
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Ecol. Evol., 10 October 2023
Sec. Conservation and Restoration Ecology
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1214176
This article is part of the Research TopicHuman Impacts on Bats in Tropical Ecosystems: Sustainable Actions and AlternativesView all 10 articles
 Bruna Silva Xavier1*
Bruna Silva Xavier1* Ana Rainho2
Ana Rainho2 Ana M. C. Santos3,4
Ana M. C. Santos3,4 Marcus Vinícius Vieira1
Marcus Vinícius Vieira1 William Douglas Carvalho3,4,5
William Douglas Carvalho3,4,5Introduction: The conversion of natural habitats to agricultural systems is one of the main global threats to bats. Here, we aimed to develop a systematic mapping to identify publication trends and research gaps in studying bats and agricultural systems.
Methods: We reviewed 309 studies published between 1990 and 2021 that sampled bats in agricultural systems or evaluated the effect of these systems on these animals.
Results: We found that most studies were conducted in the Palearctic and Neotropical regions (55.3%) and forest biomes (66.0%). Grassland-cropland systems (50.2%) and forest plantations that do not require cutting during the extraction of their products (47.9%) were more studied than forest plantations that require cutting (19.7%). Additionally, acoustic recordings (41.1%) and mist nets (34.3%) were the primary sampling methods used, with few studies combining these methods (7.1%). Also, most studies were conducted on a local scale (77.7%). The number of landscape-scale studies was smaller (34.3%) and concentrated in the Palearctic region (39.6%). Most studies assessed how agricultural systems affect biodiversity (62.1%). However, the phylogenetic and functional dimensions and b-diversity were little explored, with 2.5% and 23.3% of the biodiversity studies, respectively. Of the proposed mitigation measures, the most cited was including natural/semi-natural/potential bat habitats in cultivated landscapes (59.5%).
Discussion: In summary, our findings highlight the need for attention to the Afrotropic and Indo-Malaysia regions; predominantly non-forested biomes; plantations that require cutting during the extraction of their products; combined use of different sampling methods, as well as other methods as telemetry; use of multiple biodiversity descriptors and others biological descriptors, such as ecological services; landscape-scale studies and the role of conservation policies in promoting their conservation and raising awareness of their importance among producers and local communities. Filling these knowledge gaps is necessary to understand the factors influencing bat survival in cultivated landscapes. This is the only way to develop management and conservation strategies in these landscapes.
Agricultural systems have already replaced millions of hectares of native vegetation in the world (Food and Agriculture Organization, 2020a; Potapov et al., 2022). This conversion changed the structure (e.g., canopy cover) and composition (e.g., plant species present) of local vegetation, and transformed the landscape into a patchy mosaic where the original habitat has become fragmented or reduced (Wiegand et al., 2005; Fischer and Lindenmayer, 2007; May et al., 2019). These changes decrease the availability of shelter and food resources, thus affecting animals’ presence, abundance, and behavior (Gibson et al., 2011; Tuck et al., 2014; Newbold et al., 2020; Outhwaite et al., 2022). For bats, for example, the conversion of natural habitats to agricultural systems represent one of the main global threats to their conservation (Frick et al., 2019).
Bats provide several ecological services which are beneficial in both natural and anthropogenic habitats (Boyles et al., 2013; Castillo-Figueroa, 2020; Regolin et al., 2020). Some of these services are directly related to the productivity of agricultural systems (e.g., pollination, pest control), while others are related with the restoration of degraded habitats (e.g., seed dispersal) that appear as a consequence of the establishment of plantations, grasslands or croplands (Frick et al., 2019; Castillo-Figueroa, 2020). Previous studies show that bats’ responses to habitat conversion can vary according to the type of the original habitat replaced (Carballo-Morales et al., 2021), type of agricultural systems that replaces it (Law et al., 2016; Farneda et al., 2020; Carballo-Morales et al., 2021) and the intensity of management (Park, 2015; Williams-Guillén et al., 2016). Furthermore, bat responses are influenced by their traits such as their diet, wing shape, body mass, and echolocation type (García-Morales et al., 2013; Farneda et al., 2020; Loeb, 2020; Mendes and Srbek-Araujo, 2021). Due to this variability, it is necessary to understand the consequences of replacing natural environments with agricultural systems, filling existing knowledge gaps. This represents a crucial initial step in the development of effective mitigation strategies to ensure the preservation of the ecosystem services provided by these animals (Aizpurua et al., 2018; Ongole et al., 2018; Costa et al., 2020; Charbonnier et al., 2021).
Bat responses to the presence of plantation or the conversion of their habitats to agricultural systems have been partially reviewed in previous studies (e.g., Park, 2015; Williams-Guillén et al., 2016; Frick et al., 2019). However, most of these reviews are focused on a particular group of bats (e.g., Aziz et al., 2016; Law et al., 2016; Carballo-Morales et al., 2021), country or geographical region (e.g., Maas et al., 2016; Meyer et al., 2016; Farneda et al., 2020; Carballo-Morales et al., 2021). Therefore, such studies do not provide a clear overall picture of current research publication trends and potential knowledge gaps, nor identify general future research venues. Here, we employ a systematic mapping approach which aims to describe the publication trends and find potential gaps in existing literature, allowing to identify aspects of a particular research question that are still missing additional empirical research (James et al., 2016). Particularly, we focus on the research performed on bats and agricultural systems, evaluating the geographic regions that have been studied, the methods used, the research objectives, and the proposed mitigation strategies for.
We systematically reviewed in three databases (Google Scholar, Web of Science and SCOPUS). Data collection was carried out in June 2020 and updated in August 2021. We did not add year of publication restriction to searches. We search for the following keywords and their variations in all studies fields: “bat”, “Chiroptera”, “farming”, “farm”, “farmland”, “plantation”, “planting”, “sylviculture”, “silviculture”, “agriculture”, “forestry”, “crop”, “agroforestry” (Supplementary Table 1). This initial search resulted in 16,842 records. We screened the title and abstract of each study, and retained those studies that followed these eligibility criteria: 1) were written in English; 2) were peer-reviewed scientific articles; and 3) included sampling of bats in agricultural systems or considered the effect of agricultural systems on bats. Here we define agricultural systems as areas cultivated by humans for the purpose of commercial production and subsequent sale and which encompass areas cultivated with crops, forestry, groves, and grassland (for more details see Curveira-Santos et al., 2021). Thus, we also included cultivated pastures (grassland) used locally to feed livestock, analyzing this type of agriculture system together with croplands (hereinafter simply grassland-cropland system), since both systems do not present vertical/arboreal strata. Finally, review studies and studies that mixed the effects of the agricultural system considered by us with other land uses, so that it was not possible to know which disturbance the bats were responding to, or mixed the response of bats with other organisms, so that it was not possible to know what was the response of the bats to the analyzed variables, were excluded. In a second stage, we excluded duplicates and assessed the eligibility of studies by screening the entire text, using the same criteria described above. The flowchart in Figure 1 illustrates the procedure applied for article screening, that was built according to the PRISMA (Preferred Reported Items for Systematic Reviews and Meta-Analyses statement; Moher et al., 2009). After following these steps, we retained a total of 309 publications, which are listed in Supplementary Table 2.
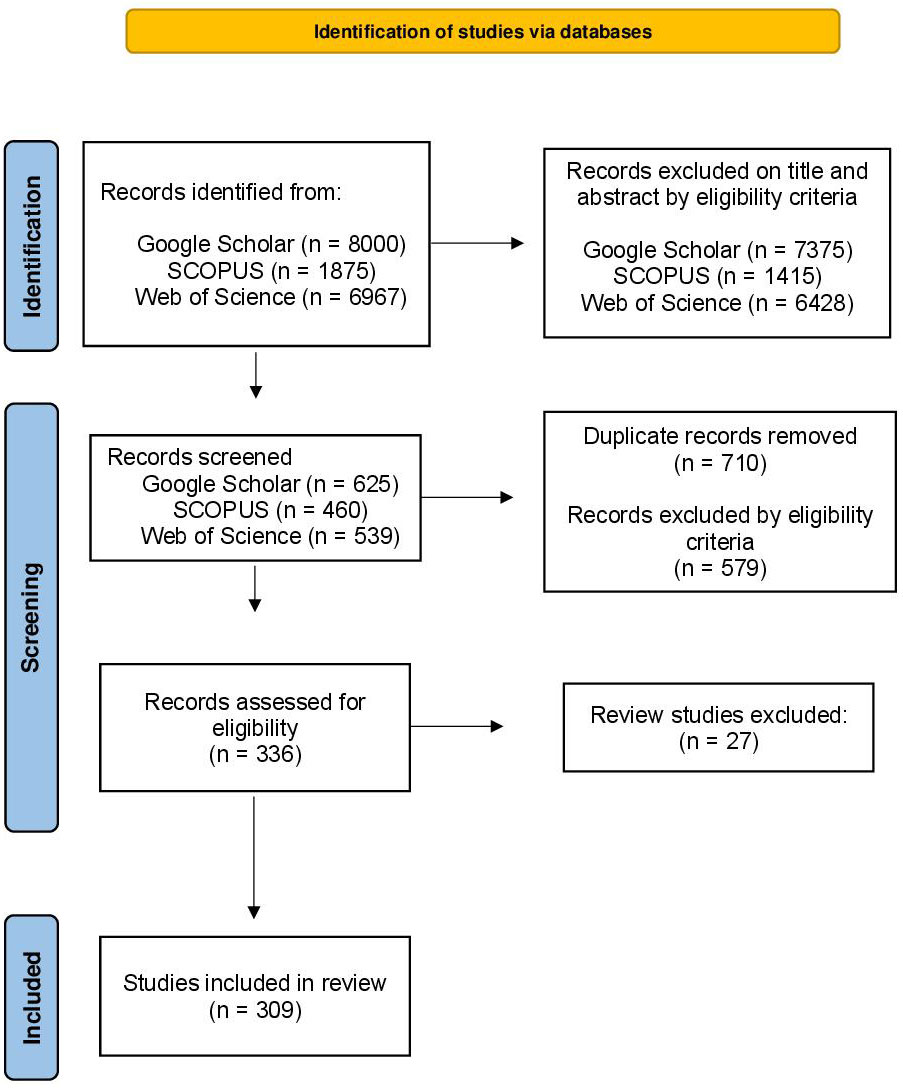
Figure 1 PRISMA flowchart of the systematic review of studies focusing on bats that were sampled in agricultural systems or that evaluate the effect of these agricultural systems on bats.
We extracted the following information from the selected studies: 1) publication year; 2) region where the study was conducted, namely: Australasia, Afrotropic, Indo-Malaysia, Nearctic, Neotropics, and Palearctic (following the Ecoregions2017©Resolve, available in ecoregions2017.appspot.com); 3) structure type of the original biome, as defined by Dinerstein et al. (2017): predominantly forested, and predominantly non-forested; 4) type of agricultural systems sampled: grassland-cropland (agricultural systems that do not present vertical/arboreal strata; i.e., are herbaceous or shrubby), forest plantation that require cutting during the extraction of its products (tree-dominated plantations where product extraction leads to the cutting of trees), forest plantation that does not require cutting during the extraction of its products (tree-dominated plantations where product extraction does not lead to the cutting of trees); 5) focal taxa (species or group of bats studied); 6) sampling method; 7) spatial scale of the study (local: studies conducted within of the agricultural systems and studies comparing habitats; landscape: studies related to an area and studies comparing landscapes); 8) predictors used to assess the effect of agricultural systems (e.g., comparison between types of agricultural systems, comparison between agricultural systems and natural/semi-natural area, use of agrochemicals, percentage of cultivated area in the landscape); 9) biological descriptors used in the study (e.g., biodiversity, ecological services, mortality); 10) biodiversity descriptor of the groups of bats studied (e.g., species richness, taxonomic diversity, total abundance) and; 11) proposals for mitigating the impact of agricultural systems on bats. For more details see the Supplementary Table 3. We quantified studies (n) in each of these categories; however, when a study fell into more than one category, it was counted in all categories to which it belonged. Also, in some situations we refer to cases, which were different situations evaluated within the studies. For example, the same study that compares abundance and richness between two different types of agricultural systems and between agricultural systems and other land uses, has four cases (one comparing abundance between different types of agricultural systems, one comparing abundance between agricultural systems and another land use, one comparing richness between different types of agricultural systems, and lastly, one comparing richness between agricultural system and another land use). Likewise, four different studies comparing abundance between different types of agricultural systems also constituted four different cases.
The systematic review resulted in 309 studies (Figures 1, 2; Supplementary Table 2) that either used empirical data of bat collected from agricultural systems or that considered the effect of these agricultural systems on bats. These studies were published between 1990 and 2021 and showed an increasing publication rate over the years (r² = 0.78; p<0.001; Figure 2). However, our data revealed an unequal geographic distribution of knowledge across the globe (Figure 3A). The Palearctic (n = 89) and Neotropical (n = 82) regions concentrated most of the studies (55.3% of total), followed by Indo-Malaysia (n = 43, 13.9%), Nearctic (n = 38, 12.3%), Afrotropic (n = 35, 11.3%), and Australasia (n = 22, 7.1%; Figure 3A). Each region has unique characteristics, such as climate, species diversity, and economic development, which can impact how bat communities are affected by agricultural systems (Williams-Guillén et al., 2016; Frick et al., 2019; Outhwaite et al., 2022). For example, in cases comparing abundance, richness, and taxonomic diversity between natural/semi-natural habitats and agricultural systems or assessing the effect of the presence/amount of the cultivated area on these descriptors, most communities and individuals across all regions did not respond to the presence/amount of the agricultural systems (66.3% of 1779 cases). However, cases that respond significantly revealing that communities and individuals were negatively affected by the presence of agricultural systems, are best represented in the Australasia, Indo-Malaysia, Nearctic and Palearctic regions (88.5% of 61 cases, 79.6% of 49 cases, 58.6% of 29 cases, and 52.4% of 206 cases, respectively). In the Afrotropics and Neotropics, most studies show positive responses to the agriculture systems (81.4% of 43 cases and 58.3% of 211 cases, respectively). This means that knowledge acquired in one region may not apply to other areas, so is it is essential to study all regions. However, some regions (Neotropics, Afrotropic and Indo-Malaysia) are considered a priority for bat conservation due to their high bat diversity and prevalence of endemic, threatened, or poorly studied species (Frick et al., 2019). Despite this, our analysis found that only the Neotropics received significant attention. Therefore, it is crucial to increase efforts to understand the impact of agricultural systems on bat populations in the less studied regions of Afrotropic and Indo-Malaysia.
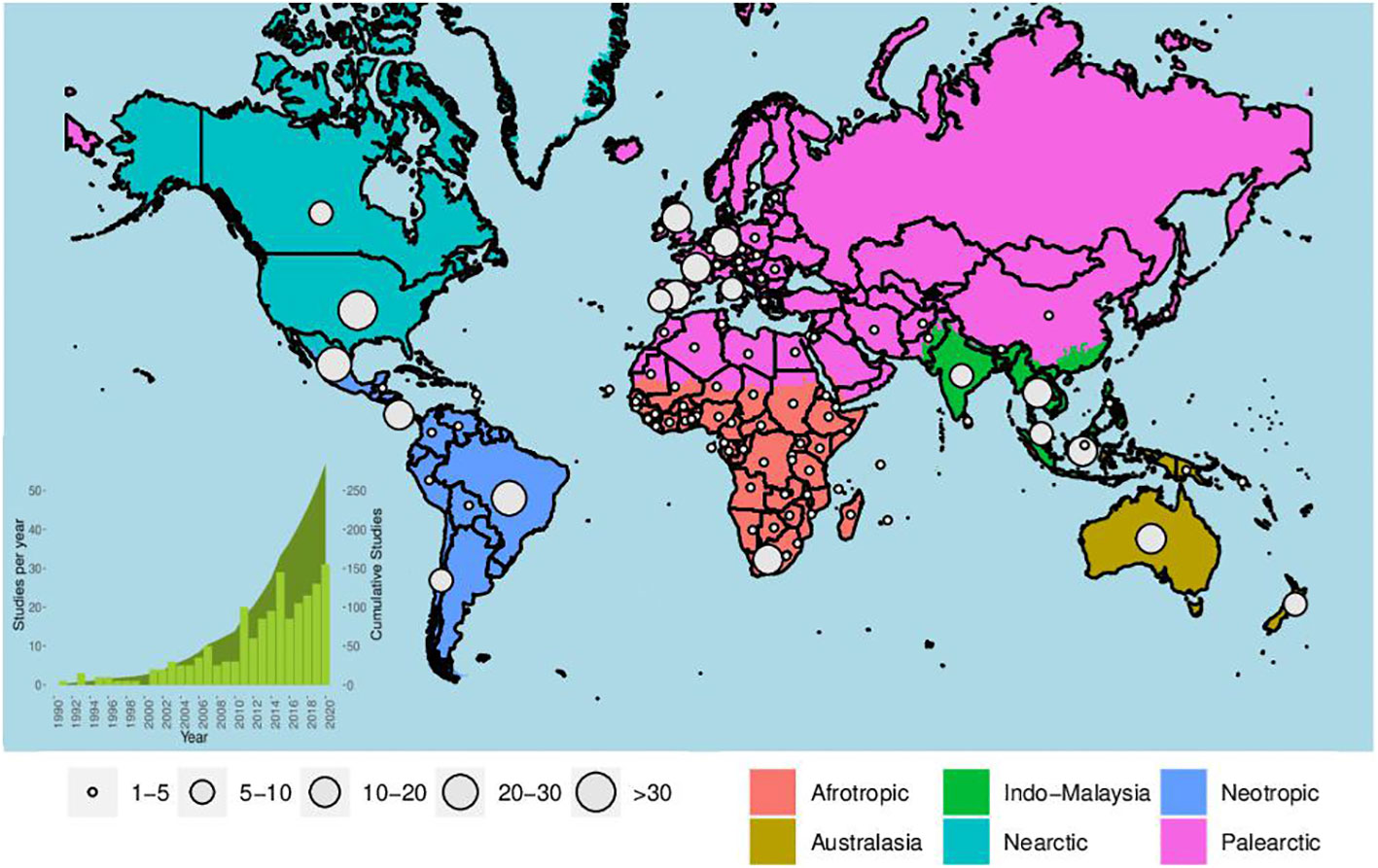
Figure 2 Worldwide distribution of studies published in which bats were sampled in agricultural systems or that assess the effect of these agricultural systems on bats. The circles on the map indicate the number of studies per country and the colors demarcate the global regions, according to Ecoregions2017©Resolve (available in ecoregions2017.appspot.com). The green bars in the graph (lower left side of the map) represent the number of studies published per year and the curve shaded in green represents the cumulative number of published studies.
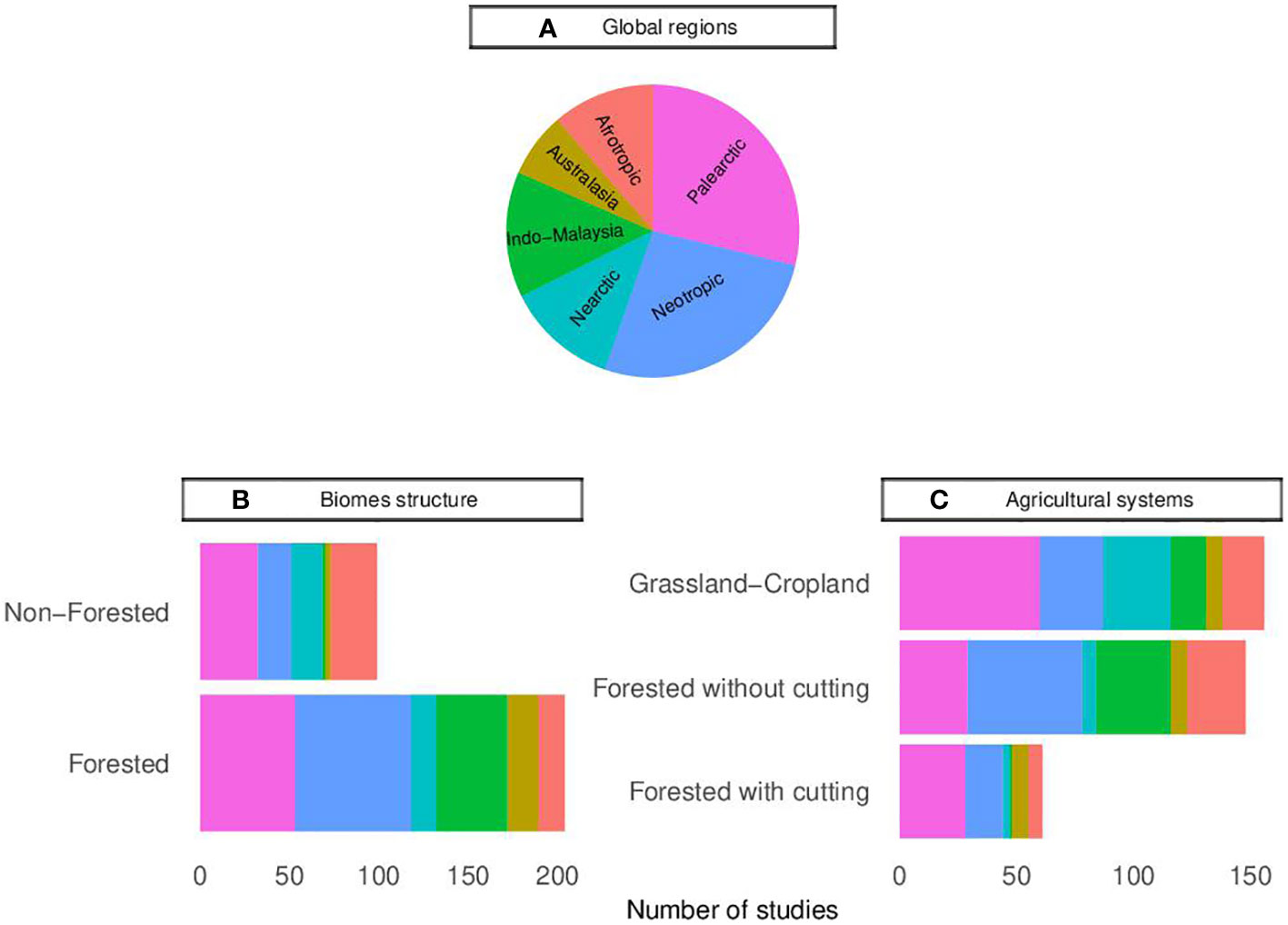
Figure 3 Distribution of studies published worldwide in which bats were sampled in agricultural systems or that evaluate the effect of these agricultural systems on bats, according to the (A) global regions, (B) type of structure of the original biomes, and (C) types of agricultural systems.
Most studies (n = 204, 66.0%) were carried out in predominantly forested biomes, while only 31.7% (n = 98) were carried out in predominantly non-forested biomes (Figure 3B, Supplementary Table 3). Additionally, among the 128 studies that examined the impact of the presence of natural/semi-natural habitats on bats in agricultural systems or compared bats in agricultural systems with those in natural/semi-natural habitats, only 27 studies investigated non-forested habitats (e.g., flooded and natural fields), while 118 studies focused on forested habitats. Bats are often forest dependent organisms (Meyer et al., 2016; Williams-Guillén et al., 2016; Mendes and Srbek-Araujo, 2021), which may explain the bias in the type of habitats and biomes that have been studied more often. However, the presence of non-forested habitats in the landscape can also influence bat communities, acting as complementary habitats for resource and movement (Lentini et al., 2012; Weier et al., 2018; Muñoz et al., 2019; Carvalho et al., 2021). Indeed, of the 27 studies that considered non-forested natural/semi-natural habitats, 20 showed the effect of these natural areas on bats present in agricultural ecosystems, with 13 of these indicate positive effects (e.g., Taylor et al., 2011; Rodríguez-San Pedro et al., 2019; Carvalho et al., 2020). Replacing non-forested habitats with agricultural systems can change the dynamics of these landscapes, which in turn might have a negative on bat diversity and activity (see Weier et al., 2018; Muñoz et al., 2019; Rodríguez-San Pedro et al., 2019; Carvalho et al., 2020). Thus, further studies investigating the replacement of these habitats with agricultural systems are needed.
Not all types of agricultural systems have been equally studied (Figure 3C; Supplementary Table 3). Grassland-cropland systems (e.g., soybean, rice, cultivated pasture) have been studied more often than others (n = 155, 50.2%). Most of the studies on this type of agricultural system took place in vineyards (n = 25, 16.1%), corn fields (n = 22, 14.2%) and rice (n = 21, 13.5%). Globally, cultivated areas are mainly represented by this type of agricultural systems (Food and Agriculture Organization, 2020a), which may explain our result. However, despite forest plantations representing a small percentage of the world’s cultivated area, the number of studies conducted in forest plantations that do not require cutting (e.g., agroforestry, orchards, oil palm; n = 148, 47.9%) is comparable to the number of studies in grassland-cropland systems. Most of the studies on this type of agricultural system took place in banana plantations (n = 32, 21.6%), shaded cafe (n = 27, 18.2%) and cacao (n = 22, 14.9%). Forest plantations may represent permeable matrices for bats or even work as complementary habitat for these organisms, while grassland-cropland systems may represent the opposite (Kunz and Lumsden, 2003; Law et al., 2016; Meyer et al., 2016; Williams-Guillén et al., 2016). Of the cases that compared abundance, richness, and taxonomic diversity between natural/semi-natural habitats and agricultural systems, most of those who responded significantly found negative effects of grassland-cropland systems and forest plantations on these descriptors. However, the percentage of studies that responded negatively to grassland-cropland systems (81.1% of 106 cases) is higher than those that responded to forest plantations (57.3% of 293 cases). In addition, even within forest plantations, the differences between plantations that require cutting for the extraction of their products (e.g., acacia, eucalyptus, heart of palm) and those that do not require cutting can be relevant. This occurs because in the latter case, the forest structure is generally maintained all the time, while in plantations that require cutting, mainly in a short-time rotation system, the matrix or habitat can change from a permeable matrix and/or suitable habitat to a matrix that can even be very limiting to bat movements (Stephens and Wagner, 2007; Zhang and Stanturf, 2008; Law et al., 2016). We also found that negative relationships between abundance, richness, or taxonomic diversity were reported more often in forest plantations that require cutting (87.9% of 78 cases) than on those that do not require cutting (48.5% of 204 cases). However, forest plantations that require cutting was the least studied (n = 61, 19.7%), with most studies focusing on pine (n = 29, 47.5%) and eucalyptus (n = 23, 37.7%) plantations. With the representation of these plantations in the world’s cultivated area increasing in recent years (Food and Agriculture Organization, 2020b), there is a growing need for a better understanding of the potential impact this type of agricultural systems can have on bats.
The Neotropical region had the highest number of studies in predominantly forest biomes (n = 65, 31.9%; Supplementary Table 3) and in forest plantations that do not require cutting (n = 48, 33.1%; Supplementary Table 3) when compared to other global regions. This region has a larger area covered originally by forested biomes than by non-forested biomes (Ecoregions2017©Resolve; ecoregions2017.appspot.com), which may explain this difference. Furthermore, it has already been found in other reviews (e.g., Meyer et al., 2016; Williams-Guillén et al., 2016), that the Neotropics have many studies on agroforestry, included here as forest plantations that do not require cutting, which may be driving this pattern. Agroforestry, which can maintain the structural complexity of native forests, has been found to mitigate the negative effects of agricultural systems (Williams-Guillén et al., 2016). Therefore, it is important to conduct studies on this type of plantation in other regions as well. The Palearctic leads in studies in predominantly non-forest biomes (n = 32, 32.7%; Supplementary Table 3), in grassland-cropland systems (n = 57, 37.3%; Supplementary Table 3) and forest plantations that require cutting (n = 28, 43.8%; Supplementary Table 3) compared to other global regions. In addition to being the region with more studies, the Palearctic has the largest area of originally non-forested biome (Ecoregions2017©Resolve; ecoregions2017.appspot.com), and of grassland-cropland cultivated areas (Food and Agriculture Organization, 2020a).
Twenty data collection methods were used in the studies reviewed (Figure 4; Supplementary Table 3). Most studies sampled bats with either acoustic recorders (n = 127, 41.1%) or mist nets (n = 106, 34.3%). Acoustics were mainly used in the Palearctic region (n = 61, 48.0%) while mist nets were used mainly in the Neotropics (n = 55, 51.9%). Using exclusively one of these methods can lead to sampling bias, as acoustic recordings perform better at recording insectivorous bats that echolocate, while mist nets are a more effective method for capturing bats that are unable to avoid interception traps (MacSwiney-G et al., 2008; Yoh et al., 2020; Appel et al., 2021; Carvalho et al., 2023). Ultimately, this can lead to an incorrect interpretation of the results found. For example, Heer et al. (2015) compared forest fragments with rubber-cacao plantations and found that the species richness recorded with acoustics did not differ between these habitats, while the number of species was higher in forest fragments when considering only the species sampled with mist nets. Thus, an approach combining several sampling methods, such as mist nets and acoustic recorders, would be ideal to reduce the risk of sampling bias for certain bat groups and, thus, gain a better understanding of their response to replacement of natural areas with agricultural systems (MacSwiney-G et al., 2008; Appel et al., 2021; Carvalho et al., 2023). Another option would be to combine mist nets with harp-traps or with captures in roost (e.g., Flaquer et al., 2007; Pech-Canche et al., 2011; Xavier et al., 2018; Appel et al., 2021). Among the selected studies, 33 (10.7%) combine various bat sampling methods, with 24 of them using one of the aforementioned combinations.
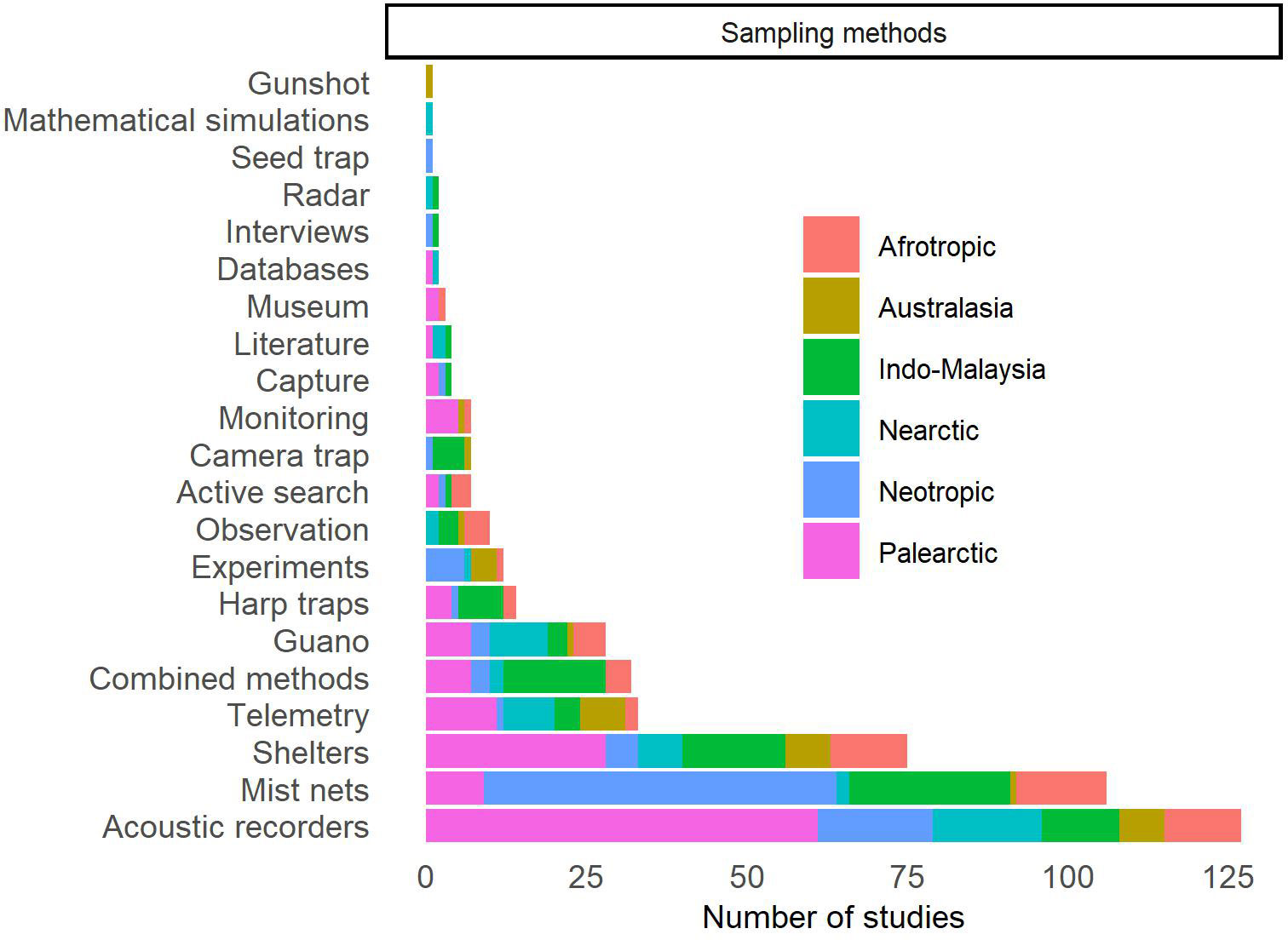
Figure 4 Distribution of studies published worldwide in which bats were sampled in agricultural systems or that evaluate the effect of these agricultural systems on bats, according to the sampling methods.
Bat roosts were used in 75 studies (24.3%), of which 27 were intended to study the roosts themselves and 48 studies used roosts to capture bats or collect information about them (e.g., ectoparasites, guano for food diet studies). Roost captures facilitate sampling of high-flying bats, that are not often caught in interception traps (e.g., mist nets, harp traps). This is very important to enrich acoustic databases that can help understand the consequences of agricultural systems on bats (Taylor et al., 2013). The fourth most used method was telemetry (n = 33, 10.7%). Telemetry is crucial for understanding bat behavior and ecology, including their habitat selection and home range (Clerc et al., 2021). New technologies that make telemetry less costly and more suitable for different bat species may increase its general use in research (O’Mara et al., 2014). The fifth most used method was the collection of guano of bats (n = 28, 9.1%), a method mainly associated with foraging studies (diet composition, diversity, richness and selection) and ecological services and disservices. Harp traps were used only in 14 studies (4.5%). Studies that used an experimental approach (n = 12, 3.9%) had the main objective of evaluating ecological services. Capture by gunshot was mentioned in one study; however, it is important to note that this approach raises ethical concerns, and we strongly advise against its use. Other methods, such as seed traps, camera traps, mathematical simulations, and interviews, were also rarely used. Studies based on information obtained from databases, literature and museum collections were used to complement the collected data or for macroecological studies.
Certain bat groups (e.g., guilds and taxa) and species may have different responses to landscape changes (Farneda et al., 2020; Loeb, 2020; Carballo-Morales et al., 2021; Mendes and Srbek-Araujo, 2021). Thus, the choice of bat group will influence the results of the studies, being important consider the communities and subgroups or functional characteristics of these communities when evaluating the effects of agricultural systems on bats to plan conservation strategies in cultivated areas. Most studies are described as being developed at the community level (n = 167, 54.0%; Supplementary Table 3). However, although these studies claim to assess the whole community, some of these focused on a particular taxon. This is mainly due to the methods used which are often selective and bias sampling towards some species. Some of these studies (n = 79), for example, are focused on insectivores by using acoustic recording as the only methods sampling, including in the Neotropics where insectivorous bats are not predominant.
Single species are the second most common focal taxa studied (n = 157, 50.8%). This type of study tend to focus on behavior, hunting, abundance, contamination, populations, ecological services and disservices and, in nine of these studies, a single species was used to represent a guild (based on body mass and use of space). The third most common focal taxa was the trophic guilds (n = 66, 21.4%), with studies of this type being conducted mainly in the Neotropics (n = 30, 45.5%). Diet or trophic level are recurrently evaluated in the Neotropics, as bats have a diverse diet in this region (Kalko et al., 1996; Kalko, 1998; Kalka et al., 2008; Kunz et al., 2011). In addition, diet reflects other important characteristics in bats, such as taxonomic group, morphological characteristics and type of echolocation (Jones et al., 2013; Castillo-Figueroa and Pérez-Torres, 2021; Potter et al., 2021). Other types of guilds were also studied (n = 36, 10.7%), mainly those based on the use of space (bats classified as open, closed or edge foragers; n = 20). In addition, six studies classified bats by their foraging strategy (aerial or gleaners), two by echolocation parameters, four by specialization in habitat use (foraging, general use and roost), two differentiated bats as rare and dominant, two used size and body mass and one used maneuverability.
Most studies were carried out on a local scale, comparing two distinct habitats (n = 135, 43.7%; Supplementary Table 3). The analyzes carried out on this scale show significant responses in 39.4% of cases. The second most common type was the studies on a local scale carried out only within of some agricultural systems (n = 128, 41.4%), with the analyzes made in this scale presenting significant responses in 36.4% of the cases. Landscape studies relating to an area were the third most common (n = 94, 30.4%), with few analyzes made on this scale showing significant responses (14.1%). Lastly, a few studies compared landscapes (n = 14, 4.5%) and found significant responses in 41.8% of cases. The scale at which bat populations are being studied is crucial, since some variables may affect bats on one scale but not on another. For example, Pina et al. (2013) found that bat composition changes with the amount of eucalyptus plantation in the landscape, but it is similar when locally comparing eucalyptus plantations with forest patches in Cerrado (Brazilian savanna) landscapes. Furthermore, bat responses on one scale may be influenced by variables acting at another scale (e.g., Herrera et al., 2016; Mendes et al., 2017; Schoeman and Monadjem, 2018; Luz et al., 2020). For example, in studies that consider the local scale and compare natural habitats with agricultural systems, bat responses to agriculture systems may vary according to the proximity to natural forest habitats or the amount of these habitats in the landscape (Park, 2015; Meyer et al., 2016). Despite this, only 38 studies (12.3%) evaluated both local and landscape scales simultaneously. Therefore, to better understand how agricultural systems affect bats, further studies are needed incorporating multiple scales of analysis, when possible.
In addition to the importance of carrying out more studies on more than one scale, using an appropriate size scale for these studies is also essential. When examining the most used scale size, specifically the circular buffer with a radius of 1000 m (842 cases), bats exhibited a significant response to the variable in question in 9.4% of cases. The other scale sizes used in more than ten studies (500, 2000, 3000, and 5000 m radius circular buffer, 422 to 486 cases) had similar percentages (from 7.1% to 13.6%). Studies that use scales size relative to the home range of bats (126 cases) are the exception, as bats responded significantly to the analyzed variables in 75.4% of cases. Based on our findings, it is plausible that the ideal size of a study area is relative to the home range of the bats under investigation. However, determining the home range of all species in a community can be challenging. Furthermore, it is essential to consider factors such as seasonality that can impact bat home range size (Meyer et al., 2016; Peixoto et al., 2018). Thus, we consider that studies using multiple scale sizes are of higher value to understand how variables in cultivated landscapes will affect bat communities (Gonthier et al., 2014). An alternative would be to compare landscapes. For example, Rodríguez-San Pedro et al. (2021) compared vineyard farms adjacent to natural habitats with vineyard farms neighboring cultivated habitats and showed that taxonomic diversity and total insectivore foraging activity were similar between these landscapes. Still, the foraging activity of Lasiurus varius, Lasiurus villosissimus, and Myotis chiloensis was higher in vineyard farms adjacent to cultivated habitats (Rodríguez-San Pedro et al., 2021). Shapiro and Bordignon (2014) found higher richness, evenness, and taxonomic diversity and lower abundance in fragmented Cerrado surrounded by small agricultural matrix fields when compared to fragmented Cerrado surrounded by urban matrix. Also, Faria et al. (2006) and Faria and Baumgarten (2007) found greater taxonomic richness of bats in shade cocoa plantations within well-preserved landscapes with a higher amount of forest compared to isolated shade cocoa plantations within less conserved landscapes. These studies aimed to compare cultivated landscapes with matrices of different cover qualities or to compare more preserved landscapes (e.g., with a higher amount of natural habitat) with less preserved landscapes. These studies constitute a qualitative way of evaluating patch and matrix contrast in the first case and the amount of land use in the second case. They may be helpful ways to assess the effects of agricultural systems on bat communities at the landscape scale.
Our analysis found a relatively even distribution of studies conducted at various scales, except for those comparing landscapes, which were fewer in number (Supplementary Table 3). However, when considering global regions, notable biases in research efforts emerged (Supplementary Table 3). The Afrotropic and Australasia regions had limited studies across all scales, highlighting the need for more comprehensive studies in these regions. The Nearctic region would benefit from additional studies incorporating reference habitats. In the Indo-Malaysia region, studies should focus on enhancing our understanding of plantation landscapes and studies incorporating reference habitats. Similarly, the Neotropics would benefit from more comprehensive studies examining the impacts of cultivated landscapes on bats.
Most studies carried out on a local scale and comparing two distinct habitats, compared agricultural systems with natural/semi-natural habitats (n = 106, 78.5%; Supplementary Table 3). These studies were mostly performed in the Neotropics (n = 50, 47.2%; Supplementary Table 3), region where the high availability of natural environments facilitate this type of comparisons (Food and Agriculture Organization, 2020b). Although the Palearctic region had the highest number of total studies, the highly urbanized and developed landscape limits the number of comparisons that can be made between natural habitats and agricultural systems (Park, 2015; Food and Agriculture Organization, 2020b). In addition to comparisons between agricultural systems and natural/semi-natural habitats, comparisons were also made between two types of agricultural systems (n = 63, 46.7%) and between other land uses (e.g., urban area, pasture) and agricultural systems (n = 29, 21.5%). Comparing different types of agricultural systems can provide valuable insights into the effects of different management practices on bats and, thus, help create strategies to balance biodiversity conservation and crop production (e.g., Barré et al., 2017; Froidevaux et al., 2017; McFadden and Dirzo, 2018; Chaiyarat et al., 2020). For example, we found that among the selected studies, there are more cases (81.1% of 106 cases) reporting significant negative effects of grassland-cropland systems on abundance, species richness, and diversity compared to forest plantations (57.3% of 293 cases). This indicates that forest plantations maintain a higher structural complexity that is less detrimental to bat communities. On the other hand, comparing different land uses can provide valuable insights into the specific anthropogenic changes that pose the greatest threat to bats, thereby enabling targeted efforts to mitigate these impacts (Frick et al., 2019). Here, we found a higher percentage (65.2% of 92 cases) of significant negative responses in terms of abundance, species richness, and diversity to agricultural systems when compared to other land uses, such as urban areas and pastures. This suggests that agricultural systems may have a more pronounced impact on bat communities, highlighting the need for specific attention to mitigate their effects.
We identified 10 different types of predictors used in studies in local scale carried out within the agricultural systems (Supplementary Table 3). Most these studies (n = 37, 28.9%) assessed structure and/or composition of the agricultural systems (e.g., basal area, canopy cover, vegetation height, presence of water) and abiotic variables (e.g., rain, seasonality, humidity) appear as the second most used type of predictors (n = 25, 19.5%). Bats were also used as predictors of ecological services and disservices (n = 25, 19.5%). The direct effect of agricultural pests on bats was also evaluated (n = 6, 4.7%), and a few studies have also evaluated how the vegetative/reproductive stage of the plantation affects bats (n = 5, 3.9%). Despite the importance of knowing how these parameters of agricultural systems affect bats, the analysis of some variables, such as structural complexity and vegetation composition, are only meaningful when compared to reference habitats (Park, 2015; Meyer et al., 2016). In addition, 34 studies (26.6%) carried out within agricultural systems were only descriptive and did not test descriptors such as biodiversity, foraging, and home range with any predictors of the agriculture systems (e.g., abiotic variables, composition of the plantation). These studies represent 23.3% (n = 10) and 21.1% (n = 8) of the studies on bats and croplands published in Indo-Malaysia and Nearctic, respectively. Thus, it is necessary, mainly in these regions, to test hypotheses and use control habitats, especially natural ones, to enhance our understanding and conservation efforts for bats in agricultural systems.
Landscape studies are of crucial importance for this highly mobile taxon, as they provide valuable insights on how the surrounding composition and configuration influence these species in their habitats (Meyer et al., 2016; Peixoto et al., 2018; Schoeman and Monadjem, 2018; Outhwaite et al., 2022). We identified 15 different predictors at this scale, most these studies have examined the presence/amount of agricultural systems on bat populations within a particular landscape (n = 79, 84.0%; Supplementary Table 3). In contrast, only 26 studies (27.7%) have examined the presence/amount of natural/semi-natural habitat, and 22 studies (23.4%) have examined the presence/amount of other land uses in the landscape (Supplementary Table 3). Therefore, even in the Palearctic, where the number of landscape studies is the largest (43.7%) compared to other regions, there is still a scarcity of research on how the presence/amount of natural/semi-natural habitats and other land uses (e.g., urban areas, roads, managed forests) in landscapes with some agricultural systems impact bat populations. We also found studies that took into consideration the landscape configuration (e.g., distance to some landscape elements, such as linear elements, water, buildings; landscape complexity; isolation/connectivity/fragmentation). These studies were scarcer (n = 27, 28.7%) and more concentrated in the Palearctic (n = 11, 40.7%). To effectively conserve bat populations in cultivated landscapes, it is crucial to comprehend how these respond to the composition and arrangement of the surrounding landscape (Park, 2015; Williams-Guillén et al., 2016; McFadden and Dirzo, 2018). However, our knowledge about bat responses to landscape variables may be region-specific and not broadly applicable to other regions. Lastly, studies that compare landscapes include comparisons (i) between some agricultural systems and natural/semi-natural area (n = 5, 35.7%) or (ii) other land uses (n = 4, 28.6%), (iii) between types of agricultural systems (n = 3, 21.4%), (iv) presence or amount of area natural/semi-natural (n = 3, 21.4%), and (v) landscape management (n = 1, 7.1%).
Predictors such as abiotic variables (n = 26, 8.4%), presence/number of insects (n = 12, 3.9%) and agricultural system management approaches (n = 47, 15.2%) were occasionally studied in more than one scale (Supplementary Table 3). Abiotic variables can affect bat responses to agricultural systems (Cisneros et al., 2015; Heim et al., 2016; Shapiro et al., 2020). In cultivated landscapes in the Neotropics, for example, bats may respond positively to forest patch size in the dry season and have no relationship with this variable in the wet season (Cisneros et al., 2015). In the Afrotropic, insectivores respond more strongly to landscape metrics in the dry season, while in the wet season they respond more strongly to local metrics (Shapiro et al., 2020). To better understand the response of bats to agricultural systems, it is important to analyse how abiotic variables interact with other factors in both the agriculture system and the surrounding landscape. Of the few studies that have evaluated the effect of insect abundance or availability on bats, all have done so at the local scale (n = 12), with only one study also evaluating it at the landscape level. Understanding food availability is of utmost importance in agricultural systems and cultivated landscapes, as it plays a vital role in determining the presence of bats in these areas. This includes investigating whether the required food resources are found within the agricultural system itself or in nearby habitats (Park, 2015; Meyer et al., 2016; Williams-Guillén et al., 2016). In addition to these studies, three others evaluated food availability as a response variable (descriptors). In all cases, the amount of fruit or plants potentially consumed by bats was estimated. Another predictor assessed at more than one scale was management. Management is essential to mitigate the impacts of agricultural systems on bats (Park, 2015; Williams-Guillén et al., 2016; McFadden and Dirzo, 2018), and can be evaluated and cultivated at different scales. For instance, both plantations with less intensive uses and landscapes that preserve potential habitats for bats while maintaining connectivity between them, hold the potential to conserve bat populations, even in the presence of agricultural systems (Park, 2015; Meyer et al., 2016). However, these studies were concentrated in the Palearctic (n = 23, 48.9%) and Neotropical (n = 15, 31.9%) regions.
The biological descriptors used in the selected studies were grouped into 22 categories (Figure 5; Supplementary Table 3). The most used descriptor was biodiversity (n = 192, 62.1%), with most studies being carried out in the Neotropical (n = 65, 33.9%) and Palearctic (n = 54, 28.1%). Of these studies that assess biodiversity, 82.8% (n = 159) assessed the biodiversity of bat communities and guilds (Figure 6), with the other studies assessing the abundance of a species. All studies that used biodiversity to describe guilds and communities explored the taxonomic dimension, while the phylogenetic and functional dimensions of biodiversity were addressed in four studies (2.5%) each. However, among the studies that use taxonomic descriptors, 35.2% (n = 56) estimated these descriptors for a guild and 5.7% (n = 9) aimed to study one or more families of Chiroptera. Thus, 135 studies (84.9%) aimed to assess the taxonomic biodiversity of all bats recorded in the studied area. The indices used to assess biodiversity are important tools for comparing habitats and, thus, assessing the effect of disturbances, such as the replacement of natural habitat by agricultural systems. However, it is important to note that these indices may only capture part of the changes occurring in communities, particularly when solely relying on taxonomic approaches (Pellens and Grandcolas, 2016; Moreno et al., 2018; Peixoto et al., 2018; Presley et al., 2018). Species will play different ecological functions in a habitat, but when there is some redundancy, the loss of a species may not necessarily lead to the loss of a function (Fonseca and Ganade, 2001; Blakey et al., 2019). Furthermore, species’ responses to environmental factors may be driven by their functional traits and/or evolutionary histories (Pellens and Grandcolas, 2016; Presley et al., 2018; Blakey et al., 2019). Thus, functional and phylogenetic studies can add valuable information about the consequences of converting a natural habitat to an agricultural systems or about the presence of these agricultural systems in landscapes. For example, Carvalho et al. (2020) described that while taxonomic and functional diversity is lower in acacia plantations than in forest patches, phylogenetic diversity is similar. In Olivier et al. (2020), phylogenetic diversity was found to respond negatively to the amount of agriculture in the landscape, but taxonomic diversity is unaffected. Additionally, most studies that used biodiversity to describe guilds and communities, employed α-diversity descriptors (n = 157, 98.7%), whereas β-diversity was less frequently utilized (n = 37, 23.3%). While α-diversity measures are crucial for understanding local biodiversity patterns, it is equally important to investigate how species composition, as described by β-diversity and its components (turnover or species replacement and nestedness or richness diference), is affected by agricultural ecosystems. For example, Alpízar et al. (2019) showed that taxonomic diversity was similar between pineapple cultivation and forests, but community composition differed between these habitats. The same pattern was observed by Olimpi and Philpott (2018) when comparing forest fragments with organic and conventional agriculture. We highlight that such investigations are essential for identifying conservation strategies for fauna in cultivated landscapes (Cardoso et al., 2014; Socolar et al., 2016).
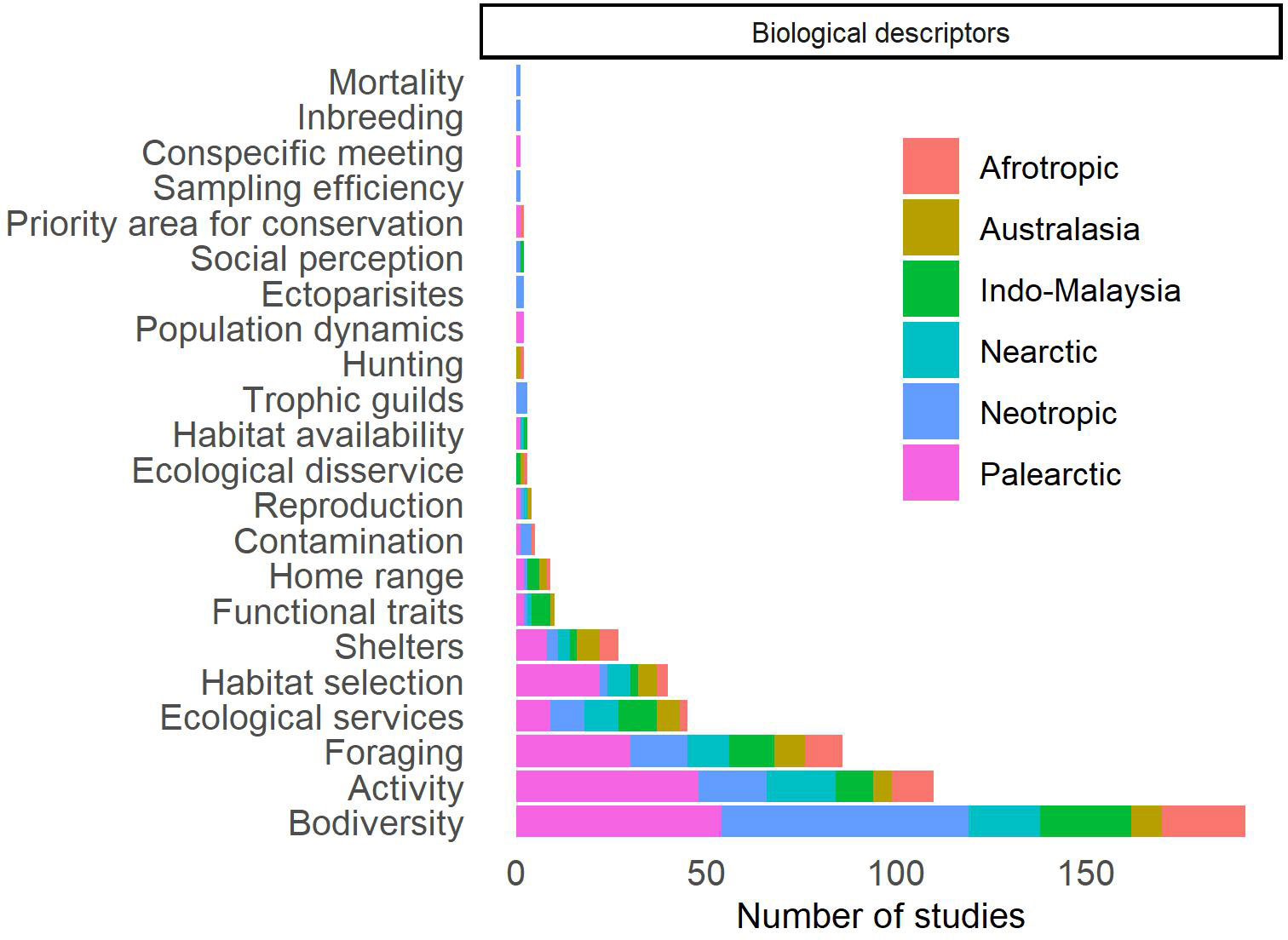
Figure 5 Distribution of studies published worldwide in which bats were sampled in agricultural systems or that assess the effect of these agricultural systems on bats, according to the biological descriptors used in these studies.
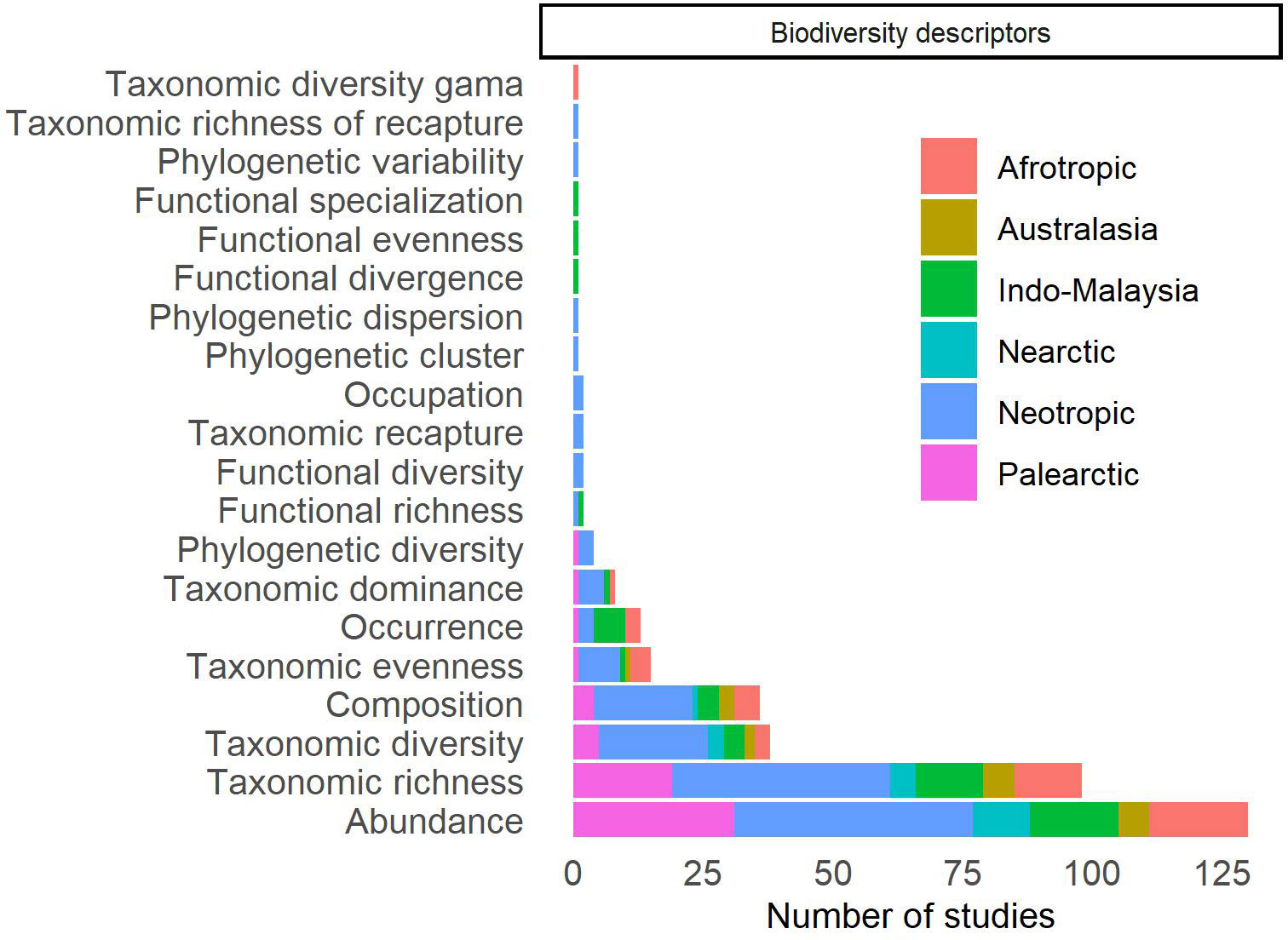
Figure 6 Distribution of studies published worldwide in which bats were sampled in agricultural systems or that assess the effect of these agricultural systems on bats, according to the biodiversity descriptors used in these studies.
We found that 20 biodiversity descriptors were used to describe bat communities and guilds (Figure 6). Of these, abundance was the most used (n = 130, 81.8%), followed by taxonomic richness (n = 98, 61.6%). Thus, many quantitative reviews use only these descriptors to assess disturbance effects on bat communities, at best, evaluating guilds separately (e.g., Cunto and Bernard, 2012; García-Morales et al., 2013; Williams-Guillén et al., 2016; Carballo-Morales et al., 2021). Abundance and taxonomic richness may show opposite trends when compared to other biodiversity indices. For example, Dawson et al. (2012) did not find a difference in abundance and taxonomic richness when comparing primary forests to plantations but found a decrease in taxonomic diversity in plantations. The scarcity of studies using other indices limits our understanding of how plantations affect bats. This gap in knowledge includes large-scale comparisons and meta-analyses, due to the limited information provided by existing studies.
The second most used biological descriptor was bats activity (n = 110, 35.6%; Figure 5; Supplementary Table 3). However, most studies (n = 105, 94.5%) used activity as a proxy for abundance. Thus, analyzes such as changes in the duration and peak of activity and social activity received less attention (n = 5, 8.2%). In addition to these few behavioral studies on bat activity, bat behavior has been evaluated in foraging studies (n = 86, 27.8%), habitat selection (n = 40, 12.9%), roosts selection (n = 27, 8.7%) and home range (n = 9, 2.9%), of which only the last was little represented. While we found several aspects of bat behavior to be well-studied, it is important to note that most of these studies were conducted in the Palearctic region (Supplementary Table 3). Therefore, future studies focusing on these topics should be conducted in regions other than the Palearctic, to gain a comprehensive understanding of bat behavior in different environments.
Out of the 45 studies (14.6%; Supplementary Table 3) that investigated the ecological services provided by bats, 33 focused on pest control/suppression, nine on pollination, three on seed dispersal, and one on litter decomposition. However, it is possible that our review was not able to locate all studies on this topic, as some may not mention the agricultural systems themselves. For example, despite the fact that seed dispersal is often cited as one of the most important and more studied ecosystem services provided by bats, according to recent studies that do not focus on agricultural systems (Castillo-Figueroa, 2020; Regolin et al., 2020), our analysis revealed that it was one of the least studied services among the studies reviewed. However, other comprehensive reviews, such as Ramírez-Fráncel et al. (2022), found that most ecosystem services studies aim to study pest suppression, and other ecosystem services, such as seed dispersal, are less studied. Eight studies assigned monetary value to the service provided, of which six of them dealt with pest control and two with pollination. Although challenging, it is essential to value the benefits of bats to ecosystem functioning, and ultimately to humans, in order to raise awareness about their importance and ensure their protection and conservation (Kunz et al., 2011; Boyles et al., 2013). We recommend further efforts to investigate the impact of agricultural systems on ecological services beyond agricultural pest control/suppression, as well as to improve the quantification of the monetary value of these services.
Few studies used trophic guilds (n = 3) or other guilds (n = 10) as biological descriptors (Supplementary Table 3). Those which did, evaluated the changes in composition or proportion of functional (n =3) and trophic guild (n = 10) characteristics. The functional traits used in these cases were: morphological aspects (n = 8), mainly from the bat wings (n = 7); echolocation parameters (n = 4); body mass (n = 3); fertility (n = 2); growth rate (n = 2); specialization in habitat use (n = 1); and diet specialization (n = 1). However, as already discussed, many studies used these guilds as focal taxa, analyzing them with abundance and taxonomic descriptors, and, thus, the guilds ended up being extensively explored in the studies selected here.
Contamination of bats by pesticides has received little attention (n = 5, 1.6%; Supplementary Table 3). However, it is possible that our review was not able to locate all studies on this topic, as some may not mention the agricultural systems themselves. Oliveira et al. (2021), identified 28 recent studies on the contamination of bats by pesticides, indicating a higher number compared to our study, although the total number of studies remains small considering the importance of this topic. Contamination studies were the only ones to consider physiological responses. These studies, along with those the assessment of the presence and prevalence of ectoparasites (n = 2, 0.6%), were the sole investigations that encompassed the assessment of potential diseases affecting bats within agricultural systems or cultivated landscapes. No other diseases or cases of zoonoses were evaluated in the studies selected. Sex ratio or other issues related to reproduction were also little studied (n = 4, 1.3%). In addition to these, four other studies (on home range, diet composition and diversity, and roost characteristics) considered the gender of the studied individuals. Apart from studies that assess the abundance of a single species, populations were analyzed in only a few studies (two studies on population dynamics, one on inbreeding, one on conspecific encounter, one on differentiation and genetic diversity, and one on allelic richness). Apart from these few population studies, none other used genetic descriptors (Supplementary Table 3).
Mitigation proposals were made in 205 studies (66.3%), and these were categorized into 16 types (Figure 7; Supplementary Table 3). The measure proposed more often in the selected studies was the conservation or restoration of natural, semi-natural, or potential areas for bats in cultivated landscapes (n = 122, 59.5%) These studies refer mainly to forested habitats, but wetlands, karstic areas and savannas are also cited. Natural forested habitats vary from continuous and well-preserved forests, with the potential to maintain the original bat community, to riparian forests and forest fragments that can function as corridors and promote connectivity (Yoshikura et al., 2011; Akasaka et al., 2012; Toffoli and Rughetti, 2020). The preservation or restoration of secondary forests have also been suggested to provide resources for some less sensitive bat species (Louzada et al., 2010; Ervis et al., 2021). The second most cited measure was less intensive management (n = 85, 41.5%). These studies mention “friendly” agriculture, agroforestry, organic agriculture, traditional agriculture, polyculture, fallow practices, shaded plantations, presence of trees, presence of forest characteristics, maintenance or clearance of the understory in forest plantations, decrease and regulation of pesticide use, and increased heterogeneity and structural complexity of plantations (e.g., Fuller et al., 2005; Faria et al., 2006; Dietz et al., 2013). These measures are intended to make these environments less hostile for bats, increasing their roosting and foraging potential and facilitating displacement between adjacent habitats (Law and Chidel, 2006; Cortes-Delgado and Sosa, 2014; Rodríguez-San Pedro and Simonetti, 2015; Fill et al., 2021). In third place, studies propose that increasing or maintaining connectivity is an important measure to mitigate the presence of agricultural systems in landscapes (n = 60, 29.3%). These studies propose that the presence of riparian forests, forest fragments, trees in plantations, networks of water bodies and the presence of linear elements (e.g., living fences) can promote connectivity (Davy et al., 2007; Akasaka et al., 2012; Heer et al., 2015; Heim et al., 2016). In addition to the aforementioned recommendations, several studies emphasize the importance of increasing roost availability (n = 41, 20.0%), both artificial and natural, as well as protecting and restoring foraging areas (n = 26, 12.7%). To achieve this, it may be necessary to maintain natural or semi-natural areas that have the potential to support bat populations, promote less intensive land management practices, utilize artificial roosts, and increase connectivity to ensure that bats can access roosts and food resources within cultivated landscapes (Koschnicke et al., 2010; Wordley et al., 2017). Restoration of water bodies (n = 21, 10.2%) to maintain connectivity and foraging areas, and other landscape-scale management, such as preserving heterogeneity (n = 24, 11.7%) and original landscape structure (n = 2, 1.0%) were also mentioned. Some studies also proposed other mitigations, such as preserving the quality of remaining natural habitats (n = 2, 1.0%), using degraded areas for planting (n = 3, 1.5%), prohibiting bat hunting (n = 4, 2.0%), and avoiding large-scale planting (n = 6, 2.9%).
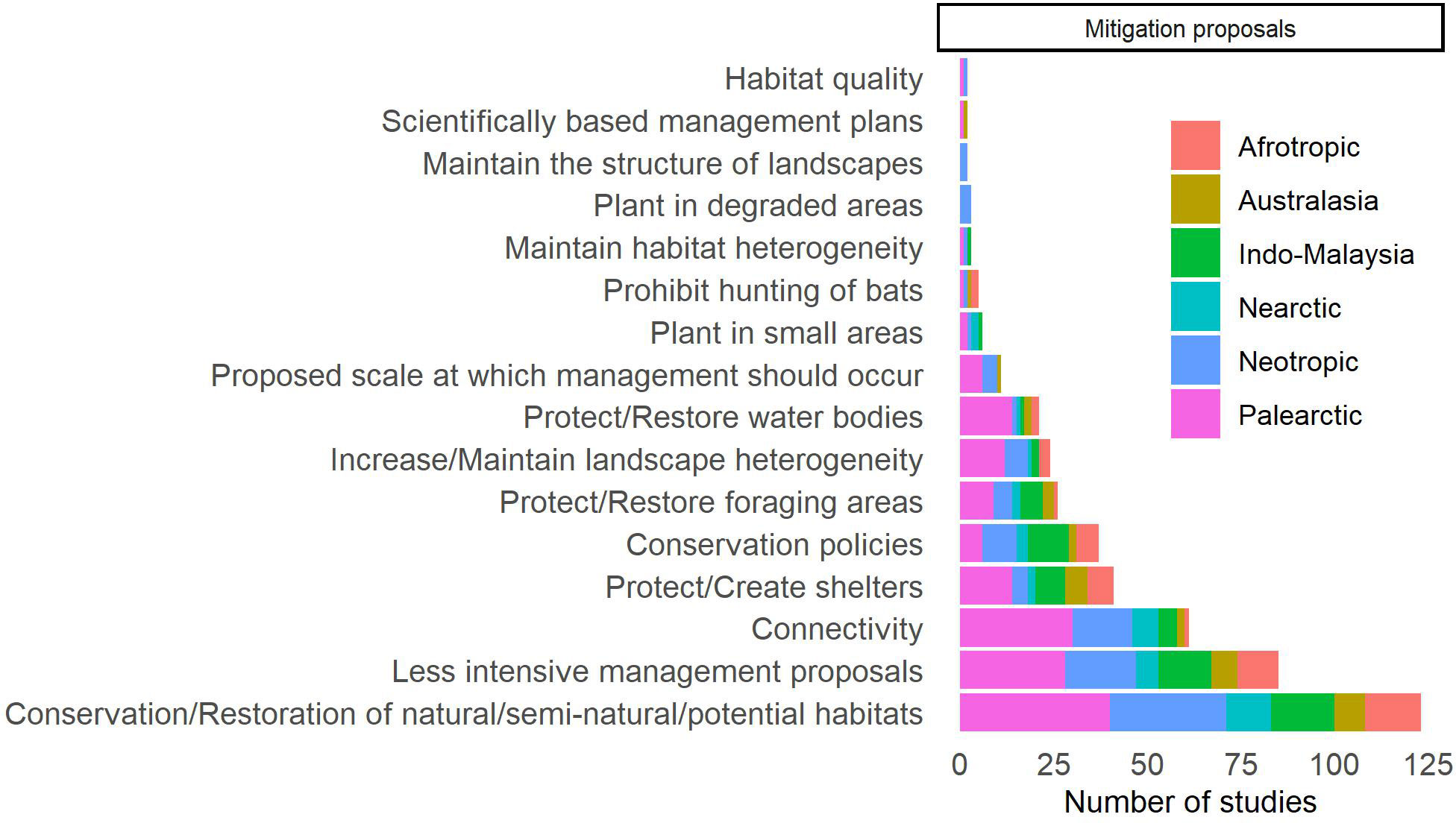
Figure 7 Distribution of studies published worldwide in which bats were sampled in agricultural systems or that assess the effect of these agricultural systems on bats, according to the proposals for mitigating the consequences of agricultural systems for bats.
Some studies also mentioned the importance of implementing policies and regulations (n = 36, 17.6%). In most cases, the measures aim to create actions that satisfy both producers and bat conservation efforts simultaneously. Among these, include the promotion of environmental education to highlight the importance of bats in providing ecological services, incorporating local and ecological scientific knowledge in mitigation measures, and fostering collaboration between scientists, legislators, and producers. Furthermore, financial incentives were also proposed that add value to sustainable products pollinated or dispersed by bats, which can benefit everyone, including small producers. Additionally, other actions were suggested, like involving local residents in inspections, promoting ecotourism, and exploring technologies to reduce conflicts between humans and bats (e.g., using protection nets in fruit production). Finally, some studies recommended funding research focused on developing effective mitigation measures (MacDonald et al., 2018; Sheherazade et al., 2019; Sow et al., 2020; Oleksy et al., 2021).
Our review resulted in a considerable number of studies (n = 309) related with bats and agricultural systems. These studies revealed an uneven distribution of knowledge across global regions, with a predominant focus on the Palearctic and Neotropical regions. This highlights the need to direct more resources on other geographical regions. Specifically, we recommend prioritizing the Indo-Malaysia and Afrotropic regions. These areas have received limited attention in previous studies, and they are critical regions for bat conservation. To understand how bats are affected by agricultural systems, we need filling this and others knowledge gaps that we identified here, such as the groups of bats studied, and the methods applied for data collection. For example, although most studies aimed to assess the bat community, few have combined sampling methods that allows knowing the whole community, which has biased results towards certain groups (e.g., insectivorous bats, phyllostomid bats). Combining sampling methods is important for a more comprehensive sampling of these bat communities. Complementary methods are more often needed in regions where bats that fly high and/or have efficient echolocation to perceive interception traps (e.g., mist nets) coexist with bats that do not echolocate or do not have enough variation in echolocation to be identified by acoustic recordings. Another critical aspect that can impact our comprehension of the effects of commercial plantations is the type of descriptors used. Most studies rely on taxonomic biodiversity as the primary descriptor, with a particular emphasis on abundance and taxonomic richness. However, there is a need to explore other biodiversity dimensions, such as phylogenetic and functional diversity, and β-diversity. Furthermore, it is worth noting that relying solely on abundance and taxonomic richness can lead to incomplete interpretations, as these indices can show opposing trends when compared to other biodiversity indices. Therefore, we highly recommend utilizing multiple biodiversity indices to gain a more holistic understanding of the effects of agricultural systems on bat populations.
Our analysis uncovered several other significant gaps in current research on the effects of agricultural systems on bat populations. For instance, few studies have examined how the presence or number of trophic resources, or the influence of abiotic factors, impact bats in agricultural systems. We recommend evaluating these variables, particularly outside the Palearctic region, where these studies are most often performed. Additionally, we found that studies that investigated contamination, disease, and other physiological responses, as well as those that assessed sex ratio, reproduction, and genetic diversity, were severely lacking. Only less than 10 studies have focused on these crucial factors. While many studies focused on a single bat species, most primarily aimed to understand bat abundance response, other critical population parameters such as population dynamics, inbreeding, and conspecific encounter have often been overlooked. Therefore, we urge researchers to broaden their focus and explore these important population parameters. Furthermore, although there is a balanced number of studies using different scales (except for those that make comparisons between landscapes), studies at the landscape scale are concentrated in the Palearctic region. Thus, in addition to the need for more studies at all scales outside the Palearctic and Neotropical regions, it is necessary to concentrate efforts on landscape-scale studies in the Neotropical region. Our analysis suggests that the optimal scale size in landscape studies depends on the home range of the bats under investigation. As such, we recommend that greater efforts be made to understand this parameter, for instance, by using telemetry studies. In the absence of such data, we believe that studies that utilize multiple scale sizes or make comparisons between different landscapes can provide a more comprehensive understanding of bats in cultivated landscapes.
We also found gaps in the type of agricultural systems studied. There are still few studies that assess the consequences that forest plantations that require the cutting of trees have on bats, in all regions. These type of plantations may have different impacts from other agricultural systems (grassland-cropland systems and forest that do not require cutting) on bat communities, as they are subject to cyclical changes in their structure; we recommend further studies in these type of agricultural systems. Furthermore, predominantly non-forested biomes (e.g., African and South American savannas), as well as non-forested habitats (e.g., flooded and natural fields), have been understudied compared to forested ones. The most frequently proposed mitigation measure in the selected studies was the conservation or restoration of natural, semi-natural, or potential bat habitats in cultivated landscapes. Therefore, understanding what these potential habitats are and how they benefit bats in agricultural systems will help to outline strategies for the conservation of these organisms. This reinforces the importance of studies that evaluate natural non-forest habitats as well as forest ones. Non-forest habitats can influence bat communities by functioning as complementary habitats for resource and promote movement and their replacement by agricultural systems can change the dynamics of these landscapes. In addition, studies of bat behavior, such as those using telemetry, were more common in the Palearctic region and are extremely relevant to understand what these potential areas are. These studies can provide information on home range size, roosting and foraging habitat selection, dispersal patterns and, in addition, can help determine the ideal scale size for landscape studies. The relevance of the conservation or restoration of natural, semi-natural or potential areas for bats also reinforces the importance of focusing on studies that considered natural/semi-natural areas (as the amount and presence of natural/semi-natural areas affects bats in cultivated landscapes) in regions where these studies are scarce (outside the Neotropical region). In addition to this landscape composition variable, studies evaluating the configuration variables of cultivated landscapes were scarce in all regions. Connectivity and other landscape configuration variables, such as heterogeneity, were cited in many studies as mitigating measures. Thus, we reinforce the importance of studying this landscape parameter. In addition to conservation or restoration of natural bat habitats, many studies also proposed less intensive agricultural system management as a mitigation measure. Therefore, a better understanding of how bats respond to different types of management, particularly outside the Palearctic and Neotropics, where these studies are most often performed, is essential in creating effective strategies for improving connectivity in these landscapes. This understanding will also enable the identification of ways to increase roost and food availability within plantations, thereby making them complementary habitats for bats.
Raising awareness of the importance of bats is crucial for their conservation. However, few studies have investigated producers’ perception of bats. Furthermore, the ecological service studies included in this analysis have predominantly focused on agricultural pest control/suppression, with limited research exploring the impact of agricultural systems on other ecological services provided by bats. Moreover, only a few studies have attributed a monetary value to these services. Therefore, we recommend increased efforts to understand the impact of agricultural systems on ecological services other than pest control/suppression and to assign economic value to these services. Finally, conservation policies are vital for the success of these strategies, as they can promote environmental education, raise awareness among local populations and producers about the importance of bats, provide financial incentives to producers for their conservation efforts, and fund research to fill knowledge gaps.
In summary, our findings highlight the urgent need for a more comprehensive understanding of the impact of agricultural systems on bat conservation, particularly in the Afrotropic and Indo-Malaysia regions. In addition, future studies should focus on forest plantations that require cutting, predominantly non-forested biomes (e.g., African and South American savannas), and non-forested habitats (e.g., flooded and natural fields). Also, studies should use a combination of different sampling methods (e.g., mist net and acoustic recorders), telemetry, and multiple biodiversity descriptors. Further research is essential to gain a comprehensive understanding of the impact of trophic resources and abiotic factors, contamination, disease prevalence, physiological responses, landscape-scale effects, population ecology, and the ecological services provided by bats. Additionally, there is a need to enhance the role of conservation policies in promoting bat conservation, as well as raise awareness of their importance among producers and local communities. Only by filling these gaps and understanding all the factors that influence bat occurrence and survival in these human impacted habitats, it becomes possible to develop science-supported and effective management and conservation strategies, in areas affected by agricultural systems.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
BSX, WC, and MV originally formulated the idea. BSX conducted the review and compilation of data. BSX performed the qualitative analysis of this review. BSX wrote the drafts of the main manuscript and the online resources. All authors contributed article and approved the submitted version.
This work was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (88882.42497512019-01), Spanish Ministry of Universities (CA3/RSUE/2021-00197), Neotropical Grassland Conservancy (01098803-5), National Geographic Society (EC-59186R-19 and NGS-83254R-20), Bat Conservation International (2434131) e Rufford Foundation (25585-1).
We thank all authors of the studies included in this review. We thank to Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil for the PhD scholarship granted to BdSX and for the postdoctoral fellowship granted to WC. AMCS was supported by the Ramón y Cajal program (grant RYC2020-029407-I), funded by MCIN/AEI/ 10.13039/501100011033 and by “ESF Investing in your future”. WC receives a postdoctoral fellowship from ‘Ayudas Maria Zambrano’ (CA3/RSUE/2021-00197) founded by Spanish Ministry of Universities. We thank to Neotropical Grassland Conservancy (01098803-5), National Geographic Society (EC-59186R-19 and NGS-83254R-20), Bat Conservation International (2434131) and Rufford Foundation (25585-1) for funding the BSX doctorate.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1214176/full#supplementary-material
Aizpurua O., Budinski I., Georgiakakis P., Gopalakrishnan S., Ibanez C., Mata V., et al. (2018). Agriculture shapes the trophic niche of a bat preying on multiple pest arthropods across Europe: Evidence from DNA metabarcoding. Mol. Ecol. 27 (3), 815–825. doi: 10.1111/mec.14474
Akasaka T., Akasaka M., Nakamura F. (2012). Scale-independent significance of river and riparian zones on three sympatric Myotis species in an agricultural landscape. Biol. Conserv. 145, 15–23. doi: 10.1016/j.biocon.2011.08.017
Alpízar P., Rodríguez-Herrera B., Jung K. (2019). The effect of local land use on aerial insectivorous bats (Chiroptera) within the two dominating crop types in the Northern Caribbean lowlands of Costa Rica. PloS One 14 (1), e0210364. doi: 10.1371/journal.pone.0210364
Appel G., Capaverde-Jr. U. D., De Oliveira L. Q., Pereira L. G. A., Tavares V. C., López-Baucells A., et al. (2021). Use of complementary methods to sample bats in the Amazon. Acta Chiropterologica 23 (2), 499–511. doi: 10.3161/15081109ACC2021.23.2.017
Aziz S. A., Olival K. J., Bumrungsri S., Richards G. C., Racey P. A. (2016). “The conflict between pteropodid bats and fruit growers: species, legislation and mitigation,” in The anthropocene: conservation of bats in a changing world. Eds. Voight C. C., Kingston T. (London, UK: Springer), 377–426.
Barré K., Le Viol I., Julliard R., Chiron F., Kerbiriou C. (2017). Tillage and herbicide reduction mitigate the gap between conventional and organic farming effects on foraging activity of insectivorous bats. Ecol. Evol. 8, 1496–1506. doi: 10.1002/ece3.3688
Blakey R. V., Webb E. B., Kesler D. C., Siegel R. B., Corcoran D., Johnson M. (2019). Bats in a changing landscape: Linking occupancy and traits of a diverse montane bat community to fire regime. Ecol. Evol. 9, 5324–5337. doi: 10.1002/ece3.5121
Boyles J. G., Sole C. L., Cryan P. M., McCracken G. F. (2013). “On estimating the economic value of insectivorous bats: prospects and priorities for biologists,” in Bat evolution, ecology, and conservation. Eds. Adams R. A., Pedersen S. C. (New York, USA: Springer Science+Business Media), 501–515.
Carballo-Morales J. D., Saldaña-Vázquez R. A., Villalobos F. (2021). Trophic guild and forest type explain phyllostomid bat abundance variation from human habitat disturbance. Global Ecol. Conserv. 25, e01425. doi: 10.1016/j.gecco.2020.e01425
Cardoso P., Rigal F., Carvalho J. C., Fortelius M., Borges P. A. V., Podani J., et al. (2014). Partitioning taxon, phylogenetic and functional beta diversity into replacement and richness difference components. J. Biogeography 41, 749–761. doi: 10.1111/jbi.12239
Carvalho W. D., Meyer C. F. J., Xavier B. S., Mustin K., Castro I. J., Silvestre S. M., et al. (2020). Consequences of replacing native savannahs with acacia plantations for the taxonomic, functional, and phylogenetic α- and β-diversity of bats in the northern Brazilian amazon. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.609214
Carvalho W. D., Miguel J. D., Xavier B. S., López−Baucells A., Castro I. J., Hilário R. R., et al. (2023). Complementarity between mist−netting and low−cost acoustic recorders to sample bats in Amazonian rainforests and savannahs. Community Ecol. 24, 47–60. doi: 10.1007/s42974-022-00131-5
Carvalho W. D., Mustin K., Farneda F. Z., Castro I. J., Hilário R. R., Martins A., et al. (2021). Taxonomic, functional and phylogenetic bat diversity decrease from more to less complex natural habitats in the Amazon. Oecologia. 196, 1–17. doi: 10.1007/s00442-021-05009-3
Castillo-Figueroa D. (2020). Why bats matters: a critical assessment of bat mediated ecological processes in the neotropics. Eur. J. Ecol. 6 (1), 77–101. doi: 10.17161/eurojecol.v6i1.13824
Castillo-Figueroa D., Pérez-Torres J. (2021). On the development of a trait-based approach for studying Neotropical bats. Papéis Avulsos Zoologia 61, e20216124. doi: 10.11606/1807-0205/2021.61.24
Chaiyarat R., Boonma W., Koedrith P. (2020). The role of pteropodid bats in pollination of durian (Durio zibethinus) in managed orchards in suburban habitat of Thailand. Urban Ecosyst. 23, 97–106. doi: 10.1007/s11252-019-00919-w
Charbonnier Y., Papura D., Touzot O., Rhouy N., Sentenac G., Rusch A. (2021). Pest control services provided by bats in vineyard landscapes. Agriculture Ecosyst. Environ. 306, 107207. doi: 10.1016/j.agee.2020.107207
Cisneros L. M., Fagan M. E., Willig M. R. (2015). Effects of human-modified landscapes on taxonomic, functional and phylogenetic dimensions of bat biodiversity. Diversity Distributions 21 (5), 523–533. doi: 10.1111/ddi.12277
Clerc J., Brigham R. M., Boyles J. G., McGuire L. P. (2021). “A NASBR history of radiotelemetry: how technology has contributed to advances in bat biology,” in 50 years of bat research. Eds. Lim B. K., Fenton M. B., Brigham R. M., Mistry S., Kurta A., Gillam E. H., et al. (Cham, CH: Springer), 241–253.
Cortes-Delgado N., Sosa V. J. (2014). Do bats roost and forage in shade coffee plantations? A perspective from the frugivorous bat sturnira hondurensis. Biotropica 46 (5), 624–632. doi: 10.1111/btp.12142
Costa A., Silva B., Jiménez-Navarro G., Barreiro S., Melguizo-Ruiz N., Rodríguez-Pérez J. (2020). Structural simplification compromises the potential of common insectivorous bats to provide biocontrol services against the major olive pest Prays oleae. Agriculture Ecosyst. Environ. 287, 106708. doi: 10.1016/j.agee.2019.106708
Cunto G. C., Bernard A. E. (2012). Neotropical bats as indicators of environmental disturbance: what is the emerging message? Acta Chiropterologica 14 (1), 143–151. doi: 10.3161/150811012X654358
Curveira-Santos G., Santos M. J., Santos-Reis M., Rosalino L. M. (2021). Global patterns of carnivore spatial ecology research in agroecosystems. Biodiversity Conserv. 30, 257–273. doi: 10.1007/s10531-020-02093-4
Davy C. M., Russo D., Fenton M. B. (2007). Use of native woodlands and traditional olive groves by foraging bats on a Mediterranean island: consequences for conservation. J. Zoology 273, 397–405. doi: 10.1111/j.1469-7998.2007.00343.x
Dawson J., Turner C., Pileng O., Farmer A., McGary C., Walsh C., et al. (2012). The bats (Mammalia: Chiroptera) of the lower Waria Valley, Morobe Province, Papua New Guinea: a survey and comparison across habitat types using mist nets. Aust. Mammalogy 34, 234–244. doi: 10.1071/AM11030
Dietz M., Pir J. B., Hillen J. (2013). Does the survival of greater horseshoe bats and Geoffroy’s bats in Western Europe depend on traditional cultural landscapes? Biodiversity Conserv. 22, 3007–3025. doi: 10.1007/s10531-013-0567-4
Dinerstein E., Olson D., Joshi A., Vynne C., Burgess N. D., Wikramanayake E. (2017). An ecoregion-based approach to protecting half the terrestrial realm. BioScience 67 (6), 534–545. doi: 10.1093/biosci/bix014
Ervis M. D., Eric-Moise B. F., Aaron M. M., Jules A. P., Fernand-Nestor T. F. (2021). Diversity pattern of bats (Mammalia: Chiroptera) in a modified tropical environment in the western region of Cameroon. Afr. Zoology 56 (2), 133–145. doi: 10.1080/15627020.2021.1932588
Faria D., Baumgarten J. (2007). Shade cacao plantations (Theobroma cacao) and bat conservation in southern Bahia, Brazil. Biodiversity Conserv. 16, 291–312. doi: 10.1007/s10531-005-8346-5
Faria D., Laps R. R., Baumgarten J., Cetra M. (2006). Bat and bird assemblages from forests and shade cacao plantations in two contrasting landscapes in the Atlantic Forest of southern Bahia, Brazil. Biodiversity Conserv. 15, 587–612. doi: 10.1007/s10531-005-2089-1
Farneda F. Z., Meyer C. F. J., Grelle C. E. V. (2020). Effects of land-use change on functional and taxonomic diversity of Neotropical bats. Biotropica 52, 120–128. doi: 10.1111/btp.12736
Fill C. T., Allen C. R., Benson J. F., Twidwell D. (2021). Roost use and movements of northern long-eared bats in a southeast nebraska agricultural landscape. Am. Midland Nat. J. 185, 241–248. doi: 10.1674/0003-0031-185.2.241
Fischer J., Lindenmayer D. B. (2007). Landscape modification and habitat fragmentation: a synthesis. Global Ecol. Biogeography 16 (3), 265–280. doi: 10.1111/j.1466-8238.2007.00287.x
Flaquer C., Torre I., Arrizabalaga A. (2007). Comparison of sampling methods for inventory of bat communities. J. Mammalogy 88 (2), 526–533. doi: 10.1644/06-MAMM-A-135R1.1
Fonseca C. R., Ganade G. (2001). Species functional redundancy, random extinctions and the stability of ecosystems. J. Ecol. 89, 118–125. doi: 10.1046/j.1365-2745.2001.00528.x
Food and Agriculture Organization. (2020a). FAOSTAT. https://www.fao.org/faostat/en/#data/EL (Accessed February 22, 2023).
Food and Agriculture Organization (2020b). Global forest resources assessment 2020 (Rome: Food and Agriculture Organization). doi: 10.4060/ca9825en
Frick W., Kingston T., Flanders J. (2019). A review of the major threats and challenges to global bat conservation. Ann. New York Acad. Sci. 1469, 5–25. doi: 10.1111/nyas.14045
Froidevaux J. S. P., Louboutin B., Jones G. (2017). Does organic farming enhance biodiversity in Mediterranean vineyards? A case study with bats and arachnids. Agriculture Ecosyst. Environ. 249, 112–122. doi: 10.1016/j.agee.2017.08.012
Fuller R. J., Norton L. R., Feber R. E., Johnson P. J., Chamberlain D. E., Joys A. C., et al. (2005). Benefits of organic farming to biodiversity vary among taxa. Biol. Lett. 1, 431–434. doi: 10.1098/rsbl.2005.0357
García-Morales R., Badano E. I., Moreno C. E. (2013). Response of neotropical bat assemblages to human land use. Conserv. Biol. 27 (5), 1096–1106. doi: 10.1111/cobi.12099
Gibson L., Lee T. M., Koh L. P., Brook B. W., Gardner T. A., Barlow J., et al. (2011). Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 478, 378–381. doi: 10.1038/nature10425
Gonthier D. J., Ennis K. K., Farinas S., Hsieh H.-Y., Iverson A. L., Batáry P., et al. (2014). Biodiversity conservation in agriculture requires a multi-scale approach. Proc. R. Soc. B 281, 20141358. doi: 10.1098/rspb.2014.1358
Heer K., Helbig-Bonitz M., Fernandes R. G., Mello M. A. R., Kalko E. K. V. (2015). Effects of land use on bat diversity in a complex plantation-forest landscape in northeastern Brazil. J. Mammalogy 96 (4), 720–731. doi: 10.1093/jmammal/gyv068
Heim O., Schröder A., Eccard J., Jung K., Voigt C. C. (2016). Seasonal activity patterns of European bats above intensively used Farmland. Agriculture Ecosyst. Environ. 233, 130–139. doi: 10.1016/j.agee.2016.09.002
Herrera J. M., Salgueiro P. A., Medinas D., Costa P., Encarnação C., Mira A. (2016). Generalities of vertebrate responses to landscape composition and configuration gradients in a highly heterogeneous Mediterranean region. J. Biogeography 43, 1203–1214. doi: 10.1111/jbi.12720
James K. L., Randall N. P., Haddaway N. R. (2016). A methodology for systematic mapping in environmental sciences. Environ. Evidence 5, 7. doi: 10.1186/s13750-016-0059-6
Jones G., Teeling E. C., Rossiter S. J. (2013). From the ultrasonic to the infrared: molecular evolution and the sensory biology of bats. Front. Physiolog 4. doi: 10.3389/fphys.2013.00117
Kalka M. B., Smith A. R., Kalko E. K. V. (2008). Bats limit arthropods and herbivory in a tropical forest. Science 320 (5872), 71. doi: 10.1126/science.1153352
Kalko E. K. V. (1998). Organisation and diversity of tropical bat communities through space and time. Zoology 101, 281–297.
Kalko E. K. V., Handley C. O., Handley D. (1996). “Organization, diversity, and long-term dynamics of a neotropical bat community,” in Long-term studies of vertebrate communities. Eds. Martin L. C., Jeffrey A. S. (Los Angeles, USA: Academic Press), 503–553.
Koschnicke S., Franke L., van Doormaal F., Kuipers H., Weterings M., Gloza-Rausch F., et al. (2010). Bat boxes as a tool for biological insect pest control on cocoa plantations in Ghana. Nyctalus (N.F.) 4, 357–366.
Kunz T. H., de Torrez E. B., Bauer D., Lobova T., Fleming T. H. (2011). Ecosystem services provided by bats. Ann. New York Acad. Sci. 1223, 1–38. doi: 10.1111/j.1749-6632.2011.06004.x
Kunz T. H., Lumsden L. F. (2003). “Ecology of cavity and foliage roosting bats,” in Bat ecology. Eds. Kunz T. H., Fenton M. B. (Chicago, USA: University of Chicago Press), 3–89.
Law B. S., Chidel M. (2006). Eucalypt plantings on farms: Use by insectivorous bats in south-eastern Australia. Biol. Conserv. 133, 236–249. doi: 10.1016/j.biocon.2006.06.016
Law B. S., Park K. J., Lacki M. J. (2016). “Insectivorous bats and silviculture: balancing timber production and bat conservation,” in The anthropocene: conservation of bats in a changing world. Eds. Voight C. C., Kingston T. (London, UK: Springer), 105–150.
Lentini P. E., Gibbons P., Fischer J., Law B., Hanspach J., Martin T. G. (2012). Bats in a farming landscape benefit from linear remnants and unimproved pastures. PloS One 7 (11), e48201. doi: 10.1371/journal.pone.0048201
Loeb S. C. (2020). Qualitative synthesis of temperate bat responses to silvicultural treatments - where do we go from here? J. Mammalogy 101 (6), 1513–1525. doi: 10.1093/jmammal/gyaa089
Louzada J., Gardner T., Peres C., Barlow J. (2010). A multi-taxa assessment of nestedness patterns across a multiple-use Amazonian forest landscape. Biol. Conserv. 143, 1102–1109. doi: 10.1016/j.biocon.2010.02.003
Luz J. L., Costa L. M., Esbérard C. E. L. (2020). Influence of banana plantations on bat assemblages (Chiroptera). Austral Ecol. 46 (3), 349–358. doi: 10.1111/aec.12989
Maas B., Karp D. S., Bumrungsri S., Darras K., Gonthier D., Huang J. C.-C., et al. (2016). Bird and bat predation services in tropical forests and agroforestry landscapes. Biol. Rev. 91 (4), 1081–1101. doi: 10.1111/brv.12211
MacDonald M. A., Angell R., Dines T. D., Dodd S., Haysom K. A., Hobson R., et al. (2018). Have Welsh agri-environment schemes delivered for focal species? Results from a comprehensive monitoring programme. J. Appl. Ecol. 56, 812–882. doi: 10.1111/1365-2664.13329
MacSwiney-G M. C., Clarke F. M., Racey P. A. (2008). What you see is not what you get: the role of ultrasonic detectors in increasing inventory completeness in Neotropical bat assemblages. J. Appl. Ecol. 45, 1364–1371. doi: 10.1111/j.1365-2664.2008.01531.x
May F., Rosenbaum B., Schurr F. M., Chase J. M. (2019). The geometry of habitat fragmentation: Effects of species distribution patterns on extinction risk due to habitat conversion. Ecol. Evol. 9, 2775–2790. doi: 10.1002/ece3.4951
McFadden T. N., Dirzo R. (2018). Opening the silvicultural toolbox: A new framework for conserving biodiversity in Chilean timber plantations. For. Ecol. Manage. 425, 75–84. doi: 10.1016/j.foreco.2018.05.028
Mendes E. S., Fonseca C., Marques S. F., Maia D., Pereira M. J. R. (2017). Bat richness and activity in heterogeneous landscapes: guild-specific and scale-dependent? Landscape Ecol. 32, 295–311. doi: 10.1007/s10980-016-0444-0
Mendes P., Srbek-Araujo N. A. (2021). Effects of land-use changes on Brazilian bats: a review of current knowledge. Mammal Rev. 51, 127–142. doi: 10.1111/mam.12227
Meyer C. F., Struebig M. J., Willig M. R. (2016). “Responses of tropical bats to habitat fragmentation, logging, and deforestation Bats,” in The anthropocene: conservation of bats in a changing world. Eds. Voight C. C., Kingston T. (London, UK: Springer), 63–103.
Moher D., Liberati A., Tetzlaff J., Altman D. G., PRISMA Group (2009). Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PloS Med. 6 (7), e1000097. doi: 10.1136/bmj.n71
Moreno C. E., Calderón-Patrón J. M., Martín-Regalado N., Martínez-Falcon A. P., Ortega-Martínez I. J., Rios-Díaz C. L., et al. (2018). Measuring species diversity in the tropics: a review of methodological approaches and framework for future studies. Biotropica 50 (6), 929–941. doi: 10.1111/btp.12607
Muñoz A. E., Ossa G., Zaviezoa T., Bonacic C. (2019). Diversity and foraging activity of bats in cultivated and uncultivated areas in agroecosystems of a Mediterranean-climate hotspot. Neotropical Biodiversity 5 (1), 36–40. doi: 10.1080/23766808.2019.1621626
Newbold T., Bentle L. F., Hill S. L. L., Edgar M. J., Horton M., Su G., et al. (2020). Global effects of land use on biodiversity differ among functional groups. Funct. Ecol. 34, 684–693. doi: 10.1111/1365-2435.13500
Oleksy R. Z., Ayady C. L., TATAYAH V., Froidevaux C.J.J.S.P., Racey P. A., Jones G. (2021). The impact of the Endangered Mauritian flying fox Pteropus Niger on comercial fruit farms and the efficacy of mitigation. Oryx 55 (1), 114–121. doi: 10.1017/S0030605318001138
Olimpi E. M., Philpott S. M. (2018). Agroecological farming practices promote bats. Agriculture Ecosyst. Environ. 265, 282–291. doi: 10.1016/j.agee.2018.06.008
Oliveira J. M., Destro A. L. F., Freitas M. B., Oliveira L. L. (2021). How do pesticides affect bats? – A brief review of recent publications. Braz. J. Biol. 81 (2), 499–507. doi: 10.1590/1519-6984.225330
Olivier T., Thébault E., Elias M., Fontaine B., Fontaine C. (2020). Urbanization and agricultural intensification destabilize animal communities differently than diversity loss. Nat. Commun. 11, 2686. doi: 10.1038/s41467-020-16240-6
O’Mara M. T., Wikelski M., Dechmann D. K. N. (2014). 50 years of bat tracking: device attachment and future directions. Methods Ecol. Evol. 5, 311–319. doi: 10.1111/2041-210X.12172
Ongole S., Sankaran M., Karanth K. K. (2018). Responses of aerial insectivorous bats to local and landscape-level features of coffee agroforestry systems in Western Ghats, India. PloS One 13 (8), e0201648. doi: 10.1371/journal.pone.0201648
Outhwaite C. L., Ortiz A. M. D., Spooner F. E. B., Dalin C., Newbold T. (2022). Availability and proximity of natural habitat influence cropland biodiversity in forest biomes globally. Global Ecol. Biogeography 31 (8), 1589–1602. doi: 10.1111/geb.13532
Park K. J. (2015). Mitigating the impacts of agriculture on biodiversity: bats and their potential role as bioindicators. Mamm. Biol. 80, 191–204. doi: 10.1016/j.mambio.2014.10.004
Pech-Canche J. M., Estrella E., López-Castillo D. L., Hernández-Betancourt S. F., Moreno C. E. (2011). Complementarity and efficiency of bat capture methods in a lowland tropical dry forest of Yucatán, Mexico. Rev. Mexicana Biodiversidad 82, 896–903. doi: 10.22201/ib.20078706e.2011.3.683
Peixoto F. P., Braga P. H. P., Mendes P. (2018). A synthesis of ecological and evolutionary determinants of bat diversity across spatial scales. BMC Ecol. 18, 18. doi: 10.1186/s12898-018-0174-z
Pellens R., Grandcolas P. (2016). “Phylogenetics and conservation biology: drawing a path into the diversity of life,” in Biodiversity conservation and phylogenetic systematics: preserving our evolutionary heritage in an extinction crisis. Eds. Hawksworth D. L., Chaurasia A. (Berlin, Germany: Springer Open), 1–15.
Pina S. M. S., Meyer C. F. J., Zortéa M. (2013). A comparison of habitat use by phyllostomid bats (Chiroptera: Phyllostomidae) in natural forest fragments and Eucalyptus plantations in the Brazilian Cerrado. Chiroptera Neotropical 19 (3), 14–30.
Potapov P., Turubanova S., Hansen M. C., Tyukavina. A., Zalles V., Khan A., et al. (2022). Global maps of cropland extent and change show accelerated cropland expansion in the twenty-first century. Nat. Food 3, 19–28. doi: 10.1038/s43016-021-00429-z
Potter J. H. T., Davies K. T. J., Yohe L. R., Sanchez M. K. R., Rengifo E. M., Struebig M., et al. (2021). Dietary diversification and specialization in neotropical bats facilitated by early molecular evolution. Mol. Biol. Evol. 38 (9), 3864–3883. doi: 10.1093/molbev/msab028
Presley S. J., Cisneros L. M., Higgins C. L., Klingbeil B. T., Scheiner S. M., Willig M. R. (2018). Phylogenetic and functional underdispersion in Neotropical phyllostomid bat Communities. Biotropica 50 (1), 135–145. doi: 10.1111/btp.12501
Ramírez-Fráncel L. A., García-Herrera L. V., Losada-Prado S., Reinoso-Flórez G., Sánchez-Hernández A., Estrada-Villegas S., et al. (2022). Bats and their vital ecosystem services: a global review. Integr. Zoology 17 (1), 2–23. doi: 10.1111/1749-4877.12552
Regolin A. L., Muylaert R. L., Crestani A. C., Dáttilo W., Ribeiro M. C. (2020). Seed dispersal by Neotropical bats in human-disturbed Landscapes. Wildlife Res. 48 (1), 1–6. doi: 10.1071/WR19138
Rodríguez-San Pedro A., Ávila F., Chaperon P. N., Beltrán C. A., Allendes J. L., Grez A. A. (2021). The role of the adjacent habitat on promoting bat activity in vineyards: a case study from central Chile. Acta Chiropterologica 23 (1), 177–187. doi: 10.3161/15081109ACC2021.23.1.014
Rodríguez-San Pedro A., Rodríguez-Herbach C., Allendes J. L., Chaperon P. N., Beltrán C. A., Grez A. A. (2019). Responses of aerial insectivorous bats to landscape composition and heterogeneity in organic vineyards. Agriculture Ecosyst. Environ. 277, 74–82. doi: 10.1016/j.agee.2019.03.009
Rodríguez-San Pedro A., Simonetti J. A. (2015). The relative influence of forest loss and fragmentation on insectivorous bats: does the type of matrix matter? Landscape Ecol. 30, 1561–1572. doi: 10.1007/s10980-015-0213-5
Schoeman M. C., Monadjem A. (2018). Community structure of bats in the savannas of southern Africa: influence of scale and human land-use. Hystrix Ital. J. Mammalogy 29 (1), 3–10. doi: 10.4404/hystrix-00038-2017
Shapiro J. T., Bordignon M. O. (2014). Bat (Chiroptera) assemblages in three Cerrado fragments of Mato Grosso do Sul, southwestern Brazil. Check List 10 (6), 1380–1386. doi: 10.15560/10.6.1380
Shapiro J. T., Monadjem A., Rödere T., McCleery R. A. (2020). Response of bat activity to land cover and land use in savannas is scale-, season-, and guild-specific. Biol. Conserv. 241, 108245. doi: 10.1016/j.biocon.2019.108245
Sheherazade, Ober H. K., Tsang S. M. (2019). Contributions of bats to the local economy through durian pollination in Sulawesi, Indonesia. Biotropica 51, 913–922. doi: 10.1111/btp.12712
Socolar J. B., Gilroy J. J., Kunin W. E., Edwards D. P. (2016). How should beta-diversity inform biodiversity conservation? Trends Ecol. Evol. 31 (1), 67–80. doi: 10.1016/j.tree.2015.11.005
Sow A., Seye D., Faye E., Benoit L., Galan M., Haran J., et al. (2020). Birds and bats contribute to natural regulation of the millet head miner in tree-crop agroforestry systems. Crop Prot. 132, 105127. doi: 10.1016/j.cropro.2020.105127
Stephens S. S., Wagner M. R. (2007). Forest plantations and biodiversity: A fresh perspective. J. Forestry 105 (6), 307–313. doi: 10.1093/jof/105.6.307
Taylor P. J., Mkhari D., Mukwevho T., Monadjem A., Schoeman M. C., Schoeman C., et al. (2011). Bats as potential biocontrol agents in an agricultural landscape, Levubu Valley: diet, activity and species composition of bats in macadamia orchards and neighbouring natural habitats. South Afr. avocado growers’ Assoc. Yearbook 34, 54–64.
Taylor P. J., Sowler S., Schoeman M. C., Monadjem A. (2013). Diversity of bats in the Soutpansberg and Blouberg Mountains of northern South Africa: complementarity of acoustic and non-acoustic survey methods. South Afr. J. Wildlife Res. 43 (1), 12–26. doi: 10.3957/056.043.0117
Toffoli R., Rughetti M. (2020). Effect of water management on bat activity in rice paddies. Paddy Water Environ. 18, 687–695. doi: 10.1007/s10333-020-00811-w
Tuck S. L., Winqvist C., Mota F., Ahnstrom J., Turnbull L. A., Bengtsson J. (2014). Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis. J. Appl. Ecol. 51 (3), 746–755. doi: 10.1111/1365-2664.12219
Weier S. M., Grass I., Linden V. M. G., Tscharntke T., Taylor P. J. (2018). Natural vegetation and bug abundance promote insectivorous bat activity in macadamia orchards, South Africa. Biol. Conserv. 226, 16–23. doi: 10.1016/j.biocon.2018.07.017
Wiegand T., Revilla E., Moloney K. A. (2005). Effects of habitat loss and fragmentation on population dynamics. Conserv. Biol. 19 (1), 108–121. doi: 10.1111/j.1523-1739.2005.00208.x
Williams-Guillén K., Olimpi E., Maas B., Taylor P. J., Arlettaz R. (2016). “Bats in the anthropogenic matrix: challenges and opportunities for the conservation of chiroptera and their ecosystem services in agricultural landscapes,” in The anthropocene: conservation of bats in a changing world. Eds. Voight C. C., Kingston T. (London, UK: Springer), 151–186.
Wordley C. F. R., Sankaran M., Mudappa D., Altringham J. D. (2017). Bats in the Ghats: Agricultural intensification reduces functional diversity and increases trait filtering in a biodiversity hotspot in India. Biol. Conserv. 210, 48–55. doi: 10.1016/j.biocon.2017.03.026
Xavier B. S., Carvalho W. D., Dias D., Tabosa L. O., Santos C. E. L., Esbérard C. E. L. (2018). Bat richness (Mammalia: Chiroptera) in an area of montane Atlantic Forest in the Serra da Mantiqueira, state of Minas Gerais, southeast Brazil. Biota Neotropica 18 (2), e20170496. doi: 10.1590/1676-0611-BN-2017-0496
Yoh N., Syme P., Rocha R., Meyer C. F. J., López-Baucells A. (2020). Echolocation of Central Amazonian ‘whispering’ phyllostomid bats: call design and interspecific variation. Mammal Res. 65, 583–597. doi: 10.1007/s13364-020-00503-0
Yoshikura S., Yasui S., Kamijo T. (2011). Comparative study of forest-dwelling bats’ abundances and species richness between old-growth forests and conifer plantations in Nikko National Park, central Japan. Mammal Study 36 (4), 189–198. doi: 10.3106/041.036.0402
Keywords: bat sampling, Chiroptera, communities, ecological indices, guilds, global regions, mitigations, scale
Citation: Xavier BS, Rainho A, Santos AMC, Vieira MV and Carvalho WD (2023) Global systematic map of research on bats in agricultural systems. Front. Ecol. Evol. 11:1214176. doi: 10.3389/fevo.2023.1214176
Received: 29 April 2023; Accepted: 26 September 2023;
Published: 10 October 2023.
Edited by:
Hugo Rebelo, University of Lisbon, PortugalReviewed by:
Maurício Silveira, University of Brasilia, BrazilCopyright © 2023 Xavier, Rainho, Santos, Vieira and Carvalho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bruna Silva Xavier, YnJ1bmF4YXZpZXI2QGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.