
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol., 18 August 2023
Sec. Population, Community, and Ecosystem Dynamics
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1258185
This article is part of the Research TopicEcological Links between Aboveground and Underground Ecosystems under Global ChangeView all 7 articles
 Zhenhui Ding1
Zhenhui Ding1 Ao Guo1Meng Lian2Yan Wang3Wenjia Ying1Hao Jiang1Xingyu Zhou1Cheng Qian1Jiangshan Lai2*Jiajie Cao1*
Ao Guo1Meng Lian2Yan Wang3Wenjia Ying1Hao Jiang1Xingyu Zhou1Cheng Qian1Jiangshan Lai2*Jiajie Cao1*Introduction: Understanding the birds’ breeding strategies in urban habitats is vital for ensuring their continued existence. Therefore, more research must be conducted on bird breeding and urban adaptation strategies in urban green spaces. This study aimed to address this gap by investigating the influence of landscape factors on the selection of bird nest sites.
Methods: Data on the presence and absence of magpie (Pica pica) and gray magpie (Cyanopica cyana) nests were collected through field surveys conducted in the campus of Nanjing Forestry University during the 2023 breeding season. Generalized additive models (GAMs) incorporating landscape variables were employed to assess the effects of these predictors on nest occurrence. The model with the lowest Akaike’s information criterion value was selected among the candidate GAMs.
Results: Below is a summary of the main results. Nest tree height (TH), distance from the central lawn (D), and tree coverage (TC) within the sampled area were identified as the primary landscape factors influencing nest site choice. Conversely, factors such as the shortest distance to the water source, herb coverage, shrub coverage, percentage of buildings, and percentage of hard pavement did not significantly impact on nest site selection. Furthermore, the nesting potential of magpies and grey magpies initially increased with tree height, reaching a maximum at ca. TH=25 meters after which it began to decline. The nesting occurrence rate showed an initial decrease tendency with increasing distance from the central lawn, reaching a minimum at D=400 meters, and then increased with further distance. Additionally, nesting potential decreased initially with increasing of TC in the range of 0–20%, fluctuated evenly between 20–60% TC, and decreased rapidly when TC exceeded 60%.
Discussion: This study provides valuable insights into the selection of nest sites by birds in urban habitats, specifically with respect to landscape factors. The understanding of the impact of urban green spaces on urban birds and the underlying mechanisms of their behavior contributes to the conservation of wild birds and promotes the harmonious development of urban areas.
Understanding the impact of the urban environment on wildlife is becoming increasingly urgent and important, as more and more species are settling in urban areas. Urbanization is an inevitable consequence of human economic and social development. The expansion of urban areas is considered to be one of the most important anthropogenic impacts on Earth’s ecosystems (Foley et al., 2005), resulting in substantial alterations to natural habitat structure, ecosystem function and biodiversity (Gaston et al., 2010; Gil and Brumm, 2013; Forman, 2014). Numerous life-history strategies of animals have undergone modifications in response to the anthropogenic changes associated with urban environments (Arroyo-Solís et al., 2013; Kivelä et al., 2014). Urbanization presents numerous challenges for organisms due to extensive and dramatic changes in urban habitats, including replacement and fragmentation of natural vegetation, shifts in predators and food resources in communities, and increased human disturbance and pollution (Lowry et al., 2012; Sol et al., 2013). While high-density urbanization provides convenience for humans, high-density built environments have adverse effects on wildlife. Although urbanization often negatively affects some native species and even leads to their extinction (Soulé et al., 1988; McKinney, 2006; Husté and Boulinier, 2007; Sol et al., 2014), there are increasing total abundances of bird species that are successfully occupying and adapting to these new urban environments (Luniak, 2004; Francis and Chadwick, 2012).
The ecological habitats, distribution characteristics, and community structure of birds, as an important indicator species of the urban environment, constitute the main content of basic research on urban birds. Understanding urban birds and exploring the relationship between birds and the urban environment is based on the structure and dynamics of bird communities (Zheng et al., 2008). This study specifically focuses on the bird guild and community structure of cities, bird community surveys, bird community comparisons, and the seasonal or inter annual dynamics of urban bird community structure. A notable research topic is the comparison of bird communities in different urban green spaces or habitats (Zhang and Huang, 2018). Significant differences in bird diversity have been found between habitat types. The richness and diversity of bird communities in cities depend on the richness and diversity of the urban green spaces. The distribution of bird communities in urban green spaces is determined by the bioclimatic area, as well as the type and degree of urbanization (Clergeau et al., 2006). However, it is not necessarily the case that the maximum richness and diversity of bird communities are achieved in less urbanized areas (Jokimäki and Suhonen, 1993; Carbó-Ramírez and Zuria, 2011). Bird communities select habitats of different degrees of urbanization according to their habits. For instance, anthropophilic species take advantage of human activities in highly urbanized areas, while bird species inhabiting agroforestry vegetation are present in low urbanized areas (Burger et al., 2004; Marzluff and Rodewald, 2008; Parker and Nilon, 2012).
The design and management of urban green spaces play a vital role in determining the diversity and richness of bird communities. Two key factors that affect these communities are the composition and structure of the vegetation within these spaces (MacGregor-Fors and Schondube, 2011). On the one hand, the composition of plant communities is closely related to bird diversity (James and Wamer, 1982; Huang et al., 2015). The researchers found significant correlations between the heterogeneity and abundance of trees with the richness and density of birds. This study also revealed a positive correlation between bird diversity and richness of flowering plants (Blinkova and Shupova, 2017). On the other hand, the volume and density of plants in urban green areas positively impact bird richness and diversity (Savard et al., 2000; Mella and Loutit, 2007). Therefore, thinning of trees and shrubs is counterproductive (Camprodon and Brotons, 2006). Inappropriate vegetation structure in urban green spaces can also lead to a further reduction in bird diversity (Ge et al., 2005; Xu et al., 2007; Yang et al., 2015). Therefore, by providing suitable microclimates, refuges, abundant food resources, and nesting areas with reduced competition and predation, urban green spaces offer birds agreeable living conditions (Magre et al., 2019).
Among these elements, trees are considered as one of the most important elements to increase bird richness and diversity in urban green spaces (Palomino and Carrascal, 2006; Yang et al., 2015; Weaving et al., 2016). Tree canopies provide sheltering, nesting sites and feeding opportunities (Munyenyembe et al., 1989). Specially, birds use dense tree canopies, tree trunk with holes and branches that produce fruits or seeds. The presence of these resources for bird’ refuge, nesting and breeding promotes the access of adjacent flora and fauna into the urban green spaces (Boada and Sánchez, 2012). In summary, promoting urban green management actions that lead to a suitable composition and structure of vegetation in cities can potentially enhance the diversity and richness of birds residing in these areas (Camprodon and Brotons, 2006; Shanahan et al., 2011).
Urbanization has brought about changes in the resources upon which birds depend, such as habitat types and availability (Camprodon and Brotons, 2006; Ibáñez-Álamo and Soler, 2010). To survive and breed successfully in urban environments, birds have had to adapt to trade-offs between the pressures specific to urban settings and the benefits that these new habitats offer (Grinnell, 1924; Wang et al., 2015). Species living in urban environments have to adapt their behavior and life histories to the new environment. Urbanization has resulted in differences in genetic, reproductive and ecological characteristics of bird populations compared to those found in natural habitats (Luniak and Mulsow, 1988; Partecke et al., 2004; Wysocki, 2004; Partecke et al., 2006). Breeding, as the most critical stage in a bird’s life history, is an important issue for birds in adapting to the disturbances of the urban environment and successfully breeding their offspring (Meillère et al., 2015). Adaptations in nesting behaviour reflect the ability of birds to adapt to urban ecosystems and have been demonstrated in a variety of bird species (Luniak, 2004; Wang et al., 2009). Nest site selection is a significant aspect of breeding as it minimizes the impact of natural predators, disturbance from similar species, and other factors, thus enhancing breeding success. It is an important means by which birds can adapt to a complex and changing environment (Lack, 1969). Various environmental features can influence nest site selection, which in turn affects bird survival, breeding, and adaptation (Kivelä et al., 2014; Sepp et al., 2018).
Despite the increasing knowledge about urban nesting behavior, there is still relatively little understanding of the predictability of nesting behavioral shifts in urban areas. Most studies on the richness and diversity of birds in urban environments primarily focus on investigating e breeding success by analyzing nest depredation rates. These studies have shown that nest depredation rates tend to be higher when nests are associated with specific mammals and predatory birds (Miller et al., 1998; Matthews et al., 1999; Jokimäki and Huhta, 2000; Phillips et al., 2005; Reale and Blair, 2005; Bakermans and Rodewald, 2006; Burhans and Thompson, 2006; Smith-Castro, 2008). In Mediterranean cities, cats (Stracey, 2011) and magpies (Bonnington et al., 2015) are the main cause of nest depredation. However, the only magpie species found in urban areas (Pica pica) actually prefers nearby rural areas (Andren, 1992). Other studies conducted in peri-urban areas have found that nests located at low heights experience higher rates of depredation due to the influx of domestic animals (Miller et al., 1998), while those at higher heights remain better conserved (Smith-Castro, 2008). Nonetheless, research on the effects of specific landscape factors on bird nesting in urban green spaces is limited.
The main objective of this paper is to test the influence of various landscape factors associated with urban habitats on the selection of nest sites for magpie and gray magpies in green spaces on the campus of Nanjing Forestry University. A field survey was conducted during the 2023 breeding season, resulting in a collection of 152 presence–absence data points. Generalized additive models (GAMs) (Hastie and Tibshirani, 1987; Wood, 2004) were employed to assess the effects of landscape variables on the occurrence of nests. The possibilities of bird nest site selection were compared and analyzed in relation to different landscape factors. These models incorporated various predictor variables related to the surrounding landscape. To determine the best model, several candidate GAMs were employed, considering all significant predictor variables and selecting the model with the lowest Akaike’s information criterion (AIC) value. The findings of this study may serve as a valuable tool for urban green managers and technicians aiming to enhance urban biodiversity. Furthermore, the study contributes to a deeper understanding of the mechanisms through which birds adapt to urbanized habitats.
Nanjing, residing in the southwestern part of Jiangsu Province, at the central region of the lower Yangtze River, is located between 31°14′N to 32°37′N latitude and 118°22′E to 119°14′E longitude. The city experiences a subtropical monsoon climate, characterized by four distinct seasons. It has cold winters and hot summers, with abundant precipitation resulting in an average annual rainfall of 1005.9mm, and a relative humidity average of 76%. The city encounters maximum wind speeds reaching up to 25 m/s, with prevailing winds originating from the southeast and east during the summer months, and from the northeast and east during winter. The annual absolute minimum temperature plunges to −14°C, while the annual absolute maximum temperature reaches 43°C. The yearly average temperature is recorded at 15.7°C, with the coldest month averaging at −1.5°C, and the hottest month averaging at 28.1°C (Wang et al., 2016). In fact, Nanjing holds significant importance as an essential node in the East Asia–Australasia bird migration route. Additionally, Nanjing’s geographic location, particularly its proximity to the lower Yangtze River and Xuanwu Lake, makes it an important wintering site for various waterfowl species.
Field work was conducted on the Xinzhuang campus of Nanjing Forestry University (NJFU), which covers an area of 83.8 hectares with a green space ratio of 53.8% and a green coverage of 78% (Wang et al., 2016). The campus is well endowed with natural resources, including a large number of large and long-standing trees and a water system connecting the various landscape spaces, resulting in a high degree of habitat continuity and a rich variety of habitat types. It is surrounded by an urban matrix including commercial development, residential communities, and urban green space (Figure 1). Moreover, the campus has a close relationship with the core areas of Nanjing’s main urban area, Zijin Mountain and Xuanwu Lake, and is located between these core habitat sources (Ding et al., 2023). As a result, it is considered one of the larger urban forests in Nanjing. The known flora of the campus forest consists of 603 species (including subspecies and varieties) in 91 families, showing a high species richness. The campus also exhibits a variety of vegetation types, with the natural secondary forest dominated by deciduous broad-leaved and mixed evergreen and deciduous broad-leaved forests, while the planted forest consists of fewer plant species and a simpler hierarchy with a single species of street trees. In terms of species diversity, the herbaceous layer shows the highest diversity, followed by the shrub layer and the tree layer. The campus is particularly rich in herbaceous plants. The available records indicate the presence of 70 bird species from 29 families, including both resident and migratory species. Among them, magpie (Pica pica) and grey magpie (Cyanopica cyana), belonging to the family Corvidae, and included in the IUCN Red List, are the most widespread species in the study area. They are ideal species to study the relationship between landscape factors and urban nesting preference. The areas of observation activity mainly focus on open sites where birds are in close contact with human activity.
Figure 1 Schematic of study area: Nanjing Forestry University.
The field survey was conducted during the spring of 2023, spanning from February to July, which coincided with the breeding season. The primary objective of the survey was to investigate the landscape factors that influence nest site selection. The survey covered the entire campus area, and all nests within the sample area were located and recorded. Weekly nest site surveys were repeated to identify new nests, based on the characteristics of the birds’ life history and breeding processes in the study area. The survey was conducted in favorable weather between 09:00 and 16:00, using binoculars, a DJI Mavic air drone, and a digital camera for observation and photography (Gao et al., 2014). Upon the initial discovery of a new nest, the bird species occupying the nest was identified and the latitude, longitude, and altitude of the nest were recorded using a handheld GPS (eTrex30, Garmin, China). After nesting concluded, these sites were revisited to identify the vegetation of nesting substrate. The current usage status of each nest site was ensured through a minimum of 30 minutes of observation in the vicinity of the nest site. The center of the nesting trees was selected, and a 50 m×50 m square sample plot was used as a standard plot to count the area surrounding the sample square. Moreover, various landscape factors within the presence–absence sample square of the nest site was measured and recorded. This study collected a total of eight landscape factors that are related to bird nest site selection (see Table S1).
In order to explore the impact of landscape architecture elements on bird nest site selection in urban habitats, we employed semi-parametric generalized additive models with a logit link function (Hastie and Tibshirani, 1987; Wood, 2004). These models were utilized to analyze the effects of eight landscape variables on 152 presence–absence data points (Y=1 or 0) at each site. The landscape variables considered were tree height (TH), distance from the central lawn (D), tree coverage (TC), shortest distance to the water source, herbaceous coverage, shrub coverage, percentage of buildings, and percentage of hard pavement. Non-significant landscape variables were removed, and candidate GAMs (Table S2) were constructed using the remaining significant landscape variables. The model with the lowest Akaike’s Information Criterion (AIC) value was selected as the best model. To assess the nonlinear effects of each landscape variable on the occurrence of bird nest selection, we examined the partial residual plots of these variables within the best model. The ‘gam’ function in the ‘mgcv’ package (version 1.8-40; Wood, 2004) was used to carried out the GAM fit.
During the 2023 breeding season in Nanjing Forestry University, a comprehensive survey on the campus identified a total of 76 nests attributed to magpie and grey magpie (Figure 2). These nests were exclusively located in trees and were continuously monitored throughout the breeding season for subsequent statistical analysis. Among the 76 nests, the predominant nesting tree species were Metasequoia glyptostroboides, Platanus acerifolia, Pterocarya stenoptera, Populus euramevicana, and Liquidambar formosana, which collectively accounted for 60.54% of the total nests. Specifically, Metasequoia glyptostroboides accounted for 17.11% of the nests, while Platanus acerifolia and Pterocarya stenoptera accounted for 13.16%, Populus euramevicana accounted for 9.21%, and Liquidambar formosana accounted for 7.89% (Figure 3). Additionally, nests of magpies and grey magpies were found in various tree species, including representatives from Fagaceae, Magnoliaceae, Sapindaceae, Hamamelidaceae, Ginkgoaceae, Pinaceae, and Lauraceae families. The average height of trees hosting nests was determined to be 19.43 m, ranging from a minimum of 7.7 m to a maximum of 30.00 m. The average nest height was 16.86 m, with the highest nest recorded at 27 m and the lowest at 6.8 m. The minimum distance of nests from water sources was measured to be 171.85 m. Notably, the selection of nest sites for magpies and grey magpies in the sampled locations did not show a significant relationship with the type of nest tree. The majority of the nesting trees belonged to the Taxodiaceae and Salicaceae families, collectively representing 36.85% of all nesting trees, with the Taxodiaceae family accounting for 18.43% and the Salicaceae family accounting for 18.42%.
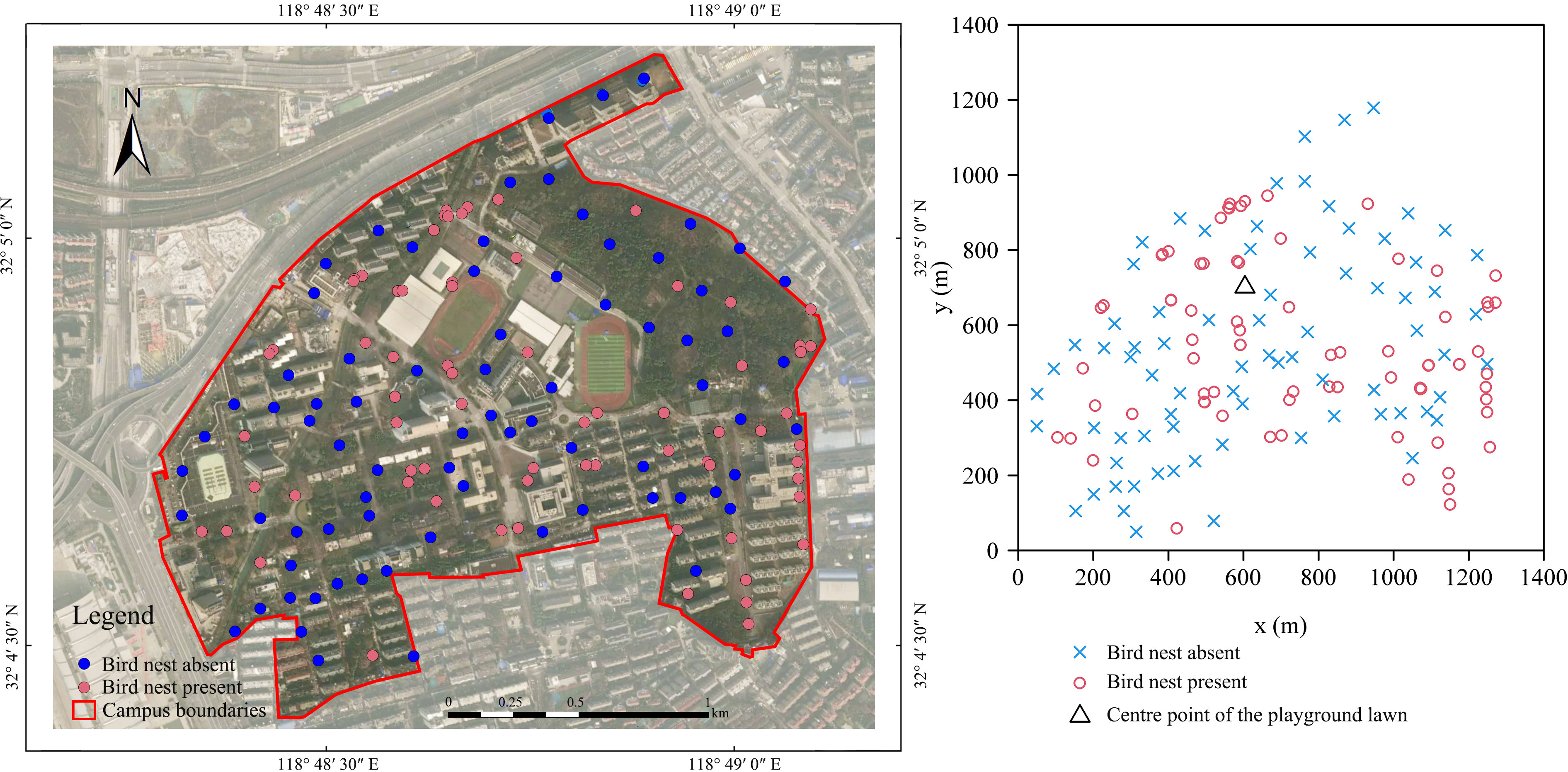
Figure 2 Spatial distribution of bird nests (presence–absence) at Nanjing Forestry University.
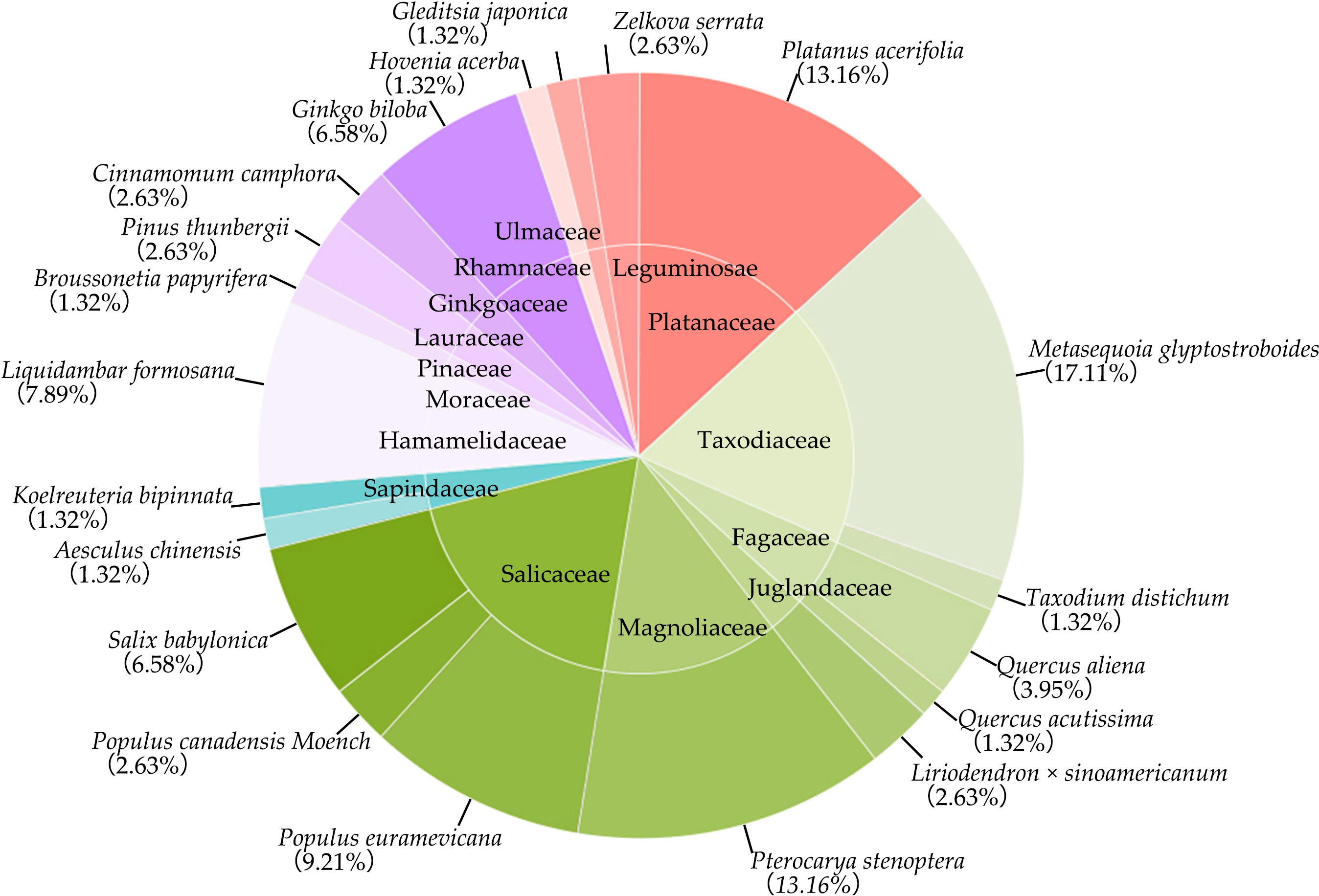
Figure 3 Composition and proportion of nesting tree species.
Model 4[y ~ s(TH) + s(d) + s(TC); AIC=183.64] (see Table S2) had the lowest AIC value among the seven candidate GAMs tested, thus it was chosen as the best model. The results of model 4 indicated significant effects of TH, D, TC on the occurrence of bird nest selection.
The odds of nesting by magpies and grey magpies exhibited a notable non-linear relationship with tree height, as illustrated in Figure 4. Initially, there was an upward trend in nesting probability with increasing tree height, reaching its peak at 25 m. However, beyond this point, the odds of nesting started to decline, indicating a diminishing trend. Moreover, as illustrated in Figure 4A, higher tree heights were found to inhibit nesting frequency.

Figure 4 For the presence–absence data, plots of the relationship between each feature and the response in the fitted model. Each plot displays the fitted function and pointwise standard errors. Three functions are all cubic splines in (A) tree height (TH), (B) distance (D) and (C) tree coverage (TC).
Figure 4B presents the influence of distance (D) from the central lawn of the playground on nest emergence odds. Initially, an increase in distance hindered nest emergence, but as the distance increased further, it began to promote nest emergence. The odds of nesting exhibited an initial decrease with increasing distance from the central lawn, reaching its minimum at D=400. Beyond this threshold, the odds of nesting started to increase with distance. The impact of tree coverage on nesting potential is clearly demonstrated in Figure 4C. It displayed an overall decreasing trend with some fluctuations, eventually leading to a sharp decline. The odds of nesting decreased as tree coverage increased from 0–20%, remained relatively stable between 20–60%, and then experienced a rapid decline for coverage exceeding 60%. Figure 4 demonstrates that the three primary influencing factors (tree height, distance from the central lawn, and tree coverage) exhibited significant non-linear relationships with the odds of nesting, displaying distinct phases of fluctuation.
Urban environments pose significant challenges to species adapted to natural ecosystems due to high population densities (Kight and Swaddle, 2007), intense artificial light (Dominoni et al., 2013; Gaston et al., 2013), chemical pollution (Hui, 2002), noise levels (Slabbekoorn and Ripmeester, 2008), and fragmented habitats (Grimm et al., 2008; Alberti et al., 2017; Johnson and Munshi-South, 2017). Successful nesting and breeding of bird species in urban habitats depend on the trade-offs they make between inputs and benefits in these environments. In this study conducted on the campus, 76 tree nests belonging to magpies and grey magpies were identified and analyzed. The results revealed no significant preference for specific tree species, but tree height, distance from the central lawn of the playground, and tree coverage emerged as the main factors influencing nest site selection. Nest site selection in urban habitats can be influenced by predator habits. Birds in cities have adapted their nesting strategies to mitigate predation risks, employing various tactics to minimize breeding losses. The findings of this study indicate that optimal nesting sites for magpies and grey magpies on campus were characterized by a tree height of 25 m, proximity to the central lawn of the playground, and tree coverage below 60%. Choosing such locations helps reduce the risk of predation during breeding, leading to successful reproduction. Several factors may contribute to the observed optimal strategy. Nesting at greater heights reduces the detection chances by ground predators such as snakes and rodents, as accessing higher nest locations becomes more challenging for these predators. (Rendell and Robertson, 1989). However, nests situated above 25 m height become vulnerable to aerial predators, which can lead to reproductive failure. Notably, the absence of the crested eagle (Cao et al., 2023), a known aerial predator of magpies and grey magpies, in previous studies conducted at Southern Forest University supports this finding. The nest site selection model of this study further suggests that the odds of nest site selection increases with increasing tree height between 0–25 m and decreases with increasing tree height between 25–30 m, allowing magpies and grey magpies to mitigate the risk of predation from ground and aerial predators.
Foliage surrounding the nest provides shade and reduces the odds of predation on the nesting birds (Martin, 1993). In addition to predator-related factors, magpies and grey magpies in this study showed a preference for nesting near the top of trees in high-density tree species in open green spaces. This finding suggests a trade-off between concealment and light requirements in nest site selection. The birds choose nest sites that offer sufficient concealment while also ensuring adequate light, which is beneficial for the normal development of chick feathers and the maintenance of nest microhabitat temperatures. Dense foliage also provides protection against adverse weather conditions such as wind, rain, and strong light (Deng et al., 2001). The study’s results indicate that the odds of nest site selection by magpies and grey magpies displayed an overall decreasing trend with some fluctuations, eventually leading to a sharp decline. The odds of nesting decreased as tree coverage increased from 0–20%, remained relatively stable between 20–60%, and then experienced a rapid decline for coverage exceeding 60%.
Furthermore, the study has shown that the choice of nest site for magpies and grey magpies decreases with increasing distance from the center of the playground in the 0–400 m range, with the odds reaching a minimum at a distance of 400 m. At distances greater than 400 m, the odds of nesting begins to increase with distance. The main school playground, with its real grass turf and central location on the campus, has been observed as a favorable foraging site for birds, providing grazing opportunities and access to insects. The anomalous decrease in nesting probability beyond 400 m may be attributed to the omnivorous nature of magpies and grey magpies (Kaplan, 2019), as they may find better foraging sites within and around the school grounds. Magpies exhibit a complex relationship with humans in urban environments, depending on and maintaining a certain distance from human activities, enabling them to access abundant food resources (Hao et al., 1992). They are considered active urban adapters.
Distance from water sources and human activity intensity were not identified as primary factors influencing nest site selection by magpies and grey magpies in this study. This finding can be attributed to the adaptability and flying abilities of these bird species, their larger activity range (Claramunt et al., 2022), the proximity of Xuanwu Lake (which offers better and easily accessible water supply) to the campus, and their overall strong resource access capabilities and adaptability to urban habitats.
Birds in urban environments face challenges that require them to carefully evaluate costs and benefits of their habitat choices. To thrive in cities, birds must weigh that the advantages of the urban ecosystem against its disadvantages. An ideal nesting site provides improved protection, stable materials for nest construction, and proximity to food-rich foraging habitats that support breeding breeding (Hafner and Fasola, 1992). However, urban environments also pose negative impacts on birds, necessitating additional energy expenditure and resource competition compared to their natural habitats. The increasing urban population and urbanization have led to higher levels of artificial light (Hölker et al., 2010; Falchi et al., 2016) and human noise (Job et al., 2016), demanding avian adaptation during crucial breeding periods that require additional resources. In response to urbanization, birds actively seek suitable nesting sites and develop optimal breeding strategies for their survival and prosperity in cities (Sepp et al., 2018).
Magpies and grey magpies are representative of bird species that can coexist with humans in urban ecosystems, displaying the ability to adapt to human activities to a certain extent. However, they also possess specific requirements that may conflict with human activities. For example, the preferred nesting environment provided by poplar trees presents challenges due to the adverse effects of these trees and their globular double-hanging fruit on the human environment. Some researchers and authorities suggest using lower evergreen trees like Magnolia grandiflora as alternatives (Liu et al., 2019). However, this poses a challenge to the survival of magpies and grey magpies, as they prefer nesting in deciduous trees at greater heights. Additionally, the impact of tree planting types, tree characteristics, institutional building patterns, and green space distribution on magpie and grey magpie nesting in green spaces warrants further investigation (Ding and Feeley, 2010).
This study aims to enhance the protection and cultivation of tall trees in highly urbanized areas and urban habitats such as schools, with the goal of safeguarding wildlife such as magpies and grey magpies. Achieving this requires increasing the number of tall iconic landscape structures while considering their compatibility with green space design. Urban planning and green space landscape design should prioritize retaining a mix of “high-medium-low” levels of tree species in new institutions or the renovation and upgrading of existing ones. Furthermore, various plant configuration patterns should be considered in the design process to ensure a proportionate balance of open grassland and open forest in the green space landscape. These measures will reduce damage to the native ecological base, promote ecological continuity within green spaces, mitigate habitat fragmentation, and contribute to the development of better and sustainable urban ecosystems where humans and nature coexist harmoniously.
In this study, we conducted field surveys, collecting 152 presence–absence data of bird nests, which were subjected to rigorous analysis using generalized additive models. The primary objective was to investigate the impact of landscape factors on bird nest site selection in urban green spaces and uncovering the adaptive strategies employed by birds in urban environments. Our findings have led to several significant conclusions:
Firstly, the distribution of bird nests in the campus habitat was strongly influenced by three key factors: nest tree height (TH), distance from the central lawn (D), and tree cover (TC). Notably, the proximity to water sources, herb coverage, shrub coverage, percentage of buildings, and percentage of hard pavement did not show a significant effect on nest site selection.
Secondly, for magpies and grey magpies, the odds of nesting initially increased with rising tree height, reaching a maximum at ca. TH=25 meters, after which it gradually declined with further increases in tree height.
Thirdly, the odds of magpies and grey magpies nesting displayed an initial decrease with increasing distance from the central lawn, reaching its lowest point at D=400 meters, and subsequently started to rise with greater distances.
Lastly, the odds of nesting decreased as tree cover increased within the range of 0–20% TC, exhibited relatively consistent fluctuations between 20–60% TC, and experienced a rapid decline beyond 60% TC.
To deepen our understanding of the landscape and environmental factors influencing breeding success in urban areas, future research should focus on quantifying nesting site preferences and success at finer scales. Specifically, it is crucial to consider environmental factors such as the size, shape, and proximity to human traffic of plant patches, as well as the contributions of parental breeding inputs. These further investigations will contribute to a more comprehensive understanding of avian nesting dynamics in urban habitats.
Further research should quantify both nest-site preference and success at finer scales, such as plant patch size, shape, isolation and distance to human traffic to better understand the landscape factors impacting nest-site selection strategies in urban areas. The examination of “urban adapter”, exemplified by magpie and gray magpie, offers a unique opportunity to explore the nesting strategies adopted by birds thriving in urban environment. Concurrently, there exist other avian species known as “urban avoider”, which have also ventured into urban areas. Investigating the nesting preferences and adaptive mechanisms of these “urban avoider” is a compelling line of research. Understanding the factors that influence these birds to thrive in urban environments can provide valuable insights into the broader dynamics of urban ecology and bird conservation. By delving into the very different strategies and adaptations of “urban pioneers” and “urban escapees”, we can gain a fuller understanding of bird responses to urbanization and contribute to the sustainable coexistence of wildlife and human communities in urban landscapes.
The original contributions presented in the study are included in the article/Supplementary material. Further inquiries can be directed to the corresponding authors.
ZD: Conceptualization, Data curation, Formal Analysis, Investigation, Software, Validation, Visualization, Writing – original draft. AG: Formal Analysis, Investigation, Methodology, Resources, Validation, Writing – original draft. ML: Data curation, Formal Analysis, Resources, Validation, Visualization, Writing – original draft. YW: Conceptualization, Investigation, Visualization, Writing – original draft. WY: Conceptualization, Investigation, Resources, Writing – original draft. HJ: Investigation, Resources, Validation, Writing – original draft. XZ: Investigation, Resources, Validation, Writing – original draft. CQ: Investigation, Resources, Validation, Writing – original draft. JL: Conceptualization, Formal Analysis, Methodology, Resources, Writing – review & editing. JC: Conceptualization, Funding acquisition, Methodology, Resources, Validation, Writing – review & editing.
The authors declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by the National Natural Science Foundation of China (grant number 32071832). And the APC was funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), Postgraduate Research & Practice Innovation Program of Jiangsu Province (KYCX23_1206) and Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University.
We are deeply thankful to two reviewers for their invaluable comments that have largely improved the quality of this manuscript. We sincerely thank Chengkang Wang, Peijian Shi, Hanchong Wu, Jiaying Ye, Zewen Gu, Yifan Yang and Lukai Liu for their valuable help in the preparation of this work.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1258185/full#supplementary-material
Alberti M., Correa C., Marzluff J. M., Hendry A. P., Palkovacs E. P., Gotanda K. M., et al. (2017). Global urban signatures of phenotypic change in animal and plant populations. Proc. Natl. Acad. Sci. 114 (34), 8951–8956. doi: 10.1073/pnas.1606034114
Andren H. (1992). Corvid density and nest predation in relation to forest fragmentation: a landscape perspective. Ecology 73 (3), 794–804. doi: 10.2307/1940158
Arroyo-Solís A., Castillo J. M., Figueroa E., López-Sánchez J. L., Slabbekoorn H. (2013). Experimental evidence for an impact of anthropogenic noise on dawn chorus timing in urban birds. J. Avian. Biol. 44 (3), 288–296. doi: 10.1111/j.1600-048x.2012.05796.x
Bakermans M. H., Rodewald A. D. (2006). Scale-Dependent Habitat use of Acadian Fly Catc her (Empidonax Virescens) in Central Ohio. Auk. 123 (2), 368–382. doi: 10.1093/auk/123.2.368
Blinkova O., Shupova T. (2017). Bird communities and vegetation composition in the urban forest ecosystem: correlations and comparisons of diversity indices. Ekológia (Bratislava) 36 (4), 366–387. doi: 10.1515/eko-2017-0029
Boada M., Sánchez S. (2012). Naturaleza y cultura, biodiversidad urbana. Ecoinovaçao para melhoria ambiental produtos e serviços. Experiências espanholas e brasileiras nos setores industrial urbano e agrícola 131-142.
Bonnington C., Gaston K. J., Evans K. L. (2015). Ecological traps and behavioural adjustments of urban songbirds to fine-scale spatial variation in predator activity. Anim. Conserv. 18 (6), 529–538. doi: 10.1111/acv.12206
Burger J., Jeitner C., Jensen H., Fitzgerald M., Carlucci S., Shukla S., et al. (2004). Habitat use in basking Northern water (Nerodia sipedon) and Eastern garter (Thamnophis sirtalis) snakes in urban New Jersey. Urban Ecosyst. 7, 17–27. doi: 10.1023/b:ueco.0000020169.86700.76
Burhans D. E., Thompson F. R. (2006). Songbird abundance and parasitism differ between urban and rural shrublands. Ecol. Appl. 16 (1), 394–405. doi: 10.1890/04-0927
Camprodon J., Brotons L. (2006). Effects of undergrowth clearing on the bird communities of the Northwestern Mediterranean Coppice Holm oak forests. For. Ecol. Manage. 221 (1-3), 72–82. doi: 10.1016/j.foreco.2005.10.044
Cao J., Fu J., Fan C., Qiu B., Chen X., Yu Z., et al. (2023). Research on the construction method of urban green space habitat network based on InVEST model and Least-cost path: A case study of Nanjing. Chin. Landsc. Archit. 38 (1), 53–58. doi: 10.19775/j.cla.2023.01.0053
Carbó-Ramírez P., Zuria I. (2011). The value of small urban greenspaces for birds in a Mexican city. Landscape Urban Plan. 100 (3), 213–222. doi: 10.1016/j.landurbplan.2010.12.008
Claramunt S., Hong M., Bravo A. (2022). The effect of flight efficiency on gap-crossing ability in Amazonian forest birds. Biotropica 54 (4), 860–868. doi: 10.1111/btp.13109
Clergeau P., Croci S., Jokimäki J., Kaisanlahti-Jokimäki M. L., Dinetti M. (2006). Avifauna homogenisation by urbanisation: analysis at different European latitudes. Biol. Conserv. 127 (3), 336–344. doi: 10.1016/j.biocon.2005.06.035
Deng W., Gao W., Song X. (2001). Effect of forest patch size and isolation on reproductive success of Great Tit in fragmented secondary-forests. Chin. J. Appl. Ecol. 04), 527–531.
Ding P., Feeley K. H.. (2010). Patterns of species diversity and functional diversity of breeding birds in Hangzhou across an urbanization gradient. Avian Res. 1(1), 1–8. doi: 10.5122/cbirds.2009.0020
Ding Z., Cao J., Wang Y. (2023). The construction and optimization of habitat networks for urban–natural symbiosis: a case study of the main urban area of nanjing. Forests 14 (1), 133. doi: 10.3390/f14010133
Dominoni D., Quetting M., Partecke J. (2013). Artificial light at night advances avian reproductive physiology. P. R. Soc B-Biol. Sci. 280 (1756), 20123017. doi: 10.1098/rspb.2012.3017
Falchi F., Cinzano P., Duriscoe D., Kyba C. C., Elvidge C. D., Baugh K., et al. (2016). The new world atlas of artificial night sky brightness. Sci. Adv. 2 (6), e1600377. doi: 10.1126/sciadv.1600377
Foley J. A., DeFries R., Asner G. P., Barford C., Bonan G., Carpenter S. R., et al. (2005). Global consequences of land use. Science 309 (5734), 570–574. doi: 10.1126/science.1111772
Francis R. A., Chadwick M. A. (2012). What makes a species synurbic? Appl. Geogr. 32 (2), 514–521. doi: 10.1016/j.apgeog.2011.06.013
Gao Y., Ci H., Lei X. (2014). Nest-site selection of Pica pica and Cyanopica cyana in protection forest of freeways. Ecol. Sci. 33 (05), 904–908. doi: 10.14108/j.cnki.1008-8873.2014.05.013
Gaston K. J., Bennie J., Davies T. W., Hopkins J. (2013). The ecological impacts of nighttime light pollution: a mechanistic appraisal. Biol. Rev. 88 (4), 912–927. doi: 10.1111/brv.12036
Gaston K. J., Davies Z. G., Edmondson J. L. (2010). Urban environments and ecosystem functions. Urban Ecol. 35-52. doi: 10.1017/cbo9780511778483.004
Ge Z. M., Wang T. H., Shi W. Y., Zhou L. C., Xue W. J. (2005). Impacts of environmental factors on the structure characteristics of avian community in Shanghai woodlots in spring. Zool. Res. 43 (01), 17–24.
Grimm N. B., Faeth S. H., Golubiewski N. E., Redman C. L., Wu J., Bai X., et al. (2008). Global change and the ecology of cities. Science 319 (5864), 756–760. doi: 10.2307/20053310
Hafner H., Fasola M. (1992). The relationship between feeding habitat and colonially nesting Ardeidae. Manag. Mediterr. wetlands their birds 20, 194–201.
Hao Y., An W., Ren J., Hao Y., Meng X., Li F. (1992). Food for magpies. Sichuan J. Zool. 1992 (02), 37–38.
Hastie T., Tibshirani R. (1987). Generalized additive models: some applications. J. Am. Stat. Assoc. 82 (398), 371–386. doi: 10.1080/01621459.1987.10478440
Hölker F., Wolter C., Perkin E. K., Tockner K. (2010). Light pollution as a biodiversity threat. Trends Ecol. Evol. 25 (12), 681–682. doi: 10.1016/j.tree.2010.09.007
Huang Y., Zhao Y., Li S., von Gadow K. (2015). The effects of habitat area, vegetation structure and insect richness on breeding bird populations in Beijing urban parks. Urban For. Urban Gree. 14 (4), 1027–1039. doi: 10.1016/j.ufug.2015.09.010
Hui C. A. (2002). Concentrations of chromium, manganese, and lead in air and in avian eggs. Environ. pollut. 120 (2), 201–206. doi: 10.1016/s0269-7491(02)00158-6
Husté A., Boulinier T. (2007). Determinants of local extinction and turnover rates in urban bird communities. Ecol. Appl. 17 (1), 168–180. doi: 10.2307/40061985
Ibáñez-Álamo J. D., Soler M. (2010). Does urbanization affect selective pressures and life-history strategies in the common blackbird (Turdus merula L.)? Biol. J. Linn. Soc 101 (4), 759–766. doi: 10.1111/j.1095-8312.2010.01543.x
James F. C., Wamer N. O. (1982). Relationships between temperate forest bird communities and vegetation structure. Ecology 63 (1), 159–171. doi: 10.2307/1937041
Job J. R., Kohler S. L., Gill S. A. (2016). Song adjustments by an open habitat bird to anthropogenic noise, urban structure, and vegetation. Behav. Ecol. 27 (6), 1734–1744. doi: 10.1093/beheco/arw105
Johnson M. T., Munshi-South J. (2017). Evolution of life in urban environments. Science 358 (6363), eaam8327. doi: 10.1126/science.aam8327
Jokimäki J., Huhta E. (2000). Artificial nest predation and abundance of birds along an urban gradient. Condor. 102 (4), 838–847. doi: 10.2307/1370311
Jokimäki J., Suhonen J. (1993). Effects of urbanization on the breeding bird species richness in Finland: a biogeographical comparison. Ornis. Fennica. 70, 71–71.
Kaplan G. (2019). Australian magpie: biology and behaviour of an unusual songbird (Australia: CSIRO publishing).
Kight C. R., Swaddle J. P. (2007). Associations of anthropogenic activity and disturbance with fitness metrics of eastern bluebirds (sialia sialis). Biol. Conserv. 138, 189–197. doi: 10.1016/j.biocon.2007.04.014
Kivelä S. M., Seppänen J. T., Ovaskainen O., Doligez B., Gustafsson L., Mönkkönen M., et al. (2014). The past and the present in decision-making: the use of conspecific and heterospecific cues in nest site selection. Ecology 95 (12), 3428–3439. doi: 10.1890/13-2103.1
Lack D. (1969). The numbers of bird species on islands. Bird stud. 16 (4), 193–209. doi: 10.1080/00063656909476244
Liu Y., Li Y., Jia Q., Zhang Y., Wang Y. (2019). Magpie Nest-Site Selection in Campus of Universities in Xinxiang City, Henan Province. Chin. J. Wildl. 40 (01), 103–108. doi: 10.19711/j.cnki.issn2310-1490.2019.01.015
Lowry J. H., Baker M. E., Ramsey R. D. (2012). Determinants of urban tree canopy in residential neighborhoods: household characteristics, urban form, and the geophysical landscape. Urban Ecosyst. 15, 247–266. doi: 10.1007/s11252-011-0185-4
Luniak M. (2004). “Synurbization–adaptation of animal wildlife to urban development,” in Proceedings 4th international urban wildlife symposium, Arizona, USA: Tucson, University of Arizona Press. 50–55.
Luniak M., Mulsow R. (1988). Ecological parameters in urbanization of the European Blackbird. Acta Congr. Int. Orn. 19, 1787–1793.
MacGregor-Fors I., Schondube J. E. (2011). Gray vs. green urbanization: relative importance of urban features for urban bird communities. Bas. Appl. Ecol. 12 (4), 372–381. doi: 10.1016/j.baae.2011.04.003
Magre J. M., Juncà M. B., Campanera J. M., Pagès A. B., Mallén I. R., Zaragoza R. M., et al. (2019). How urban green management is influencing passerine birds’ nesting in the Mediterranean: A case study in a Catalan city. Urban For. Urban Gree. 41, 221–229. doi: 10.1016/j.ufug.2019.03.012
Martin T. E. (1993). Nest predation and nest sites. BioScience 43 (8), 523–532. doi: 10.2307/1311947
Marzluff J., Rodewald A. (2008). Conserving biodiversity in urbanizing areas: nontraditional views from a bird’s perspective. Cities Environ. (CATE) 1 (2), 6. doi: 10.15365/cate.1262008
Matthews A., Dickman C. R., Major R. E. (1999). The influence of fragment size and edge on nest predation in urban bushland. Ecography 22 (4), 349–356. doi: 10.2307/3683227
McKinney M. L. (2006). Urbanization as a major cause of biotic homogenization. Biol. Conserv. 127 (3), 247–260. doi: 10.1016/j.biocon.2005.09.005
Meillère A., Brischoux F., Angelier F. (2015). Impact of chronic noise exposure on antipredator behavior: an experiment in breeding house sparrows. Behav. Ecol. 26 (2), 569–577. doi: 10.1093/beheco/aru232
Mella J. E., Loutit A. (2007). Ecología comunitaria y reproductiva de aves en cerros islas y parques de Santiago.
Miller S. G., Knight R. L., Miller C. K. (1998). Influence of recreational trails on breeding bird communities. Ecol. Appl. 8 (1), 162–169. doi: 10.1016/s1240-1307(98)80058-7
Munyenyembe F., Harris J., Hone J., Nix H. (1989). Determinants of bird populations in an urban area. Aust. J. Ecol. 14 (4), 549–557. doi: 10.1111/j.1442-9993.1989.tb01460.x
Palomino D., Carrascal L. M. (2006). Urban influence on birds at a regional scale: a case study with the avifauna of northern Madrid province. Landscape Urban Plan. 77 (3), 276–290. doi: 10.1016/j.landurbplan.2005.04.003
Parker T. S., Nilon C. H. (2012). Urban landscape characteristics correlated with the synurbization of wildlife. Landscape Urban Plan. 106 (4), 316–325. doi: 10.1016/j.landurbplan.2012.04.003
Partecke J., Gwinner E., Bensch S. (2006). Is urbanisation of European blackbirds (Turdus merula) associated with genetic differentiation? J. Ornithol. 147, 549–552. doi: 10.1007/s10336-006-0078-0
Partecke J., Van't Hof T., Gwinner E. (2004). Differences in the timing of reproduction between urban and forest European blackbirds (Turdus merula): result of phenotypic flexibility or genetic differences . Ser. B: Biol. Sci. 271 (1552), 1995–2001. doi: 10.1098/rspb.2004.2821
Phillips J., Nol E., Burke D., Dunford W. (2005). Impacts of housing developments on Wood Thrush nesting success in hardwood forest fragments. Condor. 107 (1), 97–106. doi: 10.2307/3247758
Reale J. A., Blair R. B. (2005). Nesting success and life-history attributes of bird communities along an urbanization gradient. Urban Habitats 3 (1), 1–24.
Rendell W. B., Robertson R. J. (1989). Nest-site characteristics, reproductive success and cavity availability for Tree Swallows breeding in natural cavities. Condor. 91 (4), 875–885. doi: 10.2307/1368072
Savard J. P. L., Clergeau P., Mennechez G. (2000). Biodiversity concepts and urban ecosystems. Landscape Urban Plan. 48 (3-4), 131–142. doi: 10.1016/s0169-2046(00)00037-2
Sepp T., McGraw K. J., Kaasik A., Giraudeau M. (2018). A review of urban impacts on avian life-history evolution: Does city living lead to slower pace of life? Global Change Biol. 24 (4), 1452–1469. doi: 10.1111/gcb.13969
Shanahan D. F., Miller C., Possingham H. P., Fuller R. A. (2011). The influence of patch area and connectivity on avian communities in urban revegetation. Biol. Conserv. 144 (2), 722–729. doi: 10.1016/j.biocon.2010.10.014
Slabbekoorn H., Ripmeester E. A. P. (2008). Birdsong and anthropogenic noise: implications and applications for conservation. Mol. Ecol. 17 (1), 72–83. doi: 10.1111/j.1365-294x.2007.03487.x
Smith-Castro J. R. (2008). Impacts of recreational trails on breeding birds in forested urban parks (State of Ohio, United States of America: The Ohio State University).
Sol D., González-Lagos C., Moreira D., Maspons J., Lapiedra O. (2014). Urbanisation tolerance and the loss of avian diversity. Ecol. Lett. 17 (8), 942–950. doi: 10.1111/ele.12297
Sol D., Lapiedra O., González-Lagos C. (2013). Behavioural adjustments for a life in the city. Anim. Behav. 85 (5), 1101–1112. doi: 10.1016/j.anbehav.2013.01.023
Soulé M. E., Bolger D. T., Alberts A. C., Wrights J., Sorice M., Hill S. (1988). Reconstructed dynamics of rapid extinctions of chaparral-requiring birds in urban habitat islands. Conserv. Biol. 2 (1), 75–92. doi: 10.2307/2386274
Stracey C. M. (2011). Resolving the urban nest predator paradox: the role of alternative foods for nest predators. Biol. Conserv. 144 (5), 1545–1552. doi: 10.1016/j.biocon.2011.01.022
Wang Y., Chen S., Blair R. B., Jiang P., Ding P. (2009). Nest composition adjustments by Chinese Bulbuls Pycnonotus sinensis in an urbanized landscape of Hangzhou (E China). Acta Ornithol. 44 (2), 185–192. doi: 10.3161/000164509x482768
Wang Y., Huang Q., Lan S., Zhang Q., Chen S. (2015). Common blackbirds Turdus merula use anthropogenic structures as nesting sites in an urbanized landscape. Curr. Zool. 61 (3), 435–443. doi: 10.1093/czoolo/61.3.435
Wang Q., Tang Y. F., Wang G. B. (2016). A preliminary investigation on forest composition and structure in campus of urban forest. J. NJFU 49 (01), 109. doi: 10.3969/j.jssn.1000-2006.2006.01.028
Weaving M. J., White J. G., Isaac B., Rendall A. R., Cooke R. (2016). Adaptation to urban environments promotes high reproductive success in the tawny frogmouth (Podargus strigoides), an endemic nocturnal bird species. Landscape Urban Plan. 150, 87–95. doi: 10.1016/j.landurbplan.2016.03.001
Wood S. N. (2004). Stable and efficient multiple smoothing parameter estimation for generalized additive models. J. Am. Stat. Assoc. 99 (467), 673–686. doi: 10.1198/016214504000000980
Wysocki D. (2004). Alternative mating strategies in the urban population of the European Blackbird Turdus merula in Szczecin(NW Poland). Ardea 92 (1), 103–112.
Xu X. J., Ge Z. M., Pei E. L., Shi W. Y., Wang Z. H., Wang T. H. (2007). Avian diversity and its affecting factors in Shanghai Expo’s site and surrounding areas. Chin. J. Ecol. 26, 1954–1958.
Yang G., Xu J., Wang Y., Wang X., Pei E., Yuan X., et al. (2015). Evaluation of microhabitats for wild birds in a Shanghai urban area park. Urban For. Urban Gree. 14 (2), 246–254. doi: 10.1016/j.ufug.2015.02.005
Zhang Z., Huang G. (2018). Urban ornithological research in China: A review. Acta Ecologica. Sinica. 38 (10), 3357–3367.
Keywords: landscape factors, magpie and gray magpie, nesting site selection, tree height, urban habitats
Citation: Ding Z, Guo A, Lian M, Wang Y, Ying W, Jiang H, Zhou X, Qian C, Lai J and Cao J (2023) Landscape factors influencing bird nest site selection in urban green spaces. Front. Ecol. Evol. 11:1258185. doi: 10.3389/fevo.2023.1258185
Received: 13 July 2023; Accepted: 26 July 2023;
Published: 18 August 2023.
Edited by:
Jie Gao, Xinjiang Normal University, ChinaReviewed by:
Wenchen Song, Minzu University of China, ChinaCopyright © 2023 Ding, Guo, Lian, Wang, Ying, Jiang, Zhou, Qian, Lai and Cao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiangshan Lai, bGFpQG5qZnUuZWR1LmNu; Jiajie Cao, Y2FvamlhamllQG5qZnUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.