
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Ecol. Evol., 06 October 2023
Sec. Behavioral and Evolutionary Ecology
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1263974
 Miklós Laczi1,2,3*
Miklós Laczi1,2,3* Gábor Herczeg1,4
Gábor Herczeg1,4 Gyula Szabó2,5
Gyula Szabó2,5 Helga Gyarmathy2,5
Helga Gyarmathy2,5 Fanni Sarkadi2,5
Fanni Sarkadi2,5 János Török1,2Gergely Hegyi1,2
János Török1,2Gergely Hegyi1,2Information on the exact nature of sexual dichromatism might be incomplete, often leading to the treatment of dichromatic species as monochromatic. This error is evident when the two sexes of a species look identical to the human eye, as in the great tit (Parus major). We measured reflectance in three sections (throat, breast, belly) of the abdominal black stripe of great tits during the pairing and nestling feeding periods and assessed differences between patch sections, sexes, and seasons using visual models. We found that the stripe, which had previously been considered a single, contiguous patch, consists of multiple patches. In males, the breast section differed markedly from the throat and belly sections in having higher total brightness and ultraviolet chroma, while the female’s breast seemed to be less bright than the two other regions, resulting in strong sexual dichromatism hidden from the human eye. Colouration was more pronounced in winter, but dichromatism was present in both periods. The hidden breast ultraviolet patch we discovered in males may act as a sexual ornament or a signal amplifier.
Sexual dichromatism is frequently invoked in evolutionary biology (Kodric-Brown, 1998; Kimball and Ligon, 1999; Pérez i de Lanuza et al., 2013; Dale et al., 2015; Shultz and Burns, 2017; Miller et al., 2021). Dichromatism could imply sexual selection on a trait, hence, its degree is often used as an index of sexual selection (Kraaijeveld et al., 2011; Seddon et al., 2013; Cally et al., 2021; but see Badyaev and Hill, 2000; Price, 2019). Cryptic dichromatism (invisible to the human eye) is widespread (Cummings et al., 2003; Hofmann et al., 2007; Font et al., 2009; Pérez i de Lanuza et al., 2013; Hernández-Palma, 2016), which is typically known to be linked to the ultraviolet (UV) component of colouration (Eaton, 2005; Stella et al., 2018). However, its investigation in the achromatic dimension (colour-independent intensity) is rare (Eaton and Lanyon, 2003; Armenta et al., 2008; Burns and Shultz, 2012). Additionally, studies generally a priori define a patch as one that has a colour distinguishable by the human eye from the rest of the adjacent areas (Eaton, 2005; Burns and Shultz, 2012; Amézquita et al., 2017; Negro et al., 2018a; but see Rossi et al., 2019).
It is known from flowers and butterflies that human-visible and human-invisible patterns can be different (Lunau, 1992; Stella et al., 2018; Koski, 2020). In birds, only Eaton and Lanyon (2003) considered the potential existence of hidden UV patterns within a seemingly contiguous colour patch, and found evidence for this in two species, namely in the UV–violet throat patch of the superb fairywren (Malurus cyaneus) and the orange nape patch of the streaked bowerbird (Amblyornis subalaris). Such observations could become especially meaningful to the inspiring studies on the origin, function, or consequences of integumental patch pattern diversity (Rojas, 2017; Mason and Bowie, 2020; Cally et al., 2021; Miller et al., 2021; Emberts and Wiens, 2022; Luro and Hauber, 2022). Another overlooked phenomenon is the temporal plasticity of colouration. In birds, feather reflectance can change without moult within a short time interval (Delhey et al., 2006; Surmacki et al., 2011; Hegyi et al., 2019; Laczi et al., 2020). These changes can differ between sexes (Örnborg et al., 2002), which may potentially affect the observed magnitude of dichromatism depending on sampling time.
If the zebra finch (Taeniopygia guttata) is ‘the ultimate Australian supermodel’ of behavioural ecology (Griffith and Buchanan, 2010), the great tit (Parus major) is the Eurasian supermodel. It has a composite plumage pattern usually marked as sexually monochromatic to the human eye (Soler and Moreno, 2012; Negro et al., 2018b). However, spectrometry revealed sexual dichromatism in the yellow breast and black crown (Hegyi et al., 2007; Evans et al., 2010; Laczi et al., 2019). Additionally, the obviously dichromatic size of the abdominal stripe (black to humans) could reflect information on social status, survival and parental abilities (Järvi and Bakken, 1984; Norris, 1990; Senar et al., 2014). We observed in the field that the middle (breast) section of the stripe looked glossy in males and dull in females, similarly to the crown (Figure 1A), which is dichromatic with regards to brightness and UV chroma (Hegyi et al., 2007; Laczi et al., 2019). Here, we tested whether cryptic dichromatism with hidden within-patch pattern and seasonal effects is present in great tits’ fitness-linked abdominal stripe, measuring three stripe regions (throat, breast, belly; Figure 1B) in two seasons (winter, spring).

Figure 1 Great tits: (A) as seen in nature, depicting that under direct sunlight, the male black breast section of the abdominal stripe is moderately glossy, in contrast to females; (B) showing the measured plumage parts with false-colouring differences in reflectance as seen by humans and tits. (Credit: ML).
Data were collected in the Pilis-Visegrádi Mountains, Hungary (47°72’N, 19°01’E), in 2022. We captured birds during winter (territory establishment and pairing period) and spring (nestling feeding period) (winter/spring n♀ = 34/44, n♂ = 48/43; 10 females and 12 males were recaptured from winter to spring). We recorded reflectance curves (Supplementary Figure 1) in the field from the throat, breast, and belly regions of the abdominal black stripe using an USB2000 spectrometer, DH-2000 light source, R400-7 detector, and WS-1-SS white standard (Ocean Optics Europe). We then calculated brightness (average intensity between 320–700 nm, R320–700) and ultraviolet chroma (R320–400/R320–700; hereafter UVC). For additional details, see the Supplementary Material.
We tested for colour differences during the pairing season using generalized linear mixed models with backward stepwise model simplification, normal error, identity link function, and Satterthwaite estimation for degrees of freedom, using the ‘lmerTest’ package (Kuznetsova et al., 2017) in R 4.2.1 (R Development Core Team, 2022). Brightness, or UVC was used as the response variable. Individual identity was fitted as a random effect. Sex and patch region (repeated measures factor) were fixed categorical effects. Two-way interactions between fixed factors were also tested. Binary age (yearling or older) had no consistent effect on any spectral characteristic examined here (results not shown), and its distribution was skewed within and inhomogeneous across groups, violating the assumptions of parametric analyses. We therefore did not include age in the models reported here.
As a next step, we performed across-season comparisons. The initial main model included sex, patch region (repeated measures factor), season and their two- and three-way interactions as fixed effects, and identity as a random effect. To avoid pseudoreplication, we excluded the spring repeats of individuals caught in both seasons. We also analyzed within-individual changes, but these are not reported here in detail due to the small numbers of recaptured birds (see Supplementary Material for details).
We estimated if statistical differences were perceptible to the birds using the ‘pavo 2’ package (Maia et al., 2019). We calculated quantum cone catch values for each spectrum by the ‘vismodel’ function using the blue tit visual system data. These parameters of the great tit are currently not known in detail, but rod and cone spectral sensitivities among songbirds show only moderate variation (Bowmaker et al., 1997; Hart et al., 2000). We then averaged the values for combinations of season, sex, and plumage region. On these averaged values, we used the ‘coldist’ function to get discriminability contrast values between patches, expressed in units of ‘just noticeable differences’ (JNDs), where JND > 1 indicates that two stimuli are distinguishable (Siddiqi et al., 2004). For details, see Supplementary Material.
Among-patch patterns differed significantly between sexes, and they were significant in both sexes for both brightness and UVC. In detail, the sex × patch interaction was significant in brightness (F2,240 = 103.75, p < 0.001) and UVC (F2,160 = 10.70, p < 0.001). The main effect of patch was significant in both sexes for both brightness (males: F2,141 = 125.58, p < 0.001; females: F2,66 = 33.24, p < 0.001) and UVC (males: F2,94 = 31.39, p < 0.001; females: F2,66 = 27.82, p < 0.001). In males, brightness was highest in the breast and lowest in the throat (all p < 0.001), and the pattern was similar for UVC (all p < 0.001). In females, brightness was highest in the belly and lowest in the breast (breast-throat p = 0.048; other comparisons p < 0.001), while UVC increased from the throat to the belly (breast–belly p < 0.01; other comparisons p < 0.001). The detailed results of the pairwise tests are shown in Table 1.
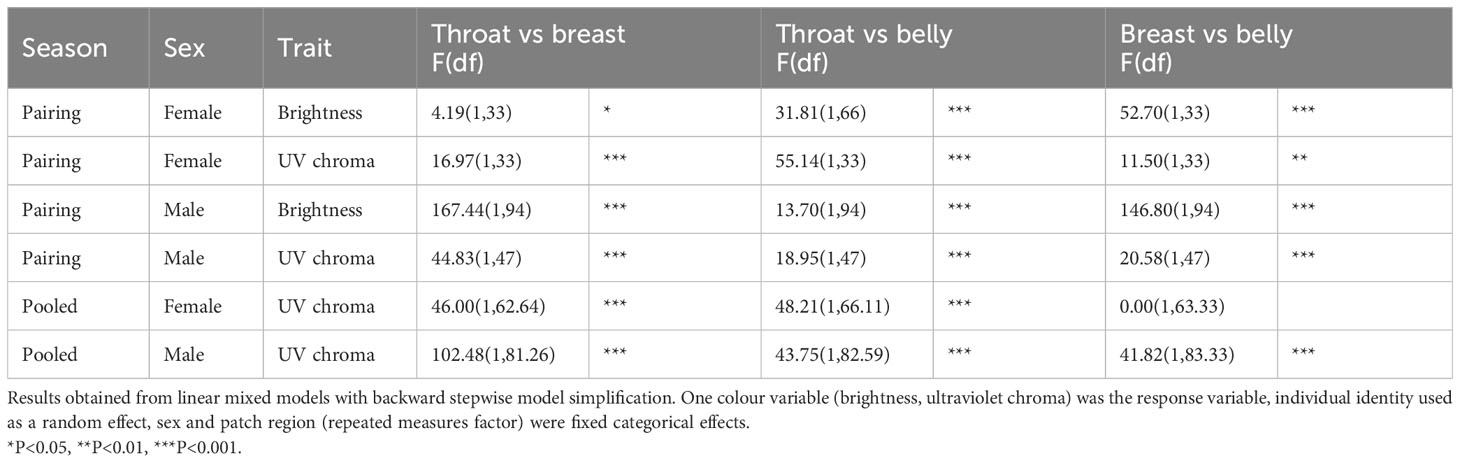
Table 1 Pairwise comparisons of abdominal stripe patches in great tits in different contexts.
The sex × season × patch interaction was significant for brightness (Supplementary Table 1). The season × patch interaction was significant in both males (F2,231 = 13.93, p < 0.001) and females (F2,133.72 = 4.73, p < 0.001). We were particularly interested in the across-season stability of the patch differences, so we ran separate tests of the season × patch interaction for patch pairs, revealing the detailed pattern behind the overall interaction.
In males, breast brightness decreased, while the brightness of the throat and belly increased from winter to spring. This resulted in lower differences in brightness across patches in spring, In females, the breast had lower brightness than either the throat or belly irrespective of season, and the pattern change concerned the relative brightness of the other two areas (Figure 2A, Supplementary Table 1). In more details, in males, the interaction was significant for the breast–throat (F1,154 = 20.12, p < 0.001) and breast–belly comparisons (F1,87.33 = 15.76, p < 0.001), but not for the throat–belly comparison (F1,96.33 = 2.31, p = 0.13). The throat was less bright (darker) than the belly, irrespective of season (main effect F1,97.73 = 11.12, p < 0.01). The breast was brighter than the other two areas, but this difference declined from winter (breast–throat: F1,94 = 167.44, p < 0.001; breast–belly: F1,94 = 146.80, p < 0.001) to spring (breast–throat: F1,60 = 25.71, p < 0.001; breast–belly: F1,30 = 26.01, p < 0.001). For females, the situation was reversed. The breast had lower brightness than the other areas (main effect breast–throat: F1,69.76 = 10.33, p < 0.01; breast–belly: F1,67.63 = 42.42, p < 0.001), with no seasonal change in the differences (season × patch interaction breast–throat: F1,68.73 = 2.75, p = 0.10; breast–belly: F1,66.701 = 2.24, p = 0.14). The season × patch interaction was significant only for the throat–belly comparison (F1,67.67 = 8.42, p < 0.01). The throat showed lower brightness than the belly in winter (main effect F1,66 = 31.81, p < 0.001), but it became brighter with time, and therefore the patch difference disappeared by spring (main effect F1,33 = 0.23, p = 0.64).
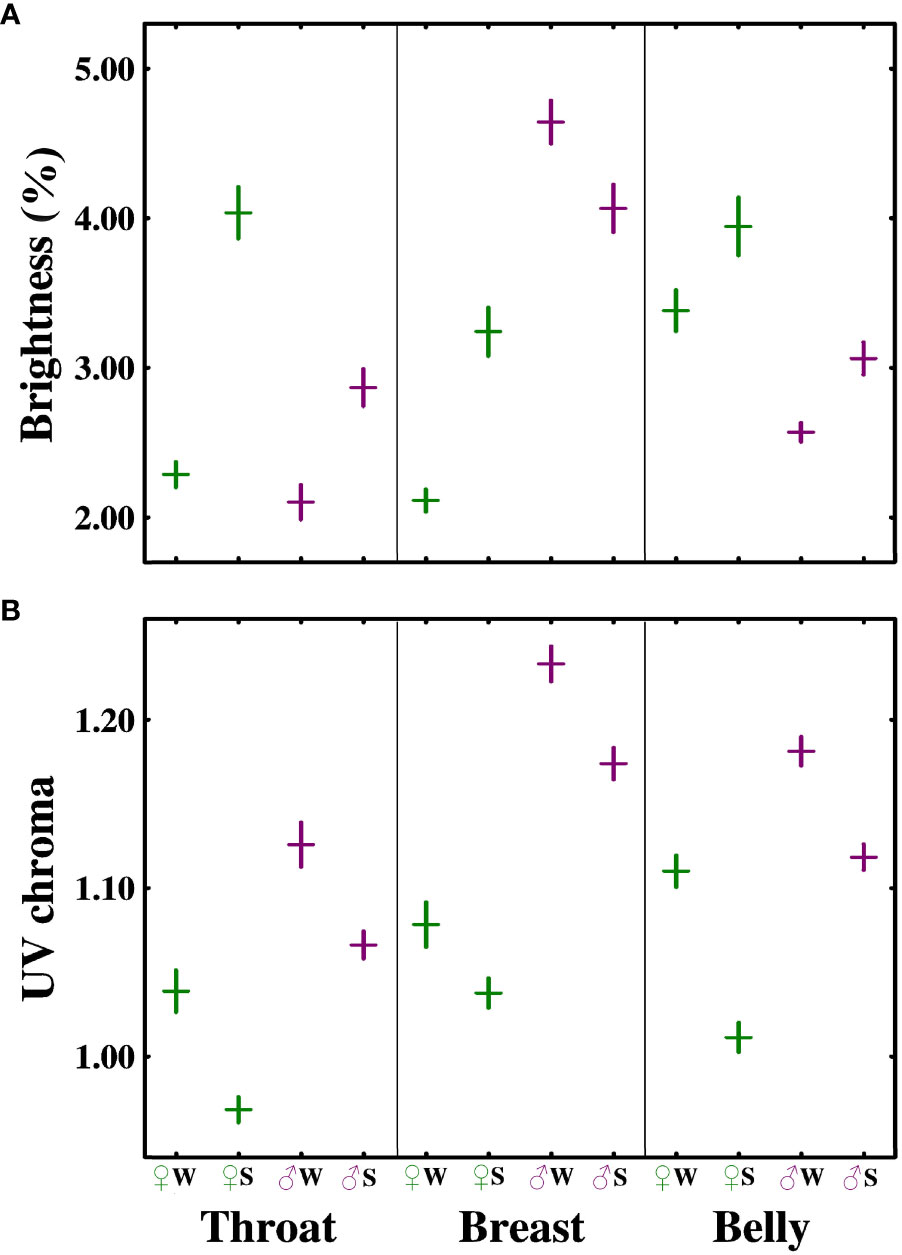
Figure 2 Differences (mean ± SE) in brightness (A) and ultraviolet (UV) chroma (B) of the throat, breast, and belly region of the black abdominal stripe of the great tit, between sexes, and winter (W) and spring (S).
Seasonal change in the among-patch pattern of UVC did not differ between sexes, and all patches declined similarly in both sexes. In details, the sex × season × patch interaction was not significant for UVC, and the only significant two-way interaction was between sex and patch (Supplementary Table S1), with a consistent seasonal change (decline) across sexes and patches (main effect of season in Supplementary Table 1). The effect of patch was significant in both males (F2,161.01 = 71.13, p < 0.001) and females (F2,125.92 = 32.85, p < 0.001). Pairwise comparisons of patches were all significant in males (all p < 0.001), with UVC increasing from the throat to the belly to the breast. In females, the throat showed lower UVC than the other areas (p < 0.001), with no difference between the breast and the belly (p = 0.98). All of the seasonal differences are presented in Figure 2B, and detailed results of the pairwise tests are shown in Table 1.
An across-seasons repeated measures analysis with the recaptured individuals was largely congruent with the above-described patterns (see Supplementary Table 3).
As judged from the JNDs (Table 2, Supplementary Table 2), the male breast was distinguishable from both the throat and the belly in terms of both brightness and chromaticity in both seasons. In females, the breast was distinguishable in terms of brightness from the belly in both seasons and from the throat only in spring. In terms of chromaticity, however, the female breast was visually similar to the throat and the belly in both seasons. Finally, the breast patches of males and females were distinguishable in both seasons in terms of both brightness and chromaticity.
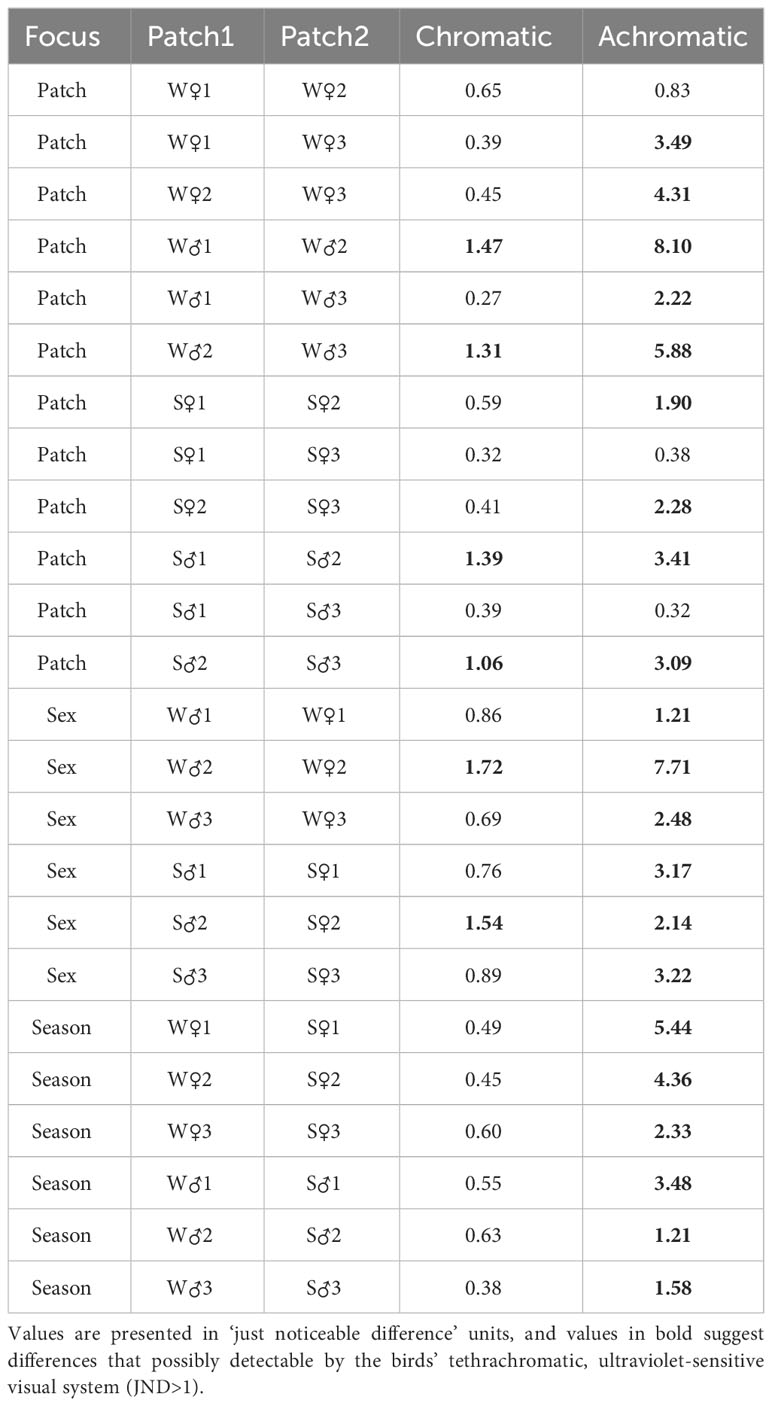
Table 2 Visual model-based chromatic (colour) and achromatic (brightness) differences between throat (1), breast (2), and belly (3) in the abdominal black stripe of great tits in winter (W) and spring (S).
Sexual dichromatism in melanic colouration is less frequent than in structural-based or other pigment-based colouration (Bradley and Mundy, 2008; Taysom et al., 2011), and comparative studies in birds suggest that it is also weaker (Delhey and Peters, 2017). Studies of the abdominal black stripe of great tits considered it as a single trait, sexually monochromatic except for its size (e.g. Norris, 1993). We found that the stripe contains distinct patches, and shows marked sexual dichromatism and seasonal changes.
From a proximate viewpoint, males may have feather barbs and barbules with higher light-absorbing melanin content and a higher density of melanin-containing barbules (Galván, 2011; D’Alba et al., 2014; Laczi et al., 2019). The glossiness (higher specular reflectance) and UV richness in the male breast may be caused by the nanostructure, where the outermost melanin granules in barbules form a continuous layer, allowing the keratin on top of the melanin layer to function as a thin layer (Maia et al., 2011), similarly to the male crown (see Laczi et al., 2019). Additionally, surface smoothness may play a role in glossiness too (Iskandar et al., 2016).
Seasonal colour changes were also prominent. Although sample size and sample composition were slightly different between the two periods, which may potentially have an effect on the results, but the high agreement between the non-redundant horizontal data set and the within-individual data suggests that the seasonal changes in across-patch reflectance patterns were due to within-individual degradation and that sampling bias played a negligible role. Previously, in great tit yellow feathers, a seasonal decline within a moult cycle was found in UVC but not in brightness (Evans et al., 2012), which could partly be explained by soiling and preen oiling (Surmacki and Nowakowski, 2007), but there could be other confounding factors as well (e.g. feather wear, photobleaching). Delhey et al. (2010) showed a parallel change between sexes across the whole year in colour of other plumage parts, with sexual dichromatism being present throughout the year. Temporal changes in plumage colouration were revealed, but this study used visual judgment and not spectrophotometry (Figuerola and Senar, 2005). Here, we found that sexual differences in the breast stripe persisted from winter to spring. Brightness became higher in all stripe parts in both sexes, except for the male breast, which was less bright in spring, whereas UVC was uniformly lower in spring. According to the visual model, it is possible that only the achromatic seasonal changes are visible for birds. The detected seasonal changes can be easily explained if we refer again to the crown. If barbs shorten due to feather wear, and barbule loss occurs along the remaining barb segments, this can lead to elevated brightness and reduced UVC due to the loss of light-absorbing components (see Laczi et al., 2019). In the special case of the male breast, the seasonal brightness reduction may occur due to barbule loss, reducing the reflective area that creates glossiness. These explanations align with the seasonal breakdown in stripe size (Adamík and Vaňáková, 2011), which can only be caused by wear.
From an ultimate perspective, the sex-related colour pattern may be meaningful as an amplifier of sex identity, combined with the stripe width differences between sexes. Such an explanation has been suggested, for example, for the whole-plumage colouration and stripe size in the green-backed tit (Parus monticolus) (Hofmann et al., 2007), or in Salamandrina species where both the gular patch morphology and brightness were sex-dependent (Ancillotto et al., 2022). Amplifiers are traits that enhance the perceived differences in traits signalling quality (Hasson, 1991). This may be particularly important for the great tit abdominal stripe considering that the true size of the stripe probably cannot be estimated reliably by conspecifics due to a high degree of error, as apparent width is significantly affected by the actual positions of individual feathers (i.e. we can ‘comb’ greater or smaller patch size to an individual in the hand by adjusting the feathers). It is also possible that the hidden UV breast patch of males serves as an ornament, and the less bright and less UV-saturated black throat and belly regions serve as amplifiers by forming heightened within-pattern contrasts (Doucet and Meadows, 2009). The inverse female colour pattern (darker breast with brighter surroundings) may specifically play a role in female–female interactions or male mate choice, for example. Systematic differences in contrast to the surrounding yellow feather areas are unlikely to confound the patterns we described here, because yellow feather reflectance seems to vary largely independently of melanin-based colour in our population (Hegyi et al., 2015).
It is important to underline that the appearance with regards to glossiness of the breast of males slightly depends on the angle of the plumage surface to the incoming light (pers. obs.), similarly to other species (Reed et al., 2020). This phenomenon gives the signaller an opportunity to change the conspicuousness of its ornament via behavioural adjustments according to the ambient light orientation (Sicsú et al., 2013; Simpson and McGraw, 2018). This could be important in great tits too, as during frontal encounters such as courtship or aggressive displaying (for display repertoire in great tits, see figures in Jones, 1968), the black stripe is elevated and it may be more exposed to iridescence-generating light, playing important roles in social interactions in both sexes (e.g. Järvi and Bakken, 1984; Wilson, 1992; Thys et al., 2020).
From the viewpoint of patterns, it could be conceivable that not only the colour of the single patch itself may have signal value, but also the contrast between integumental regions [see manakins (Heindl and Winkler, 2003), humans (Lu et al., 2022) or tamarins (Moreira et al., 2023)], as contrast may increase signal transmission and conspicuousness or attractiveness. In other animals, where sexes share the same colour pattern (as in the great tit), humans may perceive sex-differences in pattern contrast, as, for example, among vipers (Shine and Madsen, 1994). However, our findings highlight that such contrast-related dichromatism will be judged as more common if we consider cryptic colour differences, including not only its chromatic but also the overlooked achromatic dimensions. The absolute change in colouration of bird plumage between two time periods can also be a signal (e.g. Laczi et al., 2021), or the degree of its actual expression can convey different information at different times (Hegyi et al., 2019). Based on this, it is possible that, for example, at the time of pairing and territory establishment period (i.e. in winter), the breast patch could be a courtship signal, while later, during the incubation and nestling rearing period, it may reflect the parental abilities due to the variation of actual state and hence different colour properties of feathers. In addition, the coherence of expression of the components of a complex signalling system and the change in relative component expression may also be informative (Hegyi et al., 2022).
Taken together, colouration of different parts of the great tit abdominal stripe should be examined as sexual ornaments or signal amplifiers. From a methodological viewpoint, revealing the existence of (for the human eye) hidden patches may also help us to choose more accurate methods for quantifying patch size, including selecting the approximate boundaries of the trait in question (see Figuerola and Senar, 2000), and helping detect composite signals.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
The animal study was approved by the Ethical Committee of the Eötvös Loránd University. The study was conducted under a long-term research agreement with the Pilis Park Forestry, and with research permits from the regional nature conservation authority (PE-06/KTF/920–7/2018). All applicable international, national, and institutional guidelines for the use of animals were followed. The study was conducted in accordance with the local legislation and institutional requirements.
ML: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. GáH: Conceptualization, Funding acquisition, Supervision, Visualization, Writing – review & editing. GS: Data curation, Investigation, Writing – review & editing. HG: Data curation, Investigation, Writing – review & editing. FS: Data curation, Investigation, Writing – review & editing. JT: Investigation, Resources, Writing – review & editing. GeH: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Resources, Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was funded by the NKFIH (grant number: K124443, K143222) and the Doctoral School of Biology, Eötvös Loránd University (PhD Scholarship to HG, FS, GS).
We are grateful to the Behavioural Ecology Group.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1263974/full#supplementary-material
Adamík P., Vaňáková M. (2011). Feather ornaments are dynamic traits in the Great Tit Parus major. Ibis 153, 357–362. doi: 10.1111/j.1474-919X.2010.01097.x
Amézquita A., Ramos Ó., González M. C., Rodríguez C., Medina I., Simões P. I., et al. (2017). Conspicuousness, color resemblance, and toxicity in geographically diverging mimicry: The pan-Amazonian frog Allobates femoralis. Evolution 71, 1039–1050. doi: 10.1111/evo.13170
Ancillotto L., Vignoli L., Martino J., Paoletti C., Romano A., Bruni G. (2022). Sexual dichromatism and throat display in spectacled salamanders: a role in visual communication? J. Zool. 318, 75–83. doi: 10.1111/jzo.13006
Armenta J. K., Dunn P. O., Whittingham L. A. (2008). Quantifying avian sexual dichromatism: a comparison of methods. J. Exp. Biol. 211, 2423–2430. doi: 10.1242/jeb.013094
Badyaev A. V., Hill G. E. (2000). Evolution of sexual dichromatism: contribution of carotenoid- versus melanin-based coloration. Biol. J. Linn. Soc 69, 153–172. doi: 10.1111/j.1095-8312.2000.tb01196.x
Bowmaker J. K., Heath L. A., Wilkie S. E., Hunt D. M. (1997). Visual pigments and oil droplets from six classes of photoreceptor in the retinas of birds. Vis. Res. 37, 2183–2194. doi: 10.1016/S0042-6989(97)00026-6
Bradley B. J., Mundy N. I. (2008). The primate palette: the evolution of primate coloration. Evol. Anthrop. 17, 97–111. doi: 10.1002/evan.20164
Burns K. J., Shultz A. J. (2012). Widespread cryptic dichromatism and ultraviolet reflectance in the largest radiation of Neotropical songbirds: implications of accounting for avian vision in the study of plumage evolution. Auk 129, 211–221. doi: 10.1525/auk.2012.11182
Cally J. G., Stuart-Fox D., Holman L., Dale J., Medina I. (2021). Male-biased sexual selection, but not sexual dichromatism, predicts speciation in birds. Evolution 75, 931–944. doi: 10.1111/evo.14183
Cummings M. E., Rosenthal G. G., Ryan M. J. (2003). A private ultraviolet channel in visual communication. Proc. R. Soc Lond. B 270, 897–904. doi: 10.1098/rspb.2003.2334
D’Alba L., Van Hemert C., Spencer K. A., Heidinger B. J., Gill L., Evans N. P., et al. (2014). Melanin-based color of plumage: role of condition and of feathers’ microstructure. Integr. Comp. Biol. 54, 633–644. doi: 10.1093/icb/icu094
Dale J., Dey C. J., Delhey K., Kempenaers B., Valcu M. (2015). The effects of life history and sexual selection on male and female plumage colouration. Nature 527, 367–370. doi: 10.1038/nature15509
Delhey K., Burger C., Fiedler W., Peters A. (2010). Seasonal changes in colour: a comparison of structural, melanin-and carotenoid-based plumage colours. PloS One 5, e11582. doi: 10.1371/journal.pone.0011582
Delhey K., Peters A. (2017). The effect of colour-producing mechanisms on plumage sexual dichromatism in passerines and parrots. Funct. Ecol. 31, 903–914. doi: 10.1111/1365-2435.12796
Delhey K., Peters A., Johnsen A., Kempenaers B. (2006). Seasonal changes in blue tit crown color: do they signal individual quality? Behav. Ecol. 17, 790–798. doi: 10.1093/beheco/arl012
Doucet S. M., Meadows M. G. (2009). Iridescence: a functional perspective. J. R. Soc Interface 6, S115–S132. doi: 10.1098/rsif.2008.0395.focus
Eaton M. D. (2005). Human vision fails to distinguish widespread sexual dichromatism among sexually ‘monochromatic’ birds. Proc. Natl. Acad. Sci. U.S.A. 102, 10942–10946. doi: 10.1073/pnas.0501891102
Eaton M. D., Lanyon S. M. (2003). The ubiquity of avian ultraviolet plumage reflectance. Proc. R. Soc Lond. B 270, 1721–1726. doi: 10.1098/rspb.2003.2431
Emberts Z., Wiens J. J. (2022). Why are animals conspicuously colored? Evolution of sexual versus warning signals in land vertebrates. Evolution 76, 2879–2892. doi: 10.1111/evo.14636
Evans S. R., Hinks A. E., Wilkin T. A., Sheldon B. C. (2010). Age, sex and beauty: methodological dependence of age-and sex-dichromatism in the great tit Parus major. Biol. J. Linn. Soc 101, 777–796. doi: 10.1111/j.1095-8312.2010.01548.x
Evans S. R., Summers A. G., Sheldon B. C. (2012). Seasonality of carotenoid-based plumage coloration: modelling wavelength-specific change through spectral reconstruction. J. Avian Biol. 43, 234–243. doi: 10.1111/j.1600-048X.2012.05654.x
Figuerola J., Senar J. C. (2000). Measurement of plumage badges: an evaluation of methods used in the Great Tit Parus major. Ibis 142, 482–484. doi: 10.1111/j.1474-919X.2000.tb04445.x
Figuerola J., Senar J. C. (2005). Seasonal changes in carotenoid- and melanin-based plumage coloration in the Great Tit Parus major. Ibis 147, 797–802. doi: 10.1111/j.1474-919x.2005.00461.x
Font E., Pérez I de Lanuza G., Sampedro C. (2009). Ultraviolet reflectance and cryptic sexual dichromatism in the ocellated lizard, Lacerta (Timon) lepida (Squamata: Lacertidae). Biol. J. Linn. Soc 97, 766–780. doi: 10.1111/j.1095-8312.2009.01251.x
Galván I. (2011). Feather microstructure predicts size and colour intensity of a melanin-based plumage signal. J. Avian Biol. 42, 473–479. doi: 10.1111/j.1600-048X.2011.05533.x
Griffith S. C., Buchanan K. L. (2010). The zebra finch: the ultimate Australian supermodel. Emu 110, v–xii. doi: 10.1071/MUv110n3_ED
Hart N. S., Partridge J. C., Cuthill I. C., Bennett A. T. D. (2000). Visual pigments, oil droplets, ocular media and cone photoreceptor distribution in two species of passerine bird: the blue tit (Parus caeruleus L.) and the blackbird (Turdus merula L.). J. Comp. Physiol. A 186, 375–387. doi: 10.1007/s003590050437
Hasson O. (1991). Sexual displays as amplifiers: practical examples with an emphasis on feather decorations. Behav. Ecol. 2, 189–197. doi: 10.1093/beheco/2.3.189
Hegyi G., Laczi M., Boross N., Jablonszky M., Kötél D., Krenhardt K., et al. (2019). When to measure plumage reflectance: a lesson from collared flycatchers Ficedula albicollis. Ibis 161, 27–34. doi: 10.1111/ibi.12609
Hegyi G., Laczi M., Herényi M., Markó G., Nagy G., Rosivall B., et al. (2022). Functional integration of multiple sexual ornaments: signal coherence and sexual selection. Am. Nat. 200, 486–505. doi: 10.1086/720620
Hegyi G., Laczi M., Nagy G., Szász E., Kötél D., Török J. (2015). Stable correlation structure among multiple plumage colour traits: can they work as a single signal? Biol. J. Linn. Soc 114, 92–108. doi: 10.1111/bij.12412
Hegyi G., Szigeti B., Török J., Eens M. (2007). Melanin, carotenoid and structural plumage ornaments: information content and role in great tits Parus major. J. Avian Biol. 38, 698–708. doi: 10.1111/j.2007.0908-8857.04075.x
Heindl M., Winkler H. (2003). Interacting effects of ambient light and plumage color patterns in displaying wire-tailed manakins (Aves, Pipridae). Behav. Ecol. Sociobiol. 53, 153–162. doi: 10.1007/s00265-002-0562-3
Hernández-Palma A. (2016). Light matters: testing the ‘Light Environment Hypothesis’ under intra-and interspecific contexts. Ecol. Evol. 6, 4018–4031. doi: 10.1002/ece3.2188
Hofmann C., Lo W. S., Yao C. T., Li S. H. (2007). Cryptic sexual dichromatism occurs across multiple types of plumage in the Green-backed Tit Parus monticolus. Ibis 149, 264–270. doi: 10.1111/j.1474-919X.2006.00625.x
Iskandar J. P., Eliason C. M., Astrop T., Igic B., Maia R., Shawkey M. D. (2016). Morphological basis of glossy red plumage colours. Biol. J. Linn. Soc 119, 477–487. doi: 10.1111/bij.12810
Järvi T., Bakken M. (1984). The function of the variation in the breast stripe of the great tit (Parus major). Anim. Behav. 32, 590–596. doi: 10.1016/S0003-3472(84)80296-1
Jones N. G. B. (1968). Observations and experiments on causation of threat displays of the great tit (Parus major). Anim. Behav. Monogr. 1, 73–158. doi: 10.1016/S0066-1856(68)80002-0
Kimball R. T., Ligon J. D. (1999). Evolution of avian plumage dichromatism from a proximate perspective. Am. Nat. 154, 182–193. doi: 10.1086/303228
Kodric-Brown A. (1998). Sexual dichromatism and temporary color changes in the reproduction of fishes. Am. Zool. 38, 70–81. doi: 10.1093/icb/38.1.70
Koski M. H. (2020). Macroevolution of flower color patterning: biased transition rates and correlated evolution with flower size. Front. Plant Sci. 11, 945. doi: 10.3389/fpls.2020.00945
Kraaijeveld K., Kraaijeveld-Smit F. J., Maan M. E. (2011). Sexual selection and speciation: the comparative evidence revisited. Biol. Rev. 86, 367–377. doi: 10.1111/j.1469-185X.2010.00150.x
Kuznetsova A., Brockhoff P. B., Christensen R. H. (2017). lmerTest package: tests in linear mixed effects models. J. Stat. Software 82, 1–26. doi: 10.18637/jss.v082.i13
Laczi M., Balogh J., Nardou X., Török J., Hegyi G. (2021). The meaning of purely structural colour: white plumage reflectance indicates feather condition. Ibis 163, 407–416. doi: 10.1111/ibi.12902
Laczi M., Hegyi G., Kötél D., Csizmadia T., Lőw P., Török J. (2019). Reflectance in relation to macro-and nanostructure in the crown feathers of the great tit (Parus major). Biol. J. Linn. Soc 127, 113–124. doi: 10.1093/biolinnean/blz016
Laczi M., Hegyi G., Nagy G., Pongrácz R., Török J. (2020). Yellow plumage colour of Great Tits Parus major correlates with changing temperature and precipitation. Ibis 162, 232–237. doi: 10.1111/ibi.12761
Lu Y., Xiao K., Yang J., Pointer M., Li C., Wuerger S. (2022). Different colour predictions of facial preference by Caucasian and Chinese observers. Sci.Rep. 12, 12194. doi: 10.1038/s41598-022-15951-8
Lunau K. (1992). A new interpretation of flower guide colouration: absorption of ultraviolet light enhances colour saturation. Plant Syst. Evol. 183, 51–65. doi: 10.1007/BF00937735
Luro A. B., Hauber M. E. (2022). Pressure for rapid and accurate mate recognition promotes avian-perceived plumage sexual dichromatism in true thrushes (genus: Turdus). J. Evol. Biol. 35, 1558–1567. doi: 10.1111/jeb.14089
Maia R., D'Alba L., Shawkey M. D. (2011). What makes a feather shine? A nanostructural basis for glossy black colours in feathers. Proc. R. Soc Lond. B 278, 1973–1980. doi: 10.1098/rspb.2010.1637
Maia R., Gruson H., Endler J. A., White T. E. (2019). pavo 2: new tools for the spectral and spatial analysis of colour in R. Methods Ecol. Evol. 10, 1097–1107. doi: 10.1111/2041-210X.13174
Mason N. A., Bowie R. C. (2020). Plumage patterns: Ecological functions, evolutionary origins, and advances in quantification. Auk 137, ukaa060. doi: 10.1093/auk/ukaa060
Miller E. C., Mesnick S. L., Wiens J. J. (2021). Sexual dichromatism is decoupled from diversification over deep time in fishes. Am. Nat. 198, 232–252. doi: 10.1086/715114
Moreira L. A., Watsa M., Erkenswick G., Higham J. P., Melin A. D. (2023). Evaluating genital skin color as a putative sexual signal in wild saddleback (Leontocebus weddelli) and emperor (Saguinus imperator) tamarins. Am. J. Primatol. 85, e23456. doi: 10.1002/ajp.23456
Negro J. J., Figueroa-Luque E., Galván I. (2018a). Melanin-based sexual dichromatism in the Western Palearctic avifauna implies darker males and lighter females. J. Avian Biol. 49, jav–01657. doi: 10.1111/jav.01657
Negro J. J., Figueroa-Luque E., Galván I. (2018b). Data from: Melanin-based sexual dichromatism in the Western Palearctic avifauna implies darker males and lighter females. Dryad Dataset. doi: 10.5061/dryad.q8r422h
Norris K. J. (1990). Female choice and the quality of parental care in the great tit Parus major. Behav. Ecol. Sociobiol. 27, 275–281. doi: 10.1007/BF00164900
Norris K. J. (1993). Heritable variation in a plumage indicator of viability in male great tits Parus major. Nature 362, 537–539. doi: 10.1038/362537a0
Örnborg J., Andersson S., Griffith S. C., Sheldon B. C. (2002). Seasonal changes in a ultraviolet structural colour signal in blue tits, Parus caeruleus. Biol. J. Linn. Soc 76, 237–245. doi: 10.1046/j.1095-8312.2002.00061.x
Pérez i de Lanuza G., Font E., Monterde J. L. (2013). Using visual modelling to study the evolution of lizard coloration: sexual selection drives the evolution of sexual dichromatism in lacertids. J. Evol. Biol. 26, 1826–1835. doi: 10.1111/jeb.12185
Price J. J. (2019). Sex differences in song and plumage color do not evolve through sexual selection alone: new insights from recent research. J. Ornithol. 160, 1213–1219. doi: 10.1007/s10336-019-01681-8
R Development Core Team (2022). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Reed S., Simpson R. K., McGraw K. J. (2020). Feather morphological predictors of angle-dependent color changes in parrot plumage. Avian Biol. Res. 13, 108–117. doi: 10.1177/1758155920963198
Rojas B. (2017). Behavioural, ecological, and evolutionary aspects of diversity in frog colour patterns. Biol. Rev. 92, 1059–1080. doi: 10.1111/brv.12269
Rossi N., Benitez-Vieyra S., Cocucci A., Chiaraviglio M., Cardozo G. (2019). Sexual dichromatism and color diversity in the spiny lava lizard Tropidurus spinulosus using lizard visual modelling. Sci. Rep. 9, 14270. doi: 10.1038/s41598-019-50712-0
Seddon N., Botero C. A., Tobias J. A., Dunn P. O., MacGregor H. E., Rubenstein D. R., et al. (2013). Sexual selection accelerates signal evolution during speciation in birds. Proc. R. Soc Lond. B 280, 20131065. doi: 10.1098/rspb.2013.1065
Senar J. C., Conroy M. J., Quesada J., Mateos-Gonzalez F. (2014). Selection based on the size of the black tie of the great tit may be reversed in urban habitats. Ecol. Evol. 4, 2625–2632. doi: 10.1002/ece3.999
Shine R., Madsen T. (1994). Sexual dichromatism in snakes of the genus Vipera: a review and a new evolutionary hypothesis. J.Herpetol. 28, 114–117. doi: 10.2307/1564692
Shultz A. J., Burns K. J. (2017). The role of sexual and natural selection in shaping patterns of sexual dichromatism in the largest family of songbirds (Aves: Thraupidae). Evolution 71, 1061–1074. doi: 10.1111/evo.13196
Sicsú P., Manica L. T., Maia R., Macedo R. H. (2013). Here comes the sun: multimodal displays are associated with sunlight incidence. Behav. Ecol. Sociobiol. 67, 1633–1642. doi: 10.1007/s00265-013-1574-x
Siddiqi A., Cronin T. W., Loew E. R., Vorobyev M., Summers K. (2004). Interspecific and intraspecific views of color signals in the strawberry poison frog Dendrobates pumilio. J. Exp. Biol. 207, 2471–2485. doi: 10.1242/jeb.01047
Simpson R. K., McGraw K. J. (2018). Two ways to display: male hummingbirds show different color-display tactics based on sun orientation. Behav. Ecol. 29, 637–648. doi: 10.1093/beheco/ary016
Soler J. J., Moreno J. (2012). Evolution of sexual dichromatism in relation to nesting habits in European passerines: a test of Wallace’s hypothesis. J. Evol. Biol. 25, 1614–1622. doi: 10.1111/j.1420-9101.2012.02544.x
Stella D., Faltýnek Fric Z., Rindoš M., Kleisner K., Pecháček P. (2018). Distribution of ultraviolet ornaments in Colias butterflies (Lepidoptera: Pieridae). Environ. Entomol. 47, 1344–1354. doi: 10.1093/ee/nvy111
Surmacki A., Liu M., Mercadante A., Hill G. E. (2011). Effect of feather abrasion on structural coloration in male eastern bluebirds Sialia sialis. J. Avian Biol. 42, 514–521. doi: 10.1111/j.1600-048X.2011.05503.x
Surmacki A., Nowakowski J. K. (2007). Soil and preen waxes influence the expression of carotenoid-based plumage coloration. Naturwissenschaften 94, 829–835. doi: 10.1007/s00114-007-0263-x
Taysom A. J., Stuart-Fox D., Cardoso G. C. (2011). The contribution of structural-psittacofulvin-and melanin-based colouration to sexual dichromatism in Australasian parrots. J. Evol. Biol. 24, 303–313. doi: 10.1111/j.1420-9101.2010.02166.x
Thys B., Pinxten R., Eens M. (2020). Does the tie fit the female? Melanin-based colouration, aggressive personality and reproductive investment in female great tits (Parus major). Behav. Ecol. Sociobiol. 74, 1–11. doi: 10.1007/s00265-020-2828-z
Keywords: colouration, cryptic dichromatism, great tit, melanin, Parus major
Citation: Laczi M, Herczeg G, Szabó G, Gyarmathy H, Sarkadi F, Török J and Hegyi G (2023) The great tit abdominal stripe contains a sexually dichromatic colour patch hidden from the human eye. Front. Ecol. Evol. 11:1263974. doi: 10.3389/fevo.2023.1263974
Received: 20 July 2023; Accepted: 22 September 2023;
Published: 06 October 2023.
Edited by:
Jan Lindstrom, University of Glasgow, United KingdomReviewed by:
Liliana D’Alba, Naturalis Biodiversity Center, NetherlandsCopyright © 2023 Laczi, Herczeg, Szabó, Gyarmathy, Sarkadi, Török and Hegyi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Miklós Laczi, bWlrbG9zLmxhY3ppQHR0ay5lbHRlLmh1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.