; Coccinella septempunctata Linnaeus 343; Exochomus quadripustulatus Linnaeus 2134; Halyzia sedecimguttata (Linnaeus) 1; Harmonia axyridis (Pallas) 264; Harmonia quadripunctata (Pontoppidan) 442; Hippodamia variegata (Goeze) 3; Myrrha octodecimguttata (Linnaeus) 2; Myzia oblongoguttata (Linnaeus) 24; Nephus quadrimaculatus (Herbst) 5; Oenopia conglobata (Linnaeus) 3; Platynaspis luteorubra (Goeze) 4; Propylea quatuordecimpunctata (Linnaeus) 11; Psyllobora vigintiduopunctata (Linnaeus) 3; Rhyzobius chrysomeloides (Herbst) 362; Scymnus abietis (Paykull) 258; Scymnus ferrugatus (Moll) 6; Scymnus frontalis (Fabricius) 11; Scymnus interruptus (Goeze) 24; Scymnus rubromaculatus (Goeze) 16; Scymnus subvillosus (Goeze) 1; Scymnus suturalis Thunberg 7; Stethorus pusillus (Herbst) 77; Vibidia duodecimguttata (Poda) 42.
Total numbers of individuals of coccinellid species recorded at all locations in this study on blue spruce (Picea pungens).
Adalia conglomerata (Linnaeus) 87; Adalia decempunctata (Linnaeus) 11; Anatis ocellata (Linnaeus) 4; Aphidecta obliterata (Linnaeus) 782; Calvia quatuordecimguttata (Linnaeus) 2; Chilocorus bipustulatus Linnaeus 6; Coccinella septempunctata Linnaeus 113; Exochomus quadripustulatus Linnaeus 749; Harmonia axyridis (Pallas) 129; Harmonia quadripunctata (Pontoppidan) 120; Hippodamia variegata (Goeze) 1; Myrrha octodecimguttata (Linnaeus) 1; Myzia oblongoguttata (Linnaeus) 3; Oenopia conglobata (Linnaeus) 6; Platynaspis luteorubra (Goeze) 1; Propylea quatuordecimpunctata (Linnaeus) 6; Psyllobora vigintiduopunctata (Linnaeus) 3; Rhyzobius chrysomeloides (Herbst) 184; Scymnus abietis (Paykull) 78; Scymnus auritus Thunberg 1; Scymnus ferrugatus (Moll) 2; Scymnus frontalis (Fabricius) 5; Scymnus haemorrhoidalis Herbst 1; Scymnus interruptus (Goeze) 11; Scymnus rubromaculatus (Goeze) 8; Stethorus pusillus (Herbst) 43; Vibidia duodecimguttata (Poda) 12.
Keywords: ladybirds, conifers, congeneric plants, urban ecosystems, Picea abies, Picea pungens
Citation: Jauschová T, Sarvašová L, Zach P, Saniga M, Martinková Z, Honěk A, Skuhrovec J, Holecová M and Kulfan J (2024) Coccinellidae on native and introduced spruce in central Europe: conservation implications in urban areas. Front. Ecol. Evol. 12:1352625. doi: 10.3389/fevo.2024.1352625
Received: 08 December 2023; Accepted: 17 June 2024;
Published: 15 July 2024.
Edited by:
Panagiotis Theodorou, Martin Luther University of Halle-Wittenberg, GermanyReviewed by:
Olivia Sanllorente, University of Granada, SpainCopyright © 2024 Jauschová, Sarvašová, Zach, Saniga, Martinková, Honěk, Skuhrovec, Holecová and Kulfan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lenka Sarvašová, c2FydmFzb3ZhQGlmZS5zaw==; bGVua2Euc2FydmFzb3ZhMTRAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.