 Félicien Degueldre
Félicien Degueldre Serge Aron
Serge Aron- Laboratory Evolutionary Biology and Ecology, Université Libre de Bruxelles, Bruxelles, Belgium
In many species, females have multiple mates, whose sperm compete for paternity. Males may subsequently invest in the increased production of sperm and/or seminal fluid. The latter is a complex mixture of proteins, peptides, and other compounds generated by the accessory glands (AGs) and is transferred to females along with a male’s sperm. Seminal fluid is known to be a key determinant of competitive outcomes among sperm, and its production may trade off with that of sperm. We show that AG size—a proxy for seminal fluid production—has a positive and phylogenetically robust correlation with both sperm competition intensity and sperm production in nine species of Cataglyphis desert ants. These results indicate a lack of trade-off between sperm production and seminal fluid production. They underscore that sperm competition may strongly shape sperm traits and could drive reproductive performance in eusocial hymenopterans.
Introduction
Within species, female promiscuity often fosters competition among males, whose spermatozoa vie for fertilisation opportunities (Parker, 1970). This competition extends sexual selection beyond mating and can significantly affect male fitness. Extensive theoretical and empirical research has shown the evolutionary impacts of sperm competition on sperm traits, particularly (i) number (e.g., Gomendio et al., 1998; Gage and Morrow, 2003), (ii) size (e.g., Gomendio and Roldan, 1991; Immler et al., 2011; Tourmente et al., 2011), and (iii) velocity (e.g., Anderson et al., 2007; Firman et al., 2013; Nakanishi and Takegaki, 2018).
Investment in the seminal fluid is another powerful mean through which males can enhance their reproductive success (Ramm, 2020). Typically transferred to females at the same time as sperm, seminal fluid is a complex combination of secretions produced by the sex accessory glands (AGs) (Chapman, 2001; Manaskova et al., 2002; King et al., 2011; den Boer et al., 2015; Santiago-Moreno and Blesbois, 2020), and it contains diverse organic compounds, including proteins, lipids, and sugars (Owen and Katz, 2005; Avila et al., 2011; Gorshkov et al., 2015; Lu et al., 2016). Post ejaculation, seminal fluid can influence sperm performance (den Boer et al., 2009; Smith and Stanfield, 2012), competitive outcomes among rival sperm in the female reproductive tract (den Boer et al., 2010), and female reproductive behaviour and physiology (Simmons and Gwynne, 1991; Wolfner, 2002).
There thus exist two potential, mutually exclusive relationships between sperm production and seminal fluid production (Ramm, 2020). First, there may be a positive correlation if seminal fluid boosts male reproductive success by enhancing sperm function. Second, there may be a negative correlation if a trade-off exists between sperm production and seminal fluid production. For instance, if seminal fluid has effects that are independent of sperm (e.g., the fluid generates a plug that prevents or interferes with further female mating), then a greater investment in seminal fluid production may relax selection pressure on sperm production. To date, findings are mixed: the production of sperm and seminal fluid appears to be positively correlated in mammals (Dixson, 1998; Ramm et al., 2005), fishes (Mazzoldi et al., 2005), and some insects (Morrow and Gage, 2000) but negatively correlated in fungus-growing ants (Baer and Boomsma, 2004).
The mating system of eusocial hymenopterans (ants, social bees, and wasps) imposes unique selective pressures on male ejaculates that are rarely, if ever, found in other animals (Boomsma et al., 2009; Boomsma, 2013). First, males produce sperm only once in their lives. Spermatogenesis mainly occurs during the pupal stage and is usually completed shortly after males emerge. Their testes then deteriorate, and sperm are stored in the accessory testes until mating. At ejaculation, the semen and seminal fluid are combined. Males die shortly after copulation, by exhaustion or predation. Second, mating occurs during a single nuptial flight at the beginning of adult life for both sexes. During this short window of time, female reproductives copulate with one or several males, amassing a lifetime supply of sperm in their spermatheca. In some species, females live for decades, generating offspring from stored sperm. Third, polyandry has evolved repeatedly in ants, bees, and wasps (Hughes et al., 2008), creating conditions conducive to sperm competition in multiple species. In the few studies looking at how sperm competition influences sperm traits, it has been observed that sperm competition may select for (i) enhanced sperm production (ants: Baer and Boomsma, 2004; Aron et al., 2016; Degueldre and Aron, 2023), (ii) increased sperm size (bumble bees: Baer et al., 2003). Furthermore, seminal fluid can promote male fitness by (i) enhancing sperm viability (honey bees: den Boer et al., 2009); (ii) incapacitating rival sperm when females have multiple mates (ants and honey bees: den Boer et al., 2010); and (iii) reducing female promiscuity by either generating a mating plug (ants: Baer, 2011; bumble bees: Duvoisin et al., 1999; Brown et al., 2002) or by negatively affecting the eyesight of females during the mating flight (honey bees: Liberti et al., 2019).
However, it remains largely unknown whether there is a correlation between the production of sperm and seminal fluid in eusocial hymenopterans. To date, a single study on fungus-growing ants has revealed that, in species with singly mated females, males have small accessory testes and large AGs, while the opposite is true in species with multiply mated females (Baer and Boomsma, 2004). It has been suggested that males exploit AG compounds to produce mating plugs and prevent further mating by females (Baer and Boomsma, 2004; Baer, 2011; den Boer et al., 2015). Consequently, if mating plugs became incapable of serving this function, males might redirect their investment from seminal fluid (i.e., large AGs) to sperm production (Baer and Boomsma, 2004).
Desert ants of the genus Cataglyphis are well suited to explore the association between seminal fluid production and sperm production. Indeed, (i) the degree of polyandry varies almost 10-fold across species (range of the mean number of male mates: 1.07 – 8.5; Aron et al., 2016) and (ii) sperm production increases with sperm competition intensity (Aron et al., 2016; Degueldre and Aron, 2023). Using phylogenetically controlled analyses, we explored the relationship, and potential trade-off, between accessory gland size—a proxy for seminal fluid production— and sperm production in nine species of Cataglyphis ants experiencing various degrees of sperm competition.
Methods
Sampling
During regional periods of sexual reproduction, we sampled colonies of nine Cataglyphis species across several countries: C. cursor in France; C. hispanica and C. velox in Spain; C. bombycina, C. emmae, and C. viatica in Morocco, and C. livida, C. niger, and C. savignyi in Israel (Table S1). Specifically, we collected male pupae and returned them to the laboratory, where they emerged as adults. Estimates of seminal fluid production and sperm production were obtained from the same males.
Accessory gland size
Because gland volume usually correlates with production (Widdicombe and Wine, 2015), we used AG size as a proxy for seminal fluid production (see also Baer and Boomsma, 2004). Between 8- and 10- days post emergence (i.e., depending on species maturation times), males were decapitated and dissected under a Leica MZ6 stereomicroscope (Leica Microsystems, Wetzlar, Germany) in semen diluent (188.3 mM sodium chloride, 5.6 mM glucose, 574.1 nM arginine, 684.0 nM lysine, and 50 mM tris[hydroxymethyl]aminomethane, pH 8.7; Paynter et al., 2014). All the males had degenerated testes; sperm were found in the accessory testes, indicating their complete maturation.
Each male’s AGs were properly isolated, and carefully placed on a grid in the dissection plate with a drop of semen diluent. They were positioned to measure their greatest length. AGs were photographed using light microscopy (50x magnification) and then measured with a precision of 0.01 mm using IMAGEJ (version 1.8). In all our analyses, absolute AG size was controlled by male body size using analyses of covariance with maximum head width (including eyes) as a covariate (García-Berthou, 2001). Maximum head width is a reliable proxy of body size in Cataglyphis ants (Aron et al., 2016; Lecocq de Pletincx et al., 2021). Males were decapitated and their head measured using the same method as AGs.
Sperm production
Sperm production data for the nine study species were obtained from Degueldre and Aron (2023). Briefly, both accessory testes from each male were dissected in semen diluent. They were placed in 1 ml semen diluent, their membrane was removed and the resulting sperm stream carefully mixed with the diluent. 150 µl of this sperm solution was transferred to an empty 1.5 ml vial; 850 µl of sperm diluent was added to obtain a final volume of 1 ml. This step was repeated to obtain two technical replicates per male. For each sample, cells were quantified by flow cytometry. Sperm populations were identified based on characteristic forward and sideward scatter, enabling total sperm production to be quantified. We used the average of the two technical replicates to estimate sperm production for each male. A square root transformation was used to ensure normality and homoscedasticity of the data.
Paternity frequency
For each species, sperm competition intensity was inferred from paternity frequency (i.e., the number of males that father the offspring of a queen; Aron et al., 2016; Degueldre and Aron, 2023). Data on paternity frequency were obtained from previous studies in which microsatellite loci were used to determine patriline number. For each species, AG size and sperm production were measured from males collected in the same location as that in which paternity frequency was estimated (Table S1).
Phylogenetic analyses
Using species-level data, we evaluated whether paternity frequency was correlated with AG size. To account for potential trait similarities resulting from shared ancestry among species, we utilised phylogenetic generalised least squares (PGLS) regression (Pagel, 1999). In this approach, maximum likelihood ratio tests are used to estimate the degree of phylogenetic association (λ) within the data. When λ values are closer to 0, it is more likely that traits evolved independently. We performed these analyses using the pgls function in the caper package (Freckleton et al., 2002) and employed the Cataglyphis phylogeny (Figure S1; from Lecocq de Pletincx et al., 2021), which was handled using the ape package (Paradis et al., 2004). All analyses were performed in R (v. 4.2.1; R Core Team, 2020).
Results and discussion
We measured AG size for 117 males across the 9 study species (mean number of males per species ± SD = 13 ± 5.79, range: 8–23; mean number of colonies per species ± SD = 2.56 ± 1.01, range: 1–4; mean number of males per colony ± SD = 5.17 ± 2.87, range: 2–16; Table S1). Absolute AG size ranged from 1.17 to 2.23 mm.
Across species, AG size was positively associated with paternity frequency (covariance PGLS: model coefficient = 0.07; R2 = 0.7, p < 0.01; Figure 1). Similarly, AG size was positively correlated with sperm production (partial correlation: ρ = 0.64, p < 0.001; Figure 2). There was a negligible phylogenetic signal for AG size (λ < 0.001, p(Ho: λ=0) = 1; Figure S2). These results strongly suggest a lack of trade-off between sperm production and seminal fluid production. They are consistent with other studies in rodents (Ramm et al., 2005), fishes (Mazzoldi et al., 2005) and moths (Morrow and Gage, 2000) showing a positive association between sperm competition level and the relative size of accessory reproductive glands. Such a lack of trade-off was also reported in Drosophila where testis length is not correlated with accessory gland length (Kraaijeveld et al., 2008).
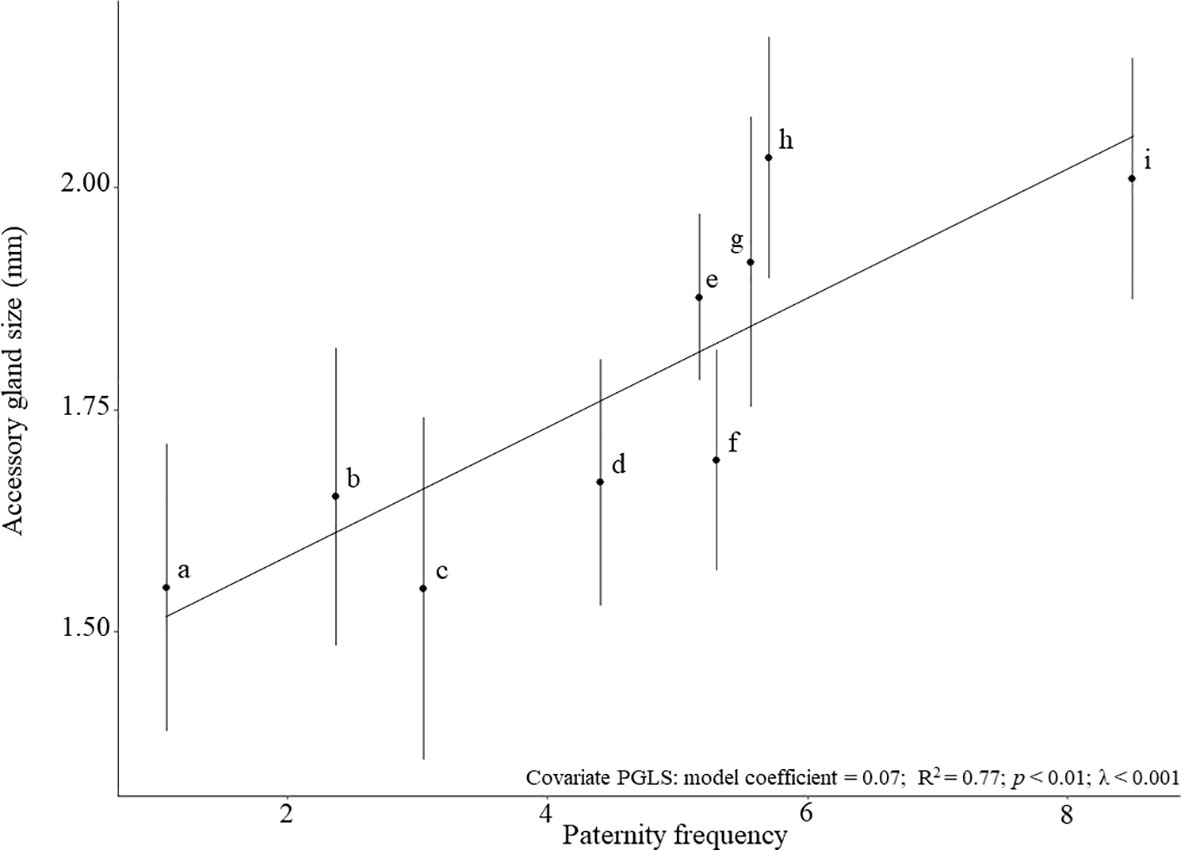
Figure 1 Relationship between paternity frequency and mean AG size (± SD) in nine species of Cataglyphis ants. The linear regression line is shown for AG size. (a) C. hispanica (paternity frequency [Mp] = 1.07; Leniaud et al., 2012); (b) C. velox (Mp = 2.37; Eyer et al., 2013); (c) C. emmae (Mp = 3.05; Jowers et al., 2013); (d) C. livida (Mp = 4.41; Timmermans et al., 2010); (e) C. niger (Mp = 5.17; Leniaud et al., 2011); (f) C. cursor (Mp = 5.3; Pearcy et al., 2009); (g) C. viatica (Mp = 5.56; Aron et al., 2013); (h) C. bombycina (Mp = 5.7; Leniaud et al., 2013); and (i) C. savignyi (Mp = 8.5; Leniaud et al., 2011).
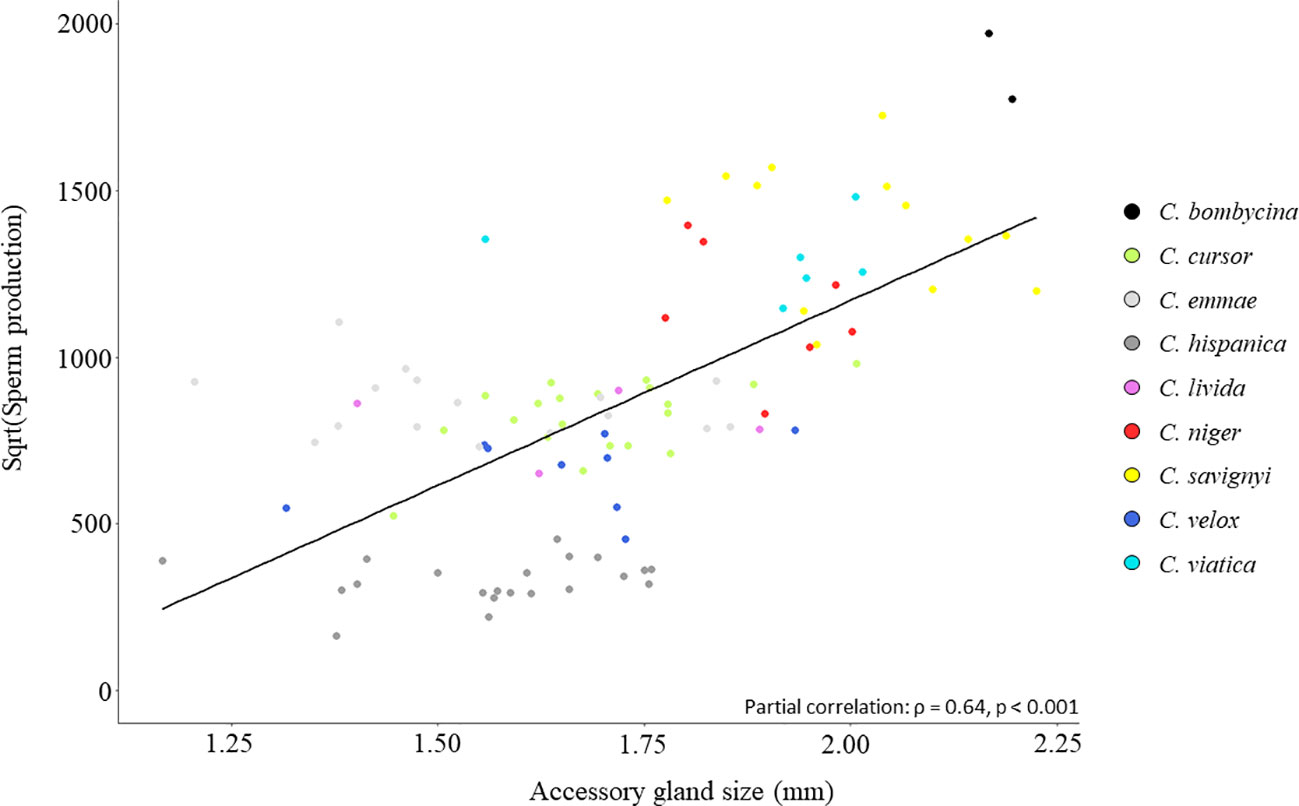
Figure 2 Relationship between sperm production and AG size across all nine Cataglyphis study species. The linear regression line is shown.
Although our results are clearly significant, they must be interpreted with caution because our estimates of paternity frequency are based on data from a single population and year for each study species. Past research has documented the existence of biogeographical variation in population-specific paternity frequencies in various ant species (e.g., Sundström, 1994; Boomsma and van der Have, 1998; Suni and Eldakar, 2011), including in the silver ant, Cataglyphis bombycina (Leniaud et al., 2015). Here, we measured AG size and sperm production for males that came from the population for which paternity frequency had been estimated. To date, there has been no dedicated work exploring interannual variation in paternity frequencies within populations of desert ants, although preliminary findings suggest that such variation is minimal in C. viatica and C. cursor (unpublished data). Beyond these results, it remains unknown how variation in paternity frequency interacts with AG size and sperm production.
In Cataglyphis desert ants, multiple mating by females could drive males to evolutionarily invest in both functional traits. Our findings contrast with those of previous research on fungus-growing ants (Baer and Boomsma, 2004). This work found that, in species where females have multiple mates, males have small AGs and large accessory testes, suggesting they face intense sperm competition and preferentially invest in sperm production. However, it also noted that, in species where females have a single mate, males could employ seminal fluid to form mating plugs, suggesting they may experience lower levels of sperm competition and thus decreased selection pressure on sperm production. Despite the strong association between AGs size, sperm production and paternity frequency in Cataglyphis, other factors than sperm competition may also influence these reproductive traits, such as the efficiency of spermatogenesis (Johnson et al., 2000) and the queen’s sperm storage capacity (Boomsma et al., 2005).
In Cataglyphis, AG size and sperm production are positively associated with sperm competition intensity which suggests that seminal fluid boosts sperm function (Ramm, 2020). In the same vein, it has been shown that sperm competition selects for increased sperm quality in Cataglyphis: in species with highly polyandrous females, males produce more viable sperm with lower levels of DNA fragmentation (Degueldre and Aron, 2023). Seminal fluid could help maintain high levels of sperm quantity and quality by furnishing (i) antioxidant enzymes that limit oxidative stress (Weirich et al., 2002; Collins et al., 2006; Baer et al., 2009); (ii) substances that sustain sperm metabolism (Blum et al., 1962; Alumot et al., 1969); and (iii) antimicrobial and antifungal enzymes (Baer et al., 2009; Peng et al., 2015; Chérasse et al., 2018; Dávila et al., 2018). Seminal fluid could also improve sperm motility after ejaculation, a trait that is essential for successfully reaching the spermatheca (Pearcy et al., 2014; Liberti et al., 2018; Yániz et al., 2020). In the honeybee (Apis mellifera) and leafcutter ants (Acromyrmex echinatior and Atta colombica), females have multiple mates, and a given male’s seminal fluid reduces the survival of sperm from rival males (den Boer et al., 2010), via the action of AG-generated serine proteases (Dosselli et al., 2019). Further research should explore quantitative and/or qualitative differences in the seminal fluid of Cataglyphis species facing differing levels of sperm competition.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because my models are desert ants. They are not species requiring ethical approval.
Author contributions
FD: Conceptualization, Data curation, Formal analysis, Methodology, Writing – original draft, Writing – review & editing. SA: Conceptualization, Funding acquisition, Methodology, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by a FRIA scholarship (FC33495 to FD), the Van Buuren–Jaumotte–Demoulin fund (to FD), and the Belgian National Fund for Scientific Research (FRS-FNRS; grant #J000420 to SA).
Acknowledgments
We thank Xim Cerda, Claude Lebas, Abraham Hefetz, Grégory Philippart and Pauline Dacquin for helping us to collect Cataglyphis samples. We are also grateful to Jessica Pearce-Duvet for her language editing services.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2024.1336015/full#supplementary-material
References
Alumot E., Lensky Y., Holstein P. (1969). Sugars and trehalase in the reproductive organs and hemolymph of the queen and drone honey bees (Apis mellifica L. var. ligustica spin.). Comp. Biochem. Physiol. 28 (3), 1419–1425. doi: 10.1016/0010-406X(69)90579-9
Anderson M. J., Chapman S. J., Videan E. N., Evans E., Fritz J., Stoinski T. S., et al. (2007). Functional evidence for differences in sperm competition in humans and chimpanzees. Am. J. Phys. Anthropol. 134 (2), 274–280. doi: 10.1002/ajpa.20674
Aron S., Darras H., Eyer P. A., Leniaud L., Pearcy M. (2013). Structure génétique des sociétés et systèmes d’accouplement chez la fourmi Cataglyphis viatica (Fabricius 1787). Bull. l’Institut Scientifique Rabat Section Sci. la Vie 35, 103–109.
Aron S., Lybaert P., Baudoux C., Vandervelden M., Fournier D. (2016). Sperm production characteristics vary with level of sperm competition in Cataglyphis desert ants. Funct. Ecol. 30, 614–624. doi: 10.1111/1365-2435.12533
Avila F. W., Sirot L. K., LaFlamme B. A., Rubinstein C. D., Wolfner M. F. (2011). Insect seminal fluid proteins: identification and function. Annu. Rev. Entomol. 56, 21–40. doi: 10.1146/annurev-ento-120709-144823
Baer B. (2011). The copulation biology of ants (Hymenoptera: Formicidae). Myrmecol. News 14, 55–68. doi: 10.25849/myrmecol.news_014:055
Baer B., Boomsma J. J. (2004). Male reproductive investment and queen mating-frequence in fungus-growing ants. Behav. Ecol. 15 (3), 426–432. doi: 10.1093/beheco/arh025
Baer B., Heazlewood L. J., Taylor L. N., Eubel H., Millar H. A. (2009). The seminal fluid of the honeybee Apis mellifera. Proteomics 9, 2085–2097. doi: 10.1002/pmic.200800708
Baer B., Schmid-Hempel P., Høeh J. T., Boomsma J. J. (2003). Sperm length, sperm storage and mating system characteristics in bumblebees. Insectes Soc 50, 101–108. doi: 10.1007/s00040-003-0641-0
Blum M. S., Glowska Z., Taber S. (1962). Chemistry of the drone honey bee reproductive system. II. carbohydrates in the reproductive organs and semen. Ann. Entomol. Soc Am. 55 (1), 135–139. doi: 10.1093/aesa/55.1.135
Boomsma J. J. (2013). Beyond promiscuity: mate-choice commitments in social breeding. Philos. Trans. R. Soc Lond. B Biol. Sci. 368, 20120050. doi: 10.1098/rstb.2012.0050
Boomsma J. J., Baer B., Heinze J. (2005). The evolution of male traits in social insects. Annu. Rev. Entomol. 50, 395–420. doi: 10.1146/annurev.ento.50.071803.130416
Boomsma J. J., Kronauer D. J. C., Pedersen J. S. (2009). “The evolution of social insect mating systems,” in Organization of Insect Societies. Eds. Gadau J., Fewelle J., Wilson E. O. (Cambridge, MA: University Press), 1–25.
Boomsma J. J., van der Have T. M. (1998). Queen mating and paternity variation in the ant Lasius Niger. Mol. Ecol. 7, 1709–1718. doi: 10.1046/j.1365-294x.1998.00504.x
Brown M., Baer B., Schmid-Hempel R., Schmid-Hempel P. (2002). Dynamics of multiple-mating in the bumble bee Bombus hypnorum. Insectes Soc 49, 315–319. doi: 10.1007/pl00012654
Chapman T. (2001). Seminal fluid-mediated fitness traits in Drosophila. Heredity 87, 511–521. doi: 10.1046/j.1365-2540.2001.00961.x
Chérasse S., Baer B., Schiott M., Boomsma J. J. (2018). Differential immune gene expression in sperm storage organs of leaf-cutting ants. J. Exp. Biol. 221, 173435. doi: 10.1242/jeb.173435
Collins A. M., Caperna T. J., Williams V., Garret W. M., Evans J. D. (2006). Proteomics analyses of male contributions to honey bee sperm storage and mating. Insect Mol. Biol. 15 (5), 541–549. doi: 10.1111/j.1365-2583.2006.00674.x
Dávila F., Botteaux A., Bauman D., Chérasse S., Aron S. (2018). Antibacterial activity of male and female sperm-storage organs in ants. J. Exp. Biol. 221 (6), jeb175158. doi: 10.1242/jeb.175158
Degueldre F., Aron S. (2023). Sperm competition increases sperm production and quality in Cataglyphis desert ants. Proc. Biol. Sci. 290 (1995), 20230216. doi: 10.1098/rspb.2023.0216
den Boer S. P., Baer B., Boomsma J. J. (2010). Seminal fluid mediates ejaculate competition in social insects. Science 327, 1506–1509. doi: 10.1126/science.1184709
den Boer S. P., Boomsma J. J., Baer B. (2009). Honey bee males and queens use glandular secretions to enhance sperm viability before and after storage. J. Insect Physiol. 55 (6), 538–543. doi: 10.1016/j.jinsphys.2009.01.012
den Boer S. P., Stürup M., Boomsma J. J., Baer B. (2015). The ejaculatory biology of leafcutter ants. J. Insect Physiol. 74, 56–62. doi: 10.1016/j.jinsphys.2015.02.006
Dixson A. F. (1998). Sexual selection and evolution of the seminal vesicles in primates. Folia Primatol. 69 (5), 300–306. doi: 10.1159/000021643
Dosselli R., Grassl J., den Boer S. P., Kratz M., Moran J. M., Boomsma J. J., et al. (2019). Protein-level interactions as mediators of sexual conflicts in ants. Mol. Cell. Proteomics 18, S34–S45. doi: 10.1074/mcp.RA118.000941
Duvoisin N., Baer B., Schmid-Hempel P. (1999). Sperm transfer and male competition in a bumblebee. Anim. Behav. 58, 743–749. doi: 10.1006/anbe.1999.1196
Eyer P. A., Leniaud L., Darras H., Aron S. (2013). Hybridogenesis through thelytokous parthenogenesis in two Cataglyphis desert ants. Mol. Ecol. 22, 947–955. doi: 10.1111/mec.12141
Firman R. C., Klemme I., Simmons L. W. (2013). Strategic adjustments in sperm production within and between two island populations of house mice. Evolution 67 (10), 3061–3070. doi: 10.1111/evo.12164
Freckleton R. P., Harvey P. H., Pagel M. (2002). Phylogenetic analysis and comparative data: a test and review of evidence. Am. Nat. 160, 712–726. doi: 10.1086/343873
Gage M. J. G., Morrow E. H. (2003). Experimental evidence for the evolution of numerous, tiny sperm via sperm competition. Curr. Biol. 13, 754–757. doi: 10.1016/S0960-9822(03)00282-3
García-Berthou E. (2001). On the misuse of residuals in ecology: testing regression residuals vs. the analysis of covariance. J. Anim. Ecol. 70, 708–711. doi: 10.1046/j.1365-2656.2001.00524.x
Gomendio M., Harcourt A. H., Roldan E. R. S. (1998). “Sperm competition in mammals,” in Sperm Competition and Sexual Selection. Eds. Birkhead T. R., Moller A. P. (San Diego, CA: Academic Press), 667–756.
Gomendio M., Roldan E. R. S. (1991). Sperm size and sperm competition in mammals. Proc. Biol. Sci. 243, 181–185. doi: 10.1098/rspb.1991.0029
Gorshkov V., Blenau W., Koeniger G., Römpp A., Vilcinskas A., Spengler B. (2015). Protein and peptide composition of male accessory glands of Apis mellifera drones investigated by mass spectrometry. PloS One 10 (5), e0125068. doi: 10.1371/journal.pone.0125068
Hughes W. O., Oldroyd B. P., Beekman M., Ratnieks F. L. W. (2008). Ancestral monogamy shows kin selection is key to the evolution of eusociality. Science 320, 1213–1216. doi: 10.1126/science.1156108
Immler S., Pitncik S., Parker G. A., Durrant K. L., Lüpold S., Calhim S., et al. (2011). Resolving variation in the reproductive tradeoff between sperm size and number. Proc. Natl. Acad. Sci. U.S.A. 108 (13), 5325–5330. doi: 10.1073/pnas.100905910
Johnson L., Varner D. D., Roberts M. E., Smith T. L., Keillor G. E., Scrutchfield W. L. (2000). Efficiency of spermatogenesis: a comparative approach. Anim. Reprod. Sci. 60-61, 471–480. doi: 10.1016/s0378-4320(00)00108-1
Jowers M. J., Leniaud L., Cerda X., Alasaad S., Caut S., Amor F., et al. (2013). Social and population structure in the ant Cataglyphis emmae. PloS One 8, e72941. doi: 10.1371/journal.pone.0072941
King M., Eubel H., Millar A. H., Baer B. (2011). Proteins within the seminal fluid are crucial to keep sperm viable in the honeybee Apis mellifera. J. Insect Physiol. 57 (3), 409–414. doi: 10.1016/j.jinsphys.2010.12.011
Kraaijeveld K., Denniff M., Baker R. H., Chapman T. (2008). The evolution of sperm and non-sperm producing organs in male Drosophila. Biol. J. Linn. Soc 94, 505–512. doi: 10.1111/j.1095-8312.2008.01030.x
Lecocq de Pletincx N., Dellicour S., Aron S. (2021). The evolution of ant worker polymorphism correlates with multiple social traits. Behav. Ecol. Sociobiol. 75 (8), 1–11. doi: 10.1007/s00265-021-03049-6
Leniaud L., Darras H., Boulay R., Aron S. (2012). Social hybridogenesis in the clonal ant Cataglyphis hispanica. Curr. Biol. 22, 1188–1193. doi: 10.1016/j.cub.2012.04.060
Leniaud L., Heftez A., Grumiau L., Aron S. (2011). Multiple mating and supercoloniality in Cataglyphis desert ants. Biol. J. Linn. Soc 104, 866–876. doi: 10.1111/j.1095-8312.2011.01772.x
Leniaud L., Pearcy M., Aron S. (2013). Sociogenetic organisation of two desert ants. Insectes Soc 60, 337–344. doi: 10.1007/s00040-013-0298-2
Leniaud L., Pearcy M., Taheri A., Aron S. (2015). Testing the genetic determination of the soldier caste in the silver ant. Insectes Soc 65, 517–524. doi: 10.1007/s00040-015-0431-5
Liberti J., Baer B., Boomsma J. J. (2018). Rival seminal fluid induces enhanced sperm motility in a polyandrous ant. BMC Evol. Biol. 18, 28. doi: 10.1186/s12862-018-1144-y
Liberti J., Görner J., Welch M., Dosselli R., Schiøtt M., Ogawa Y., et al. (2019). Seminal fluid compromises visual perception in honeybee queens reducing their survival during additional mating flights. eLife 8, e45009. doi: 10.7554/eLife.45009
Lu J. C., Jing J., Yao Q., Fan K., Wang G. H., Feng R. X., et al. (2016). Relationship between lipids levels of serum and seminal plasma and semen parameters in 631 Chinese subfertile men. PloS One 11 (1), e0146304. doi: 10.1371/journal.pone.0146304
Manaskova P., Ryslava H., Ticha M., Jonakova V. (2002). Characterization of proteins from boar prostate. Am. J. Reprod. Immunol. 48, 283–290. doi: 10.1034/j.1600-0897.2002.01138.x
Mazzoldi C., Petersen C. W., Rasotto M. B. (2005). The influence of mating system on seminal vesicle variability among gobies (Teleostei, Gobiidae). J. Zoolog. Syst. Evol. Res. 43 (4), 307–314. doi: 10.1111/j.1439-0469.2005.00331.x
Morrow E. H., Gage M. J. (2000). The evolution of sperm length in moths. Proc. Biol. Sci. 267 (1440), 307–313. doi: 10.1098/rspb.2000.1001
Nakanishi A., Takegaki T. (2018). Tactic-specific sperm traits in the dusky frillgoby (Bathygobius fuscus). J. Zool. 307, 71–77. doi: 10.1111/jzo.12619
Owen D. H., Katz D. F. (2005). A review of the physical and chemical properties of human semen and the formulation of a semen simulant. J. Androl. 26, 459–469. doi: 10.2164/jandrol.04104
Pagel M. (1999). Inferring the historical patterns of biological evolution. Nature 401, 877–884. doi: 10.1038/44766
Paradis E., Claude J., Strimmer K. (2004). APE: analyses of phylogenetics and evolution in R language. Bioinformatics 20, 289–290. doi: 10.1093/bioinformatics/btg412
Parker G. A. (1970). Sperm competition and its evolutionary consequences in the insects. Biol. Rev. Cambridge Philos. Soc 45, 525–567. doi: 10.1111/j.1469-185X.1970.tb01176.x
Paynter E., Baer-Imhoof B., Linden M., Lee-Pullen T., Heel K., Rigby P., et al. (2014). Flow cytometry as a rapid and reliable method to quantify sperm viability in the honeybee Apis mellifera. Cytometry A 85 (5), 463–472. doi: 10.1002/cyto.a.22462
Pearcy M., Delescaille N., Lybaert P., Aron S. (2014). Team swimming in ant spermatozoa. Biol. Lett. 10, 20140308. doi: 10.1098/rsbl.2014.0308
Pearcy M., Timmermans I., Allard D., Aron S. (2009). Multiple mating in the ant Cataglyphis cursor: testing the sperm limitation and the diploid male load hypotheses. Insectes Soc 56, 94–102. doi: 10.1007/s00040-008-1043-0
Peng Y., Grassl J., Millar A. H., Boris B. (2015). Seminal fluid of honeybees contains multiple mechanisms to combat infections of the sexually transmitted pathogen Nosema apis. Proc. Biol. Sci. 283, 20151785. doi: 10.1098/rspb.2015.1785
Ramm S. A. (2020). Seminal fluid and accessory male investment in sperm competition. Philos. Trans. R. Soc Lond. B Biol. Sci. 375 (1813), 20200068. doi: 10.1098/rstb.2020.0068
Ramm S. A., Parker G. A., Stockley P. (2005). Sperm competition and the evolution of male reproductive anatomy in rodents. Proc. Biol. Sci. 272 (1566), 949–955. doi: 10.1098/rspb.2004.3048
R Core Team (2020) R: A language and environment for statistical computing. Available at: https://www.r-project.org/.
Santiago-Moreno J., Blesbois E. (2020). Functional aspects of seminal plasma in bird reproduction. Int. J. Mol. Sci. 21 (16), 5664. doi: 10.3390/ijms21165664
Simmons L. W., Gwynne D. T. (1991). The refractory period of female katydids (Orthoptera: Tettigoniidae): sexual conflict over the remating interval. Behav. Ecol. 2, 276–282. doi: 10.1093/beheco/2.4.276
Smith J. R., Stanfield G. M. (2012). A seminal fluid protease activates sperm motility in C. elegans males. Worm 1 (3), 151–154. doi: 10.4161/worm.19502
Sundström L. (1994). Sex ratio bias, relatedness asymmetry and queen mating frequency in ants. Nature 367, 266–268. doi: 10.1038/367266a0
Suni S. S., Eldakar O. T. (2011). High mating frequency and variation with lineage ratio in dependent-lineage harvester ants. Insect Soc 58, 357–364. doi: 10.1007/s00040-011-0150-5
Timmermans I., Grumiau L., Hefetz A., Aron S. (2010). Mating system and population structure in the desert ant Cataglyphis livida. Insectes Soc 57, 39–46. doi: 10.1007/s00040-009-0048-7
Tourmente M., Gomendio M., Roldan E. R. S. (2011). Sperm competition and the evolution of sperm design in mammals. BMC Evol. Biol. 11, 12. doi: 10.1186/1471-2148-11-12
Weirich F. G., Collins M. A., Williams P. V. (2002). Antioxidant enzymes in the honey bee, Apis mellifera. Apidologie 33, 3–14. doi: 10.1051/apido:2001001
Widdicombe J. H., Wine J. J. (2015). Airway gland structure and function. Physiol. Rev. 95 (4), 1241–1319. doi: 10.1152/physrev.00039.2014
Wolfner M. (2002). The gifts that keep on giving: physiological functions and evolutionary dynamics of male seminal proteins in Drosophila. Heredity 88, 85–93. doi: 10.1038/sj.hdy.6800017
Keywords: sperm competition, accessory glands, sperm production, ants, eusocial hymenopterans
Citation: Degueldre F and Aron S (2024) Accessory gland size increases with sperm competition intensity in Cataglyphis desert ants. Front. Ecol. Evol. 12:1336015. doi: 10.3389/fevo.2024.1336015
Received: 09 November 2023; Accepted: 15 January 2024;
Published: 26 January 2024.
Edited by:
Francisco Garcia-Gonzalez, Spanish National Research Council (CSIC), SpainReviewed by:
Bodhisatta Nandy, Indian Institute of Science Education and Research Berhampur (IISER), IndiaBahar Patlar, Martin Luther University of Halle-Wittenberg, Germany
Copyright © 2024 Degueldre and Aron. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Félicien Degueldre, RmVsaWNpZW4uRGVndWVsZHJlQHVsYi5iZQ==