 Armando Ferreira-Nuño1*
Armando Ferreira-Nuño1* Larry J. Young2
Larry J. Young2 Adriana Morales Otal1
Adriana Morales Otal1 Francisco Camacho3
Francisco Camacho3 Néstor F. Díaz4
Néstor F. Díaz4 Raúl G. Paredes3,5
Raúl G. Paredes3,5 Alberto Prado5
Alberto Prado5 Antonio Cruz Benites1
Antonio Cruz Benites1 Wendy Portillo3*
Wendy Portillo3*- 1Departamento de Biología de la Reproducción, Área de Neurociencias, Universidad Autónoma Metropolitana-Iztapalapa, Ciudad de México, Mexico
- 2Silvio O. Conte Center for Oxytocin and Social Cognition, Center for Translational Social Neuroscience, Department of Psychiatry and Behavioral Sciences, Emory National Primate Research Center, Emory University, Atlanta, GA, United States
- 3Instituto de Neurobiología, Universidad Nacional Autónoma de México (UNAM), Querétaro, Mexico
- 4Departamento de Fisiología y Desarrollo Celular, Instituto Nacional de Perinatología, Ciudad de México, Mexico
- 5Escuela Nacional de Estudios Superiores, Unidad Juriquilla, Universidad Nacional Autónoma de México (UNAM), Querétaro, Mexico
Introduction: The prairie vole (Microtus ochrogaster) is a socially monogamous species that, after cohabitation with mating, forms an enduring pair bond. It has been suggested that female prairie voles avoid mating with fathers and siblings to prevent inbreeding depression. However, controlled laboratory tests of preferences involving males with varying degree of relatedness have not been conducted.
Methods: To address this, we employed a multiple socio-sexual preference arena consisting of four adjacent cylinders arranged in a closed circle. In each cylinder, we placed a male of varying relatedness to the experimental female (i.e., father, sibling, first-degree cousin, and unrelated males) and registered their behavior for five hours. Male socio-sexual preference was determined by the proportion of time spent in each male's chamber, which can be driven by affiliative preferences for the father and sibling and sexual attraction for the cousin and unrelated males. Mating preference was analyzed as the frequency of mating with each male. We hypothesized that receptive females would show sexual attraction and mating preferences for the unrelated males and cousins and affiliative preferences for the fathers and siblings.
Results: Our analyses showed that females spent more time with first-degree cousins and mated more often with them compared to unrelated males, siblings, or fathers. However, complete inbreeding avoidance was not observed, and some females mated with siblings, fathers, or both.
Discussion: Although our results did not support the hypothesis, they are consistent with other studies that have argued that mating with first-degree cousins optimizes the costs and benefits associated with inbreeding and outbreeding.
1 Introduction
Mate choice is a critical element for the reproductive success of many species that involves context-specific factors (Charlton, 2008). Among the evolutionary benefits of mate choice is the adaptive nature of parental genes inherited by offspring, which can influence fitness (Jennions and Petrie, 2000; Roberts and Gosling, 2003). Also, mating with kin (inbreeding) can reduce offspring fertility due to inbreeding depression (Ralls et al., 1988; Pusey and Wolf, 1996; Crnokrak and Roff, 1999). Thus, choosing the appropriate mate affects an individual’s fitness, particularly in females that have limited reproductive potential.
The prairie vole (Microtus ochrogaster), is a small rodent with a socially monogamous mating system, present only in 5% of mammals, and males actively contribute to the care of the offspring (Kleiman, 1977; Thomas and Birney, 1979; McGraw and Young, 2010; Getz et al., 1981; Walum and Young, 2018). It has been reported in mammals that if males participate in parental care, their contribution goes beyond simple genetic heritage and includes increased offspring survival (McGuire et al., 1992; Gubernick and Teferi, 2000). It has been proposed that mate choice by female prairie voles is a vital task, that may rely on many cues. For instance, females tend to prefer males who engage in more allogrooming and maintain close contact with them (Ophir et al., 2008a).
Unlike genetic monogamy, in which reproduction is limited to only one partner in life, in social monogamy, members of a couple may mate with others, despite having a tight pair bond (Ophir et al., 2008b; Lukas and Clutton-Brock, 2013; Carter and Perkeybile, 2018). For example, studies conducted in seminatural conditions, using paternity analyses, have shown that in situations where males outnumber females, multiple paternity occurs in 57% of pregnant female prairie voles (Rice et al., 2022). As a reproductive system, prairie voles behave as residents and wanderers throughout their lives (Solomon and Jacquot, 2002; Lichter et al., 2020). Indeed, male residents exhibit social monogamy due to pair bonding with the females within their territory and nest site, while wanderers are often single and promiscuous (Getz et al., 1993; Solomon and Jacquot, 2002; Solomon et al., 2004; Ophir et al., 2008b; Rice et al., 2018; Lichter et al., 2020; Solomon and Ophir, 2020),. However, sexual selection is not very strong in socially monogamous species. Social monogamous species typically have minimal sex differences, less intra-specific competition, no biased adult sex ratio, and males do not experience selective pressures for range expansion (Gaulin and Fitzgerald, 1989; Carter and Getz, 1993).
Prairie voles are capable of kin recognition through smell (Newman and Halpin, 1988). For example, in laboratory conditions, sexually naïve females tend to avoid mating with males with whom they were raised (father and siblings) (McGuire and Getz, 1981). Female prairie voles display induced sexual receptivity by exposure to a chemosignal in male urine. Because females usually do not naso-genital sniff familiar males, it has been suggested that under natural conditions, sexual receptivity is induced in prairie vole females by signals from unrelated males (Getz and Carter, 1980; Gavish et al., 1983; McGuire and Getz, 1991). Moreover, females enter sexual receptivity more quickly when exposed to males that do not have prior mating experience (Pierce and Dewsbury, 1991).
Carter et al. (1986) demonstrated that when female voles were mated with a sibling, a father, or in the presence of the father and the mother, their sexual encounters are brief and probably infertile, with a low frequency of lordosis (Getz et al., 2005). These behaviors make it possible to avoid mating with genetically related individuals.
On the other hand, mammals display adverse effects produced by inbreeding depression. For example, in the white-footed mouse (Peromyscus leucopus), it was observed that females that mated with their cousins gave birth to litters of the same size as those of females that mated with unrelated males. In addition, no significant differences were found in the weight of pups at weaning. However, litters obtained from breeding among siblings were small, and the pups’ weight at weaning was significantly lower than that of pups from females that mated with their cousins or unrelated males (Keane, 1990). In prairie voles, mating with siblings from the same litter results in offspring weighing less at birth and weaning, smaller litter sizes, increased intervals between litters, and a decrease in the proportion of pairs that produce litters (Bixler and Tang-Martinez, 2006).
Sexually naïve female prairie voles that mate with their siblings after having been kept together from birth until puberty do not breed under laboratory conditions. However, when siblings are separated after weaning and kept apart for at least 8–15 days before mating, they do breed (McGuire and Getz, 1981; Gavish et al., 1984). These data suggest that sibling affiliation among prairie voles might be due to previous contact or familiarity in early life stages rather than to a mechanism restricted to sensory recognition of genetic kinship, as is predicted by the theory of kin selection (Hamilton 1964a; Hamilton 1964b; Paz y Miño and Tang-Martínez, 1999b). Similarly, alloparental care is not based on kin relationships. Sexually immature male prairie voles performed similar alloparental care toward their siblings and unrelated neonate voles (Finton et al., 2022).
Different paradigms and conditions have been employed to evaluate mate choice in prairie voles. The most common approach is to allow the experimental subject to choose between two individuals of the opposite sex. This choice paradigm has been modified to assess bond strength in prairie voles allowing a subject to choose between a partner and a stranger (Williams et al., 1992; Beery, 2021). Such tests have involved several arena arrangements: rectangular ones with three equal compartments (Ferguson et al., 1986; Pierce and Dewsbury, 1991; Ahern et al., 2009), or arenas shaped in a “Y” (Aragona and Wang, 2004; Curtis, 2010; Shapiro and Dewsbury 1986b), a “T” (Rodriguez et al., 2013), or square (Wolff and Dunlap, 2002; Ophir et al., 2008a). In all these arenas, the experimental female is placed in a compartment where she can access the other sections and choose the stimulus male with whom she wants to mate. Under these conditions, the male she spends more time with and has more sexual encounters with during the test is designated as the preferred male (Ferguson et al., 1986; Pierce and Dewsbury, 1991; Shapiro and Dewsbury 1986b; Shapiro et al., 1986a). Until now, there are no studies using this kind of paradigm to expose sexually receptive females simultaneously with related (father, siblings, or first-degree cousins) or unrelated males to determine socio-sexual and mating preferences. The only study that mated a sexually naïve female prairie vole simultaneously with three males used unrelated adults as stimuli (Wolff and Dunlap, 2002).
Our research group developed a paradigm, the multiple socio-sexual preference test, to assess mate choice in laboratory rats, demonstrating that estrous females prefer one of four males despite being a polygamous species (Ferreira-Nuño et al., 2005). In the present study, we assess the socio-sexual preference by quantifying the time and number of visits in each male’s chamber, which can be driven by affiliative (fathers and siblings) preference and sexual attraction (first-degree cousin and unrelated). Mating preference was obtained by analyzing the number of mounts, intromissions, and ejaculations with each male.
Since prairie voles display several mechanisms to avoid inbreeding depression (Gavish et al., 1983; McGuire and Getz, 1981; McGuire and Getz, 1991; Getz and Carter, 1998), we hypothesized that under the multiple socio-sexual preference test sexually receptive females would exhibit a socio-sexual preference (sexual attraction) and mating preference for the unrelated males and cousins and socio-sexual preference (affiliative) for the fathers and siblings. In addition, we sought to determine whether huddling behavior (side-by-side contact), which usually occurs among prairie voles once the pair bond is established, is a predictive marker with mating preference in this test. Finally, we investigated the temporal development of social interactions with the four males, including how mating and huddling emerged over the five hours of the test under the multiple socio-sexual preference test.
2 Methods
2.1 Animal housing and husbandry
Prairie voles were bred in the vivarium at the Instituto de Neurobiología, at the Universidad Nacional Autónoma de México (UNAM). The colony started with animals kindly provided by Dr. Larry J. Young at Emory University. Prairie voles were housed in 40 × 20 × 20 cm acrylic cages, with pine chip bedding and recycled paper, in a room with a 14/10 L/D cycle (lights were turned on at 08:00 h), at 20–25°C with ad libitum access to water and food (Rabbit high fiber diet 5326 LABDIET, oats, and sunflower seeds). All experiments were performed in accordance with the “Reglamento de la Ley General de Salud en Materia de Investigación para la Salud” of the Mexican Health Ministry and followed the National Institutes of Health guide for the care and use of Laboratory animals. The experiments were approved by the Animal Care Committee of the Instituto de Neurobiología (072) and the Ethics Committee of the Instituto Nacional de Perinatología (212250-3230-21216-05-15).
To obtain the experimental animals, adult male and female prairie voles (3–4 months old) were mated in pairs inside clear acrylic cages with wood shavings for bedding. Once the offspring were born, they were separated at weaning at 21 days of age and housed in home acrylic cages in groups of 3–4 voles of the same sex and age per cage. Voles were marked for identification with a metal ear tag and were kept in these conditions until multiple socio-sexual preference tests were performed. Female and male siblings were separated for more than two months.
2.2 Behavioral procedures
For the socio-sexual and mating preference test, we used the multiple socio-sexual preference arena, with four clear acrylic cylinders arranged in a closed circle side by side, as shown in Figure 1, forming a neutral chamber in the center. Each cylinder had a 45 cm diameter, was 20 cm high, and had an access of 4 × 4 cm hole at the bottom, facing the central chamber. The hole allowed the experimental females to move freely from each cylinder into the central compartment to choose with which male or males to interact. On the day before the experiments, the stimulus males underwent a 30 minutes habituation period in the arena. They were placed and tethered within their respective cylinders using a neck harness made with a 4 mm diameter aquarium airline tubing. This setup allowed the males to move freely inside their cylinders but prevented them from entering each other’s chambers. Females were not present in the arena during male’s habituation.
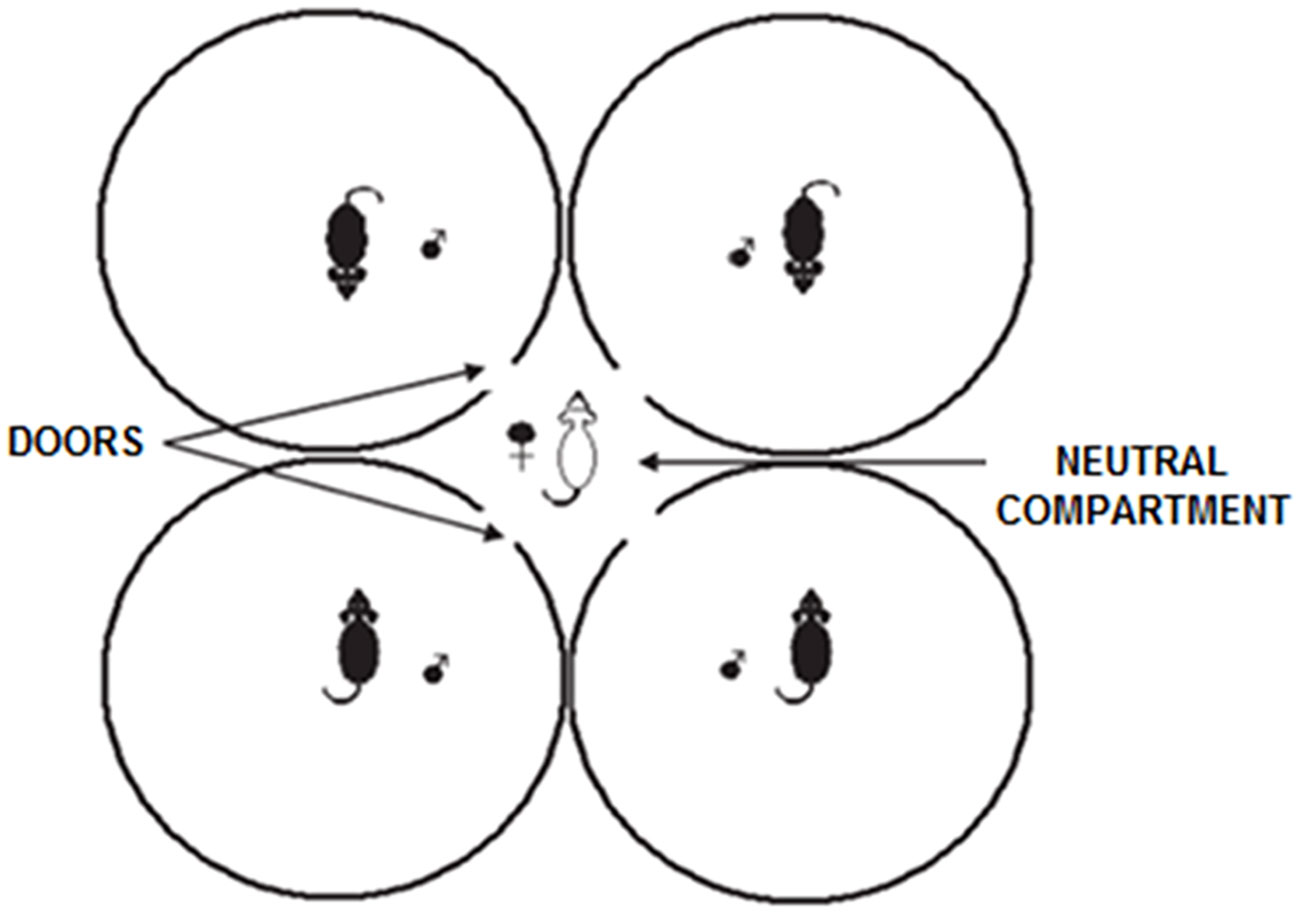
Figure 1 Multiple socio-sexual preference test. The arena consists of four transparent acrylic cylinders arranged in a closed circle side by side, forming a neutral compartment in the center. Each cylinder had an access hole at the bottom that allowed the experimental animal to move freely from each cylinder into the central chamber to choose with which stimulus voles to interact. The stimulus animals were placed and tethered within their respective cylinder; they could move and interact with the experimental subject but could not leave their cylinder.
The females were also habituated to the arena for 30 minutes with free access to the four cylinders without any males present. On the day of the test, both males and females were given five minutes to acclimate to the apparatus before the trial commenced. The position of the four stimulus males was randomized across the test.
In each multiple socio-sexual preference test, one experimental female had the opportunity to interact with four stimulus males: father, sibling, first-degree cousin, and unrelated male. The fathers were 7–9 months old, while the cousins, siblings, and unrelated males were sexually naïve and age-matched to the experimental females (3–4 months old). In total, 20 experimental females were tested, along with 13 fathers, 20 siblings (female’ s litter mates), 20 first-degree cousins (same grandparents), and 20 genetically unrelated males. All fathers were pair-bonded with another female in a mating cage at the time of the multiple socio-sexual preference test.
The experimental females were injected daily, for four consecutive days, with 0.5 µg of estradiol benzoate (Sigma-Aldrich) to induce sexual receptivity (Smale et al., 1985; Roberts et al., 1998). Each mate choice test lasted five hours and was recorded with a Logitech video camera placed on top of the multiple socio-sexual preference arena focusing on the four cylinders. One observer that was placed 1.5 meters from the testing arena, performed the procedures and remained in the experimental room throughout the trial. Multiple socio-sexual preference tests were performed during the light part of the photoperiod cycle. Each video recording was screened, and the following parameters were recorded: each male visited by the females and the frequency and duration of the visits. A female was considered to have visited a male when she entered or placed her front paws and head within the male’s cylinder. We determined the frequency and duration of visits to each male in hourly blocks. To obtain the time spent per visit each hour, the time the female spent during visits was divided by the total number of visits. We also recorded copulatory behavior, including number of mounts, intromissions, and ejaculations (Gray and Dewsbury, 1973); huddling behavior when both members of a pair remained side by side for at least 60 sec; and aggressive behaviors, including biting, boxing, and chasing.
Socio-sexual preference was determined by the proportion of time spent in each male’s chamber, and mating preference was analyzed as the frequency of mating with each male. After the multiple socio-sexual preference test, females that did not copulate were subsequently tested with a sexually experienced male, distinct from the experimental males. This text was conducted immediately, using one of the cylinders of this arena as a testing cage. The aim was to confirm whether these females displayed heightened levels of sexual receptivity in response to copulation.
3 Statistical analysis
Socio-sexual preference for the males was evaluated with chi-squared tests for given probabilities to assess if the time each female spent with males was equivalent. When the chi-squared tests revealed that females were not spending their time homogeneously with the four males, post hoc comparisons were carried out between the 1st and 2nd place stimulus male with a Bonferroni adjustment of the p-values. The percentages of preferred prairie voles and males that mated were analyzed using a chi-squared test for given probabilities. Subsequently, a post hoc test was conducted to compare the first and second-place preference, with p-values adjusted using the Bonferroni method.
Differences in the time spent with the males during the five hours were analyzed with a linear mixed-effect model in which the female was considered a random factor (Pinheiro et al., 2023). The variations in the total number of visits were analyzed with a negative binomial regression using the glm.nb function from the MASS R-package (Venables and Ripley, 2002). Another negative binomial regression was fitted to the variations in the number of visits to all males per hour.
Differences in copulatory events (number of mounts, intromissions, and ejaculations) were analyzed using mixed effects models with a Poisson distribution of errors fitted by maximum likelihood and considering the female as a random factor using the pscl R-package (Zeileis et al., 2008; Zuur et al., 2009). Differences in the time spent huddling and aggressive behaviors during the five hours were analyzed with a linear mixed-effect model in which the female was considered a random factor (Pinheiro et al., 2023). Given that females frequently spent more time with their cousins (the preferred male) and fathers were the only ones with sexual experience, the cousin was considered the baseline level of the factor “male” in all mixed effect models.
Data from the time spent (duration) per visit for each hour to all male compartments did not have a normal distribution and was compared with the Friedman One-Way Repeated Measure Analysis of Variance by Ranks, followed by Student-Newman-Keuls post hoc test.
4 Results
4.1 Socio-sexual preference
Two parameters can be used to determine socio-sexual preference: the time spent with each stimulus male and the number of visits to each male compartment.
4.1.1 Time spent with the four stimulus males
The average time experimental females spent with the stimulus males is shown in Figure 2. Female voles spent more time with their cousin than their sibling and unrelated males. Table 1s shows the negative binomial generalized linear model coefficients.
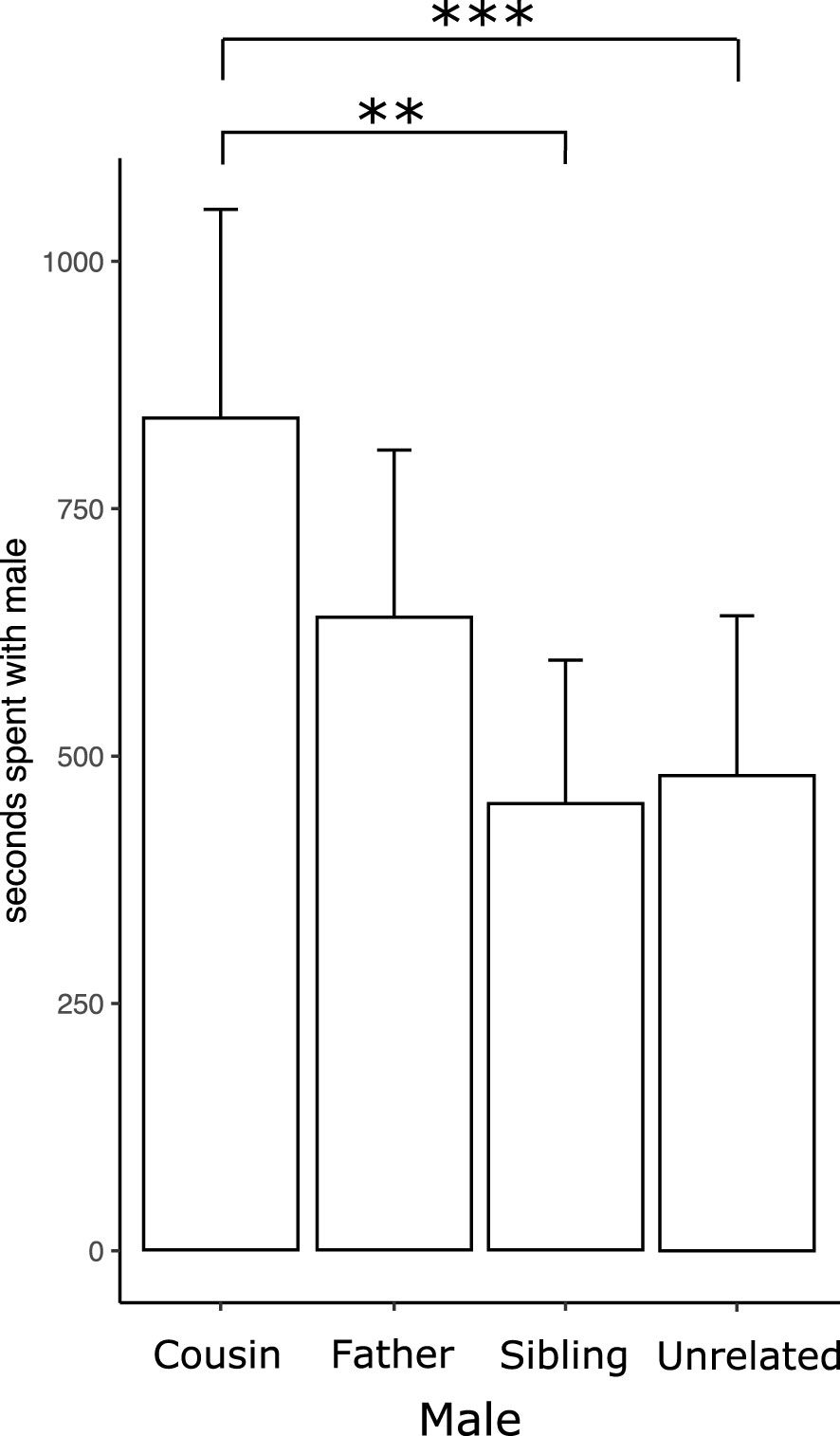
Figure 2 Time experimental females spent with stimulus males (first-degree cousin, father, sibling, and unrelated male) during the five-hour multiple socio-sexual preference test. The bars represent the mean values, and error bars indicate the standard error (**p < 0.01, ***p < 0.001, linear mixed-effect model).
To better assess the female’s socio-sexual preference throughout the test, Figure 3A shows the proportion of time spent by the experimental females each hour with the four stimulus males. Data from the total proportion of time spent with males showed that 14 females preferred a male. Of these seven females (50%) preferred the first-degree cousin, four females preferred the father (28.6%), two preferred the sibling (14.3%), and one preferred the unrelated male (7.1%). Significant differences were found in the percentage of females that preferred a male (chi-squared=600, df=3, p<0.0001). The post hoc test (chi-squared=7.44, df=1, Bonferroni-corrected p=0.013) revealed variations in the percentage of females that preferred the cousin (1st place) compared to those that chose the father (2nd place) Figure 3B.
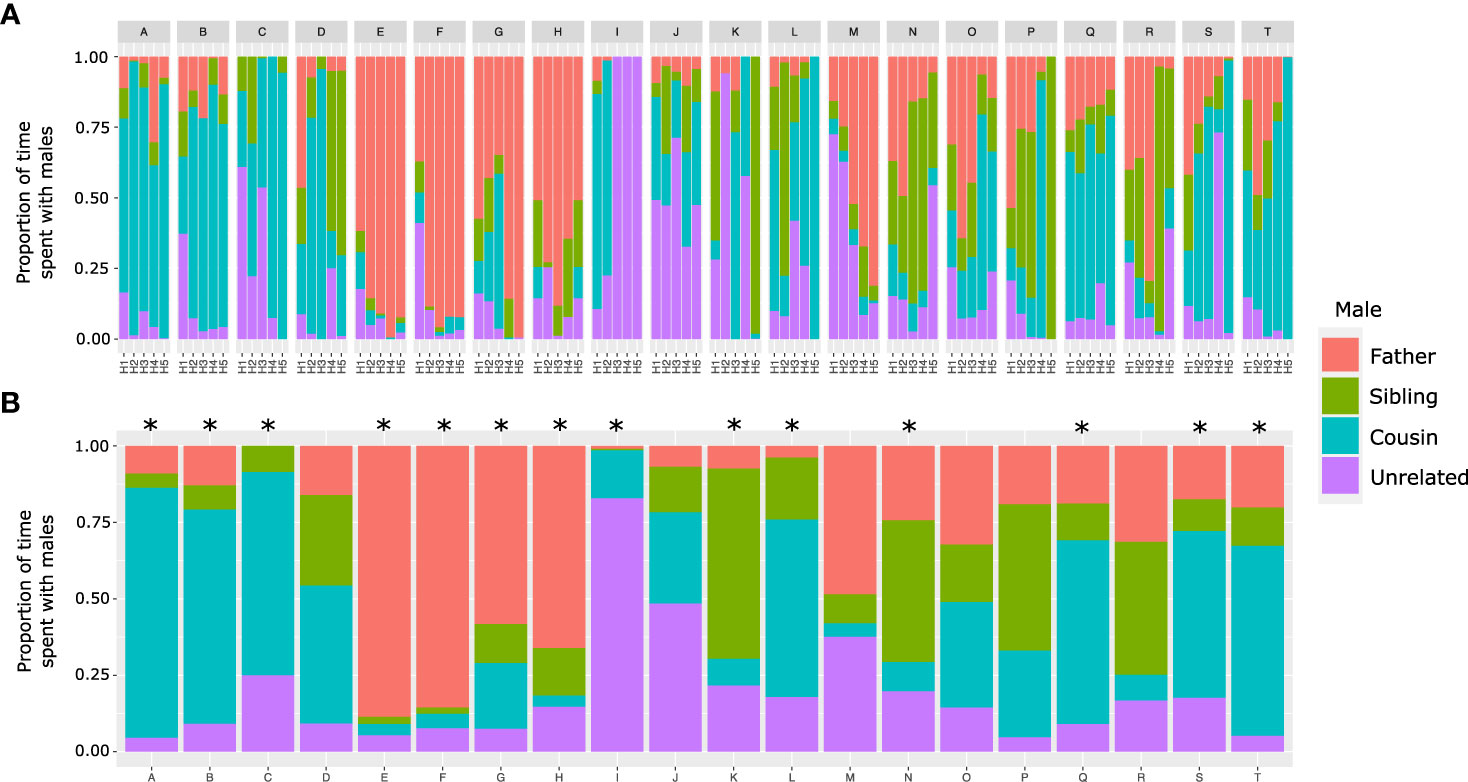
Figure 3 Proportion of time experimental females spent with stimulus males (father, sibling, first-degree cousin, and unrelated) during the multiple socio-sexual preference test lasting five hours (H1–H5). (A) Hourly time spent with males. (B) Total time spent with males. Asterisks indicate chi-squared post hoc comparisons for given probabilities between the 1st and 2nd place males (*p < 0.05). Females that preferred father (E–H), sibling (K, N), cousin (A–C, L, Q, S and T), and unrelated male (I).
4.1.2 Frequency and duration of visits
Figure 4A shows, by the hour, the mean total number of visits by the females to the males throughout the test. Females visited the four stimulus males significantly more frequently during the first hour (Table 2s). This parameter indicates the female’s willingness to explore each male compartment during the first hour to choose the preferred male. No significant differences were found in the number of visits to the stimulus males (Figure 4B). Table 2s shows the negative binomial generalized linear model coefficients for the variations in the total number of visits and Table 3s for the number of visits made to the different males.
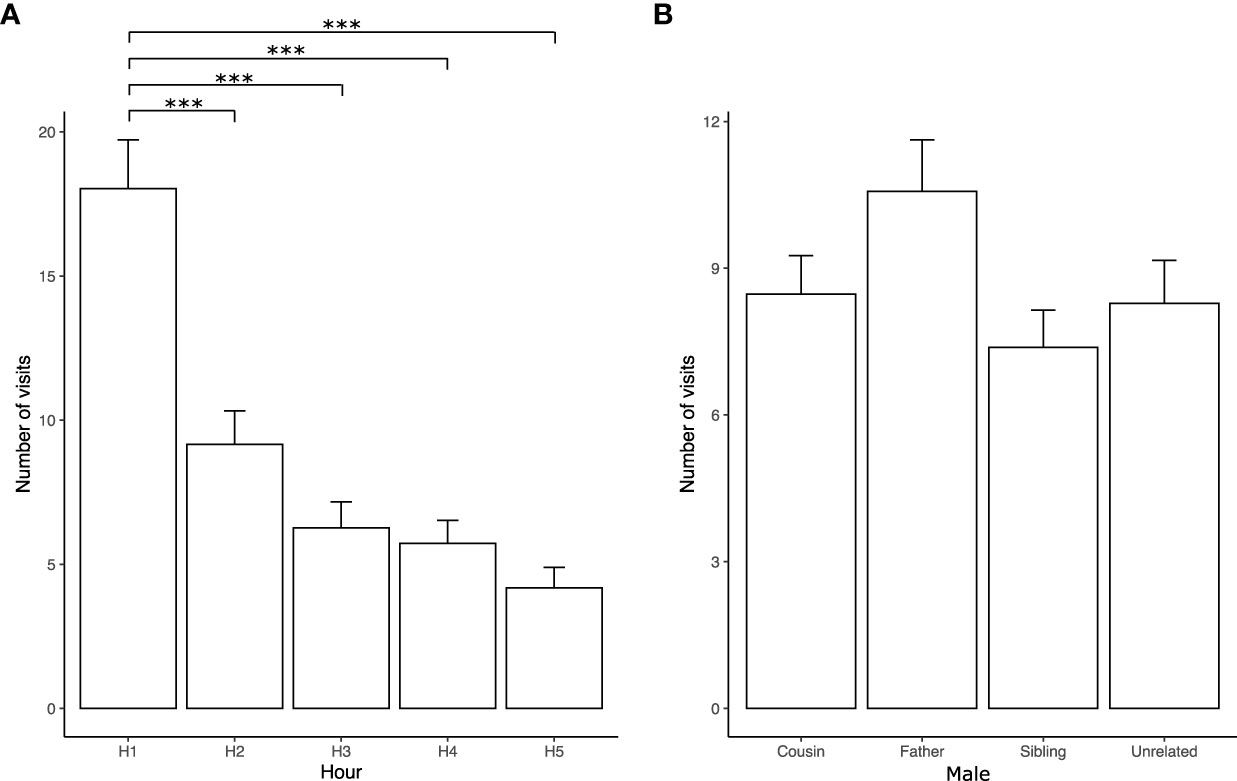
Figure 4 Number of visits to stimulus males. (A) Number of visits per hour to all males. (B) Number of times females visited the stimulus males (first-degree cousin, father, sibling, and unrelated) during the five-hour multiple socio-sexual preference test (H1–H5). The bars represent the mean values, and error bars indicate the standard error (***p < 0.0001, negative binomial regressions).
Significant differences (chi-square=48.1, 4df, p<0.001) were found in the time spent per visit (Table 1). The time spent per visit increases by the hour (except for the third hour vs. the fourth hour).
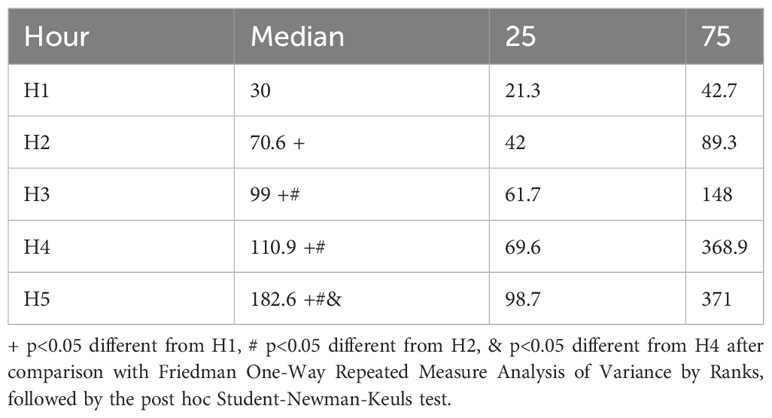
Table 1 Time spent per visit (median and 25 and 75 interquartile ranges) per hour, made by 20 females to stimulus males during the multiple socio-sexual preference test.
4.2 Mating preference
Table 2 presents the results of sexual behavior and huddling during the multiple socio-sexual preference test. Out of the 20 females, 16 (80%) engaged in copulation during the multiple socio-sexual preference test. Among these, eight (50%) mated exclusively with one of the males, who was also their socio-sexual preferred male (except one female; ID “N”). Six females (37.5%) copulated with two males, while two females (12.5%) mated with three males. Therefore, the majority of female prairie voles copulated with only one of the four stimulus males in the multiple socio-sexual preference arena. Of the 14 females that preferred a male, eight females mated with the cousin, four with the father, three with the unrelated male, and three copulated with the sibling. Statistical analysis revealed that the cousin was the preferred male for mating (chi-squared=377.78, df=3, p<0.0001). The post hoc test further confirmed the difference between the number of females that copulated with the cousin (1st place) and those that copulated with the father (2nd place) after Bonferroni correction (chi-squared 11.1, df=1, Bonferroni corrected p=0.0017) (Table 2).
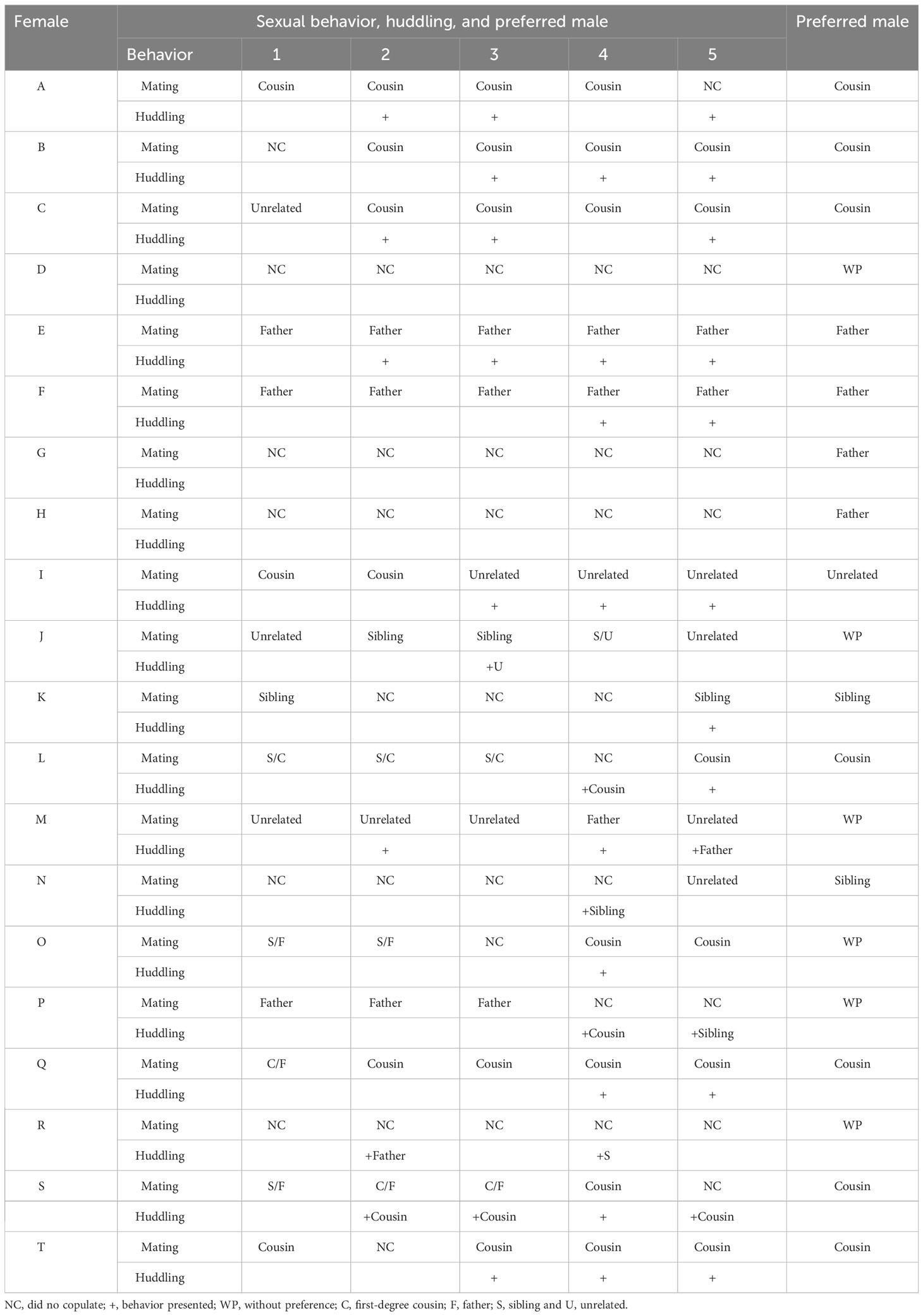
Table 2 Sexual behavior, huddling, and socio-sexual preferred male in female prairie voles during each hour of the multiple socio-sexual preference test.
Sexual parameters showed that cousins displayed more mounts, intromissions, and ejaculations than the sibling and unrelated males and more intromissions than the fathers (Figures 5A–C and Table 4s).
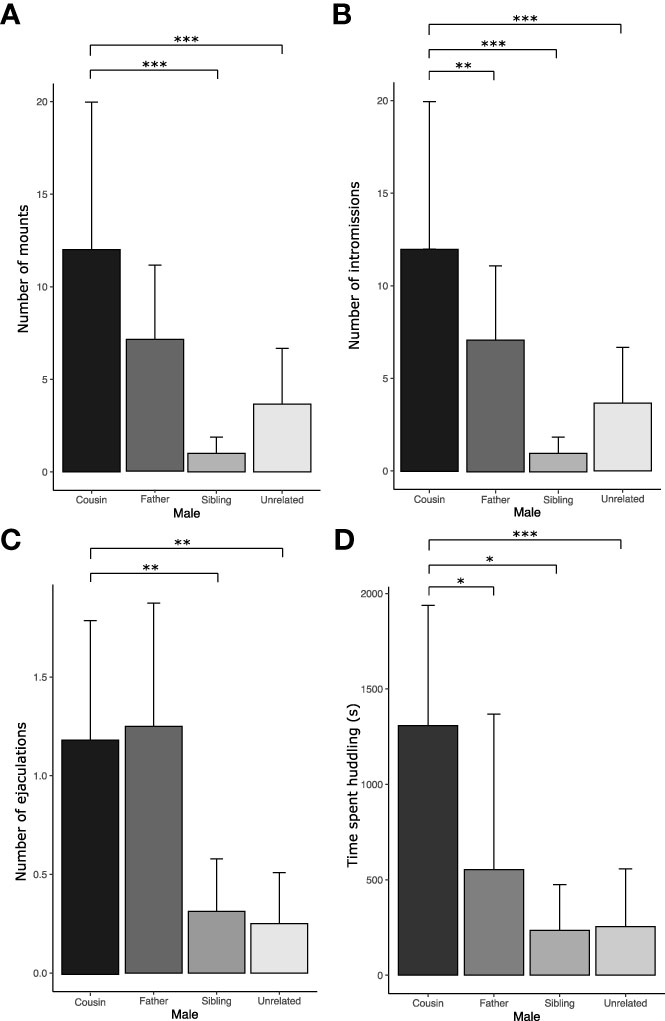
Figure 5 Sexual behavior during the multiple socio-sexual preference test. (A) Number of mounts. (B) Number of intromissions. (C) Number of ejaculations. (D) Time spent huddling, in seconds. The bars represent the mean values, and the error bars represent the standard error (** p<0.01, *** p<0.0001; linear mixed-effect models with Poisson distribution for (A–C), linear mixed-effect model for (D)). Nine females mated with their cousins, seven with their fathers, five with their brothers, and five with unrelated males. Since huddling is an affiliative behavior, this parameter was analyzed only in females who had a socio-sexual preference for a male (n=14).
As for huddling, this behavior only occurred with the females that mated during the test (except female R), and this behavior was only displayed with the socio-sexual preferred male, even with females that copulated with two males (Table 2). Interestingly, significant differences were found in the time spent huddling. Females spent less time huddling with fathers, siblings, and unrelated males than with cousins (Figure 5D and Table 4s). Given the majority of females that copulated during the test huddled with the socio-sexual preferred male, huddling serve as a valuable indicator for determining the preferred male in the multiple socio-sexual preference test. Interestingly, even though four females chose the father as the socio-sexual preferred male, two of them did not copulate with him, suggesting that this may be a filial social preference rather than a sexual preference. Aggressive behaviors also were recorded during the multiple socio-sexual preference test. Fathers were the males who displayed more aggressive behaviors (biting, boxing, and chasing) toward the females (Figure 6, Table 5s).
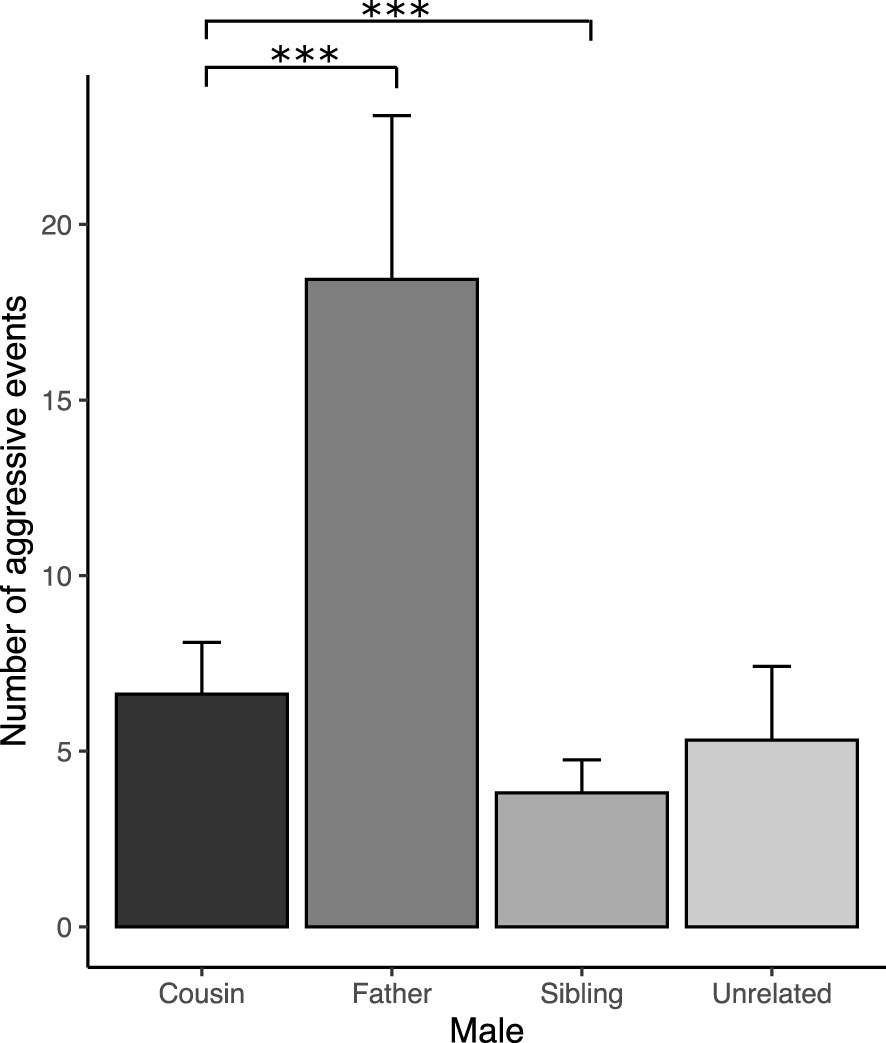
Figure 6 Number of aggressive events of stimulus males (cousin, father, sibling, and unrelated) towards females during the multiple socio-sexual preference test. The bars represent the mean values, and the error bars represent the standard error (*** p<0.001; linear mixed-effect models with Poisson distribution).
At the end of the test, the four females that did not copulate after five hours in the multiple socio-sexual preference arena were evaluated in the home cage of a sexually experienced male who did not participate in this study to determine if they were sexually receptive. The females did show sexual receptivity (lordosis) in response to the male mount.
5 Discussion
Socio-sexual preference, evaluated as the time spent with each stimulus male, revealed that the females spent more time with their cousins than their siblings and unrelated males (Figure 2, Table 1s). Additionally, a higher percentage of females selected their cousin as the socio-sexual preferred male (Figure 3B).
Regarding mating preference, we observed that among the 14 females who preferred a male, most mated with their cousin (8 females) (Table 2). Furthermore, we found that cousins exhibited more mounts, intromissions, and ejaculations than siblings and unrelated males. Notably, cousins also displayed more intromission than fathers (Figure 5). Females who preferred a male also spent more time engaged in huddling behavior with their cousins as opposed to their fathers, siblings, and unrelated males.
We believe the degree of kinship could have significantly influenced the females’ preference for the cousin. Unlike the father, the three other males (sibling, cousin, and stranger) had similar characteristics: they were the same age and had no previous sexual experience. In most studies that suggest that prairie voles avoid inbreeding (Getz and Carter, 1980; McGuire and Getz, 1981; Gavish et al., 1983; McGuire and Getz, 1991), parents have been paired with daughters and sons to demonstrate incest avoidance, but only a few studies have carried out pairings among cousins. However, none of these studies assessed the sexually naïve females’ preference for the cousin using a multiple socio-sexual preference test. For example, under laboratory and seminatural conditions, studies have conducted inbreeding among cousins to assess its effect on inbreeding depression, as well as on the survival and reproductive success of the offspring (Barnard and Fitzsimons, 1988; Keane, 1990; Pillay and Rymer, 2017).
Mating among individuals with an intermediate kinship has been described for some rodent species that tend to reject reproduction with related congeners to avoid inbreeding depression, as is the case of the mouse (Mus musculus), (Barnard and Fitzsimons, 1988) and the white-footed mouse (Peromyscus leucopus), (Keane, 1990). For example, Barnard and Fitzsimons (1988), using a cross-fostered system in the house mouse, assessed preference as a function of relatedness with opposite sex individuals, including full siblings, half-siblings, cousins in the first, and second-degree cousins. These authors demonstrated that males preferred the odor and interaction of females of intermediate kinship (cousins). Something similar was demonstrated for the white-footed mouse; females in estrus spent more time sniffing soiled bedding from cousins in the first-degree than soiled bedding from siblings or unrelated males, and the females also interacted longer with their cousins than with the sibling or an empty compartment. In addition, females in estrus showed more amicable behaviors and were less antagonistic or aggressive with their cousins than with siblings and unrelated males (Keane, 1990). Finally, in the whistling rat (Paratomys littledalei), it was also observed that the time spent by inbred females in estrus sniffing the wood-shaving beddings of their first-degree cousins was similar to the time spent sniffing the unrelated males’ bedding. Time sniffing the unrelated male’s odors was significantly longer than the time dedicated to smell full siblings of the same litter, or half siblings bedding (Pillay and Rymer, 2017). The above suggests that the females’ levels of preference for the cousins and their reproductive success could be similar to unrelated males.
We anticipated that sexually naive female voles would be demonstrate a preference for the unrelated (stranger) males. Surprisingly, only one chose the unrelated vole as the socio-sexual preferred male. Outbreeding can be disadvantageous due to the breakdown of locally adapted or co-adapted gene complexes (Tregenza and Wedell, 2000). Individuals with a sexual preference for conspecifics of intermediate relatedness had an optimal balance between inbreeding and outbreeding (Bateson, 1982; Loyau et al., 2012). Male and female Japanese quail spent more time with the first-degree cousin than with familiar and novel siblings. Also, they prefer the cousin over the unrelated bird (Bateson, 1982). Loyau et al. (2012) demonstrated that female fruit flys (D. melanogaster) mate more often with their bothers than with unrelated males.
Our hypothesis was that females would exhibit socio-sexual preference (affiliative) for the father and sibling but not mating preferences. Our results suggested that female prairie voles did not avoid incest in the multiple socio-sexual preference test. Four females showed socio-sexual preference for their father, and two chose their siblings. In mating preference, four females mate with their fathers and three with their siblings. These findings disagree with previous studies that demonstrated that females prevent inbreeding depression by rejecting sexual contact with the father or the siblings under laboratory and seminatural conditions (Getz and Carter, 1980; Gavish et al., 1983; McGuire and Getz, 1991).
As expected, females spend less time with their siblings, and only a few mate with them. Only two sexually naïve female prairie voles chose the sibling as the socio-sexual preferred male (Figure 3B; Table 2), even though subjects were separated from their siblings for two months, from weaning until the test was conducted. Moreover, females spent less time with their siblings and avoided huddling and copulation (Figures 2 and 5).
McGuire and Getz (1981) and Gavish et al. (1984) reported that under laboratory conditions that do not allow the female to choose between different males, sexually naïve female prairie voles will copulate and reproduce with their siblings, as long as they have been separated since weaning (21 days of age) and kept apart for at least 15 days before mating. Getz and Carter (1998) suggested that when female prairie voles are separated and isolated from their siblings, they are incapable of kin recognition.
Thus, incest avoidance would be mainly explained by the familiarity produced by the previous contact with the siblings during the nursing period rather than by genetic kinship (Getz and Carter, 1998). However, in this multiple socio-sexual preference, our results suggest that genetic kinship could still be detected by the female prairie vole and thus prevented them from mating despite being separated from their siblings from weaning to the moment of testing. In the studies performed by McGuire and Getz (1981) and Gavish et al. (1983), female prairie voles copulated and mated with a sibling because he was the only available male, whereas, in our case, the female may have rejected the sibling because she had access to three other males in the multiple socio-sexual preference arena. In nature, the situation is even more complex, with much more space to occupy. The avoidance we observed in our study may be sufficiently robust to avoid sibling incest in a natural environment. In our study, the female voles could distinguish and avoid mating with siblings among the various stimulus males, with only a few females choosing to mate with them, suggesting that a combination of smell, visual and auditory cues may be involved in their affiliations.
In addition, mate with a sibling can decrease offspring survival. Using establishing seminatural confinements (0.1 hectares) with related (opposite-sex siblings) or unrelated breeding pairs of adult prairie voles, Lucia-Simmons and Keane (2015) demonstrated that inbreeding produced a decrease in the survival of the young in the first three generations. However, after three generations, the number of litters conceived by the related males and unrelated males was similar (Lucia-Simmons and Keane, 2015). This outcome is explained because the males born from endogamic breeding voles stayed close to the nest where they were raised, participating in the care and protection of the young that were bred afterward, which increased their survival (Lucia-Simmons and Keane, 2015).
Another result in this study was that four out of the 20 females chose the father as the socio-sexual preferred male (Figure 3B, Table 2). Mating preference data showed that only two of these four females copulated with the father (Table 2). In the multiple socio-sexual preference test, females spent more time with a male (socio-sexual preference), but they did not necessarily mate with him (mating preferences). Hence, a mating preference of the female prairie vole in the multiple socio-sexual preference arena was not necessarily associated with the sexual interest to copulate with a male.
Our results could be explained if we consider that the father was pair-bonded with another female in a mating cage during the multiple socio-sexual preference test. Therefore, he was the only one with different traits (sexually experienced, older, aggressiveness) that might have influenced the females’ mate choice. Female prairie voles avert incest by avoiding olfactory exploration of the father and the sibling (McGuire and Getz, 1981), but the fathers can also reject being sniffed by the daughters. For example, in studies conducted with family groups of this species, in larger enclosures, the fathers do not try to mount their daughters (McGuire and Getz, 1991).
The fathers’ aggressiveness likely also had an influence on preference in our study; attacks did occur in most cases when the female visited the father and started to interact with him (Figure 6). One of the most remarkable behaviors in this monogamous species is rejection and attacks by a member of a mated pair towards others of their kind once pair bonding is established (Getz and Carter, 1980). Thus, it is not rare that the fathers were the males that most attacked the experimental females, given that they were mated with another female at the testing time. In accordance, it has been reported that female voles did not show a preference for aggressive males (Ophir et al., 2008a).
As for social status, in dyadic encounters, sexually naïve receptive females avoid sniffing and copulatory activity with males that are already mated with another female at the time of testing and instead prefer to copulate with unmated males (Pierce and Dewsbury, 1991). Also, it was shown that breeder males tried to mount the virgin females fewer times than sexually experienced non-paired males (Getz et al., 1981). In our study, these factors might have adversely influenced the fathers since only two females mated with their fathers.
On the other hand, fathers exhibited characteristics that could have influenced female preference, such as dominance and large size, which male prairie voles tend to have with older age. For example, sexually naïve female prairie voles prefer to copulate with large males (Aschemeier et al., 2008; Solomon, 1993), and sexually experienced females prefer males of higher social status or dominance (Shapiro and Dewsbury, 1986b), traits that possibly characterize the fathers in our study.
5.1 Frequency and duration of visits
In our study, females spent more time with their cousins than siblings and unrelated males in the multiple socio-sexual preference test (Figure 2, Table 1s). Females showed variable exploratory activity since the number of visits to the males was higher in the first hour of testing compared to hours 2–5 (see Figure 4A, Table 2s). This parameter indicates the female’s willingness to explore each male compartment during the first hour to choose the socio-sexual preferred male. It must be noted that, despite this decrease, most of the experimental females continued visiting the four stimulus males throughout the test. No significant differences were found in the number of visits to each male. Therefore, the time spent with the males determines socio-sexual preferences in our test conditions.
The time spent per visit was significantly shorter during the first hour (Table 1) compared to the second through fifth hours. The intense exploratory activity at the beginning of the test is possibly helpful for the female to spatially identify where the four males are located in the cylinders. This behavior also contributes to sampling the males. Indeed, intense exploratory activity and a high number of visits during the first minutes have also been observed in the female rat when mated in a multiple socio-sexual preference arena with four males (Ferreira-Nuño et al., 2005).
5.2 Mating preferences
In our study, 16 of the 20 females mated with at least one stimulus male. Analysis of the copulatory parameters showed that the cousins displayed more mounts, intromissions, and ejaculations than their siblings and unrelated males, and they performed more intromissions than the fathers.
The percentages of those that copulated with only one and two males were 50% and 37.5%, respectively. Thus, under the multiple socio-sexual preference test, mating with only one male was the most common pattern of female mating behavior (Table 2). These results differ from other studies in which one female was mated simultaneously with three males under laboratory conditions. One of the studies used nulliparous females (Wolff and Dunlap, 2002), while the other study separated females from their sexual partners for 30 days prior to testing after their first time giving birth (Wolff et al., 2002). In both cases, most females (55%) copulated with more than one male; in the studies by Wolf and collaborators, mating sessions lasted 24 hours (Wolff and Dunlap, 2002; Wolff et al., 2002), while in our study, they lasted five hours. Furthermore, as the duration of the tests increased, the number of males with whom the females copulated also increased. During the first two hours, only 7% of the females copulated with more than one male, while 18% mated with two males and 25% mated with three males in 24 hours (Wolff et al., 2002). It could be that socially monogamous female prairie voles copulate with several males before choosing a mate if these males are found in the immediate surroundings.
5.3 Huddling behavior reflects a mating preference
Both in dyadic encounters and in preference tests, huddling increases selectively in male–female couples with a history of cohabitation and/or mating (Williams et al., 1992). In our study, only the females that mated also huddled with the socio-sexual preferred male (Table 2), suggesting that huddling in the prairie vole is a clear indication of socio-sexual and mate preference for a male (Williams et al., 1992; Winslow et al., 1993; Carter et al., 1995; Insel and Hulihan, 1995; Lim et al., 2004; Young and Wang, 2004). When the time spent huddling was analyzed, we found that females spent more time huddling with their cousins than with their fathers, siblings, and unrelated males (Figure 2). Taken together, our data indicate that females demonstrate both ad overall preferences to affiliate and mate with first-degree cousins.
5.4 Conclusion
Using the multiple socio-sexual preference test, we found that sexually naïve females spent more time with their cousins; a higher percentage of females chose their cousin as the preferred male and spent more time huddling with them. Although the sexually naïve female voles did not show absolute inbreeding avoidance, socio-sexual preferences for siblings was low. Moreover, cousins performed more mounts, intromissions, and ejaculations than siblings and unrelated males. These data are consistent with other studies that have indicated that mating with first-degree cousins represent an optimal balance between the cost and benefits of inbreeding and outbreeding (Bateson, 1982; Tregenza and Wedell, 2000; Loyau et al., 2012). We also found that females only displayed huddling behavior with the males with whom they mated, which were also their socio-sexual preferred males. This suggests that this behavior is a relevant sign to identify that mate choice has occurred.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
All experiments were performed in accordance with the “Reglamento de la Ley General de Salud en Materia de Investigación para la Salud” of the Mexican Health Ministry and followed the National Institutes of Health guide for the care and use of Laboratory animals. The experiments were approved by the Animal Care Committee of the Instituto de Neurobiología (072) and the Ethics Committee of the Instituto Nacional de Perinatología (212250-3230-21216-05-15). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
AF-N conceptualization, methodology, formal analysis, investigation, writing original draft and review and editing; LY conceptualization, writing review and editing, funding acquisition; AM conceptualization, methodology, formal analysis; FC takes care of our animal colony, methodology; ND proper analysis, writing review and editing, founding acquisition; RP conceptualization, writing review and editing, funding acquisition; AP statistical analysis; AC methodology, formal analysis; WP conceptualization, writing original draft and review and editing, funding acquisition. All authors contributed to the article and approved the submitted version.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was supported by grants UNAM-DGAPA-PAPIIT IN202818, IN208221 and IN206521, INPER 212250-3230-21216-05-15, 2018-1-163, 2022-1-13, CF-2023-G-206 CONAHCYT, and NIH grants P51OD011132 and P50MH100023.
Acknowledgments
We thank Deisy Gasca, Martín García, Alejandra Castilla, Paula Bozo, and Jessica Norris for their excellent technical assistance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1154800/full#supplementary-material
References
Ahern T. H., Modi M. E., Burkett J. P., Young L. J. (2009). Evaluation of two automated metrics for analyzing partner preference tests. J. Neurosci. Methods 182 (2), 180–188. doi: 10.1016/j.jneumeth.2009.06.010
Aragona B. J., Wang Z. (2004). The prairie vole (Microtus ochrogaster): an animal model for behavioral neuroendocrine research on pair bonding. ILAR J. 45 (1), 35–45. doi: 10.1093/ilar.45.1.35
Aschemeier L. M., Keane B., Solomon N. G. (2008). Body size has a greater influence on female mate choice than length of microsatellite region of the avpr1a gene in male prairie voles. J. Young Investigators 18.
Barnard C. J., Fitzsimons J. (1988). Kin recognition and mate choice in mice: the effects of kinship, familiarity and social interference on intersexual interaction. Anim. Behav. 36, 1078–1090. doi: 10.1016/S0003-3472(88)80067-8
Beery A. K. (2021). Familiarity and mate preference assessment with the partner preference test. Curr. Protoc. 1, e173. doi: 10.1002/cpz1.173
Bixler A., Tang-Martinez Z. (2006). Reproductive performance as a function of inbreeding in prairie voles (Microtus ochrogaster). J. Mammal. 87, 944–949. doi: 10.1644/05-MAMM-A-353R2.1
Carter C. S., DeVries A. C., Getz L. L. (1995). Physiological substrates of mammalian monogamy: the prairie vole model. Neurosci. Biobehav. Rev. 19, 303–314. doi: 10.1016/0149-7634(94)00070-h
Carter S. C., Getz L. L. (1993). Monogamy and the prairie vole. Sci. Am. 268 (6), 100–106. doi: 10.1038/scientificamerican0693-100
Carter C. S., Getz L. L., Cohen-Parsons M. (1986). “Relationships between social organization and behavioral endocrinology in a monogamous mammal,” in Advances in the Study of Behavior, vol. 16 . Eds. Rosenblat J. S., Beer C., Busnel M. C., Slater P. J. B. (New York, NY: Academic Pres), 109–145. doi: 10.1016/S0065-3454(08)60189-8
Carter C. S., Perkeybile A. M. (2018). The monogamy paradox: what do love and sex have to do with it? Front. Ecol. Evol. 6. doi: 10.3389/fevo.2018.00202
Charlton B. D. (2008). “Female mate choice in nonhuman mammals,” in Animal Behavior: New Research. Eds. Weber E. A., Krause L. H. (New York: Nova Science Publishers, Inc), 35–56.
Crnokrak P., Roff D. (1999). Inbreeding depression in the wild. Heredity 83, 260–270. doi: 10.1038/sj.hdy.6885530
Curtis J. T. (2010). Female prairie vole mate-choice is affected by the males' birth litter composition. Physiol. Behav. 101 (1), 93–100. doi: 10.1016/j.physbeh.2010.04.020
Ferguson B., Fuentes S. M., Sawrey D. K., Dewsbury D. A. (1986). Male preferences for unmated versus mated females in two species of voles (Microtus ochrogaster and Microtus montanus). J. Comp. Psychol. 100, 243–247. doi: 10.1037/0735-7036.100.3.243
Ferreira-Nuño A., Morales-Otal A., Paredes R. G., Velázquez-Moctezuma J. (2005). Sexual behavior of female rats in a multiple-partner preference test. Horm. Behav. 47 (3), 290–296. doi: 10.1016/j.yhbeh.2004.11.012
Finton C. J., Kelly A. M., Ophir A. G. (2022). Support for the parental practice hypothesis: Subadult prairie voles exhibit similar behavioral and neural profiles when alloparenting kin and non-kin. Behav. Brain Res. 417, 113571. doi: 10.1016/j.bbr.2021.113571
Gaulin S. J. C., Fitzgerald R. W. (1989). Sexual selection for spatial-learning ability. Anim. Behav. 37, 322–331. doi: 10.1016/0003-3472(89)90121-8
Gavish L., Carter C. S., Getz L. L. (1983). Male-female interactions in prairie voles. Anim. Behav. 31, 511–517. doi: 10.1016/S0003-3472(83)80073-6
Gavish L., Hofmann J. E., Getz L. L. (1984). Sibling recognition in the prairie vole, Microtus ochrogaster. Anim. Behav. 32, 362–366. doi: 10.1016/S0003-3472(84)80269-9
Getz L. L., Carter C. S. (1980). Social organization in Microtus ochrogaster populations. Biologist. 62, 56–69.
Getz L. L., Carter S. C. (1998). Inbreeding avoidance in the prairie vole, Microtus ochrogaster. Ethol Ecol. Evol. 2, 115–127. doi: 10.1080/08927014.1998.9522861
Getz L. L., Carter C. S., Gavish L. (1981). The mating system of the prairie vole, Microtus ochrogaster: Field and laboratory evidence for pair-bonding. Behav. Ecol. Sociobiol. 8, 189–194. doi: 10.1007/BF00299829
Getz L. L., McGuire B., Carter C. S. (2005). Social organization and mating system of free-living prairie voles Microtus ochrogaster: A review. Act Zool Sinca 51 (2), 178–186.
Getz L. L., McGuire B., Pizzuto T., Hofmann J. E., Frase B. (1993). Social organization of the prairie vole (Micrrotus ochrogaster). J. Mammal 74 (1), 44–58. doi: 10.2307/1381904
Gray G. D., Dewsbury D. A. (1973). A quantitative description of copulatory behavior in prairie voles (Microtus ochrogaster). Brain Behav. Evol. 8, 437–452. doi: 10.1159/000124368
Gubernick D. J., Teferi T. (2000). Adaptive significance of male parental care in a monogamous mammal. Proc. Biol. Sci. 267 (1439), 147–150. doi: 10.1098/rspb.2000.0979
Hamilton W. D. (1964a). The genetical evolution of social behaviour. I. J. Theoret Biol. 7 (1), 1–16. doi: 10.1016/0022-5193(64)90038-4
Hamilton W. D. (1964b). The genetical evolution of social behaviour. II. J. Theoret Biol. 7 (1), 17–52. doi: 10.1016/0022-5193(64)90038-4
Insel T. R., Hulihan T. J. (1995). A gender-specific mechanism for pair bonding: oxytocin and partner preference formation in monogamous voles. Behav. Neurosci. 109 (4), 782–789. doi: 10.1037/0735-7044.109.4.782
Jennions M. D., Petrie M. (2000). Why do females mate multiply? A review of the genetic benefits. Biol. Rev. Camb Philos. Soc 75 (1), 21–64. doi: 10.1017/s0006323199005423
Keane B. (1990). The effect of relatedness on reproductive success and mate choice in the white-footed mouse, Peromyscus leucopus. Anim. Behav. 39, 264–273. doi: 10.1016/S0003-3472(05)80870-X
Lichter J. B., Lambert C. T., Solomon N. G., Keane B. (2020). Breeding patterns of female prairie voles (Microtus ochrogaster) displaying alternative reproductive tactics. J. Mammal 101 (4), 990–999. doi: 10.1093/jmammal/gyaa058
Lim M. M., Wang Z., Olazabal D. E., Ren X., Terwilliger E. F., Young L. J. (2004). Enhanced partner preference in a promiscuous species by manipulating the expression of a single gene. Nature 429, 754–757. doi: 10.1038/nature02539
Loyau A., Cornuau J. H., Clobert J., Danchin E. (2012). Incestuous Sisters: Mate Preference for Brothers over Unrelated Males in Drosophila melanogaster. PLoS One 7 (122), 1–6. doi: 10.1371/journal.pone.0051293
Lucia-Simmons K. E., Keane B. (2015). Behavioral plasticity in nest residency compensates for inbreeding depression in male prairie voles. Behav. Ecol. 26, 1060–1070. doi: 10.1093/beheco/arv053
Lukas D., Clutton-Brock T. H. (2013). The evolution of social monogamy in mammals. Science 341 (6145), 526–530. doi: 10.1126/science.1238677
McGraw L. A., Young L. J. (2010). The prairie vole: an emerging model organism for understanding the social brain. Trends Neurosci. 33 (2), 103–109. doi: 10.1016/j.tins.2009.11.006
McGuire M. R., Getz L. L. (1981). Incest taboo between sibling Microtus ochrogaster. J. Mammal. 62, 213–215. doi: 10.2307/1380502
McGuire B., Getz L. L. (1991). Response of young female prairie voles Microtus ochrogaster to nonresident males: implications for population regulation. Can. J. Zool. 69, 1 348–1 355. doi: 10.1139/z91-190
McGuire B., Russell K. D., Mahoney T., Novak M. (1992). The effects of mate removal on pregnancy success in prairie voles (Microtus ochrogaster) and meadow voles (Microtus pennsylvanicus). Biol. Reprod. 47, 37–42. doi: 10.1095/biolreprod47.1.37
Newman K., Halpin Z. T. (1988). Individual odours and mate recognition in the prairie vole, Microtus ochrogaster. Anim. Behav. 36, 1779–1787. doi: 10.1016/S0003-3472(88)80117-9
Ophir A. G., Crino O. L., Wilkerson Q. C., Wolff J. O., Phelps S. M. (2008a). Female-directed aggression predicts paternal behavior, but female prairie voles prefer affiliative males to paternal males. Brain Behav. Evol. 71, 32–40. doi: 10.1159/000108609
Ophir A. G., Phelps S. M., Sorin A. B., Wolff J. O. (2008b). Social but not genetic monogamy is associated with greater breeding success in prairie voles. Anim. Behav. 75, 1143–1154. doi: 10.1016/j.anbehav.2007.09.022
Paz y Miño G., Tang-Martínez Z. (1999b). Social interactions, crossfostering, and sibling recognition in prairie voles, Microtus ochrogaster. Can. J. Zool. 77, 1631–1636. doi: 10.1139/z99-109
Pierce J. D., Dewsbury D. A. (1991). Female preferences for unmated versus mated males in two species of voles (Microtus ochrogaster and Microtus montanus). J. Comp. Psychol. 105, 165–171. doi: 10.1037/0735-7036.105.2.165
Pillay N., Rymer T. (2017). Preference for outbreeding in inbred littledale's whistling rats parotomys littledalei. Evol. Biol. 44, 21–30. doi: 10.1007/s11692-016-9383-6
Pinheiro J., Bates D., R Core Team. (2023). nlme: Linear and Nonlinear Mixed Effects Models. R package version 3. Available at: https://CRAN.R-project.org/package=nlme.
Pusey A., Wolf M. (1996). Inbreeding avoidance in animals. Trends Ecol. Evol. 11 (5), 201–206. doi: 10.1016/0169-5347(96)10028-8
Ralls K., Ballou J. D., Templeton A. (19881988). Estimates of lethal equivalents and the cost of inbreeding in mammals. Conserv. Biol. 2 (2), 185–193.
Rice M. A., Galindez S. M., Garner J. T., Ophir A. G. (2022). Effects of social and environmental contexts on multi-male mating and mixed paternity in socially monogamous female prairie voles. R Soc. Open Sci. 9 (10), 220298. doi: 10.1098/rsos.220298
Rice M. A., Restrepo L. F., Ophir A. G. (2018). When to cheat: modeling dynamics of paternity and promiscuity in socially monogamous prairie voles ( Microtus ochrogaster). Front. Ecol. Evol. 6. doi: 10.3389/fevo.2018.00141
Roberts R. L., Cushing B. S., Carter C. S. (1998). Intraspecific variation in the induction of female sexual receptivity in prairie voles. Physiol. Behav. 64, 209–212. doi: 10.1016/s0031-9384(98)00042-0
Roberts S. C., Gosling L. M. (2003). Genetic similarity and quality interact in mate choice decisions by female mice. Nat. Genet. 35 (1), 103–106. doi: 10.1038/ng1231
Rodriguez N. A., Legzim K. M., Aliou F., Al-Naimi O. A., Bamshad M. (2013). Does mating prevent monogamous males from seeking other females? A study in prairie voles (Microtus ochrogaster). Behav. Processes. 100, 185–191. doi: 10.1016/j.beproc.2013.10.002
Shapiro L. E., Austin D. J., Ward S. E., Dewsbury D. A. (1986a). Familiarity and female mate choice in two species of voles (Microtus ochrogaster and M. montanus). Anim. Behav. 34, 90–97. doi: 10.1016/0003-3472(86)90010-2
Shapiro L. E., Dewsbury D. A. (1986b). Male dominance, female choice and male copulatory behavior in two species of voles (Microtus ochrogaster and Microtus montanus). Behav. Ecol. Sociobiol 18, 267–274. doi: 10.1007/BF00300003
Smale L., Nelson R. J., Zucker I. (1985). Neuroendocrine responsiveness to oestradiol and male urine in neonatally androgenized prairie voles (Microtus ochrogaster). J. Reprod. Fertil. 74, 491–496. doi: 10.1530/jrf.0.0740491
Solomon N. G. (1993). Body size and social preferences of male and female prairie voles, Microtus ochrogaster. Anim. Behav. 45, 1031–1033. doi: 10.1006/anbe.1993.1122
Solomon N. G., Jacquot J. J. (2002). Characteristics of resident and wandering prairie voles, Microtus ochrogaster. Can. J. Zool. 80 (5), 951–955. doi: 10.1139/z02-053
Solomon N. G., Keane B., Knoch L. R., Hogan P. J. (2004). Multiple Paternity in social monogamous prairie voles Microtus ochrogaster. Can. J. Zool. 82, 1667–1671. doi: 10.1139/z04-142
Solomon N. G., Ophir A. G. (2020). Editorial: what's love got to do with it: the evolution of monogamy. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.00110
Thomas J. A., Birney E. C. (1979). Parental care and mating system of the prairie vole, microtus ochrogaster. Behav. Ecol. Sociobiol. 5 (2), 171–186. doi: 10.1007/BF00293304
Tregenza T., Wedell N. (2000). Genetic compatibility, mate choice and patterns of parentage: invited review. Mol. Ecol. 9 (8), 1013–1027. doi: 10.1046/j.1365-294x.2000.00964.x
Venables W. N., Ripley B. D. (2002). Modern Applied Statistics with S. 4th ed. (New York: Springer). Available at: https://www.stats.ox.ac.uk/pub/MASS4/.
Walum H., Young L. J. (2018). The neural mechanisms and circuitry of the pair bond. Nat. Rev. Neurosci. 19, 643–654. doi: 10.1038/s41583-018-0072-6
Williams J. R., Catania K. C., Carter C. S. (1992). Development of partner preferences in female prairie voles (Microtus ochrogaster): The role of social and sexual experience. Horm. Behav. 26, 339–349. doi: 10.1016/0018-506x(92)90004-f
Winslow J. T., Hastings N., Carter C. S., Harbaugh C. R., Insel T. R. (1993). A role for central vasopressin in pair bonding in monogamous prairie voles. Nature 365, 545–548. doi: 10.1038/365545a0
Wolff J. O., Dunlap A. S. (2002). Multi-male mating, probability of conception, and litter size in the prairie vole (Microtus ochrogaster). Behav. Processes 58 (1-2), 105–110. doi: 10.1016/s0376-6357(02)00022-0
Wolff J., Mech S., Dunlap A. S., Hodges K. (2002). Multi-male mating by paired and unpaired female prairie voles (Microtus ochrogaster). Behaviour 139 (9), 1147–1160. doi: 10.1163/15685390260437308
Young L. J., Wang Z. (2004). The neurobiology of pair bonding. Nat. Neurosci. 7, 1048–1054. doi: 10.1038/nn1327
Zeileis A., Kleiber C., Jackman S. (2008). Regression Models for Count Data in R. Journal of Statistical Software. Available at: http://www.jstatsoft.org/v27/i08/.
Keywords: prairie voles, socio-sexual preferences, mating preferences, kinship, inbreeding avoidance, huddling
Citation: Ferreira-Nuño A, Young LJ, Morales Otal A, Camacho F, Díaz NF, Paredes RG, Prado A, Cruz Benites A and Portillo W (2023) Kin affiliation, socio-sexual and mating preference in the female Microtus ochrogaster in a multiple socio-sexual preference test. Front. Ecol. Evol. 11:1154800. doi: 10.3389/fevo.2023.1154800
Received: 31 January 2023; Accepted: 28 November 2023;
Published: 14 December 2023.
Edited by:
Sebastien Lebreton, Cearitis, FranceReviewed by:
Alexander G. Ophir, Cornell University, United StatesNicole Lee, University of California, Berkeley, United States
Copyright © 2023 Ferreira-Nuño, Young, Morales Otal, Camacho, Díaz, Paredes, Prado, Cruz Benites and Portillo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Armando Ferreira-Nuño, ZmVuYUB4YW51bS51YW0ubXg=; Wendy Portillo, cG9ydGlsbG9AdW5hbS5teA==