 Hugh D. Loxdale
Hugh D. Loxdale- School of Biosciences, Cardiff University, Cardiff, United Kingdom
Some colourful European wild birds display contrasting bright colours. These are often striking black and white or primary and secondary colours, and with the same basic plumage livery shared by both sexes. These contrasting colours are, one assumes, very obvious to predators when these birds forage diurnally, or continue other aspects of their life history, including courting, nest building and rearing their brood. Here, I posit that such birds may be displaying aposematic warning colouration, possibly enhanced by chemical noxious substances in their flesh and/or feathers, as is already known in certain bird species, including colourful as well as cryptic species. The warning colouration may be Müllerian or Batesian in nature, or maybe is a ruse to suggest to predators that they are in some way noxious, and thus to be avoided. Even if not actually noxious as such, this may give the intended prey time to escape. Certainly, birds like the very obviously blue-black and white patterned Eurasian magpie, Pica pica, are largely avoided by the Eurasian sparrowhawk, Accipiter nisus, although this of course could be a size-related avoidance, as other larger raptorial birds do predate it. These various possibilities are discussed in the present article.
Introduction
As is well-established (Cott, 1940), many examples of cryptic camouflage occur in the natural world, with some animals mimicking stones, flowers, twigs and branches, or the ground itself. Such camouflaged animals abound in many groups, especially insects, but also including fish, amphibians, reptiles, mammals and birds. They do so either to escape predation or in the case of predators, to capture prey. For example, the Eurasian stone-curlew, Burhinus oedicnemus (L.), disappears against the dry, often stony ground on which it lives and nests. Similarly, the tawny frog mouth, Podargus strigoides (Latham) of Australia, appears to be a part of a branch on a tree, until that is, it opens its mouth wide to catch a flying insect.
Many animals also show aposematic warning colouration: bright contrasting colours, such as yellow and orange, yellow and red or black and white, often displayed as sudden flash colouration, as in the hind wings of moths like the red underwing moth, Catocala nupta (L.) (Lepidoptera: Erebidae; Rothschild and Lane, 1960). This is to warn would-be attackers that they are to be avoided; that they are in some way chemically defended, either with noxious chemical sprays (e.g., skunks), poisonous venom (e.g., certain snakes like the coral snake), toxic skin or flesh (e.g., certain tropical frog species), or the warning is a ruse (Cott, 1940; Baker and Parker, 1979; Howse, 2014, 2021; Hedley and Caro, 2022). Perhaps then it is not surprising that certain bird species also apparently display warning colours, except that we have perhaps often failed to recognise the true purpose of such colourful plumage. We assess such birds as being harmless and hence ignore these warning signs, as they are apparently not a threat to us. But this may well not be true of other, smaller creatures like Passerine birds, reptiles and predatory arthropods which see – and duly take notice of – such signals (Howse, 2014, 2021, 2022).
Are some birds toxic including European species?
So is it true? Do some bird species, including European species, show warning colours and signals and are they potentially toxic if eaten, not only by us but by other animals? It is known that a few tropical birds display warning colours and are toxic (Baker and Parker, 1979; Hedley and Caro, 2022). One of the best examples is the hooded pitohui, Pitohui dichrous (Bonaparte) of New Guinea (Figure 1A), a medium-sized songbird with rich chestnut and black plumage, which contains a range of batrachotoxin (BTX) compounds (extremely potent cardio- and neurotoxic steroidal alkaloids found in certain species of beetles, birds, and frogs) in its skin, feathers and other tissues (cf. Bodawatta et al., 2023 and references therein). These are obtained from its diet, a mix of fruit, seeds and insects and other arthropods. The sequestered toxins in its feathers and flesh are used by the bird to provide it with some degree of protection against predators, probably mainly birds and snakes, since only one larger predatory mammal, a marsupial, exists in New Guinea (Leary et al., 2016). Both sexes have similar plumage. The toxins are especially concentrated in the breast and belly feathers, such that these not only protect the adult birds, but may also rub off on eggs and young in the nest (Dumbacher et al., 2000; Yeung et al., 2022). That such warning colouration is shown by other, poisonous pitohuis in New Guinea (Jønsson et al., 2008), as well as bird species of other genera, e.g., the blue-capped ifrit, Ifrita kowaldi (De Vis) (Dumbacher et al., 2000; Bodawatta et al., 2023) appears to be a case of convergent evolution leading to Müllerian mimicry, i.e., gaining additional protection by mimicking other contrastingly, brightly coloured noxious/venomous animals (Cott, 1940; Baker and Parker, 1979; Hedley and Caro, 2022). The Hooded Pitohui’s defence is reinforced by a strong odour (Dumbacher et al., 1992). Its livery is also apparently mimicked by other un-related, non-toxic birds, these birds hence showing Batesian mimicry,1 where a non-toxic animal mimics a toxic or dangerous model organism (cf. Cott, 1940).

Figure 1. (A-M): Left to right, from top: (A) Hooded Pitohui; (B) Goldfinch; (C) Great Spotted Woodpecker; (D) Great Tit; (E) Pied Wagtail; (F) Kingfisher; (G) Barn Swallow; (H) Eurasian Magpie; (I) Eurasian Nuthatch; (J) Eurasian Oystercatcher; (K) European Bee-eater; (L) Hoopoe; (M) Roller. Photo credit: (A) eBird,;Frédérick Pelsy (B) eBird, Eitan Altman; (C) Pixabay, Kurt Bouda; (D) Pixabay, Christiane; (E) Nature Spot, David Nicholls; (F) Pixabay, Timo Schlüter; (G) iStock, Trevor Jones; (H) NautreSpot, Barbara Cooper; (I) eBird, Santiago Caballero Carrera; (J) Pixabay, Mabel Amber; (K) eBird, Josep del Hoyo; (L) Pixabay, Harald Landsrath; (M) eBird, Rafael Merchante.
With this in mind, it occurred to me whist watching the wild birds coming to the bird table and feeders in my garden in North Devon, SW England that the many European Goldfinches, Carduelis carduelis (L.) (Figure 1B) that visit with their red, black and white face masks, black and white wing and tail feathers and bright yellow flash colouration on the wings, may be displaying warning colouration (Loxdale, 2022; cf. also Howse, 2014, p. 85). In the same article, I also surmised that other common garden birds like the Great Spotted woodpecker, Dendrocopos major (L.), tits, e.g., Great tit, Parus major L., and even wagtails, e.g., Pied wagtail, Motacilla alba L. (Figures 1C–E), may also be showing warning colouration. If their feathers and flesh are to some degree toxic to would-be predators, these could be further examples of avian Müllerian mimicry. On the other hand, if the birds are not toxic, then such examples could be evidence of Batesian mimicry. Besides the pitohuis and blue-capped ifrit of New Guinea, other birds from other taxa and geographical regions are known to have toxic flesh as a result of sequestering toxins from their seed and insect diet. These include the cryptically-coloured European or Common quail, Coturnix coturnix (L.) (Korkmaz et al., 2011) and the Ruffed grouse, Bonasa umbellus (L.) of North America (Causey, 2020). Goldfinches consume seeds and feed insects to their young, whilst the diet of tits and wagtails is largely insectivorous. Herbivorous insects like caterpillars and aphids, the principal food of many small insectivorous Passerine birds, are known to harbour toxins from their plant-based diet (Opitz and Müller, 2009). Cott (1947, p 489) considers these to be a likely source of some of the toxins found in distasteful birds.
In addition to the aforementioned European garden birds, other species may be displaying warning colouration related to Müllerian or Batesian mimicry, and like the Hooded pitohui and goldfinch, show a lack of sexual dimorphism in plumage colouration. In this light, the Eurasian kingfisher, Alcedo atthis (L.), Barn swallow, Hirundo rustica L., Eurasian magpie, Pica pica (L.), Eurasian nuthatch, Sitta europaea L. (Pallas), Eurasian oystercatcher, Haematopus ostralegus L., and the rare visitors to northern Europe, including the British Isles, the European bee-eater, Merops apiaster L., Eurasian hoopoe, Upupa epops L., and Roller, Coracias garrulus L. come to mind (Figures 1F–M). There are probably other gaudy species also that fit the category of noxious species, as Cott (1947) implies. Perhaps we just do not see it, but other animals do, notably would-be predators. These are suitably impressed enough, if only momentarily, to generally avoid these birds when they come across them, allowing the intended prey to escape. Certainly, the largely insectivorous hoopoe, which shows aposematic warning colouration in the form of contrasting black and white striped wings and tail, and a striking black and white-tipped orange crest surmounting its salmon pink head and upper torso (Figure 1L), is known to have anti-predator chemical defences in the nest. The uropygial gland of the incubating and brooding female produces a foul-smelling liquid, as do the glands of nestlings, these secretions being rubbed into the plumage. The secretion smells like rotting meat and is used to deter predators and parasites and possibly act as an antibacterial agent (Martin-Platero et al., 2006). The secretions stop just prior to the young leaving the nest (Fry, 2003).
Discussion and conclusions
An excellent pioneering insight into the edibility of different bird species was provided in the 1940s by Cott (1947). This concerned a 4-year comparative quantitative study of the attractiveness and acceptability of the flesh, especially the breast meat, of a range of bird species inhabiting Egypt and the Lebanon, whilst the author was stationed out there in the military during World War 2. Most of the birds tested in the study (38 spp.) were directly shot. Their edibility was assessed by exposing the flesh of the various birds tested (a lump 5–7 mm. in diameter) to the predations of the Oriental hornet, Vespa orientalis L. (Hymenoptera: Vespidae), an omnivorous social insect. The number of insects attending the flesh over a given 5-min period was assessed visually and the data collated and represented both graphically and in tabular form (cf. Cott, 1947 for further details).
A major finding of this study was that there was a clear inverse relationship between edibility and visibility: the more cryptic species were generally more desirable to the hornets than the more visible species. The latter often appeared in broad daylight and were seen to display aposematic warning colouration to a greater or lesser degree, depending on the species concerned. To do this, Cott prepared a list of bird species vs. edibility (E), arbitrarily giving each species an edibility ranking of 1–20, where 20 was the most edible. Also, a list of species vs. visibility (V) with a ranking of 1–10, where 10 was the most visible (see his Table LVIII, p. 414 and Figure 30, p. 437). He stated that visibility is in many ways dependent on the habitat that the particular bird species inhabits (desert, semi-arable, arable), some species of course showing severe habitat fidelity (e.g., those highly adapted species inhabiting deserts). Other brightly coloured species (e.g., Barn swallow, H. rustica), are seen to move between habitats, especially when water is present (oases, streams and rivers, etc.). Cott made the additional point that some visible species like the bee-eater are not so visible when they are seen against – say – a background of mixed vegetation, whist, as we now know, some birds have hues not necessarily visible to the human eye, i.e., ultraviolet (Cuthill et al., 2000; Withgott, 2000; see below). Lastly, he suggested that birds with pale underparts are actually showing a certain degree of crypsis as they move around the habitat, especially one assumes with the sky above them.
So the picture that emerges is generally clear, but with some level of variation associated with it. Certainly, some anomalies in the rankings are apparent. Of the aforementioned species inhabiting Europe as residents and tested by Cott (1947), i.e., Kingfisher (Mediterranean form); Barn swallow, Goldfinch Carduelis carduelis ssp. niediecki Reichenow (Middle-Eastern form), Great tit, Parus major ssp. terrae-sanctae Hartert. (Palestinian form), Pied (White) wagtail, or very occasional visitors from Africa, i.e., the bee-eater [Cott tested the closely related and colourful species, the Little bee-eater, Merops orientalis cleopatra (Nicoll)], and the closely related and similarly marked hoopoe subspecies, Upupa epops major (Brehm; Cocker and Mabey, 2005), these were found to have high V and low E rankings, respectively, as shown in Table 1. Although not directly tested in the various feeding experiments, he cites Neumann (1905) concerning the edibility of the Great spotted woodpecker, D. major, describing it as “coarse, and retaining some of the repulsive smell of the bird itself, even when roast, so that it does not make pleasant eating” (cf. p. 283 in Neumann, Band 4, pp. 432). Interestingly, one of the most edible of the 38 species tested by Cott was the Wryneck, Jynx torquilla (L.), a very cryptically-camouflaged species with a visibility and edibility ranking of V = 1 and E = 20, respectively. The odour and edibility of the nuthatch was not tested nor seemingly by other authors subsequently, and therefore awaits such investigation.
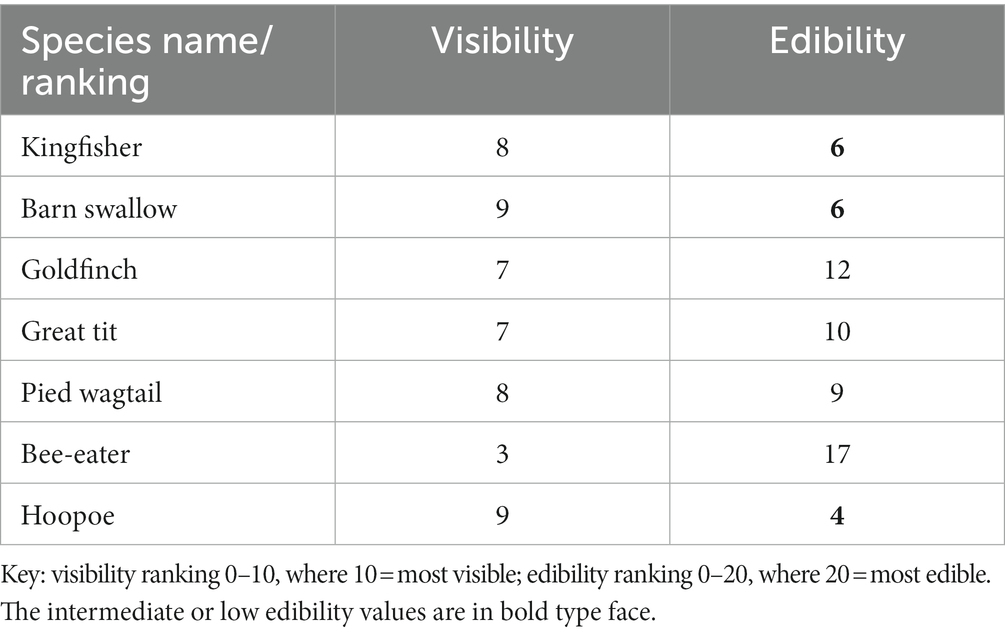
Table 1. Relative visibility and edibility rankings of birds assumed to display aposematic warning colouration and to have intermediate or low (bold numbering) edibility, as originally tested by Cott (1947).
In a further series of experiments, edibility tests were performed with domestic cats, Felis catus L. and humans. In the case of cats, Cott (1947) states that (p. 455):
Neither the number of experiments carried out nor the number of species tested is sufficient to yield a detailed comparison between food preferences of cat and hornet. The available evidence does appear, however, to show a general agreement on the likes and dislikes of these two unrelated meat-eaters.
With humans, rather than direct tests of edibility, he cites various published sources. He concludes that:
When we turn to compare the preferences of man, cat and hornet, unfortunately the available evidence only covers seven genera, namely Chloris, Passer, Merops, Streptopelia, Parus, Upupa, and Ceryle – in which the same or closely allied species were tested by all three meat-eaters. It is a striking fact that in none does the evidence suggest a considerable difference in the edibility rating: this concurrence of taste appears all the more remarkable when found, as here, in three creatures so utterly differing in organisation and habits.
In a follow-up study performed in Zambia, Cott and Benson (1969) continued investigation of the relative palatability vs. visibility of birds using 200 species belonging to 57 families, tested using a panel of human volunteers. Many of these birds were “not normally eaten by man,” but again an inverse relationship was found between edibility and visibility. Interestingly, the authors noted that the palatability of the very visibly marked Lilac-breasted roller, Coracias caudatus L. was given a moderate value of 6.1 (range 2 = inedible to 9 = excellent), whereas its visibility was scored at 5 on a scale of seven grades ranging from 1 (highly cryptic) to 7 (highly conspicuous; Cott and Benson, 1969).
Lastly in this context, Götmark (1994) re-analysed Cott’s data and showed that in 30 South European passerines there was, irrespective of sex, a negative correlation between visibility/conspicuousness and edibility. Similarly, in 87 non-passerine birds from southern Africa, a similar trend was noted across species, including closely-related species. However, in 105 female passerines tested, also from southern Africa, visibility/conspicuousness were negatively correlated with edibility, but only significantly so in relation to conspicuousness. Conversely, male breeding plumage conspicuousness of these species was not negatively correlated with edibility in any analysis. Thus, in such sexually dimorphic species, visibility and conspicuousness may, in males, be sexually selected, or signal other aspects of prey unprofitability rather than being related to edibility as such.
From these various studies and analyses, there is sometimes an obvious and clear negative association between visibility/conspicuousness and edibility, whereas in other studies and tests, including using live raptorial birds and either stuffed prey species or their flesh, the correlation is not clear. Even so, on the basis that nothing in nature exists if it has not a purpose (or once had) à la Dobzhansky (1973), then an explanation for bright colours in sexually monomorphic birds needs to be found, or at least posited.
This is something that Weldon and Rappole (1997) have investigated further by asking field and museum ornithologists which bird species they considered odorous (from intact or skinned exemplars) or unpalatable (from cooked birds). Likewise, Dumbacher and Pruett-Jones (1996) review the available evidence, direct and apocryphal, as to the nature and extent of avian toxins amongst different bird species representing widely different taxa, which they duly tabulate in terms of their odour or palatability. These studies show that some of the aforementioned bird groups are malodourous or unpalatable, including species of woodpecker, kingfisher, tits and the Eurasian oystercatcher, the last having a very prominent black and white livery and a crimson bill, pink legs and feet (Neumann, 1905; Dumbacher and Pruett-Jones, 1996; Figure 1J). Empirical studies need to be performed to test this hypothesis further and determine the chemical nature of the toxins involved for the various species. Very recently, Yeung et al. (2022) have reviewed the nature of the toxicity of some well-known poisonous birds, such as the pitohui and hoopoe, including the structure of the toxins themselves (cf. also Bartram and Boland, 2001 and Ligabue-Braun and Carlini (2015) in this respect).
Of the aforementioned colourful, sexually monomorphic birds displaying aposematic warning colouration, what would be of further interest is to ascertain whether this is Müllerian or Batesian in nature (in this context, cf. Table 1 in Hedley and Caro, 2022). The fact that these birds fly and feed in daytime in full sight of would-be predators suggests that they do indeed possess a certain degree of toxicity. Birds such as the Goldfinch may not be especially toxic (according to Cott’s scale of edibility the species is intermediate; Table 1), but as with the harmless scarlet king snake, Lampropeltis elapsoides (Holbrook) of the eastern and south eastern USA, a Batesian mimic of the deadly coral snake, Micrurus fulvius (L.), it is a false aposematic mimic. Their plumage livery might be enough to deter many, if not most, predators, but not all (Unprofitable Prey Hypothesis, UPH; cf. Hedley and Caro, 2022).
Raptorial birds, including Eurasian sparrowhawk, Peregrine falcon, Falco peregrinus Tunstall, Goshawk, Accipiter gentilis (L.), Merlin, Falco columbarius L. and Hobby, Falco subbuteo L. are known to predate some or all of the following prey species: Goldfinch, Barn swallow, Great tit, Pied wagtail, Magpie and Great spotted woodpecker (Owen, 1932; Selås, 1993; Zawadzka and Zawadzki, 2001; López-López et al., 2009; Ivanovskij and Sidorovich, 2018). In the USA, the American Oystercatcher, Haematopis palliates Temminck, which has a similar livery to the Eurasians species, is predated by skunks and racoons and some larger birds, including raptors (Hardin, 2014). However, according to Uttendörfer (1939; cited in Cott, 1947), of 43,211 prey items taken by Eurasian sparrowhawk, only 5% comprised Great tit and 5% Barn swallow (cf. Cott, 1947, p. 491 for further details). In the case of the Eurasian magpie, a strikingly obvious diurnal feeder, it is rarely, if ever, attacked by the Sparrowhawk (Owen, 1932), but is regularly attacked by the larger Goshawk in urban areas of Germany (Rutz, 2004), Peregrines in Spain (López-López et al., 2009) and Common Buzzard, Buteo buteo (L.) in the northeast of Ireland (Rooney and Montgomery, 2013). As quoted in Cott (1947; p. 482), the flesh of the magpie as eaten by one person was described as being “pretty awful.” In a quantitative study by Götmark (1997) carried out in Scandinavia in which he tested stuffed Eurasian magpies and Eurasian jay, Garrulus glandarius (L.) susceptibly to attacks by Goshawk, he found the attack rate was similar, and overall, the tests suggested frequency dependent selection of the prey birds by the predator. The author concluded that the livery of Eurasian magpies is not aposematic in nature and that whilst it may increase the likelihood of predation, may instead favour sexual or social selection. Despite this conclusion, the bird is very obvious, with a colour scheme normally associated with warning colouration in other animals like the Common European adder, Vipera berus (L.) and Striped skunk, Mephitis mephitis (Schreber) of North America.
In other experiments (Götmark and Unger, 1994) involving stuffed birds, in this case, Pied wagtail paired with cryptic Meadow pipit, Anthis pratensis (L.) and Great Spotted woodpecker paired with cryptic (brown) female Blackbird, Turdus merula L., the attacks by avian predators (mostly Accipiters and filmed with automatic cameras) on the wagtail and pipit were similar, whereas the blackbird were much more frequently attacked than the woodpecker. To test the palatability of woodpecker vs. blackbird, the authors also conducted experiments feeding flesh of these birds directly to captive falcons. No clear evidence was found that woodpeckers are especially distasteful to these particular avian predators. A stuffed bird is of course not supplying all the potential deterrents that a living bird may do, i.e., unpleasant odour and/or unpalatable or toxic flesh.
Perhaps, as with the North American Blue Jay, Cyanocitta cristata (L.) during a first encounter with the Monarch butterfly, Danaus plexippus (L.) (Lepidoptera: Nymphalidae), the larvae of which feed on Asclepias curassavica L. (Gentianales: Apocynaceae) which contain toxic cardiac glycosides (Brower et al., 1967), naïve predatory birds and mammals may initially attack such prey bird species. However, if these are too big to easily subdue or if subdued and eaten, have an unpleasant aftertaste or cause an unpleasant reaction, then the predator soon learns to avoid them. Conjecture yes, but then again, how does one account for such obvious livery and diurnal foraging behaviour of these colourful birds, which overtly display their presence in the landscape? Because both sexes show the same or largely the same bright plumage (e.g., Great spotted woodpecker), this is probably not for purposes of sexual display and reproduction (Loxdale, 2022). Therefore, it does not involve a trade-off between negative selection incurred by predators due to the visibility and “showiness” of the male vs. crypsis and greater survival of the female, as seen in sexually dimorphic birds like the Common Chaffinch, Fringilla coelebs L. and Eurasian Bullfinch, Pyrrhula pyrrhula (L.) (cf. Götmark, 1993 for conflicting experimental evidence for this assumption).
Other aspects of interest when considering the putative palatability – or not – of the aforementioned colourful bird species is whether they are universally toxic to would-be predators, or only to some of them, which may have evolved a certain tolerance to the innate toxins. Besides the fact that different species of raptor have different food preferences and diet breadth, even known noxious birds such as the hoopoe are predated by some raptors in some locations (e.g., Peregrine in eastern Spain; López-López et al., 2009). The fact that the Eurasian Magpie, whilst being shunned by many predatory birds, is taken by Buzzard, Goshawk and Peregrine, perhaps relates to the fact that these are larger than the Sparrowhawk and can thus subdue it more easily. It is also possible that the prey is not uniformly toxic, and the predator selects the flesh that is less so, or they have become tolerant to the toxins the species contain, as has the Peregrine to those harboured by Hoopoe. The abundance of potential prey (availability) must also play some part in whether or not a particular predator is likely to tackle a given prey species, as shown experimentally in other predator–prey scenarios (Blouin, 1990; Marples, 1993; Hossie et al., 2021).
That we see the birds, the topic of this article, as very obvious within the environment, also does not mean that they are universally seen as such. For a start, as aforementioned, birds can see into the UV spectrum, which means that their perception of these colours and hues may be somewhat different than those we humans or other mammals/animals perceive. Furthermore, with any discussion concerning aposematically coloured animals vs. cryptic ones, this has to be seen in context to the habit they are living in, in turn affected by environmental conditions of habitat complexity, prevailing light conditions and weather/climate. Thus for example, it is known that distance-dependent factors are associated with the perception of putative prey by would-be predators (Barnet and Cuthill, 2014; Barnett et al., 2018). There are in addition, trade-offs between prey obviousness in terms of aposematism (and hence the possibly of predation by naïve birds) and crypsis, such that an intermediate state is found to be adaptively favoured (Tullberg et al., 2005 in the case of swallowtail butterfly larvae, Papilio machaon L.). In further support of this idea, Loeffler-Henry et al. (2023) suggest that species (here amphibians) also display an intermediate stage between crypsis and bright colouration involving facultative exposure of concealed colours (e.g., under the belly) when attacked. In this way, they gain the best of both worlds in terms of crypsis and warning defence. Ultimately, however, if the selective pressure is great enough, the costs of producing full-on warning colouration, perhaps with concomitant changes in warning behaviour, must outweigh the intermediate strategy in terms of adaptive and hence survival strategy.
Another aspect worthy of consideration is the variation in the toxicity of the prey and how this might affect predation. Speed et al. (2012) argue that over the course of the evolution, toxin variation is of adaptive significance and depends on the dose of toxin borne by a particular species geno-phenotype in relation to their abundance in the population, hence leading to the frequency-dependent selective response of predators. In a similar vein, Endler and Mappes (2004), using mathematical modelling, explore the effects of predators on prey in relation to frequency-dependent, frequency-independent, and negative frequency-dependent predation. Their main conclusion is that “weak signalling of aposematic species can evolve if predators vary in their tendency to attack defended prey,” again presumably in relation to the cost benefits of producing the toxin with accompanying warning signalling, chemical and behavioural. Less effort in responding to predators in terms of signals/signalling may also be adaptively advantageous.
Certainly, in the case of the presently discussed colourful birds, the fact that they are predated by some of the common bird predators like sparrowhawk and goshawk, despite showing clear aposematic warning coloration, argues that these particular predators, especially naïve ones, may indeed be functioning on a frequency-depended basis. Over time, they learn of the non-profitability of such prey, and avoid them, If this were not so, why do such bright warning colours persist, suggesting that they have been positively selected over the course of evolution. This does not help much in determining whether such colouring really is Müllerian or Batesian in nature, but probably it is the former, a broad warning signal, as with venomous wasps and bees. If it were Batesian, then models would have to be identified, and none are obvious.
An additional factor to consider in the context of the attack vs. avoidance of aposematically-coloured birds includes social learning. Thus individuals of conspecific/congeneric predator species, or even those of very different taxa, rapidly learn of the effect on their fellow predators when these attack a particular kind of prey, e.g., discarding the prey or, if eaten, vomiting it up shortly afterwards (Hämäläinen et al., 2020), as in the Blue Jay/Monarch butterfly scenario.
Lastly, in all discussion of the obviousness of a potential prey item within its habit, aspects of habitat matching and/or disruptive coloration must be considered (Cott, 1940). This topic is discussed by Stevens and Merilaita (2009), who revise earlier categorisations, especially by Thayer (1909) and Cott (1940), and re-assesses the range of principles and sub-categories that govern the phenomena involved (cf. Stevens and Merilaita, 2009 for details; also Honma et al., 2015 and Kang et al., 2015). Only in light of these various factors and influences can the true nature of the colourful livery of the birds be more fully understood.
In conclusion, the available evidence strongly suggests that some familiar European birds are displaying aposematic warning colouration and as such, may be noxious-toxic to would-be predators, although this is not to say that naïve birds may initially predate them, but learn to avoid them in future. The mimicry could well be Müllerian in nature, i.e., a broad warning signal, but may also be a ruse to some extent (so-called “cheats”; Speed et al., 2012), as in the case of the Goldfinch, possibly making some predators (but clearly not all, i.e., some Accipiters) wary of attacking the bird in the first place. If this is so, the gaudy livery, rather than being of no purpose, and apparently not involved in sexual selection by the female since both sexes share the same or nearly the same livery, argues that the plumage colouring is of selective advantage (Endler and Mappes, 2004; Speed et al., 2012). It is unlikely, but not impossible, that in some species with low toxicity status, the mimicry is Batesian in nature, but what the exact model might be remains conjectural.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
The author confirms being the sole contributor of this work and has approved it for publication.
Acknowledgments
I thank the referee for their most helpful comments which have improved the manuscript and Philip Howse OBE for kindly sending me an original copy of Cott’s (1947) paper. I also thank Cornell Laboratory of Ornithology for permission to reproduce certain photos as originally displayed on their eBird website (https://ebird.org/home), and David Nicholls of NatureSpot (https://www.naturespot.org.uk/) for allowing me to use photos of the Pied wagtail and Eurasian magpie. Other bird photos were downloaded free from the Pixabay website (https://pixabay.com/), whilst the photo of the Barn swallow was purchased from iStock (https://www.istockphoto.com/stock-photos).
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
References
Baker, R. R., and Parker, G. A. (1979). The evolution of bird coloration. Philos. Trans. R. Soc. B: Biol. Sci. 287, 63–130.
Barnet, J. B., and Cuthill, I. C. (2014). Distance-dependent defensive coloration. Curr. Biol. 24, R1157–R1158. doi: 10.1016/j.cub.2014.11.015
Barnett, J. B., Cuthill, I. C., and Scott-Samuel, N. E. (2018). Distance-dependent aposematism and camouflage in the cinnabar moth caterpillar (Tyria jacobaeae, Erebidae). R. Soc. Open Sci. 5:171396. doi: 10.1098/rsos.171396
Bartram, S., and Boland, W. (2001). Chemistry and ecology of toxic birds. Chembiochem 2, 809–811. doi: 10.1002/1439-7633(20011105)2:11<809::AID-CBIC809>3.0.CO;2-C
Blouin, M. S. (1990). Evolution of palatability differences between closely-related tree-frogs. J. Herpetol. 24, 309–311. doi: 10.2307/1564399
Bodawatta, K. H., Hu, H., Schalk, F., Daniel, J.-M., Maiah, G., Koane, B., et al. (2023). Multiple mutations in the Nav1.4 sodium channel of new Guinean toxic birds provide autoresistance to deadly batrachotoxin. Mol. Ecol. doi: 10.1111/mec.16878
Brower, L. P., van Brower, J., and Corvino, J. M. (1967). Plant poisons in a terrestrial food chain. PNAS 57, 893–898. doi: 10.1073/pnas.57.4.893
Causey, K.F. (2020). Dietary roulette: Human poisoning from ruffed grouse (Bonasa umbellus). California, USA: University of California, Davis, pp. 21.
Cott, H. B. (1947). The edibility of birds: illustrated by five years' experiments and observations (1941–1946) on the food preferences of the hornet, cat and man; and considered with special reference to the theories of adaptive coloration. Proc. Zool. Soc. London 116, 371–524. doi: 10.1111/j.1096-3642.1947.tb00131.x
Cott, H. B., and Benson, C. W. (1969). The palatability of birds, mainly based upon observations of a tasting panel in Zambia. Ostrich. J. African Ornithol. 40, 357–384. doi: 10.1080/00306525.1969.9639135
Cuthill, I. C., Partridge, J. C., Bennett, A. T. D., Church, S. C., Hart, N. S., and Hunt, S. (2000). Ultraviolet vision in birds. Adv. Study Behav. 29, 159–214. doi: 10.1016/S0065-3454(08)60105-9
Dobzhansky, T. (1973). Nothing in biology makes sense except in the light of evolution. Am. Biol. Teach. 35, 125–129. doi: 10.2307/4444260
Dumbacher, J. P., Beehler, B., Spande, T., Garraffo, H., and Daly, J. (1992). Homobatrachotoxin in the genus Pitohui: chemical defense in birds? Science 258, 799–801. doi: 10.1126/science.1439786
Dumbacher, J. P., and Pruett-Jones, S. (1996). Avian chemical defense. Curr. Ornithol. 13, 137–174. doi: 10.1007/978-1-4615-5881-1_4
Dumbacher, J. P., Spande, T. F., and Daly, J. W. (2000). Batrachotoxin alkaloids from passerine birds: a second toxic bird genus (Ifrita kowaldi) from New Guinea. PNAS 97:12970. doi: 10.1073/pnas.200346897
Endler, J. A., and Mappes, J. (2004). Predator mixes and the conspicuousness of aposematic signals. Am. Nat. 163, 532–547. doi: 10.1086/382662
Fry, H. C. (2003). “Hoopoe,” in Firefly Encyclopedia of birds. ed. C. Perrins Toronto, Canada: Firefly Books.
Götmark, F. (1993). Conspicuous coloration in male birds is favoured by predation in some species and disfavoured in others. Proc. R. Soc. B: Biol. 253, 143–146. doi: 10.1098/rspb.1993.0094
Götmark, F. (1994). Are bright birds distasteful? A re-analysis of H.B. Cott’s data on the edibility of birds. J. Avian Biol. 25, 184–197. doi: 10.2307/3677074
Götmark, F. (1997). Bright plumage in the magpie: does it increase or reduce the risk of predation? Behav. Ecol. Sociobiol. 40, 41–49. doi: 10.1007/s002650050314
Götmark, F., and Unger, U. (1994). Are conspicuous birds unprofitable prey? Field experiments with hawks and stuffed prey species. Auk 111, 251–262. doi: 10.2307/4088590
Hämäläinen, L., Mappes, J., Rowland, H. M., Teichmann, M., and Thorogood, R. (2020). Social learning within and across predator species reduces attacks on novel aposematic prey. J. Anim. Ecol. 89, 1153–1164. doi: 10.1111/1365-2656.13180
Hardin, M. (2014). Haematopus palliatus (On-line), Animal Diversity Web. Available at https://animaldiversity.org/accounts/Haematopus_palliatus/ (Accessed March 3, 2023).
Hedley, E., and Caro, T. (2022). Aposematism and mimicry in birds. Ibis 164, 606–617. doi: 10.1111/ibi.13025
Honma, A., Mappes, J., and Valkonen, J. K. (2015). Warning coloration can be disruptive: aposematic marginal wing patterning in the wood tiger moth. Ecol. Evol. 5, 4863–4874. doi: 10.1002/ece3.1736
Hossie, T. J., Chan, K., and Murray, D. L. (2021). Increasing availability of palatable prey induces predator-dependence and increases predation on unpalatable prey. Sci. Rep. 11, 1, 6763–6712. doi: 10.1038/s41598-021-86080-x
Howse, P. (2014). Seeing butterflies: New perspectives on colour, patterns and mimicry. London & Winterbourne, Berkshire, UK: Papadakis.
Howse, P. (2021). Bee tiger: The death’s head hawk-moth through the looking glass. Taunton, Somerset, UK: Brambleby Books Ltd.
Howse, P. (2022). The spider-winged cupid and the platypus. Poole, Dorset, UK: Redfern Natural History Productions Ltd.
Ivanovskij, V.V., and Sidorovich, A.A. (2018). Niche separation between the Merlin (Falco columbarius) and the Eurasian Hobby (Falco subbuteo) nested on pine bogs of northern Belarus. Bulletin of TVGU. Series ‘Biology and Ecology’ 1, 103–112.
Jønsson, K. A., Bowie, R. C. K., Norman, J. A., Christidis, L., and Fjeldså, J. (2008). Polyphyletic origin of toxic Pitohui birds suggests widespread occurrence of toxicity in corvoid birds. Biol. Lett. 4, 71–74. doi: 10.1098/rsbl.2007.0464
Kang, C., Stevens, M., Moon, J. -y., Lee, S. -I., and Jablonskia, P. G. (2015). Camouflage through behavior in moths: the role of background matching and disruptive coloration. Behav. Ecol. 26, 45–54. doi: 10.1093/beheco/aru150
Korkmaz, I., Kukul Güven, F.M., and Eren, Ş.H. and, Dogan, Z. (2011). Quail consumption can be harmful. J. Emerg. Med. 41, 499–502. doi: 10.1016/j.jemermed.2008.03.045
Leary, T., Seri, L., Flannery, T., Wright, D., Hamilton, S., Helgen, K., et al. (2016). Dasyurus spartacus. IUCN Red List of Threatened Species. 2016, e.T6301A21947093. Available at: www.gbif.org/species/176661403 (Accessed 24 April, 2023).
Ligabue-Braun, R., and Carlini, C. R. (2015). Poisonous birds: a timely review. Toxicon 99, 102–108. doi: 10.1016/j.toxicon.2015.03.020
Loeffler-Henry, K., Kang, C., and Sherratt, T. N. (2023). Evolutionary transitions from camouflage to aposematism: hidden signals play a pivotal role. Science 379, 1136–1140. doi: 10.1126/science.ade5156
López-López, P., Verdejo, J., and Barba, E. (2009). The role of pigeon consumption in the population dynamics and breeding performance of a peregrine falcon (Falco peregrinus) population: conservation implications. Eur. J. Wildl. Res. 55, 125–132. doi: 10.1007/s10344-008-0227-2
Loxdale, H. D. (2022). Insights from watching wild birds at bird feeders: biodiversity, behaviour and warning colouration. Linnean 38, 26–33.
Marples, N. M. (1993). Do wild birds use size to distinguish palatable and unpalatable prey types? Anim. Behav. 46, 347–354. doi: 10.1006/anbe.1993.1196
Martin-Platero, A. M., Valdivia, E., Ruiz-Rodriguez, M., Soler, J. J., Martin-Vivaldi, M., Maqueda, M., et al. (2006). Characterization of antimicrobial substances produced by Enterococcus faecalis MRR 10-3, isolated from the Uropygial gland of the hoopoe (Upupa epops). Appl. Environ. Microbiol. 72, 4245–4249. doi: 10.1128/AEM.02940-05
Opitz, S. E. W., and Müller, C. (2009). Plant chemistry and insect sequestration. Chemoecology 19, 117–154. doi: 10.1007/s00049-009-0018-6
Rooney, E., and Montgomery, W. I. (2013). Diet diversity of the common buzzard (Buteo buteo) in a vole-less environment. Bird Study 60, 147–155. doi: 10.1080/00063657.2013.772085
Rothschild, M., and Lane, C. (1960). Warning and alarm signals by birds seizing aposematic insects. Ibis 102, 328–330. doi: 10.1111/j.1474-919X.1960.tb07120.x
Rutz, C. (2004). Breeding season diet of northern goshawks Accipiter gentilis in the city of Hamburg, Germany. Corax 19, 311–322.
Selås, V. (1993). Selection of avian prey by breeding Sparrowhawks Accipiter nisus in southern Norway: the importance of size and foraging behaviour of prey. Ornis Fennica 70, 144–154.
Speed, M. P., Ruxton, G. D., Mappes, J., and Sherratt, T. N. (2012). Why are defensive toxins so variable? An evolutionary perspective. Biol. Rev. 87, 874–884. doi: 10.1111/j.1469-185X.2012.00228.x
Stevens, M., and Merilaita, M. (2009). Defining disruptive coloration and distinguishing its functions. Philos. Trans. R. Soc. B 364, 481–488. doi: 10.1098/rstb.2008.0216
Thayer, G. H. (1909). Concealing-coloration in the animal kingdom: An exposition of the laws of disguise through color and pattern: Being a summary of Abbott H. Thayer’s discoveries. New York: Macmillan.
Tullberg, B. S., Merilaita, S., and Wiklund, C. (2005). Aposematism and crypsis combined as a result of distance dependence: functional versatility of the colour pattern in the swallowtail butterfly larva. Proc. R. Soc. B 272, 1315–1321. doi: 10.1098/rspb.2005.3079
Uttendörfer, O. (1939). Die Ernährung der deutschen Raubvögel und Eulen und ihre Bedeutung in der heimischen Natur. Sachsen-Anhalt, Germany: Neumann-Neudamm.
Weldon, P. J., and Rappole, J. H. (1997). A survey of birds odorous or unpalatable to humans: possible indications of chemical defense. J. Chem. Ecol. 23, 2609–2633. doi: 10.1023/B:JOEC.0000006670.79075.92
Withgott, J. (2000). Taking a Bird's-eye view … in the UV: recent studies reveal a surprising new picture of how birds see the world. Bioscience 50, 854–859. doi: 10.1641/0006-3568(2000)050[0854,TABSEV]2.0.CO;2
Yeung, K. A., Chai, P. R., Russell, B. L., and Erickson, T. B. (2022). Avian toxins and poisoning mechanisms. J. Med. Toxicol. 18, 321–333. doi: 10.1007/s13181-022-00891-6
Keywords: aposematic, Batesian mimicry, birds, Müllerian mimicry, plumage, predators, toxicity, warning colouration
Citation: Loxdale HD (2023) Are some brightly coloured European wild birds toxic? Front. Ecol. Evol. 11:1150576. doi: 10.3389/fevo.2023.1150576
Edited by:
Shahram Niknafs, The University of Queensland, AustraliaReviewed by:
Carita Lindstedt, University of Jyväskylä, FinlandCopyright © 2023 Loxdale. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hugh D. Loxdale, aHVnaGRsb3hkYWxlQGdtYWlsLmNvbQ==