 Michal Hradec
Michal Hradec Gudrun Illmann
Gudrun Illmann Martina Prikrylová
Martina Prikrylová Petra Bolechová
Petra Bolechová Hana Vostrá-Vydrová1
Hana Vostrá-Vydrová1- 1Department of Ethology and Companion Animal Science, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Prague, Czechia
- 2Department of Ethology, Institute of Animal Science, Praha Uhríněves, Prague, Czechia
All gibbons (Primates: Hylobatidae) are well known for emitting loud vocalizations specific for species and sex. The songs of paired and unpaired male southern yellow-cheeked gibbons (Nomascus gabriellae) are characterized by the presence of staccato notes and multi-modulation phrases with two or more extremely rapid frequency modulations in the second note of each phrase. In addition, paired males also produce a coda vocalization, which is similar to the multi-modulation phrase of male calls but lacks the staccato notes and always occurs directly following the female great call as part of a pair-specific duet. The aim of this study was first to assess whether the songs of paired and unpaired males can be acoustically distinguished from one another and second, whether the coda vocalization differs from the multi-modulation phrase of the male call in paired males. To assess these issues, we analyzed 616 songs obtained from a long-term study of vocal development in 14 captive adult males (>7 years old), half of which were unpaired and significantly younger than paired subjects. For each song, we quantified nine acoustic features, for which we applied a suite of linear mixed effects models with social status as a fixed variable and age as a regression coefficient. This allowed us to compare (1) the structure of male calls (staccato notes and multi-modulation phrase) between paired and unpaired subjects, (2) the muti-modulation phrase of unpaired subjects to the coda vocalization of paired subjects, and (3) the multi-modulation phrase of paired males to the coda vocalization. We found that the male call of younger-unpaired subjects had a longer duration, broader frequency range, higher maximum frequency, and fewer staccato notes than their counterparts in paired subjects. The coda vocalization of older-paired males exhibited a larger number of frequency modulations than the multi-modulation phrase of all males. While the male call of younger-unpaired males differs from both the male call and the coda vocalization of older-paired males, further studies are necessary to disentangle the effects of age and pairing status.
1. Introduction
Vocal communication, unlike visual signaling, can convey information about the sender over long distances. Such information includes identity (Owren and Rendall, 2003; Price et al., 2009), social rank (Fischer et al., 2004), sex, and body size (Pfefferle and Fischer, 2006; Ey et al., 2007). Along with physical growth, changes in vocalization with increasing age were confirmed in birds (Nemeth et al., 2012) as well as in aquatic (Sanvito et al., 2008; Umeed et al., 2018) and terrestrial mammals (Reby and McComb, 2003; Charlton et al., 2009; Briefer et al., 2010).
Although various forms of vocal signaling have been documented in several mammalian taxa, such as primates, rodents, bats, and cetaceans (for recent reviews see Banerjee et al., 2019; Vernes and Wilkinson, 2020; Janik and Knornschild, 2021; De Gregorio et al., 2022; Vanderhoff and Hoverud, 2022), the critical acoustic features involved during inter-individual vocal exchanges, duetting included, are little known. Some advances have been made in singing primates, notably in Sulawesi tarsiers (genus Tarsius, Clink et al., 2020), indris (Indri indri, Torti et al., 2013), coppery titi monkeys (Plecturocebus cupreus, Lau et al., 2020), and gibbons (Geissmann, 2002). Additional studies have focused on song dynamics (e.g., rhythm, pitch, and degree of overlap) (Gamba et al., 2016), song flexibility (Clarke et al., 2006; Terleph et al., 2018a; Hradec et al., 2021a), song individuality (Sun et al., 2011; Clink et al., 2021), song ontogeny (Hauser, 1989; Pistorio et al., 2006; Hradec et al., 2017, 2021b; De Gregorio et al., 2021), song recognition (Raemaekers and Raemaekers, 1985; Mitani, 1987; Caselli et al., 2015), song taxonomy, and genetic relatedness (Konrad and Geissmann, 2006; Torti et al., 2017). However, the question of whether songs by singing primates transmit information about pairing status (i.e., paired vs. unpaired individuals) has received surprisingly little attention.
Gibbons (family Hylobatidae) are a uniform group of territorial and mostly pair-living apes that are well known for emitting loud, stable patterns of vocalizations specific for species and sex (Geissmann, 2002; but see Reichard et al., 2016; De Gregorio et al., 2022). The vocalizations are often referred to as “songs”, uttered in succession and forming a recognizable temporal sequence of pure, melodic sounds (Thorpe, 1961; Haimoff, 1984a; Geissmann, 2002; Supplementary Table 1).
Gibbon songs are relatively stereotyped and are thought to be under strong genetic constraints (Brockelman and Schilling, 1984; Geissmann, 1984). For most gibbon species, paired individuals (i.e., adult male and adult female) often combine their respective songs into coordinated duets, except for the silvery gibbon (Hylobates moloch) and the Kloss's gibbon (Hylobates klossii) in which paired individuals produce only solo songs (Tenaza, 1976; Geissmann and Nijman, 2006). Duet songs are an alternation of sex-specific vocalizations contributed to by the female and her mate (Haimoff, 1984a). A typical male-female gibbon duet begins with a few introductory notes from both mates, followed by the female great call. During the build-up phase of the female great call, the paired male ceases his song and, after the completion of the great call, adds a coda vocalization. The coda vocalization is produced only by paired males and always occurs directly following the female great call (Geissmann, 2002). Subsequently, the paired male repeats several male calls during the so-called interlude sequence (Figure 1) until the female begins her next great call (Geissmann, 2002; Konrad and Geissmann, 2006).
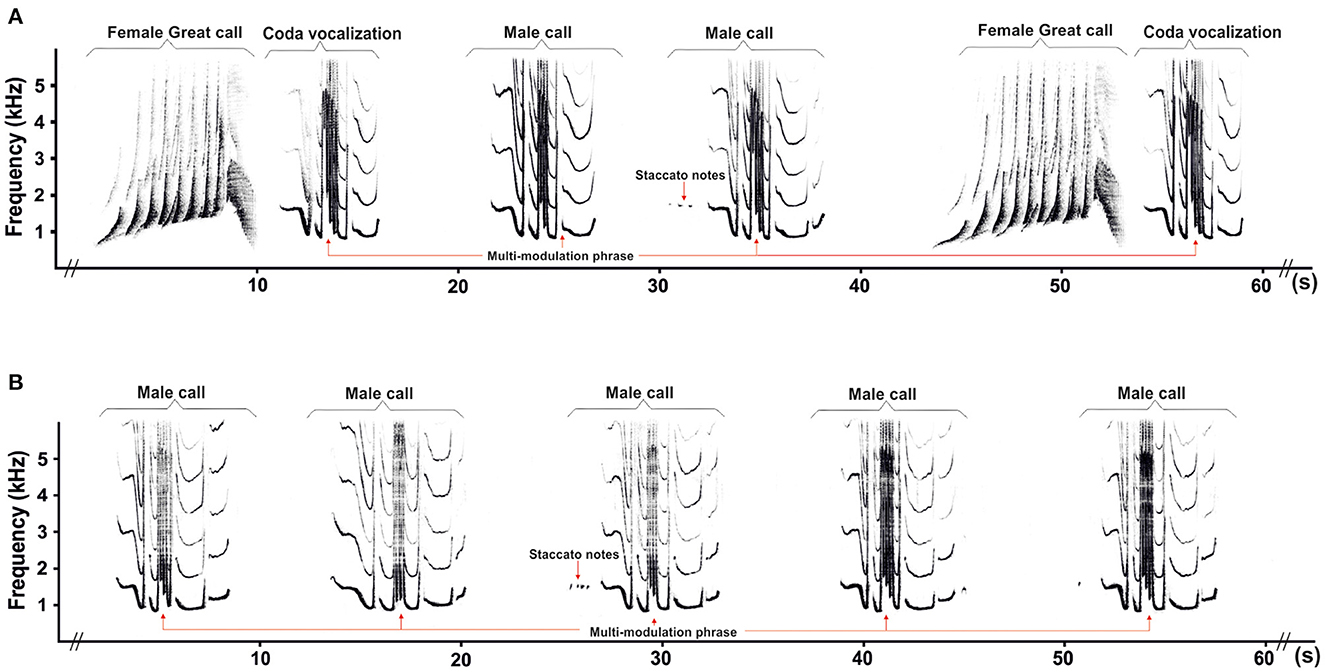
Figure 1. Representative spectrograms showing a duet song (A) and an interlude sequence of a solo song (B) in southern yellow-cheeked gibbons.
The vocal structure of the coda vocalization and the male call has been studied in several gibbon species (Geissmann, 2002; Konrad and Geissmann, 2006; Terleph et al., 2018b). The stable structure of the coda vocalization has been found to be particularly useful in determining individuality and taxonomy in male gibbons, both in the absence of the male call (Sun et al., 2011; Lau et al., 2018; Clink et al., 2021) and in conjunction with the male call (Wanelik et al., 2013). However, recent studies in white-handed gibbons (Hylobates lar) have shown that the stable structure of both the male call and the female great call declines with age (Barelli et al., 2013; Terleph et al., 2016).
Wild unpaired Kloss's gibbons (Hylobates klossii) and white-handed gibbons (Hylobates lar) aged 8–9 years disperse from their natal group and become “floaters”, i.e., individuals in search of a mate and territory (Tilson, 1981; Brockelman and Gittins, 1984; Cowlishaw, 1992). These unpaired adult males emit loud calls, exclusively as a solo song, without the coda vocalizations (Brockelman and Srikosamatara, 1984; Mitani, 1988; Cowlishaw, 1992). This raises the question of whether male calls differ according to pairing status, which could then be used by conspecific (female) receivers to assess the emitter's social status. The only available study (Mitani, 1988) did not find any evidence of such differences between the songs of paired and unpaired male agile gibbons (Hylobates agilis). In addition, playback experiments revealed that females (and other individuals) failed to distinguish paired males from unpaired males, based solely on vocal cues (Mitani, 1988). In non-human primates, paired males are usually older than unpaired males, at least in captivity, and it is also known that the acoustic parameters of vocalization change with advancing age (Simakobu monkeys: Erb et al., 2013; and titi monkeys: Clink et al., 2019; e.g., Japanese macaques: Inoue, 1988) or age range (e.g., baboons: Fischer et al., 2002, 2004). In fact, an age-related decline in acoustic parameters (i.e., duration and frequency) has been shown in paired males and females of white-handed gibbons (Barelli et al., 2013; Terleph et al., 2016). However, the influence of age on acoustic structure in unpaired males is completely unknown.
Given such limited information on male calls, it might be worth exploring, in other gibbon species, whether male songs differ according to pairing status or because of aging. To do this, we analyzed available data from a long-term study of vocalizations in captive southern yellow-cheeked gibbons (Nomascus gabriellae). In both paired and unpaired individuals of this species, the male call is characterized by the presence of irregular series of soft staccato notes that sometimes precede a multi-modulation phrase (Figures 1A, B). A previous report in the same species established that, when reaching 7.1 years of age, unpaired males produce the complete male call as a solo song (Hradec et al., 2021b), while paired males include it in duets with females (Haimoff, 1984a; Geissmann, 2002; Konrad and Geissmann, 2006).
In addition, paired males also produce a coda vocalization (Figure 1A) which is similar to the multi-modulation phrase in solo and duet songs of a male call but has a more stereotyped structure, and it is always produced immediately after the female great call without staccato notes (Geissmann, 2002). The multi-modulation phrase is the predominant acoustic structure of the male call and is divided into several notes. The second note of the multi-modulation phrase and the coda vocalization feature two or more extremely rapid frequency modulations (a steep up-and-down sweeping) that set southern yellow-cheeked gibbons apart from other species of the genus Nomascus (Konrad and Geissmann, 2006).
It has been suggested that the extremely rapid changes in the frequency modulations in the second note of the multi-modulation phrase and coda vocalization (also called “roulade”; Demars and Goustard, 1978 or “roll”; Konrad and Geissmann, 2006) are reminiscent of songbird trills. The high repetition rate of those trills is costly to produce and might be considered an accurate signal of male quality (Gil and Gahr, 2002; Ballentine, 2009; Cramer, 2013) but we do not know whether this applies to females. Among non-human primates, a trade-off between call rate and frequency bandwidth has been first reported in the trills of the female great call of Bornean gibbons (Hylobates muelleri) but whether these performance constraints inform recipients of female quality remains to be investigated (Clink et al., 2018). At the same time, it has been shown that acoustic signals of longer durations, higher frequencies, and wider bandwidths are reliable indicators of the unpaired status of male rufous-and-white wrens, (Thryothorus rufalbus), that, in turn, could prove attractive to females (Hennin et al., 2009).
To date, it is still unclear whether structural differences exist in the multi-modulation phrase between paired and unpaired southern yellow-cheeked gibbon males. For instance, male calls displaying a wider frequency range and longer duration might be an indicator of unpaired status or else might reflect an age difference. Furthermore, it is essentially unknown whether the coda vocalization—as an integral part of the duet—structurally differs from the multi-modulation phrase of both paired (in duet song) and unpaired (in solo song) individuals. Such differences have been suggested based on the visual inspection of the calls. For instance, the coda vocalization of the Hainan black gibbon, Hylobates concolor hainanus—since renamed Hainan gibbon (Nomascus hainanus)—has a longer duration and displays a higher number of frequency modulations in the second note than that found in the multi-modulation phrase of the male call (Haimoff, 1984a,b). This indicates that, in paired individuals, the coda vocalization is structurally more complex. However, an in-depth analysis of the acoustical structure comparing the coda vocalization to male calls is still lacking.
The present study is based on a unique but unequally distributed dataset in which unpaired adult males were significantly younger (age: 7.7–10.10 years) than paired males (age: 11.4–34 years), which may not necessarily reflect the situation in the wild (Mitani, 1988). Such artificial distribution in the group composition of zoo-housed animals relies on the breeding management recommendations, as stated by the European Association of Zoos and Aquaria (EAZA) Ex-situ Program (EEP). The possibility of obtaining vocal samples from both paired and unpaired males that would overlap in age distribution is thus limited by the number of animals kept and the EEP regulations. This study is the first of its kind to compare the acoustical structure of unpaired and paired southern yellow-cheeked gibbon males while considering the age factor.
Our aim is to assess (1) whether social status and age synergically influence the song structure in these two groups of captive male southern yellow-cheeked gibbons namely by comparing a set of acoustic features in the male call and coda vocalizations of older-paired vs. younger-unpaired males and (2) whether the coda vocalization, indeed, differs from the male calls among older-paired males of the same species, as was found in the Hainan gibbon (Haimoff, 1984a,b). First, we predicted that, in comparison to older-paired males, younger-unpaired males would produce songs characterized by a higher pitch and longer duration, predominantly in the second note of the multi-modulation phrase. Second, we also predicted that focusing only on paired males, the coda vocalization would differ from the multi-modulation phrase in having a higher maximum frequency, a longer total duration, and a larger number of frequency modulations in the second note.
2. Materials and methods
2.1. Subjects under study
This study was conducted in Czech, Slovak, and Slovenian zoological parks and involved 14 southern yellow-cheeked gibbons (all adult males) distributed in nine groups (Supplementary Table 2, Figure 1). Four of the 14 subjects were born in the wild (Vietnam) and transported as juveniles to the European zoological parks in the late 1980s. The remaining 10 individuals were born in captivity. Seven of the 14 subjects were unpaired males, and the remaining seven were paired males that were housed with their mate (group 4) or as a family group (an adult pair and offspring). The Jihlava and Olomouc zoological parks each had two groups of gibbons that remained in visual and auditory contact with each other. Group 2 at the Jihlava Zoo comprised one adult female and her two male offspring of different ages; the adult male died in 2009. Group 7 at the Košice Zoo comprised two unpaired males (brothers); both were raised in a family group up to 5.5 years of age and 7.5 years of age, respectively (Birot, 2022). Although some individuals were related, in this study, the sample size was too small (N = 7 unpaired males) to include relatedness in the statistical model. Each group had permanent access to an indoor and outdoor enclosure or was confined to an island (group 9). The gibbons were fed four times per day, and their diet consisted of fruits, vegetables, seeds, leaves, cereals, and eggs. Water was available ad libitum. Further information about indoor and outdoor enclosure, as well as the overall composition of family groups, is available in Supplementary Tables 3, 4.
2.2. Data collection and acoustic analysis
This research was part of a long-term study focusing on the vocal behavior of captive Nomascus gibbons (Hradec et al., 2016, 2017, 2021a,b). Acoustic data were collected on the first and sixth visitations that took place between 2014 and 2021. All of the adult males were observed for 1 to 3 days during each observation. We classified all of the males as either older-paired or younger-unpaired (Supplementary Table 2, Figure 1). In our study, unpaired males are younger (7.7–10.10 years), and paired males are older (11.4–34 years), which indicates that the age of paired and unpaired males did not overlap. Consequently, the social effect cannot be differentiated from the age effect. Older-paired males and their female partner would regularly emit a duet song, typically in the morning (5:00 to 10:00 a.m.), which lasted approximately 10–25 min. Younger-unpaired males emitted only solo songs (i.e., male call) in the morning (5:20–11:00 a.m.), and which lasted 8–30 min.
Younger-unpaired males who lived in family groups emitted solo singing after the parental duets. Where two brothers shared the same enclosure (groups 2 and 6; Supplementary Table 2), these males sang solo songs and produced them independently from each other. All 14 subjects produced a fully matured male vocal pattern, as revealed by the production of staccato notes followed by a multi-modulation phrase exhibiting two or more frequency modulations on the second note (Hradec et al., 2021b). Our dataset derives from 34 recording sessions targeting 7 adult pairs and 17 recording sessions targeting 7 younger-unpaired males, making for a total of 51 recording sessions. From this pool of recordings, we extracted 616 male songs (older-paired: N = 374 songs comprising 184 multi-modulation phrases and 190 coda vocalizations; younger-unpaired: N = 242 songs). In older-paired subjects, we analyzed 190 codas and 184 male calls, 101 of which were preceded by staccato notes. In younger-unpaired subjects, we analyzed 242 male calls, 29 of which were preceded by staccato notes (Table 1).
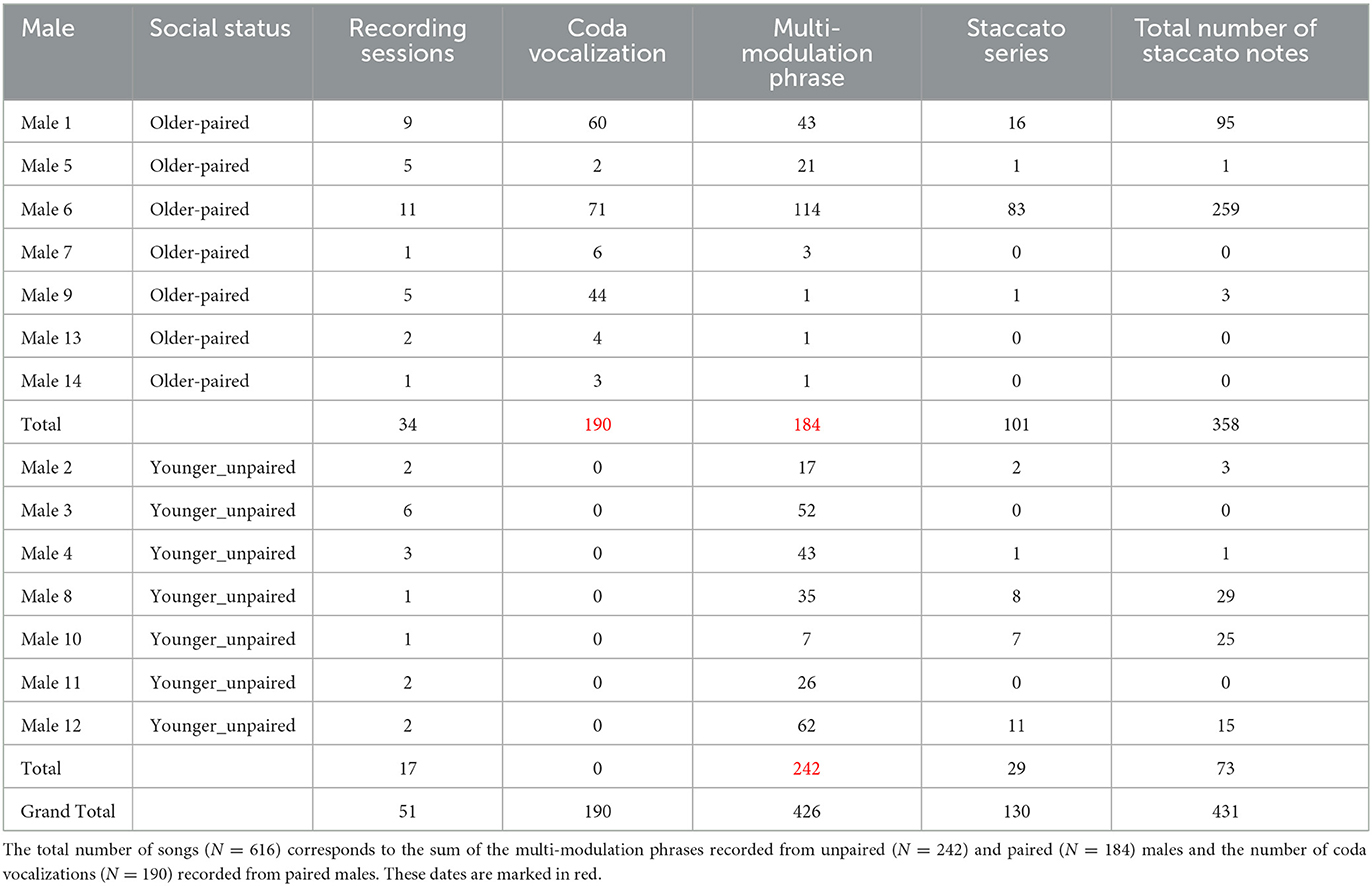
Table 1. Information on the number of songs (i.e., staccato notes, multi-modulation phrases, and coda vocalizations) in older-paired and younger-unpaired males.
We recorded the male vocalizations on an M-Audio Micro Track II recorder or a Marantz PMD661 with a Rode NTG-2 semi-directional microphone (in mono at 16-bit resolution, 44.1 kHz sampling rate) at a distance of 2 to 10 m in an outdoor enclosure or island. All of the recordings were saved as waveform audio files. Acoustic analysis was carried out using Avisoft SASLab Pro version 5.2 software (Avisoft Bioacoustics, Berlin, Germany). Spectrograms were generated under the following settings: FFT length = 1,024; frequency resolution = 12 Hz; temporal resolution = 21.3 ms; overlap = 75%; and window type = Hamming.
To examine the structural differences between male calls (staccato notes and multi-modulated phrases) and coda vocalization as a function of pairing status, we selected the nine acoustic parameters presented in Figure 2. We quantified these features in Avisoft SASLab Pro following the prior annotations of each note and call via visual inspection of spectrograms. Previous studies in the genus Nomascus (Schilling, 1984; Konrad and Geissmann, 2006; Thinh et al., 2011) have established that both the male call and the coda are made up of three notes, i.e., Note 1, Note 2, and Last notes (Figure 2). The Last notes consist of one or several notes. If more notes were produced, we did not divide them further; they were treated as one entity. Importantly, the second note of the multi-modulation phrase includes rapid frequency modulations consisting of a steep up-and-down sweep (Konrad and Geissmann, 2006; Hradec et al., 2021b), which is more flexible than the overall pattern of a multi-modulation phrase. Therefore, our analysis of frequency parameters focused on this second note and included only those notes that had two or more modulation frequencies. From this pool of notes, we measured the maximum frequency (kHz), minimum frequency (kHz), and frequency range (kHz), i.e., the difference between the maximum and minimum frequency of the second note. As for temporal parameters, we measured the total duration of the multi-modulation phrase (all three notes included) and the duration of the second note. Finally, the presence or absence of staccato notes in male calls was also quantified due to their considerable individual variation during singing. When present, we counted the number of staccato notes at the beginning of a song.
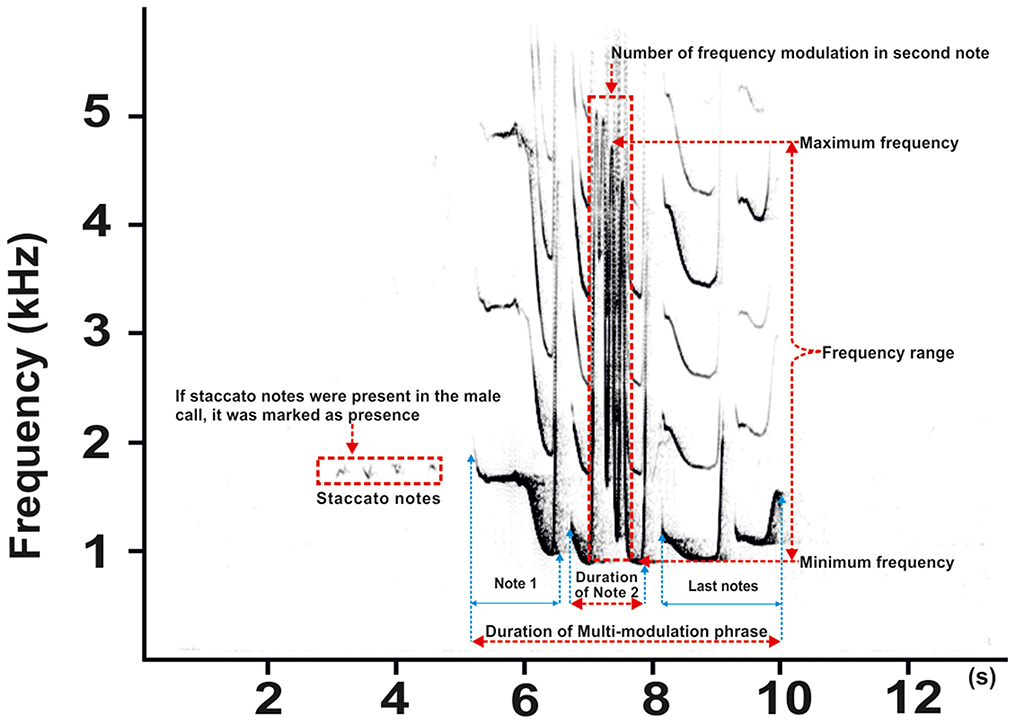
Figure 2. Representative spectrogram male call of southern yellow-cheeked gibbons (adapted from Hradec et al., 2017) displaying staccato notes and a multi-modulation phrase (divided into three notes). The most rapid change in frequency modulation is evident in the second note of the steep up-and-down sweeping sound (red dashed rectangle). Red dashed lines indicate the measured parameters of the multi-modulation phrase.
2.3. Statistical analysis
All data were analyzed with the aid of SAS software version 9.4 (SAS Institute Inc., Cary, NC). Results with a p-value of less than 0.05 (P ≤ 0.05) were considered statistically significant. We applied six statistical Linear Mixed Models fit by REML (restricted maximum likelihood) to test for differences in the vocal structure of the following: (1) male calls in younger-unpaired and older-paired males. Seven features were tested (Figure 2): total duration of multi-modulation phrase (s); duration of the second note (s); maximum frequency (kHz); minimum frequency (kHz); frequency range (kHz); the number of frequency modulations in the second note; and presence of staccato notes; (2) the multi-modulation phrases in younger-unpaired males and coda vocalizations. Six features were tested: total duration of multi-modulation phrase (s); duration of the second note (s); maximum frequency (kHz); minimum frequency (kHz); frequency range (kHz); and the number of frequency modulations in the second note; and (3) the multi-modulation phrases in older-paired males and coda vocalizations. Six features were tested: total duration of multi-modulation phrase (s); duration of the second note (s); maximum frequency (kHz); minimum frequency (kHz); frequency range (kHz); and the number of frequency modulations in the second note. The structure of the multi-modulation phrase in the male calls of both younger-unpaired and older-paired males was similar to that of the coda vocalizations in older-paired males. Because most of our subjects were sampled multiple times, statistical differences in the multi-modulation phrase and coda vocalizations between younger-unpaired and older-paired males were analyzed by a mixed linear model procedure (PROC MIXED) with repeated measurements. The data were analyzed in two steps: In the first step, the effect of social status (older-paired vs. younger-unpaired) was analyzed by the following model:
In the second step, the data set was divided into two subsets based on the determined differences in social status. Additionally, we first carried out a visual data inspection, which indicated a possible different age effect for the acoustical parameters in older-paired and younger-unpaired males. Subsequently, the effect of age on the acoustic parameters was analyzed separately for paired and younger-unpaired males using the following model:
In each of these models, yij is a dependent variable (maximum and minimum frequency, frequency range, number of frequency modulations, duration of the multi-modulation phrase, and duration of the second note), the social status is a fixed effect of the ith social status (i = younger-unpaired, older-paired), bAge is a regression coefficient for the age of individuals, a is a random effect of jth animals, and e is a residual error.
We determined statistical differences between younger-unpaired and older-paired males for the multi-modulation phrase of dependent variable “staccato” with a generalized linear mixed model (GLMM) using PROC GLIMMIX with repeated statements for the purpose of repeated measurements. Here, the dependent variable staccato notes (yi) has a value of 1 (presence) with a probability of πi or 0 (absence) and with a probability of 1 − πi for observation i. The logistic model uses a link function g(pi) linking the expected value to the following non-linear function:
where πi is the probability of “staccato” being recorded
where social status is a fixed effect of the ith (i = older-paired, younger-unpaired), bAge presents a regression coefficient for the age of individuals, and a is a random effect of jth animals. The goodness of fit of each model (homoscedasticity, normality of errors, and independence) was checked by visually inspecting residuals using the SAS statement “PLOTS = PEARSON PANEL” and testing for normality according to the Kenward–Roger test.
3. Results
3.1. The structure of the male call differs between younger-unpaired and older-paired males
3.1.1. Social status as a fixed effect
Four of the seven acoustic parameters were significantly longer and higher for younger-unpaired males than they were for older-paired males (Table 2). Younger-unpaired males had a longer total duration of the multi-modulation phrase (P < 0.0001), a longer duration of the second note (P < 0.0001), a higher maximum frequency (P < 0.0001), and a wider frequency range (P < 0.0001). On the contrary, older-paired males had a higher minimum frequency (P < 0.0001). Younger-unpaired males exhibited a lower occurrence of staccato notes (11.98 %; N = 73; P < 0.0001) than their older-paired counterparts (50.54 %; N = 358; P < 0.0001). In younger-unpaired males, there were shorter series of staccato notes (range: 0–8 notes) than in older-paired males (range: 0–18 notes). Among the latter, two subjects (Male 1 and Male 6) mainly contributed to this difference (Table 1).

Table 2. Results based on GLMM showing a comparison between male calls (staccato notes and multi-modulation phrases) by younger-unpaired males and those of older-paired males.
3.1.2. Age as a regression coefficient
In younger-unpaired males, two of the six acoustic parameters were significantly influenced by age: the total duration of the multi-modulation phrase (P < 0.0001) and the minimum frequency (P = 0.0026). The total duration of the multi-modulation phrase decreased with age, while the minimum frequency increased with age (Table 2, Figure 3). In older-paired males, five of the six acoustic parameters increased with age: the total duration of the multi-modulation phrase, the duration of the second note, the maximum frequency, the minimum frequency, and the frequency range (all p < 0.0001).
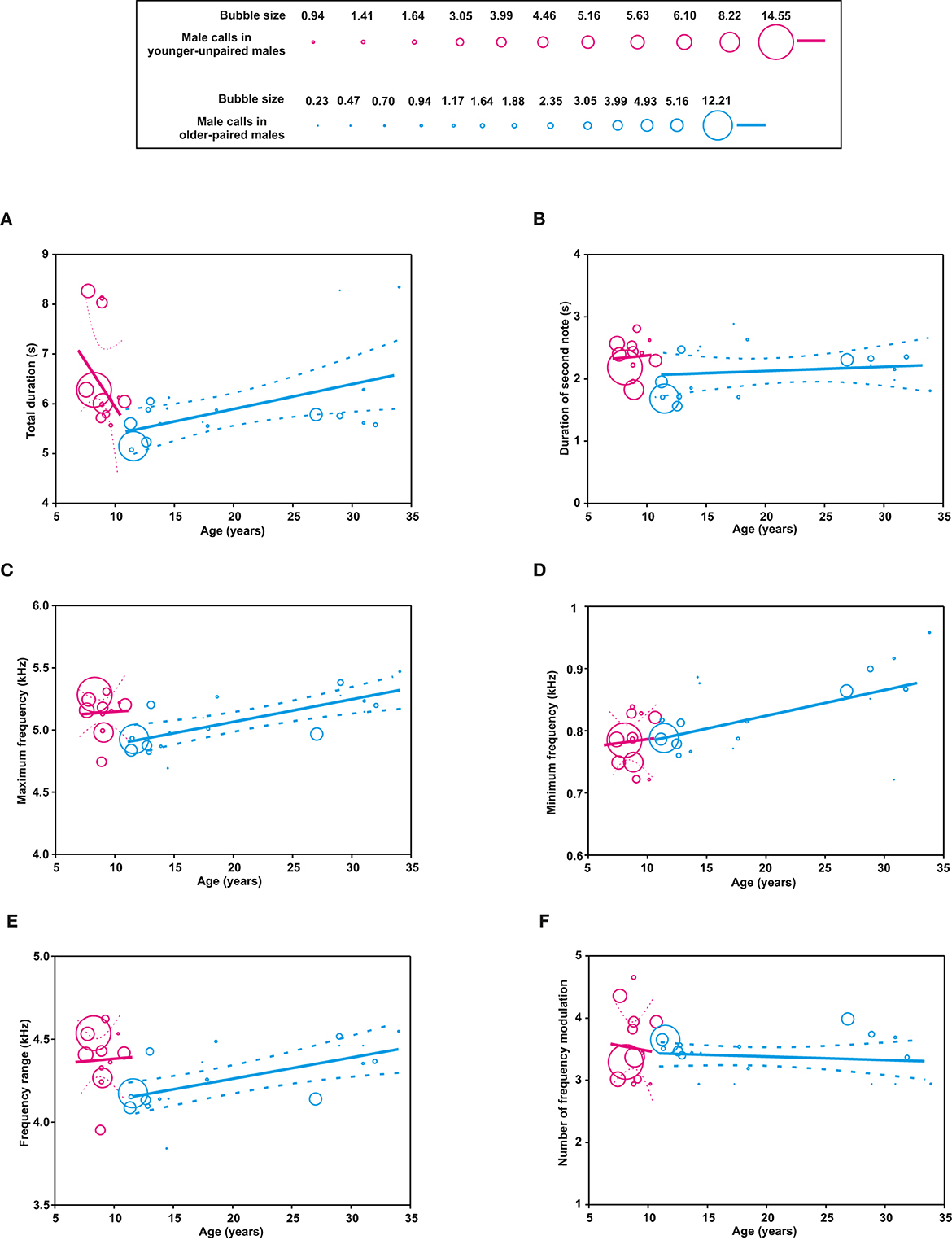
Figure 3. Bubble plots showing the influence of age (years) and social status on the acoustic parameters of male calls (A–F) in younger-unpaired males (pink bubbles) and older-paired males (blue bubbles). In each panel, the size of the bubbles denotes the proportion of recording samples obtained from each subject at a given age. For each group of subjects, the regression lines are plotted with 95% confidence intervals.
3.2. The coda vocalization structure in older-paired males compared to the multi-modulation phrase structure of the male call in younger-unpaired males
3.2.1. Social status as a fixed effect
All six acoustic parameters differed significantly when comparing the multi-modulation phrase of younger-unpaired males with the coda vocalizations of older-paired males (Table 3). These consisted of a longer total duration of the multi-modulation phrase (P < 0.0001), a longer duration of the second note (P = 0.0025), a higher maximum frequency (P = 0.0075), and a wider frequency range (P < 0.0001). On the contrary, the coda vocalization of paired males indicated a higher minimum frequency (P = 0.0006) and a higher number of frequency modulations in the second note (P = 0.0002), than that of the multi-modulation phrase emitted by unpaired males. Both acoustic parameters increased with age (Table 3).

Table 3. Results based on GLMM showing a comparison between the multi-modulation phrase in younger-unpaired males and the male coda vocalization in older-paired males.
3.2.2. Age as a regression coefficient
For the multi-modulation phrase, two of the six acoustic parameters were significantly influenced by age: the total duration decreased with age (P < 0.0001), whereas the minimum frequency increased with age (P = 0.0026; Table 3, Figure 4). For the coda vocalization, all six acoustic parameters were significantly influenced by age: the total duration of the multi-modulation phrase (P = 0.0014), the duration of the second note (P < 0.0001), the maximum frequency (P < 0.0001), the minimum frequency (P = 0.0309), the frequency range (P < 0.0001), and the number of frequency modulations in the second note (P = 0.0422). The total duration of the multi-modulation phrase, the duration of the second note, the maximum and minimum frequency, and the frequency range increased with age, while the number of frequency modulations in the second note decreased with age (Table 3, Figure 4).
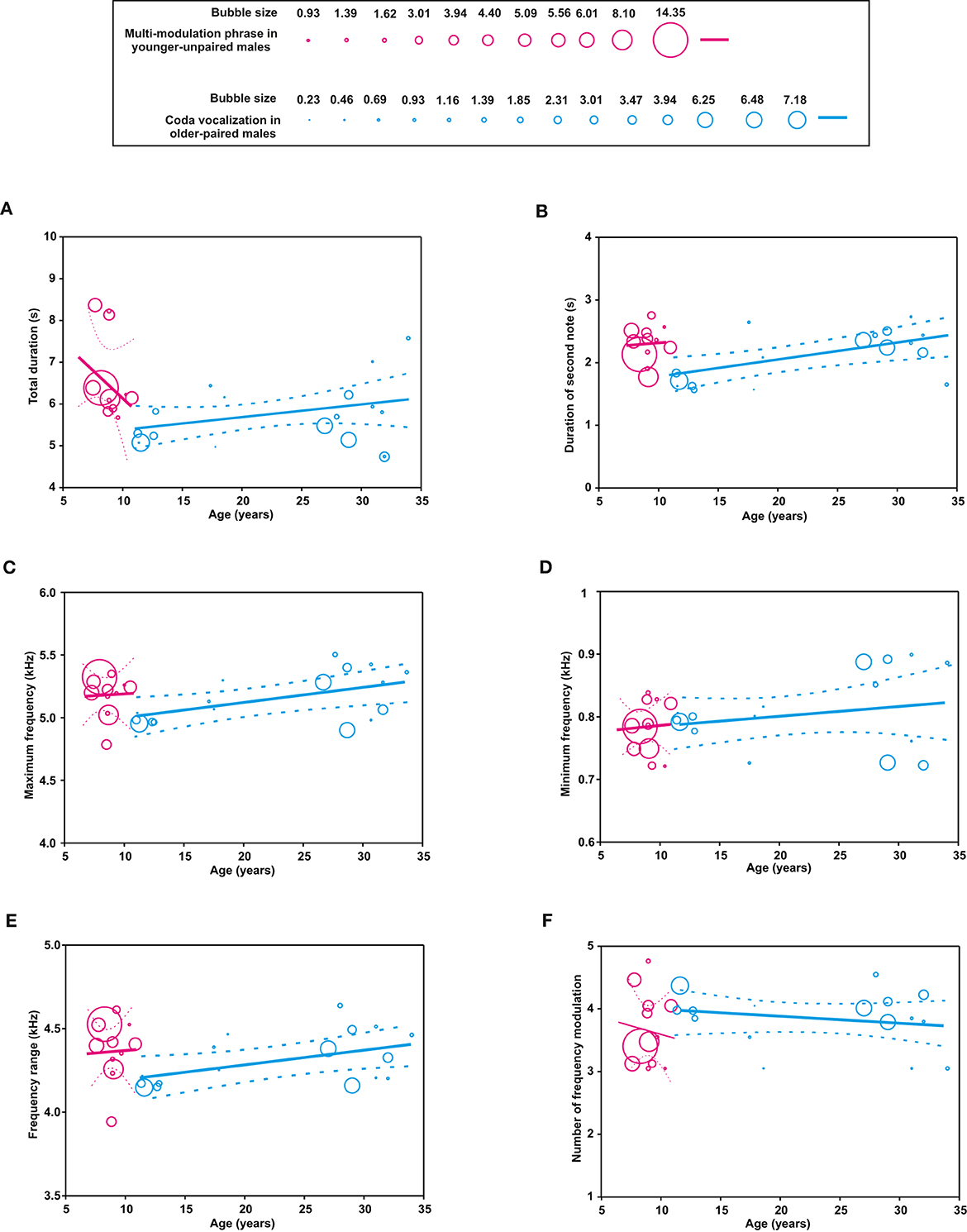
Figure 4. Bubble plots showing the influence of age (years) and social status on the acoustic parameters of male calls (A–F) in multi-modulation phrase of younger-unpaired males (pink bubbles) and coda vocalization of older-paired males (blue bubbles). In each panel, the size of the bubbles denotes the proportion of recording samples obtained from each subject at a given age. For each group of subjects, the regression lines are plotted with 95% confidence intervals.
3.3. Comparing the structure of coda vocalization to that of the multi-modulation phrase in older-paired males
In older-paired males, only one of the six acoustic parameters was significantly higher for the coda vocalization than for the multi-modulation phrase of the male call, namely the number of frequency modulations in the second note (P < 0.0001, Table 4). On the contrary, the multi-modulation phrase of the male call of older-paired males had a longer total duration (P < 0.0001) and a higher minimum frequency (P < 0.0084) than the coda vocalization.
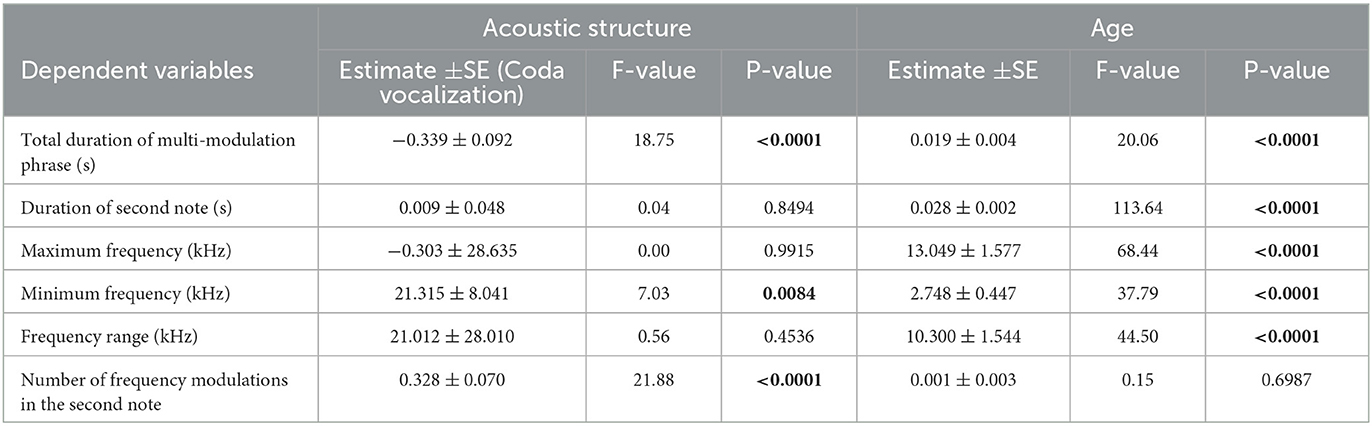
Table 4. Results based on GLMM showing a comparison between the multi-modulation phrase and the male coda vocalization in older-paired males.
3.3.1. Age as a regression coefficient
Five of the six acoustic parameters significantly increased with age: the total duration of the multi-modulation phrase, the duration of the second note, the maximum frequency, the minimum frequency, and the frequency range (all P < 0.0001; Table 4).
4. Discussion
Overall, our analysis of the songs uttered by captive adult males of southern yellow-cheeked gibbons provides three new results. (1) We confirmed our first prediction that younger-unpaired males produce songs that differ in various acoustical features from older-paired males. However, it is unclear which features were influenced by social status or age. (2) We partly confirmed our second prediction that in older-paired males, the coda vocalization structurally differs from the multi-modulation phrase of male calls. (3) Compared with younger-unpaired males, longer series of staccato notes were found to precede the multi-modulation phrase in older-paired males.
4.1. Synergic effects of social status and age on male call and coda vocalization
Our analysis focused on the second note of the multi-modulation phrase of southern yellow-cheeked gibbons, which is the predominant acoustic structure that sets them apart from other species of the genus Nomascus (Konrad and Geissmann, 2006). Compared to the male call and coda vocalization of older-paired males, we found that the multi-modulation phrase of the male calls in younger-unpaired males was characterized by a longer total duration, with the second note having a longer duration, a higher maximum frequency, a lower minimum frequency, and a broader frequency range. In contrast, the coda vocalization in older-paired males was characterized by a higher minimum frequency and a larger number of frequency modulations in the second note compared to that same note in the multi-modulation phrase of male calls.
While the male call of unpaired, younger males can be clearly distinguished from the male call and the coda vocalization of paired older males, the effect of social status (paired vs. unpaired males) could not be dissociated from the effect of age (younger vs. older males) since both variables were confounded. The breeding of gibbons in European zoological gardens needs to follow the recommendations of the relevant EEP coordinator with respect to the needs of the population as a whole and the specific conditions inherent to each zoo. Due to the long-term surplus of males, the tendency is to extend the stay of a male in his natal group for as long as possible, ideally up to 10 years of age. The possibility of obtaining vocal samples from both paired and unpaired males that overlap in age distribution is thus limited by the number of animals kept and the EEP breeding recommendations.
Nonetheless, the effect of social status on the song structure might be supported by the fact that male gibbons below 8 years of age have been known to mate and produce offspring (Geissmann, 1991). It is, therefore, possible that younger-unpaired males might attract females by emitting more potent songs compared with older-paired males.
As mentioned above, the multi-modulation phrase of the male call by younger-unpaired males had fewer extremely rapid frequency modulations in the second note than the coda vocalization in older-paired males. Furthermore, this number of frequency modulations did not differ from that found in the male calls of older-paired males. It might be that younger-unpaired males invest more energy in other acoustic features, which might provide more information to receivers about the singer's attributes than the higher number of frequency modulations in the second note.
The multi-modulation phrase of the male call exhibited minimum frequency values that increased with age, irrespective of the pairing status. Our results are in disagreement with previous findings in adults of non-human primates, showing that the minimum frequency decreases (e.g., Inoue, 1988) or remains similar (Terleph et al., 2016) with increasing age. The proximate mechanisms responsible for the age-dependent increase in the minimum frequency of vocal output that we report are not yet known.
In both older-paired and younger-unpaired males, a series of staccato notes sometimes precede the multi-modulation phrase of the male call. We found that staccato notes had a lower probability of occurrence in younger-unpaired males and exhibited shorter series of notes compared with older-paired males. This is in agreement with Huang et al. (2020), who also observed fewer staccato notes in unpaired Cao-vit gibbons (Nomascus nasutus) and black-crested gibbons (Nomascus concolor) males than in their paired counterparts. The function of staccato notes is not yet fully known. Given their very subtle structures, low volume, and high numbers in older-paired males, we hypothesize that staccato notes may, for example, play a role in song coordination between males and females during duets.
We found no support for our first prediction in two hylobatid studies, one focusing on social status (Mitani, 1988) and the other one on the effect of age (Barelli et al., 2013). Mitani (1988) compared the vocal structure of unpaired and paired males in agile gibbons. In that report, two of the four unpaired adult males were still resident within their family group and the two others were floaters of unknown age. The study did not reveal any obvious acoustical difference in the solo songs of those males compared to paired males. One possible explanation for the difference in results between our study and that of Mitani (1988) is that paired Nomascus gibbon males usually produce the male song only in combination with the female great call, i.e., as part of a non-overlapping duet (Haimoff, 1984a; Geissmann, 2002), whereas paired agile gibbon males produce male songs either as solos or as a contribution to the duet song. It is possible that paired agile gibbon males produce solo songs in different situations, and the vocal structure of their solo songs might differ from that of their duets.
However, our results are in agreement with previous findings, showing that in birds, unpaired males sing higher frequency songs compared to paired males (Staicer, 1996; Martínez and Zuberogoitia, 2002; Staicer et al., 2006; Sung and Handford, 2019). Our findings are also consistent with those of Hennin et al. (2009) in the rufous-and-white wrens (Thryothorus rufalbus), in which the trills of bachelor (unpaired) males displayed a higher maximum frequency, broader bandwidths (i.e., frequency range), and longer call durations than paired males. In this study, three solitary males were recorded both before and after pairing with a female, while 13 others were recorded both before and after losing a mate. In both groups, the bachelors produced songs that were more potent than those of paired males. These results suggest that songs of rufous-and-white wrens encode information about their paired status (Hennin et al., 2009).
An effect of age might be another possible explanation for the more salient songs of younger-unpaired males. It has been shown that the development of the male pattern in the southern yellow-cheeked gibbon is completed by the age of 7.1 years (Hradec et al., 2021b). Androgen levels probably contribute to larynx growth (changes in laryngeal muscle) during the development of the male call; androgen receptors are located on the laryngeal cartilage (Newman et al., 2000), which has a permanent impact on the length or tension of the vocal folds (Fitch and Hauser, 1995). Higher levels of androgens in gibbons may persist even after the development of the adult male pattern at the age of 7 years (Barelli et al., 2013), thus further impacting the components of the vocal apparatus—larynx morphology and tension of the vocal folds. This could explain our finding of the wider frequency range and longer duration in the calls of younger-unpaired males. Subsequently, androgen levels in older males may decline and stabilize at a lower level (Barelli et al., 2013).
To our knowledge, there is no other study that focuses on the effect of age on the song structure in unpaired male gibbons. At first glance, visual inspection of the data for younger-unpaired males showed a decline of several acoustical parameters, but subsequent analysis revealed a significant decrease only for the total duration of the multi-modulation phase, not for the other parameters. A study on white-handed gibbons examined the effect of age in paired males but not in unpaired males (Barelli et al., 2013). In that study, two age categories were compared (adult males: 8–25 years of age; senior males: over 25 years of age), revealing an age-related decline in acoustic features of male calls. That study states that senior males emit calls having a lower pitch and a shorter duration than their (younger) adult counterparts. Our results showed the opposite effect for all analyzed acoustical parameters except for the number of frequency modulations in the second note (i.e., there was no age effect). It is difficult to interpret the age effect from our study when compared to that of Barelli et al. (2013) in which only the solo songs of paired males were investigated but not their duet songs. In addition, the white-handed gibbon belongs to the genus Hylobates, which differs from the genus Nomascus by the absence of frequency modulations (Geissmann, 2002).
It is known that female white-handed gibbons exhibit an age-related decline in the acoustic parameters of the great call, suggesting that call features correlate with physical conditions (Terleph et al., 2016). However, females reproduce up to the age of 40 years in the wild (Reichard et al., 2012). To the best of our knowledge, such data are not available for males, but it can be assumed that 40-year-old males are still fertile and may even reproduce beyond that age. It is not clear how the song features change during senescence in gibbons. It has been shown that acoustical parameters of calls in male baboons (Papio cynocephalus ursinus) change with social rank and increasing age (Fischer et al., 2004). Higher-ranking males had a higher call rate and produced longer bouts (Fischer et al., 2002; Kitchen et al., 2003). However, the social system of these primates (multilevel society) differs from the mostly monogamous gibbon species.
Our results are to some extent supported by a study by Wich et al. (2003) in Thomas langurs (Presbytis thomasi), which showed that a change in the social environment (unpaired vs. paired males), rather than an age effect, is related to the changes in acoustic parameters of the male call. The male loud call consists of harsh non-tonal elements (N-units) and tonal elements (T-units), but their meaning is not yet known. That study compared males of the same age (about 7 years) and found that in paired males, not in unpaired males, the total duration and number of N-units increased, while the number of T-units decreased. These findings compare to our results despite the differing age compositions of our two groups of subjects (older-paired and younger-unpaired males).
4.2. Structural differences between the coda vocalization and the multi-modulation phrase of the male call in older-paired males
Our study revealed that, in older-paired males, the coda vocalization differed from the male call by a larger number of frequency modulations in the second note. Conversely, the multi-modulation phrase had a longer total duration than the coda vocalization. To our knowledge, this study is the first of its kind that compares the structure of the multi-modulation phrase of the male call to the coda vocalization in older-paired males.
The higher number of frequency modulations found in the second note of the coda vocalization is in agreement with the results obtained from the visual inspection of the coda vocalization in Hainan gibbons (Haimoff, 1984b). It is possible that the emission of trill-like vocalizations, both in the coda and the multi-modulation phrase, requires a significant amount of energy and might be perceived by other males as a reliable signal of the sender's physical quality, as has been suggested in some birds and mammals (Podos, 1997; Illes et al., 2006; Pasch et al., 2011). In contrast, the shorter duration of the coda vocalization relative to the multi-modulation phrase of the male call is inconsistent with the findings in Hainan gibbons (Haimoff, 1984b).
We also found that the acoustical parameters for coda vocalizations and multi-modulation phrases in older-paired males increased significantly with age, with only one exception, i.e., the number of frequency modulations in the second note. It is possible that after the completion of male vocal development, this acoustic feature remains stable in all types of calls (i.e., multi-modulation phrase and coda vocalization) produced in adult males, as found in the current study. This indicates that the number of frequency modulations in the second note may largely be genetically determined and might potentially be included in future studies of the vocal individuality of Nomascus gibbons.
The purpose of the coda vocalization and the male call is still not fully understood. Several hypotheses have been proposed for the male song, namely territorial advertisement, resource/mate defense, strengthening (cohesion) pair and/or family bonds (social cohesion), and mate attraction (Cowlishaw, 1992; Geissmann, 2002). It is possible that these two calls do not share the same function due to the difference in spectro-temporal parameters (e.g., frequency range and duration of the second note). It is generally known that the coda vocalization is always emitted immediately following a female great call (Konrad and Geissmann, 2006), which suggests that it functions as a means to strengthen pair and/or family bonds. It is also possible that a higher number of frequency modulations in the second note of the coda vocalization may be an indicator of mate guarding in male gibbons. This is supported by the fact that the male emits the coda vocalization immediately after the female great call, perhaps as a sign of his paired status. The greater ability of older-paired males to rapidly modulate the vocal tract may deter other males from approaching the female. In future studies, it would be useful to determine whether the structure of the multi-modulation phrase in unpaired or paired males is more attractive to females in playback settings.
5. Conclusion
Our study is the first of its kind to provide evidence of structural differences in the male calls and coda vocalizations of southern yellow-cheeked gibbons. Calls uttered by younger-unpaired males differed from both the male calls and coda vocalizations of older-paired males by a higher maximum frequency, wider frequency range, and longer duration of the multi-modulation phrase. In contrast, the male calls of older-paired subjects were preceded by a relatively larger number of staccato notes, and a salient feature of the coda vocalization in those males was the emission of a higher number of frequency modulations in the second note together with a lower minimum frequency. Our findings demonstrate that the male call of younger-unpaired males can be clearly distinguished from the male call and the coda vocalization of older-paired males, but further studies are needed to disentangle the effects of social status and age.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The research conducted herein was approved by the Ethics and Animal Care Committee at the Czech University of Life Sciences, Prague (reference number: CZU/1606) and was performed in accordance with relevant guidelines. This study was fully non-invasive and approved by the management of zoos. All zoological institutions employ rigorous standards for animal welfare and are accredited by the European Association of Zoos and Aquaria (EAZA) and and the Union of Czech and Slovak Zoos (UCSZOO). This study fully complied with the legal requirements of the Czech Republic, the Slovak Republic, and the Republic of Slovenia, and those provided by the European Directive 2010/63/EU. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
MH: conceptualizing the experiment, conducting and collecting all data, writing and editing the manuscript, and constructing the figures and tables. GI: conceptualizing the experiment and writing and editing the manuscript. MP: writing and editing the manuscript and constructing the figures. PB: editing the manuscript. HV-V: statistical analyses and editing the manuscript. All authors contributed to the article and approved the submitted version.
Funding
GI was supported by the Ministry of Agriculture of the Czech Republic (MZE-RO0723).
Acknowledgments
The authors would like to thank Helena Chaloupková and Richard Policht for their helpful comments on an earlier draft of the study. Furthermore, we wish to thank all the curators and keepers from the Czech, Slovak, and Slovenian zoos involved in this study for their support and gracious permission for data collection.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.956922/full#supplementary-material
Supplementary Figure 1. Distribution of data collection in younger-unpaired and paired males (number and age of subjects).
Supplementary Table 1. Acoustic terms and definitions for gibbon song.
Supplementary Table 2. Overview of subject males.
Supplementary Table 3. Information on the dimensions of the indoor and outdoor enclosures.
Supplementary Table 4. Overall composition of family groups.
References
Ballentine, B. (2009). The ability to perform physically challenging songs predicts age and size in male swamp sparrows (Melospiza georgiana). Anim. Behav. 77, 973–978. doi: 10.1016/j.anbehav.2008.12.027
Banerjee, A., Phelps, S. M., and Long, M. A. (2019). Singing mice. Curr. Biol. 29, 190–191. doi: 10.1016/j.cub.2018.11.048
Barelli, C., Mundry, R., Heistermann, M., and Hammerschmidt, K. (2013). Cues to androgens and quality in male gibbon songs. PLoS ONE. 8, e82748. doi: 10.1371/journal.pone.0082748
Birot, H. (2022). 16th European Studbook Southern Yellow-Cheeked Gibbon (Nomascus gabriellae). Mulhouse: Mulhouse Zoo.
Briefer, E., Vannoni, E., and McElligott, A. G. (2010). Quality prevails over identity in the sexually selected vocalisations of an ageing mammal. BMC Biol. 8, 35. doi: 10.1186/1741-7007-8-35
Brockelman, W. Y., and Gittins, S. P. (1984). “Natural hybridization in the Hylobates lar species group: Implications for speciation in gibbons,” in The Lesser Apes: Evolutionary and Behavioural Biology, Preuschoft, H., Chivers, D.J., Brockelman, W.Y., and Creel, N. (eds). (Edinburgh University: Edinburgh Press).
Brockelman, W. Y., and Schilling, D. (1984). Inheritance of stereotyped gibbon calls. Nature. 312, 634–636. doi: 10.1038/312634a0
Brockelman, W. Y., and Srikosamatara, S. (1984). “Maintenance and evolution of social structure in gibbons,” in The Lesser Apes: Evolutionary and Behavioural Biology, Preuschoft, H., Chivers, D.J., Brockelman, W.Y., and Creel, N. (eds). (Edinburgh University: Edinburgh Press).
Caselli, C. B., Mennill, D. J., Gestich, C. C., Setz, E. Z. F., and Bicca-Marques, J. C. (2015). Playback responses of socially monogamous black-fronted titi monkeys to simulated solitary and paired intruders. Am. J. Primatol. 77, 1135–1142. doi: 10.1002/ajp.22447
Charlton, B. D., Zhihe, Z., and Snyder, R. J. (2009). The information content of giant panda, Ailuropoda melanoleuca, bleats: Acoustic cues to sex, age and size. Anim. Behav. 78, e8981. doi: 10.1016/j.anbehav.2009.06.029
Clarke, E., Reichard, U. H., and Zuberbuehler, K. (2006). The syntax and meaning of wild gibbon songs. PLoS ONE. 1, e73. doi: 10.1371/journal.pone.0000073
Clink, D. J., Carif, R. A., Crofoot, M. C., and Marshall, A. J. (2018). Evidence of vocal performance constraints in a female non-human primate. Anim. Behav. 141, 85–94. doi: 10.1016/j.anbehav.2018.05.002
Clink, D. J., Lau, A. R., and Bales, K. L. (2019). Age-related changes and vocal convergence in titi monkey duet pulses. Behaviour. 56, 1471–1494. doi: 10.1163/1568539X-00003575
Clink, D. J., Tasirin, J. S., and Klinck, H. (2020). Vocal individuality and rhythm in male and female duet contributions of a nonhuman primate. Curr. Zool. 66, 173–186. doi: 10.1093/cz/zoz035
Clink, D. J., Zafar, M., Ahmad, A. H., and Lau, A. R. (2021). Limited evidence for individual signatures or site-level patterns of variation in male northern gray gibbon (Hylobates funereus) duet codas. Int. J. Primatol. 42, 896–914. doi: 10.1007/s10764-021-00250-2
Cowlishaw, G. (1992). Song function in gibbons. Behaviour. 121, 131–153. doi: 10.1163/156853992X00471
Cramer, E. R. A. (2013). Physically challenging song traits, male quality, and reproductive success in house wrens. PLoS ONE. 8, e59208. doi: 10.1371/journal.pone.0059208
De Gregorio, C., Carugati, F., Estienne, V., Valente, D., Raimondi, T., Torti, V., et al. (2021). Born to sing! Song development in a singing primate. Curr. Zool. 67, 597–608. doi: 10.1093/cz/zoab018
De Gregorio, C., Carugati, F., Valente, D., Raimondi, T., Torti, V., Miaretsoa, L., et al. (2022). Notes on a tree: reframing the relevance of primate choruses, duets, and solo songs. Ethol. Ecol. Evol. 34, 205–219. doi: 10.1080/03949370.2021.2015451
Demars, C., and Goustard, M. (1978). Le grand chant d'Hylobates concolor leucogenys. Comparison avec les émissions sonores homologues d'H. c. gabriellae et d'H. klossii (iles Mentawai, ouest Sumatra). Behaviour. 65, 1–26. doi: 10.1163/156853978X00503
Erb, W. M., Hodges, J. K., and Hammerschmidt, K. (2013). Individual, Contextual, and Age-Related Acoustic Variation in Simakobu (Simias concolor) Loud Calls. Plos One. 8:e83131. doi: 10.1371/journal.pone.0083131
Ey, E., Pfefferle, D., and Fischer, J. (2007). Do age- and sex-related variations reliably reflect body size in non-human primate vocalizations? A review. Primates. 48, 253–267. doi: 10.1007/s10329-006-0033-y
Fischer, J., Hammerschmidt, K., Cheney, D. L., and Seyfarth, R. M. (2002). Acoustic features of male baboon loud calls: influences of context, age, and individuality. J. Acoust. Soc. Am. 111:1465–1474. doi: 10.1121/1.1433807
Fischer, J., Kitchen, D. M., Seyfarth, R. M., and Cheney, D. L. (2004). Baboon loud calls advertise male quality: acoustic features and their relation to rank, age, and exhaustion. Behav. Ecol. Sociobiol. 56, 140–148. doi: 10.1007/s00265-003-0739-4
Fitch, W. T., and Hauser, M. D. (1995). Vocal production in non-human primates—acoustics, physiology, and functional constraints on honest advertisement. Am. J. Primatol. 37, 191–219 doi: 10.1002/ajp.1350370303
Gamba, M., Torti, V., Estienne, V., Randrianarison, R. M., Valente, D., Rovara, P., et al. (2016). The indris have got rhythm! Timing and pitch variation of a primate song examined between sexes and age classes. Fron. Neurosci. 10, 249. doi: 10.3389/fnins.2016.00249
Geissmann, T. (1984). Inheritance of song parameters in the gibbon song, analyzed in 2 hybrid gibbons (Hylobates pileatus x H. lar). Folia Primatol. 42, 216–235. doi: 10.1159/000156165
Geissmann, T. (1991). Reassessment of age of sexual maturity in gibbons (Hylobates spp.). Am. J. Primatol. 23, 11–22. doi: 10.1002/ajp.1350230103
Geissmann, T. (2002). Duet-splitting and the evolution of gibbon songs. Biol. Rev. 77, 57–76. doi: 10.1017/S1464793101005826
Geissmann, T., and Nijman, V. (2006). Calling in wild silvery gibbons (Hylobates moloch) in Java (Indonesia): Behavior, phylogeny, and conservation. Am. J. Primatol. 68, 1–19. doi: 10.1002/ajp.20203
Gil, D., and Gahr, M. (2002). The honesty of bird song: multiple constraints for multiple traits. Trends Ecol. Evol. 17, 133–141. doi: 10.1016/S0169-5347(02)02410-2
Haimoff, E. H. (1984a). “Acoustic and organizational features of gibbon song”, in The Lesser Apes: Evolutionary and Behavioural Biology, eds H. Preuschoft, W. Y. Chivers, W. Y. Brockelman, and N. Creel (Edinburgh University: Edinburgh Press).
Haimoff, E. H. (1984b). The organization of song in the Hainan black gibbon (Hylobates concolor hainanus). Primates. 25, 225–235. doi: 10.1007/BF02382394
Hauser, M. D. (1989). Ontogenic changes in the comprehension and production of vervet monkey (Cercopithecus aethiops) vocalizations. J. Comp. Psychol. 103, 149–158. doi: 10.1037/0735-7036.103.2.149
Hennin, H. L., Barker, N. K. S., Bradley, D. W., and Mennill, D. J. (2009). Bachelor and paired male rufous-and-white wrens use different singing strategies. Behav. Ecol. Sociobiol. 64, 151–159. doi: 10.1007/s00265-009-0832-4
Hradec, M., Bolechova, P., and Svobodová, I. (2016). Production of a female-specific great call in an immature male gibbon, the Nomascus genus. Primates 57, 445–448 doi: 10.1007/s10329-016-0569-4
Hradec, M., Illmann, G., Bartos, L., and Bolechova, P. (2021b). The transition from the female-like great calls to male calls during ontogeny in southern yellow-cheeked gibbon males (Nomascus gabriellae). Sci. Rep. 11, 22040. doi: 10.1038/s41598-021-01648-x
Hradec, M., Illmann, G., and Bolechova, P. (2021a). A first report of separation calls in southern yellow-cheeked gibbons (Nomascus gabriellae) in captivity. Primates 62, 5–10. doi: 10.1007/s10329-020-00870-x
Hradec, M., Linhart, P., Bartos, L., and Bolechova, P. (2017). The traits of the great calls in the juvenile and adolescent gibbon males Nomascus gabriellae. PLoS ONE. 12, e0173959. doi: 10.1371/journal.pone.0173959
Huang, M. P., Ma, H. G., Ma, C. Y., Garber, P. A., and Fan, P. F. (2020). Male gibbon loud morning calls conform to Zipf's law of brevity and Menzerath's law: insights into the origin of human language. Anim. Behav. 160, 145–155. doi: 10.1016/j.anbehav.2019.11.017
Illes, A. E., Hall, M. L., and Vehrencamp, S. L. (2006). Vocal performance influences male receiver response in the banded wren. Proc. R. Soc. B:.Biol. Sci. 273, 1907–1912. doi: 10.1098/rspb.2006.3535
Inoue, M. (1988). Age gradations in vocalization and body weight in Japanese monkeys (Macaca fuscat). Folia Primatol. 51:76–86. doi: 10.1159/000156359
Janik, V. M., and Knornschild, M. (2021). Vocal production learning in mammals revisited. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 376, 20200244. doi: 10.1098/rstb.2020.0244
Kitchen, D. M., Seyfarth, R. M., Fischer, J., and Cheney, D. L. (2003). Loud calls as indicators of dominance in male baboons (Papio cynocephalus ursinus). Behav. Ecol. Sociobiol. 53, 374–384. doi: 10.1007/s00265-003-0588-1
Konrad, R., and Geissmann, T. (2006). Vocal diversity and taxonomy of Nomascus in Cambodia. Int. J. Primatol. 27, 713–745. doi: 10.1007/s10764-006-9042-3
Lau, A. R., Clink, D. J., and Bales, K. L. (2020). Individuality in the vocalizations of infant and adult coppery titi monkeys (Plecturocebus cupreus). Am. J. Primatol. 82:e23134. doi: 10.1002/ajp.23134
Lau, A. R., Clink, D. J., Crofoot, M. C., and Marshall, A. J. (2018). Evidence for High Variability in Temporal Features of the Male Coda in Muller's Bornean Gibbons (Hylobates muelleri). Int. J. Primatol. 39, 670–684. doi: 10.1007/s10764-018-0061-7
Martínez, J. A., and Zuberogoitia, I. (2002). Factors affecting the vocal behaviour of eagle owls Bubo bubo: effects of season, density and territory quality. Ardeola. 50, 255−258
Mitani, J. C. (1987). Territoriality and monogamy among agile gibbons (Hylobates agilis). Behav. Ecol. Sociobiol. 20, 265–269. doi: 10.1007/BF00292179
Mitani, J. C. (1988). Male gibbon (Hylobates agilis) singing behavior - natural history, song 548 variations and function. Ethology. 79, 177–194. doi: 10.1111/j.1439-0310.1988.tb00710.x
Nemeth, E., Kempenaers, B., Matessi, G., and Brumm, H. (2012). Rock sparrow song reflects male age and reproductive success. PLoS ONE. 7:e43259. doi: 10.1371/journal.pone.0043259
Newman, S. R., Butler, J., Hammond, E. H., and Gray, S. D. (2000). Preliminary report on hormone receptors in the human vocal fold. J. Voice 14, 72–81. doi: 10.1016/S0892-1997(00)80096-X
Owren, M. J., and Rendall, D. (2003). Salience of caller identity in rhesus monkey (Macaca mulatta) coos and screams: Perceptual experiments with human (Homo sapiens) listeners. J. Comp. Psychol. 117, 380–390. doi: 10.1037/0735-7036.117.4.380
Pasch, B., George, A. S., Campbell, P., and Phelps, S. M. (2011). Androgen-dependent male vocal performance influences female preference in neotropical singing mice. Anim. Behav. 82, 177–183. doi: 10.1016/j.anbehav.2011.04.018
Pfefferle, D., and Fischer, J. (2006). Sounds and size: identification of acoustic variables that reflect body size in hamadryas baboons, Papio hamadryas. Anim. Behav. 72, 43–51. doi: 10.1016/j.anbehav.2005.08.021
Pistorio, A. L., Vintch, B., and Wang, X. (2006). Acoustic analyses of vocal development in a new world primate, the common marmoset (Callithrix jacchus). J. Acoust. Soc. Am. 120, 1655–1670. doi: 10.1121/1.2225899
Podos, J. (1997). A performance constraint on the evolution of trilled vocalizations in a songbird family (Passeriformes:Emberizidae). Evolution. 51, 537–551. doi: 10.2307/2411126
Price, T., Arnold, K., Zuberbuhler, K., and Semple, S. (2009). Pyow but not hack calls of the male putty-nosed monkey (Cercopithcus nictitans) convey information about caller identity. Behaviour. 146, 871–888. doi: 10.1163/156853908X396610
Raemaekers, J. J., and Raemaekers, P. M. (1985). Field playback of loud calls to gibbons (Hylobates lar) - Territorial, sex-specific and species-specific responses. Anim. Behav. 33, 481–493. doi: 10.1016/S0003-3472(85)80071-3
Reby, D., and McComb, K. (2003). Anatomical constraints generate honesty: acoustic cues to age and weight in the roars of red deer stags. Anim. Behav. 65, 519–530. doi: 10.1006/anbe.2003.2078
Reichard, U. H., Barelli, C., Hirai, H., and Nowak, M. G. (2016). “Evolution of gibbons and siamang phylogeny, morphology, and cognition”, in The Evolution of Gibbons and Siamang, Reichard, U. H., Hirai, H., and Barelli, C. (Springer, New York, NY). doi: 10.1007/978-1-4939-5614-2
Reichard, U. H., Ganpanakngan, M., and Barelli, C. (2012). “White-handed gibbons of Khao Yai: social flexibility, reproductive strategies, and a slow life history,” in Long-term Field Studies of Primates, Kappeler, P. M., and Watts, D. (eds). (Springer, Berlin). p. 237–258. doi: 10.1007/978-3-642-22514-7_11
Sanvito, S., Galimberti, F., and Miller, E. H. (2008). Development of aggressive vocalizations in male southern elephant seals (Mirounga leonina): maturation or learning? Behaviour. 145:137–170. doi: 10.1163/156853907783244729
Schilling, D. (1984). “Song bouts and duetting in the concolor gibbon,” in The Lesser Apes: Evolutionary and Behavioural Biology, Preuschoft, H., Chivers, D. J., Brockelman, W. Y., and Creel, N. (eds). (Edinburgh University: Edinburgh Press).
Staicer, C. A. (1996). Honest advertisement of pairing status: evidence from a tropical resident woodwarbler. Anim. Behav. 51, 375–390. doi: 10.1006/anbe.1996.0036
Staicer, C. A., Ingalls, V., and Sherry, T. A. (2006). Singing behavior varies with breeding status of American redstarts (Setophaga ruticilla). Wilson J. Ornithol. 118, 439–451. doi: 10.1676/05-056.1
Sun, G. Z., Huang, B., Guan, Z. H., Geissmann, T., and Jiang, X. L. (2011). Individuality in male songs of wild black crested gibbons (Nomascus concolor). Am. J.Primatol. 73, 431–438. doi: 10.1002/ajp.20917
Sung, H., and Handford, P. (2019). Seasonal and diurnal variation of singing activity: their possible relationship with intra- and inter-sexual function in the savannah sparrow (Passerculus sandwichensis). Zool. Sci. 36, 173–181. doi: 10.2108/zs180139
Tenaza, R. R. (1976). Song, choruses and countersinging of Klosi gibbons (Hylobates klossii) in Siberut island, Indonesia. Z. Tierpsychol.-J.Compar. Ethol. 40, 37–52. doi: 10.1111/j.1439-0310.1976.tb00924.x
Terleph, T. A., Malaivijitnond, S., and Reichard, U. H. (2016). Age related decline in female lar gibbon great call performance suggests that call features correlate with physical condition. Bmc Evol. Biol. 16,4 doi: 10.1186/s12862-015-0578-8
Terleph, T. A., Malaivijitnond, S., and Reichard, U. H. (2018a). Male white-handed gibbons flexibly time duet contributions. Behav. Ecol. Sociobiol. 72:16. doi: 10.1007/s00265-017-2432-z
Terleph, T. A., Malaivijitnond, S., and Reichard, U. H. (2018b). An analysis of white-handed gibbon male song reveals speech-like phrases. Am. J. Phys. Anthropol. 166, 649–660. doi: 10.1002/ajpa.23451
Thinh, V. N., Hallam, C., Roos, C., and Hammerschmidt, K. (2011). Concordance between vocal and genetic diversity in crested gibbons. Bmc Evol. Biol. 11,36 doi: 10.1186/1471-2148-11-36
Thorpe, W. H. (1961). “Bird-Song. The Biology of Vocal Communication and Expression in Birds,” in Cambridge Monographs in Experimental Biology No. 12. New York: Cambridge University Press.
Tilson, R. L. (1981). Family formation strategies of Kloss's gibbons. Folis primatol. 35, 259–297. doi: 10.1159/000155979
Torti, V., Bonadonna, G., De Gregorio, C., Valente, D., Randrianarison, R. M., Friard, O., et al. (2017). An intra-population analysis of the indris' song dissimilarity in the light of genetic distance. Sci. Rep. 7:10140. doi: 10.1038/s41598-017-10656-9
Torti, V., Gamba, M., Rabemananjara, Z. H., and Giacoma, C. (2013). The songs of the indris (Mammalia: Primates: Indridae): contextual variation in the long-distance calls of a lemur. Ital. J.Zool. 80, 596–607. doi: 10.1080/11250003.2013.845261
Umeed, R., Attademo, F. L. N., and Bezerra, B. (2018). The influence of age and sex on the vocal repertoire of the antillean manatee (Trichechus manatus manatus) and their responses to call playback. Mar. Mammal. Sci. 34, 577–594. doi: 10.1111/mms.12467
Vanderhoff, E. N., and Hoverud, N. B. (2022). Perspectives on antiphonal calling, duetting and counter-singing in non-primate mammals: an overview with notes on the coordinated vocalizations of bamboo rats (Dactylomys spp., Rodentia: Echimyidae). Front. Ecol. Evol. 10:906546. doi: 10.3389/fevo.2022.906546
Vernes, S. C., and Wilkinson, G. S. (2020). Behaviour, biology and evolution of vocal learning in bats. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 375:20190061. doi: 10.1098/rstb.2019.0061
Wanelik, K. M., Azis, A., and Cheyne, S. M. (2013). Note-, phrase- and song-specific acoustic variables contributing to the individuality of male duet song in the Bornean southern gibbon (Hylobates albibarbis). Primates 54, 159–170. doi: 10.1007/s10329-012-0338-y
Keywords: gibbon, Nomascus, vocalization, coda, male call
Citation: Hradec M, Illmann G, Prikrylová M, Bolechová P and Vostrá-Vydrová H (2023) Difference in the songs of paired and unpaired southern yellow-cheeked gibbon males (Nomascus gabriellae): social status or age? Front. Ecol. Evol. 11:956922. doi: 10.3389/fevo.2023.956922
Received: 30 May 2022; Accepted: 31 May 2023;
Published: 28 July 2023.
Edited by:
Patrice Adret, Universidad Autónoma Gabriel René Moreno, BoliviaReviewed by:
Sofya Dolotovskaya, German Primate Center, GermanyChiara De Gregorio, University of Turin, Italy
Copyright © 2023 Hradec, Illmann, Prikrylová, Bolechová and Vostrá-Vydrová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michal Hradec, aHJhZGVjbUBhZi5jenUuY3o=