 Cheng-Ren Zhao1,2†
Cheng-Ren Zhao1,2† Binghua Sun
Binghua Sun Dong-Po Xia
Dong-Po Xia
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 09 May 2023
Sec. Behavioral and Evolutionary Ecology
Volume 11 - 2023 | https://doi.org/10.3389/fevo.2023.1144932
Immigrant males employed a range of species-specific social strategies to facilitate integration into a new social group for establishing stable dominant ranks and affiliative relationships with unfamiliar competitors who require them to adapt their behavior. However, less is known whether the social strategies of integration were individual-specific. In this study, a case that nine adult males immigrated into the subject social group provided a natural experiment to investigate the variation of social strategies between the bluff and unobtrusive immigrant males during integration into a new social group. We followed the wild YA1 Tibetan macaque group and collected the behavioral data from August 2021 to May 2022. The results showed that the bluff and unobtrusive immigrant males faced similar social pressure from intrasexual individuals in the target social group during both the prospecting and integrating stages. During the prospecting stage, the bluff and unobtrusive immigrant males formed “attempt strategies” to seek and establish social relationships with unfamiliar individuals in a new group. The bluff immigrant males formed affiliative relationships with high-ranking females and resident males whereas the unobtrusive immigrant males formed affiliative relationships with low-ranking females during the integrating stage. Our results firstly provided quantitative methods to distinguish prospecting and integrating stages during immigration processes. This case study also demonstrated that the bluff and unobtrusive immigrant males formed varied social strategies. Our results provided new insight into understanding the individual-specific social strategies of immigrant males during integration into a new social group.
Dispersal, defined as the movement of an individual from a social group to another group, is an important consequence of long-term ecological adaptation for individual fitness because it limits the risks of inbreeding (Perrin and Goudet, 2001). It may enable an individual to increase mating opportunities and improve their reproductive success (Pusey and Packer, 1987; Teichroeb et al., 2011; Wei et al., 2016). In non-human primates, male-biased dispersal with female philopatry is the most common pattern, especially in Asian and African cercopithecine species (Greenwood, 1980; Borries, 2000; Clutton-Btock and Lukas, 2012). Dispersal involves three-step processes: emigration from the natal or residential group, movement between groups and home ranges, and immigration into a new target social group (Wolff, 1994; Bowler and Benton, 2005). Dispersal individuals often experience a stressful stage to face increased predation risks, restricted access to known food, and elevated attack from target group members (Alberts and Altmann, 1995a; Bonte et al., 2012; Marty et al., 2017a).
The causes of dispersal and the determination of successful dispersal are multifactorial (Greenwood, 1980; Alberts and Altmann, 1995b; Xia et al., 2019; L'Allier et al., 2022), whereas immigration is a critical and stressful process to be a long-term residency in a new social group (Marty et al., 2017b). Upon arrival in a new target social group, immigrant males faced elevations in aggression from intrasexual conspecifics of a new group (Ydenberg et al., 1988; Teichroeb et al., 2011). For example, in a study of male dispersal and female philopatric Japanese macaques (Macaca fuscata), Horiuchi (2005) found that the frequency of aggression between immigrant males and resident males was higher than that among resident males.
More and more evidence showed that immigrant males employed a range of social strategies to facilitate integration into a new social group for establishing stable dominant ranks and affiliative relationships with unfamiliar competitors who require them to adapt their behavior accordingly (Smale et al., 1997; Xia et al., 2021). Some strategies are species-specific. For example, in species living in a multilevel society (e.g., Rhinopithecus bieti and Rhinopithecus roxellana), individuals immigrated from a one-male unit to another within a social group (Xia et al., 2019; Fang et al., 2022). Some species formed parallel immigration with familiar or kin individuals to share coalition against resident males (e.g., Alouatta seniculus, Pope, 2000; Cebus capucinus, Schoof et al., 2009), whereas in some species dispersal individuals formed immigration alone (e.g., Macaca thibetana, Xia et al., 2021; Macaca nigra, Marty et al., 2017a). Immigration strategies were also individual-specific. Some individuals are natal dispersers (dispersal from the natal group to another), whereas some individuals are secondary dispersers (from the breeding group to another breeding group) (Pusey and Packer, 1987). Young immigrant males would likely to form short-duration immigration by immediately challenging a dominant male (Teichroeb et al., 2011), whereas secondary immigrant males were more likely to form slow and gradual immigration by avoiding intensive aggression and starting from lower social rank (Xia et al., 2021). Immigrant males with high fighting ability often challenged resident males to achieve a high social rank in the new group, whereas immigrants with lower fighting ability often achieve a lower social rank in the new group (Marty et al., 2016, 2017a).
van Noordwijk and van Schaik (1985) provided a criterion to distinguish two types of immigrant males, bluff and unobtrusive immigrants. According to this criterion, the bluff immigrants attempted to take over the dominant male of the new social group and were all young adult males, whereas unobtrusive immigrants entered with low social dominance rank and were males of all age classes (van Noordwijk and van Schaik, 1985). It indicated that immigrant males with high fighting ability and low fighting ability could be distinguished into the bluff and unobtrusive immigrants, respectively. However, limited by sample size (because immigration was a rare event for a certain social group), less was known whether there were variations of social strategies between bluff and unobtrusive immigrant males.
In this study, nine adult males were found to immigrate into the study group. It provided a case and natural experiment to investigate the variation of partnerships between bluff and unobtrusive immigrant males. Tibetan macaques (Macaca thibetana) are excellent primate model species to investigate the variation of social strategies between the bluff and unobtrusive immigrant males during integrating into a new social group. Male Tibetan macaques are known to disperse from one group to another (e.g., natal dispersal, secondary dispersal) several times in their life (Li et al., 2020). Males who achieve a high social rank upon immigration (i.e., bluff with high fighting ability) are mainly young adult males around the age of sexual maturity dispersing from their natal group for the first time (natal dispersal, Li et al., 2020). Middle-age or old adult immigrant males mainly achieve a low social rank upon immigration (i.e., unobtrusive with social experience and low fighting ability) during dispersing from one breeding group to another breeding group (secondary dispersal, personal observation, Dong-Po Xia), these males preferred to form a partnership with a small set of females and form a stable partnership with these resident females to facilitate integrating into a new social group (Xia et al., 2021). In this species, grooming accounts for approximately 20% of an individual's daily activity budget (see Xia et al., 2012 and Xia et al., 2013). Previous studies indicate that both male and female Tibetan macaques use grooming as an important social tool in forming and maintaining social relationships (Berman et al., 2008; Xia et al., 2012, 2013). Social bridging serves as a positive affiliative interaction, such as buffering agonistic interaction and strengthening social bonds, especially among male intrasexual dyads (Bauer et al., 2014; Kalbitz et al., 2017).
Within the framework of determining bluff and unobtrusive immigrant males according to van Noordwijk and van Schaik (1985), we hypothesized that bluff and unobtrusive immigrant males would form varied social strategies to integrate into a new social group. The following predictions were tested. If immigrants faced elevations in aggression from intrasexual conspecifics of a new group, then the amount of aggression received from resident males of the bluff and unobtrusive immigrant males would be higher than that from resident females and immigrants (Prediction 1). Accordingly, to decrease the potential social pressures from residents of the new social group, both bluff and unobtrusive immigrants would participate in more affiliative interaction (such as social grooming) with resident females than resident males (Prediction 2). If the partnerships with the high-ranking females were potentially beneficial to its competition for the high-ranking, then the bluff immigrants would engage in more social grooming with high-ranking resident females than that with low-ranking resident females during the integrating stage, whereas the unobtrusive immigrant males would engage in more social grooming with low-ranking resident females than that with high-ranking resident females (Prediction 3). If the partnership with high-ranking resident males represented social strategies to facilitate obtaining high-ranking for immigrant males, then the bluff immigrant males would participate more in social bridging with high-ranking resident males than unobtrusive immigrant males (Prediction 4). In addition, if the bluff immigrant males had a higher possibility to obtain higher social rank, then the frequency of copulation of the bluff immigrant males would be higher than the unobtrusive immigrant males (Prediction 5).
This study was conducted at the Mt. Huangshan National Reserve located in Anhui Province, China. Mt. Huangshan (118.3E, 30.2N, elevation 1841m) is a scenic area and tourist destination in east-central China, which consists of steep, sparsely treed peaks at high elevations and mixed deciduous and evergreen forests in the middle and lower elevations. The study site is the Valley of the Wild Monkeys, the Fuxi village, which is adjacent to Mt. Huangshan. More information about the study site can be found in Li et al. (2020). There are several groups of Tibetan macaques living throughout the reserve (Berman and Li, 2002).
The study group was the Yulinkeng A1 group (YA1), which was observed since 1986. The yearly demographic data have been collected since 1987. We can identify all the individuals according to their natural features (such as fur, facial features, body size, and body characteristics), including the resident males, resident females, and immigrant males. We can also identify the age classes of immigrant males according to the criterion from Li et al. (2020).
The study group was provisioned daily with 3–4 kg of corn by reserve staff to maintain their presence at designated tourist-viewing sites during 2004 and 2014, and thus it is habituated to humans (i.e., <1 m). Since 2015, the tourist-viewing sites were closed, and the study group was still provisioned by reserve staff in the same manner. After feeding, the monkeys continue their natural and undisturbed activities in the forest.
During the study period, there were 62 individuals, including 15 adult males, 12 adult females, 25 juveniles, and 10 infants. We defined an immigrant male as an adult male who had immigrated into the group and resided in the group for <2 years and a resident male as an adult male who had resided in the group for more than 2 years. A total of 15 adult males were in the study group during the study period, including 9 immigrant males and 6 resident males (see Table 1). The subject individuals were 15 adult males and 12 adult females.
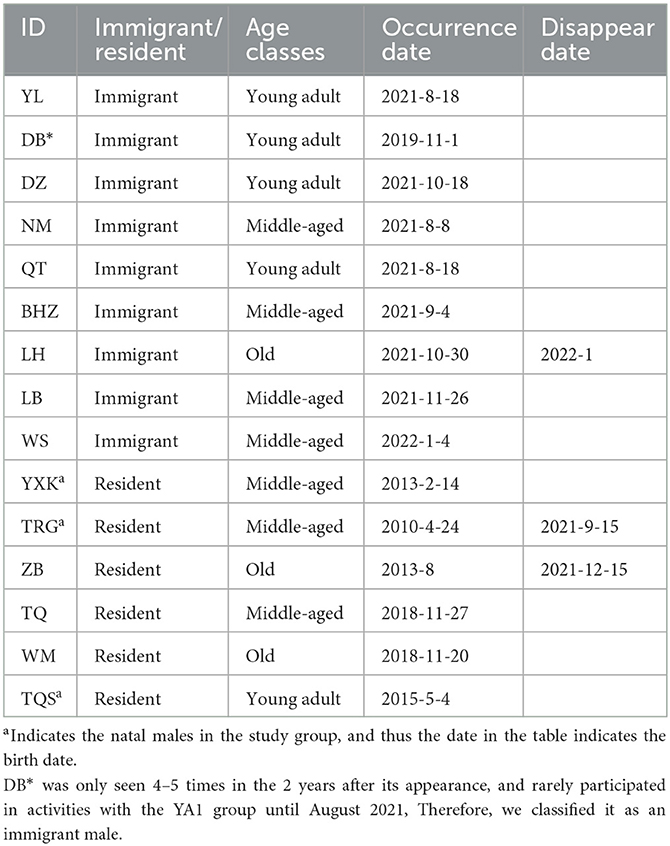
Table 1. The identity of adult males during the study period.
During August 2021 and May 2022, to collect the behavioral data, we followed the study group from dawn to dusk beginning at approximately 7 a.m. to 8 a.m. and ending at 5 p.m. to 6 p.m. each day (depending on the time of year). All the behavioral data were collected by a single observer (C.R.Z.) from a distance of 5–10 m from the monkeys. Focal animal sampling and continuous recording (using a digital voice recorder, model News my V19) were used to score the daily activities of all adult males (Altmann, 1974). The focal sample duration was set at 20 min (Xia et al., 2010, 2012). We collected all the behavioral data when the subjects were in the forest, away from the viewing platforms. If the focal animal could not be followed or was lost from view during the sampling interval, then another individual was randomly selected (Li et al., 2007; Shutt et al., 2007). We made an effort to locate and record the behavior of the lost adult macaque during the next 20 min sampling period (Li et al., 2005, 2007; Xia et al., 2010).
Focal sampling yielded a total of 175.66 h of data. There are differences in the time of appearance of immigrant males, and the sampling time of each individual is also different (11.71 ± 2.71 h, n = 15, range: 1–40.33 total h per individual). We defined grooming as any act in which an individual (groomer) used their hand or mouth to touch, clean, or manipulate the fur of another individual (groomer) for a continuous period lasting at least 5 s (Berman et al., 2008; Xia et al., 2012). For each bout, we recorded the identities of the participants and the time spent grooming and being groomed by each partner to the nearest second. We defined social bridging as two individuals simultaneously holding or picking up one infant (Ogawa, 1995; Li et al., 2020). Aggressive interactions were defined as an individual threatening, chasing, slapping, grabbing, or biting another individual (Berman et al., 2004). We defined submissive behaviors as an individual exhibiting a fearful response, such as a fearful grin, cowering, mock leaving, avoiding, fleeing, or screaming (Berman et al., 2004).
According to van Noordwijk and van Schaik (1985), we used the comprehensive standard (based on unprovoked fear, aggression, coalition against, and branch shaking) to categorize immigrant males into either bluff or unobtrusive individuals. Using ad libitum sampling, we collected approach and live, attacks, and branch shaking of immigrant and resident males. Unprovoked fear was defined as one male showing submission not preceded by aggression by the other. Aggression was defined as an individual threatening, chasing, slapping, grabbing, or biting another individual (Berman et al., 2004). Coalition against was defined as after individual attacks another individual, one or more individuals attack the same individual within 5 s. Branch shaking was defined as the male quickly moving to the branch and bobbing up and down. Accordingly, in this study, the three bluff males (YL, DB, and DZ) and six unobtrusive males (QT, NM, BHZ, LH, LB, and WS) were identified (see Table 2).
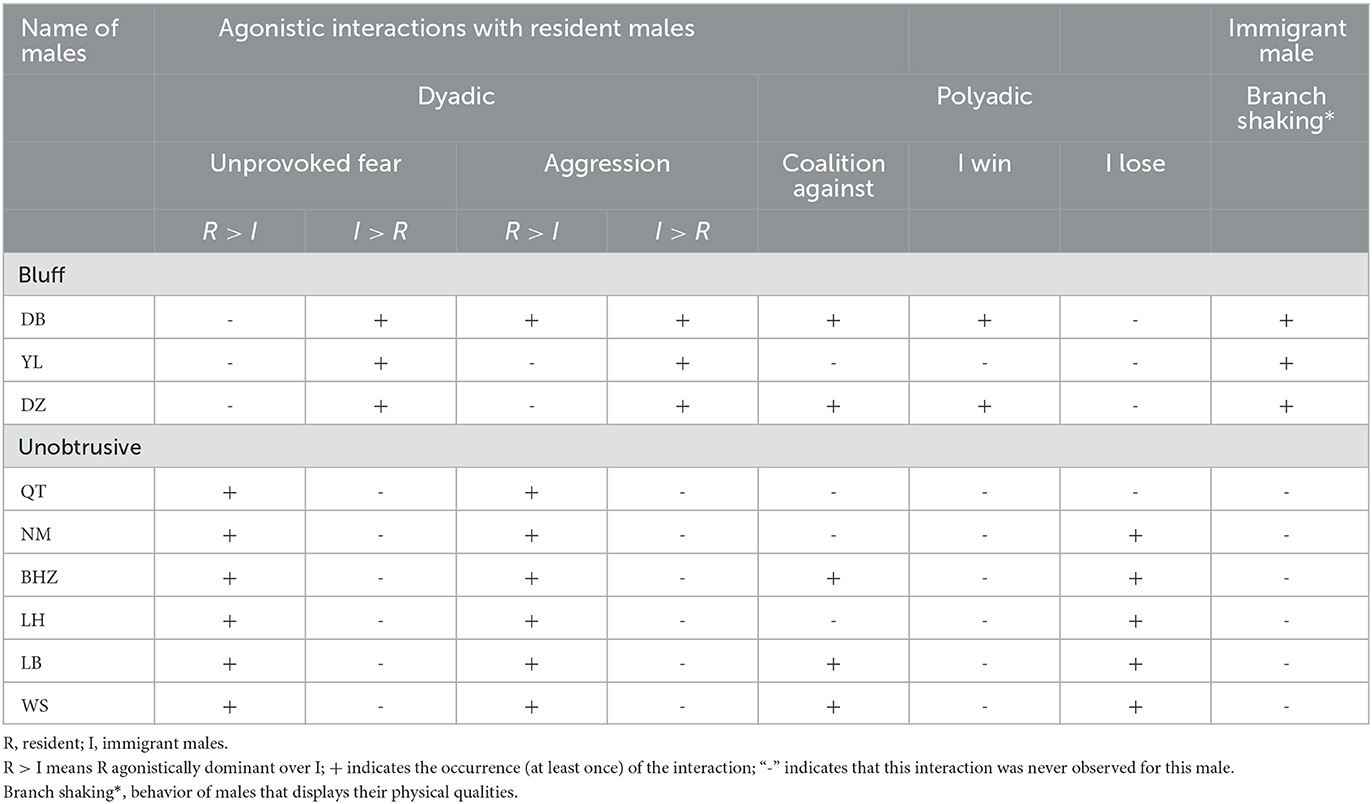
Table 2. Categorization of bluff and unobtrusive immigrant males.
Marty et al. (2016) used a change-point analysis to determine whether immigrant males were randomly integrated into a new social group or not. In crested macaques (Macaca nigra), based on the finding that immigration of a new male into a group occurred on average 81 days, a change-point analysis was used to determine the immigration occurred non-randomly with 74% of all male immigrants with 25 days of another male (before or after). Within the framework of this idea, we calculated affinity based on the amount of grooming duration and used this analysis method to separate the prospecting stage and integrating stage based on the weekly variation of affinity during integration into a new social group. A changepoint analysis (Change Point Analyzer 2.3; Taylor Enterprises, Inc.) was conducted to reveal significant changes in the mean squared error distribution of male immigration. 10,000 boot-straps with replacements were run and the confidence interval was set at 99% (Marty et al., 2016).
For example, an immigrant male YL was first observed in the subject social group on August 18th, and the number of affinities increased since the second week of October 2021 (about October 10th). Based on the changing pattern of individual affinity, we separated the immigrant period of YL into the prospecting stage (from the third week of August 2021 to the second week of October 2022) and the immigrant stage (from the third week of October 2021 to the fourth week of May 2022). For another immigrant male WS, although the amount of grooming duration decreased, the number of affinities did not show variation since the first observed day (January 4th, 2022) to the ending date of this study. As such, we clarified the prospecting stage of WS as the first week of January 2022 and the fourth week of May 2022. The more detail of prospecting stage and integrating stage of all nine immigrant males can be found in Table 3. All the analysis results and the figures can be found in the Supplement material (Result of Change-Point Analyzer).
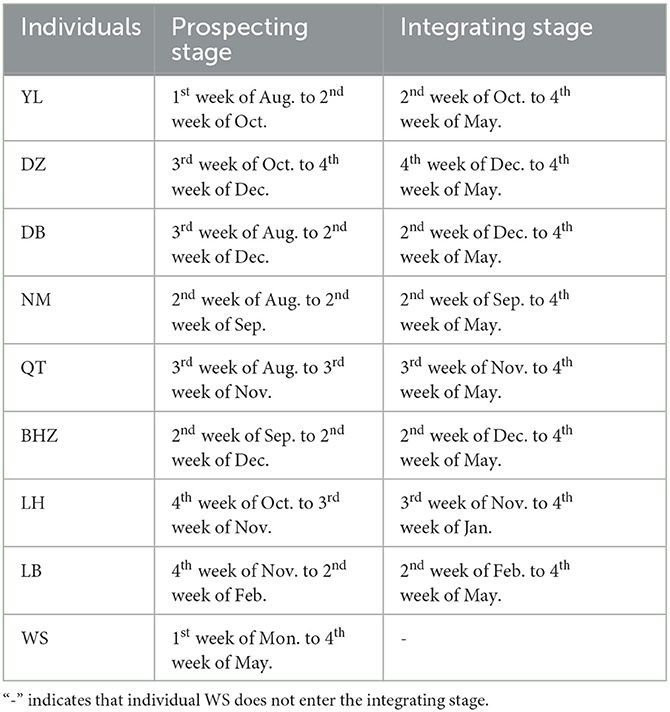
Table 3. Prospecting and integrating stage of immigrant males.
Using ad libitum sampling, we collected the number and direction of dyadic aggressive and submissive behaviors among intrasexual dyads (either male-male dyads or female-female dyads). We scored the total number of dyadic aggressive and submissive interactions and built an aggressive/submissive matrix according to the direction of agonistic interactions given and received. Based on this matrix, we calculated the standard David's Score (DS) to determine the individual social rank of adult males and adult females, respectively (Gammell et al., 2003; de Vries et al., 2006). The higher the DS value, the higher the social rank. Aggressive interactions were defined as an individual threatening, chasing, slapping, grabbing, or biting another individual (Berman et al., 2007). We defined submissive behaviors as an individual exhibiting a fearful response, such as a fearful grin, cowering, mock leaving, avoiding, fleeing, or screaming, according to Berman et al. (2004).
We classify each resident adult male into the following rank classes: high ranking (YXK, ZB), and low ranking (WM, TQ, TQS). We also classify each resident adult female into the following rank classes: high-ranking (YXX, YH, YCY, YXY, YCH, TXH, and TQL), and low-ranking (TH, YCL, TXX, THY, THX, TQY, TFH, and TQG) by used k-means cluster analysis (100 iterations).
All data were calculated as the mean (±SE) of grooming frequency (bouts/h), grooming duration (min/h), bridging frequency (episodes/h), and aggressive frequency (episodes/h). We calculated the grooming frequency as the number of bouts a target individual initiates or receives grooming from another individual per hour (Xia et al., 2012). We calculated the grooming duration as the total number of minutes of grooming per pair per hour (Xia et al., 2012). We calculated the aggressive as the number of bouts a target individual initiates or receives aggressively from another individual per hour (Berman et al., 2007).
Firstly, the generalized linear mixing models (GLMMs) was used to explore which factors were related to the strategy of immigrant males integrate into a new group. To test the prediction 1, resident gender was included as a fixed factor and individual social rank and age of residents were treated as random factors. To test the prediction 2, resident gender was included as a fixed factor and individual social rank and age of residents were treated as random factors. To test the prediction 3, resident social rank was included as a fixed factor and resident age was treated as random factors. To test the prediction 4, resident social rank was included as a fixed factor and resident age was treated as random factors.
When we found there was only one predictor to influence the variable, we used a univariate analysis for further analysis. We used the Mann-Whitney U-test and Friedman test to analyze differences in the amount of aggression received from resident males, resident females, and immigrants during both the prospecting and integrating stages (testing Prediction 1). We used the Wilcoxon matched-pairs signed-ranks to analyze the difference in grooming initiated by the bluff and unobtrusive males to resident males or resident females during the prospecting and integrating stages (testing Prediction 2). We used the Wilcoxon matched-pairs signed-ranks test to analyze the difference in social grooming initiated by the bluff and unobtrusive immigrant males to resident high-ranking vs. low-ranking females (testing Prediction 3). We used the Mann-Whitney U-test and Kruskal-Wallis to analyze the differences in frequency of social bridging the bluff and unobtrusive immigrant males initiated with high-ranking and low-ranking resident males (testing the Prediction 4). All analyses were two-tailed and analyses were performed using SPSS 27.0 (SPSS Inc., Norusis, 2005) and R software with a prespecified significance level of 0.05.
In addition, we used GLMM to test whether the type of immigrant males (e.g., the bluff and unobtrusive) related to mating success (testing Prediction 5). In this model, the type of immigrant males (categories: the bluff immigrant male, the unobtrusive immigrant male) was included as a fixed factor, whereas the male social rank, the type of males (categories: immigrant male, resident male), age class were treated as random factors. We ran the GLMM in R 4.1.3 using the function Elmer of the R package lme4 (Bates et al., 2012).
The GLMM model showed that the gender of resident adults was a positive predictor of the aggression frequency that immigrant males received from resident adults (estimate ± SE = 5.127 ± 1.076, Z = 4.766, P < 0.001). The effects of age and individual social rank of resident adults was excluded (estimate ± SE = −0.050 ± 0.117, Z = –0.433, P = 0.67 for age; estimate ± SE = −0.160 ± 0.955, Z = –0.168, P = 0.87 for social rank). As indicated in Figure 1 (left), during the prospecting stage, for both the bluff and unobtrusive immigrant males, the amount of aggression received from resident males (bluff: 0.30 ± 0.15 episodes/h; unobtrusive: 1.43 ± 0.27 episodes/h) was higher than that from resident females (bluff: 0.03 ± 0.03 episodes/h; unobtrusive: 0.07 ± 0.04 episodes/h) and immigrant males (bluff: 0.27 ± 0.14 episodes/h; unobtrusive: 0.13 ± 0.06 episodes/h) (χ2 = 11.677, P = 0.003). The bluff immigrant males were lower frequently attacked by resident males than the unobtrusive immigrant males (Z = –2.324, P < 0.05). Similarly, during the integrating stage, as indicated in Figure 1 (right), for both the bluff and unobtrusive immigrant males, the amount of aggression received from resident males (bluff: 0.12 ± 0.06 episodes/h, unobtrusive: 1.09 ± 0.19 episodes/h) was higher than that from resident females (bluff: 0 episodes/h; unobtrusive: 0.20 ± 0.07 episodes/h) and immigrant males (bluff: 0.15 ± 0.12 episodes/h; unobtrusive: 0.14 ± 0.08 episodes/h) (χ2 = 11.032, P = 0.004). The bluff immigrant males were lower frequently attacked by resident males than the unobtrusive immigrant males (Z = –2.324, P < 0.05).
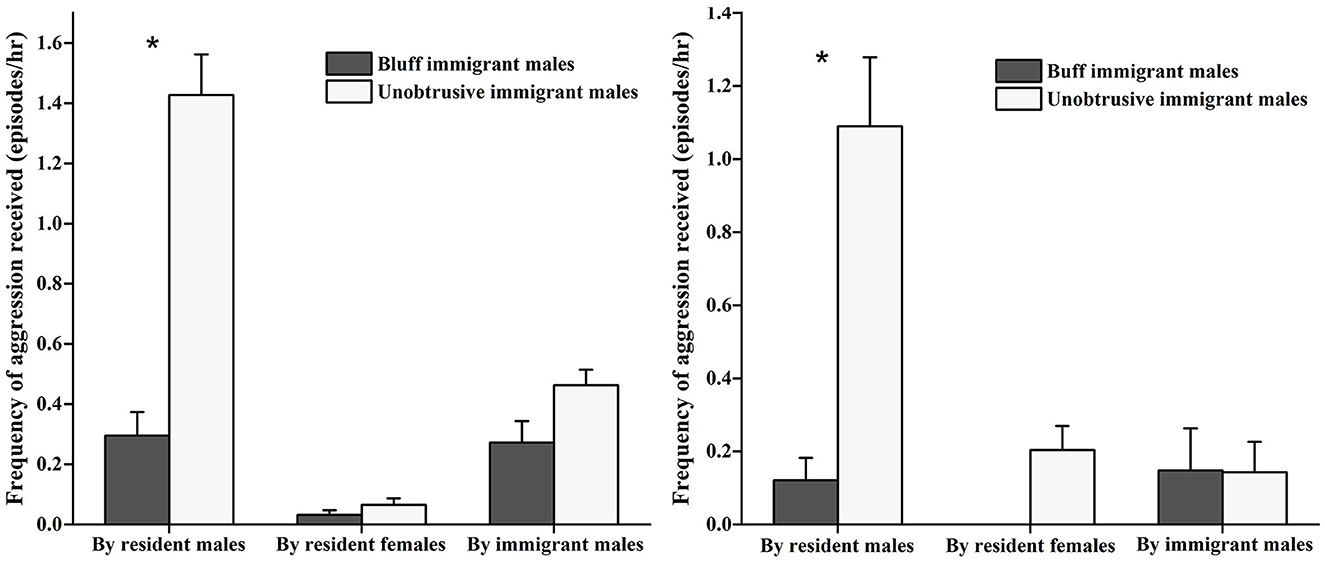
Figure 1. Frequency of aggression immigrants received during the prospecting (left) and integrating stage (right). *means that the aggression by resident males to bluff and unobtrusive immigrant males was significant difference.
The GLMM model showed that the gender of resident adults was a positive predictor of the grooming frequency which immigrant males initiated to the resident adults (estimate ± SE = −4.014 ± 0.639, Z = –6.286, P < 0.001). The effect of the age of resident adults was excluded (estimate ± SE = −0.007 ± 0.057, Z = –0.509, P = 0.61). As shown in Figure 2, during the prospecting stage, we found that the amount of grooming initiated by the bluff immigrant males to resident females (frequency: 0.03 ± 0.01) was higher than that to resident males (frequency: 0) (Z = –2.116, P < 0.05 for the grooming frequency; Z = –2.114, P < 0.05 for the grooming duration). However, there was no diffident in grooming amount initiated by the unobtrusive immigrant males to resident females and males (Z = 0, P > 0.05 for grooming frequency; Z = –0.116, P > 0.05 for the grooming duration). During the integrating stage, we found that the amount of grooming initiated by the bluff immigrant males to resident females (frequency: 0.19 ± 0.07 bouts/h; duration: 0.78 ± 0.26 min/h) was higher than that to resident males (frequency: 0.03 ± 0.01 bouts/h; duration: 0.06 ± 0.03 min/h, Z = –3.517, P < 0.001 for grooming frequency, Z = –3.517, P < 0.001 for grooming duration). Similarly, the amount of grooming initiated by the unobtrusive immigrants to resident females (frequency: 0.23 ± 0.06 bouts/h; duration: 1.07 ± 0.26 bouts/h) was higher than that of resident males (frequency: 0.04 ± 0.03 bouts/h; duration: 0.11 ± 0.07 min/h, Z = –3.629, P < 0.001 for grooming frequency, Z = –3.483, P < 0.001 for grooming duration).
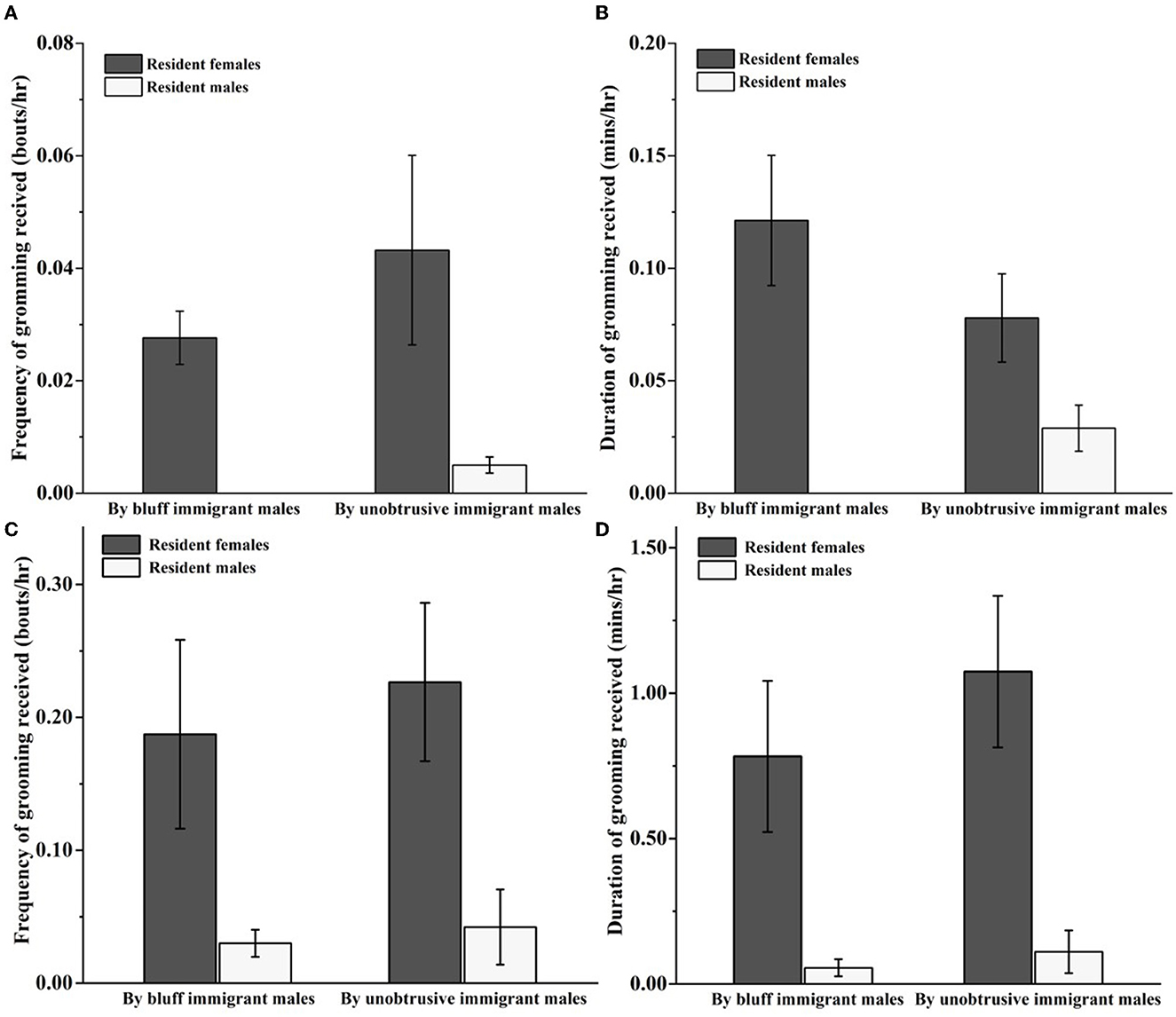
Figure 2. Grooming between immigrants and resident adults during the prospecting (A, B) and integrating stage (C, D).
The GLMM model showed that the social rank of resident females was a positive predictor of the grooming frequency which bluff/unobtrusive immigrant males initiated to the resident females (estimate ± SE = −0.181 ± 0.060, Z = –3.031, P = 0.002 for bluff immigrant males; estimate ± SE = 0.134 ± 0.065, Z = 2.055, P = 0.04 for unobtrusive males). The effect of age of resident females was excluded (estimate ± SE = −0.044 ± 0.059, Z = –0.737, P = 0.46 for bluff immigrant males; estimate ± SE = 0.064 ± 0.066, Z = 0.975 P = 0.33 for unobtrusive immigrant males). As shown in Figure 3, during the prospecting stage, there was no difference in the amount of social grooming initiated by the bluff immigrant males to high-ranking resident females (frequency: 0.01 ± 0.004, duration: 0.02 ± 0.01) and low-ranking females (frequency: 0.04 ± 0.02, duration: 0.21 ± 0.11; Z = –1.594, P >0.05 for grooming frequency; Z = –1.532, P > 0.05 for grooming duration). Similarly, there was also no difference in the amount of social grooming initiated by the unobtrusive immigrant males to high-ranking resident females (frequency: 0.07 ± 0.07, duration: 0.33 ± 0.31) and low-ranking females (frequency: 0.02 ± 0.01, duration: 0.07 ± 0.04; Z = –0.344, P >0.05 for grooming frequency; Z = –0.207, P > 0.05 for grooming duration). During the integrating stage, the amount of grooming initiated by the bluff immigrant males to high-ranking resident females (frequency: 0.10 ± 0.02 bouts/h, duration: 0.46 ± 0.11 min/h) was higher than those initiated to low-ranking resident females (frequency: 0.03 ± 0.01 bouts/h, duration: 0.10 ± 0.03 min/h; Z = – 3.008, P = 0.003 for grooming frequency, Z = –3.258, P < 0.001 for grooming duration). The amount of grooming initiated by the unobtrusive immigrant males to high-ranking resident females (frequency: 0.05 ± 0.02 bouts/h, duration: 0.11 ± 0.03 min/h) was lower than those initiated to low-ranking resident females (frequency: 0.15 ± 0.03 bouts/h, duration: 0.82 ± 0.23 min/h, Z = –3.258, P < 0.001 for grooming frequency; Z = –3.774, P < 0.001 for grooming duration).
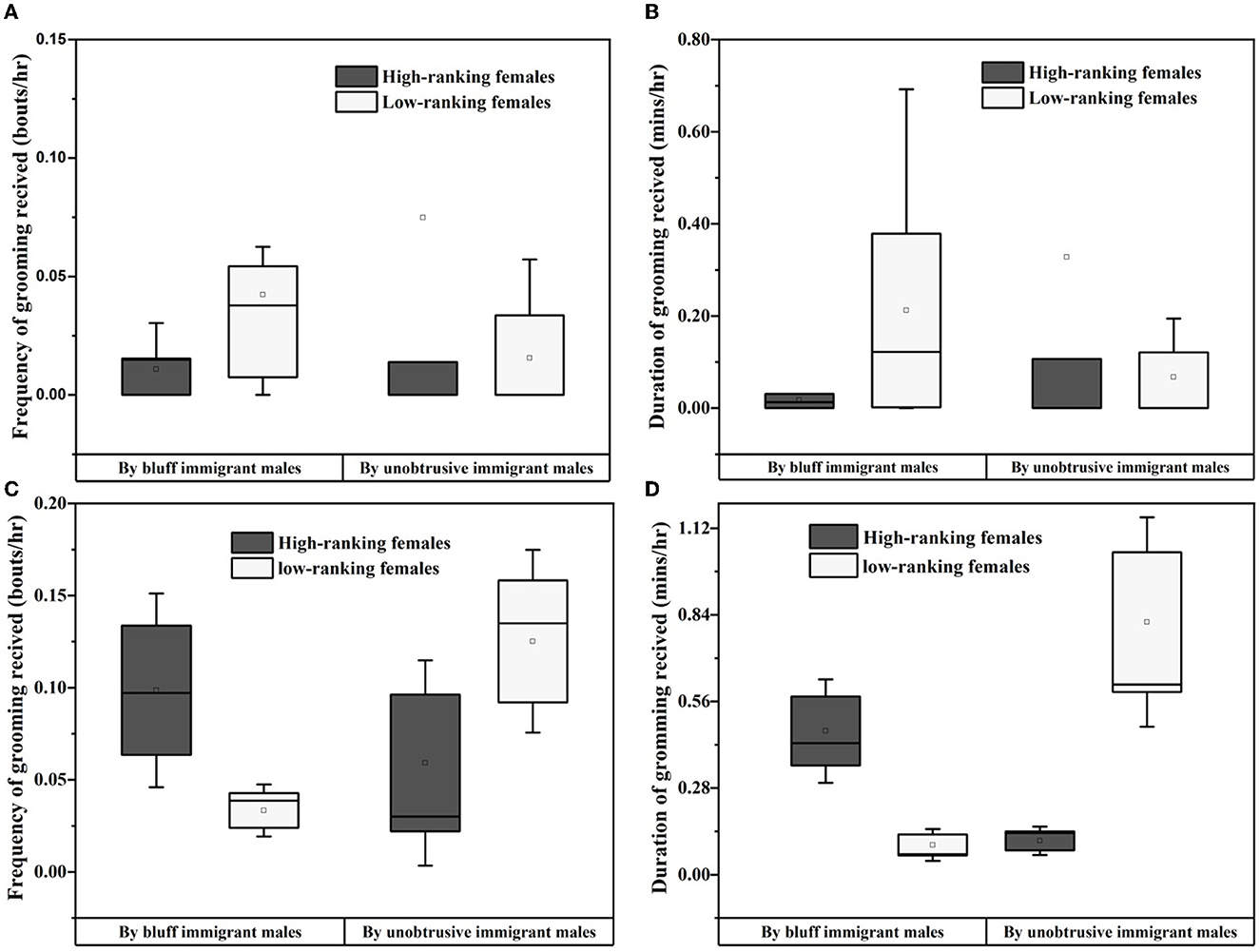
Figure 3. Grooming between immigrant males and resident females during the prospecting (A, B) and integrating stage (C, D).
The GLMM model showed that the social rank of resident males was a positive predictor of the bridging frequency which immigrant males initiated to the resident males (estimate ± SE = −0.478 ± 0.115, Z = –4.142, P < 0.001). The effect of the age of resident males was excluded (estimate ± SE = −0.070 ± 0.054, Z = –1.296, P = 0.20). As indicated in Figure 4, during the prospecting stage, the frequency of social bridging to resident males initiated by the bluff immigrant males (0.02 ± 0.01 episodes/h) was higher than the unobtrusive immigrant males (0 episodes/h) (Z = –2.207, P < 0.05). However, the frequency of social bridging initiated by the bluff immigrant males was found no difference among different types of resident males (resident high-ranking males: 0.11 ± 0.09 episodes/h, resident low-ranking males: 0.02 ± 0.01 episodes/h, the bluff immigrant males: 0.02 ± 0.01 episodes/h, the unobtrusive immigrant males: 0 episodes/h, H = 7.463, P > 0.05). During the integrating stage, the frequency of social bridging initiated by the bluff immigrant males to resident males (0.11 ± 0.04 episodes/h) was higher than the unobtrusive immigrant males (0.01 ± 0.01 episodes/h, Z = –3.298, P < 0.001). The frequency of social bridging initiated by the bluff immigrant males to resident high-ranking males (0.42 ± 0.18 episodes/h) was higher than that of other adult males (resident low-ranking males: 0.11 ± 0.03 episodes/h, the bluff immigrant males: 0.07 ± 0.03 episodes/h, the unobtrusive immigrant males: 0 episodes/h, H = 27.172, P < 0.001).
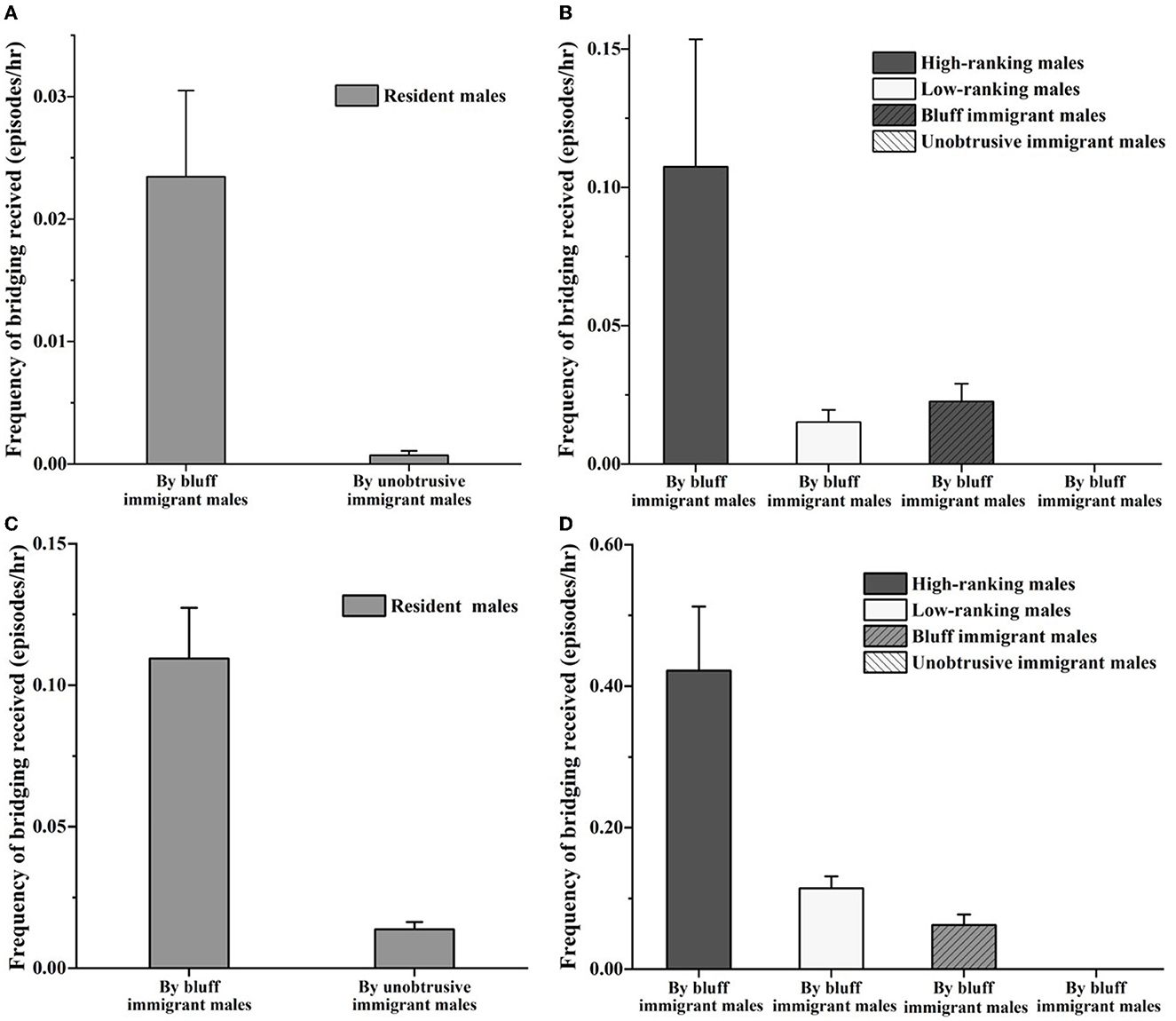
Figure 4. Frequency of bridging between immigrant males and resident males during the prospecting (A, B) and integrating stage (C, D).
The results showed that the type of immigrant males (e.g., the bluff immigrant males, the unobtrusive immigrant males) was a positive predictor of the frequency of copulation. The bluff immigrant males engaged in a higher frequency of copulation. The results also showed a positive effect of male social rank on the frequency of copulation. The higher the social rank, the more frequency of copulation (Table 4).
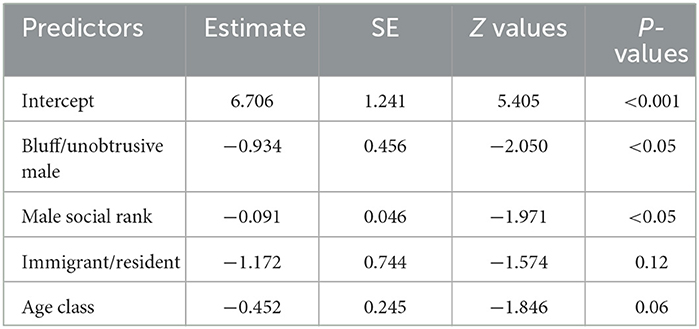
Table 4. Results of the generalized linear mixed model testing whether the type of immigrant males was related to the copulation frequency.
In this case study, we firstly investigated the variation of social strategies between the bluff and unobtrusive immigrant males during both prospecting and integrating stages of integration into a new social group in male dispersal and female philopatric Tibetan macaques. The results showed that, for both the bluff and unobtrusive immigrant males, aggressions received from resident males were higher than that from resident females during both the prospecting and integrating stages. It indicated that immigrant males experienced similar stressful stages to face increased aggression from intrasexual conspecifics from a new social group. However, during the integrating stage, the bluff immigrant males formed higher frequency and longer duration social grooming with high-ranking resident females, whereas the unobtrusive immigrant males formed higher frequency and longer duration social grooming with low-ranking females. Moreover, the bluff immigrant males engaged in higher frequency with high-ranking resident males than the unobtrusive immigrant males during the integrating stage. Our results suggested that, compared to the unobtrusive immigrant males, the bluff immigrant males employed varied social strategies for establishing affiliative relationships with unfamiliar competitors who require them to adapt their behaviors and partnerships during integration into a new social group.
Previous studies proposed that integration into a new social group was the last process of dispersal and had been considered as a great challenge for dispersal individuals, all the dispersal individuals had to experience this process. Our results found that, upon arrival in a new target social group, both the bluff and unobtrusive immigrant males faced elevations in aggressions from intrasexual conspecifics from new group. It provided supportive evidence that immigrant males were frequently attacked by resident males in macaque society (Bercovitch, 1997; van Schaik and van Noordwijk, 2001; Schoof et al., 2009; Port and Johnstone, 2013; Georgiev et al., 2016). It suggested that, during integration into a new social group, forming new partnerships with resident males could be stressful and the presence of new males could represent an increase in male–male competition for immigrant males. Thus, females' acceptance and male-female grooming had positive effects in male integration strategies (Kawazoe and Sosa, 2019; Xia et al., 2021). To facilitate the integration, the alternative strategies was to prefer to form partnerships with resident females and form stable partnerships with a small set of resident females (e.g., Macaca fuscata, Horiuchi, 2005; Macaca thibetana, Xia et al., 2021). Our results provided further evidence that, either during the prospecting or integrating stages, both the bluff and unobtrusive immigrant males engaged more frequently or longer duration of social grooming with resident females in a new group. In terms of immigration, to some extent, all the immigrant individuals might experience the similar social pressure and form strong affiliative relationships with resident females during integration into a new social group, regardless the type of immigrants (e.g., the bluff immigrants or unobtrusive immigrants), the stage of immigration (e.g., the prospecting stage or integrating stage).
Furthermore, our results provided quantitative criteria, based on the change of social relationships, to separate gradual processes of integration into a new social group as prospecting and integrating stages. Tenue length (e.g., the first day a male was seen in a new group in Macaca mulatta, Wei et al., 2016; 30 days in Chlorocebus pygerythrus, L'Allier et al., 2022, 2 years in Macaca thibetana, Xia et al., 2021) was used to determine the immigrant males in a new social group. However, previous studies more or less neglected the variation of social strategies between the prospecting stage and the integrating stage during the consecutive immigration processes. Our results first proposed the social strategies of immigrant males during the prospecting stage. To locate the target new group, immigrant males were preceded by males “visiting” or “prospecting” other groups for up to several days or longer duration before their emigration (such as Chlorocebus pygerythrus and Macaca thibetana, Cheney and Seyfarth, 1983; Li et al., 2020). Upon arrival at a new social group, the first step is to determine when and how they should attempt to interact with group members of new social group. The more important is that immigrant males might have sufficient ability to decide which kinds of individuals would be potential partners and how to promote relationships with those partners based on their characteristics. In this study, our results showed that there was no difference in the amount of social grooming initiated by both the bluff and unobtrusive immigrant males to high-ranking resident females and low-ranking females during the prospecting stage. It indicates that to seek a target social partner and establish a social relationship with unfamiliar individuals, both the bluff and unobtrusive immigrant males might form “attempt strategies” to interact and establish social relationships with members in a target social group at the beginning of the arrival of a new group. Moreover, during the prospecting stage, although the frequency of social bridging to resident males initiated by the bluff immigrant males was higher than the unobtrusive immigrant males, the frequency of social bridging initiated by the bluff immigrant males was found no difference among different types of resident males. It suggests that, compared to the unobtrusive immigrant males, the prospecting stage might be the beginning to form varied social strategies for the bluff immigrant males.
In addition, our results suggested that immigration into a new social group was an individual-specific process, especially during the integrating stage. Successful immigration strategies involved in the consecutive integration processes represent complex processes that are also contained by individual-specific characteristics. It includes the costs of reproductive competition, the benefits of social cooperation, the advantages of collective action, and the strength of female mate choice within a new social group. Compared to immigration strategies in other species, such as parallel immigration in white-faced capuchins (Cebus capucinus, Jack and Fedigan, 2004a,b) and Japanese macaques (Macaca fuscata, Kawazoe and Sosa, 2019), Tibetan macaques were generally found to disperse as lone or solitary individuals (Li et al., 2020). Thus, individual-specific social strategies would be more important in this species. Previous studies suggested that several factors influence individual social strategies, including the age of the immigrant males (Teichroeb et al., 2011), fighting ability (Marty et al., 2016), and mother ranking (Wei et al., 2016). In Tibetan macaque society, previous case studies showed that some young immigrant males with high fighting ability often challenge and replace the alpha male by engaging in short periods of severe aggression (Li et al., 2020), whereas middle-aged or old immigrant males with limited fighting ability preferred to form stable partnerships with a small set of resident females to facilitate integration into a new social group (Xia et al., 2021). In this study, our results show that, during the integrating stage, the bluff immigrant males mainly form partnerships with resident high-ranking females and males, whereas the unobtrusive immigrant males mainly form partnerships with resident low-ranking females. It provided positive evidence of individual-specific social strategies. It also suggested that the integrating stage was a more important phase than the prospecting stage. Among all the nine immigrant males, the bluff immigrant males were mainly young adult males (7–10 years of age) and showed stronger aggression, such as branch shaking. However, the unobtrusive immigrant males are mainly other middle or old males and show less aggression. Based on the attempted strategies during the prospecting stage, differentiated strategies emerged between the bluff immigrant males and the unobtrusive immigrant males. It suggests that immigrant males can adjust their integration strategies according to their physical characteristics, and attempt strategies with resident individuals during the prospecting stage.
Finally, we also found that the types of immigrant males (e.g., the bluff and unobtrusive) and social rank were the positive predictors of the mating opportunity. It indicated that the bluff immigrant males would be likely to have more mating opportunities, whereas the unobtrusive immigrant males would have fewer mating opportunities. However, previous studies argued that immigration strategies were driven to a greater extent in an attempt to avoid costs rather than to obtain potential benefits (e.g., Macaca nigra, Marty et al., 2016). The alternative explanation was that the bluff immigrant males established affiliative relationships with high-ranking resident males and obtained high-ranking, whereas the unobtrusive immigrant males often had low rankings (e.g., Young et al., 2014; Biggs et al., 2021). Future studies need to pay greater attention to the benefits strategies used by different immigrant males, such as reproductive success and individual fitness during the integration and when settling in a new social group.
In conclusion, the present case study provided quantitative methods and new insight into distinguishing prospecting and integrating stages during immigration processes. It also firstly demonstrated, during the integrating stage, the bluff immigrant males formed affiliative relationships with high-ranking females and resident males whereas the unobtrusive immigrant males formed affiliative relationships with low-ranking females. Our results provided new insight into understanding the individual-specific social strategies of immigrant males during integration into a new social group.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The study was solely observational, involving no interference with the animals and thus no review from the Ethics Committee in China was required. We collected the behavioral data according to the rules of the Wildlife Protection Law of the People's Republic of China and all research reported here adhered to the regulatory requirements of the Huangshan Garden Forest Bureau, China, where the study was conducted. The observational protocols adhered to the rules and regulations published in Frontiers in Ecology and Evolution for the ethical treatment of primates.
Designed the study: D-PX. Performed the experiments and contributed analysis: C-RZ, S-WC, H-WT, and Q-XZ. Contributed to writing and editing the manuscript: C-RZ, S-WC, and D-PX. All authors contributed to the article and approved the submitted version.
This study was supported in part by grants from the National Natural Science Foundation of China (Nos. 32070455 and 31772475), the Anhui Provincial Natural Science Foundation (2108085Y12), and the Special fund for Anhui Provincial Central Government to Guide Local Science and Technology Development (2019b11030018).
We are very grateful to the Huangshan Garden Forest Bureau for their permission and support of this study. We gratefully acknowledge H.B. Cheng's family for their outstanding logistic support of our study and J.H. Li's supervision and suggestions to highly improve this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1144932/full#supplementary-material
Alberts, S. C., and Altmann, J. (1995a). Balancing costs and opportunities: dispersal in male baboons. Am. Nat. 145, 279–306. doi: 10.1086/285740
Alberts, S. C., and Altmann, J. (1995b). Maturity of age at reproductive determinants and activation: preparation in male baboons. Behav. Ecol. Sociobiol. 36, 397–406. doi: 10.1007/BF00177335
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour 49, 227–267. doi: 10.1163/156853974X00534
Bates, D., Maechler, M., and Bolker, B. (2012). lme4: Linear Mixed-Effects Models Using S4 Classes. R package version 0.999999-0. Available online at: http://CRAN.R-project.org/package=lme4
Bauer, B., Sheeran, L. K., Matheson, M. D., Li, J. H., and Wagner, R. S. (2014). Male Tibetan macaques' (Macaca thibetana) choice of infant bridging partners. Zool. Res. 35, 222–230. doi: 10.11813/j.issn.0254-5853.2014.3.222
Bercovitch, F. B. (1997). Reproductive strategies of rhesus macaques. Primates 38, 247–263. doi: 10.1007/BF02381613
Berman, C. M., Ionica, C., and Li, J. H. (2004). Dominance style among Macaca thibetana on Mt. Huangshan, China. Int. J. Primatol. 25, 1283–1312. doi: 10.1023/B:IJOP.0000043963.77801.c3
Berman, C. M., Ionica, C., and Li, J. H. (2007). Supportive and tolerant relationships among male Tibetan macaques at Huangshan, China. Behaviour 144, 631–661. doi: 10.1163/156853907781347790
Berman, C. M., and Li, J. H. (2002). Impact of translocation, provisioning and range restriction on a group of Macaca thibetana. Int. J. Primatol. 23, 383–397. doi: 10.1023/A:1013891730061
Berman, C. M., Ogawa, H., Ionica, C., Yin, H. B., and Li, J. H. (2008). Variation in kin bias over time in a group of Tibetan macaques at Huangshan, China: contest competition, time constraints or risk response? Behaviour 145, 863–896. doi: 10.1163/156853908784089252
Biggs, N., Gartland, K. N., and White, F. J. (2021). Personality and alternative mating strategy in male Japanese macaques (Macaca fuscata). Am. J. Phys. Anthropol. 174, 10–10.
Bonte, D., van Dyck, H., Bullock, J. M., Coulon, A., Delgado, M., Gibbs, M., et al. (2012). Costs of dispersal. Biol. Rev. 87, 290–312. doi: 10.1111/j.1469-185X.2011.00201.x
Borries, C. (2000). Male Dispersal and Mating Season Influxes in Hanuman Langurs Living in Multi-Male Groups. Cambridge: Cambridge University Press.
Bowler, D. E., and Benton, T. G. (2005). Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamics. Biol. Rev. 80, 205–225. doi: 10.1017/S1464793104006645
Cheney, D. L., and Seyfarth, R. M. (1983). Nonrandom dispersal in free-ranging vervet monkeys: social and genetic consequences. Am. Nat. 122, 392–412. doi: 10.1086/284142
Clutton-Btock, T. H., and Lukas, D. (2012). The evolution of social philopatry and dispersal in female mammals. Mol. Ecol. 21, 472–492. doi: 10.1111/j.1365-294X.2011.05232.x
de Vries, H., Stevens, J. G., and Vervaecke, H. (2006). Measuring and testing the steepness of dominance hierarchies. Anim. Behav. 71, 585–592. doi: 10.1016/j.anbehav.2005.05.015
Fang, G., Jiao, H. T., Wang, M. Y., Huang, P. Z., Liu, X. M., Qi, X. G., et al. (2022). Female demographic changes contribute to the maintenance of social stability within a primate multilevel society. Anim. Behav. 192, 101–108. doi: 10.1016/j.anbehav.2022.07.018
Gammell, M. P., De-Vries, H., Jennings, D. J., Carlin, C. M., and Hayden, T. J. (2003). David's score: a more appropriate dominance ranking method than Clutton-Brock et al.'s index. Anim. Behav. 66, 601–605. doi: 10.1006/anbe.2003.2226
Georgiev, A. V., Christie, D., Rosenfield, K. A., Ruiz-Lambides, A. V., Maldonado, E., Thompson, M. E., et al. (2016). Breaking the succession rule: the costs and benefits of an alpha-status take-over by an immigrant rhesus macaque on Cayo Santiago. Behaviour 153, 325–351. doi: 10.1163/1568539X-00003344
Greenwood, P. J. (1980). Mating systems, philopatry and dispersal in birds and mammals. Anim. Behav. 28, 1140–1162. doi: 10.1016/S0003-3472(80)80103-5
Horiuchi, S. (2005). Affiliative relations among male Japanese macaques (Macaca fuscatayakui) within and outside a troop on Yakushima Island. Primates 46, 191–197. doi: 10.1007/s10329-005-0131-2
Jack, K. M., and Fedigan, L. (2004a). Male dispersal patterns in white-faced capuchins, Cebus capucinus, part 1: patterns and causes of natal emigration. Behaviour 67, 761–769. doi: 10.1016/j.anbehav.2003.04.015
Jack, K. M., and Fedigan, L. (2004b). Male dispersal patterns in white-faced capuchins, Cebus capucinus, part 2: patterns and causes of secondary dispersal Behaviour 67, 771–782. doi: 10.1016/j.anbehav.2003.06.015
Kalbitz, J., Schülke, O., and Ostner, J. (2017). Triadic male-infant-male interaction serves in bond maintenance in male Assamese macaques. PLoS ONE 12, e0183981. doi: 10.1371/journal.pone.0183981
Kawazoe, T., and Sosa, S. (2019). Social networks predict immigration success in wild Japanese macaques. Primates 60, 213–222. doi: 10.1007/s10329-018-0702-7
L'Allier, S., Schwege, M. A., Filazzola, A., Mastromonaco, G., Chapman, C. A., and Schoof, V. A. M. (2022). How individual, social, and ecological conditions influence dispersal decisions in male vervet monkeys. Am. J. Primatol. 84, e23426. doi: 10.1002/ajp.23426
Li, J.-H., Kappeler, P. M., and Sun, L. (2020). The Behavioral Ecology of the Tibetan Macaque. New York, NY: SpringerLink.
Li, J.-H., Yin, H.-B., and Zhou, L. Z. (2007). Non-reproductive copulation behaviour among Tibetan macaques (Macaca thibetana) at Huangshan, China. Primates 48, 64–72. doi: 10.1007/s10329-006-0002-5
Li, J.-H., Yin, H. B., and Wang, Q. S. (2005). Seasonality of reproduction and sexual activity in female Tibetan macaques (Macaca thibetana) at Huangshan, China. Acta Zoologica Sinica 51, 365–375.
Marty, P. R., Hodges, K., Agil, M., and Engelhardt, A. (2016). Determinants of immigration strategies in male Crested macaques (Macaca nigra). Sci. Rep. 6, 32028. doi: 10.1038/srep32028
Marty, P. R., Hodges, K, Heistermann, M., Agil, M., and Engelhardt, A. (2017a). Alpha male replacement and delayed dispersal in crested macaques (Macaca nigra). Am. J. Primatol. 79, e22448. doi: 10.1002/ajp.22448
Marty, P. R., Hodges, K, Heistermann, M., Agil, M., and Engelhardt, A. (2017b). Is social dispersal stressful? A case study in male crested macaques (Macaca nigra). Horm. Behav. 87, 62–68. doi: 10.1016/j.yhbeh.2016.10.018
Norusis, M. (2005). SPSS 13.0 Advanced Statistical Procedures Companion. Hoboken, NJ: Prentice Hall.
Ogawa, H. (1995). Recognition of social relationships in bridging behavior among Tibetan macaques (Macaca thibetana). Am. J. Primatol. 35, 305–310. doi: 10.1002/ajp.1350350406
Perrin, N., and Goudet, J. (2001). Inbreeding, kinship, and the evolution of natal dispersal. In: Clobert, J., Danchin, E., Dhondt, A. A., Nichols, J. D., editors. Dispersal. Oxford: Oxford University Press.
Pope, T. R. (2000). The evolution of male philopatry in neotropical monkeys. In: Kappeler, P.M., editor. Primate Males: Causes and Consequences of Variation in Group Composition. Cambridge: Cambridge University Press.
Port, M., and Johnstone, R. A. (2013). Facing the crowd: intruder pressure, within-group competition, and the resolution of conflicts over group-membership. Ecol. Evol. 3, 1209–1218. doi: 10.1002/ece3.533
Pusey, A. E., and Packer, C. (1987). Dispersal and Philopatry. In: Smuts, B. B., Cheney, D. L., Seyfarth, R. M., Wrangham, R. W., and Struhsaker, T. T., editors. Chicago: Primate societies University of Chicago Press.
Schoof, V. A. M., Jack, K. M., and Isbell, L. A. (2009). What traits promote male parallel dispersal in primates? Behaviour 146, 701–726. doi: 10.1163/156853908X399086
Shutt, K. A., MacLarnon, A., Heistermann, M., and Semple, S. (2007). Grooming in Barbary macaques: better to give than to receive? Biol. Lett. 3, 231–233. doi: 10.1098/rsbl.2007.0052
Smale, L., Nunes, S., and Holekamp, K. (1997). Sexually dimorphic dispersal in mammals: patterns, causes, and consequences. Adv. Study Behav. 26, 181–250. doi: 10.1016/S0065-3454(08)60380-0
Teichroeb, J. A., Wikberg, E. C., and Sicotte, P. (2011). Dispersal in male ursine colobus monkeys (Colobus vellerosus): influence of age, rank and contact with other groups on dispersal decisions. Behaviour 148, 765–793. doi: 10.1163/000579511X577157
van Noordwijk, M. A., and van Schaik, C. P. (1985). Male migration and rank acquisition in wild long-tailed macaques (Macaca fascicularis). Anim. Behav. 33, 849–861. doi: 10.1016/S0003-3472(85)80019-1
van Schaik, C. P., and van Noordwijk, M. A. (2001). Career moves: transfer and rank challenge decisions by male long-tailed macaques. Behaviour 138, 359–395. doi: 10.1163/15685390152032505
Wei, B. M., Kulik, L., Ruiz-Lambides, A. V., and Widdig, A. (2016). Individual dispersal decisions affect fitness via maternal rank effects in male rhesus macaques. Sci. Rep. 6, 32212. doi: 10.1038/srep32212
Xia, D. P., Garber, P. A., Sun, L. X., Sun, B. H., Wang, X., and Li, J. H. (2021). Social strategies used by dispersing males to integrate into a new group in Tibetan macaques (Macaca thibetana). Am. J. Primatol. 2021, e23306. doi: 10.1002/ajp.23306
Xia, D. P., Kyes, R. C., Wang, X., Sun, B. H., Sun, L. X., and Li, J. H. (2019). Grooming networks reveal intra- and intersexual relationships in Tibetan macaques (Macaca thibetana). Primates 60, 223–232. doi: 10.1007/s10329-018-00707-8
Xia, D. P., Li, J. H., Garber, P. A., Matheson, M. D., Zhu, Y., and Sun, B. H. (2013). Grooming reciprocity in male Tibetan macaques. Am. J. Primatol. 75, 1009–1020. doi: 10.1002/ajp.22165
Xia, D. P., Li, J. H., Garber, P. A., Sun, L. X., Zhu, Y., and Sun, B. H. (2012). Grooming reciprocity in female Tibetan macaques (Macaca thibetana). Am. J. Primatol. 74, 569–579. doi: 10.1002/ajp.21985
Xia, D. P., Li, J. H., Zhu, Y., Sun, B. H., Sheeran, L. K., and Matheson, M. D. (2010). Seasonal variation and synchronization of sexual behaviours in free-ranging male Tibetan macaques (Macaca thibetana) at Huangshan, China. Zoological Research 31, 509–515.
Ydenberg, R., Giraldeau, L., and Falls, J. (1988). Neighbours, strangers, and the asymmetric war of attrition. Anim. Behav. 36, 343–347. doi: 10.1016/S0003-3472(88)80004-6
Keywords: Macaca thibetana, immigrant male, bluff, unobtrusive, partnership
Citation: Zhao C-R, Chen S-W, Tian H-W, Sun B, Zhang Q-X and Xia D-P (2023) Variation of social strategies between bluff and unobtrusive immigrant males during integration into a new social group: a case study in Macaca thibetana. Front. Ecol. Evol. 11:1144932. doi: 10.3389/fevo.2023.1144932
Received: 15 January 2023; Accepted: 17 April 2023;
Published: 09 May 2023.
Edited by:
Anindita Bhadra, Indian Institute of Science Education and Research Kolkata, IndiaReviewed by:
Arrilton Araujo, Federal University of Rio Grande do Norte, BrazilCopyright © 2023 Zhao, Chen, Tian, Sun, Zhang and Xia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dong-Po Xia, ZHB4aWFAYWh1LmVkdS5jbg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.